
Sustainable Management of Diseases in Horticulture: Conventional and New Options

Research Centre for Olive, Fruit and Citrus Crops, Council for Agricultural Research and Economics (CREA), Via di Fioranello, 52, 00134 Roma, Italy
Horticulturae 2022, 8(6), 517; https://doi.org/10.3390/horticulturae8060517
Submission received: 11 April 2022
/
Revised: 3 June 2022
/
Accepted: 8 June 2022
/
Published: 13 June 2022
(This article belongs to the Special Issue Sustainable Control Strategies of Plant Pathogens in Horticulture)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:To reduce the impact of chemical pesticides on the environment, there are relevant efforts to enhance the possibility of controlling plant diseases using environmentally friendly biocontrol agents or natural products that show pathogen control capacity. The European Union, FAO, and the United Nations largely promote and finance projects and programs in order to introduce crop protection principles that can attain sustainable agriculture. Preventive measures related to the choice of cultivars, soil fertility, integrated pest management (IPM), and organic farming strategies are still the basis for obtaining satisfactory crop yields and reducing classical pesticide utilisation through the application of commercially available and ecofriendly control agents. Effective pathogen detection at borders to avoid quarantine pathogens is mandatory to reduce the risk of future epidemics. New technical support for the development of sustainable pathogen control is currently being provided by forecasting models, precision farming, nanotechnology, and endotherapy. New biocontrol agents and natural products, disease management through plant nutrition, systemic resistance inducers, and gene-silencing technology will provide solutions for obtaining satisfactory disease control in horticulture. The “multi-stakeholder partnership” strategy can promote the implementation of sustainable crop protection.
1. Introduction
The concepts that illustrate sustainable agriculture have been posed and defined decades ago and can be summarised by the principles and approaches described by F.A.O. “Building a common vision for sustainable food and agriculture” (https://www.fao.org/3/i3940e/i3940e.pdf, accessed on 22 May 2022) as “an integrated system of plant and animal production practices having a site-specific application that over the long-term will: (a) satisfy human food and fiber needs; (b) enhance environmental quality and the natural resources; (c) make the most efficient use of nonrenewable resources and on-farm resources and integrate natural biological cycles and control; (d) sustain the economic viability of farm operations; (e) enhance the quality of life for farmers and society as a whole”.
Considering that the complete achievement of all such goals still requires a relevant effort [1], the success of sustainable agriculture mainly depends on the acceptance of these principles by the farmers, which should actively identify strategies for maintaining, enhancing, and developing their on-site resources (i.e., soil, water, air, biodiversity, and landscape) for future generations [2]. The successful application of such goals can be assessed by indicators that measure the percentage of the agricultural area which satisfies the specified criteria of sustainability regarding water, soil, and biodiversity, and achieving a specific level of productivity [3]. However, the need for a continuously widespread application of sustainability criteria in agriculture with less impact on the environment is also necessary in a world where food demand is increasing.
For the European Union, sustainable agriculture, through the “Farm to Fork” strategy, is one of the main objectives of the European Green Deal (Annex to the European Green Deal, 2019). The aim of agriculture before the year 2030 should be: (a) at least 25% of agriculture in Europe being organic; (b) reduction by 50% of chemical pesticide utilisation; (c) reduction by 50% of more hazardous pesticide utilisation; (d) reduction by 20% of fertiliser utilisation; (e) reduction of soil nutrient losses by at least 50%. In addition, to diminish the utilisation of copper in agriculture, the executive regulation 2018/1981 of the European Commission reduced the maximum limit of usable copper to 4 kg per hectare, and a maximum of 28 kg per hectare in seven years to minimise the potential accumulation of copper in soil and the exposure of non-target organisms. Similarly, the United Nations 2030 agenda promotes sustainability in agriculture through Sustainable Development Goals (SDG) (https://sdgs.un.org/goals, accessed on 22 May 2022). Within this scenario, there are already examples of communities that, upon a referendum, decided to ban the use of chemical pesticides to protect the local environment and obtain pesticide-free food [4].
It should be noted that the use of traditional pesticides showed a 2% decrease per year owing to the application of regulatory restriction laws, compared to the 15% increase per year in favour of biopesticide utilisation [5]. Within this context, the future of agriculture will be based on environmentally friendly agronomical techniques that, at the same time, can assure a profit to the farmer and the sustainability of the farm itself [6].
The success of obtaining satisfactory pathogen management according to sustainable agriculture principles requires parallel actions to prevent the spread of phytopathogens. From this perspective, effective quarantine measures are necessary to avoid the introduction of destructive plant pathogens into new areas of cultivation. Currently, this aspect is particularly relevant because of the extensive global circulation of plant materials and climate change [7]. Modern diagnostic tools should be implemented at the points of plant material circulation (i.e., airports and ports) and the local entry points (i.e., regional phytosanitary services) [8]. Local quarantine agencies can be assisted by climate-matching tools and geographical information systems that can predict the possibility of pathogen spread in a new area [9].
Many reviews have been published on the different aspects of sustainable agriculture, including basic knowledge on the control of phytopathogens [10,11,12,13,14]. The principles that rule out agro-ecology and organic farming are not discussed. This review attempts to provide a broad overview of sustainable agriculture and integrated pest management (IPM) principles applied to achieve pesticide reduction, with a focus on disease management under the regulatory framework of the European Union. There is a focus on the main strategies based on the utilisation of well-known and new biocontrol agents and products or compounds with a low impact on the environment that are already developed or undergoing achievements regarding the control of some diseases of woody and herbaceous crops. New technologies to augment the efficacy of disease control in sustainable agriculture are also presented and discussed.
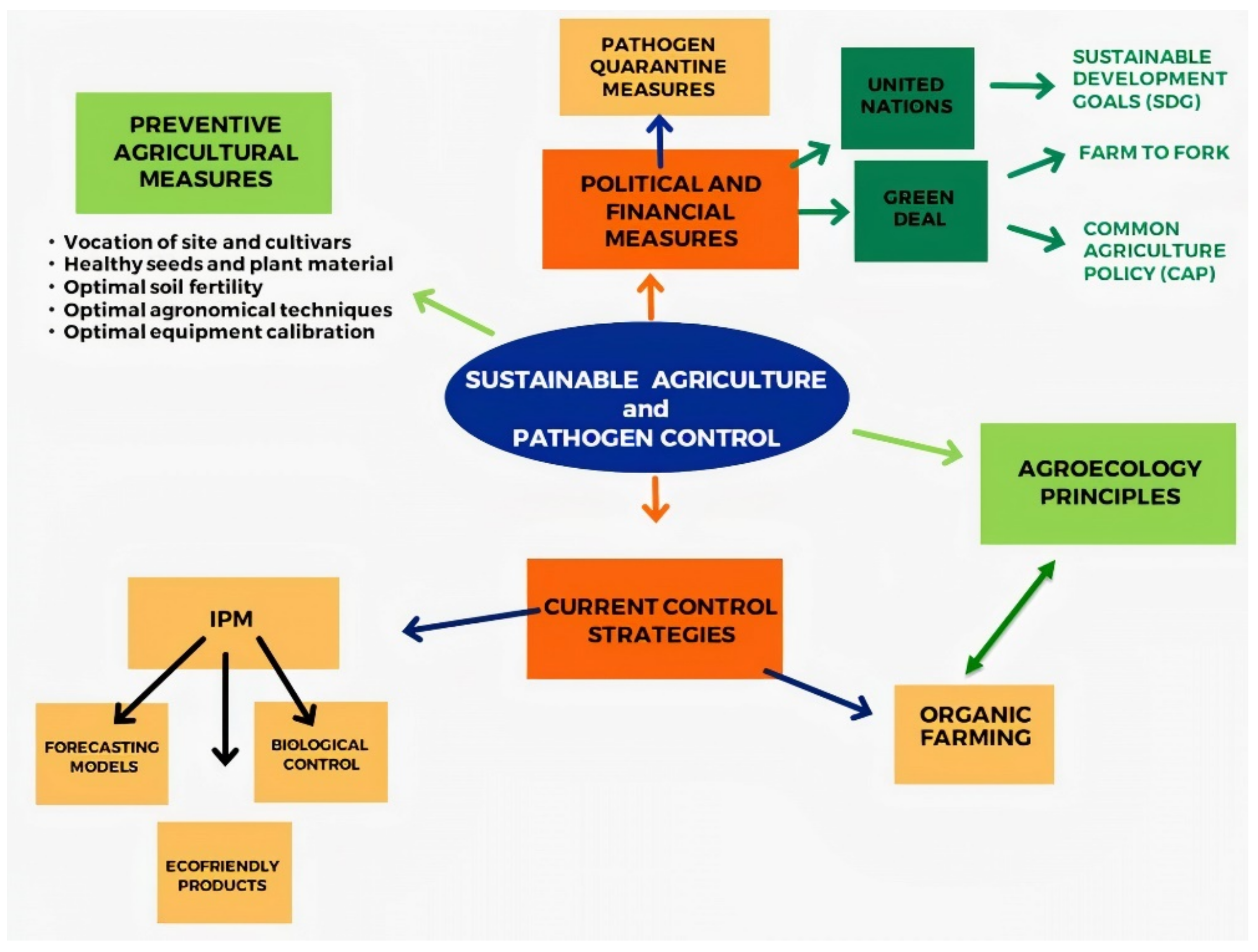
A synoptic panel of current control strategies in relation to sustainability principles and policies is shown in Figure 1.
2. The Basis for a Sustainable Disease Control: The Preventive Measures
2.1. Suitability and Selection of the Site and Cultivars
In addition to the economic aspects and infrastructural facilities, the climatic factors characterising an area must be considered for the choice of the crop to be cultivated. At present, this issue is relevant because of climatic changes that affect most areas of the world. Climate change can result in the adoption of different pathogen control strategies and agronomical techniques, owing to the possible adaptation of new pathogens to the new climatic scenario. For example, many areas with a Mediterranean climate that are traditionally free from freezing events either in winter or early spring have recently faced relevant frost damage during such periods, which seriously threatens the economic profit of crops [15,16]. In addition, for other areas such as Central and Eastern Asia, Central–North America, Northern India, Australia, and the Mediterranean Basin, the occurrence of “hot spots” (i.e., temperature > 40 °C for many consecutive days, accompanied by the absence of rainfall) during summer pose a risk to wheat cultivation [17] and can cause severe damage to heat-tolerant crops such as olive [18]. Edaphic (i.e., soil fertility, texture, and porosity) and biotic (i.e., occurrence of bees, pests, and pathogens) factors must also be considered to avoid future problems due to climate change.
In addition to area suitability, the right choice of cultivars is another basic element that can allow the success of the crop according to sustainability criteria. For woody species, the choice is of basic importance and, according to soil characteristics, should also consider the choice of rootstock. The right choice is even more critical, particularly when the crop reaches a new cultivation area [19]. The cultivar choice for herbaceous crops is also important in the context of climate change, as shown by an extensive survey performed in Germany with cereal producers. Farmers judged eco-stability, grain yield performance, and steadiness as being the most important cultivar requirements [20].
2.2. Healthy Seeds and Plant Material
The healthy phytosanitary status of seeds, tubers, plantlets, potted plants, and propagative material is a fundamental prerequisite for initiating cultural cycles. At present, this aspect is particularly important considering the extensive global circulation of these commodities. In recent years, the relevant increase in global plant circulation regarding the agricultural and forest trade has dramatically increased the possibility of pests and pathogens to rapidly reach new countries and, consequently, to colonise and infect new crops and the same crops cultivated on another continent [21]. Once introduced in a new area, phytopathogens can become part of the new environment(s) depending on a series of factors, such as the number of introduction events, the transmission rate of the pathogen, the density and spatial variation of the susceptible host, the favourability of the climatic conditions, the synchronicity between host susceptibility, and the pathogen life cycle [22]. An efficient surveillance system at the border should be developed in each country to rapidly intercept new threats before they can be established in a new territory. This issue is particularly important for countries that have not yet developed a phytosanitary regulation system based on quarantine principles. In contrast, seed companies and plant nurseries should efficiently implement all preventive measures that can reduce the colonisation of plant material (i.e., effective pathogen control strategies during plant growth and disinfection of plant material before shipping). In addition, farmers should carefully monitor crops, particularly during the first phases of growth, to observe and eliminate potential diseases.
2.3. Optimal Soil Fertility and Agronomical Techniques
One of the pillars of the European Common Agricultural Policy (CAP) is the maintenance and enhancement of soil fertility; correct soil management is one of the fundamental prerequisites for sustainability in agriculture [23]. According to agroecological principles, some effective practices can be applied to herbaceous and woody crops to maintain and augment soil fertility. Crop rotation with leguminous species and the planting of cover crops between tree rows are methods that can ensure, over a long-term period, the maintenance of natural soil fertility [24,25]. Crop rotation can also result in a better control of some soil-borne diseases, such as in potatoes affected by Rhizoctonia solani and Streptomyces scabies [26]. In addition, the application of plant growth-promoting rhizobacteria (PGPR), biofertilisers, composts, mycorrhiza, biochars, and humic and fulvic acids can augment nutrient acquisition and assimilation, improve soil texture and plant growth, and induce systemic resistance to biotic and abiotic stresses [26,27,28,29,30,31,32]. For example, the distribution to the soil of a biofertiliser that contained a mixed fungal and bacterial microflora induced conferred protection against Fusarium wilt of banana caused by Fusarium oxysporum f. sp. cubense after three years [33]. Paecilomyces variotii, a fungus obtained from agro-industrial compost, showed efficacy in the control of Fusarium wilt of melon caused by Fusarium oxysporum f. sp. melonis [34]. However, care should be given to the correct choice of compounds released into the soil to increase their overall fertility. In some circumstances, organic matter, particularly animal manure, can release antibiotics that can perturb native microflora, causing adverse effects on the crop [35]. It should be noted that balanced crop nutrition is an essential component of any integrative program for crop protection [36].
Given that the application of compounds for crop protection aims to deposit the highest amount of the active ingredient on the target plant part (i.e., buds, leaves, and canopy) in which the pathogen resides, an effective and desirable reduction of the spread of any compound in the environment can be achieved by the appropriate calibration of the sprayers. At present, it is possible to adjust the sprayer nozzles to achieve the intended target (i.e., the plant part that shows symptoms of disease), which also reduces water utilisation [37]. Soil solarisation is a well-known technique that, when properly applied, can effectively control important soil-borne pathogens. However, the utilisation of plastic covers poses a relevant concern for their subsequent removal and disposal [38]. Organic farming largely benefits from such preventive measures to obtain effective pathogen control, particularly soilborne pathogens [39].
3. Sustainable Agriculture and Pathogen Control
3.1. The Basis for an Effective Sustainable Pathogen Control
Knowledge of the genomic structure, virulence factors, and epidemiology of pathogens is the basis for developing fine-tuned strategies for the effective control of biotic diseases in crops. Selected biocontrol agents or compounds with potential curative effects should be tested against different strains of pathogens that represent the entire population structure. In addition, the disease cycle of the target pathogen should be fully understood to precisely apply the biocontrol agent/compound before and during plant colonisation or the internal multiplication of the microorganism. Some examples of the basic studies that link crop and pathogen epidemiology to the selection of biocontrol agents and fine-tune the spread of active ingredients for disease control are as follows [40,41,42,43,44,45,46]. Knowledge of the pathogen cycle of diseases is also the basis for the development and implementation of disease forecasting [47,48]. Moreover, the sustainability of modern pathogen control should be considered in addition to crop productivity, the ecological function of the crop, and the social acceptance of the strategy [49]. A report that concerns either the effectiveness or the social impact of different strategies to control fire blight of apple, caused by Erwinia amylovora, in Switzerland has been prepared. A thorough investigation performed by interviewing experts and a literature data search revealed that biological control performed with Aureobasidium pullulans is either effective or widely accepted in rural areas because of its feasibility, durability, low impact on animals, biodiversity, soil and water habitation, low cost, and acceptance by consumers [50]. In this case, the majority of the inhabitants of an area are aware of the importance and efficacy of sustainable agriculture for the maintenance and improvement of their lifestyle and environmental safety.
3.2. Current Control Strategies
Integrated pest management (IPM) is the current strategy that allows for the effective control of many plant pathogens in many cases. According to the European Union Framework Directive on the sustainable use of pesticides (Directive 2009/128/EC), IPM “means careful consideration of all available plant protection methods and subsequent integration of appropriate measures that discourage the development of populations of harmful organisms and keep the use of plant protection products and other forms of intervention to levels that are economically and ecologically justified and reduce or minimise risks to human health and the environment. IPM emphasises the growth of a healthy crop with the least possible disruption to agro-ecosystems and encourages natural pest control mechanisms”. In addition, the FAO IPM programme involves three large areas located in Asia, the Near East, and West Africa to improve farming skills, raise the awareness of smallholder farmers of the risks posed by traditional agrochemicals, and promote sustainable agriculture (https://www.fao.org/agriculture/crops/core-themes/theme/pests/ipm/en/, accessed on 22 May 2022).
However, given the diversity and complexity of agricultural scenarios, IPM can differ significantly among countries, and each crop of a definite area of cultivation should apply the IPM criteria according to the local reality and a holistic approach; thus, the combination of control tactics into a planned strategy can provide more effective and sustainable results than the single-tactic approach [51,52]. The development and utilisation of ad hoc web-based platforms illustrating control thresholds, cultural practices that can influence disease attack, pathogen virulence, and fungicide efficacy can help farmers, advisors, and researchers to better plan the control strategy according to a real-time assessment of the environmental conditions of the area [53].
The application of the IPM strategy on a large scale would benefit from ad hoc studies that provide updated information on the current control strategies applied to any crop in an area in order to identify the lack of knowledge in the field, to be resolved through future studies. These systematic maps of knowledge have proven useful in applying IPM to arable crops (i.e., wheat, barley, oat, potato, sugar beet, and oilseed rape) cultivated in large areas in Sweden [54]. A refined IPM strategy which combines all the validated methods for monitoring and reducing the impact of the diseases would allow researchers to control apple scab and apple powdery mildew, caused by Venturia inaequalis and Podosphaera leucotricha, respectively. This strategy includes disease monitoring and forecasting, ecofriendly fungicides, adequate orchard sanitation, biological control, and insect control through mating disruption. It is comparable with the results obtained by conventional pest management methods [55]. IPM is also the basis for a transition from a chemical to a biological control strategy in Canada regarding greenhouse vegetable crops [56].
In addition, IPM strategies largely benefit from the cultivation of resistant/tolerant cultivars, as observed for the more effective application of biocontrol agents such as Bacillus mycoides to a sugar beet cultivar that is more tolerant to Cercospora beticola, or Bacillus subtilis towards chickpeas infected by Fusarium oxysporum f. sp. ciceris [57]. In the Netherlands and Ireland, the utilisation of novel potato cultivars resistant to late blight, caused by Phytophthora infestans, in combination with real-time pathogen population monitoring and checking of its genetic structure allowed for a reduction of 80–90% of fungicide use (https://www.wur.nl/en/newsarticle/more-sustainable-potato-production-through-extended-ipm-for-late-blight.htm, accessed on 22 May 2022). The protection of crops starting from the seed is another relevant option for better management of diseases. Trichoderma gamsii, applied to maize kernels, has been proven effective in reducing pink ear rot caused by Fusarium verticillioides and root infection [58].
Despite the higher incidence of pathogens in the crop, the application of organic farming principles for some years can also support, in many circumstances, the effective biological control of pathogens [59]. The observed increase in ecological intensifications of the agro-ecosystem promotes a higher occurrence of beneficial microorganisms in the crop [59]. The synergism between IPM strategies and organic farming principles in relation to pathogen control could provide benefits for improving environmental quality, farm economic viability, and soil and human health [60].
At present, the success of IPM and organic farming in relation to pathogen control is based on three research sectors that are closely related to each other: disease forecasting models, biological control, and environmentally friendly natural products or compounds.
3.2.1. Disease-Forecasting Models
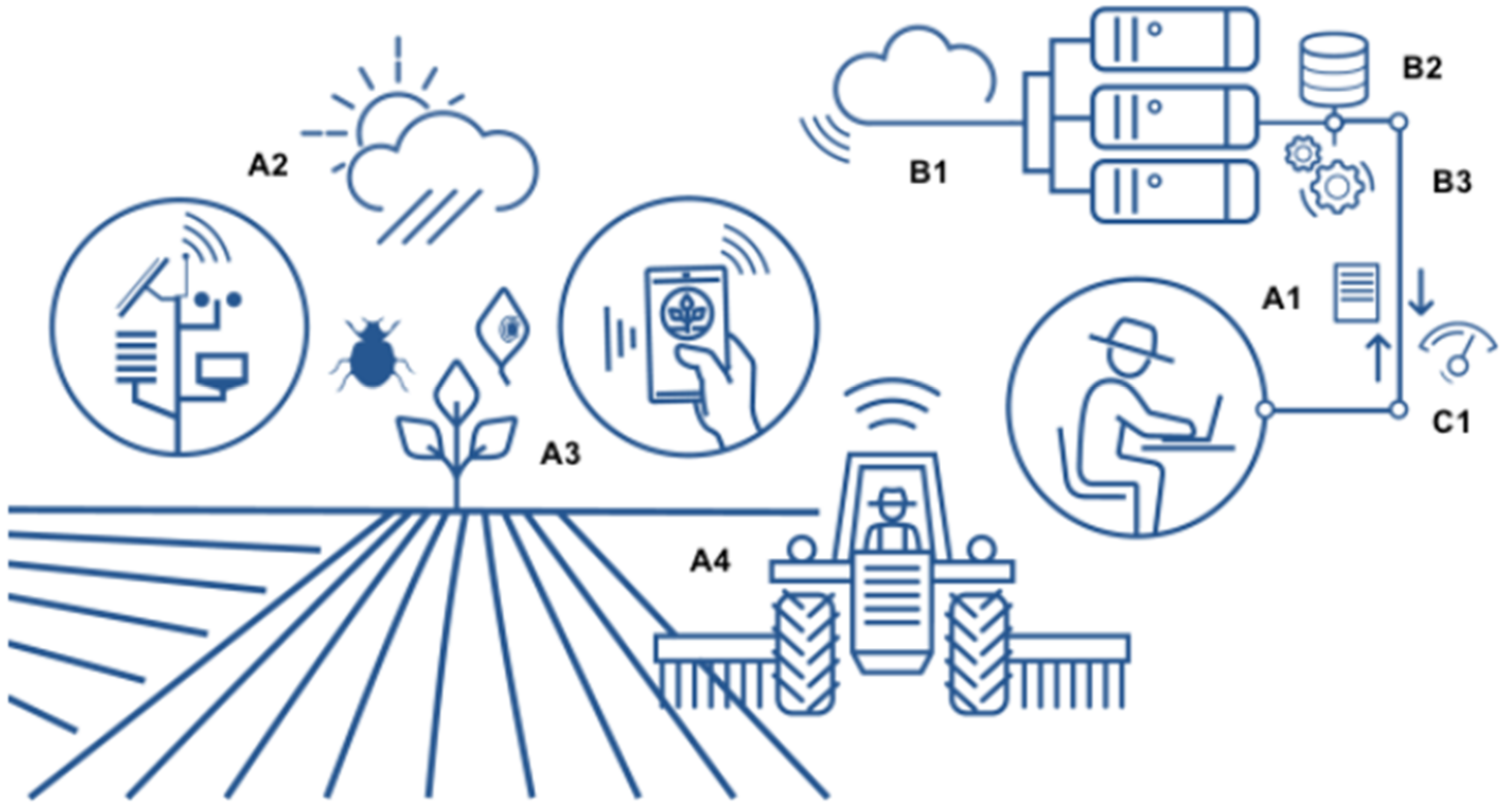
Disease forecasting is based on mechanical models designed with the input of climatic data and the pathogen cycle of disease to alert the grower on whether, when, and how to apply an agrochemical or a biocontrol agent to protect crops. Such models are dynamic because they analyse the changes in the components of an epidemic over time according to external variables (i.e., climatic data, pathogen multiplication, and plant growth stage in relation to disease development) [47] (Figure 2). An effective example of a forecasting model that allows a relevant reduction in pesticide distribution in the environment is vite.net® [61]. Based on a decision support system that calculates vineyard parameters (i.e., air, soil, plant, pathogen, and disease development) and a web-based tool that analyses such data, vite.net® provides information for Plasmopara viticola management in the vineyard. The system is flexible and can be tailored to a single vineyard or an area characterised by high similarity. This tool was largely utilised by grape growers on more than 17,000 h in 2017 in Italy, Spain, Portugal, Greece, Romania, and the United Kingdom and allowed for a reduction of approximately 50% in pesticide utilisation [47,61].
Another effective and used disease-forecasting model is BrassicaspotTM that is applied to manage Albugo candida, a causal agent of white blister of Brassica crops (i.e., radish and broccoli) in Australia [62]. The application of the model allowed for a reduction of more than 80% of the disease and reduced the number of pesticide sprays from fourteen to one or two per year. In addition, the introduction of a resistant cultivar and the simple change in the time of irrigation from 2000 h to 4000 h also decreased disease incidence [62]. Similarly, in Florida, the Strawberry Advisory System (SAS) based on local weather data allows growers to reduce the use of chemical sprays for controlling anthracnose, caused by Colletotrichum spp., and grey mould, caused by Botrytis cinerea [63], by 50%. In South Korea, the EPIRICE model was developed to assess the daily risk for the occurrence of rice blast caused by Magnaporthe oryzae [48]. The model utilises some climatic data linked to fungus multiplication, such as air relative humidity, temperature, and precipitation, and can be used to predict the risk of disease at an early stage [48].
3.2.2. Biological Control
Biological control agents for plant diseases are defined as naturally occurring microorganisms capable of suppressing the growth and proliferation of a target pathogen by different mechanisms of action (i.e., competition for space and nutrients, antibiosis, predation, induced host resistance, and lytic enzymes). In addition to living microorganisms, they sometimes utilise metabolite(s) that can be sprayed directly onto crops [64]. Beneficial microorganisms are registered as plant protection products, and they are usually applied to crops at a high density once or several times during the growing season. In the United States and Canada, government agencies are responsible for confirming the biosafety of the biocontrol agents (i.e., the Environmental Protection Agency (EPA) in the United States, and the Pest Management Regulatory Agency (ARLA) in Canada). In Europe, according to Regulation 1107/2009 for plant protection products, the authorisation for commercialising biocontrol agents is obtained through some related steps: (a) the bioactive microorganism should be approved at the European level by the European Food Safety Authority (EFSA) according to the physiochemical properties of the substance, its risk profile for human health, and its risk profile for the environment; (b) formulated products should be authorised at the member state level; (c) further scrutiny with regard to organic agriculture requirements. In addition, the Directorate General for Health and Food Safety (DG SANTE), the Standing Committee on Plants, Animal, Food, and Feed (PAFF Committee), and the Rapporteur Member State are involved in the decision according to Directive 91/414, which states that any active substance should be included in an approved EU list (Annex 1), and its further application must be authorised by member states. These procedures take a long time, thus creating an overall slowness for final approval [65].
The causal agents of plant diseases can have either a worldwide occurrence in specific crops or local distribution. For the first case, the application of the Nagoya protocol of October 2014, for the “Access and the Fair and Equitable Benefit-sharing of Genetic Resources” that restricts the international exchange of biological material, can result in a limitation on the circulation and use of biological control agents that are selected in a different geographical area [66]. Consequently, given the increase in organic food demand and the current Green Deal policy, the selection of native local biocontrol agents is of paramount importance [11]. It should be noted that this selection largely depends on the specific pathosystem under study. Two approaches illustrate how it is possible to proceed: (a) selection through consecutive screenings for testing the effectiveness of the biocontrol agent (i.e., in vitro assays for antibiosis, lytic enzymes, and antimicrobial metabolites, in planta assays for colonisation, control performance, and induced resistance); (b) selection through the assistance of genetic/genomic studies (i.e., the use of genetic markers for finding single biocontrol traits, genome-wide DNA markers for selecting complex traits) [11,67]. The durability of biocontrol agents is another important prerequisite for their long-standing efficacy. Indeed, there are documented reports of a significant reduction in the control effectiveness of Botrytis cinerea in Astilbe hybrids, as shown by Pseudomonas chlororaphis after eight treatments [68].
In some circumstances, natural selection yields biocontrol agents that are capable of displaying long-term positive effects on some diseases, such as for soybean root rot caused by Fusarium spp., Pythium spp., and Rhizoctonia solani in Northeast China. In this case, a naturally occurring suppressive soil was analysed, and Trichoderma harzianum, Pochonia clamydosporia, Paecilomyces lilacinus, and Pseudomonas fluorescens strains have been found to inhibit the fungal pathogens of soybean roots [69]. Another well-known example is the natural occurrence of mycoviruses of the family Hypoviridae, which infect Cryphonectria parasitica, the causal agent of chestnut blight. Upon infection, the virus incites hypovirulence in the fungus by reducing its parasitic growth and sporulation capacity. The virus can be isolated from chestnut cankers and utilised as a biocontrol agent to cure trees by inhibiting further canker development. In addition, the virus is capable of spreading naturally in the forest and reaching other infected trees [70].
Fungi, bacteria, and yeast are the most widely used biocontrol agents. The following species are among the most versatile: Trichoderma harzianum, Trichoderma viride, Bacillus subtilis, and Pseudomonas fluorescens. All of them, indeed, have shown control activity towards some common fungal pathogens such as Botrytis cinerea, Monilinia fructicola, Plasmopara viticola, Puccinia graminis, and Erisiphe spp. [71]. Rhizobium (Agrobacterium) radiobacter strain K84 is among the most known biocontrol agents used for many years to control crown gall caused by Agrobacterium tumefaciens [72]. A stand-alone effective biocontrol agent has been also selected for apple scab caused by Venturia inaequalis, namely Cladosporium cladosporioides H39, that, over a wide range of environment, showed a high control level and appears effective even when applied after some days from the infection event [73]. Similarly, Bacillus amyloliquefaciens strains are capable of effectively controlling Fusarium equiseti in broad bean cultivation [74]. In some circumstances, a single biocontrol agent is capable of effectively reducing two diseases caused by distantly related microorganisms, as was seen for Paecilomyces variotii, which showed control activity towards both the bacterium Xanthomonas vesicatoria, the causal agent of bacterial spot of tomato, and Fusarium oxysporum f. sp. melonis, the causal agent of Fusarium wilt of melon [34].
Trichoderma spp. are among the most studied biocontrol agents in agriculture. Through different metabolic pathways, the induction of systemic resistance to the plant, multiple adaptive mechanisms, antimicrobial molecules, and antagonistic behaviour, Trichoderma strains can either promote plant growth or act as effective biocontrol agents against fungal species under numerous agricultural conditions, including in greenhouses and nurseries [75,76]. These strains account for the greatest proportion of fungal biocontrol agents against the phytopathogenic microorganisms investigated, and many commercial formulations that contain a single Trichoderma strain or a mixture of different Trichoderma strains are available [75,77]. A series of common and widespread phytopathogenic fungi, such as Rhizoctonia solani, Fusarium oxysporum, Botrytis cinerea, Pythium spp., Sclerotinia spp., Verticillium spp., Phytophthora spp., and Alternaria spp. can be controlled by generalist Trichoderma spp. strains [75,78]. Other fungal genera that also show antagonistic activity toward phytopathogenic fungi are Alternaria, Aspergillus, Penicillium, Pichia, Candida, Talaromyces, and nonpathogenic Fusarium, Pythium, and Verticillium [79]. Pichia anomala is effective in controlling postharvest crown rot of banana caused by Colletotrichum musae, Fusarium verticillioides, and Lasidiodiplodia theobromae [80]. In addition, Ampelomyces quisqualis is commercially available for the preventive control of powdery mildew fungi in different crops, such as eggplant, cucumber, tomato, and strawberry.
Pseudomonad strains provide a very large amount of potential biocontrol agents that can be found within the following species and species complexes: Pseudomonas fluorescens, P. chlororaphis, P. putida, P. syringae, P. aureofaciens, P. protegens, P. mandelii, P. corrugata, P. koreensis, and P. gessardii [81]. However, very few commercially available products are currently available: P. fluorescens for Erwinia amylovora; nonpathogenic Pseudomonas syringae for postharvest disease of fruits, potato, and sweet potato; Pseudomonas chlororaphis for fungal diseases of ornamental crops and turf grass; Pseudomonas aureofaciens for lawn and grass management against soil-borne fungi. Genetic instability and poor shelf life are among the main causes of the limited registration of pseudomonads as biocontrol agents [81]. Bacillus species have a higher shelf life due to their possibility to form endospores, and have many potential biocontrol agents, stemming from their ample antagonism mechanisms (i.e., antibiosis, enzymes, lipopeptides, competition for space, and nutrients), are also present in this genus, with Bacillus subtilis, B. amyloliquefaciens, and B. polymixa being the richest in providing biocontrol effectiveness [82].
Among the yeasts, five species are most widely used as biocontrol agents, namely Candida oleophila, Metschnikowia fructicola, Cryptococcus albidus, Saccharomyces cerevisiae, and the yeast-like fungus Aureobasidium pullulans. The low production of toxic secondary metabolites by yeast is an important prerequisite that raises fewer biosafety concerns and increases their possibility of utilisation [83]. Candida oleophila is utilised as a postharvest biocontrol agent for banana, apple, and citrus, towards Colletotrichum musae, Botrytis cinerea, and Penicillium spp., whereas Aureobasidium pullulans is utilised as a biocontrol agent towards fire blight of apple and pear, caused by Erwinia amylovora, and for apple postharvest diseases [84]. Moreover, a satisfactory control has been also achieved in strawberry cultivated in greenhouses towards Phytophthtora cactorum and Botrytis cinerea, the causal agents of crown and root rot and grey mould, respectively [85].
In some cases, a combination of more biocontrol agents provides better control than the application of a single agent. The sprays of Bacillus mycoides and the yeast Pichia guillermondii on strawberry leaves provided a better performance for controlling Botrytis cinerea than the application of a single biocontrol agent [86]. In addition, to induce stimuli for the plant growth, plant growth-promoting rhizobacteria (i.e., Bacillus subtilis and Curtobacterium flaccumfaciens) can also act as effective biocontrol agents as the case of a multiple effectiveness against three cucumber pathogens, namely Colletotrichum orbiculare, Pseudomonas syringae pv. Lachrymans, and Erwinia tracheiphila [87]. However, the compatibility between different control agents should always be assessed because there are also cases of reduced control activity when two agents are applied without a prior compatibility assessment [88,89]. In the case of proven compatibility, the two biocontrol agents can provide satisfactory disease control activity through a multitrophic approach [90]. The addition of specific substances that promote the growth of biocontrol agents on the plant’s surface can also enhance the performance of the biocontrol agents, as in the case of lactic acid, which is used as a biocontrol agent for Erwinia amylovora [91].
3.2.3. Natural Products and Compounds
A natural product with a potential use for controlling plant diseases can be defined as a physiologically active chemical that is synthesised by plants, microorganisms, or animals. These products can act as antimicrobials or inducers of systemic resistance, are usually easily biodegradable, and do not persist in the environment [92]. Some of them can act as templates for chemical pesticides (i.e., synthetic analogues), such as the fungicide strobilurin, which was named with reference to Strobilurus tenacellus, a wood-rotting mushroom. Chitosan and its derivatives, alkaloids, flavonoids, terpenes, proteins, and phenolic compounds are the most widely studied and used dried materials [93]. These products are the result of their coevolution with the biotic environment; thus, many of them are defence compounds toward other organisms and can possess potent bioactivities and selectivity [94]. Within this category could be included also antibiotics. However, in many countries, these products are banned for their utilisation in agriculture as protectants because of the possibility of transmission of genetic traits from bacterial phytopathogens to microbes of human and animal importance, which could confer resistance to a single or multiple antibiotics [95,96]. However, these are not discussed in this paper.
Harpin, a proteinaceous elicitor of the self-protective hypersensitivity reaction in plants against pathogens, obtained from the pathogen Erwinia amylovora [97], was among the first natural products to be commercially utilised in plant protection. Protection obtained through harpin was observed for harvested apples infected by blue mould, which is caused by Penicillium expansum [98]. Currently, it is included among the biostimulants. Another well-known natural fungicide was obtained from Reynoutria sachalinensis to control powdery mildew of cucumber and tomato in glasshouse cultivations, as well as downy mildew of grapevine [99,100]. The extract induces plant defence responses such as callose papillae and an increase in salicylic acid and caffeic acid [101]. Chitosan is a derivative polymer of chitin, the primary component of the cell wall of fungi and the exoskeleton of insects and crustaceans, and is usually extracted from shrimp shells, mud crabs, and fungi for utilisation in medical and chemical science, as well as in agriculture. Chitosan-based compounds are traditionally utilised postharvest to protect fruit decay [102]. Due to the very high economic losses in crops caused by phytopathogens that show pesticide resistance [103], the discovery, validation, and registration of new natural products that are characterised by different and novel modes of action are required, and DNA sequence technologies can help in identifying gene(s) or clusters of genes of potential interest for pathogen control [94].
4. Developing Control Strategies
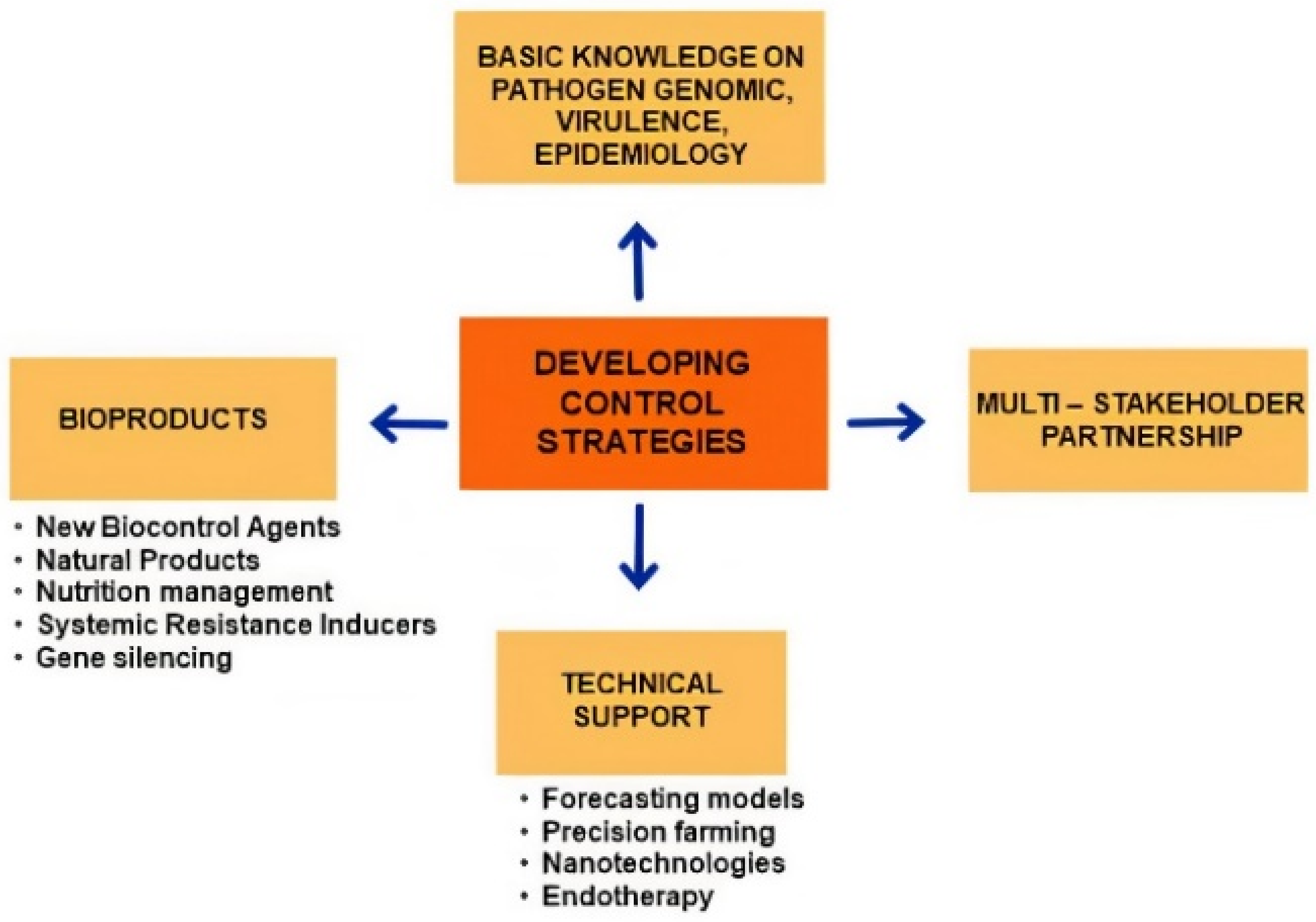
A synoptic panel concerning developing control strategies is shown in Figure 3.
4.1. Technical Support
4.1.1. Precise Timing Decision for Pathogen Control
Modern forecasting models for plant disease should be characterised by (a) flexibility and accuracy; (b) differential interactions between the analysed data and statistical representation; (c) different thresholds of infection risks in relationships with possible mixed infections and changes in crop susceptibility during the season; (d) modelling the disease dynamics, taking into account the long-distance interactions of biological parameters (e.g., spore dispersal through wind and deposition via rainfall events); (e) defining the precise timing for applying biopesticides, taking into account the different scales of climatic and epidemiological data [104]. Knowledge of the pathogenic genetic structure is also important for including such variability into the model according to a fine-tuned pathogen disease cycle. In addition, the assessment of field infection over consecutive years can also be used to calibrate and compare the current-year prediction range [104]. Because some models of the past are hindered by sparse spatial data and a limited use of field monitoring technology, predictive models for pathogen infection based on weather forecasting can now largely benefit from recent advances in the large-scale monitoring of the space provided by satellites [104]. Consequently, a further improvement could be obtained by incorporating data besides the meteorological parameters, obtained by using remote sensing, into the model, such as the leaf reflectance of the red band obtained during the crop growing season [105]. In some circumstances, remote sensing analyses allow for the detection of pre-symptomatic outbreaks and distinguish symptoms caused by different causal agents, such as Phytophthora infestans and Alternaria solani [106]. The final validation of the models in the field and their utilisation by farmers is considered a mandatory step for including the model in an effective IPM strategy [12], and the direct involvement of farmers (i.e., the end-users) in developing such tools is important [107].
Climate change projections should also be considered, so that modern forecasting models also aim to predict possible scenarios of pathogen outbreaks and provide a fine-tuned timing for applying preventive and curative biocontrol agents, or natural products, for plant protection [108]. Through different global circulation models, it is now possible to generate climatic projections of diseases in different geographical areas. Several key plant pathogens, including Fusarium spp., Puccinia recondita, Pyrenophora teres, Magnaporthe oryzae, Plasmopara viticola, Phytophthora infestans, and Xanthomonas oryzae pv. oryzae have been assessed for their potential impact on climate change [108]. These models can also predict the impact of a pathogen outbreak at the landscape level when introducing a new crop adapted to the changing climate or in the case of a widely distributed crop facing a new pathogen introduced from abroad [108].
4.1.2. Precision Farming and Pathogen Control
Strictly connected with the forecasting models, precision farming can provide methods that can be utilised at a single-farm level in order to predict pathogen outbreaks in real-time and site-specifically. The geographical information system (GIS), in combination with on-site monitoring platforms for continuously assessing pedo-climatic data, are the basis for providing alerts to the farmers in relation to the pathogen outbreak [109]. A so-called phenomic approach can assist to establish the precise time for performing the preventive or curative treatment to the crop. The measurements of crop traits such as growth and performance during the season obtained through noninvasive techniques can detect pre-symptomatic events of disease-related changes in the crop that are not visually apparent [110]. In these cases, the “Internet-of-Things” monitoring platform utilises an artificial intelligence algorithm for emulating the decision-making ability of humans regarding the choice of the precise timing for controlling the pathogen [111]. An agro-weather station is installed in the field for measuring many climatic data and additional data such as leaf wetness, soil temperature and water content, and solar radiation in order to detect the early stages of the pathogen infection and to provide an alert to the farmer for applying control measures [111]. Another advantage of precision farming for pathogen control is the management of the disease according to its possible occurrence only in some area of the farm, this resulting in a reduction of pesticide distribution. The monitoring platforms can also detect soilborne diseases by recording the changes in the foliage characteristics [109]. An on-going application for monitoring disease spread both on a large scale and at the single-farm level is the agricultural research outcomes system (AgCROS), developed in Florida to manage the citrus greening disease of citrus crops, caused by the non-cultivable bacterium Candidatus Liberibacter asiaticus. AgCROS is a GIS-based monitoring platform for sharing research data between farmers and researchers and to provide decisions for a better management of the disease [112]. One of the most important outcomes of the platform is the early detection of trees infected by the pathogen. This result can be achieved by analysing, in each farm, some plant physiological indexes such as NDVI and NDRE, shown by the trees during the season and captured by remote sensing through unmanned aerial vehicles (UAV) [112]. Moreover, to reduce the cost of the analyses for large areas, data obtained from satellite can be also utilised [112].
Another application of precision farming coupled with remote sensing data analyses is currently performed to control cotton root rot, caused by the soilborne fungus Phymatotrichopsis omnivora, in Texas [113]. Field multispectral images are acquired though satellite sensors, and field maps are created to monitor the crop. A classification map distinguishes infected field parts from the healthy ones based on different colors pointing out the diseased plants. Consequently, different rates of fungicides and fertilisers are applied to any single farm according to the precise occurrence of diseased plants in a specific area, thus allowing for a relevant reduction of pesticide spread [113].
4.1.3. Nanotechnology
Nanomaterials released within a plant through nanotechnology can provide a relevant contribution to the reduction of agrochemicals spread in the environment. However, we must first assess the interactions of the nanoparticles with the plant tissue [114], as well as the bioavailability and durability of nanoparticles [115], their potential ecotoxicological risks and accumulation in food [116], and the relative costs for their application [116]. In addition, nano-biopesticides should have refined technical properties such as a high solubility of low-solubility active ingredients, a slow targeted release, and the non-premature degradation of active ingredients [117]. Additionally, some traditional pesticides have been re-formulated by developing nanoparticles to reduce the dispersal of the active ingredients in the environment [116]. Among the preventive measures for pathogen control to be applied in sustainable agriculture, copper and zinc-based nanoparticles could be used as seed coating substances or in foliar applications to improve the overall growth of plantlets [118]. The nanomaterials that show potential for application as nanopesticides include silver, copper, zinc, carbon, magnesium, manganese, silicon, calcium carbonate, and chitosan [114]. Bioactive products can also be encapsulated within biodegradable nanomaterials or, alternatively, loaded with plant extracts possessing antimicrobial activities. These nanocarriers can release the active product selectively to the plant. Among these nanocarriers, chitosan oligomers and methacrylated lignin could be effectively utilised [119]. PGPR strains could also be utilised as nano-fertilisers and/or inducers of systemic resistance to phytopathogens [120].
Greenhouse studies have shown the relevant activity of copper-based nanoparticles towards Fusarium oxysporum f. sp. niveum, the causal agent of watermelon root rot [121]. Similarly, sprayed copper-based nanoparticles showed relevant antifungal activities in eggplant grown in a glasshouse on a soilless medium and infected with Verticillium dahliae, whereas copper, zinc, and manganese nanoparticles significantly reduced the pathogenic activity of Fusarium oxysporum f. sp. lycopersici in tomato grown on a soilless medium [122]. Nanoparticles obtained from crude extracts of Chaetomium cochlioides, a fungus that possesses antimicrobial compounds, reduced the severity of rice blast caused by Magnaporthe oryzae by 60% [123]. It should be stressed that, to date, most applications of nanoparticles have been restricted to basic laboratory studies and some field applications [114].
Due to the low degradability of nanoparticles in the natural environment [49], the full development of this sector still requires in-depth studies on the impacts of the different active ingredients used as nanoparticles to determine the long-term effects of nanoparticles on the environment and their residual content in food. The field applications of silver nanoparticles retained as effective antimicrobials are the most striking example of this cautious approach [124]. The need to formulate precise guidelines for nanoparticle utilisation in the field is also required [114,125]. Notably, however, chitosan-derived nanoparticles offer a good avenue for assessing the biosafety risks, since these nanoparticles are created from biodegradable bioactive polymers, are not toxic to humans and animals, and offer good antimicrobial activity towards both fungi and bacteria [117,126].
Antimicrobial peptides obtained from natural compounds or that are designed and synthetically obtained de novo are small peptides that show the potential capacity for controlling plant diseases, as replacements for traditional pesticides once we have verified their impacts on the ecosystem (i.e., the epiphytic microbiota of leaves and fruit) [127]. A current limitation of the wide utilisation of such peptides is the high cost of field treatments. These peptides offer the possibility to control plant diseases by either inactivating the pathogenicity of the target phytopathogen or inciting plant defense mechanisms in a multitarget approach. This possibility was verified by applying a bifunctional synthetic peptide to tomato plants and observing its control capacity towards Pseudomonas syringae pv. tomato, the causal agent of tomato bacterial speck; Xanthomonas campestris pv. vesicatoria, the causal agent of tomato bacterial spot; Botrytis cinerea, the causal agent of tomato grey mould [128].
4.1.4. Endotherapy
The release of specific products within the xylem tissue of trees through trunk injection or trunk infusion is another technique that could allow for a notable reduction of agrochemical dispersal in the environment, reduce the risk of toxicity for farmers, and possibly reduce the overall cost of the protective treatment [129]. The products to be released through endotherapy should be preliminarily verified for their suitability in apoplastic transport within the plant, for the absence of phytotoxic effects [130], and to determine the most suitable plant port for effective injection [131]. There is also a need to reduce the trunk wounds that are caused by devices utilised for the injection [132,133,134]. Apple trees infected with Erwinia amylovora, the causal agent of fire blight, and injected with products that incite the induction of systemic acquired resistance, such as acibenzolar-S-methyl and potassium phosphite, were found to significantly reduce both blossom and shoot blight. This injection also induced the expression of some proteins related to plant defense [133]. Similarly, a study in Apulia (Italy) on evaluating the possibility of controlling Xylella fastidiosa subsp. pauca, the causal agent of olive quick decline syndrome, showed that the trunk injection of a curative biocomplex containing zinc, copper, and citric acid incited an increase in some plant defense-related metabolites, such as oleuropein and polyphenols, as well as a simultaneous decrease in other metabolites related to the disease, such as quinic acid and mannitol [135]. Endotherapy performed with glutaraldehyde on grapevine cuttings provided a satisfactory control of Phaeoacremonium minimum, one of the fungi involved in the esca disease complex [136]. In addition, endotherapy performed on grapevine plants showing symptoms of esca disease complex using chitosan oligomers as nanocarriers loaded with the extracts of some plants offering antimicrobial properties, such as Rubia tinctorum, Equisetum arvensis, Urtica dioica, and Silybum marianum, yielded a significant reduction of foliar symptoms [119].
4.2. New Bioproducts and Sustainability
4.2.1. Biocontrol Agents
The potential use of biocontrol agents in sustainable plant protection is currently limited by factors such as the fragmentation of biocontrol sub-disciplines and crop site-specific factors; unwieldy regulatory processes; increasingly bureaucratic barriers to access biocontrol agents; insufficient engagement and communication with the public, stakeholders, growers, and politicians in regards to the considerable economic benefits of biocontrol; relatively high costs [137,138]. For biosafety reasons, any microorganism released into the environment should be carefully assessed via the post-release field monitoring of putative negative environmental impacts, and well-designed ecological monitoring programs will provide data that can help regulators [139]. The cost-efficiency of the product plays a relevant role in farmer choice, and considerable attention should be devoted by the selling companies to the methods of formulation, storage, and delivery [32]. Additionally, the cost of registration for the biocontrol agents is very high, and some companies register their bioproducts as biofertilisers [140].
Apart from bureaucratic and regulatory barriers, some technical aspects for identifying potential biocontrol agents could also be improved, and the need for new high-throughput screening systems was previously suggested [141]. In particular, marker-based screening that can test, in vitro, many enzymes and/or metabolites linked to the antagonistic activity of the microorganism(s) and perform genomic-based searches for genetic traits that show antagonistic activity should be applied to reveal potential beneficial strains to be further assessed through in planta tests [11,141].
One branch of development for obtaining biocontrol agents is based on bacteriophages and phages, the viruses of bacteria. Apart from their stability in the environment [142], one of the most critical points for the utilisation of biocontrol agents is the definition of their host range. This aspect is critical since the potential dispersal of such agents into the environment should not create problems for beneficial bacterial microflora [143,144]. Moreover, phages should not interfere with the genetic material of the target plant by r eleasing traits through transduction that could be incorporated into the plant cell [143]. There are many studies demonstrating the in vitro effectiveness of phages and phage cocktails in infecting and killing plant pathogenic bacteria, even though relatively few strains have reached the commercial phase [142,143,144]. This lack of commercial applications could be due to the intrinsic difficulty in establishing a direct correlation between laboratory and field conditions [145].
In the U.S.A., Xanthomonas vesicatoria, the causal agent of bacterial spot of tomato and pepper, Pseudomonas syringae pv. tomato, the causal agent of bacterial speck of tomato, Erwinia amylovora, the causal agent of fire blight of apple and pear, and Xylella fastidiosa subsp. fastidiosa, the causal agent of Pierce’s disease of grapevine are examples of bacterial pathogens that can be controlled or mitigated using commercially available products based on phages. In Europe, Enterobacteriaceae (Pectobacterium, Dickeya), which affects potato during post-harvest, and Erwinia amylovora, which is under strict regulation during spring, are the sole pathogens for which commercially distributed phages are allowed [143,144]. Soil-borne pathogens and seeds are other targets for biocontrol through phage utilisation [143,144]. In addition, phage-derived proteins, such as endolysin for Gram-positive species, can be exploited as potential antimicrobial agents [146]. In the future, precision farming through sensor-based technology could effectively assist and improve upon the field utilisation of phages. The detection of early stages of the disease coupled with the best time to avoid leaf desiccation and U.V. stress could facilitate the success of phage activity in the field [144].
Usually, the rhizosphere and phyllosphere are the most commonly exploited resources for isolating potential bacterial strains for biological control. Plant-growth-promoting rhizobacteria are well-known active agents that enhance the plant defense system, thus conferring a general healthy status to the crop. Acinetobacter, Arthrobacter, Azospirillum, Bacillus, Bradyrhizobium, Rhizobium, Sphingomonas, Serratia, and the nonpathogenic Agrobacterium, Burkholderia, and Pseudomonas are the most common isolated genera that show growth-promoting activities [147]. Bacillus spp. and Pseudomonas spp. isolated from the rhizosphere and/or soil are among the bacterial genera most widely utilized as biocontrol agents for soil-borne fungal and bacterial phytopathogens, whereas the genus Streptomyces yielded a relevant number of strains with potential as biocontrol agents, especially towards fungal species [148]. In the phyllosphere, Pseudomonas, Bacillus, and Pantoea are the predominant genera that can be isolated as potential biocontrol agents [149]. Preliminary screening to reveal the occurrence of antimicrobial compounds and antagonistic activities (e.g., hydrolytic enzymes, ammonia, and antibiosis) related to phytopathogen biocontrol and the promotion of plant growth should be performed on a large collection of isolates to highlight the most promising strains [150]. Other important features of successful biocontrol agents include niche-adaptability, competition for nutrients, and colonisation ability [151,152]. Moreover, some commercial products based on strains of the aforementioned bacterial genera are currently used as biocontrol agents for crop protection, including soil-borne diseases.
For example, Pantoea vagans and Pseudomonas fluorescens provide many strains with antagonistic activities to both phytopathogenic bacterial and fungal species. Some strains of both species show control capacity for Erwinia amylovora, the causal agent of fire blight of apple and pear [153]. Additionally, commercial compounds have been developed as potential substitutes for streptomycin application in disease control. Pseudomonas fluorescens is among the most versatile bacterial species and can act as either a plant-growth-promoting or biocontrol agent for plant diseases, especially soil-borne diseases [154]. Similarly, Pseudomonas chlororaphis strains have been registered in commercial formulations to control leaf blight and grey mould in tomato, which are caused by Alternaria alternata and Botrytis cinerea, respectively [155].
Bacillus subtilis, Bacillus amyloliquefaciens, and Bacillus pumilus are largely employed as biocontrol agents for citrus diseases both in the field and during storage—namely, Colletothricum acutatum and Colletothricum gloeosporioides, the causal agents of anthracnose; Alternaria citri, the causal agent of black rot; Phytophthora citrophthora, the causal agent of root rot; Plenodomus thacheiphilus, the causal agent of mal secco; Penicillium digitatum, Penicillium italicum, and Geotrichum candidum, which are involved in fruit decay during storage [156,157]. To improve the effectiveness of these biocontrol agents during fruit storage, additional compounds or physical treatments (i.e., copper hydroxide, sodium carbonate, tea saponins, and hot water) are applied to provide more integrated and effective management [156]. The salt-tolerant Bacillus velezensis showed significant biocontrol activity towards Verticillium wilt of olive [158] (Figure 4).
Bacillus strains could also be effectively used in mixed compounds to exploit the synergistic effects of their diverse antagonistic modes of action towards target pathogenic bacteria and fungi [159]. Potato is another crop that largely benefits from the utilisation of biocontrol agents to prevent tuber soft rotting caused by Pectobacterium spp., Dickeya spp., and Clostridium spp. [160]. The overall strategy includes soaking potato tubers prior to sowing in a solution containing Pseudomonas fluorescens strains to prevent future colonisation by pathogens. Similarly, the soil can also be inoculated with antagonistic bacteria such as Bacillus, Pantoea, Pseudomonas, and Streptomyces to colonize the root zone where phytopathogens usually start plant colonisation.
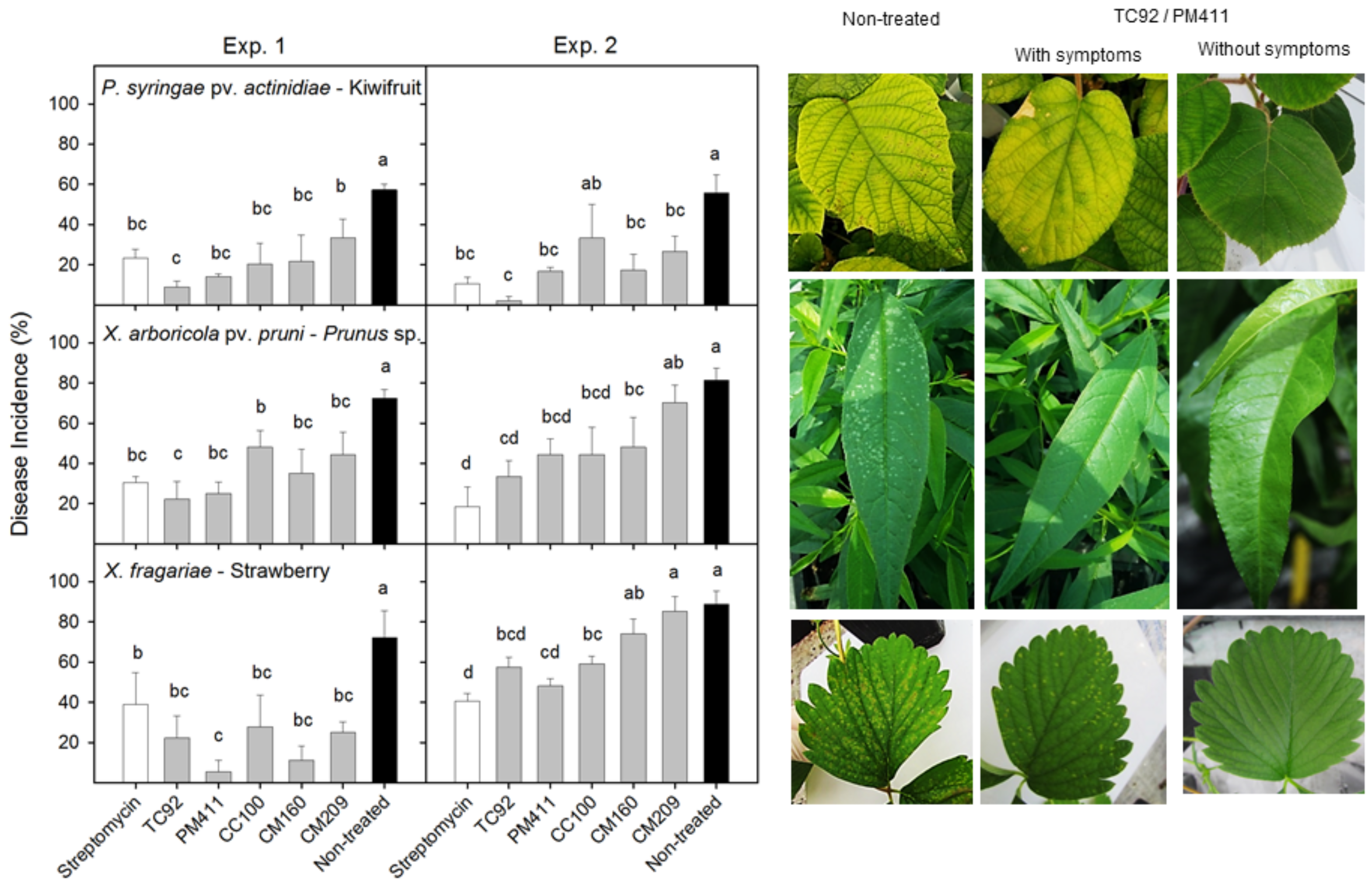
Finally, cocktails of biocontrol agents in the form of a solution or powder are used to protect harvested tubers during storage. In this case, the tubers are soaked in the solution prior to storage [160]. A significant reduction in economic losses caused by rice blast incited by Magnaporthe oryzae is expected to be obtained through the application of Streptomyces strains as biocontrol agents [161,162]. Some lactic acid bacteria, which are usually used as bioprotective agents for foodborne pathogens and spoilage microorganisms, have also shown good potential to act as biocontrol agents for plant pathogens. Due to its antimicrobial metabolites, Lactobacillus plantarum shows broad-spectrum antagonism towards some bacterial phytopathogens such as Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker; Xanthomonas arboricola pv. pruni, the causal agent of bacterial spot of stone fruit, and Xanthomonas fragariae, the causal agent of angular leaf spot of strawberry [163] (Figure 5). A consortium of endophytic lactobacilli composed of Weissella cibaria and Lactococcus lactis enabled the control of papaya dieback caused by Erwinia mallotivora [164].
The versatile behavior of Trichoderma spp. can also be exploited to provide effective control through the colonisation of additional niches besides those that are usually targeted, such as in the case of T. gamsii, which is commonly utilised to protect wheat spikes from Fusarium graminearum, the causal agent of Fusarium head blight of wheat. The addition of fungus in the soil or during sowing can enhance the overall biocontrol activity [165]. Similarly, the addition of Trichoderma polysporum spores to seeds in combination with liquid compost rich in organic matter supplied in fertigation during plant growth enhanced overall bioactivity towards melon wilt caused by Fusarium oxysporum f. sp. melonis under semiarid conditions [166]. A combination of Trichoderma strains with nanoparticles could also provide augmented control effectiveness [167].
The search for new beneficial traits within Trichoderma spp. can now be assisted through genomic prediction tools that reveal the occurrence of a single gene or cluster of genes that produce bioactive product(s) either per se or under some stimuli [168,169]. Several predicted terpenes and phytotoxins with potential bioactive behavior have been found in the genome by genomically screening publicly available Trichoderma genomes [169]. A reduction in production costs is also an important issue to resolve for this important sector of biocontrol agents, and new types of formulations exploiting low-cost substrates for producing pure fungal spores should be implemented [75].
Consortia of bacteria and fungi, so-called multi-strain biological control agents, or microbial synthetic communities, can also be utilised for the management of soil-borne diseases caused by bacteria, fungi, and oomyctes [170] or key fruit pathogens, such Botrytis cinerea, by developing niche-specific microbial interactions [171]. A microbial consortium based on Trichoderma atroviride, Aureobasidium pullulans, and Bacillus subtilis that colonises different ecological niches of the bunch during the season was proven to be effective in significantly reducing the activity of B. cinerea, the causal agent of grey mould of grapevine [171]. According to this strategy, Trichoderma atroviride, a good coloniser of dead plant tissue, provides protection at the bunch closure stage. Additionally, Aureobasidium pullulans can compete for sugar utilisation, with the fungus occurring on cracks in berries, whereas Bacillus subtilis, which produces antagonistic metabolites, should be sprayed close to harvest time [171]. Commercially available formulations contain Trichoderma strains and other bioactive species such as Pseudomonas fluorescens, Pseudomonas aureofaciens, Bacillus spp., and Streptomyces spp. [75]. Consortia of different Streptomyces strains showed the potential capability to reduce the severity of some diseases caused by Fusarium oxysporum in vegetables [172].
Non-harmful endophytic microorganisms are another source of potential biocontrol agents currently being exploited. Generally, these microbes live within plant tissues and provide useful metabolites for plant growth and tolerance to stress, including plant pathogens [13,173]. Volatile organic compounds could also be used to control plant diseases [174]. The genus Trichoderma also includes endophytic species that can be exploited as mycoparasites, as in the case of coffee leaf rust caused by Hemileia vastatrix [175]. Other fungal genera that have shown potential beneficial activities towards fungal phytopathogens include Heteroconium; Ramularia; Xylaria; Candida; and nonpathogenic strains of Fusarium oxysporum, Cladosporium, Colletotrichum, Alternaria, Phoma, Pestalotiopsis, and Botryosphaeria [13,173]. The genus Streptomyces is another source of beneficial endophytes that show antagonistic activities towards soilborne fungi such as Fusarium, Pythium, Verticillium, Rhizoctonia, and Phytophthora (directly to the phytopathogens or mediated through their metabolites). Some commercial products are also available [176]. Bacillus, Paenibacillus spp., and Rhodococcus are other good candidates for obtaining biocontrol agents [177].
4.2.2. Natural Products
Natural products that show biocidal activities are commonly obtained from plant or animal extracts or microbial metabolites. Essential oils, chitosan, some plant extracts (i.e., Yucca schidigera, Equisetum arvense, Punica granatum, Allium cepa, Urtica dioica, and Camellia sinenis), alkaloids, and bacterial lipopeptides are currently being studied, and some are already available as commercial biopesticides [178]. Seaweeds represent another potential source for obtaining biopesticides [179], especially when utilised in a well-planned IPM strategy [180]. Essential oils (i.e., mixtures of terpenes hydrocarbons, alcohols, and phenols) extracted from many plant species have been studied in detail as products with potential biocidal activities towards phytopathogenic fungi, oomycetes, and bacteria, without inciting pathogen resistance [181,182]. However, the stability and persistence of essential oils in the environment, as well as their high cost of authorisation and regulatory barriers, remain obstacles for their wide utilisation in sustainable agriculture [181,182]. Essential oils obtained from Mentha arvensis, Mentha spicata, Juniperus mexicana, Citrus x sinensis, Persicaria odorata, Piper nigrum, Canarium commune, Cinnamomum zeylanicum, Boswellia carterii, Cymbopogon flexuosus, Litsea cubeba, Artemisia alba, Cistus ladaniferus, Copaifera tree, Ferula galbaniflua, Citrus aurantium, and Schinus terebinthifolius are registered in Europe by the European Chemical Agency (Homepage-ECHA (europa.eu)) as being suitable for use in agriculture, with some approved for their biocidal activities. In addition, when applied as seed protectants against cabbage black rot caused by Xanthomonas campestris pv. campestris, essential oils obtained from Zataria multiflora yielded a significant reduction in the further occurrence of disease in the field [183], whereas clove oil obtained from Eugenia caryophyllata was observed to control pomegranate bacterial blight caused by Xanthomonas axonopodis pv. punicae [184]. Essential oils obtained from cumin, basil, and geranium, applied either as seed protectants or directly in the soil, showed effectiveness towards cumin root rot caused by Fusarium spp. [185]. New techniques such as emulsion and the encapsulation of essential oils are currently being studied to enhance the stability and persistence of such oils in the environment [181].
Chitosan is currently being studied both as a plant defense inducer and for its direct involvement in disease control [186,187]. One of chitosan’s properties is its film-forming abilities, which can be exploited to protect the surface of plants, thus avoiding colonization by pathogens. This feature can protect fruits and vegetables under postharvest conditions, and many crops benefit from treatments for controlling fungi involved in decay. These crops include table grapes, strawberry, pear, apple, citrus, peach, sweet cherry, plum, and mango [188]. The film formed by chitosan can also reduce both the incidence and severity of Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker [189]. Significant activity was also observed in rice infected with bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae [190]. Seed dressing is another potential application of chitosan to reduce disease [191]. Interestingly, chitosan showed a synergistic effect with some biocontrol agents such as Trichoderma spp., indicating both castor seed protection [192] and a increased reduction in the pathogenicity of Cercospora beticola and Fusarium oxysporum [193]. Chitosan should also be added to the soil to reduce the severity of tomato crown and root rot caused by Fusarium oxysporum f. sp. radicis lycopersici [194]. Similarly, chitosan should be applied together with essential oils to reduce the volatility of such oils and to enhance their effectiveness in the control of Aspergillus flavus on dates [195].
In a previous study, Yucca schidigera extract protected sorghum seeds through the pathogenic activity of Phoma sorghina, Curvularia lunata, Cladosporium spp., and Fusarium spp. [196]. The macerate of Equisetum arvense (common horsetail) presented control activities that were very similar to those of different copper compounds towards Phytophthora infestans, the causal agent of tomato late blight; Puccinia triticina, the causal agent of durum wheat brown rust; Fusarium graminearum, the causal agent of Fusarium head blight of durum wheat [197]. Pomegranate (Punica granatum) peel extract is another natural product that shows potential activities for the control of phytopathogens. Pomegranate fruit peel extract showed a significant reduction in the severity and incidence of disease for Pseudomonas syringae pv. tomato, the causal agent of tomato bacterial speck, and tomato damping-off caused by Fusarium oxysporum f. sp. lycopersici [198,199]. Pomegranate (Punica granatum) fruit peel extract also showed interesting activities for the postharvest control of Botrytis cinerea, Penicillium expansum, and Penicillium digitatum on sweet cherry and citrus fruits [200]. Camellia sinensis extracts presented antibacterial activities towards Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker [201].
Tetrahydro-β-carboline alkaloids, which naturally occur in fruits, showed potential inhibitory activities towards some emergent and dangerous phytopathogenic bacteria such as Xanthomonas axonopodis pv. citri, the causal agent of Asiatic citrus canker; Xanthomonas oryzae pv. oryzae, the causal agent of rice bacterial blight; Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker [202].
4.2.3. Nutrition Management
Apart from playing fundamental roles in plant growth and development, nutrients are also involved in plant pathogenesis and disease control [203]. Moreover, for pathogen control in sustainable agriculture, preventive balanced nutrition coupled with rational agronomical techniques may be more cost effective and environmentally friendly than the application of any pesticide [36,204]. However, each pathosystem (i.e., the interaction between one single pathogen and one single crop in a certain environment) has its own peculiarities that should be investigated in order to verify the relationships between the contents of each nutrient within the plant and the development of disease over the season [36]. Among the various macronutrients, nitrogen can have either a beneficial or negative effect when applied to a crop to contain disease. These differing behaviors seem to be influenced by the overall characteristics of the pathogen (i.e., an increase in disease severity with obligate pathogens and a decrease in disease severity with facultative pathogens) [205].
The role of phosphorus in crop protection appears to be inconsistent and unpredictable. Relatively high potassium content generally has positive effects in reducing disease severity [202]. In particular, potassium phosphate was found to significantly reduce the severity of barley powdery mildew [206]. After establishing that a specific dose does not reach the limit imposed for the maximum residue level of phosphites, potassium phosphite can also contribute to disease reduction, as in the case of Alternaria solani on potato and Alternaria alternata on citrus fruit [207,208]. Calcium appears to be especially effective for post-harvest treatments against gray mould caused by Botrytis cinerea [209,210], whereas magnesium showed inconsistent results in controlling diseases [211]. Further ad hoc studies should be performed to determine the potential beneficial effects [212].
Among the various micronutrients, copper has been widely used as a fungicide and bactericide for a long period of time, although its excessive utilisation can result in the accumulation of copper in the soil, which poses risks for the microbial microflora [213]. Zinc and manganese are microelements that show clear beneficial activities against plant diseases [205]. Applying a supply of manure rich in zinc prior to planting winter wheat was found to significantly reduce the severity of spring blight caused by Rhizoctonia cerealis [211]. Moreover, the higher uptake of manganese in paddy-grown rice when compared to the low uptake in upland rice cultivations yielded a greater resistance to Magnaporthe oryzae, the causal agent of rice blast [214]. The wheat root rot disease caused by Gaeumannomyces graminis can be effectively managed through the nutrition of the host plant, especially through the supply of manganese [205]. Iron can also enhance the virulence of bacteria and fungi, and soilborne pathogens can be limited by adding rhizosphere-beneficial microorganisms that, through the activation of siderophores, reduce the iron content in the soil [36].
An in-depth study to determine which nutrients can impede the multiplication of pathogens showed that the direct supply of nutrients can help directly manage disease in some circumstances [215]. Another study explored the application of a biocomplex fertiliser containing low doses of zinc, copper, and citric acid to the crowns of infected trees, providing a striking example of the importance of plant nutrition as a strategy for significantly reducing infection in olive groves with symptoms of olive quick decline syndrome caused by Xylella fastidiosa subsp. pauca [216,217] (Figure 6). In this case, zinc and copper ions, which also showed direct bactericidal activities [215], increased in olive trees grown in soils that were characterised by low contents of such ions [218]. The increases in zinc and copper content within the plant that were caused by supplying this biocomplex over a few years allowed the severely infected trees to achieve good yields [214]. Additionally, supplying some nutrients as a foliar spray—in this case, calcium and boron—enabled the better control of gray mould in strawberry, caused by Botrytis cinerea [219].
4.2.4. Systemic Resistance Inducers
The inducers of plant systemic resistance to pathogens show potential applications in IPM strategies. Such resistance can be mediated via (a) salicylic acid or its precursors, which incite systemic acquired resistance (SAR), or by (b) PGPR, which incites induced systemic resistance (ISR) by modulating the jasmonate and ethylene pathways [220]. Such inducers activate signals related to the plant defense mechanisms that are able to counteract pathogen colonisation. Acibenzolar-S-methyl, harpin, chitosan, and extracts of Reynoutria sachalinensis and Saccharomyces cerevisiae are commercially available inducers of resistance, whereas β-aminobutyric acid (BABA), probenazole, saccharin, phosphites, biochar, mycorrizhal fungi, endophytes, and algal extracts are among the most commonly studied future candidates for new commercial products [220]. Acibenzolar-S-methyl is able to induce resistance towards different kinds of pathogens in different crops, including the postharvest decay fungi of mango species such as Colletotrichum gloeosporioides [221]. Moreover, fava bean was found to be protected from Uromyces viciae-fabae and Ascochyta rabiei after acibenzolar-S-methyl treatments [222]. Within an IPM strategy, acibenzolar-S-methyl, in combination with copper treatments, reduced the severity of kiwifruit bacterial canker caused by Pseudomonas syringae pv. actinidiae [223].
Besides plant growth, PGPR mixtures showed the ability to protect crops, as in the case of pepper infected with Xanthomonas vesicatoria, the causal agent of bacterial spot [224]. Similarly, Azospirillum brasilense protected strawberry infected with Colletotrichum acutatum, the causal agent of anthracnose fruit rot [225]. Preventive BABA treatments protected lettuce and grapevine from downy mildew caused by Bremia lactucae and Plasmopora viticola, respectively [226,227]. As an inducer, biochar (i.e., a product of biomass pyrolysis obtained in the absence of oxygen) showed broad-spectrum activities when applied to the soil as an amendment, yielding protection against Botrytis cinerea, Colletotrichum acutatum, and Podosphaera aphanis in strawberry [228]. This product, obtained from olive pruning, also showed inhibitory activities towards tomato spotted wilt virus, a systemic viral agent, without negatively affecting beneficial soil biocontrol agents such as Bacillus spp. and Trichoderma spp. [229]. Marine algae also showed potential broad-spectrum activities towards plant pathogens. Extracts from Ulva fasciata reduced the activities of Colletotrichum lindemuthianum, the causal agent of anthracnose of bean [230]. Additionally, extracts of Ulva armoricana showed activities towards the powdery mildew of grapevine, bean, and cucumber [231].
4.2.5. Gene Silencing
Using RNA interference technology to achieve host-induced silencing of the pathogen gene(s) involved in mechanisms of pathogenicity and virulence is another strategy that is currently being studied to provide sustainable tools for plant disease control, including the control of viruses [232,233,234]. Gene silencing through RNA interference is a common plant strategy aimed at blocking the activities of pathogens. Phytopathogens attempt to overcome this measure through anti-silencing mechanisms aimed at inactivating the host plant’s RNA interference machinery to start the infection [232]. This technology is based on establishing small interfering host-induced (or more recently, exogenous (i.e., sprayed)) non-coding RNAs that, upon their uptake into the plant, can silence genes that are fundamental for the pathogen life cycle and/or infection (i.e., cross-kingdom RNA interference) [232,233,234]. The pathogen target genes used in developing the gene-silencing approach are numerous [234]. However, the success of this strategy largely depends on the efficiency of the pathogen in RNA uptake [235]. Some difficulties in the application of gene silencing include the degradation and uptake of RNA into the plant due to physical or biological interference, the lack of transformability among various crops, and the potential absence of genetic stability among silencing traits [236,237].
The pathogenicity of Fusarium graminearum was significantly reduced on barley leaves by using a spray that released double-stranded RNA, targeting the FgAGO and FgDCL genes in the RNA interference machinery of the fungus [238]. The pathogenic activity of Phytophthora infestans, the causal agent of potato late blight, was significantly reduced by gene-silencing targeting of the hp-PiGPB1 gene, coding for the G protein β-subunit [239]. Botrytis cinerea, the causal agent of grey mould on many crops, is potentially manageable using gene-silencing technology. Spraying fruits, foliage, and flowers with exogenous RNAs targeted to the DCL1 and DCL2 genes, coding for the fungus’ effectors involved in pathogenicity, can obtain a significant reduction of the disease on tomato, lettuce, onion, strawberry, and grapevine [240]. In addition, spray-induced gene silencing has been effectively applied to reduce the virulence of the fungus on grapevine, both on potted plants and on harvested bunches, by targeting the erg11 gene of the cytochrome P450 monooxygenase. Double-stranded RNA was sprayed under high pressure on the leaves, absorbed by the petiole, and sprayed on the bunch at postharvest [241].
5. Concluding Remarks and Perspectives
The successful control of plant diseases in horticulture has long been a pillar of both food production and the conservation of agroecosystems. At present, these two goals are becoming increasingly urgent due to the rapid increase in human population and the threats posed by climate change [242]. The global circulation of plant material, with the potential introduction in new areas of dangerous phytopathogens, poses additional risks to safeguarding crop longevity and, in some circumstances, concerns the whole landscape of a territory. Within this context, high levels of crop productivity, conservation, and improvements to agroecosystem fertility should not be viewed as contradictory, and the development and utilisation of effective environmentally sustainable strategies should be applied more consistently to integrate and substitute for chemical pesticides in the near future [243]. Achieving good quality fruits and vegetables during the postharvest period is a complementary and relevant goal to achieve [244]. This scenario is largely fostered by supranational policies that are supporting projects and developing strategies to increase either the achievement of scientific and technical results or the wide acceptance of a “greener” agriculture. Many scientists involved in various aspects of sustainable agriculture complain about delays in achieving the regulation and commercial utilisation of known and novel biopesticides and/or products that can control phytopathogens. The safety and quality checks of these formulations are compulsory, but more frequent and initiative-taking activities among regulatory institutions are required to promote the spread of new active ingredients.
The conservation of agroecosystems according to sustainability principles greatly depends on the development and application of environmentally friendly strategies on a large scale. The “multi-stakeholder partnership” is an interesting strategy that could support the spread and acceptance of sustainable agriculture. This concept highlights the possibility that diverse groups in the agro-food chain can have common problems or aspirations despite having different interests [245]. A striking example of such a strategy for sustainable agriculture came from the agreement between Barilla, a well-known Italian brand of pasta, and some private companies that produce and supply ingredients such as tomato, sugar beet, and cereals; these organisations had no previous contact. These partners, including the farmers of each single company, share mutual and diversified interests in supporting their planned activities. The network also includes agronomists, research centers, and universities. Single farms are directed to adopt crop rotation, nitrogen fixer crops or green manure, and IPM in their agronomical practices. However, these practices are ruled out in most common methods of production. Farms receive a supply contract from Barilla with some guarantees concerning the price and quantity of the product over many years [246]. These initiatives appear to be positive for improving the circulation of sustainable principles in agriculture and should be expanded to promote sustainable agriculture.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Siebrecht, N. Sustainable agriculture and its implementation gap-overcoming obstacles to implementation. Sustainability 2021, 12, 3853. [Google Scholar] [CrossRef]
- Carlisle, L.; Montenegro de Wit, M.; Delonge, M.S.; Iles, A.; Calo, A.; Getz, C.; Ory, J.; Munden-Dixon, K.; Galt, R.; Melone, B.; et al. Transitioning to sustainable agriculture requires growing and sustaining an ecologically skilled workforce. Front. Sustain. Food Syst. 2019, 3, 96. [Google Scholar] [CrossRef]
- McNeill, D. The contested discourse of sustainable agriculture. Glob. Policy 2019, 10 (Suppl. 1), 16–27. [Google Scholar] [CrossRef] [Green Version]
- Hertoge, K. Mals/Malles Venosta Referendum. 2014. Available online: http://www.marcozullo.it/wp-content/uploads/Malles-Venosta-Referendum.pdf (accessed on 22 May 2022).
- Damalas, C.A.; Koutroubas, S.D. Current status and recent development in biopesticides use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in sustainable agriculture: A critical sustainable development driver governed by green chemistry principles. Front. Sust. Food Syst. 2021, 5, 619058. [Google Scholar] [CrossRef]
- Garrett, K.A. Climate change and plant disease risk. In Global Climate Change and Extreme Weather Events: Understanding the Contributions to Infectious Disease Emergence; Relman, D.A., Hamburg, M.A., Choffnes, E.R., Mack, A., Eds.; National Academies Press: Washington, DC, USA, 2008; pp. 143–155. [Google Scholar]
- Juroszek, P.; Von Tiedemann, A. Potential strategies and future requirements for plant disease management under a changing climate. Plant Pathol. 2011, 60, 100–112. [Google Scholar] [CrossRef]
- Soussana, J.-F.; Graux, A.-I.; Tubiello, F.N. Improving the use of modelling for projections of climate change impacts on crops and pastures. J. Exp. Bot. 2010, 61, 2217–2228. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.; Thrall, P.H.; Burdon, J.J. Achieving sustainable plant disease management through evolutionary principles. Trends Plant Sci. 2014, 19, 570–575. [Google Scholar] [CrossRef]
- Leung, K.; Ras, E.; Ferguson, K.B.; Ariens, S.; Babendreier, D.; Bijma, P.; Bourtzis, K.; Brodeur, J.; Bruins, M.A.; Centurion, A.; et al. Next-generation biological control: The need for integrating genetics and genomics. Biol. Rev. 2020, 95, 1838–1854. [Google Scholar] [CrossRef]
- Fenu, G.; Malloci, F.M. Forecasting plant and crop disease: An explorative study on current algorithms. Big Data Cogn. Comput. 2021, 5, 2. [Google Scholar] [CrossRef]
- Fontana, D.C.; De Paula, S.; Torres, A.G.; Moura de Souza, V.H.; Pascholati, S.F.; Schmidt, D.; Dourado Neto, D. Endophytic fungi: Biological control and induced resistance to phytopathogens and abiotic stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- He, D.-C.; He, M.-H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economics consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Ferrante, P.; Scortichini, M. Frost promotes the pathogenicity of Pseudomonas syringae pv. actinidiae in Actinidia chinensis and A. deliciosa plants. Plant Pathol. 2014, 63, 12–19. [Google Scholar]
- Nuttall, J.G.; Perry, E.M.; Delahunty, A.J.; O’Leary, G.J.; Barlow, K.M.; Wallace, A.J. Frost response in wheat and early detection using proximal sensors. J. Agron. Crop Sci. 2019, 205, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Barlow, K.M.; Christy, B.P.; O’Leary, G.J.; Riffkin, P.A.; Nuttall, J.G. Simulating the impact of heat and frost events on wheat crop production: A review. Field Crop Res. 2015, 171, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Haworth, M.; Marino, G.; Brunetti, C.; Killi, D.; De Carlo, A.; Centritto, M. The impact of the heat stress and water deficit on the photosynthetic and stomatal physiology of olive (Olea europea L.)—A case study of the 2017 heat wave. Plants 2018, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Silvestri, C.; Bacchetta, L.; Bellincontro, A.; Cristofori, V. Advances in cultivar choice, hazelnut orchard management, and nut storage to enhance product quality and safety: An overview. Sci. Food Agric. 2021, 101, 27–43. [Google Scholar] [CrossRef]
- Macholdt, J.; Honermeier, B. Importance of variety choice: Adapting to climate change in organic and conventional farming system in Germany. Outlook Agric. 2017, 46, 178–184. [Google Scholar] [CrossRef]
- Hulme, P.E. Unwelcome exchange: International trade as direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, 666–679. [Google Scholar] [CrossRef]
- Garbelotto, M.; Pautasso, M. Impacts of exotic forest pathogens on Mediterranean ecosystems: Four case studies. Eur. J. Plant Pathol. 2012, 133, 101–116. [Google Scholar] [CrossRef]
- European Commission. Proposal for a regulation of the european parliament and of the council establishing rules on support for strategic plans to be drawn up by Member States under the Common agricultural policy (CAP Strategic Plans) and financed by the European Agricultural Guarantee Fund (EAGF) and by the European Agricultural Fund for Rural Development (EAFRD) and repealing Regulation (EU); No 1305/2013 of the European Parliament and of the Council and Regulation (EU) No 1307/2013 of the European Parliament and of the Council; COM/2018/392 final—2018/0216 (COD). 2018. [Google Scholar]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.-F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Peeters, A.; Ambhul, E.; Barberi, P.; Migliorini, P.; Ostermann, O.; Goris, M.; Donham, J.; Wezel, A.; Batello, C. Integrating Agroecology into European Agricultural Policies. Position Paper and Recommendations to the European Commission on Eco-Schemes. 2021. Available online: https://www.agroecology-europe.org/wp-content/uploads/2021/07/AEEU_Positionpaper_Ecoschemes_FINAL_english.pdf (accessed on 22 May 2022).
- Larkin, R.P.; Lynch, R.P. Use and effects of different Brassica and other rotation crops on soilborne diseases and yield of potato. Horticulturae 2018, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Muscolo, A.; Sidari, M.; Nardi, S. Humic substance: Relationships between structure and activity. Deeper information suggests univocal findings. J. Geochem. Explor. 2013, 129, 57–63. [Google Scholar] [CrossRef]
- Rillig, M.C.; Sosa-Hernandez, M.A.; Roy, J.; Aguilar-Trigueros, C.A.; Valyi, K.; Lehmann, A. Towards an integrated mycorrhizal technology: Harnessing mycorrhiza for sustainable intensification in agriculture. Front. Plant Sci. 2016, 7, 1625. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslikova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanism of action, and roadmap to commercialization of biostimulant for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Semida, W.M.; Beheiry, H.R.; Sétamou, M.; Simpson, C.R.; Abd El-Mageed, T.A.; Rady, M.M.; Nelson, S.D. Biochar implications for sustainable agriculture and environment: A review. S. Afr. J. Bot. 2019, 127, 333–347. [Google Scholar] [CrossRef]
- Seenivasagan, R.; Babalola, O.O. Utilization of microbial consortia as biofertilizers and biopesticides for the production of feasible agricultural product. Biology 2021, 10, 1111. [Google Scholar] [CrossRef]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The use of microbial inoculants for biological control, plant growth promotion, and sustainable agriculture: A review. Eur. J. Plant Pathol. 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Suarez-Estrella, F.; Arcos-Nievas, M.A.; Lopez, M.J.; Vargas-Garcia, M.C.; Moreno, J. Biological control of plant pathogens by microorganisms isolated from agro-industrial composts. Biol. Control 2013, 67, 509–515. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Lu, C.; Liao, Q.; Gudda, F.O.; Ling, W. Antibiotics in animal manure and manure-based fertilizers: Occurrence and ecological risk assessment. Chemosphere 2020, 255, 127006. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Michael, C.; Gil, E.; Gallart, M.; Stavrinides, M.C. Influence of spray technology and application rate on leaf deposit and ground losses in mountain viticulture. Agriculture 2020, 10, 615. [Google Scholar] [CrossRef]
- Gamliel, A. Application of soil solarization in the open field. In Soil Solarization THEORY and Practice; Gamliel, A., Katan, J., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2017; pp. 175–180. [Google Scholar]
- Van Bruggen, A.H.C.; Gamliel, A.; FinckH, M.R. Plant disease management in organic farming systems. Pest Sci. Manag. 2016, 72, 30–44. [Google Scholar] [CrossRef]
- Hansen, Z.R.; Everts, K.L.; Fry, W.E.; Gevens, A.J.; Grunwald, N.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; Knaus, B.J.; McGrath, M.T.; et al. Genetic variation within clonal lineages of Phytophthora infestans revealed through genotyping-by-sequencing, and implications for late blight epidemiology. PLoS ONE 2016, 11, e065690. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Y.; Ben Naim, Y.; Falach, L.; Rubin, A.E. Epidemiology of basil downy mildew. Phytopathology 2017, 107, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Newberry, A.A.; Babu, B.; Roberts, P.D.; Dufault, N.S.; Goss, E.M.; Jones, J.B.; Paret, M.L. Molecular epidemiology of Pseudomonas syringae pv. syringae causing bacterial leaf spot of watermelon and squash in Florida. Plant Dis. 2018, 102, 511–518. [Google Scholar]
- Pegg, K.G.; Coates, L.M.; O’Neill, W.T.; Turner, D.W. The epidemiology of Fusarium wilt of banana. Front. Plant Sci. 2019, 10, 1395. [Google Scholar] [CrossRef] [Green Version]
- Pruvost, O.; Boyer, K.; Ravigné, V.; Richard, D.; Vernière, C. Deciphering how plant pathogenic bacteria disperse and meet: Molecular epidemiology of Xanthomonas citri pv. citri at a microgeographic scales in a tropical area of Asiatic citrus canker endemicity. Evol. Appl. 2019, 12, 1523–1538. [Google Scholar]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balestra, G.M.; Spinelli, F. Pseudomonas syringae pv. actinidiae: Ecology, infection dynamics and disease epidemiology. Environ. Microbiol. 2020, 80, 81–102. [Google Scholar]
- Ostos, E.; Garcia-Lopez, M.T.; Porras, R.; Lopez-Escudero, F.J.; Trapero-Casas, A.; Michaelides, T.J.; Moral, J. Effect of cultivar resistance and soil management on spatial-temporal development of Verticillium wilt of olive: A long-term study. Front. Plant Sci. 2020, 11, 584496. [Google Scholar] [CrossRef]
- Caffi, T.; Rossi, V. Fungicide models are components of multiple modeling approaches for decision-making in crop protection. Phytopathol. Medit. 2018, 57, 153–169. [Google Scholar]
- Kim, K.-H.; Jung, I. Development of a daily epidemiological model for rice blast tailored for seasonal disease early warning in South Korea. Plant Pathol. J. 2020, 36, 406–417. [Google Scholar] [CrossRef]
- He, X.; Fu, P.; Aker, W.G.; Hwang, H.-M. Toxicity of engineered nanomaterials mediated by nano–bio–eco interactions. J. Environ. Sci. Health Part C 2018, 36, 21–42. [Google Scholar] [CrossRef]
- Gusberti, M.; Klemm, U.; Meier, M.S.; Maurhofer, M.; Hunger-Glaser, I. Fire blight control: The struggle goes on. A comparison of different fire blight control methods in Switzerland with respect to biosafety, efficacy and durability. Int. J. Environ. Res. Public Health 2015, 12, 11422–11447. [Google Scholar] [CrossRef] [Green Version]
- Moser, R.; Pertot, I.; Elad, Y.; Raffaelli, R. Farmers’ attitudes toward the use of biocontrol agents in IPM strawberry production in three countries. Biol. Control 2008, 47, 125–132. [Google Scholar] [CrossRef]
- Barzmann, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Jorgensen, L.N.; Hovmoller, M.S.; Hansen, J.G.; Lassen, P.; Clark, B.; Bayles, R.; Rodemann, B.; Flath, K.; Jahn, M.; Goral, T.; et al. IPM strategies and their dilemmas including an introduction to www.eurowheat.org. J. Integr. Agric. 2014, 13, 265–281. [Google Scholar] [CrossRef]
- Berlin, A.; Nordström Kälström, H.; Lindgren, A.; Olson, A. Scientific evidence for sustainable plant disease protection strategies for the main arable crops in Sweden. A systematic map protocol. Environ. Evid. 2018, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Holb, I.J.; Abpnyi, F.; Bourma, J.; Heijne, B. On-farm and on-station evaluations of three orchard management approaches against apple scab and apple powdery mildew. Crop Prot. 2017, 97, 109–118. [Google Scholar] [CrossRef]
- Shipp, L.; Elliott, D.; Gillespie, D.; Brodeur, J. From chemical to biological control in Canadian greenhouse crops. In Biological Control: A Global Perspective; Vincent, C., Goettel, M.S., Lazarovitis, C., Eds.; CABI: Wallingford, UK, 2007; pp. 118–127. [Google Scholar]
- Jacobsen, B.J.; Zidack, N.K.; Larson, B.J. The role of Bacillus-based biological control agents in integrated pest management systems: Plant diseases. Phytopathology 2004, 94, 1272–1275. [Google Scholar] [CrossRef] [Green Version]
- Galletti, S.; Paris, R.; Cianchetta, S. Selected isolates of Trichoderma gamsii induce different pathways of systemic resistance in maize upon Fusarium verticillioides challenge. Microbiol. Res. 2020, 233, 126406. [Google Scholar] [CrossRef]
- Muneret, L.; Mitchell, M.; Seufert, V.; Aviron, S.; Djoud, E.A.; Pétillon, J.; Plantegenest, M.; Thiéry, D.; Rusch, A. Evidence that organic farming promotes pest control. Nat. Sustain. 2018, 1, 361–368. [Google Scholar] [CrossRef]
- Baker, B.P.; Green, T.A.; Cooley, D.; Futrell, S.; Garling, L.; Gershuny, G.; Moyer, J.; Rajotte, E.G.; Seaman, A.J.; Young, S.L. Organic Agriculture and Integrated Pest Management: A Synergistic Partnership to Improve Sustainable Agriculture and Food Systems. 2015. Available online: https://organicipmwg.files.wordpress.com/2015/07/white-paper.pdf (accessed on 22 May 2022).
- Rossi, V.; Salinari, F.; Poni, S.; Caffi, T.; Bettati, T. Addressing the implementation problem in decision support systems: The example of vite.net®. Comput. Electron. Agric. 2014, 100, 88–99. [Google Scholar] [CrossRef]
- Minchinton, E.J.; Auer, D.P.F.; Thomson, F.M.; Trapnell, L.; Petkowski, J.E.; Galea, V.J.; Faggian, R.; Kita, N.; Murdoch, C.; Kennedy, R. Evaluation of the efficacy and economics of irrigation management, plant resistance and BrassicaspotTM models for management of white blister on Brassica crops. Austalasian Plant Pathol. 2013, 42, 169–178. [Google Scholar] [CrossRef]
- Pavan, W.; Fraisse, C.W.; Peres, N.A. Development of a web-based disease forecasting system for strawberry. Comput. Electron. Agric. 2011, 75, 169–175. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sund, I.; Eilenberg, J. Why has the authorization of microbial biocontrol agents been slower in the EU than in comparable jurisdictions? Pest Manag. Sci. 2021, 77, 2170–2178. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Hinz, H.; Mulema, J.; Weyl, P.; Ryan, M.J. Biological control and the Nagoya Protocol on access and benefit sharing-a case of effective due diligence. Biocontrol Sci. Technol. 2018, 28, 914–926. [Google Scholar] [CrossRef]
- Pliego, C.; Ramos, C.; De Vicente, A.; Cazorla, F.M. Screening for candidate bacterial biocontrol agents against soilborne fungal pathogens. Plant Soil 2011, 340, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Ajouz, S.; Nicot, P.C.; Bardin, M. Adaptation to pyrrolnitrin of Botrytis cinerea and cost of resistance. Plant Pathol. 2010, 59, 556–566. [Google Scholar] [CrossRef]
- Wei, W.; Xu, Y.; Li, S.; Zhu, L.; Song, J. Developing suppressive soil for root disease of soybean with continuous long-term cropping of soybean in black soils of Northeast China. Acta Agric. Scand. B Soil Plant Sci. 2015, 65, 279–285. [Google Scholar] [CrossRef]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Annu. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicot, P.C.; Bardin, M.; Alabouvette, C.; Köhl, J.; Ruocco, M. Potential of biological control based on published research. 1. Protection against plant pathogens of selected crops. In Classical and Augmentative Biological Control against Diseases and Pests: Critical Status Analysis and Review of Factors Influencing Their Success; Nicot, P.C., Ed.; IOBC/WPRS, 2011; pp. 1–11. [Google Scholar]
- Moore, L.W.; Warren, G. Agrobacterium radiobacter strain K84 and biological control of crown gall. Annu. Rev. Phytopathol. 1979, 17, 163–179. [Google Scholar] [CrossRef]
- Köhl, J.; Scheer, C.; Holb, I.J.; Masny, S.; Molhoek, W.M.L. Toward an integrated use of biological control of Cladosporium cladosporioides H39 in apple scab (Venturia inaequalis) management. Plant Dis. 2015, 99, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Haddoudi, I.; Cabrefiga, J.; Mora, I.; Mhadhbi, H.; Montesinos, E.; Mrabet, M. Biological control of fusarium wilt caused by Fusarium equiseti in Vicia faba with broad spectrum antifungal plant-associated Bacillus spp. Biol. Control 2021, 160, 104671. [Google Scholar] [CrossRef]
- Woo, S.L.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Sood, M.; Kapoor, D.; Sheteiwy, M.; Ramakhrisnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell Infect. Microbiol. 2020, 10, 604293. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S.; Norelli, J.; Liu, J.; Schena, L. Alternative management technologies for postharvest disease control: The journey from simplicity to complexity. Postharvest Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Höfte, M. The use of Pseudomonas spp. as bacterial biocontrol agents to control plant diseases. In Microbial Bioprotectants for Plant Disease Management; Köhl, J., Ravensberg, V., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; p. 74. [Google Scholar]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapon for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 3. [Google Scholar] [CrossRef] [Green Version]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [Green Version]
- Kunz, S.; Schmitt, A.; Haug, P. Field testing of strategies for fire blight control in organic fruit growing. Acta Hortic. 2011, 896, 431–436. [Google Scholar] [CrossRef]
- Iqbal, M.; Jamshaid, M.; Zahid, M.A.; Andreasson, E.; Vetukuri, R.R.; Stenberg, J. Biological control of strawberry crown rot, root rot and grey mould by the beneficial fungus Aureubasidium Pullulans. BioControl 2021, 66, 535–545. [Google Scholar] [CrossRef]
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Dinoor, A. Combining biocontrol agents to reduce the variability of biological control. Phytopathology 2001, 91, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Raupach, G.S.; Kloepper, J.W. Mixtures of plant growth-promoting rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology 1998, 88, 1158–1164. [Google Scholar] [CrossRef] [Green Version]
- Leibinger, W.; Breuker, B.; Hahn, M.; Mendgen, K. Control of postharvest pathogens and colonization of the apple surface by antagonistic microorganisms in the field. Phytopathology 1997, 87, 1103–1110. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.-M.; Jeffries, P.; Pautasso, M.; Jeger, M.J. Combined use of biocontrol agents to manage plant disease in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [Green Version]
- Sarrocco, S.; Valenti, F.; Manfredini, S.; Esteban, P.; Bernardi, R.; Puntoni, G.; Baroncelli, R.; Haidukowski, M.; Moretti, A.; Vannacci, G. Is exploitation competition involved in a multitrophic strategy for the biocontrol of Fusarium head blight? Phytopathology 2019, 109, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Roselló, G.; Francés, J.; Daranas, N.; Montesinos, E.; Bonaterra, A. Control of fire blight of pear trees with mixed inocula of two Lactobacillus plantarum strains and lactic acid. J. Plant Pathol. 2017, 99, 111–120. [Google Scholar]
- Slusarenko, A.J.; Patel, A.; Portz, D. Control of plant diseases by natural products: Allicin from garlic as a case study. Eur. J. Plant Pathol. 2008, 121, 313–322. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Dhangial, D.S.; Singh, J. Plant disease management by bioactive natural products. In Natural Bioactive Products in Sustainable Agriculture; Singh, J.l., Yadav, A.l., Eds.; Springer: Singapore, 2022; pp. 15–19. [Google Scholar]
- Yan, Y.; Liu, Q.; Jacobsen, S.E.; Tang, Y. The impact and prospect of natural product discovery in agriculture. EMBO Rep. 2018, 19, e46824. [Google Scholar] [CrossRef]
- Sundin, G.W.; Wang, N. Antibiotic resistance in plant pathogenic bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef]
- Taylor, P.; Reeder, R. Antibiotics use on crops in low and middle-income countries based on recommendations made by agricultural advisors. CABI Agric. Biosci. 2020, 1, 1. [Google Scholar] [CrossRef]
- Wei, Z.-M.; Beer, S. Harpin from Erwinia amylovora induces plant resistance. Acta Hortic. 1996, 411, 223–226. [Google Scholar] [CrossRef]
- De Capdeville, G.; Beer, S.V.; Wilson, C.L.; Aist, J.R. Alternative disease control agents induce resistance to blue mold in harvested “Red Delicious” apple fruit. Phytopathology 2002, 92, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daayf, F.; Schmitt, A.; Bèlanger, R.R. The effects of plant extracts of Reynouria sachalinensis on powdery mildew development and leaf physiology of long English cucumber. Plant Dis. 1995, 79, 577–580. [Google Scholar] [CrossRef]
- Trottin-Caudal, Y.; Fournier, C.; Leyre, J.-M.; Decognet, V.; Nicot, P.C.; Bardin, M. Efficiency of Plant Extract from Reynoutria Sachalinensis (Milsana) to Control Powdery Mildew on Tomato (Oidium Neolycopersici); ffhal-02764311f; Colloque International: Avignon, France, 2003. [Google Scholar]
- Margaritopoulos, T.; Toufexi, E.; Kizis, D.; Balayiannis, G.; Anagnostopoulos, C.; Theocharis, A.; Rempelos, L.; Troyanos, Y.; Leifert, C.; Markellou, E. Reynoutria sachalinensis elicits SA-dependent defense responses in courgetti genotypes against powdery mildew caused by Podosphaera xanthii. Sci. Rep. 2020, 10, 3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Ghaouth, A.; Arul, J.; Asselin, A.; Benhamou, N. Antifungal activity of chitosan on post-harvest pathogens: Induction of morphological and cytological alterations in Rhizopus stolonifer. Mycol Res. 1992, 96, 769–779. [Google Scholar] [CrossRef]
- Gould, F.; Brown, Z.S.; Kuzma, J. Wicked evolution: Can we address the sociobiological dilemma of pesticide resistance? Science 2018, 360, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Newlands, N.K. Model-based forecasting of agricultural crop disease risk at the regional scale, integrating airborne inoculum, environmental, and satellite-based monitoring data. Front. Environ. Sci. 2018, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Pu, R.; Yuan, L.; Huang, W.; Nie, C.; Yang, G. Integrating remotely sensed and meteorological observations to forecast wheat powdery mildew at a regional scale. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 4328–4339. [Google Scholar] [CrossRef]
- Gold, K.M.; Townsend, P.A.; Chlus, A.; Hermann, I.; Couture, J.J.; Larson, E.R.; Gevens, A.J. Hyperspectral measurements enable pre-symptomatic detection and differentiation of contrasting physiological effects of late blight and early blight in potato. Remote Sens. 2020, 12, 286. [Google Scholar] [CrossRef] [Green Version]
- Lindblom, J.; Lundstrom, C.; Ljung, J.; Jonsson, A. Promoting sustainable intensification in precision agriculture: Review of decision support systems development and strategies. Precis. Agric. 2017, 18, 309–331. [Google Scholar] [CrossRef] [Green Version]
- Newbery, F.; Qi, A.; Fitt, B.D.L. Modelling impacts of climate change on arable crop diseases: Progress, challenge and applications. Curr. Opin. Plant Biol. 2016, 32, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.P.; Short, N.M., Jr.; Sill, J.; Lakshman, D.K.; Hu, X.; Buser, M. Precision agriculture and geospatial techniques for sustainable disease control. Ind. Phytopathol. 2021, 74, 287–305. [Google Scholar] [CrossRef]
- Simko, I.; Jimenez-Berni, J.A.; Sirault, X.R.R. Phenomic approach and tool for phytopathologists. Phytopathology 2017, 107, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Khattab, A.; Habib, S.E.D.; Ismail, H.; Zayan, S.; Fahmy, Y.; Khairy, M.M. An IoT-based cognitive monitoring system for early plant disease forecast. Comput. Electron. Agric. 2019, 166, 105028. [Google Scholar] [CrossRef]
- Lu, X.; Lee, W.; Minzan, L.; Ehsani, R.; Mishra, A.; Yang, C.; Mangan, R. Feasibility study on huanglongbing (citrus greening) detection based on worldview-2 satellite imagery. Biosyst. Eng. 2015, 132, 28–38. [Google Scholar]
- Yang, C. Remote sensing and precision agriculture technologies for crop disease detection and management with a practical application example. Engineering 2020, 6, 528–532. [Google Scholar] [CrossRef]
- Mittal, D.; Kaur, G.; Singh, P.; Yadav, K.; Ali, S.A. Nanoparticle-based sustainable agriculture and food science: Recent advances and future overlook. Front. Nanotechnol. 2020, 2, 579954. [Google Scholar] [CrossRef]
- Kah, M.; Hofmann, T. Nanopesticide research: Current trend and future priority. Environ. Int. 2014, 63, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, L.M.; Pourzahedi, L.; Laughton, S.; Gao, X.; Zimmermann, J.B.; Theis, T.L.; Westerhoff, P.; Lowry, G.V. Guiding the design space for nanotechnology to advance sustainable crop production. Nature Nanotechnol. 2020, 15, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Fortunati, E.; Mazzaglia, A.; Balestra, G.M. Sustainable control strategies for plant protection and food packaging sectors by natural substances and novel nanotechnologies approaches. J. Sci. Food Agric. 2019, 99, 986–1000. [Google Scholar] [CrossRef]
- De Oliveira, J.L. Nano-biopesticides: Present concepts and future perspectives in integrated pest management. In Advances in Nano-Fertilizers and Nano-Pesticides in Agriculture. A Smart Delivery System for Crop Improvement; Jogaiah, S., Singh, H.B., Fraceto, L.F., De Lima, R., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 2021; pp. 1–27. [Google Scholar]
- Sanchez-Hernandez, E.; Langa-Lomba, N.; Gonzalez-Garcia, V.; Casanova-Gascon, J.; Martin-Gil, J.; Santiago-Aliste, A.; Torres-Sanchez, S.; Martin-Ramos, P. Lignin-chitosan nanocarriers for the delivery of bioactive natural products against wood-decay phytopathogens. Agronomy 2022, 12, 461. [Google Scholar] [CrossRef]
- Kalia, A.; Kaur, H. Nano-biofertilizers: Harnessing dual benefts of nano-nutrient and bio-fertilizers for enhanced nutrient use efficiency and sustainable productivity. In Nanoscience for Sustainable Agriculture; Pudake, R., Chauhan, N., Kole, C., Eds.; Springer: New York, NY, USA, 2019; pp. 51–73. [Google Scholar]
- Borgatta, J.; Ma, C.; Hudson-Smith, N.; Elmer, W.; Plaza Peréz, C.D.; De La Torre-Roche, R.; Zuverza-Mena, N.; Haynes, C.L.; White, J.C.; Hamers, R.J. Copper based nanomaterials suppress root fungal disease in watermelon (Citrullus lanatus): Role of particle morphology, composition and dissolution behaviour. ACS Sustain. Chem. Eng. 2018, 6, 11. [Google Scholar] [CrossRef]
- Elmer, W.H.; White, J.C. The use of metallic oxide nanoparticles to enhance growth of tomatoes and eggplants in diseased infected soil or soilless medium. Environ. Sci. Nano 2016, 3, 1072–1079. [Google Scholar] [CrossRef]
- Song, J.J.; Soytong, K.; Kanokmedhakul, S.; Poeaim, S. Natural products of nanoparticles constructed from Chaetomium spp. to control rice blast disease caused by Magnaporthe oryzae. Int. J. Agron. Biol. 2020, 23, 1013–1020. [Google Scholar]
- Sharma, J.; Singh, V.K.; Kumar, A.; Shankarayan, R.; Mallubhotla, S. Role of silver nanoparticles in treatment of plant diseases. In Microbial Biotechnology; Patra, J., Das, G., Shin, H.S., Eds.; Springer: Singapore, 2018; pp. 435–454. [Google Scholar]
- Kookana, R.S.; Boxall, A.B.A.; Reeves, P.T.; Ashauer, R.; Beulke, S.; Chaudry, K.; Cornelis, G.; Fernandes, T.F.; Gan, J.; Kah, M.; et al. Nanopesticides: Guiding principles for regulatory evaluation of environmental risk. J. Agric. Food Chem. 2014, 62, 4227–4240. [Google Scholar] [CrossRef] [Green Version]
- Freepons, D. Chitosan, does it have a place in agriculture? Proc. Plant Growth Regul. Soc. Am. 1991, 10, 11–19. [Google Scholar]
- Montesinos, E.; Badosa, E.; Cabrefiga, J.; Planas, M.; Feliu, L.; Bardají, E. Antimicrobial peptides for plant disease control. From discovery to application. In Small Wonders: Peptides for Disease Control; Rajasekaran, K., Cary, J., Jaynes, J., Montesinos, E., Eds.; Blackwell: Oxford, UK, 2012; pp. 235–261. [Google Scholar]
- Montesinos, L.; Gascon, B.; Ruz, L.; Badosa, E.; Planas, M.; Feliu, L.; Montesinos, E. A bifunctional synthetic peptide with antimicrobial and plant elicitation properties that protect tomato plants from bacterial and fungal infections. Front. Plant Sci. 2021, 12, 756357. [Google Scholar] [CrossRef]
- Berger, C.; Laurent, F. Trunk injection of plant protection products to protect trees from pests and pathogens. Crop Prot. 2019, 124, 104831. [Google Scholar] [CrossRef]
- Akinsamni, O.A.; Drenth, A. Phosphite and metalaxyl rejuvenate macadamia trees in decline caused by Phytophtora cinnamomi. Crop Prot. 2013, 53, 29–36. [Google Scholar] [CrossRef]
- Archer, L.; Albrecht, U.; Crane, J. Trunk injection to deliver crop protection materials: An overview of basic principles and practical considerations. University of Florida, Horticultural Science Department, IFAS Extension. EDIS 2021, 221, HFS1426. [Google Scholar]
- Montecchio, L. A Venturi effect can help cure our trees. J. Vis. Exp. 2013, 80, e51199. [Google Scholar] [CrossRef] [Green Version]
- Acimovic, S.G.; Cregg, B.M.; Sundin, G.; Wise, J.C. Comparison of drill- and needle-based tree injection technologies in healing of trunk injection ports on apple trees. Urban For. Urban Green 2016, 19, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Scortichini, M.; Loreti, S.; Pucci, N.; Scala, V.; Tatulli, G.; Verweire, D.; Oehl, M.; Widmer, U.; Massana Codina, J.; Hertl, P.; et al. Progress towards a sustainable control of Xylella fastidiosa subsp. pauca in olive groves of Salento (Apulia, Italy). Pathogens 2021, 10, 668. [Google Scholar] [CrossRef]
- Girelli, C.R.; Hussain, M.; Verweire, D.; Oehl, M.; Massana-Codina, J.; Avendano, M.S.; Migoni, D.; Scortichini, M.; Fanizzi, F.P. Agro-active endo-therapy treated Xylella fastidiosa subsp. pauca-infected olive trees assessed by the first 1H-NMR-based metabolomic study. Sci. Rep. 2022, 12, 5973. [Google Scholar]
- Del Frari, G.; Costa, J.; Oliveira, H.; Boavida Ferreira, R. Endotherapy of infected grapevine cuttings for the control of Phaeomoniella chlamydospora and Phaeoacremonium minimum. Phytopathol. Medit. 2018, 57, 439–445. [Google Scholar]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; Van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2017, 63, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, J.R.; Bischoff-Schaefer, M.; Bluemel, S.; Dachbrodt-Saaydeh, S.; Dreux, L.; Jansen, J.-P.; Kiss, J.; Kohl, J.; Kudsk, P.; Malausa, T.; et al. Identifying obstacles and ranking common biological research priorities for Europe to manage most economically important pests in arable, vegetable and perennial crops. Pest Manag. Sci. 2017, 73, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Bonaterra, A.; Badosa, E.; Cabriefiga, J.; Francés, J.; Montesinos, E. Prospects and limitations of microbial pesticides for control of bacterial and fungal pomefruit tree diseases. Trees 2012, 26, 215–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velivelli, S.L.S.; De Vos, P.; Kromann, P.; Declerck, S.; Prestwich, B.D. Biological control agents: From field to market, problems, and challenges. Trends Biotechnol. 2014, 32, 493–496. [Google Scholar] [CrossRef]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P.A. Screening for new biocontrol agents applicable in plant disease management—A review. Biol. Cont. 2020, 144, 104240. [Google Scholar] [CrossRef]
- Stefani, E.; Obradovic, A.; Gasic, K.; Altin, I.; Nagy, I.K.; Kovacs, T. Bacteriophage-mediated control of phytopathogenic xanthomonads: A promising green solution for the future. Microorganisms 2021, 9, 1056. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef]
- Balogh, B.; Nga, N.T.T.; Jones, J.B. Relative level of bacteriophage multiplication in vitro or phyllosphere may not predict in planta efficacy for controlling bacterial leaf spot on tomato caused by Xanthomonas perforans. Front. Microbiol. 2018, 9, 2176. [Google Scholar] [CrossRef] [Green Version]
- Vu, N.T.; Oh, C.-S. Bacteriophage usage for bacterial disease management and diagnosis in plant. Plant Pathol. J. 2020, 36, 204–217. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Wang, X.; Wang, C.; Li, Q.; Zhang, J.; Ji, C.; Sui, J.; Liu, Z.; Song, X.; Liu, X. Isolation and characterization of antagonistic bacteria with the potential for biocontrol of soil-borne wheat disease. Plant Pathol. 2018, 125, 1868–1880. [Google Scholar]
- Legein, M.; Smets, W.; Vandenheuvel, D.; Eilers, T.; Muyshondt, B.; Prinsen, E.; Samson, R.; Leeber, S. Modes of action of microbial biocontrol in the phyllosphere. Front. Microbiol. 2020, 11, 1619. [Google Scholar] [CrossRef]
- Mota, M.S.; Gomes, C.B.; Souza, I.T., Jr.; Bittencourt Moura, A. Bacterial selection for biological control of plant disease: Criterion determination and validation. Braz. J. Microbiol. 2017, 48, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lama Cabanas, C.; Legarda, G.; Ruano-Rosa, D.; Pizarro-Tobias, P.; Valverde-Corredor, A.; Niqui, J.L.; Trivino, J.C.; Roca, A.; Mercado-Blanco, J. Indigenous Pseudomonas spp. strains from the olive (Olea europea L.) rhizosphere as effective biocontrol agents against Verticillium dahliae: From the host roots to the bacterial genome. Front. Microbiol. 2018, 9, 277. [Google Scholar] [CrossRef] [Green Version]
- Salvatierra-Martinez, R.; Arancibia, W.; Araya, M.; Aguilera, S.; Olalde, V.; Bravo, J.; Stoll, A. Colonization ability as an indicator of enhanced biocontrol capacity—An example using two Bacillus amyloliquefaciens strains and Botrytis cinerea infection of tomatoes. J. Phytopathol. 2018, 166, 601–612. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Control of fire blight by Pseudomonas fluorescens A506 and Pantoea vagans C9-1 applied as single strains and mixed inocula. Phytopathology 2010, 100, 1330–1339. [Google Scholar] [CrossRef] [Green Version]
- Couillerot, O.; Prigent-Combaret, C.; Caballero-Mellado, J.; Moenne-Loccoz, Y. Pseudomonas fluorescens and closely related fluorescent pseudomonads as biocontrol agents of soil-borne phytopathogens. Lett. Appl. Microbiol. 2009, 48, 505–512. [Google Scholar] [CrossRef]
- Nam, H.S.; Anderson, A.J.; Kim, Y.C. Biocontrol efficacy of formulated Pseudomonas chlororaphis O6 against plant diseases and root-knot nematodes. Plant Pathol. J. 2018, 34, 241–249. [Google Scholar] [CrossRef]
- Chen, K.; Tian, Z.; He, H.; Long, C.; Jiang, F. Bacillus species as potential biocontrol agents against citrus diseases. Biol. Cont. 2020, 151, 104419. [Google Scholar] [CrossRef]
- Aiello, D.; Leonardi, G.R.; Di Petro, C.; Vitale, A.; Polizzi, G. A new strategy to improve management of Citrus mal secco disease using bioformulates based on Bacillus amyloliquefaciens strains. Plants 2022, 11, 446. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.; Torres, M.; Sampedro, I.; Martinez-Checa, F.; Torres, B.; Béjar, V. Biological control of Verticillium wilt on olive trees by the salt-tolerant strain Bacillus velezensis XT1. Microorganisms 2020, 8, 1080. [Google Scholar] [CrossRef] [PubMed]
- Fira, D.; Dimkic, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Osei, R.; Yang, C.; Cui, L.; Wei, L.; Jin, M.; Wei, X. Antagonistic bioagent mechanisms of controlling potato soft rot. Plant Prot. Sci. 2022, 58, 18–30. [Google Scholar] [CrossRef]
- Law, J.W.-F.; Ser, H.-L.; Khan, T.M.; Chuah, L.-H.; Pusparajah, P.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Chaiharn, M.; Theantana, T.; Pathom-aree, W. Evaluation of biocontrol activities of Streptomyces spp. against rice blast disease fungi. Pathogens 2020, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Daranas, N.; Rossello, G.; Cabrefiga, J.; Donati, I.; Francés, J.; Badosa, E.; Spinelli, F.; Montesinos, E.; Bonaterra, A. Biological control of bacterial plant diseases with Lactobacillus plantarum strains selected for their broad spectrum activity. Ann. Appl. Biol. 2020, 174, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Mohd Taha, M.D.; Mohd Jain, M.F.; Saidi, N.B.; Abdul Rahim, R.; Shah, U.K.; Mohd Hashim, A. Biological control of Erwinia mallotivora, the causal agent of papaya dieback disease by indigenous seed-borne endophytic lactic acid bacteria consortium. PLoS ONE 2019, 14, e0224431. [Google Scholar] [CrossRef]
- Sarrocco, S.; Esteban, P.; Vicente, I.; Bernardi, R.; Plainchamp, T.; Domenichini, S.; Puntoni, G.; Baroncelli, R.; Vannacci, G.; Dufresne, M. Straw competition and wheat root endophytism of Trichoderma gamsii T6085 as useful traits in the biological control of Fusarium head blight. Phytopathology 2021, 111, 1129–1136. [Google Scholar] [CrossRef]
- Tuão, C.A.; Pinto, J.M. Biocontrol of melon wilt caused by Fusarium oxysporum Schlect f. sp. melonis using seed treatment with Trichoderma spp. and liquid compost. Biol. Control 2016, 97, 13–20. [Google Scholar]
- Lahuf, A.A.; Kareem, A.A.; Al-Sweedi, T.M.; Alfarttoosi, H.A. Evaluation the potential of indigenous biocontrol agent Trichoderma harzianum and its interactive effect with nanosized ZnO particles against the sunflower damping-off pathogen, Rhizoctonia Solani. IOP Conf. Ser. Earth Environ. Sci. 2019, 365, 012033. [Google Scholar] [CrossRef]
- Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Altomare, C. Genomic characterization of Trichoderma atrobrunneum (T. harzianum species complex) ITEM 908: Insight into genetic endowment of a multi-target biocontrol strain. BMC Genom. 2018, 19, 662. [Google Scholar] [CrossRef]
- Rush, T.A.; Sherstha, H.K.; Meena, M.G.; Spangler, M.K.; Ellis, J.C.; Labbè, J.L.; Abraham, P.E. Bioprospecting Trichoderma: A systematic roadmap to screen genomes and natural products for biocontrol applications. Front. Fung. Biol. 2021, 2, 716511. [Google Scholar] [CrossRef]
- Niu, B.; Wang, W.; Yuan, Z.; Sederoff, R.R.; Chaing, V.L.; Borriss, R. Microbial interactions within multiple-strain biological control agents impact soil-borne plant disease. Front. Microbiol. 2020, 11, 585404. [Google Scholar] [CrossRef]
- Pertot, I.; Giovannini, O.; Benanchi, M.; Caffi, T.; Rossi, V.; Mugnai, L. Combining biocontrol agents with different mechanism of action in a strategy to control Botrytis cinerea on grapevine. Crop Prot. 2017, 97, 85–93. [Google Scholar] [CrossRef]
- Bubici, G. Streptomyces spp. as biocontrol agents against Fusarium species. CAB Rev. 2018, 13, 050. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Sdiri, Y.; Lopes, T.; Rodrigues, N.; Silva, K.; Rodrigues, I.; Pereira, J.A.; Baptista, P. Biocontrol ability and production of volatile organic compounds as a potential mechanism of action of olive endophytes against Colletotrichum Acutatum. Microorganisms 2022, 10, 571. [Google Scholar] [CrossRef]
- Del Carmen Rodríguez, H.; Evans, H.C.; De Abreu, L.M.; De Macedo, D.M.; Ndacnou, M.K.; Bekele, K.B.; Barreto, R.W. New species and records of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Sci. Rep. 2021, 11, 5671. [Google Scholar]
- Vurucunda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.E.; Park, J.M. Endophytic bacteria as biocontrol agents against plant pathogens: Current state-of-the-art. Plant Biotechnol. Rep. 2016, 10, 353–357. [Google Scholar] [CrossRef]
- Andrivon, D.; Bardin, M.; Bertrand, C.; Brun, L.; Daire, X.; Fabre, F.; Gary, C.; Montarry, J.; Nicot, P.; Reignault, P.; et al. Can organic agriculture cope without copper for disease control? In Synthesis of the Collective Scientific Assessment Report; INRA: Paris, France, 2018; p. 64. [Google Scholar]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop protection. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- Chinnadurai, C.; Ramkissoon, A.; Rajendran, R.; DeAspa, S.; Ramsubhag, A.; Jayaraj, J. Integrated disease management in pumpkin in the southern Caribbean. Trop. Agric. Univ. West Indies 2018, 95, 132–140. [Google Scholar]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahrauoi, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Gato, M.; Astray, G.; Mejuto, J.C.; Simal-Gandara, J. Essential oils as antimicrobials in crop protection. Antibiotics 2021, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Amini, L.; Soudi, M.R.; Saboora, A.; Mobasheri, H. Effect of essential oil from Zataria multiflora on local strains of Xanthomonas campestris: An efficient antimicrobial agent for decontamination of seeds of Brassica oleracea var. capitata. Sci. Hort. 2018, 236, 256–264. [Google Scholar] [CrossRef]
- Kumar, P.; Lokesh, V.; Doddaraju, P.; Kumari, A.; Singh, P.; Meti, B.S.; Sharma, J.; Gupta, K.J.; Manjunatha, G. Greenhouse and field experiments revealed that clove oil can effectively reduce bacterial blight and increase yield in pomegranate. Food Energy Secur. 2021, 10, e305. [Google Scholar] [CrossRef]
- Hashem, M.; Moharam, A.M.; Zaied, A.A.; Saleh, F.E.M. Efficacy of essential oils in the control of cumin root rot disease caused by Fusarium spp. Crop Protect. 2010, 29, 1111–1117. [Google Scholar] [CrossRef]
- Katijar, D.; Hemantaranjan, A.; Singh, B.; Nishant Bhanu, A. A future perspective in crop protection: Chitosan and its oligosaccharides. Adv. Plants Agric. Res. 2014, 1, 23–30. [Google Scholar]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Romanazzi, G.; Feliziani, E.; Sivakumar, D. Chitosan, a biopolymer with triple action on postharvest decay of fruit and vegetables: Eliciting, antimicrobial and film-forming properties. Front. Microbiol. 2018, 9, 2745. [Google Scholar] [CrossRef]
- Scortichini, M. Field efficacy of chitosan to control Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker. Eur. J. Plant Pathol. 2014, 140, 887–892. [Google Scholar]
- Stanley-Raja, V.; Senthil-Nathan, S.; Chantini, K.M.P.; Sivanesh, H.; Ramasubramanian, R.; Karthi, S.; Shyam-Sundar, N.; Vasantha-Srinivasan, P.; Kalaivani, K. Biological activity of chitosan inducing resistance efficiency of rice (Oryza sativa L.) after treatment with fungal based chitosan. Sci. Rep. 2021, 11, 20488. [Google Scholar] [CrossRef]
- Nandeeshkumar, P.; Sudisha, J.; Ramachandra, K.K.; Prakash, H.S.; Niranjana, S.R. Shekar, S.H. Chitosan induced resistance to downy mildew in sunflower caused by Plasmopara halstedii. Physiological. Mol. Plant Pathol. 2008, 72, 188–194. [Google Scholar] [CrossRef]
- Chandrika, K.S.V.P.; Prasad, R.D.; Godbole, V. Development of chitosan-PEG blended films using Thrichoderma: Enhancement of antimicrobial activity and seed quality. Int. J. Biol. Macromol. 2019, 126, 282–290. [Google Scholar] [CrossRef]
- Kappel, L.; Kosa, N.; Gruber, S. The multilateral efficacy of chitosan and Trichoderma on sugar beet. J. Fungi 2022, 8, 137. [Google Scholar] [CrossRef]
- Lafontaine, J.P.; Benhamou, N. Chitosan treatment: An emerging strategy for enhancing resistance of greenhouse tomato plants to infection by Fusarium oxysporum f. sp. radicis lycopersici. Biocontrol Sci. Technol. 1996, 6, 111–124. [Google Scholar] [CrossRef]
- Aloui, H.; Khwaldia, K.; Licciardello, F.; Mazzaglia, A.; Muratore, G.; Hamdi, M.; Restuccia, C. Efficacy of the combined application of chitosan and locust bean gum with different citrus essential oils to control postharvest spoilage caused by Aspergillus flavus in dates. Int. J. Food Microbiol. 2014, 170, 21–28. [Google Scholar] [CrossRef]
- Wullf, E.G.; Zida, E.; Torp, J.; Lund, O.S. Yucca schidigera extract: A potential biofungicide against seedborne pathogens of sorghum. Plant Pathol. 2012, 61, 331–338. [Google Scholar] [CrossRef]
- Trebbi, G.; Negri, L.; Bosi, S.; Dinelli, G.; Cozzo, R.; Marotti, I. Evaluation of Equisetum arvense (horsetail macerate) as a copper substitute for pathogen management in field-grown organic tomato and durum wheat cultivation. Agriculture 2021, 11, 5. [Google Scholar] [CrossRef]
- Quattrucci, A.; Ovidi, E.; Tiezzi, A.; Vinciguerra, V.; Balestra, G.M. Biological control of tomato bacterial speck using Punica granatum fruit peel extract. Crop Prot. 2013, 46, 18–22. [Google Scholar] [CrossRef]
- Mohamad, T.G.; Khalil, A.A. Effect of agriculture waste: Pomegranate (Punica granatum L.) fruits peel on some important phytopathogenic fungi and control of tomato damping-off. J. Appl. Life Sci. Int. 2015, 3, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Li Destri Nicosia, M.G.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of postharvest fungal rots on citrus fruit and sweet cherries using a pomegranate peel extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Lovato, A.; Pignatti, A.; Vitulo, N.; Vandelle, E.; Polverari, A. Inhibition of virulence-related traits in Pseudomonas syringae pv. actinidiae by gunpowder green tea extracts. Front. Microbiol. 2019, 10, 2362. [Google Scholar] [PubMed]
- Liu, H.-W.; Ji, Q.-T.; Ren, G.-G.; Wang, F.; Su, F.; Wang, P.-Y.; Zhou, X.; Wu, Z.-B.; Li, Z.; Yang, S. Antibacterial functions and proposed modes of action of novel 1,2,3,4-tetrahydro-β-carboline derivatives that possess an attractive 1,3-diaminopropan-2-ol pattern against rice bacterial blight, kiwifruit bacterial canker, and citrus bacterial canker. J. Agric. Food. Chem. 2020, 68, 12558–12568. [Google Scholar] [CrossRef] [PubMed]
- Agrios, N.G. Plant Pathology, 5th ed.; Elsevier-Academic Press: Amsterdam, The Netherlands, 2005; p. 635. [Google Scholar]
- Huber, D.M.; Haneklaus, S. Managing nutrition to control plant diseases. Landbauforsch. Völkenrode 2007, 57, 313–322. [Google Scholar]
- Gupta, N.; Debhnat, S.; Sharma, S.; Sharma, P.; Purohit, J. Role of nutrients in controlling the plant diseases in sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer Nature: Singapore, 2017; pp. 217–261. [Google Scholar]
- Mitchell, A.F.; Walters, D.R. Potassium phosphate induces systemic protection in barley to powdery mildew infection. Pest Manag. Sci. 2004, 60, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Yogev, E.; Sadowsky, A.; Solei, Z.; Oren, Y.; Orbach, Y. The performance of potassium phosphite for controlling Alternaria brown spot of citrus fruit. J. Plant Dis. Prot. 2006, 113, 207–213. [Google Scholar] [CrossRef]
- Liljeroth, E.; Lankinen, A.; Andreasson, E.; Alexandersson, E. Phosphite integrated the late blight treatment strategies in starch potato does not cause residue in the starch product. Plant Dis. 2020, 104, 3026–3032. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Vasilakakis, M.; Diamantidis, G.; Migani, I. The effect of postharvest calcium application on tissue calcium concentration, quality attributes, incidence of flesh browning and cell wall physicochemical aspects of peach fruits. Food Chem. 2007, 100, 1385–1392. [Google Scholar] [CrossRef]
- Sun, C.; Zhu, C.; Tang, Y.; Ren, D.; Cai, Y.; Zhou, G.; Wang, Y.; Xu, L.; Zhu, P. Inhibition of Botrytis cinerea and control of gray mold of table grapes by calcium propionate. Food Qual. Saf. 2021, 5, fyab016. [Google Scholar] [CrossRef]
- Huber, D.M.; Jones, J.B. The role of magnesium in plant disease. Plant Soil 2013, 368, 73–85. [Google Scholar] [CrossRef]
- Moreira, W.R.; Bispo, W.M.S.; Rios, J.A.; Debona, D.; Nascimento, C.W.A.; Rodrigues, F.A. Magnesium induced alterations in the photosynthetic performance and resistance of the plants infected with Bipolaris oryzae. Sci. Agric. 2015, 72, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Fagnano, M.; Agrelli, D.; Pascale, A.; Adamo, P.; Fiorentino, N.; Rocco, C.; Pepe, O.; Ventorino, V. Copper accumulation in agricultural soils: Risks for the food chain and the soil microbial populations. Sci. Total Environ. 2020, 734, 139434. [Google Scholar] [CrossRef]
- Pearson, C.J.; Jacobs, B.C. Elongation and retarded growth of rice during short-term submergence in three stages of development. Field Crops Res. 1986, 13, 331–344. [Google Scholar] [CrossRef]
- Navarrete, F.; De La Fuente, L. Zinc detoxification is required for full virulence and modification of the host leaf ionome by Xylella fastidiosa. Mol. Plant Microbe Interact. 2015, 28, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Scortichini, M.; Chen, J.; De Caroli, M.; Dalessandro, G.; Pucci, N.; Modesti, V.; L’Aurora, A.; Petriccione, M.; Zampella, L.; Mastrobuoni, F.; et al. A zinc, copper and citric acid biocomplex shows promise for control of Xylella fastidiosa subsp. pauca in olive trees of Apulia region (southern Italy). Phytopathol. Medit. 2018, 57, 48–72. [Google Scholar]
- Tatulli, G.; Modesti, V.; Pucci, N.; Scala, V.; L’Aurora, A.; Lucchesi, S.; Salustri, M.; Scortichini, M.; Loreti, S. Further in vitro assesment and mid-term evaluation of control strategy of Xylella fastidiosa subsp. pauca in olive groves of Salento (Apulia, Italy). Pathogens 2021, 10, 85. [Google Scholar] [CrossRef]
- Del Coco, L.; Migoni, D.; Girelli, C.R.; Angilè, F.; Scortichini, M.; Fanizzi, F.P. Soil and leaf ionome heterogeneity in Xylella fastidiosa subsp. pauca-infected, non infected and treated olive groves in Italy. Plants 2020, 9, 760. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, R.R.; Tyagi, S.K. Pre-harvest foliar application of calcium and boron influences physiological disorders, fruit yield and quality of strawberry (Fragaria × ananassa Duch.). Sci. Hort. 2007, 112, 215–220. [Google Scholar] [CrossRef]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Gong, D.; Zhu, S.; Zhang, L.; Zhang, L. Expression of PPO and POD genes and contents of polyphenolic compounds in harvested mango fruits in relation to benzathiadiazole-induced defense against anthracnose. Sci. Hort. 2011, 130, 85–89. [Google Scholar] [CrossRef]
- Sillero, J.C.; Rojas-Molina, M.M.; Avila, C.M.; Rubiales, D. Induction of systemic acquired resistance against rust, ascochyta blight and broomrape in faba bean by exogenous application of salicylic acid and benzothiadiazole. Crop Prot. 2012, 34, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Monchiero, M.; Gullino, M.L.; Pugliese, M.; Spadaro, D.; Garibaldi, A. Efficacy of different chemical and biological products in the control of Pseudomonas syringae pv. actinidiae on kiwifruit. Eur. J. Plant Pathol. 2015, 44, 13–23. [Google Scholar]
- Hahm, M.-S.; Sumayo, M.; Hwang, Y.-J.; Jeon, S.-A.; Park, S.-J.; Lee, J.Y.; Ahn, J.-H.; Kim, B.-S.; Ryu, C.-M.; Ghim, S.-Y. Biological control and plant growth promoting capacity of rhizobacteria on pepper under greenhouse and field conditions. J. Microbiol. 2012, 50, 380–385. [Google Scholar] [CrossRef]
- Tortora, M.L.; Díaz-Ricci, J.C.; Pedraza, R.O. Protection of strawberry plants (Fragaria ananassa Duch.) against anthracnose disease induced by Azospirillum brasilense. Plant Soil 2012, 356, 279–290. [Google Scholar] [CrossRef]
- Cohen, Y.; Rubin, A.E.; Kilfin, G. Mechanisms of induced resistance in lettuce against Bremia lactucae by DL-β-amino-butyric acid (BABA). Eur. J. Plant Pathol. 2010, 126, 553–573. [Google Scholar] [CrossRef]
- Tamm, L.; Thürig, B.; Fliessbach, A.; Goltlieb, A.E.; Karavani, S.; Cohen, Y. Elicitors and soil management to induce resistance against fungal plant diseases. NJAS Wagening. J. Life Sci. 2011, 58, 131–137. [Google Scholar] [CrossRef]
- Harel, Y.M.; Elad, Y.; Rav-David, D.; Borenstein, M.; Shulchani, R.; Lew, B.; Graber, E.R. Biochar mediates systemic response of strawberry to foliar fungal pathogens. Plant Soil 2012, 357, 245–257. [Google Scholar] [CrossRef]
- Faggioli, F.; Luigi, M.; Mangili, A.; Dragone, I.; Antonelli, M.G.; Contarini, M.; Speranza, S.; Bertin, S.; Tiberini, A.; Gentili, A. Effects of biochar on the growth and development of tomato seedlings, and on the response of tomato plants to the infection of systemic viral agents. Front. Microbiol. 2022, 13, 862075. [Google Scholar]
- Paulert, R.; Talamini, V.; Cassolato, J.E.F.; Duarte, M.E.R.; Noseda, M.D.; Smania, A.; Stadnik, M.J., Jr. Effects of sulphated polysaccharide and alcoholic extracts from green seaweed Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Protect. 2009, 116, 263–270. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.-F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Koch, A.; Kogel, K.-H. New wind in the sails: Improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol. J. 2014, 12, 821–831. [Google Scholar] [CrossRef]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA interference: A natural immune system of plants to counteract biotic stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Gebremichael, D.E.; Mehari Haile, Z.; Negrini, F.; Sabbadini, S.; Capriotti, L.; Mezzetti, B.; Baraldi, E. RNA interference strategies for future management of plant pathogenic fungi: Prospects and challenges. Plants 2021, 10, 650. [Google Scholar] [CrossRef]
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Sanchez, J.N.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of the pathogen RNA uptake. Plant Biotechnol. J. 2021, 19, 1756–1768. [Google Scholar] [CrossRef]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abtellatef, E.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Goodfellow, S.; Zhang, D.; Wang, M.-B.; Zhang, R. Bacterium-mediated RNA interference: Potential application in plant protection. Plants 2019, 8, 572. [Google Scholar] [CrossRef] [Green Version]
- Werner, B.T.; Gaffar, F.Y.; Schuemann, J.; Biedenkopf, D.; Koch, A.M. RNA-spray-mediated silencing of Fusarium graminearum AGO and DCL genes improve barley disease resistance. Front. Plant Sci. 2020, 11, 476. [Google Scholar] [CrossRef]
- Jahan, S.N.; Asman, A.K.M.; Corcoran, P.; Fogelqvist, J.; Vetukuri, R.R.; Dixelius, C. Plant-mediated gene silencing restricts growth of the potato late blight pathogen Phytophthora infestans. J. Exp. Bot. 2015, 66, 2785–2794. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.-M.; Thomma, B.; Huang, H.-D.; Jin, H. Bidirection cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- Nerva, L.; Sandrini, M.; Gambino, G.; Chitarra, W. Double-stranded RNAs (dsRNAs) as a sustainable tool against gray mold (Botrytis cinerea) in grapevine: Effectiveness of different application methods in an open-air environment. Biomolecules 2020, 10, 200. [Google Scholar] [CrossRef] [Green Version]
- Legreve, A.; Duveiller, E. Preventing potential diseases and pest epidemics under a changing climate. In Climate Change and Crop Production; Reynolds, M.P., Ed.; CABI Publishing: Wallingford, UK, 2010; pp. 50–70. [Google Scholar]
- Rippa, M.; Battaglia, V.; Cermola, M.; Sicignano, M.; Lahoz, E.; Mormile, P. Monitoring the copper persistence on plant leaves using pulsed thermography. Environ. Minit. Assess. 2022, 194, 160. [Google Scholar] [CrossRef] [PubMed]
- Wisniewsky, M.; Wilson, C.; Droby, S.; Chalutz, E.; El-Ghaouth, A.; Stevens, C. Postharvest biocontrol: New concepts and applications. In Biological Control: A Global Perspective; Vincent, C., Goettel, M.S., Lazarovitis, C., Eds.; CABI: Wallingford, UK, 2007; pp. 262–273. [Google Scholar]
- Brouwer, H.; Woodhill, J.; Hemmati, M.; Verhoosel, K.; Van Vugt, S. The MSP Guides: How to Design and Facilitate Multistakeholder Partnerships; Centre for Development and Innovation: Wageningen, The Netherlands, 2015. [Google Scholar]
- Pancino, B.; Blasi, E.; Rappoldt, A.; Pascucci, S.; Ruini, L.; Ronchi, C. Partnering for sustainability in agri-food supply chains: The case of Barilla sustainable farming in the Po valley. Agric. Food. Econ. 2019, 7, 13. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Synoptic panel that shows the current strategies and policies related to the achievement of sustainable disease control in horticulture.
Figure 1.
Synoptic panel that shows the current strategies and policies related to the achievement of sustainable disease control in horticulture.

Figure 2.
Scheme of a modern decision support system based on a forecasting model for plant disease management. Information about crop-specific characteristics (A1), environmental conditions (A2), crop and plant status (A3), and agricultural operations (A4) flows from the crop to a remote server (B1), and it is stored in database (B2). This information is then used as an input for running mathematical models and decision algorithms (B3), which generate decision supports and alerts to the grower for deciding when and how apply a protective agent (C1). (Reproduced from [47]).
Figure 2.
Scheme of a modern decision support system based on a forecasting model for plant disease management. Information about crop-specific characteristics (A1), environmental conditions (A2), crop and plant status (A3), and agricultural operations (A4) flows from the crop to a remote server (B1), and it is stored in database (B2). This information is then used as an input for running mathematical models and decision algorithms (B3), which generate decision supports and alerts to the grower for deciding when and how apply a protective agent (C1). (Reproduced from [47]).

Figure 3.
Synoptic panel that illustrates the developing disease control strategies for sustainable agriculture.
Figure 3.
Synoptic panel that illustrates the developing disease control strategies for sustainable agriculture.

Figure 4.
Olive Verticillium wilt, caused by Verticillim dahliae, treated with Bacillus velezensis: olive trees in the field experiment before and after the treatment with B. velezensis strain XT1. (Reproduced from [158]).
Figure 4.
Olive Verticillium wilt, caused by Verticillim dahliae, treated with Bacillus velezensis: olive trees in the field experiment before and after the treatment with B. velezensis strain XT1. (Reproduced from [158]).

Figure 5.
Effect of the treatments with lactobacilli strains (grey bars) on Pseudomonas syringae pv. actinidiae, Xanthomonas arboricola pv. pruni, and Xanthomonas fragariae infections in kiwifruit, Prunus, and strawberry plants, respectively, under greenhouse conditions. The effect of strains on disease incidence (%) was compared with streptomycin (white bars) and a non-treated control (black bars). Different letters indicate statistical significance (Reproduced from [163]).
Figure 5.
Effect of the treatments with lactobacilli strains (grey bars) on Pseudomonas syringae pv. actinidiae, Xanthomonas arboricola pv. pruni, and Xanthomonas fragariae infections in kiwifruit, Prunus, and strawberry plants, respectively, under greenhouse conditions. The effect of strains on disease incidence (%) was compared with streptomycin (white bars) and a non-treated control (black bars). Different letters indicate statistical significance (Reproduced from [163]).

Figure 6.
Two olive groves in Nardò (Lecce province, Apulia, Italy): the trees on the right were abandoned upon the infection by Xylella fastidiosa subsp. pauca, whereas the trees on the left were treated with the zinc–copper–citric acid biocomplex described in [216,217]. The treated trees continue to yield despite the occurrence of the pathogen in the surrounding areas.
Figure 6.
Two olive groves in Nardò (Lecce province, Apulia, Italy): the trees on the right were abandoned upon the infection by Xylella fastidiosa subsp. pauca, whereas the trees on the left were treated with the zinc–copper–citric acid biocomplex described in [216,217]. The treated trees continue to yield despite the occurrence of the pathogen in the surrounding areas.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scortichini, M. Sustainable Management of Diseases in Horticulture: Conventional and New Options. Horticulturae 2022, 8, 517. https://doi.org/10.3390/horticulturae8060517
AMA Style
Scortichini M. Sustainable Management of Diseases in Horticulture: Conventional and New Options. Horticulturae. 2022; 8(6):517. https://doi.org/10.3390/horticulturae8060517
Chicago/Turabian StyleScortichini, Marco. 2022. "Sustainable Management of Diseases in Horticulture: Conventional and New Options" Horticulturae 8, no. 6: 517. https://doi.org/10.3390/horticulturae8060517
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.