
MAC Family Transcription Factors Enhance the Tolerance of Mycelia to Heat Stress and Promote the Primordial Formation Rate of Pleurotus ostreatus
 and
and 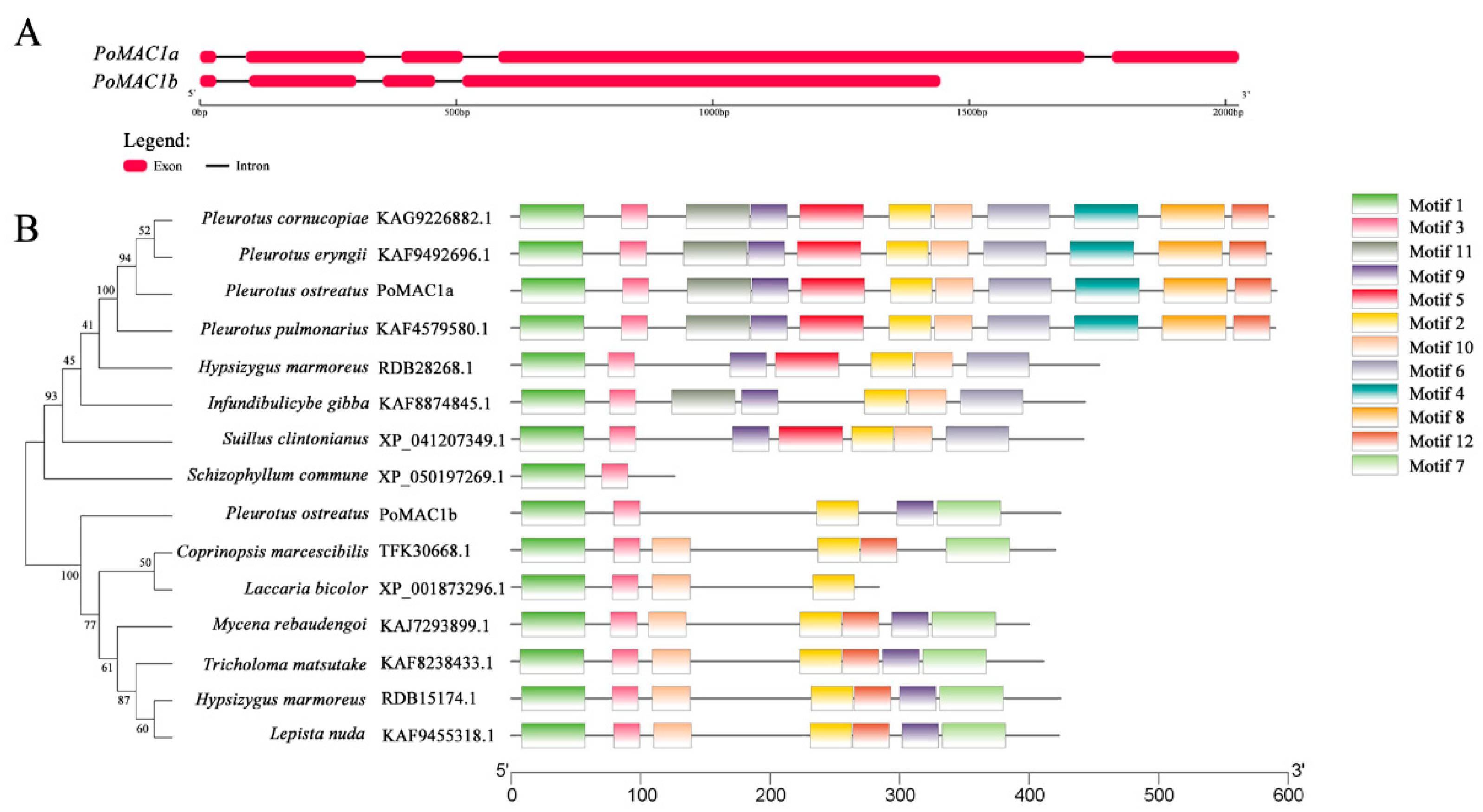{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Identification, Cloning, and Sequence Analysis of PoMAC1 Genes
2.3. Subcellular Localization of PoMAC1a and PoMAC1b
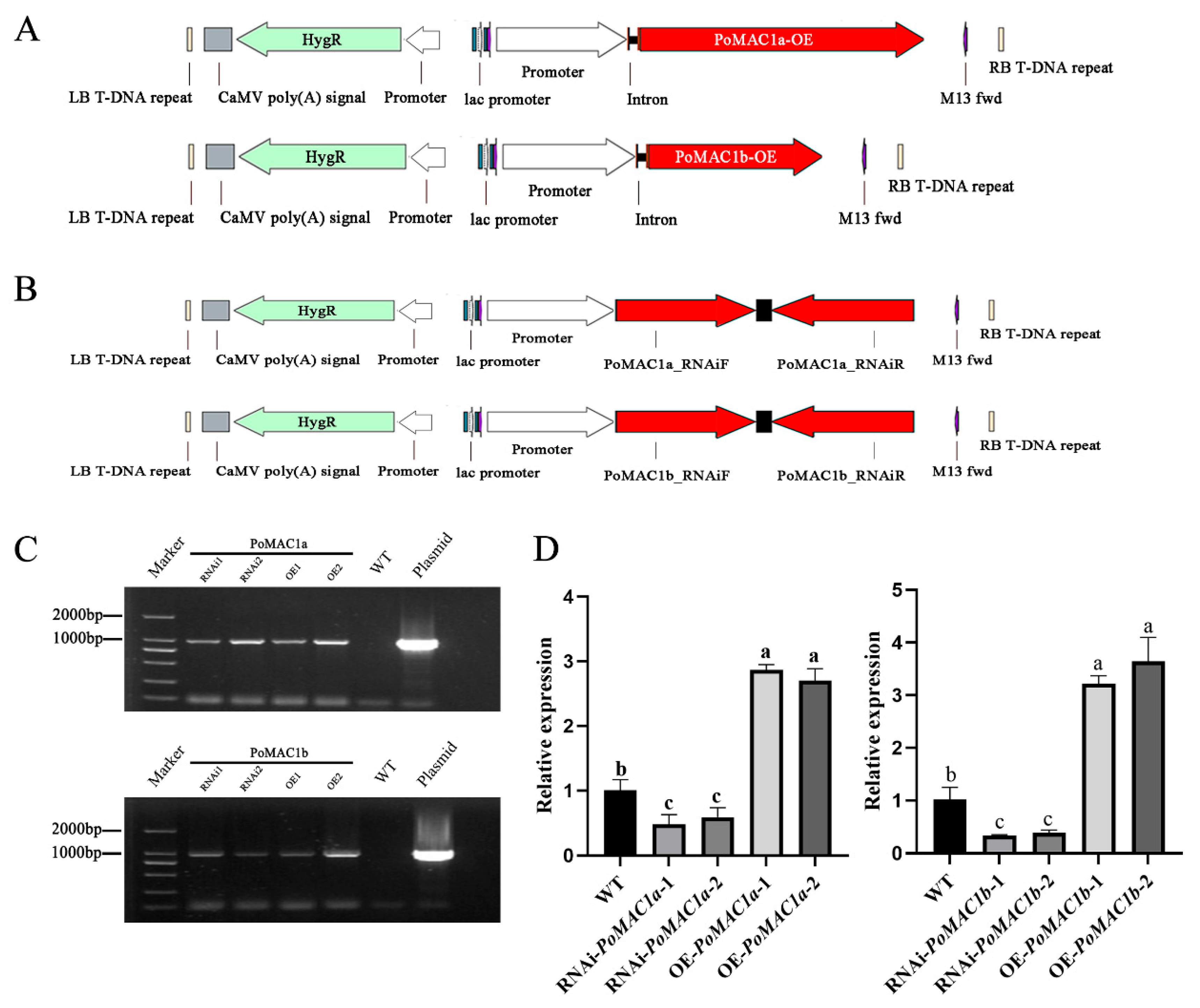
2.4. Construction of OE and RNAi Plasmids
2.5. Acquisition of Mutant Strains
2.6. Mushroom Production
2.7. Heat Stress Treatment
2.8. Growth Susceptibility Assay
2.9. Quantitative PCR (qPCR)
2.10. Data Analysis
3. Results
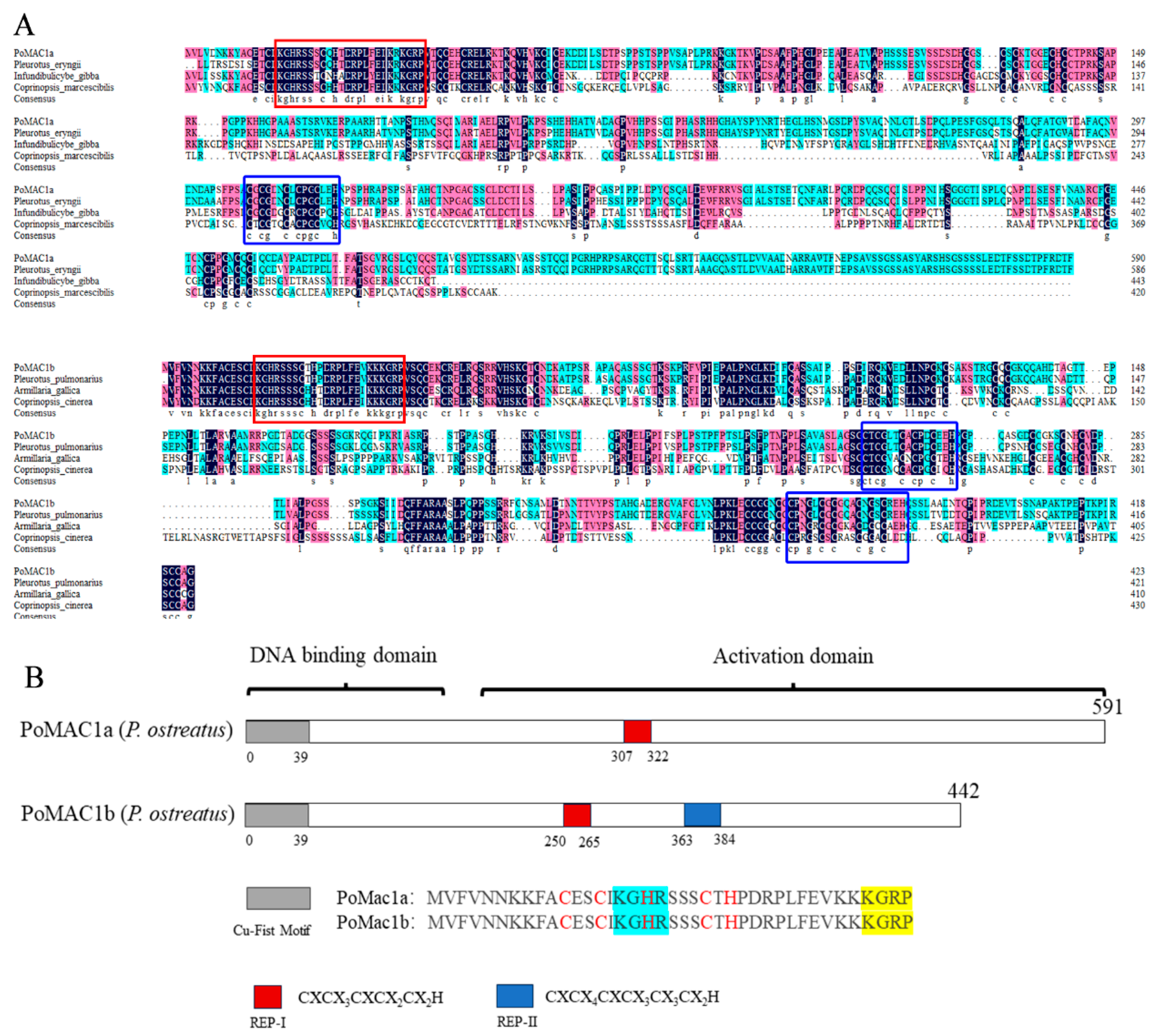
3.1. Cloning and Bioinformatics Analysis of PoMAC1a and PoMAC1b
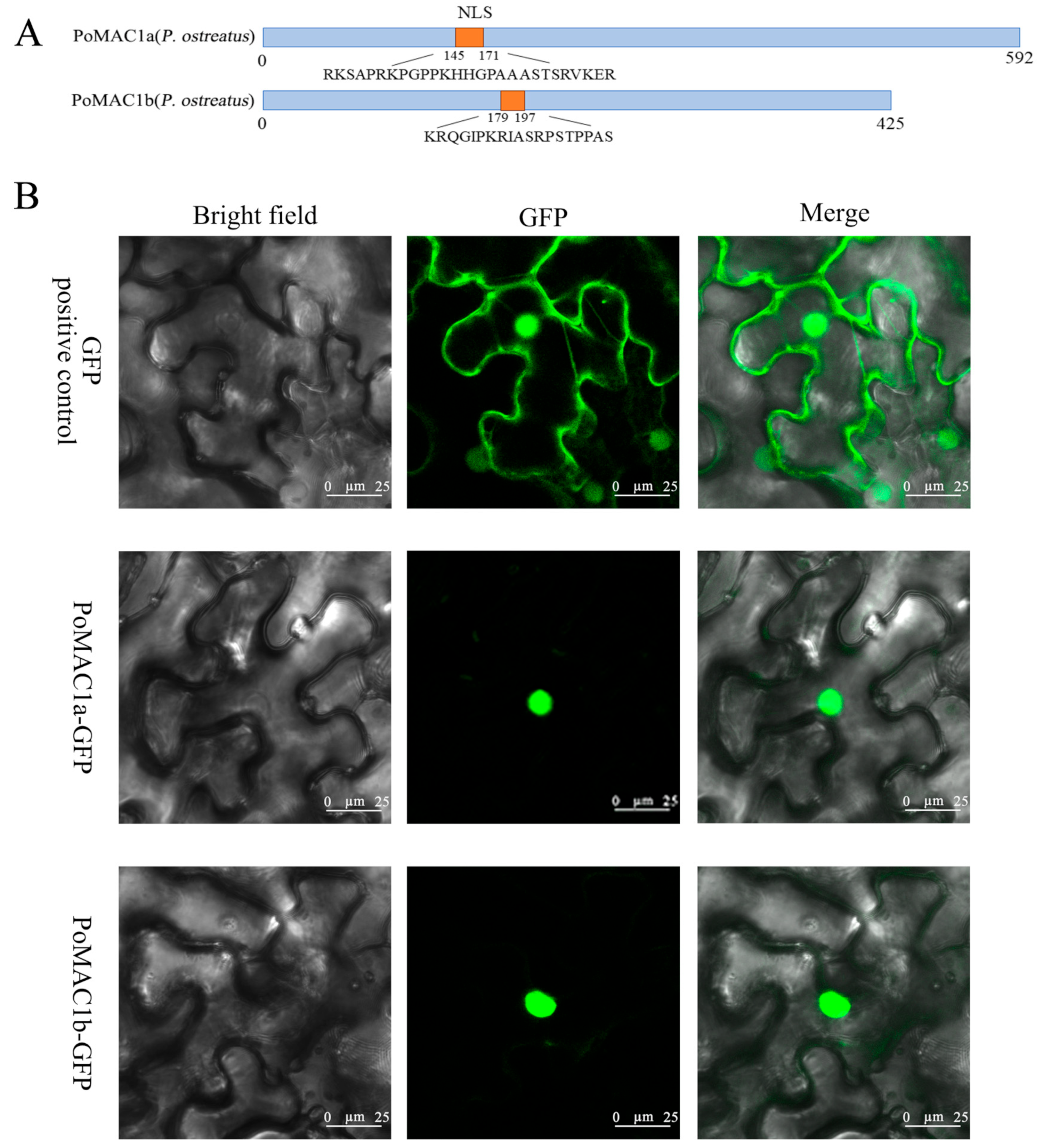
3.2. Subcellular Localization of PoMAC1
3.3. Acquisition of PoMAC1a and PoMAC1b Mutant Strains
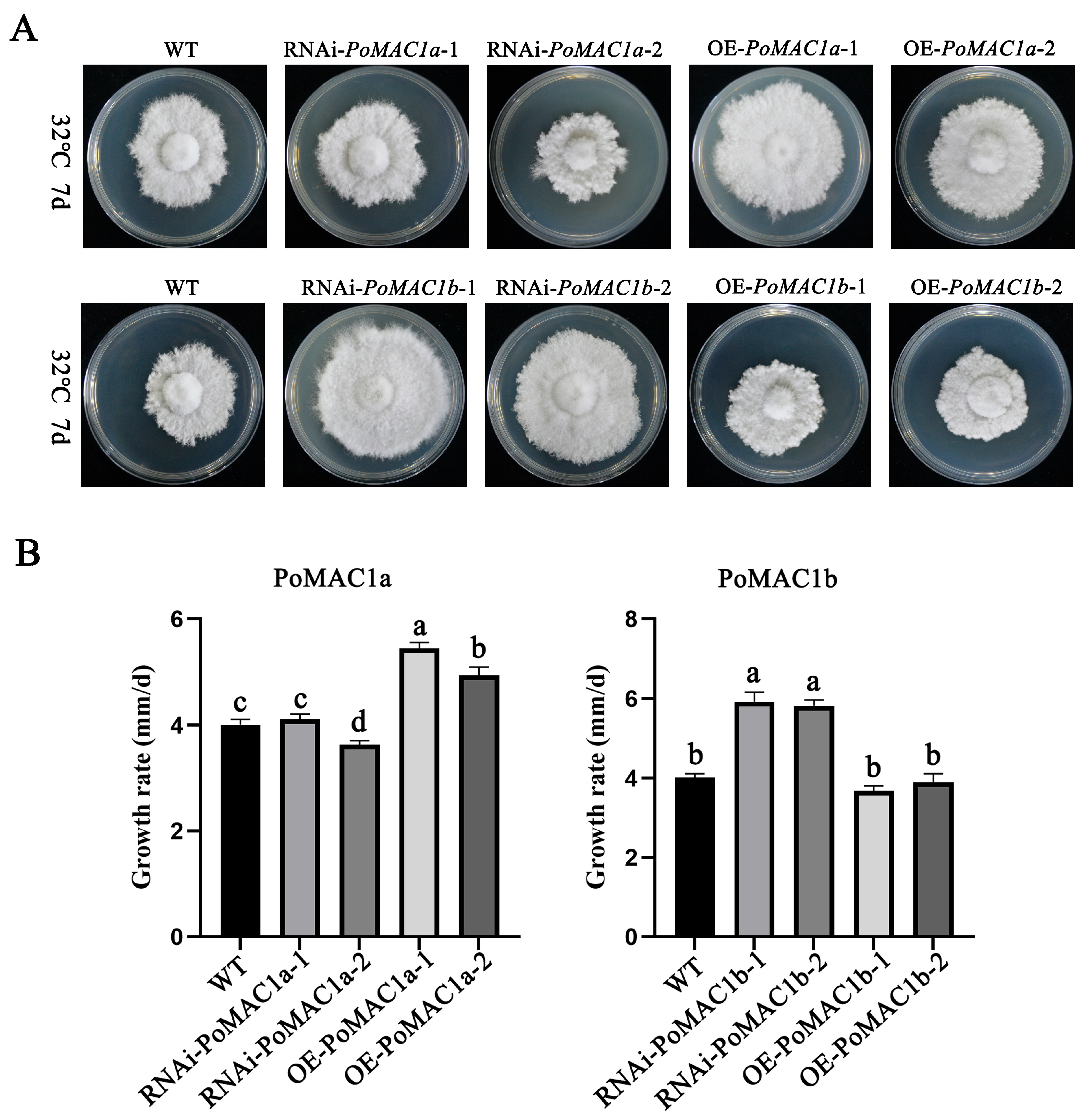
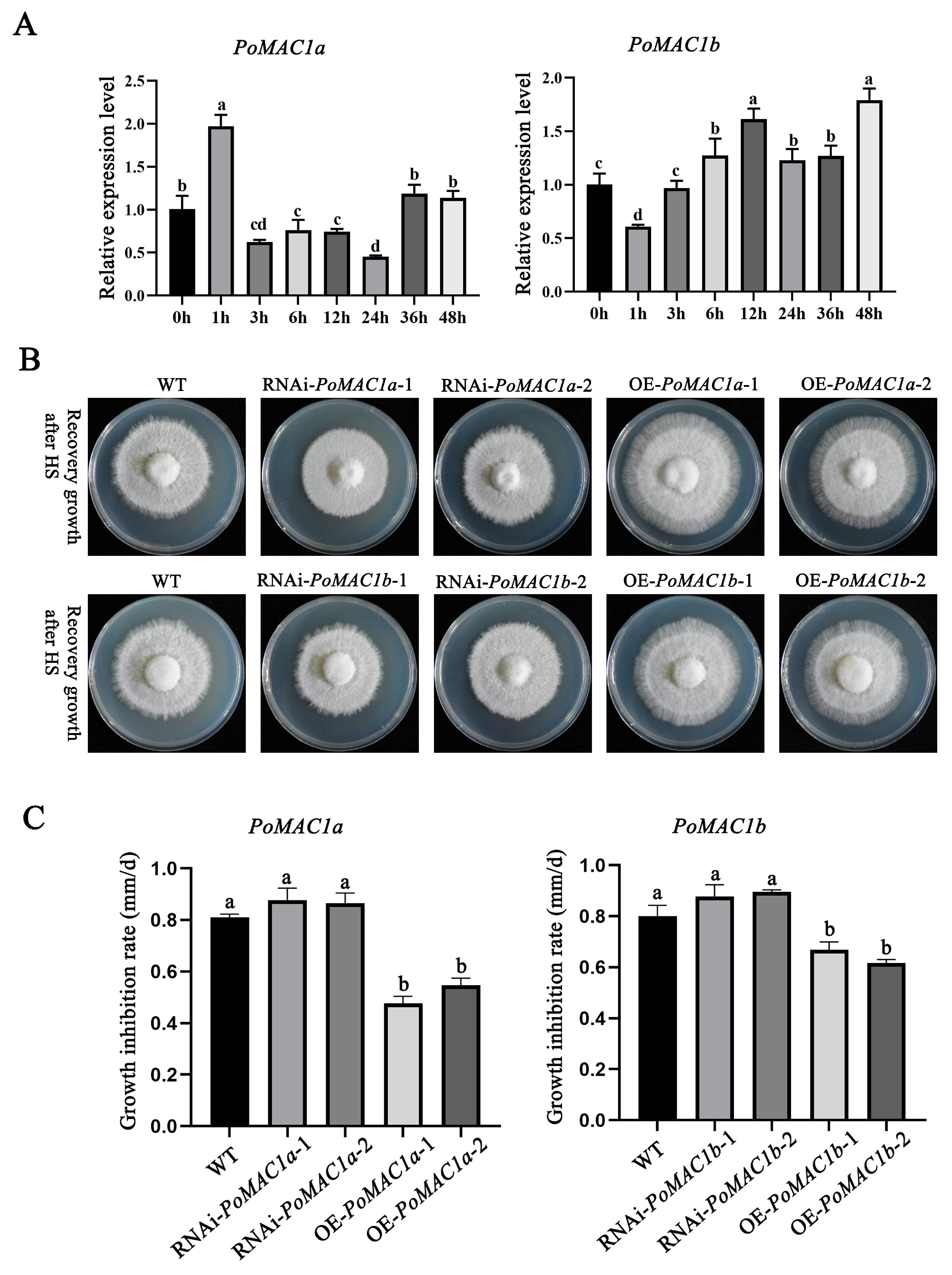
3.4. PoMAC1 Participates in the Regulation of the Mycelial Response to Different Heat Stresses
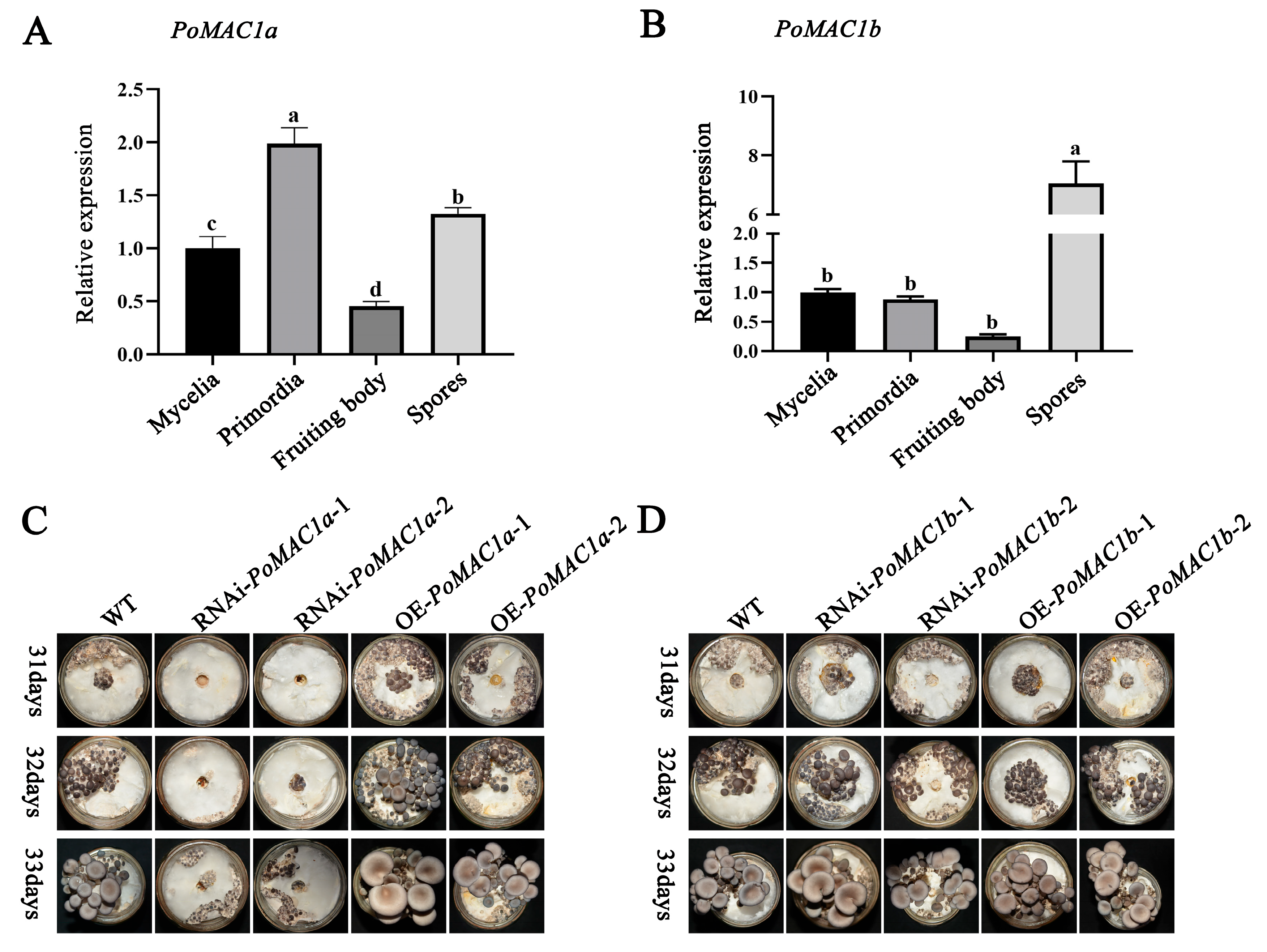
3.5. PoMAC1a Positively Regulates the Formation of Primordia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, F.; Fan, J.; Zhu, M.; Tong, H. Antioxidant Effects of a Water-Soluble Proteoglycan Isolated from the Fruiting Bodies of Pleurotus ostreatus. J. Taiwan Inst. Chem. Eng. 2011, 42, 402–407. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, T.; Zhou, H.; Zhang, Y.; Jin, G.; Yang, Y. Antidiabetic Effect of Polysaccharides from Pleurotus ostreatus in Streptozotocin-Induced Diabetic Rats. Int. J. Biol. Macromol. 2016, 83, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Wisbeck, E.; Facchini, J.M.; Alves, E.P.; Silveira, M.L.L.; Gern, R.M.M.; Ninow, J.L.; Furlan, S.A. A Polysaccharide Fraction Extracted from Pleurotus ostreatus Mycelial Biomass Inhibit Sarcoma 180 Tumor. An. Acad. Bras. Cienc. 2017, 89, 2013–2020. [Google Scholar] [CrossRef] [PubMed]
- BBarbosa, J.R.; Freitas, M.M.S.; Oliveira, L.C.; Martins, L.H.S.; Almada-Vilhena, A.O.; Oliveira, R.M.; Pieczarka, J.C.; Davi do Socorro, B.; Junior, R.N.C. Obtaining Extracts Rich in Antioxidant Polysaccharides from the Edible Mushroom Pleurotus ostreatus Using Binary System with Hot Water and Supercritical CO2. Food Chem. 2020, 330, 127173. [Google Scholar] [CrossRef] [PubMed]
- Othman, A.; Elshafei, A.M.; Hassan, M.M.; Haroun, B.M.; Farrag, A.A. Purification, Biochemical Characterization and Applications of Pleurotus ostreatus ARC280 Laccase. Br. Microbiol. Res. J. 2014, 4, 1418–1439. [Google Scholar] [CrossRef]
- Jedinak, A.; Dudhgaonkar, S.; Wu, Q.-L.; Simon, J.; Sliva, D. Anti-Inflammatory Activity of Edible Oyster Mushroom Is Mediated through the Inhibition of NF-κB and AP-1 Signaling. Nutr. J. 2011, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C. Cultivation of Pleurotus ostreatus and Other Edible Mushrooms. Appl. Microbiol. Biotechnol. 2010, 85, 1321–1337. [Google Scholar] [CrossRef]
- Lyu, X.; Jiang, S.; Wang, L.; Chou, T.; Wang, Q.; Meng, L.; Mukhtar, I.; Xie, B.; Wang, W. The Fvclp1 Gene Regulates Mycelial Growth and Fruiting Body Development in Edible Mushroom Flammulina velutipes. Arch. Microbiol. 2021, 203, 5373–5380. [Google Scholar] [CrossRef]
- Sardar, H.; Ali, M.A.; Ayyub, C.M.; Ahmad, R. Effects of Different Culture Media, Temperature and pH Levels on the Growth of Wild and Exotic Pleurotus Species. Pak. J. Phytopathol. 2015, 27, 139–145. [Google Scholar]
- Shen, Y.; Cai, W.; Zhou, S.; Jin, Q.; Li, L. Phenotype Analysis of Mycelium Growth Regeneration after HeatStress in a Lentinula edodes F2 Population. J. Hortic. Sci. Biotechnol. 2017, 92, 1–7. [Google Scholar] [CrossRef]
- Song, C.; Chen, Q.; Wu, X.; Zhang, J.; Huang, C. Heat Stress Induces Apoptotic-Like Cell Death in Two Pleurotus Species. Curr. Microbiol. 2014, 69, 611–616. [Google Scholar] [CrossRef]
- Yan, Z.; Zhao, M.; Wu, X.; Zhang, J. Metabolic Response of Pleurotus Ostreatus to Continuous Heat Stress. Front. Microbiol. 2019, 10, 3148. [Google Scholar] [CrossRef]
- Hou, L.; Wang, L.; Wu, X.; Gao, W.; Zhang, J.; Huang, C. Expression Patterns of Two pal Genes of Pleurotus ostreatus across Developmental Stages and under Heat Stress. BMC Microbiol. 2019, 19, 231. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Zheng, L.; Zhu, J.; Chen, L.; Ma, A. Enhancing Stress Tolerance by Overexpression of a Methionine Sulfoxide Reductase A (MsrA) Gene in Pleurotus ostreatus. Appl. Microbiol. Biotechnol. 2015, 99, 3115–3126. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gu, B.; Huang, J.; Jiang, S.; Chen, Y.; Yin, Y.; Pan, Y.; Yu, G.; Li, Y.; Wong, B.H.C. Transcriptome and Proteome Exploration to Provide a Resource for the Study of Agrocybe aegerita. PLoS ONE 2013, 8, e56686. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tian, Y.-S.; Peng, R.-H.; Xiong, A.-S.; Zhu, B.; Jin, X.-F.; Gao, J.-J.; Hou, X.-L.; Yao, Q.-H. Yeast Copper-Dependent Transcription Factor ACE1 Enhanced Copper Stress Tolerance in Arabidopsis. BMB Rep. 2009, 42, 752–757. [Google Scholar] [CrossRef]
- Wu, T.; Hu, C.; Xie, B.; Zhang, L.; Li, S. A Single Transcription Factor (PDD1) Determines Development and Yield of Winter Mushroom (Flammulina velutipes). Appl. Environ. Microbiol. 2019, 85, e01735-19. [Google Scholar] [CrossRef]
- Wu, T.; Hu, C.; Xie, B.; Wei, S.; Zhang, L.; Zhu, Z.; Zhang, Z.; Li, S. A Putative Transcription Factor LFC1 Negatively Regulates Development and Yield of Winter Mushroom. Appl. Microbiol. Biotechnol. 2020, 104, 5827–5844. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, R.; Wei, X.; Lv, M.; Jiang, Z. Metalloimmunology: The Metal Ion-Controlled Immunity. Adv. Immunol. 2020, 145, 187–241. [Google Scholar] [CrossRef]
- Jungmann, J.; Reins, H.A.; Lee, J.; Romeo, A.; Hassett, R.; Kosman, D.; Jentsch, S. MAC1, a Nuclear Regulatory Protein Related to Cu-Dependent Transcription Factors Is Involved in Cu/Fe Utilization and Stress Resistance in Yeast. EMBO J. 1993, 12, 5051–5056. [Google Scholar] [CrossRef]
- Jensen, L.T.; Posewitz, M.C.; Srinivasan, C.; Winge, D.R. Mapping of the DNA Binding Domain of the Copper-Responsive Transcription Factor Mac1 from Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 23805–23811. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.R.; Keller, G.L.; Pickering, I.J.; Harris, H.H.; George, G.N.; Winge, D.R. Structures of the Cuprous-Thiolate Clusters of the Mac1 and Ace1 Transcriptional Activators. Biochemistry 2002, 41, 6469–6476. [Google Scholar] [CrossRef] [PubMed]
- Wegner, S.V.; Sun, F.; Hernandez, N.; He, C. The Tightly Regulated Copper Window in Yeast. Chem. Commun. 2011, 47, 2571–2573. [Google Scholar] [CrossRef] [PubMed]
- Thiele, D.J. ACE1 Regulates Expression of the Saccharomyces Cerevisiae Metallothionein Gene. Mol. Cell. Biol. 1988, 8, 2745–2752. [Google Scholar] [CrossRef] [PubMed]
- Polanco, R.; Canessa, P.; Rivas, A.; Larrondo, L.F.; Lobos, S.; Vicuña, R. Cloning and Functional Characterization of the Gene Encoding the Transcription Factor Ace1 in the Basidiomycete Phanerochaete chrysosporium. Biol. Res. 2006, 39, 641–648. [Google Scholar] [CrossRef]
- Kusuya, Y.; Hagiwara, D.; Sakai, K.; Yaguchi, T.; Gonoi, T.; Takahashi, H. Transcription Factor Afmac1 Controls Copper Import Machinery in Aspergillus fumigatus. Curr. Genet. 2017, 63, 777–789. [Google Scholar] [CrossRef]
- Cai, Z.; Du, W.; Liu, L.; Pan, D.; Lu, L. Molecular Characteristics of the Conserved Aspergillus nidulans Transcription Factor Mac1 and Its Functions in Response to Copper Starvation. mSphere 2019, 4, e00670-18. [Google Scholar] [CrossRef]
- Grene, A.R.; Neval, E.; Heath, L.S. Role of Superoxide Dismutases (SODs) in Controlling Oxidative Stress in Plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Nasuno, R.; Aitoku, M.; Manago, Y.; Nishimura, A.; Sasano, Y.; Takagi, H. Nitric Oxide-Mediated Antioxidative Mechanism in Yeast through the Activation of the Transcription Factor Mac1. PLoS ONE 2014, 9, e113788. [Google Scholar] [CrossRef]
- Qu, J.; Zhao, M.; Hsiang, T.; Feng, X.; Zhang, J.; Huang, C. Identification and Characterization of Small Noncoding RNAs in Genome Sequences of the Edible Fungus Pleurotus ostreatus. BioMed Res. Int. 2016, 2016, 2503023. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, X.; Gao, W.; Zhao, M.; Zhang, J.; Huang, C. Differential Expression Patterns of Pleurotus ostreatus Catalase Genes during Developmental Stages and under Heat Stress. Genes 2017, 8, 335. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Zhao, M.; Huang, C.; Wu, X.; Zhang, J. Nitric Oxide Improves the Tolerance of Pleurotus ostreatus to Heat Stress by Inhibiting Mitochondrial Aconitase. Appl. Environ. Microbiol. 2019, 86, e02303-19. [Google Scholar] [CrossRef] [PubMed]
- Laskey, R.A.; Dingwall, C. Nuclear Shuttling: The Default Pathway for Nuclear Proteins? Cell 1993, 74, 585–586. [Google Scholar] [CrossRef]
- Zhu, Z.; Labbé, S.; Peña, M.M.O.; Thiele, D.J. Copper Differentially Regulates the Activity and Degradation of Yeast Mac1 Transcription Factor. J. Biol. Chem. 1998, 273, 1277–1280. [Google Scholar] [CrossRef]
- Cai, Z.; Du, W.; Zeng, Q.; Long, N.; Dai, C.; Lu, L. Cu-Sensing Transcription Factor Mac1 Coordinates with the Ctr Transporter Family to Regulate Cu Acquisition and Virulence in Aspergillus fumigatus. Fungal Genet. Biol. 2017, 107, 31–43. [Google Scholar] [CrossRef]
- Beaudoin, J.; Mercier, A.; Langlois, R.; Labbe, S. The Schizosaccharomyces pombe Cuf1 Is Composed of Functional Modules from Two Distinct Classes of Copper Metalloregulatory Transcription Factors. J. Biol. Chem. 2003, 278, 14565–14577. [Google Scholar] [CrossRef]
- Culbertson, E.M.; Bruno, V.M.; Cormack, B.P.; Culotta, V.C. Expanded Role of the Cu-sensing Transcription Factor Mac1p in Candida Albicans. Mol. Microbiol. 2020, 114, 1006–1018. [Google Scholar] [CrossRef]
- Keller, G.; Gross, C.; Kelleher, M.; Winge, D.R. Functional Independence of the Two Cysteine-Rich Activation Domains in the Yeast Mac1 Transcription Factor. J. Biol. Chem. 2000, 275, 29193–29199. [Google Scholar] [CrossRef]
- Voutsina, A.; Fragiadakis, G.S.; Boutla, A.; Alexandraki, D. The Second Cysteine-Rich Domain of Mac1p Is a Potent Transactivator That Modulates DNA Binding Efficiency and Functionality of the Protein. Febs Lett. 2001, 494, 38–43. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, K.; Guo, L.; Zhang, B.; Chang, M.; Meng, J.; Deng, B.; Liu, J.; Hou, L. MAC Family Transcription Factors Enhance the Tolerance of Mycelia to Heat Stress and Promote the Primordial Formation Rate of Pleurotus ostreatus. J. Fungi 2024, 10, 13. https://doi.org/10.3390/jof10010013
Yan K, Guo L, Zhang B, Chang M, Meng J, Deng B, Liu J, Hou L. MAC Family Transcription Factors Enhance the Tolerance of Mycelia to Heat Stress and Promote the Primordial Formation Rate of Pleurotus ostreatus. Journal of Fungi. 2024; 10(1):13. https://doi.org/10.3390/jof10010013
Chicago/Turabian StyleYan, Kexing, Lifeng Guo, Benfeng Zhang, Mingchang Chang, Junlong Meng, Bing Deng, Jingyu Liu, and Ludan Hou. 2024. "MAC Family Transcription Factors Enhance the Tolerance of Mycelia to Heat Stress and Promote the Primordial Formation Rate of Pleurotus ostreatus" Journal of Fungi 10, no. 1: 13. https://doi.org/10.3390/jof10010013