
Biocontrol Potential of Endophytic Actinobacteria against Fusarium solani, the Causal Agent of Sudden Decline Syndrome on Date Palm in the UAE

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Culture
2.2. Isolation of Endophytic Actinobacteria from Date Palm Roots
2.3. Screening of Biological Control Agents (BCAs) for the Production of CWDEs
2.4. Screening of BCAs for the Production of Diffusible Antifungal Metabolites
2.5. Bioassays for the Production of Volatile Antifungal Compounds, Siderophores, and Hydrogen Cyanide (HCN)
2.6. Determination of the Population Densities of Endophytic Actinobacteria Isolated from Date Palm
2.7. Assays of Antifungal Metabolites and CWDEs Activities
2.8. Evaluation of Crude Culture Filtrates of BCA on F. solani
2.9. Identification of the BCAs
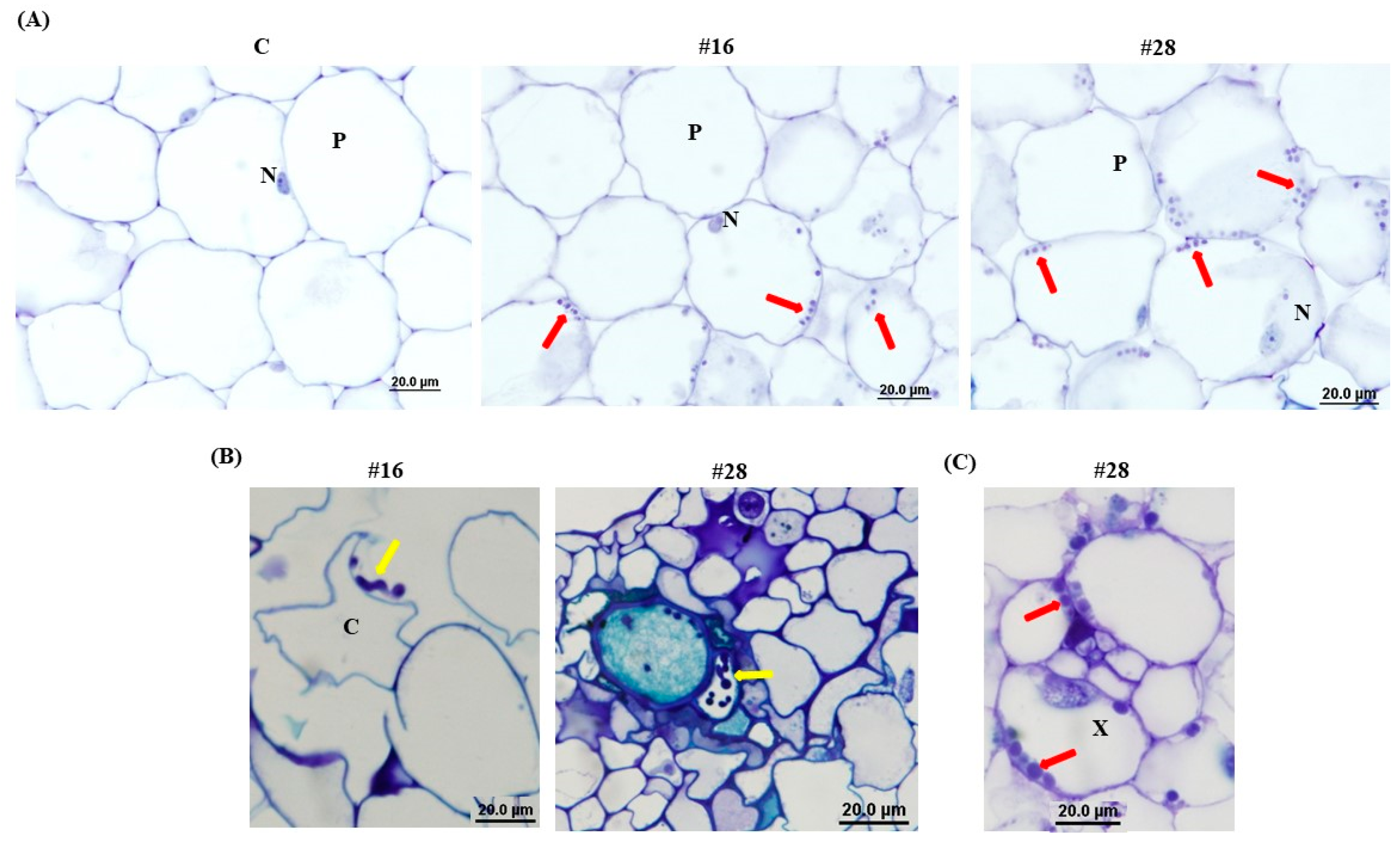
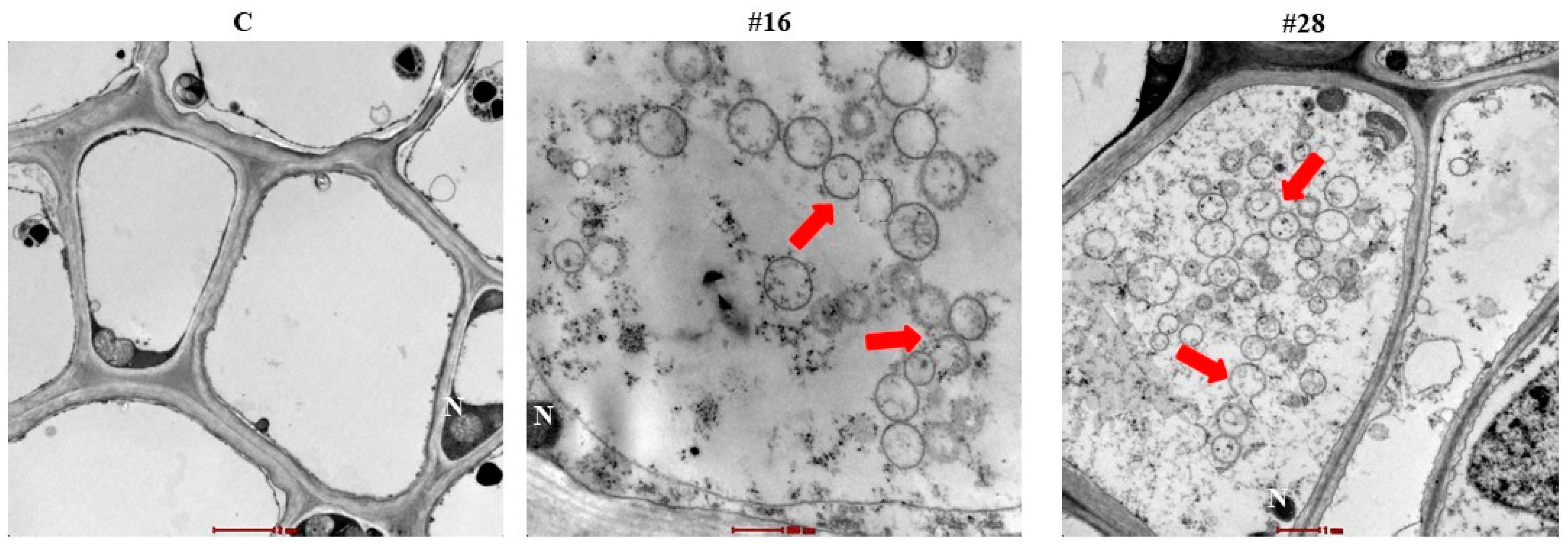
2.10. Light and Transmission Electron Microscopy
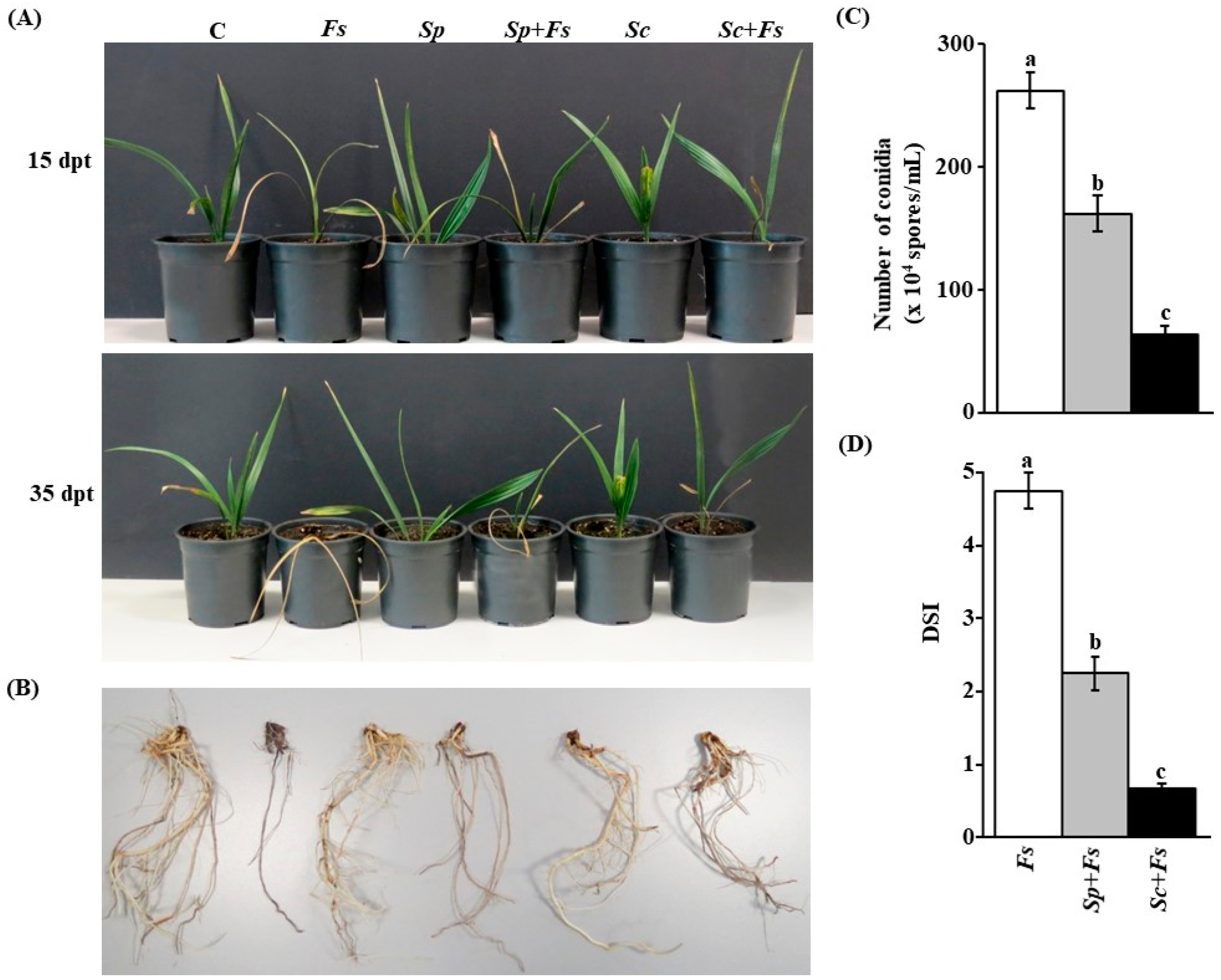
2.11. Disease Assays, Spore Counts, and Disease Severity Index (DSI) in the Greenhouse
- (i).
- Healthy controls (C): Non-inoculated seedlings with any of the BCAs or F. solani;
- (ii).
- Diseased controls (Fs): Inoculated-seedlings with F. solani only;
- (ii).
- BCA controls (Sp or Sc): Inoculated-seedlings with either S. polychromogenes UAE2 (isolate #16) (BCA1) or S. coeruleoprunus UAE1 (isolate #28) (BCA2) only; and
- (iv).
- Sp + Fs or Sc + Fs: Inoculated-seedlings with S. polychromogenes UAE2 (isolate #16) (BCA1) or S. coeruleoprunus UAE1 (isolate #28) (BCA2) at 5 days before F. solani inoculation.
2.12. Statistical Analyses
3. Results
3.1. In vitro Screening of Endophytic Actinobacteria for CWDEs Activities and Antibiosis on F. solani
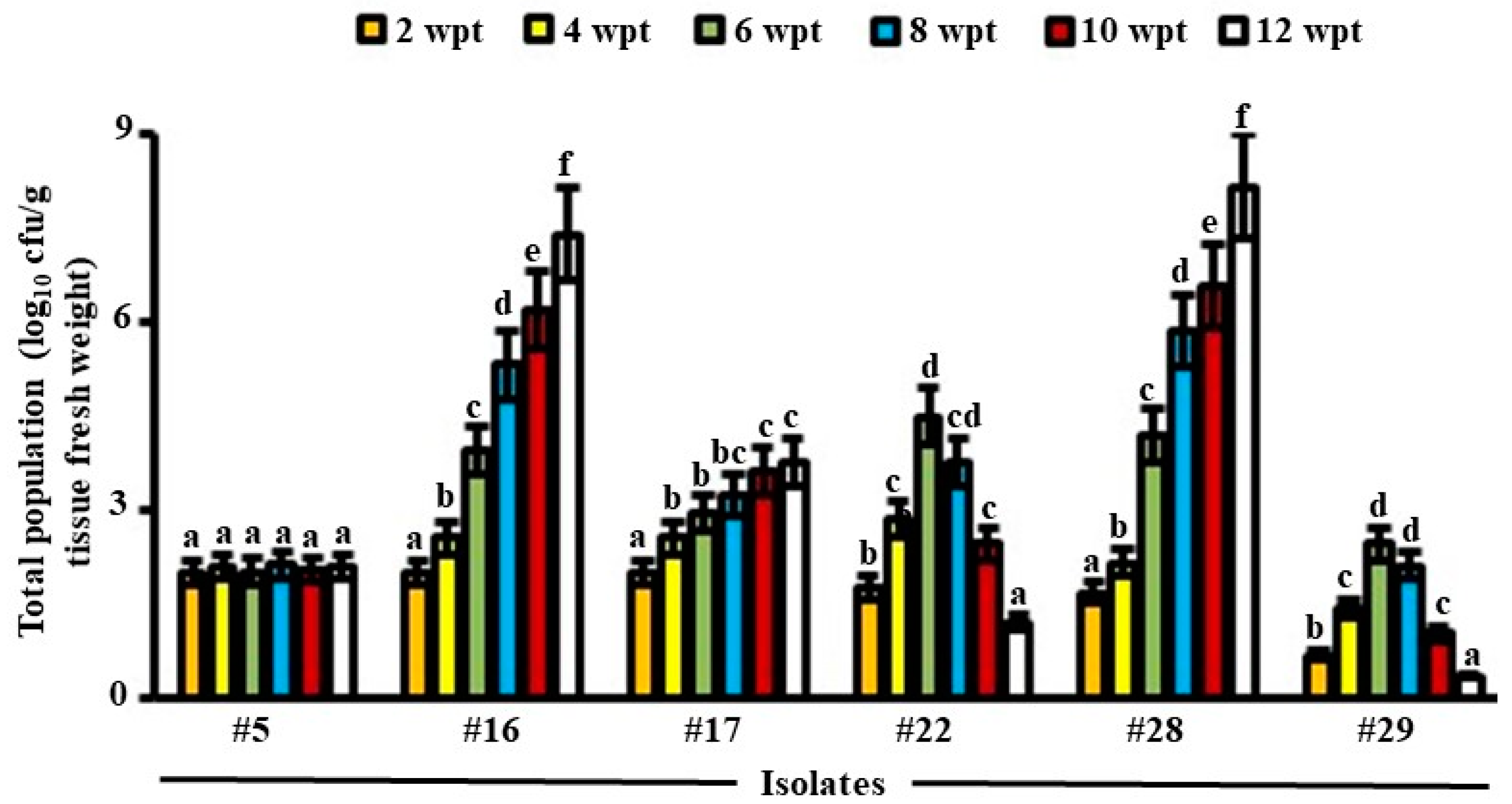
3.2. Total Population of Endophytic Candidates of BCA in Date Palm Roots
3.3. In vitro Evaluation of Antagonistic Properties of the BCA
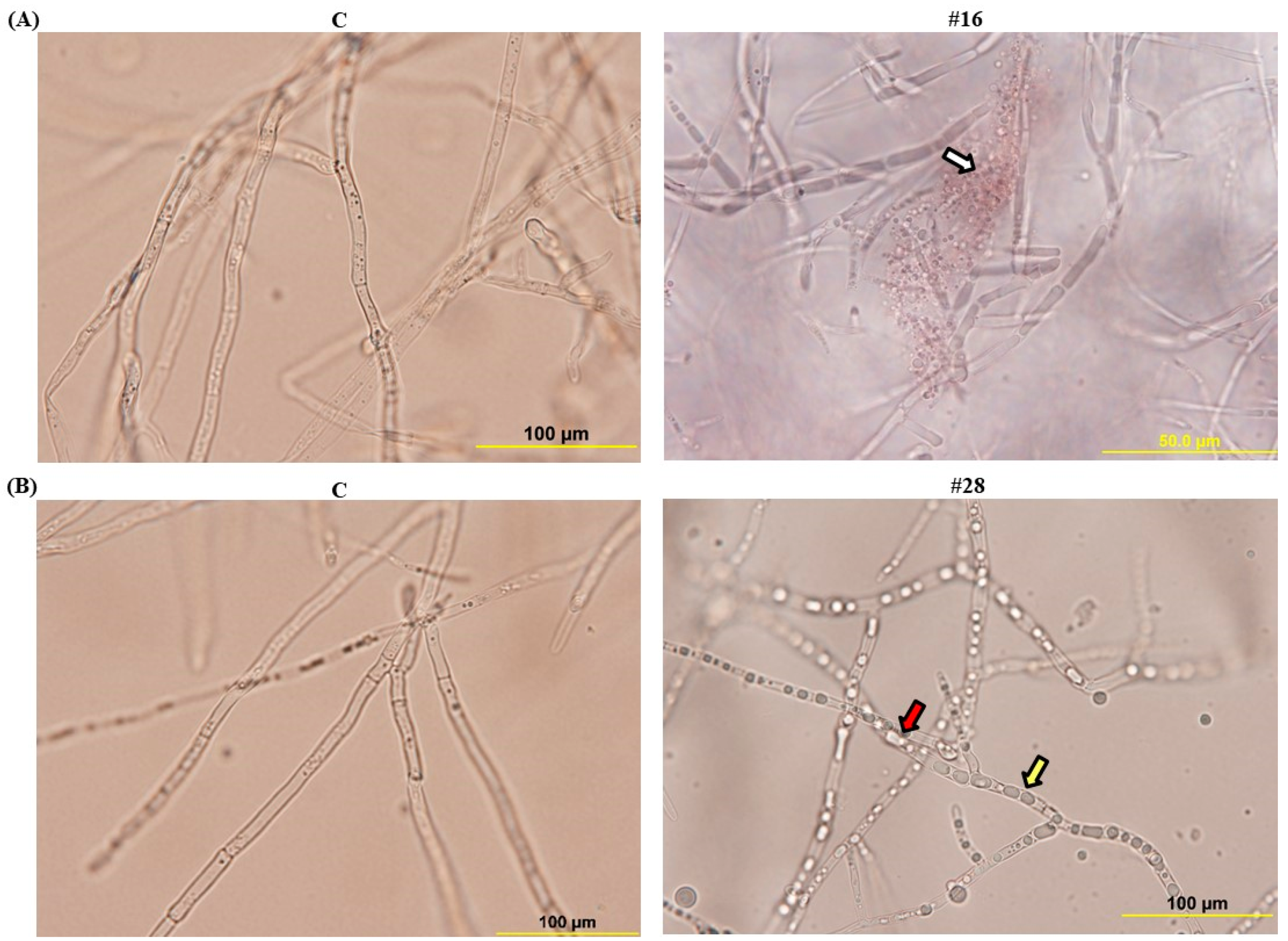
3.4. Effect of BCA Candidates on Hyphal and Cytoplasmic Integrity of F. solani
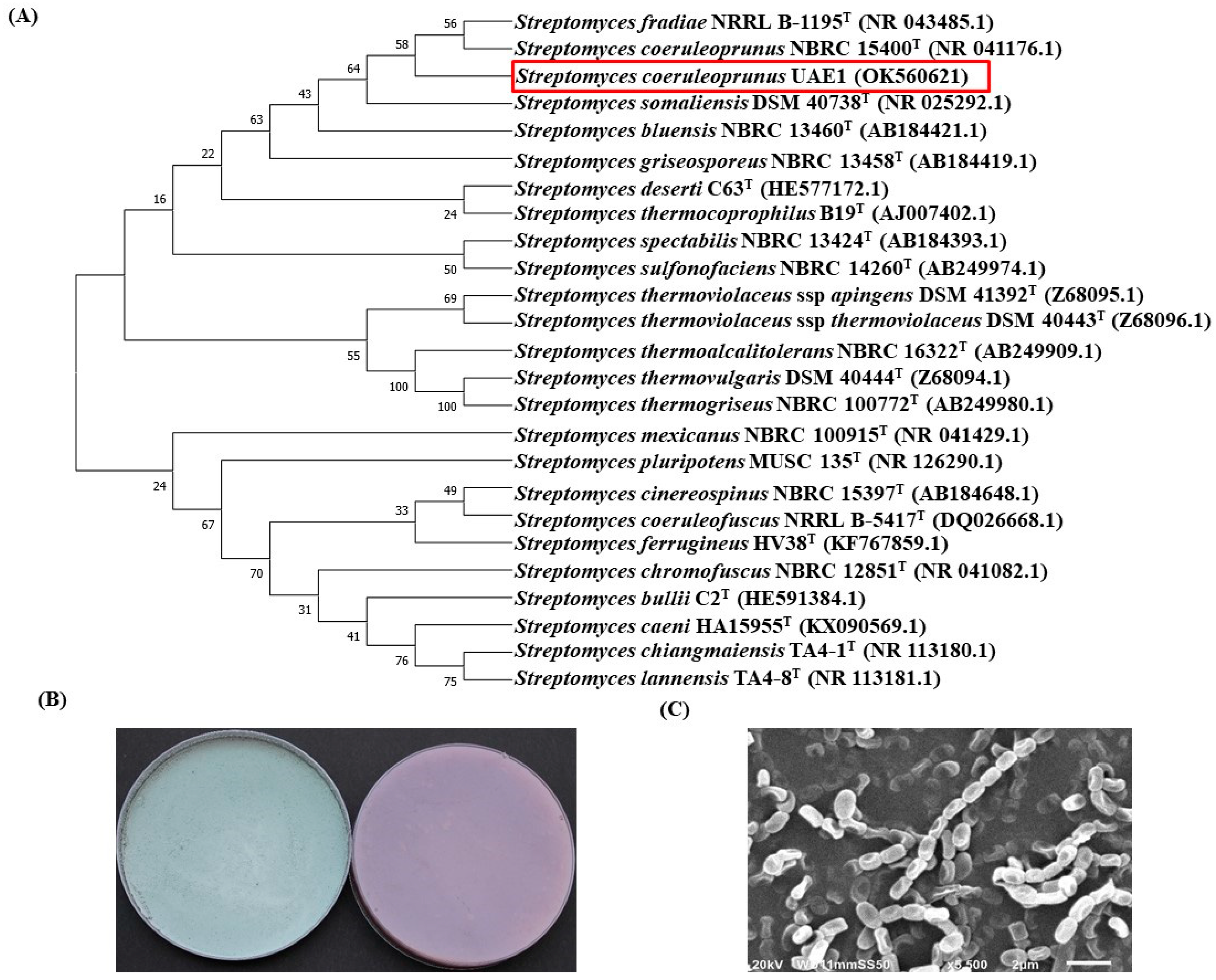
3.5. Identification of the BCA Candidates to the Species Level
3.6. Anatgonism of the Tested Streptomyces spp. on F. solani under Greenhouse Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeed, E.E.; Sham, A.; El-Tarabily, K.A.; Abu Elsamen, F.; Iratni, R.; AbuQamar, S.F. Chemical control of black scorch disease on date palm caused by the fungal pathogen, Thielaviopsis punctulata in United Arab Emirates. Plant Dis. 2016, 100, 2370–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, E.E.; Sham, A.; AbuZarqa, A.; Al Shurafa, K.A.; Al Naqbi, T.S.; Iratni, R.; El-Tarabily, K.; AbuQamar, S.F. Detection and management of mango dieback disease in the United Arab Emirates. Int. J. Mol. Sci. 2017, 18, 2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Raish, S.M.; Saeed, E.E.; Sham, A.; Alblooshi, K.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular characterization and disease control of stem canker on royal poinciana (Delonix regia) caused by Neoscytalidium dimidiatum in the United Arab Emirates. Int. J. Mol. Sci. 2020, 21, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behlau, F. An overview of citrus canker in Brazil. Trop. Plant Pathol. 2021, 46, 1–12. [Google Scholar] [CrossRef]
- Saeed, E.E.; Sham, A.; Salmin, Z.; Abdelmowla, Y.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Streptomyces globosus UAE1, a potential effective biocontrol agent for black scorch disease in date palm plantations. Front. Microbiol. 2017, 8, 1455. [Google Scholar] [CrossRef]
- Kamil, F.H.; Saeed, E.E.; El-Tarabily, K.A.; AbuQamar, S.F. Biological control of mango dieback disease caused by Lasiodiplodia theobromae using streptomycete and non-streptomycete actinobacteria in the United Arab Emirates. Front. Microbiol. 2018, 9, 829. [Google Scholar] [CrossRef]
- Zapata-Sarmientoa, D.H.; Palacios-Palaa, E.F.; Rodríguez-Hernándezb, A.A.; Melchora, D.L.M.; Rodríguez-Monroya, M.; SepúlvedaJiménez, G. Trichoderma asperellum, a potential biological control agent of Stemphylium vesicarium, on onion (Allium cepa L.). Biol. Control 2019, 140, 104105. [Google Scholar] [CrossRef]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary metabolites of actinomycetes and their antibacterial, antifungal and antiviral properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Valli, S.; Suvathi, S.S.; Aysha, O.S.; Nirmala, P.; Vinoth, K.P.; Reena, A. Antimicrobial potential of actinomycetes species isolated from marine environment. Asian Pac. J. Trop. Biomed. 2012, 2, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Al-Ansari, M.; Alkubaisi, N.; Vijayaragavan, P.; Murugan, K. Antimicrobial potential of Streptomyces sp. to the Gram positive and Gram negative pathogens. J. Infect. Public Heal. 2019, 12, 861–866. [Google Scholar] [CrossRef]
- Maiti, P.K.; Das, S.; Sahoo, P.; Mandal, S. Streptomyces sp. SM01 isolated from Indian soil produces a novel antibiotic picolinamycin effective against multi drug resistant bacterial strains. Sci. Rep. 2020, 10, 10092. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; Nassar, A.H.; Hardy, G.E.S.J.; Sivasithamparam, K. Plant growth promotion and biological control of Pythium aphanidermatum, a pathogen of cucumber, by endophytic actinomycetes. J. Appl. Microbiol. 2009, 106, 13–26. [Google Scholar] [CrossRef] [PubMed]
- El-Tarabily, K.A.; ElBaghdady, K.Z.; AlKhajeh, A.S.; Ayyash, M.M.; Aljneibi, R.S.; El-Keblawy, A.; AbuQamar, S.F. Polyamine-producing actinobacteria enhance biomass production and seed yield in Salicornia bigelovii. Biol. Fertil. Soils 2020, 56, 499–519. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Ramadan, G.A.; Elbadawi, A.A.; Hassan, A.H.; Tariq, S.; Ghazal, E.W.; Abo Gamar, M.I.; AbuQamar, S.F. The marine endophytic polyamine-producing Streptomyces mutabilis UAE1 isolated from extreme niches in the Arabian Gulf promotes the performance of mangrove (Avicennia marina) seedlings under greenhouse conditions. Front. Mar. Sci. 2021, 8, 710200. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Sham, A.; Elbadawi, A.A.; Hassan, A.H.; Alhosani, B.K.K.; El-Esawi, M.A.; AbuQamar, S.F. A consortium of rhizosphere-competent actinobacteria with multiple plant growth promoting traits contributes to improve growth and performance of Avicennia marina. Front. Mar. Sci. 2021, 8, 715123. [Google Scholar] [CrossRef]
- Al Hamad, B.M.; Al Raish, S.M.; Ramadan, G.A.; Saeed, E.E.; Alameri, S.S.A.; Al Senaani, S.S.; AbuQamar, S.F.; El-Tarabily, K.A. Effectiveness of augmentative biological control of Streptomyces griseorubens UAE2 depends on 1-aminocyclopropane-1-carboxylic acid deaminase activity against Neoscytalidium dimidiatum. J. Fungi 2021, 7, 885. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Hardy, G.E.S.J.; Sivasithamparam, K.; Hussein, A.M.; Kurtböke, D.I. The potential for the biological control of cavity spot disease of carrots caused by Pythium coloratum by streptomycete and non-streptomycete actinomycetes in Western Australia. New Phytol. 1997, 137, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J.; Baker, K.F. The Nature and Practice of Biological Control of Plant Pathogens; APS Press: St. Paul, MN, USA, 1983. [Google Scholar]
- Martínez-Hidalgo, P.; García, J.M.; Pozo, M.J. Induced systemic resistance against Botrytis cinerea by Micromonospora strains isolated from root nodules. Front. Microbiol. 2015, 6, 922. [Google Scholar] [CrossRef] [Green Version]
- Ayswaria, R.; Vasu, V.; Krishna, R. Diverse endophytic Streptomyces species with dynamic metabolites and their meritorious applications: A critical review. Crit. Rev. Microbiol. 2000, 46, 750–758. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Nassar, A.H.; Sivasithamparam, K. Promotion of growth of bean (Phaseolus vulgaris L.) in a calcareous soil by a phosphate-solubilizing, rhizosphere-competent isolate of Micromonospora endolithica. Appl. Soil Ecol. 2008, 39, 161–171. [Google Scholar] [CrossRef]
- Williams, S.T.; Wellington, E.M.H. Actinomycetes. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keency, O.R., Eds.; American Society of Agronomy/Soil Science Society of America: Madison, WI, USA, 1982; pp. 969–987. [Google Scholar]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Rothrock, C.S.; Gottlieb, D. Role of antibiosis in antagonism of Streptomyces hygroscopicus var. geldanus to Rhizoctonia solani in soil. Can. J. Microbiol. 1984, 30, 1440–1447. [Google Scholar] [CrossRef]
- Xiao, K.; Kinkel, L.L.; Samac, D.A. Biological control of Phytophthora root rots on alfalfa and soybean with Streptomyces. Biol. Control 2002, 23, 285–295. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Davide Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoudi, H.; Hosseininia, G.; Azadi, H.; Fatemi, M. Enhancing date palm processing, marketing and pest control through organic culture. J. Org. Syst. 2008, 3, 29–39. [Google Scholar]
- Alhammadi, M.S.; Al-Shariqi, R.; Maharachchikumbura, S.; Al-Sadi, A.M. Molecular identification of fungal pathogens associated with date palm root diseases in the United Arab Emirates. J. Plant Pathol. 2019, 99, 141–147. [Google Scholar] [CrossRef]
- Alwahshi, K.J.; Saeed, E.E.; Sham, A.; Alblooshi, A.A.; Alblooshi, M.M.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular identification and disease management of date palm sudden decline syndrome in the United Arab Emirates. Int. J. Mol. Sci. 2019, 20, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardi, P.; Saracchi, M.; Quaroni, S.; Petrolini, B.; Borgonovi, G.E.; Merli, S. Isolation of endophytic Streptomyces strains from surface-sterilized roots. Appl. Environ. Microbiol. 1992, 58, 2691–2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennie, R.J.; De Freitas, J.R.; Ruschel, A.P.; Vose, P.B. Isolation and identification of N2-fixing bacteria associated with sugar cane (Saccharum sp.). Can. J. Microbiol. 1982, 28, 462–467. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Küster, E. Outline of a comparative study of criteria used in characterization of the actinomycetes. Int. Bull. Bact. Nomen. Taxon. 1959, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Cross, T. Growth and examination of actinomycete-some guidelines. In Bergey’s Manual of Systematic Bacteriology; Williams, S.T., Sharpe, M.E., Holt, J.G., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1989; pp. 2340–2343. [Google Scholar]
- Wellington, E.M.H.; Williams, S.T. Preservation of actinomycete inoculum in frozen glycerol. Microbios Lett. 1977, 6, 151–157. [Google Scholar]
- Valois, D.; Fayad, K.; Barbasubiye, T.; Garon, M.; Dery, C.; Brzezinski, R.; Beaulieu, C. Glucanolytic actinomycetes antagonistic to Phytophthora fragariae var. rubi, the causal agent of raspberry root rot. Appl. Environ. Microbiol. 1996, 62, 1630–1635. [Google Scholar]
- Gupta, R.; Saxena, R.K.; Chaturvedi, P.; Virdi, J.S. Chitinase production by Streptomyces viridificans: Its potential in fungal cell wall lysis. J. Appl. Bacteriol. 1995, 78, 378–383. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered chitin agar as a selective medium for enumeration of actinomycetes in water and soil. Appl. Microbiol. 1975, 29, 422–426. [Google Scholar] [CrossRef]
- Pridham, T.G.; Lindenfelser, L.A.; Shotwell, O.L.; Stodola, F.H.; Benedict, R.G.; Foley, C.; Jacksom, P.W.; Zaumeyer, W.J.; Perston, W.H.; Mitchell, J.W. Antibiotics against plant disease. I. Laboratory and greenhouse survey. Phytopathology 1956, 46, 568–575. [Google Scholar]
- Payne, C.; Bruce, A.; Staines, H. Yeast and bacteria as biological control agents against fungal discolouration of Pinus sylvestris blocks in laboratory-based tests and the role of antifungal volatiles. Holzforschung 2000, 54, 563–569. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp-mediated plant-growth stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Castric, P.A. Hydrogen cyanide, a secondary metabolite of Pseudomonas aeruginosa. Can. J. Microbiol. 1975, 21, 613–618. [Google Scholar] [CrossRef]
- Misaghi, I.J.; Donndelinger, I.J. Endophytic bacteria in symptom-free cotton plants. Phytopathology 1990, 80, 808–811. [Google Scholar] [CrossRef] [Green Version]
- El-Tarabily, K.A.; AlKhajeh, A.S.; Ayyash, M.M.; Alnuaimi, L.H.; Sham, A.; ElBaghdady, K.Z.; Tariq, S.; AbuQamar, S.F. Growth promotion of Salicornia bigelovii by Micromonospora chalcea UAE1, an endophytic 1-aminocyclopropane-1-carboxylic acid deaminase-producing actinobacterial isolate. Front. Microbiol. 2019, 10, 1694. [Google Scholar] [CrossRef] [PubMed]
- Musson, G.; McInroy, J.A.; Kloepper, J.W. Development of delivery systems for introducing endophytic bacteria into cotton. Biocontrol Sci. Technol. 1995, 5, 407–416. [Google Scholar] [CrossRef]
- Bacharach, A.L.; Cuthbertson, W.F.J. The cup-plate method in microbiological assay, with special reference to riboflavine and aneurine. Analyst 1948, 73, 334–340. [Google Scholar] [CrossRef]
- Gibbs, J.N. A study of the epiphytic growth habit of Fomes annosus. Ann. Bot. 1967, 31, 755–774. [Google Scholar] [CrossRef]
- Tweddell, R.J.; Jabaji-Hare, S.H.; Charest, P.M. Production of chitinases and ß-1,3-glucanases by Stachybotrys elegans, a mycoparasite of Rhizoctonia solani. Appl. Environ. Microbiol. 1994, 60, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.P.; Shin, Y.C.; Park, C.S.; Chung, Y.R. Biological control of Fusarium wilt of cucumber by chitinolytic bacteria. Phytopathology 1999, 89, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissig, J.L.; Strominger, J.L.; Leloir, L.F. A modified colorimetric method for the estimation of N-acetylamino sugars. J. Biol. Chem. 1955, 27, 959–966. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Lorito, M.; Di Pietro, A.; Hayes, C.K.; Woo, S.L.; Harman, G.E. Antifungal, synergistic interaction between chitinolytic enzymes from Trichoderma harzianum and Enterobacter cloacae. Phytopathology 1993, 83, 721–728. [Google Scholar] [CrossRef]
- Sneh, B. Use of rhizosphere chitinolytic bacteria for biological control of Fusarium oxysporum f. sp. dianthi in carnation. J. Phytopathol. 1981, 100, 251–256. [Google Scholar] [CrossRef]
- Locci, R. Streptomycetes and related genera. In Bergey’s Manual of Systematic Bacteriology; Williams, S.T., Sharpe, M.E., Holt, J.G., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1989; pp. 2451–2508. [Google Scholar]
- Rainey, F.A.; Ward-Rainey, N.; Kroppenstedt, R.M.; Stackebrandt, E. The genus Nocardiopsis represents a phylogenetically coherent taxon and a distinct actinomycete lineage: Proposal of Nocardiopsaceae fam. nov. Int. J. Syst. Bacteriol. 1996, 46, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogentic trees. Mol. Biol. Evol. 1987, 4, 406–525. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Millonig, G. Laboratory Manual of Biological Electron Microscopy; Saviolo, M., Ed.; Tipografia Edizioni Saviolo: Vercelli, Italy, 1976; pp. 1–67. [Google Scholar]
- Molan, Y.Y.; Al-Obeed, R.S.; Harhash, M.M.; El-Husseini, S. Decline of date-palm offshoots with Chalara paradoxa in Riyadh region. J. King Saud Univ. 2004, 16, 79–86. [Google Scholar]
- Hagemann, G.; Penasse, L.; Teillon, J. Zur Systematik der Actinomyceten. 9. Streptomyceten mit cinnamoneus Luftmycel. In Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg., Volume 117. Hutter, Ed.; Abteilung II. 1964; 603–661. [Google Scholar]
- Gauze, G.F.; Preobrazhenskaya, T.P.; Sveshnikova, M.A.; Terekhova, L.P.; Maximova, T.S. A Guide for the Determination of Actinomycetes. Genera Streptomyces, Streptoverticillium, and Chainia; Nauka: Moscow, Russia, 1983. [Google Scholar]
- Mengiste, T.; Laluk, K.; AbuQamar, S. Mechanisms of induced resistance against B. cinerea; Chapter 2. In Post-Harvest Pathology: Volume 2; Prusky, D., Gullino, M.L., Eds.; Springer Science + Business Media: Berlin, Germany, 2010; pp. 13–30. [Google Scholar]
- Sham, A.; Moustafa, K.; Al-Shamisi, S.; Alyan, S.; Iratni, R.; AbuQamar, S.F. Microarray analysis of Arabidopsis WRKY33 mutants in response to the necrotrophic fungus Botrytis cinerea. PLoS ONE 2017, 12, e0172343. [Google Scholar] [CrossRef]
- Sham, A.; Al-Ashram, H.; Whitley, K.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Metatranscriptomic analysis of multiple environmental stresses identifies RAP2.4 gene associated with Arabidopsis Immunity to Botrytis cinerea. Sci. Rep. 2019, 9, 17010. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Jebakumar, S.R.D. Non-streptomycete actinomycetes nourish the current microbial antibiotic drug discovery. Front. Microbiol. 2013, 4, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.W. Effects of actinobacteria on plant disease suppression and growth promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control. 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Al Raish, S.M.; Saeed, E.E.; Alyafei, D.M.; El-Tarabily, K.A.; AbuQamar, S.F. Evaluation of streptomycete actinobacterial isolates as biocontrol agents against royal poinciana stem canker disease caused by Neoscytalidium dimidiatum. Biol. Control. 2021, 164, 104783. [Google Scholar] [CrossRef]
- Srivastava, D.A.; Harris, R.; Breuer, G.; Levy, M. Secretion-based modes of action of biocontrol agents with a focus on Pseudozyma aphidis. Plants 2021, 10, 210. [Google Scholar] [CrossRef]
- Osherov, N.; Yarden, O. The cell wall of filamentous fungi. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K., Ebbole, D., Eds.; ASM Press: Washington, DC, USA, 2010; pp. 224–237. [Google Scholar]
- Berini, F.; Katz, C.; Gruzdev, N.; Casartelli, M.; Tettamanti, G.; Marinelli, F. Microbial and viral chitinases: Attractive biopesticides for integrated pest management. Biotechnol. Adv. 2018, 36, 818–838. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Soliman, M.H.; Nassar, A.H.; Sivasithamparam, K.; McKenna, F.; Hardy, G.E.S.J. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol. 2000, 49, 573–583. [Google Scholar] [CrossRef]
- Mouloud, G.; Samir, M.; Laid, B.; Daoud, H. Isolation and characterization of rhizospheric Streptomyces spp. for the biocontrol of Fusarium wilt (bayoud) disease of date palm (Phoenix dactylifera L.). J. Sci. Agric. 2017, 1, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.M.; Crawford, D.L. Characterization of Streptomyces lydicus WYEC108 as a potential biocontrol agent against fungal root and seed rots. Appl. Environ. Microbiol. 1995, 61, 3119–3128. [Google Scholar] [CrossRef] [Green Version]
- Getha, K.; Vikineswary, S. Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f.sp. cubense race 4: Indirect evidence for the role of antibiosis in the antagonistic process. J. Ind. Microbiol. Biotechnol. 2002, 28, 303–310. [Google Scholar]
- He, D.-C.; He, M.-H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-Economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.-S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The use of microbial inoculants for biological control, plant growth promotion, and sustainable agriculture: A review. Eur. J. Plant Pathol. 2022. in press. [Google Scholar]
- Goodfellow, M.; Williams, S. Ecology of actinomycetes. Annu. Rev. Microbiol. 1983, 37, 189–216. [Google Scholar] [CrossRef]
- AbuQamar, S.F.; Moustafa, K.; Tran, L.S. ‘Omics’ and plant responses to Botrytis cinerea. Front. Plant Sci. 2016, 7, 1658. [Google Scholar] [CrossRef] [Green Version]
- AbuQamar, S.F.; Moustafa, K.; Tran, L.S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate | Clearing Zone Diameter a | Inhibition Zone Diameter b | Production of | ||
|---|---|---|---|---|---|---|
| (mm) | VC c | Siderophore d | HCN d | |||
| SA | ||||||
| Streptomyces | #1 | 0.0 | 34.3 ± 0.7 ab | + | - | - |
| #5 | 32.2 ± 1.0 a | 0.0 | + | + | - | |
| #7 | 0.0 | 41.6 ± 1.7 c | - | + | - | |
| #9 e | 0.0 | 0.0 | - | - | - | |
| #13 | 41.4 ± 2.1 b | 0.0 | - | + | - | |
| #15 | 0.00 | 37.3 ± 0.8 b | + | - | - | |
| #16 e | 55.4 ± 2.2 c | 0.0 | + | + | - | |
| #22 | 0.0 | 47.3 ± 1.2 d | + | + | - | |
| #24 | 37.1 ± 1.1 ab | 0.0 | - | - | - | |
| #25 | 0.0 | 31.4 ± 0.4 a | - | + | - | |
| #28 e | 0.0 | 48.2 ± 2.0 d | + | + | - | |
| #31 | 42.7 ± 1.3 b | 0.0 | - | + | - | |
| NSA | ||||||
| Actinoplanes | #3 | 0.0 | 32.8 ± 0.9 a | + | - | - |
| Dactylosporangium | #8 | 0.0 | 42.2 ± 1.6 c | - | + | - |
| Microbispora | #21 | 33.8 ± 2.0 a | 0.0 | + | - | - |
| Micromonospora | #29 | 0.0 | 47.8 ± 1.1 d | + | + | - |
| #17 | 54.6 ± 1.9 c | 0.0 | + | + | - | |
| Media | Isolate | Culture Filtrate (%) | Colony Diameter (mm) | Mycelial Dry Weight (g) | Conidia Germination (%) | Germ Tube Length (µm) |
|---|---|---|---|---|---|---|
| CCB | #16 | 0 | 97.7 ± 0.4 d | 76.4 ± 2.8 d | 87.8 ± 0.5 d | 63.4 ± 1.2 d |
| 25 | 56.3 ± 4.2 c | 45.2 ± 2.2 c | 55.1 ± 5.3 c | 33.3 ± 4.1 c | ||
| 50 | 15.5 ± 2.8 b | 13.8 ± 1.1 b | 17.6 ± 2.4 b | 7.9 ± 1.0 b | ||
| 100 | 0.0 a | 0.0 a | 1.0 ± 0.2 a | 0.0 a | ||
| FMEB | #28 | 0 | 98.0 ± 0.6 d | 74.5 ± 1.4 d | 86.6 ± 1.0 d | 50.2 ± 1.7 d |
| 25 | 61.9 ± 3.8 c | 39.9 ± 4.4 c | 51.7 ± 3.5 c | 25.7 ± 5.7 c | ||
| 50 | 13.2 ± 1.6 b | 7.8 ± 1.3 b | 14.3 ± 2.2 b | 11.2 ± 3.2 b | ||
| 100 | 0.0 a | 0.0 a | 0.9 ± 0.3 a | 1.9 ± 0.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alblooshi, A.A.; Purayil, G.P.; Saeed, E.E.; Ramadan, G.A.; Tariq, S.; Altaee, A.S.; El-Tarabily, K.A.; AbuQamar, S.F. Biocontrol Potential of Endophytic Actinobacteria against Fusarium solani, the Causal Agent of Sudden Decline Syndrome on Date Palm in the UAE. J. Fungi 2022, 8, 8. https://doi.org/10.3390/jof8010008
Alblooshi AA, Purayil GP, Saeed EE, Ramadan GA, Tariq S, Altaee AS, El-Tarabily KA, AbuQamar SF. Biocontrol Potential of Endophytic Actinobacteria against Fusarium solani, the Causal Agent of Sudden Decline Syndrome on Date Palm in the UAE. Journal of Fungi. 2022; 8(1):8. https://doi.org/10.3390/jof8010008
Chicago/Turabian StyleAlblooshi, Aisha A., Gouthaman P. Purayil, Esam Eldin Saeed, Gaber A. Ramadan, Saeed Tariq, Amna S. Altaee, Khaled A. El-Tarabily, and Synan F. AbuQamar. 2022. "Biocontrol Potential of Endophytic Actinobacteria against Fusarium solani, the Causal Agent of Sudden Decline Syndrome on Date Palm in the UAE" Journal of Fungi 8, no. 1: 8. https://doi.org/10.3390/jof8010008