


Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique
by
 , , ,
, , ,
Victorino Américo Buramuge
1,*,
Natasha Sofia Ribeiro
1,
Lennart Olsson
2,
Romana Rombe Bandeira
1 and
Sá Nogueira Lisboa
1,3 1
Department of Forest Engineering, Faculty of Agronomy and Forest Engineering, Eduardo Mondlane University, Maputo 3453, Mozambique
2
LUCSUS—Lund University Centre for Sustainability Studies, 22362 Lund, Sweden
3
N’Lab, Maputo 77, Mozambique
*
Author to whom correspondence should be addressed.
Fire 2023, 6(1), 26; https://doi.org/10.3390/fire6010026
Submission received: 31 October 2022
/
Revised: 12 December 2022
/
Accepted: 29 December 2022
/
Published: 11 January 2023
(This article belongs to the Special Issue Effects of Wildfire on Biodiversity)
Abstract
:Fire strongly impacts the composition and structure of ecosystems, with consequences yet to be understood. We used Moderate Resolution Imaging Spectroradiometer (MODIS) data to map fire frequency and fire intensity and investigate their effects on miombo woodlands (MW) of central Mozambique. Tree species diversity was evaluated and compared using rarefaction curves. Non-metric multidimensional scaling (NMDS) ordination was used to identify patterns of species composition occurrence. The indicator value index method was applied to verify the occurrence of fire indicator species. In general, tree communities responded differently to varied fire regimes. We found low tree density in Intermediate fire frequency and intensity (IfIi) (180 trees ha−1) and High-frequency and Low intensity (HfLi) (316 trees ha−1) areas. The IfIi fire regime had the lowest carbon stocks (9.1 Mg ha−1), when compared to the rest of fire regimes. The species diversity decreased as fire intensity increased. IfIi areas had the maximum species diversity. The NMDS showed a varied species composition according to fire regime. We found a strong relationship between the species diversity and composition, and the pattern of fire occurrence in each fire regime. Our results are critical in supporting fire management policies and understanding fire regimes and their effects on miombo trees’ structure and composition.
1. Introduction
This disturbance factor is dependent on many different conditions such as vegetation type, climate [3] and topography [4]. In addition, the socio-economic context, land management, and policies [5,6] are also important determinants of anthropogenic fires. Changes in any of those factors have direct implications on fire regimes.
The term fire regime refers to the multiple spatial and temporal scale variations of fire [7] such as timing, size, magnitude, frequency, and intensity [7,8]. The study of fire regimes is key for a better understanding of the relationship between fire and ecosystems [9]. Fire frequency (time between fires) and intensity (energy released) [10], are among the most critical factors of natural and anthropogenic fire regimes [11]. Fire frequency determines the length of time plants have to return to their pre-burn state, before the next fire event occurs [12]. Fire intensity is a measure of fire damage to an ecosystem such as impact on adult trees, suppression of regeneration and decline in fauna. These impacts are more evident in the end of dry season [13].
In many landscapes, human activities must be considered for a robust and comprehensive understanding of a fire regime [7], as they represent the dominant source of ignitions and the main drivers of fuel loads [14,15]. Therefore, land use controls the impact of fire on a landscape [16], by changing ecosystem structure and species composition, and potentially increasing the fuel material available, which may perpetuate fires [17]. The link between land uses and fire regimes are thus crucial for generating information on which to base critical management decisions, such as the conservation of biodiversity [18].
Miombo woodlands, covering about 1.9 million km2, are the largest and most important tropical dry forest type in sub-Saharan Africa [19]. These ecosystems are known for their high plant species diversity [19], but with an overwhelming dominance of the Detarioideae subfamily, especially the tree genera Brachystegia, Julbernadia, and Isoberlinia [20]. However, the grass layer and leaf litter comprise about 5% of the total biomass and represent the main fuel load [13,21], making this ecosystem prone to fire [22,23]. The woodlands provide essential goods and services, such as food, medicines, shelter, and energy, supporting the livelihoods of people in the world’s poorest countries [24,25]. In addition, the miombo woodlands play a crucial role in the global carbon balance.
Miombo woodlands are historically related to disturbances such as herbivory and fires and most of the plant species regenerate vigorously after those disturbances through root or stump sprouting [26]. However, despite their high resilience to disturbances [27], the woodlands are under considerable human pressure. Land uses such as charcoal production and agriculture are two key activities in the region, which use fire as a management tool [19]. Human population growth is only amplifying the use of fire. This is particularly important in some remote areas [19] where the fuel load is high [28], which in turn may modify the landscape [14].
Changes occurring in the landscape as a result of fire may eventually lead to a mosaic of different forest patches, which are described as a “fire regime mosaic” [29]. These types of fire regimes can induce varied responses in the ecosystem [30,31]. The rate of fuel load accumulation is also an essential factor in fire spread across a landscape [32]. In addition, fire is part of the miombo ecology and is the only available management tool for rural communities [19,33]. Most of the fuel load accumulates during the wet season and dries up in the dry season when it becomes available to burn [13]. High fuel load and reduced humidity in vegetation and soils [19] in the late dry season favour more intense and destructive fires. The energy emitted by fires determines the degree of effects on the structure and composition of the ecosystem [34], and in fact, species composition may reflect historical fire regime adaptations [35].
The growing human population and climate change are modifying the fire regimes in miombo woodlands [24,36,37], with consequences that are yet to be understood. Fire regimes have been intensively investigated [18,23,38], but only a few studies discuss the impact of fire on ecosystem structure and species composition [23,39]. Furthermore, many of these studies are limited to investigating only one fire regime parameter (e.g., fire frequency) [12,31,38,40]. As a result, little is known about the combined effect of fire frequency and fire intensity on floristic composition and vegetation structure, which is key to informing the development of integrated management action.
In this study, we investigated 18 years of fire history and its effects on the complex land use landscape characterized by the miombo woodlands of the Manica district, in the Beira Corridor of central Mozambique. The corridor has one of the highest deforestation rates (1.8% annually), compared to 0.79%, the national level [41]. Agriculture and charcoal production are the main activities in this region. However, inadequate use of fire results in uncontrolled wildfire propagation that affects extensive areas. The study area lies in the fire-prone region, where over 70% of this fire-prone region burns annually [42] (Figure S1).
Our study aimed to investigate the response of miombo woodlands structure, composition and species diversity to different combinations of fire frequency and fire intensity across the miombo woodlands of the Manica district.
2. Materials and Methods
2.1. Study Area Description
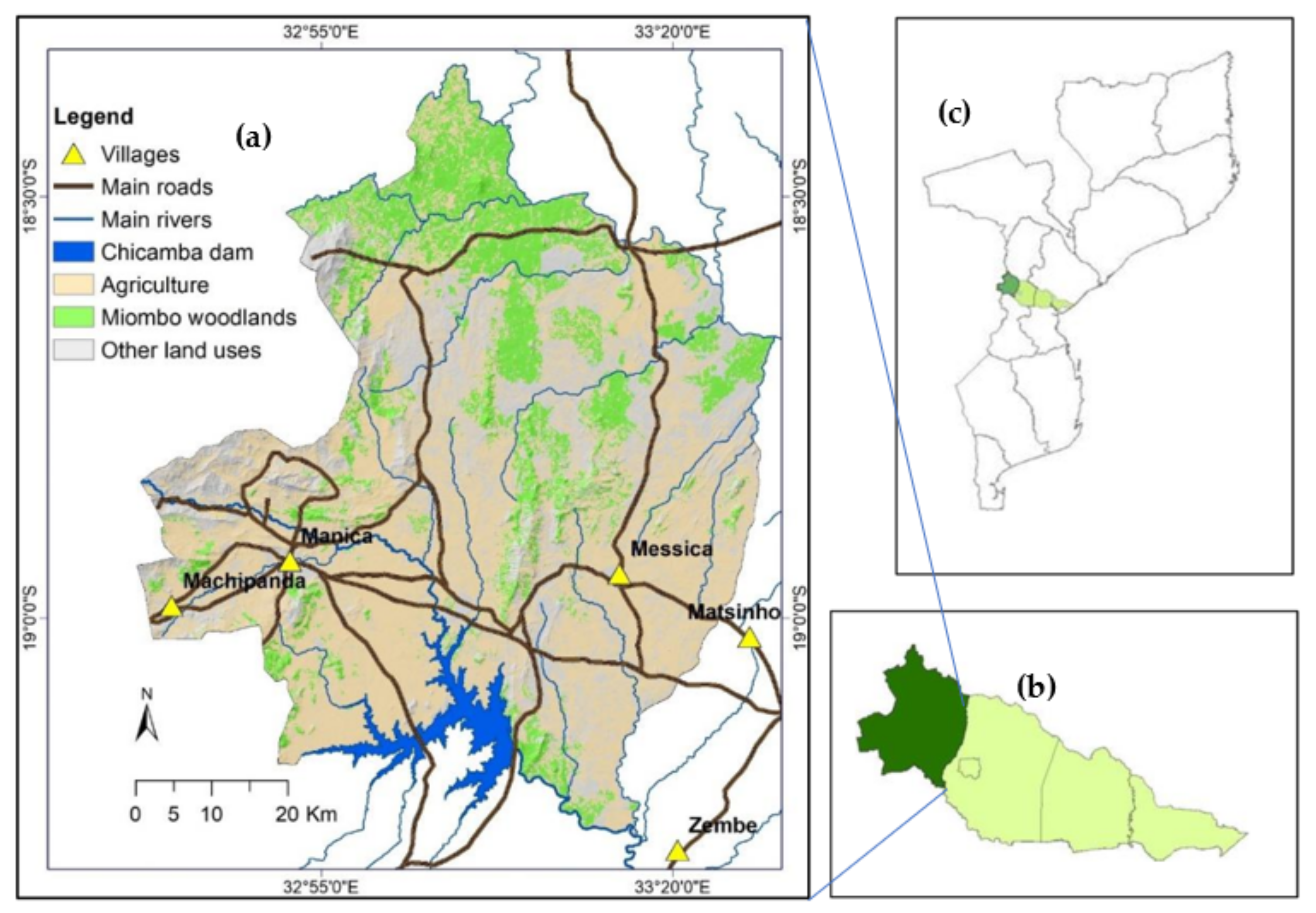
This study was conducted in the miombo woodlands of Manica district, central Mozambique. This district is part of the Beira Development Corridor, one of Mozambique’s six development corridors, linking Mozambique to hinterland countries (Zimbabwe, Malawi, and Zambia). The corridor integrates two provinces of central Mozambique, namely Sofala (Nhamatanda and Dondo districts) and Manica (Gondola and Manica districts). Manica district covers an area of 4594 km2 and borders Barué district in the north, the Sussundenga district in the south, and Gondola district in the east (Figure 1). The Republic of Zimbabwe borders the west for about 120 kilometers [43]. Manica district had about 220,000 inhabitants in 2017, with a density of 76 inhabitants per km2 [44].
The district is drained by the Revué River and its tributaries, which drain into the Búzi River. The topography is gently undulating, but it can reach altitudes of 1500–2000 m, especially on the western side. The soils are reddish brown and deep, have low fertility, and are susceptible to erosion [43]. According to the Köppen classification, the climate is tropical humid, with means annual rainfall ranging from 1000 to 1020 mm between December and April. The mean annual temperature is 21.2 °C, with a maximum temperature of 30.9 °C and a minimum temperature of 14.0 °C [43].
Manica province is rich in floristic diversity, with mosaics of dense forests, open forests, thickets, and shrubs [45]. Forest formations occupy about 27% of the province. Manica district is covered by semi-deciduous forests, including miombo woodlands [41] and other forest formations [45].
Figure 1.
Location of the study area (a) within the Beira Corridor (b); the centre part of the country (c). Land use and land cover data source: Government of Mozambique [41].
Figure 1.
Location of the study area (a) within the Beira Corridor (b); the centre part of the country (c). Land use and land cover data source: Government of Mozambique [41].

2.2. Data Acquisition and Fire Mapping
In this study we used the monthly burned area (Level-3 gridded 500 m MCD64A1) [46] and the daily active fire (Level-5 gridded 1000 m MCD14ML) [47] products of the Moderate Resolution Imaging Spectroradiometer (MODIS) sensor, for the period 2001 to 2018 obtained from https://earthexplorer.usgs.gov/ (accessed on 11 June 2019) and https://firms.modaps.eosdis.nasa.gov/ (accessed on 28 June 2019), respectively. The MODIS MCD64A1 product contains a monthly gridded product of burned areas at 500 m resolution by combining the Terra and Aqua MODIS sensors. The MCD64A1 collection is an improvement of the MCD45A1 product adapted to different conditions in various ecosystems [46]. The MODIS MCD14ML is part of Collection 6, containing information for all Terra and Aqua MODIS fire pixels in daily ASCII files [30]. We retained the active fire products with a high confidence level above 80% to generate the dataset. A high level of active fires confidence is required to reduce false detections [47].
We converted the monthly burnt area data from HDF to GEOTIF format in ArcGIS 10.1 (ESRI, Redlands, CA, USA). We then successively classified the monthly burned area product to produce a burned and non-burned layer. Next, we summed the derived monthly layer to generate an annual dataset. Finally, the resulting yearly burned area layers were aggregated into a final map of fire frequency for 18 years.
To validate the fire frequency map, we performed an accuracy assessment using the error matrix method [48]. The assessment was carried out in 107 field plots over two seasons: dry (October) and wet (April). For each sampled area, visual observations of signs of fire occurrence were conducted, such as charred litter, partially or completely consumed stems, and indications of a charred soil layer [49]. We computed metrics of overall accuracy, user accuracy, and the Kappa statistic. The validation of the burned areas MODIS product, based on confusion matrices, resulted in an overall accuracy of 94.7% and a Kappa statistic of 0.84. The producer’s accuracy in detecting burned-area pixels was 81.3%, where 18.7% of all burned pixels were wrongly classified as non-burned.
From the derived map of fire frequency, we stratified the data into the following three categories of fire frequency, based on the number of times a pixel burned during the 18-year period [31,40,50]: (i) low frequency (every 13–18 years), (ii) intermediate frequency (every 8–12 years), and (iii) high frequency (every 3–7 years).
Using MODIS-derived fire radiative power from the active fire dataset (MCD14ML), we mapped fire intensity by applying the inverse distance weighted method [51]. This method is a non-geostatistical spatial prediction, which estimates the variable over space, weighting the nearest points. It is a function of the inverse of the power of distance. Thus, the closer the sample point is to the point to be estimated, the greater the weight attributed to the sampled point [52]. Since the MODIS active fire dataset provides values, we defined limit ranges of fire intensity. We generated the fire intensity map in the same format (classes) as the fire frequency map to allow us to combine the two class groups of fire regimes. The fire intensity was classified into the following categories: (i) high intensity (137.2–681.5 MW), (ii) intermediate intensity (66.2–137.1 MW), and (iii) low intensity (11.0–66.1 MW).
This procedure resulted in nine combinations of fire frequency and fire intensity (hereafter fire stratum), but only those with representative field plots (n > 4) were used in this study, namely, (i) High-frequency vs. High-intensity (HfHi), (ii) High-frequency vs. Low-intensity (HfLi), (iii) Intermediate-frequency vs. Intermediate-intensity (IfIi), (iv) Low-frequency vs. High-intensity (LfHi) and (v) Low-frequency vs. Low-intensity (LfLi). We determined the categories of land use and land cover of each stratum through field observations carried out during the wet and dry seasons (figure). We obtained the estimated period of fallow land (age of regrowth areas) based on information provided by the landowner. We corroborated the information provided with field observations (size of trees, canopy cover, indications of abandoned settlement areas, signs of decommissioned charcoal kilns, croplands abandonment, and indications of human accessibility).
However, there were limitations in identifying non-burned areas. Fire is part of Miombo management, and it affects most of the areas. Despite this limitation, low fire frequency areas were used to compare to intermediate and high fire occurrence areas.
2.3. Field Data Collection
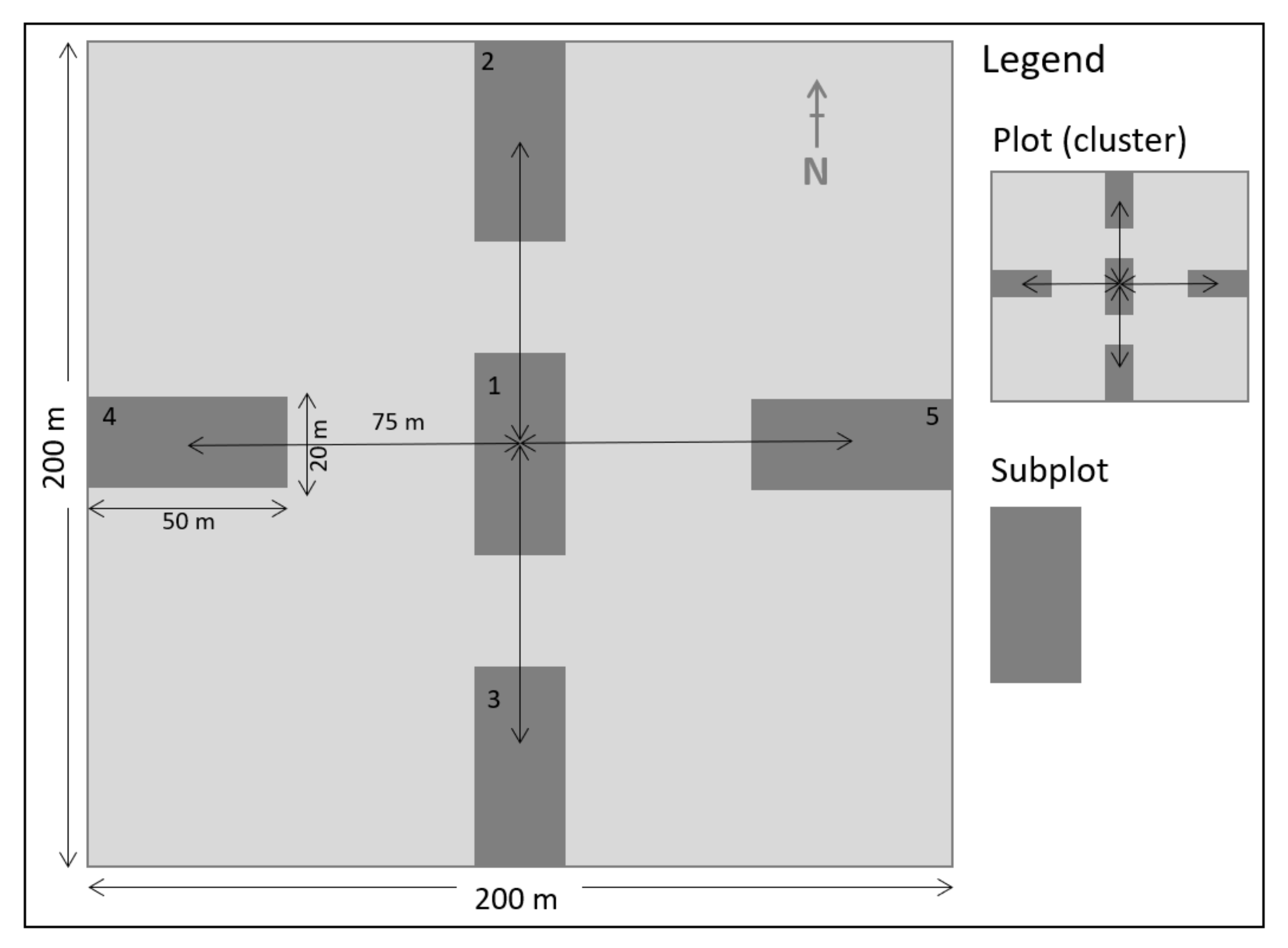
In each fire stratum (Table 1), we applied a random sampling strategy. At each sample point, we set up two 200 m perpendicular transects (north–south and east–west) and established a cluster consisting of five rectangular (50 m × 20 m) subplots: One in the center and four subplots at the end of the transects (Figure 2). Following this design, we established 20 subplots in HfHi, 25 in HfLi, 17 in IfIi, 15 in LfHi, and 30 in LfLi, totaling 107 subplots. All live trees with a diameter at breast height (DBH, 1.30 m) ≥ 5 cm, were included in the survey and measured for DBH with a diametric tape. The total tree height (m) was estimated using graduated sticks. The botanical identification made in the field relied on local botanists. Species with dubious identification had samples collected and tagged for later identification and registration in the herbarium of Eduardo Mondlane University.
2.4. Data Analysis
We carried out a vegetation structure and composition analysis for each stratum. First, we applied a rarefaction approach to compare the species richness and diversity among the fire strata, using iNEXT package [53] available in R software. We then constructed rarefaction and extrapolation curves for the five strata, using Hill numbers to obtain the effective number of species. In the rarefaction approach, q = 0 is for species richness, q = 1 is for Shannon’s diversity index, and q = 2 is for Simpson’s diversity index [53]. The rarefaction curves were tested for statistical differences, using the overlapping criteria at 95% of confidence [54].
In addition, we employed the indicator value index (IndVal) method to verify the occurrence of habitat site indicator species. Indicator species preferentially occur in specific conditions, such as disturbance sites [55]. The IndVal method is based on the species’ degree of habitat specificity or exclusivity to a given habitat, and the fidelity or sensitivity of the species as an indicator of the target site group [56]. IndVal values are expressed as percentages.
Moreover, we used the importance value index (IVI) to assess the ecological weight of tree species. The IVI (Equation (4)) was calculated as the sum of relative frequency (Equation (1)), relative density (Equation (2)), and relative dominance (Equation (3)), for adult trees with DBH ≥ 5 cm [57]
We also computed average tree density (N ha−1) and aboveground biomass (AGB, Mg ha−1). The AGB was derived from the allometric equation for miombo woodlands of the study area developed by Guedes et al. [58] as follows:
where tDW is the total aboveground dry weight (Mg ha−1) and DBH is the diameter at breast height (cm).
Based on Equation (5), according to Guedes et al. [58], we estimate the carbon stock in Mg ha−1 (Equation (6)):
To compare the means of the vegetation parameters (tree density and carbon stock), we used the Kruskal–Wallis test (for non-normally distributed data) followed by Dunn’s test at the 5% significance level.
We examined the similarity patterns of tree species among strata, using non-metric multidimensional scaling (NMDS), based on the Bray–Curtis similarity index. The NMDS analysis identifies patterns of species composition occurrence in different locations and determines which environments are (dis)similar [59]. In addition, we used a nonparametric permutation procedure of analysis of similarities (ANOSIM) to test for differences in the compositions of the groups formed by the NMDS analysis (9999 permutations) [59].
3. Results
3.1. Tree Species Composition and Structure across Fire Regimes
Our findings reveal that in general, the ecosystem was diverse. More than 5400 trees with DBH > 5 cm were sampled, representing 112 species and 33 families. The richest families were Fabaceae (36 species); Combretaceae (nine species); Anacardiaceae, Phyllanthaceae, and Rubiaceae, each with six species.
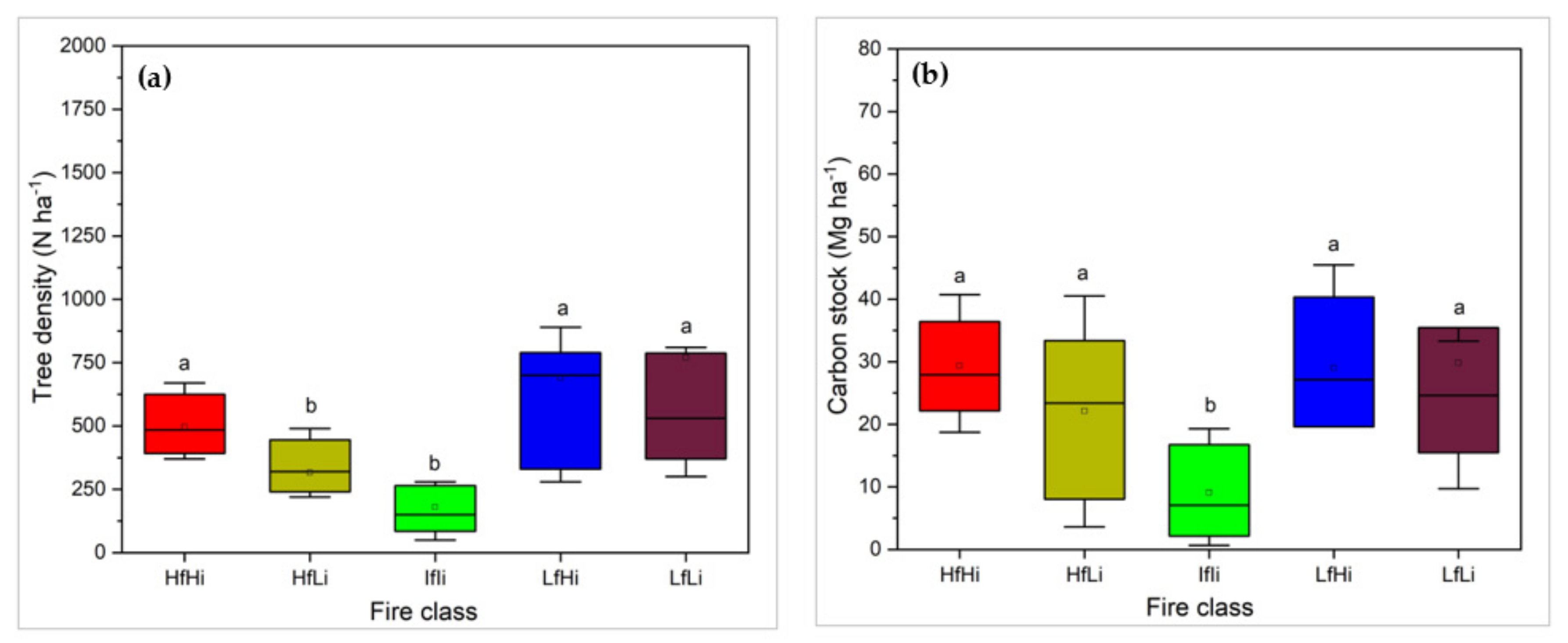
Results revealed significant differences in tree density (𝝌2 = 51.19, df = 4, p < 0.001) and carbon stock (𝝌2 = 27.46, df = 4, p < 0.001) among strata. According to Figure 3a, HfLi and IfIi had lower tree density (316 and 180 trees ha−1, respectively) than the other strata. However, no significant differences (p < 0.05) were found between HfHi and the low fire frequency areas (LfHi and LfLi). The tree density in the LfLi was 530 trees ha−1, 700 trees ha−1 in LfHi, 495 trees ha−1 in HfHi and 316 trees ha−1 in HfLi strata.
The carbon stock value is significantly lower under the IfIi stratum (9.1 Mg ha−1). However, significantly higher values were found in mature forests under HfHi (29.3 Mg ha−1) and HfLi (22.1 Mg ha−1) and the regrowth areas under 28.9 Mg ha−1 for LfHi and 29.9 Mg ha−1 for LfLi (Figure 3b).
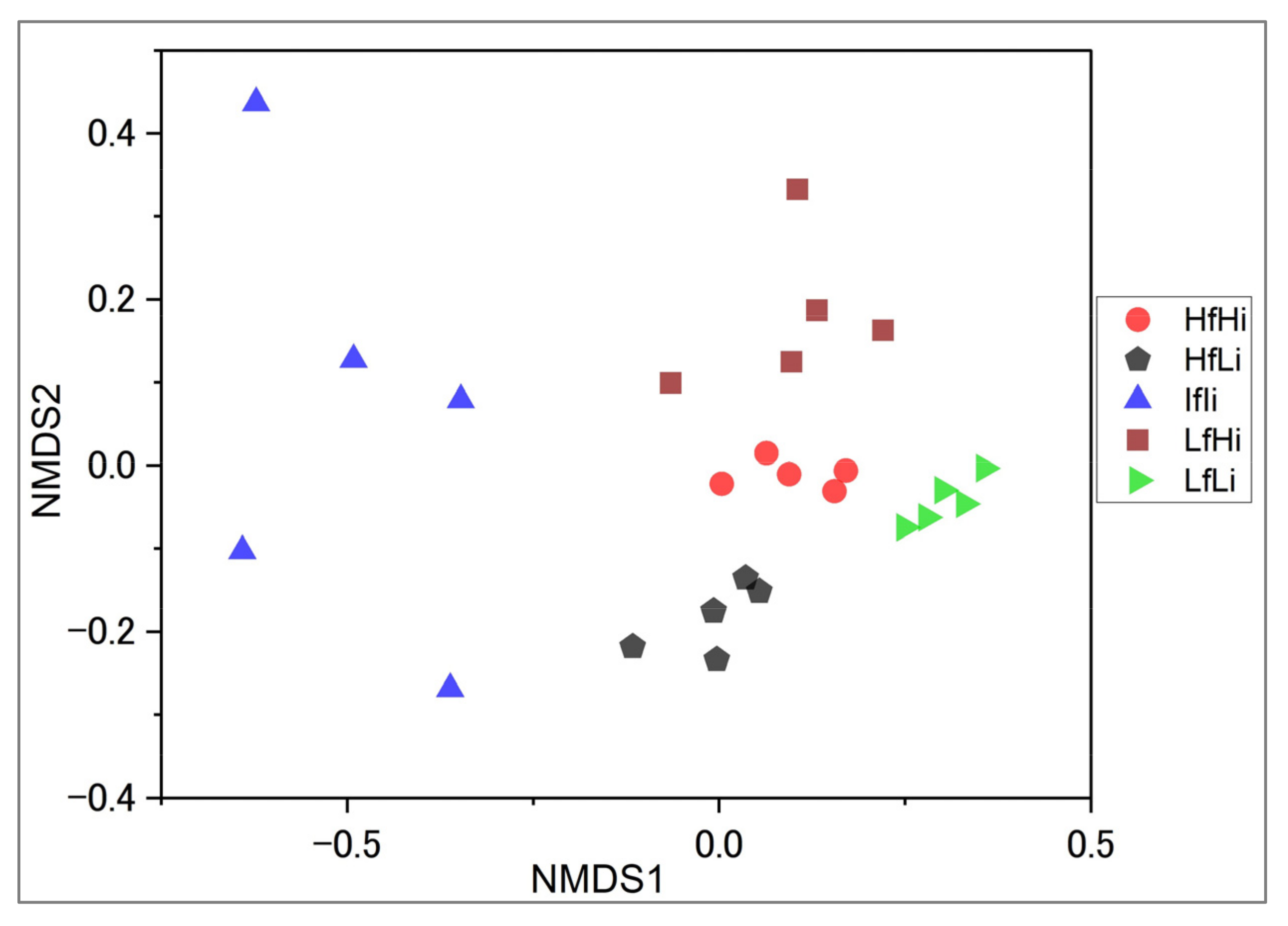
The NMDS ordination of species composition grouped samples from different strata. Overall, separations among all fire regimes were significantly expressed, as shown in Figure 4. In addition, the species composition varied substantially between fire strata (ANOSIM: Stress = 0.16; R2 = 0.63, p < 0.001, 9999 permutations). Of the 111 species, 82 contributed significantly to the variation in tree community ordination in NMDS.
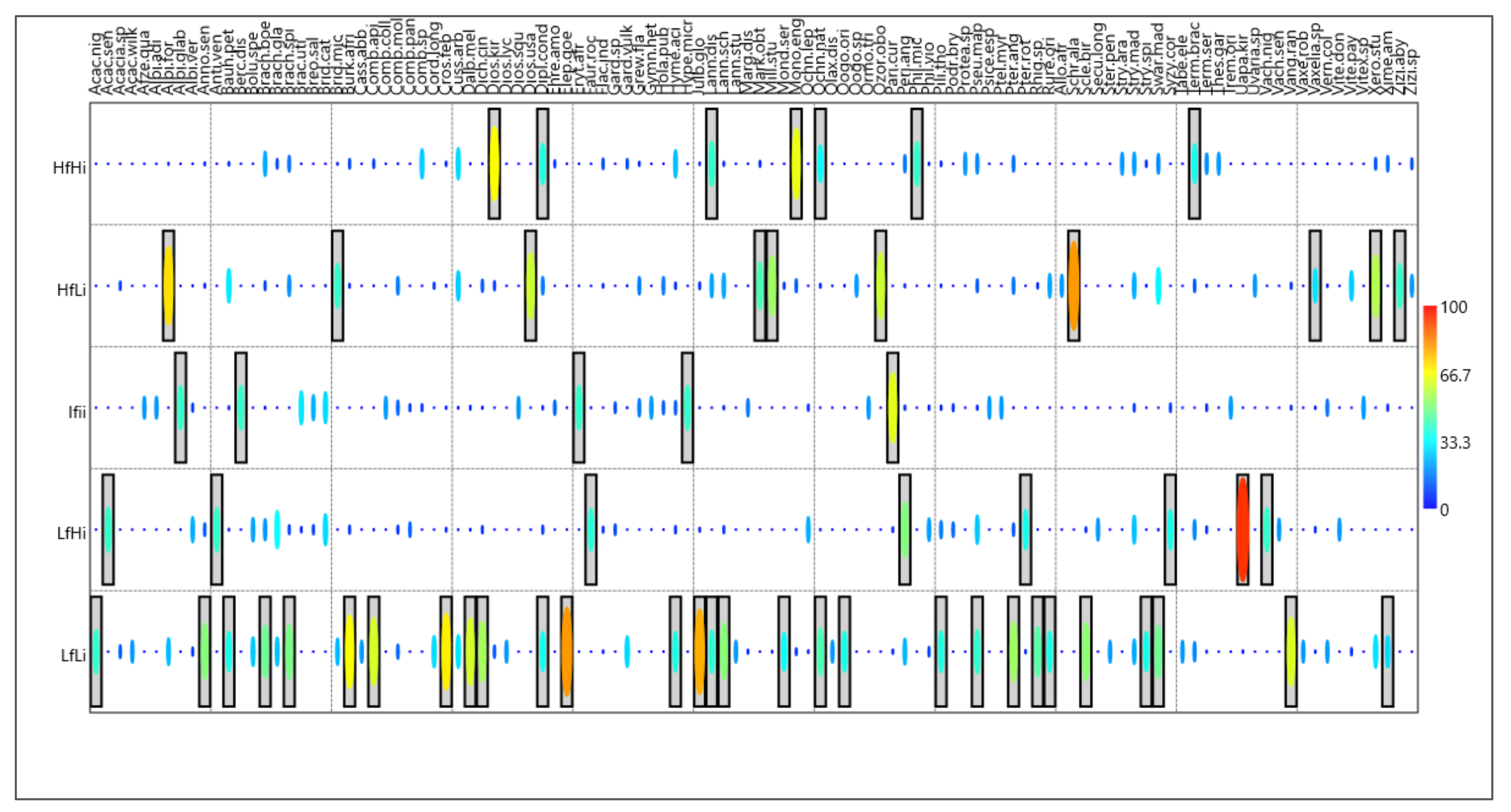
The IndVal method revealed indicator species associated with different strata (Figure 5, species codes Table S4). Seven species were significantly associated with the HfHI fire stratum, 10 with HfLi, five with IfIi, eight with LfHi, and the highest number of species were found for LfLi (29 species). Diplorhynchus condylocarpon, Terminalia brachystemma, Monotes glaber, Parinari curatellifolia, Albizia forbesii, Lannea discolor, and Swartzia madagascariensis are all associated with high fire frequency areas. Brachystegia spiciformis, Brachystegia glaucescens, Brachystegia utilis, and Julbernardia globiflora, the main canopy species in miombo woodlands, are usually indicators in areas where fire occurrence is low (LfLi). These species are assumed to be fire-sensitive as seedlings. Uapaca kirkiana appears to be more important in areas in which fires are infrequent.
3.2. Tree Species Diversity
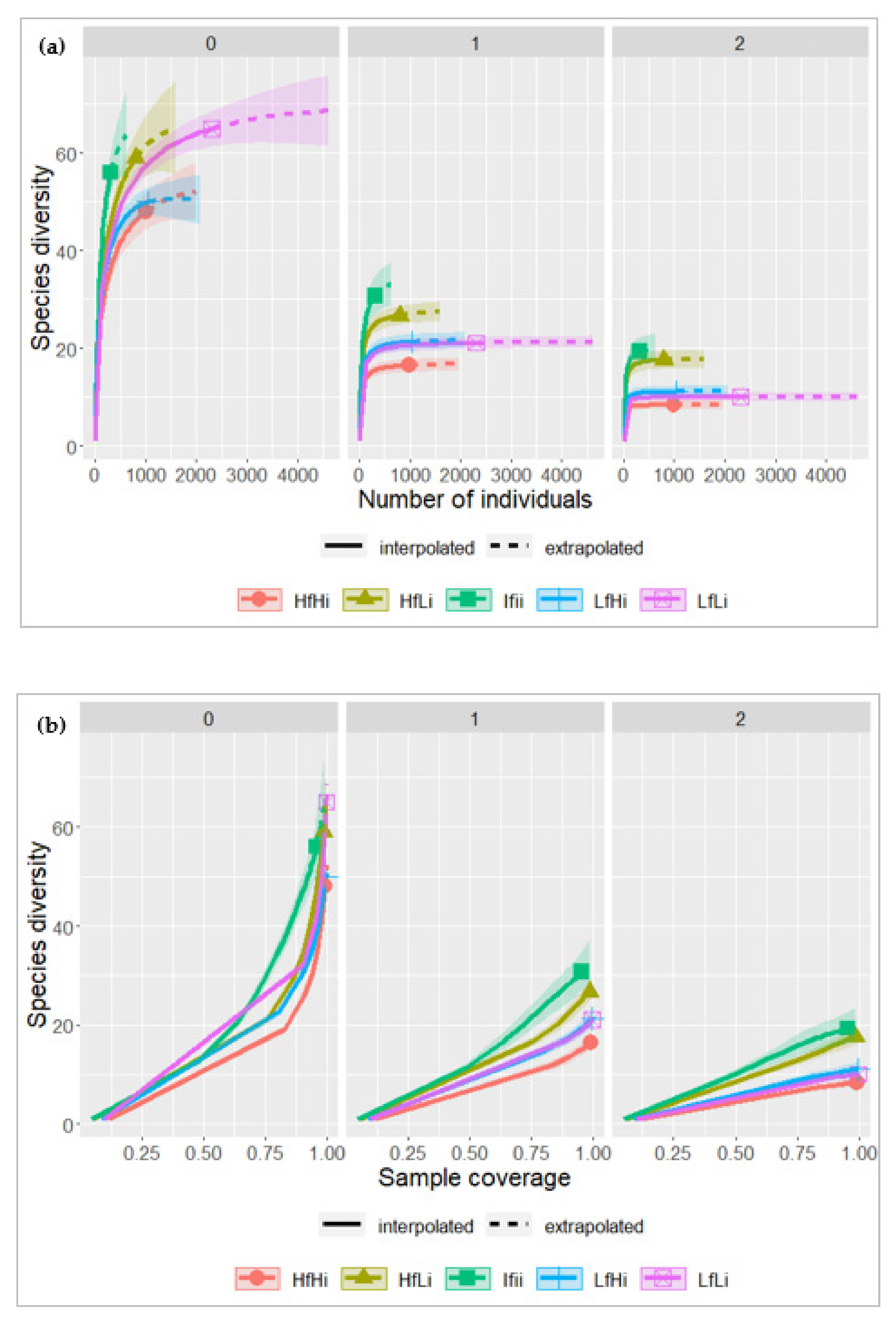
We evaluated tree species diversity based on the total number of species in each fire regime and performed a rarefaction curve of species richness and diversity as shown in Figure 6. The rarefaction and extrapolation curves showed higher species richness in the IfIi stratum, where 56 taxa (q = 0) were detected, Shannon’s diversity index was 30.8 (q = 1) and the inverse Simpson diversity index was 19.4 (q = 2). The HfHi and LfHi strata showed lower species richness, with 48 and 50 taxa (q = 1), respectively. The results reveal that when the intensity of a fire increases, the species richness decreases, both for low and high frequency, despite the lack of stabilization of the rarefaction curve. The Shannon and Simpson diversity indices followed the same pattern, with the highest values in the IfIi areas and the lowest in the HfHi areas.
3.3. Tree Species Importance Value across Fire Regimes
The most dominant species are represented by nine families, of which the Fabaceae were the most important, represented by 12 species, followed by Phyllanthaceae with three species. The IVI values in Table 2, show that different species responded differently to the variation in fire occurrence. Brachystegia boehmii, D. condylocarpon, Pseudolachnostylis maprouneifolia and Pterocarpus angolensis were represented in all strata. Due to their fire adaptations, these species are considered fire-tolerant trees in adult stages [60].
We analyzed the IVI by including all fire strata as a single assemblage and found that B. boehmii had the highest relative ecological importance in all strata. B. boehmii is one of the emblematic species of miombo woodland, and we expected to record it everywhere. A relatively low IVI of B. boehmii was reported under the HfLi (47%) and IfIi (31.8%) strata, while the highest value was found under the LfLi (75.8%) and HfHi (74.7%) fire regimes.
The same trend reported for B. boehmii was also observed for D. condylocarpon, P. maprouneifolia, and P. angolensis, where the lowest values were found in HfLi strata. Nevertheless, P. maprouneifolia was more dominant in LfHi areas (28.0%). Pericopsis angolensis, a valuable commercial wood, was prevalent in all fire strata except HfLi. J. globiflora and B. spiciformis, two of the most important species of miombo woodlands, were dominant under the LfLi areas.
4. Discussion
4.1. Effects of Fire Regime on Woody Structure and Composition
Our results reveal that fire strongly affected the species composition and vegetation structure of the miombo woodlands in Manica district, central Mozambique. Frequent disturbances may determine successional responses by altering the species composition and vegetation structure [61,62]. Fabaceae and Combretaceae were among the most important families in the study area. The Fabaceae family dominates miombo woodlands, distinguishing them from other types of forests [13].
Observed similarities in tree density between mature forests (HfHi) and regrowth areas (LfHi and LfLi), suggest a rapid woodland recovery in the areas that had been previously disturbed [26]. Regeneration by root or stump sprouting is the primary recovery process of miombo woodlands when disturbed [26,27]. However, root or stump sprouting is associated with vigorous growth, good competitive capabilities and a high stem survival rate [27]. The ability of many miombo species to recover by root or stump sprouting following disturbance has been reported by several authors [27,63].
Areas under the HfLi and IfIi strata may have good human accessibility for charcoal production, cultivation and other extractive activities, contributing to the relatively low tree density and carbon stock. Human accessibility is likely to represent an additional disturbance factor that exerts intense human pressure on the remaining forest fragments. Miombo woodlands are consistently under pressure because of the increasing human demand for forest resources. Resource use is often unsustainable, resulting in forest loss and degradation [64]. The impact of fire may involve the loss and degradation of terrestrial ecosystems, affecting vegetation composition and structure [2,65]. When the fires are repeated, seed source depletion can affect the forest’s regenerative capacity [65]. Intensive and long-term land use may reduce the ecosystem’s ability to recover and have a negative impact on tree density [19].
Carbon stock was significantly lower under intermediate fire frequency and intensity (IfIi). Human pressure from logging and clearing for agriculture, in combination with fire, may have contributed to the significant decrease in carbon stock in this stratum. Long-term fire frequency has the potential to reduce forest carbon stock [66] and tree density, affecting the capacity of miombo to sequester carbon [19]. Nevertheless, values in low-fire-frequency areas (regrowth areas) are similar to those found in mature forests. This indicates the ecological importance of regrowing miombo woodlands in the carbon balance, suggesting that low fire occurrence combined with fallow periods positively affects the recovery of miombo woodland. These findings are consistent with the results of [67]. They observed that areas previously used for charcoal production and agriculture showed remarkable natural recovery and the potential for carbon sequestration.
The description and characterization of tree communities using NMDS ordination analysis were demonstrated to be effective in confirming the occurrence and association of species, according to different fire strata. Despite the influence of fire, it is worthwhile highlighting other factors that may have influenced the occurrence of species associations. For instance, human activities heavily affect miombo woodlands [64] through deforestation and forest degradation activities. The division of the communities into groups in the NMDS analysis confirms the existence of a mosaic of different fire regimes.
The indicator species analysis showed that some species were indicative of the specific characteristics of a stratum. Generalist species that occur in multiple groups are not considered good indicators since a perfect indicator species must occur and be restricted to one particular group [55]. Typical species such as D. condylocarpon (26.5% IVI) and T. brachystemma (Combretaceae; 11% IVI), were significant in areas of frequent burning (HfHI). The presence of D. condylocarpon and species of the genus Combretum is important since they are deemed indicators of areas prone to repeated fires [19]. Both species are fire-tolerant because of their resistance to fire effects and ability to tolerate and thrive under repeated burning [60,68]. In low-burning areas, the presence of main miombo species such as B. boehmii, B. spiciformis, B. glaucescens, B. utilis, and J. globiflora was most important. These species are considered fire-sensitive; they have high mortality rates in late-season fire at young stages, and thrive under low fire occurrence [33].
The presence of U. kirkiana (IVI of 14.2%) in regeneration areas (LfHi), demonstrates the importance of this species as indicator. U. kirkiana has been used as an indicator of poor soils since it is found primarily in soils with low fertility [69]. Although it is considered semi-fire tolerant, juvenile trees and seedlings are not resistant to fire, especially at the end of the dry season [33]. This species is particularly important since its fruits are used as a food source in subsistence economies, improving food and nutritional security as livelihoods for rural inhabitants [70].
4.2. Tree Species Diversity
Fire usually causes an immediate decline in biodiversity, resulting in lower local diversity [71,72]. We found that the species diversity and richness varied with the fire regime. Tree species richness decreased as fire disturbance increased. Across the five fire strata, species richness was highest under moderate fire frequency and intensity (IfIi).
However, regardless of fire frequency, species richness decreased when fire intensity was high, as shown in Figure 6. The same pattern was observed for Shannon’s and Simpson’s inverse diversity indexes. The decline in species richness observed in areas with high fire intensity may be due to the exclusion of species that are not fire-adapted [71]. Severe fires can affect the species’ survival by reducing their ability to recolonize the area. At the same time, intermediate disturbances induce different post-fire responses, maintaining more species with varying ecological requirements [73].
Although the miombo woodlands are considered a fire-tolerant ecosystem, many species respond differently to the effect of fires. Fire-resistant species can establish rapidly under frequent and intense fire disturbance [13,60]. Conversely, many key miombo species which are fire sensitive, such as Julbernardia and Brachystegia genera, tend to be less represented in areas with frequent fires [19].
The peak of species diversity at intermediate fire frequency and intensity (IfIi) is consistent with the intermediate disturbance hypothesis, which states that species diversity is highest in communities subject to intermediate disturbance, compared with communities subject to lower or higher [74]. The occurrence of intermediate fires possibly resulted in more heterogeneous conditions leading to spatial heterogeneity and consequently sustaining high species diversity [35]. [75] found that rare or frequent fires can result in the local extinction of species in fire-prone landscapes.
This study is the first in miombo woodlands supporting the assumption that high species diversity occurs at the intermediate disturbance, as stated by the intermediate disturbance hypothesis supported in several studies [76,77,78,79]. However, there is a trend in the similarity of species diversity between mature forests and regrowth areas (HfLi and LfLi; HfHi and LfHi). This trend is expected because miombo woodlands recover quickly after disturbances, reaching similar levels to mature forest [80]. The relatively low values of species richness under the HfHi and LfHi regimes suggest that these regimes negatively impact species richness and diversity. However, despite intense disturbance, the Shannon diversity index ranged from 2.8 to 3.5, indicating high resilience and stability in species diversity and richness. The overall species diversity values in the study area were consistent with those reported by [81] in the miombo region, ranging from 3.3 to 3.59. [64] estimated species diversity values of 2.86 to 3.44 for miombo woodlands subjected to different degrees of disturbance.
4.3. Tree Species Importance Value across Fire Regimes
The fire regime analysis showed that B. boehmii, D. condylocarpon, P. maprouneifolia, and P. angolensis are four of the ten most ecologically important species that were well represented in all five fire regimes. These species are known to be fire-tolerant in adult stages [13,60], and repeated fires promote their development [13,25]. However, B. boehmii dominated all strata, showing the highest values in low fire frequency and intensity (IVI, 75.8%). Recently it has been recognized that the tolerance of B. boehmii under high-fire occurrence is mainly attributed to its genetic diversity, which contributes to its high survival rate [82]. In contrast, the juvenile trees of this species are more vulnerable to frequent fires [60].
Although the B. boehmii was well represented in all fire regime strata, the HfLi and IfIi strata had relatively low IVI values (47% and 31.8%, respectively). These areas have different land use dynamics, characterized by agriculture, charcoal production, and timber exploitation, among other activities (Table 1). B. boehmii is among the most preferred miombo species for firewood and charcoal production [83]. Charcoal production is deemed one of the leading causes of forest degradation as it is highly selective in terms of preferred species [84].
The success of the Combretaceae family in both high and low-fire frequency areas may be attributed to their ability to resist fires and colonize areas that experienced extensive land use [68]. The species of this family generally become the most dominant in the early stages of succession because of their rapid growth [85]. Indeed, [64] noted that species from this genus predominated in areas where land use was intensive and when they had been subjected to fire.
In contrast, D. condylocarpon dominated areas with frequent and intense fires. [68] observed a similar pattern: this species was more abundant in repeated burnt plots. These findings broadly support the previous evidence that this species is fire-tolerant. Its thick bark and the high moisture content of the wood play an essential role in protecting against the effects of fire, especially when the fire intensity is high [63].
It was expected that P. angolensis would be dominant in areas with frequent and intense fires since this species is fire-resistant and fire-dependent to germinate [86]. The thick bark allows the trees to tolerate adverse conditions such as fire and drought [87] and seeds only germinate after fire exposure [88]. This species is classified as producing high-quality wood and is preferred for timber exploitation in Mozambique. Selective extraction may have led to a reduction of its dominance under the HfLi and IfIi strata. In East and Southern Africa, over-exploitation as a result of its commercial value has led to a considerable decline in this species [89]. The dominance in areas where the fire was not frequent and intense (regrowing areas) suggests the potential of this species to recover under low fire disturbance. Pericopsis angolensis dominates in areas where high-intensity fires are infrequent (LfHi). This species has been considered fire-resistant [73]. Previously known as a low-quality timber species, it is currently in high demand in national and international markets. It is important to stress that HfLi stratum areas are located in a mature forest with intense human activity, which may explain the low dominance of this species because of the increased demand.
Although B. spiciformis and J. globiflora are the two key miombo woodland species, they were dominant in low-fire frequency areas. These species are considered fire-sensitive under typical fire conditions and show a marked decline, mainly when the fire occurs at the end of the dry season [25]. In Ref. [64] found that these species were absent in areas where land was intensely managed by fire.
5. Conclusions
This study showed that tree species respond differently to the effects of fires. The maximum species diversity was found in intermediate fire frequency and intensity areas. However, an inverse relationship between species diversity and fire intensity was observed. Despite controversial opinions, this finding is consistent with the intermediate disturbance hypothesis, which states that species diversity is maximized at intermediate disturbance levels [74]. The results of this study also corroborate the observation that when disturbed, miombo woodlands quickly regenerate, maintaining their capacity to sequester carbon. The NMDS analysis confirmed that the species composition was aggregated by fire regime strata, which improves our understanding of the role of fire in the distribution of tree communities. Indicator species analysis clearly showed a strong relationship between the species and the ecological condition of the strata, demonstrating the method’s potential to group species based on their fire tolerance. This finding is critical in monitoring tree community variations in response to miombo disturbances.
The management of sensitive species such as B. spiciformis and J. globiflora, the two key miombo woodland species, may overcome the challenge of managing fire in miombo woodland. However, the ecology of many miombo tree species remains poorly understood. Much of our knowledge of the response of miombo trees to fire comes from [33,60,68]. Research covering other species is needed to better understand their responses to the fire effects. Our findings suggest that varying fire regimes have different effects on miombo tree species, depending on the frequency and intensity of the fire.
Therefore, fire management policies can benefit from considering different fire regimes as distinct units, that can be used to predict vegetation responses. Furthermore, prescribed fires under varying frequencies and intensities are critical for biodiversity conservation by allowing species to persist in different ranges of fire tolerance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fire6010026/s1.
Author Contributions
Conceptualization, V.A.B., N.S.R., L.O. and R.R.B.; Formal analysis, V.A.B., N.S.R., L.O., R.R.B. and S.N.L.; Funding acquisition, N.S.R., L.O. and R.R.B.; Methodology, V.A.B., N.S.R., L.O., S.N.L. and R.R.B.; Supervision, N.S.R., L.O. and R.R.B.; Validation, N.S.R., L.O., S.N.L. and R.R.B.; Writing—original draft, V.A.B.; Writing—review & editing, V.A.B., N.S.R., L.O. and S.N.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study received financial support from Swedish International Development Cooperation Agency (SIDA), Grant number 51140073, Subprogram 123, and from Eduardo Mondlane University. Av. Julius Nyerere, 3453, Campus Universitario, Building #1, P.O. Box 257, Maputo, Mozambique.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Acknowledgments
We thank Célia Martins, Romano Guambe, and Ernesto Boane for the field data collection support; Lund University Centre for Sustainability Studies for the linguistic reviewer support on the manuscript. In addition, we thank the three anonymous reviewers for their comments.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Bond, W.J.; Keeley, J.E. Fire as a Global “Herbivore”: The Ecology and Evolution of Flammable Ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, C.E.R.; Anderson, T.M.; Sankaran, M.; Higgins, S.I.; Archibald, S.; Hoffmann, W.A.; Hanan, N.P.; Williams, R.J.; Fensham, R.J.; Felfili, J.; et al. Savanna Vegetation-Fire-Climate Relationships Differ Among Continents. Science 2014, 343, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Sharples, J.J. Review of formal methodologies for wind—slope correction of wildfire rate of spread. Int. J. Wildland Fire 2008, 17, 179–193. [Google Scholar] [CrossRef]
- Lentile, L.B.; Holden, Z.A.; Smith, A.M.S.; Falkowski, M.J.; Hudak, A.T.; Morgan, P.; Lewis, S.A.; Gessler, P.E.; Benson, N.C. Remote sensing techniques to assess active fire characteristics and post-fire effects. Int. J. Wildland Fire 2006, 15, 319–345. [Google Scholar] [CrossRef]
- Miettinen, J.; Liew, S.C. Connection between fire and land cover change in Southeast Asia: A remote sensing case study in Riau, Sumatra. Int. J. Remote Sens. 2005, 26, 1109–1126. [Google Scholar] [CrossRef]
- Whitlock, C.; Higuera, P.E.; Mcwethy, D.B.; Briles, C.E. Paleoecological Perspectives on Fire Ecology: Revisiting the Fire-Regime Concept. Open Ecol. J. 2010, 3, 6–23. [Google Scholar] [CrossRef] [Green Version]
- Krebs, P.; Pezzatti, G.B.; Mazzoleni, S.; Talbot, L.M.; Conedera, M. Fire regime: History and definition of a key concept in disturbance ecology. Theory Biosci. 2010, 129, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. BioScience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of Fire Regime and Habitat on Tree Dynamics in North Florida Longleaf Pine Savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- Gandiwa, E. Effects of Repeated Burning on Woody Vegetation Structure and Composition in a Semi-Arid Southern Africa Savanna. Int. J. Environ. Sci. 2011, 2, 458–471. [Google Scholar]
- Frost, P. The Ecology of Miombo Woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B., Ed.; Centre for International Forestry Research (CIFOR): Bogor, Indonesia, 1996; pp. 11–58. [Google Scholar]
- Eva, H.; Lambin, E. Fires and Land-Cover Change in the Tropics: A Remote Sensing Analysis at the Landscape Scale. J. Biogeogr. 2000, 27, 765–776. Available online: http://www.Jstor.Org/Stable/2656223 (accessed on 27 November 2019). [CrossRef]
- Chuvieco, E.; Mouillot, F.; van der Werf, G.R.; San Miguel, J.; Tanase, M.; Koutsias, N.; García, M.; Yebra, M.; Padilla, M.; Gitas, I.; et al. Historical background and current developments for mapping burned area from satellite Earth observation. Remote Sens. Environ. 2019, 225, 45–64. [Google Scholar] [CrossRef]
- Viedma, O.; Moreno, J.M.; Rieiro, I. Interactions between land use/land cover change, forest fires and landscape structure in Sierra de Gredos (Central Spain). Environ. Conserv. 2006, 33, 212–222. [Google Scholar] [CrossRef]
- Flasse, S.P.; Trigg, S.N.; Ceccato, P.N.; Perryman, A.H.; Hudak, A.T.; Thompson, M.W.; Brockett, B.H.; Drame, M.; Ntabeni, T.; Frost, T.E.; et al. Remote Sensing of Vegetation Fires and Its Contribution to a Fire Management Information System. In Wildland Fire Management Handbook For Sub-Sahara Africa; Goldammer, d.R., Ed.; Global Fire Monitoring Center (GFMC): Mbombela, South Africa, 2004; ISBN 191983365X. [Google Scholar]
- Archibald, S.; Scholes, R.; Roy, D.P.; Roberts, G.; Boschetti, L. Southern African fire regimes as revealed by remote sensing. Int. J. Wildland Fire 2010, 19, 861–878. [Google Scholar] [CrossRef] [Green Version]
- Dziba, L.; Ramoelo, A.; Ryan, R.; Harrison, S.; Pritchard, R.; Tripathi, H.; Sitas, N.; Selomane, O.; Engelbrecht, F.; Pereira, L.; et al. Biogeography and Ecology of Miombo Woodlands. In MW in a CES the R and S of P and Woodlands; Ribeiro, N.S., Katerere, Y., Chirwa, P.W., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Moura, I.; Maquia, I.; Rija, A.A.; Ribeiro, N.; Ribeiro-Barros, A.I. Biodiversity Studies in Key Species from the African Mopane and Miombo Woodlands. In Genetic Diversity; Bitz, L., Ed.; IntechOpen: London, UK, 2017; pp. 91–109. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.S.; Matos, C.N.; Moura, I.R.; Washington-Allen, R.A.; Ribeiro, A.I. Monitoring vegetation dynamics and carbon stock density in miombo woodlands. Carbon Balance Manag. 2013, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.; Frost, P.; Byron, N. Miombo Woodlands and Their Use: Overview and Key Issues. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B.M., Ed.; Center for International Forestry Research: Bogor, Indonesia, 1996; pp. 1–10. ISBN 9798764072. [Google Scholar]
- Saito, M.; Luyssaert, S.; Poulter, B.; Williams, M.; Ciais, P.; Bellassen, V.; Ryan, C.M.; Yue, C.; Cadule, P.; Peylin, P. Fire regimes and variability in aboveground woody biomass in miombo woodland. J. Geophys. Res. Biogeosci. 2014, 119, 1014–1029. [Google Scholar] [CrossRef] [Green Version]
- Dewees, P.A.; Campbell, B.M.; Katerere, Y.; Sitoe, A.; Cunningham, A.B.; Angelsen, A.; Wunder, S. Managing the Miombo Woodlands of Southern Africa: Policies, Incentives and Options for the Rural Poor. J. Nat. Resour. Policy Res. 2010, 2, 57–73. [Google Scholar] [CrossRef] [Green Version]
- Chidumayo, E.N.; Timberlake, J.; Sawadogo, L. Distribution and Characteristics of African Dry Forests and Woodlands. In The Dry Forests and Woodlands of Africa: Managing for Products and Services; Chidumayo, E.N., Gumbo, D.J., Eds.; Earthscan: London, UK, 2010; pp. 11–41. [Google Scholar]
- Syampungani, S.; Geldenhuys, C.J.; Chirwa, P.W. Regeneration dynamics of miombo woodland in response to different anthropogenic disturbances: Forest characterisation for sustainable management. Agrofor. Syst. 2015, 90, 563–576. [Google Scholar] [CrossRef]
- Chidumayo, E. Development of Brachystegia-Julbernardia woodland after clear-felling in central Zambia: Evidence for high resilience. Appl. Veg. Sci. 2004, 7, 237–242. [Google Scholar] [CrossRef]
- Tarimo, B.; Dick, Ø.B.; Gobakken, T.; Totland, Ø. Spatial distribution of temporal dynamics in anthropogenic fires in miombo savanna woodlands of Tanzania. Carbon Balance Manag. 2015, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, E.E.; Moss, P.; Haseler, M.; Maron, M. The influence of a variable fire regime on woodland structure and composition. Int. J. Wildland Fire 2015, 24, 59–69. [Google Scholar] [CrossRef]
- Oehl, F.; Laczko, E.; Bogenrieder, A.; Stahr, K.; Bösch, R.; van der Heijden, M.; Sieverding, E. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol. Biochem. 2010, 42, 724–738. [Google Scholar] [CrossRef]
- Makumbe, P.; Chikorowondo, G.; Dzamara, P.C.; Ndaimani, H.; Gandiwa, E. Effects of Fire Frequency on Woody Plant Composition and Functional Traits in a Wet Savanna Ecosystem. Int. J. Ecol. 2020, 2020, 1672306. [Google Scholar] [CrossRef]
- Steel, Z.L.; Safford, H.D.; Viers, J.H. The fire frequency-severity relationship and the legacy of fire suppression in California forests. Ecosphere 2015, 6, 1–23. [Google Scholar] [CrossRef]
- Trapnell, C.G. Ecological Results of Woodland and Burning Experiments in Northern Rhodisia. J. Ecol. 1959, 47, 129. [Google Scholar] [CrossRef]
- Bond, W.J.; Van Wilgen, B.W. Fire and Plants; Population and Community Biology Series 14; Chapman & Hall: London, UK, 1996; ISBN 9789401057721. [Google Scholar]
- Miller, J.E.D.; Safford, H.D. Are plant community responses to wildfire contingent upon historical disturbance regimes? Glob. Ecol. Biogeogr. 2020, 29, 1621–1633. [Google Scholar] [CrossRef]
- Jinga, P.; Ashley, M.V. Climate change threatens some miombo tree species of sub-Saharan Africa. Flora 2019, 257, 151421. [Google Scholar] [CrossRef]
- Jinga, P.; Palagi, J. Dry and wet miombo woodlands of south-central Africa respond differently to climate change. Environ. Monit. Assess. 2020, 192, 372. [Google Scholar] [CrossRef]
- Ribeiro, N.; Ruecker, G.; Govender, N.; Macandza, V.; Pais, A.; Machava, D.; Chauque, A.; Lisboa, S.N.; Bandeira, R. The influence of fire frequency on the structure and botanical composition of savanna ecosystems. Ecol. Evol. 2019, 9, 8253–8264. [Google Scholar] [CrossRef]
- Foster, C.N.; Barton, P.S.; MacGregor, C.I.; Catford, J.A.; Blanchard, W.; Lindenmayer, D.B. Effects of fire regime on plant species richness and composition differ among forest, woodland and heath vegetation. Appl. Veg. Sci. 2018, 21, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.D.; van Wilgen, B.W.; Burns, C.E.; Govender, N.; Potgieter, A.L.F.; Andelman, S.; Biggs, H.C.; Botha, J.; Trollope, W.S.W. Long-term effects of fire frequency and season on herbaceous vegetation in savannas of the Kruger National Park, South Africa. J. Plant Ecol. 2013, 6, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Governo de Moçambique. Inventário Florestal Nacional: Relatório Final; Ministério da Terra, Ambiente e Desenvolvimento Rural (MITADER): Maputo, Moçambique, 2018; 118p. [Google Scholar]
- Governo de Moçambique. Plano de Acção, Prevenção e Controlo às Queimadas Descontroladas (2008-2018); MICOA: Maputo, Moçambique, 2007; 53p. [Google Scholar]
- Ministério da Administração Estatal (Ed.) Perfil do Distrito de Muanza, Província de Sofala; MAE, Edição: Maputo, Moçambique, 2005; 60p. [Google Scholar]
- Instituto Nacional de Estatística (INE). IV Recenseamento Geral da População e Habitação; INE: Maputo, Moçambique, 2017. [Google Scholar]
- Marzoli, A. Inventário Florestal Nacional—Avaliação Integrada das Florestas de Moçambique (AIFM); DNTF, MINAG: Maputo, Mozambique, 2007. [Google Scholar]
- Giglio, L.; Boschetti, L.; Roy, D.P.; Humber, M.L.; Justice, C.O. The Collection 6 MODIS burned area mapping algorithm and product. Remote Sens. Environ. 2018, 217, 72–85. [Google Scholar] [CrossRef]
- Giglio, L.; Schroeder, W.; Hall, J.V.; Justice, C.O. MODIS Collection 6 Active Fire Product User ’s Guide Revision B. Dep. Geogr. Sci. Univ. Md. 2020, 9. [Google Scholar]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Rogan, J.; Franklin, J. Mapping Wildfire Burn Severity in Southern California Forests and Shrublands Using Enhanced Thematic Mapper Imagery. Geocarto Int. 2001, 16, 91–106. [Google Scholar] [CrossRef]
- Gandiwa, E.; Kativu, S. Influence of Fire Frequency on Colophospermum Mopane and Combretum Apiculatum Woodland Structure and Composition in Northern Gonarezhou National Park, Zimbabwe. Koedoe 2009, 51, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Achilleos, G.A. The Inverse Distance Weighted interpolation method and error propagation mechanism—Creating a DEM from an analogue topographical map. J. Spat. Sci. 2011, 2, 283–304. [Google Scholar] [CrossRef]
- Devkota, J.U. Statistical analysis of active fire remote sensing data: Examples from South Asia. Environ. Monit. Assess. 2021, 193, 608. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. In Biological Diversity: Frontiers in Measurement and Assessment; Oxford University Press: Oxford, UK, 2011; pp. 39–54. [Google Scholar]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Wiser, S.K.; Brotons, L. Using species combinations in indicator value analyses. Methods Ecol. Evol. 2012, 3, 973–982. [Google Scholar] [CrossRef]
- Curtis, J.T.; Mcintosh, R.P. The Interrelations of Certain Analytic and Synthetic Phytosociological Characters. Ecology 1950, 31, 434–455. Available online: http://Www.Jstor.Org/Stable/1931497 (accessed on 16 November 2021). [CrossRef]
- Guedes, B.S.; Sitoe, A.A.; Olsson, B.A. Allometric models for managing lowland miombo woodlands of the Beira corridor in Mozambique. Glob. Ecol. Conserv. 2018, 13, e00374. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Cauldwell, A.E.; Zieger, U. A reassessment of the fire-tolerance of some miombo woody species in the Central Province, Zambia. Afr. J. Ecol. 2000, 38, 138–146. [Google Scholar] [CrossRef]
- Peters, C.M. Sustainable Harvest of Non-Timber Plant Resources in Tropical Moist Forest: An Ecological Primer; Biodiversity Support Program c/o World Wildlife Fund: Washington, DC, USA, 1994; 45p. [Google Scholar]
- Balch, J.K.; Massad, T.J.; Brando, P.; Nepstad, D.C.; Curran, L.M. Effects of high-frequency understorey fires on woody plant regeneration in southeastern Amazonian forests. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120157. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.M.; Williams, M. How does fire intensity and frequency affect miombo woodland tree populations and biomass? Ecol. Appl. 2011, 21, 48–60. [Google Scholar] [CrossRef]
- Jew, E.K.K.; Dougill, A.J.; Sallu, S.M.; Connell, J.O.; Benton, T.G. Forest Ecology and Management Miombo Woodland under Threat: Consequences for Tree Diversity and Carbon Storage. For. Ecol. Manag. 2016, 361, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Olsson, L.; Barbosa, H.; Bhadwal, S.; Cowie, A.; Delusca, K.; Flores-Renteria, D.; Hermans, K.; Jobbagy, E.; Kurz, W.; Li, D.; et al. Land Degradation. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo, E., Buendia, V., Masson, D., Eds.; IPCC: Geneva, Switzerland, 2019; pp. 345–436. [Google Scholar]
- Thornley, J.H.M.; Cannell, M.G.R. Long-term effects of fire frequency on carbon storage and productivity of boreal forests: A modeling study. Tree Physiol. 2004, 24, 765–773. [Google Scholar] [CrossRef]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J.; Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.S.; Shugart, H.H.; Washington-Allen, R. The effects of fire and elephants on species composition and structure of the Niassa Reserve, northern Mozambique. For. Ecol. Manag. 2008, 255, 1626–1636. [Google Scholar] [CrossRef]
- Chirwa, P.W.; Akinnifesi, F.K. Ecology and Biology of Uapaca Kirkiana, Strychnos Cocculoides and Scelerocarya Birrea in Southern Africa. In Indigenous Fruit in the tropics: Domestication, Utilisation and Commercialisation; Akinnifesi, F.K., Leakey, R.R.B., Ajayi, O.C., Sileshi, G., Tchoundjeu, Z., Matakala, P., Kwesiga, F.R., Eds.; CABI: Egham, UK, 2008; p. 322. [Google Scholar]
- Chawafambira, A.; Sedibe, M.M.; Mpofu, A.; Achilonu, M. Uapaca kirkiana, an indigenous fruit tree in sub-Saharan Africa: A comprehensive review. Cogent Food Agric. 2020, 6, 1766735. [Google Scholar] [CrossRef]
- Richter, C.; Rejmánek, M.; Miller, J.E.D.; Welch, K.R.; Weeks, J.; Safford, H. The species diversity × fire severity relationship is hump-shaped in semiarid yellow pine and mixed conifer forests. Ecosphere 2019, 10, e02882. [Google Scholar] [CrossRef] [Green Version]
- Neeraja, U.V.; Rajendrakumar, S.; Saneesh, C.S.; Dyda, V.; Knight, T.M. Fire alters diversity, composition, and structure of dry tropical forests in the Eastern Ghats. Ecol. Evol. 2021, 11, 6593–6603. [Google Scholar] [CrossRef]
- Kadmon, R.; Benjamini, Y. Effects of Productivity and Disturbance on Species Richness: A Neutral Model. Am. Nat. 2006, 167, 939–946. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs High Diversity of Trees and Corals Is Maintained. Science 1978, 199, 1302–1310. Available online: http://www.jstor.org/stable/1745369 (accessed on 1 October 2022). [CrossRef] [Green Version]
- Tulloch, A.I.T.; Pichancourt, J.-B.; Gosper, C.R.; Sanders, A.; Chadès, I. Fire management strategies to maintain species population processes in a fragmented landscape of fire-interval extremes. Ecol. Appl. 2016, 26, 2175–2189. [Google Scholar] [CrossRef] [Green Version]
- Sheil, D.; Burslem, D.F. Disturbing hypotheses in tropical forests. Trends Ecol. Evol. 2003, 18, 18–26. [Google Scholar] [CrossRef]
- Bongers, F.; Poorter, L.; Hawthorne, W.D.; Sheil, D. The intermediate disturbance hypothesis applies to tropical forests, but disturbance contributes little to tree diversity. Ecol. Lett. 2009, 12, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, H.M.; Mallik, A.U. Predicting Plant Diversity Response to Disturbance: Applicability of the Intermediate Disturbance Hypothesis and Mass Ratio Hypothesis. Crit. Rev. Plant Sci. 2013, 32, 383–395. [Google Scholar] [CrossRef]
- Guitet, S.; Sabatier, D.; Brunaux, O.; Couteron, P.; Denis, T.; Freycon, V.; Gonzalez, S.; Hérault, B.; Jaouen, G.; Molino, J.-F.; et al. Disturbance Regimes Drive The Diversity of Regional Floristic Pools Across Guianan Rainforest Landscapes. Sci. Rep. 2018, 8, 3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNicol, I.M.; Ryan, C.M.; Williams, M. How resilient are African woodlands to disturbance from shifting cultivation? Ecol. Appl. 2015, 25, 2320–2336. [Google Scholar] [CrossRef]
- Hofiço, A.S.; Costa, E.A.; Fleig, F.D.; José, S.; Nanvonamuquitxo, A. Regulation of the Diametric Structure of the Miombo Woodland Using the De Liocourt Method in Mozambique Regulação Da Estrutura Diamétrica de Uma Floresta de Miombo Usando o Método De Liocourt Em Moçambique. Nativa 2018, 6, 407–414. [Google Scholar] [CrossRef]
- Maquia, I.; Ribeiro, N.S.; Silva, V.; Bessa, F.; Goulao, L.F.; Ribeiro, A.I. Genetic diversity of Brachystegia boehmii Taub. and Burkea africana Hook. f. across a fire gradient in Niassa National Reserve, northern Mozambique. Biochem. Syst. Ecol. 2013, 48, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Malimbwi, R.E.; Zahabu, E.; Monela, G.C.; Misana, S.; Jambiya, G.C.; Mchome, B. Charcoal Potential of Miombo Woodlands at Kitulangalo, Tanzania. J. Trop. For. Sci. 2005, 17, 197–210. Available online: http://www.jstor.org/stable/23616567 (accessed on 8 September 2022).
- CEAGRE & Winrock-International. Agentes e Causas de Desmatamento e Degradação Florestal em Moçambique; CEAGRE-UEM and Winrock International: Maputo, Moçambique, 2016; pp. 1–36. [Google Scholar]
- Gonçalves, F.M.P.; Revermann, R.; Gomes, A.L.; Aidar, M.P.M.; Finckh, M.; Juergens, N. Tree Species Diversity and Composition of Miombo Woodlands in South-Central Angola: A Chronosequence of Forest Recovery after Shifting Cultivation. Int. J. For. Res. 2017, 2017, 6202093. [Google Scholar] [CrossRef] [Green Version]
- Stahle, D.; Mushove, P.; Cleaveland, M.; Roig, F.; Haynes, G. Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. For. Ecol. Manag. 1999, 124, 217–229. [Google Scholar] [CrossRef]
- Geldenhuys, C.J. The effect of different regimes of annual burning on two woodland communities in kavango/die invloed van verskillende stelsels van jaarlikse brand op twee boomveldgemeenskappe in kavango. S. Afr. For. J. 1977, 103, 32–42. [Google Scholar] [CrossRef]
- Banda, T.; Schwartz, M.W.; Caro, T. Effects of fire on germination of Pterocarpus angolensis. For. Ecol. Manag. 2006, 233, 116–120. [Google Scholar] [CrossRef]
- Mojeremane, W.; Lumbile, A.U. A Review of Pterocarpus angolensis DC. (Mukwa) an Important and Threatened Timber Species of the Miombo Woodlands. Res. J. For. 2016, 10, 8–14. [Google Scholar] [CrossRef]
Figure 2.
Scheme of the cluster used for field data collection.

Figure 3.
Wood vegetation parameters under different fire strata (a) tree density and (b) above tree carbon stock. Means (± standard deviation) that do not share a letter are significantly different (Kruskal–Wallis test; α = 5%).
Figure 3.
Wood vegetation parameters under different fire strata (a) tree density and (b) above tree carbon stock. Means (± standard deviation) that do not share a letter are significantly different (Kruskal–Wallis test; α = 5%).

Figure 4.
Non-metric multidimensional scaling (NMDS) based on species composition for the fire regime strata, using the Bray–Curtis index.
Figure 4.
Non-metric multidimensional scaling (NMDS) based on species composition for the fire regime strata, using the Bray–Curtis index.

Figure 5.
Indicator Value of Species (IndVal in %) for the fire strata. The statistical significances (p < 0.05) of the indicator values are shown as boxes (species codes Table S4).
Figure 5.
Indicator Value of Species (IndVal in %) for the fire strata. The statistical significances (p < 0.05) of the indicator values are shown as boxes (species codes Table S4).

Figure 6.
Rarefaction curves according to the number of individuals (a) (95% confidence intervals); and respective sample coverage (b) under the different fire regimes. Hill numbers (effective number of species): richness (q = 0), Shannon’s Index (q = 1), and Simpson’s Index (q = 2).
Figure 6.
Rarefaction curves according to the number of individuals (a) (95% confidence intervals); and respective sample coverage (b) under the different fire regimes. Hill numbers (effective number of species): richness (q = 0), Shannon’s Index (q = 1), and Simpson’s Index (q = 2).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General characteristics of each fire stratum for the miombo woodlands of the study area.
| Stratum | Code | Description |
|---|---|---|
| High-frequency vs. High-intensity | HfHi | Sites usually occur in isolated mature forest fragments with limited human access and high seasonal fuel accumulation, consisting mainly of abundant grasses. |
| High-frequency vs. Low-intensity | HfLi | Combination of mature woodlands and multiple land uses, characterized by intense human activity such as charcoal production, agriculture, and considerable human presence and settlements. |
| Intermediate-frequency vs. Intermediate-intensity | IfIi | Transition areas between mature forest and fallow land. The presence of specific human interference (extraction of poles, firewood and wood) distinguishes this stratum from others. The local communities occasionally use these areas for charcoal and crop production. |
| Low-frequency vs. Low-intensity | LfLi | Abandoned agricultural areas (regrowth stands), with early recovery stages of miombo woodlands (10–15-year-old). |
| Low-frequency vs. High-intensity | LfHi | Similar to LfLi sites but with the natural succession process in a more advanced stage (20–25-year-old regrowth stands) and high seasonal fuel accumulation, consisting mainly of abundant grasses. |
Table 2.
Importance value index (IVI) values of the most 10 important species in each fire strata.
| Species | Family | IVI | ||||
|---|---|---|---|---|---|---|
| HfHi | HfLi | IfIi | LfHi | LfLi | ||
| Brachystegia boehmii | Fabaceae | 74.7 | 47.0 | 31.8 | 60.7 | 75.8 |
| Diplorhynchus condylocarpon | Apocynaceae | 26.5 | 15.8 | 11.3 | 10.8 | 14.2 |
| Pseudolachnostylis maprouneifolia | Phyllanthaceae | 22.2 | 19.8 | 18.2 | 28.0 | 21.5 |
| Pterocarpus angolensis | Fabaceae | 15.7 | 9.8 | 7.2 | 11.0 | 14.0 |
| Swartzia madagascariensis | Fabaceae | 13.6 | 20.3 | - | - | 15.2 |
| Pericopsis angolensis | Fabaceae | 12.6 | - | 14.3 | 20.3 | 9.4 |
| Strychnos madagascariensis | Strychnaceae | 12.3 | 12.1 | 16.2 | 13.2 | - |
| Terminalia brachystemma | Combretaceae | 11.0 | - | - | 9.3 | - |
| Lannea discolor | Anacardiaceae | 10.7 | - | - | - | - |
| Hymenocardia acida | Phyllanthaceae | 9.4 | - | 17.2 | - | 7.1 |
| Brachystegia spiciformis | Fabaceae | - | 8.5 | - | - | 9.3 |
| Albizia forbesii | Fabaceae | - | 15.8 | - | - | - |
| Xeroderris stuhlmannii | Fabaceae | - | 14.4 | - | - | - |
| Bauhinia petersiana | Fabaceae | - | 9.8 | - | - | - |
| Parinari curatellifolia | Chrysobalanaceae | - | - | 34.4 | - | - |
| Albizia glaberrima | Fabaceae | - | - | 15.4 | - | - |
| Julbernardia globiflora | Fabaceae | - | - | 7.2 | - | 20.3 |
| Uapaca kirkiana | Phyllanthaceae | - | - | - | 14.2 | - |
| Piliostigma thonningii | Fabaceae | - | - | - | 8.6 | - |
| Pterocarpus rotundifolius | Fabaceae | - | - | - | 8.0 | - |
| Crossopteryx febrifuga | Rubiaceae | - | - | - | - | 7.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Buramuge, V.A.; Ribeiro, N.S.; Olsson, L.; Bandeira, R.R.; Lisboa, S.N. Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire 2023, 6, 26. https://doi.org/10.3390/fire6010026
AMA Style
Buramuge VA, Ribeiro NS, Olsson L, Bandeira RR, Lisboa SN. Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire. 2023; 6(1):26. https://doi.org/10.3390/fire6010026
Chicago/Turabian StyleBuramuge, Victorino Américo, Natasha Sofia Ribeiro, Lennart Olsson, Romana Rombe Bandeira, and Sá Nogueira Lisboa. 2023. "Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique" Fire 6, no. 1: 26. https://doi.org/10.3390/fire6010026