
Co-regulation of Thermosensor Pathogenic Factors by C-di-GMP-Related Two-Component Systems and a cAMP Receptor-like Protein (Clp) in Stenotrophomonas maltophilia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Plasmids and Strain Construction
2.3. Biofilm Assay and Quantification
2.4. Swimming Motility and EPS Production
2.5. Analysis of Promoter Activity
2.6. Bioinformatics Analysis
2.7. Enzymatic Activity Assays
2.8. Detection of Intracellular c-di-GMP Levels
2.9. Statistical Analysis
3. Results
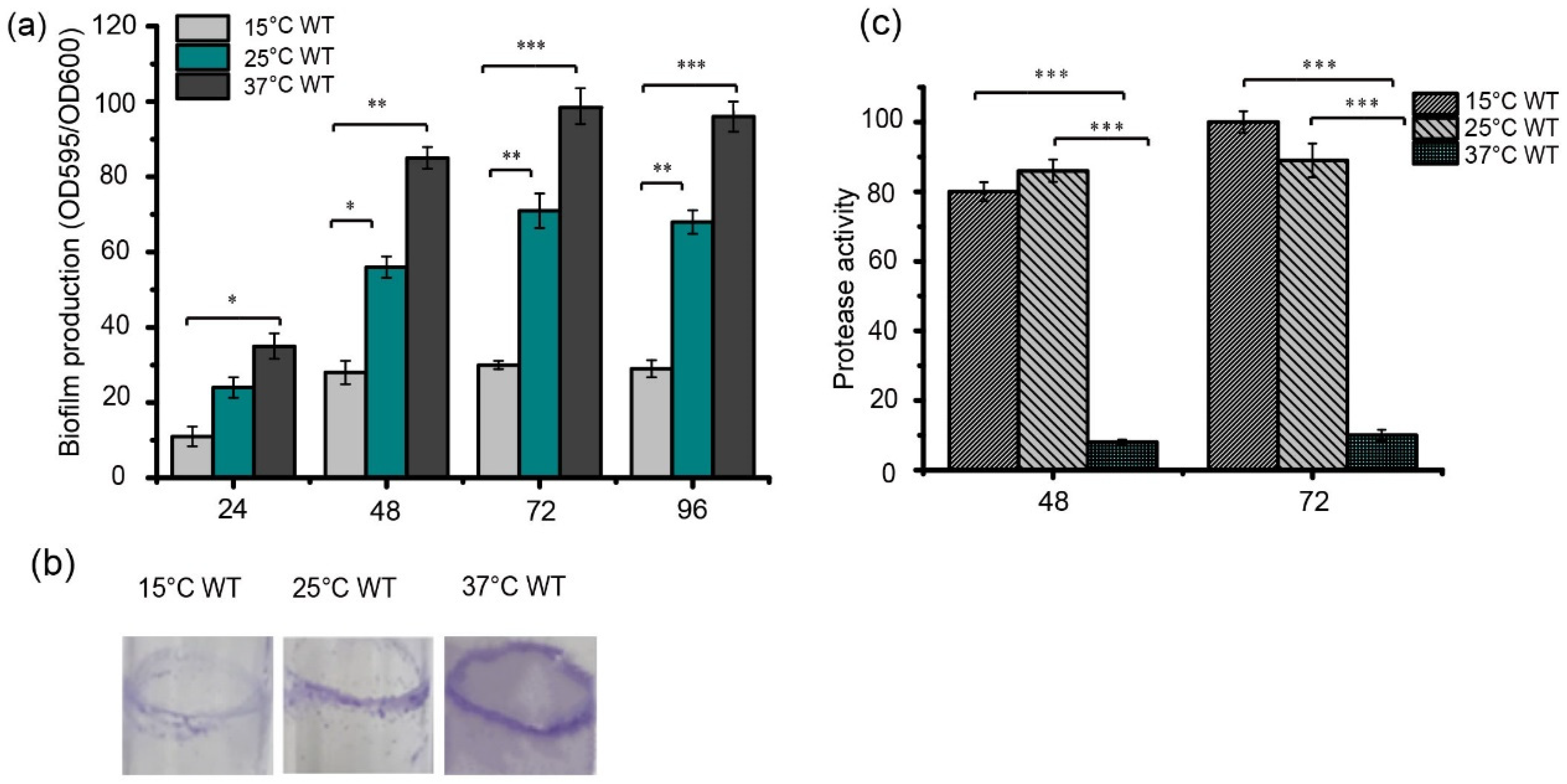
3.1. Temperature Differentially Regulates Biofilm Formation and Protease Production in S. maltophilia
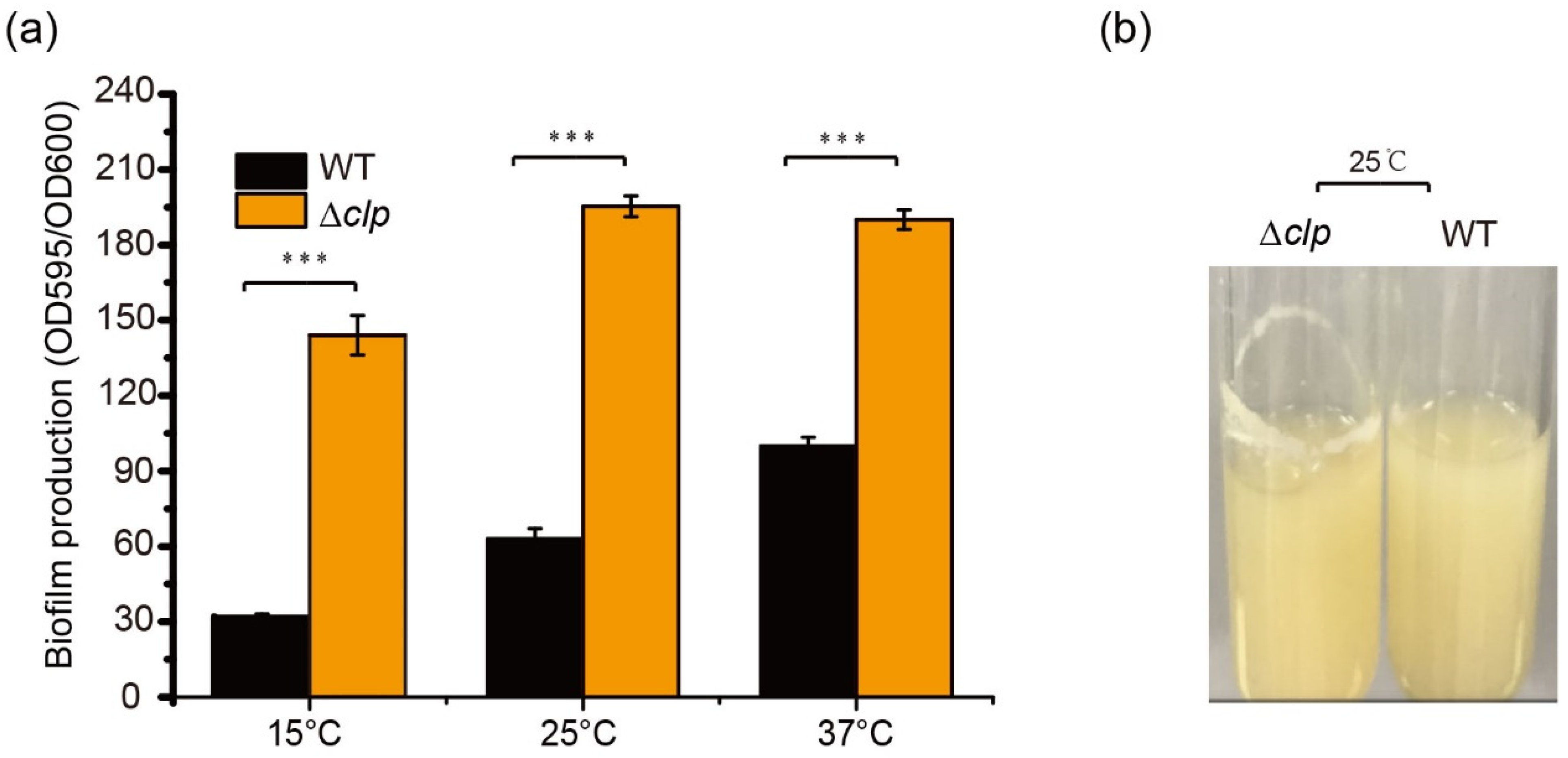
3.2. Clp Negatively Regulates Thermosensor Biofilm Formation in S. maltophilia
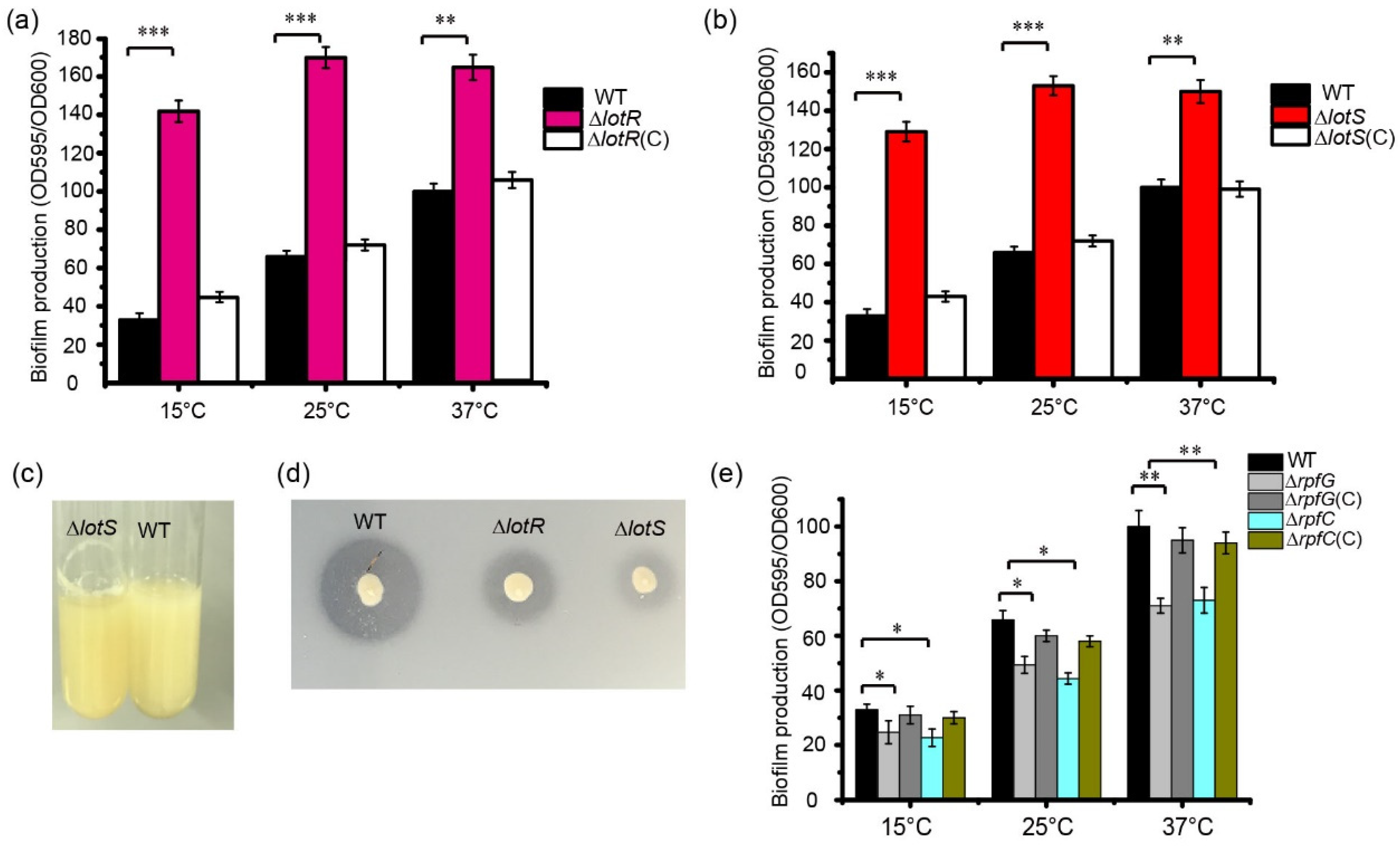
3.3. LotS/LotR Negatively Mediates Thermosensor Biofilm Formation in S. maltophilia
3.4. RpfC/RpfG Positively Regulate Thermosensor Biofilm Formation in S. maltophilia
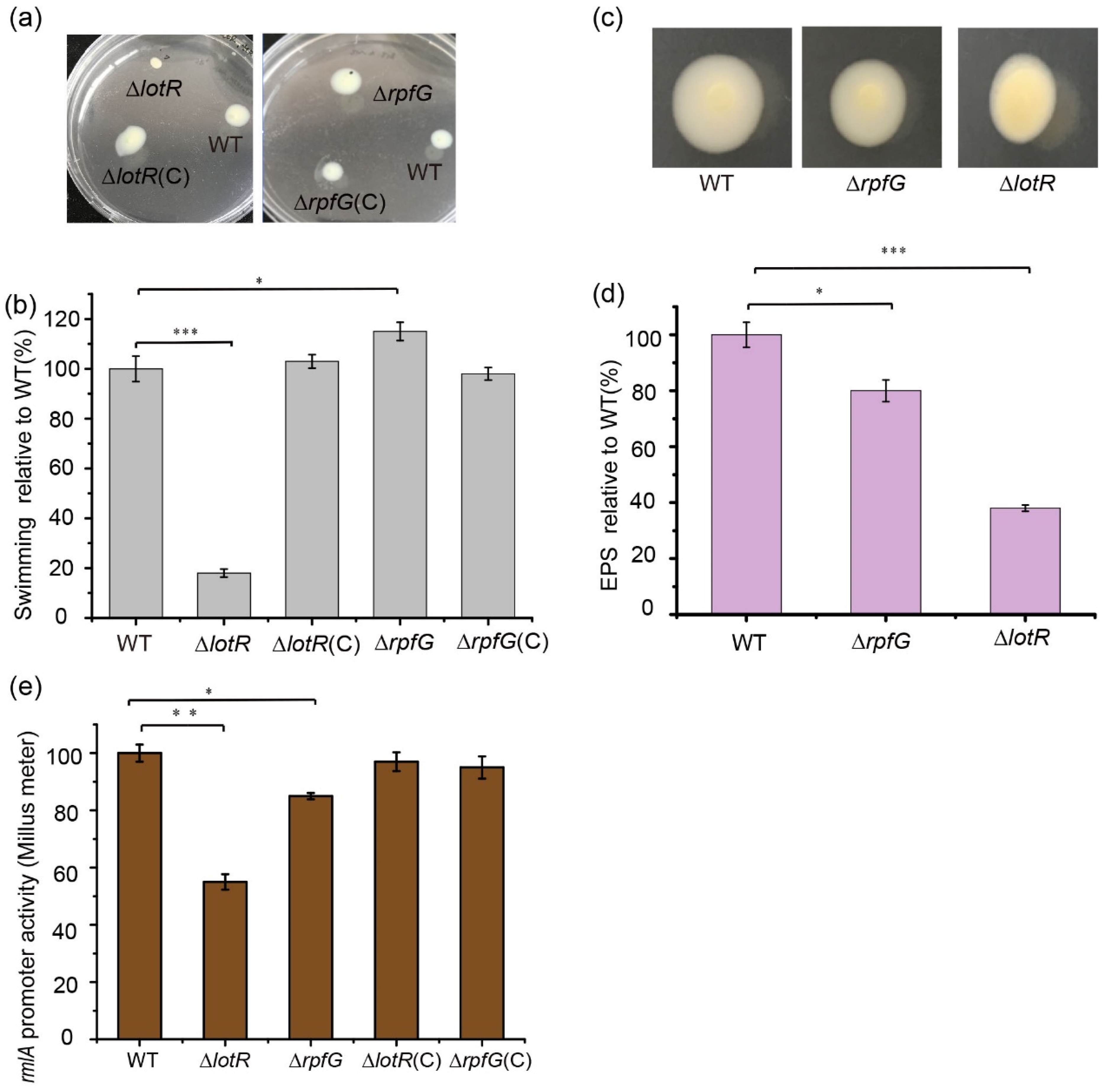
3.5. LotS/LotR Contributes More to Physiological Functions Than RpfC/RpfG
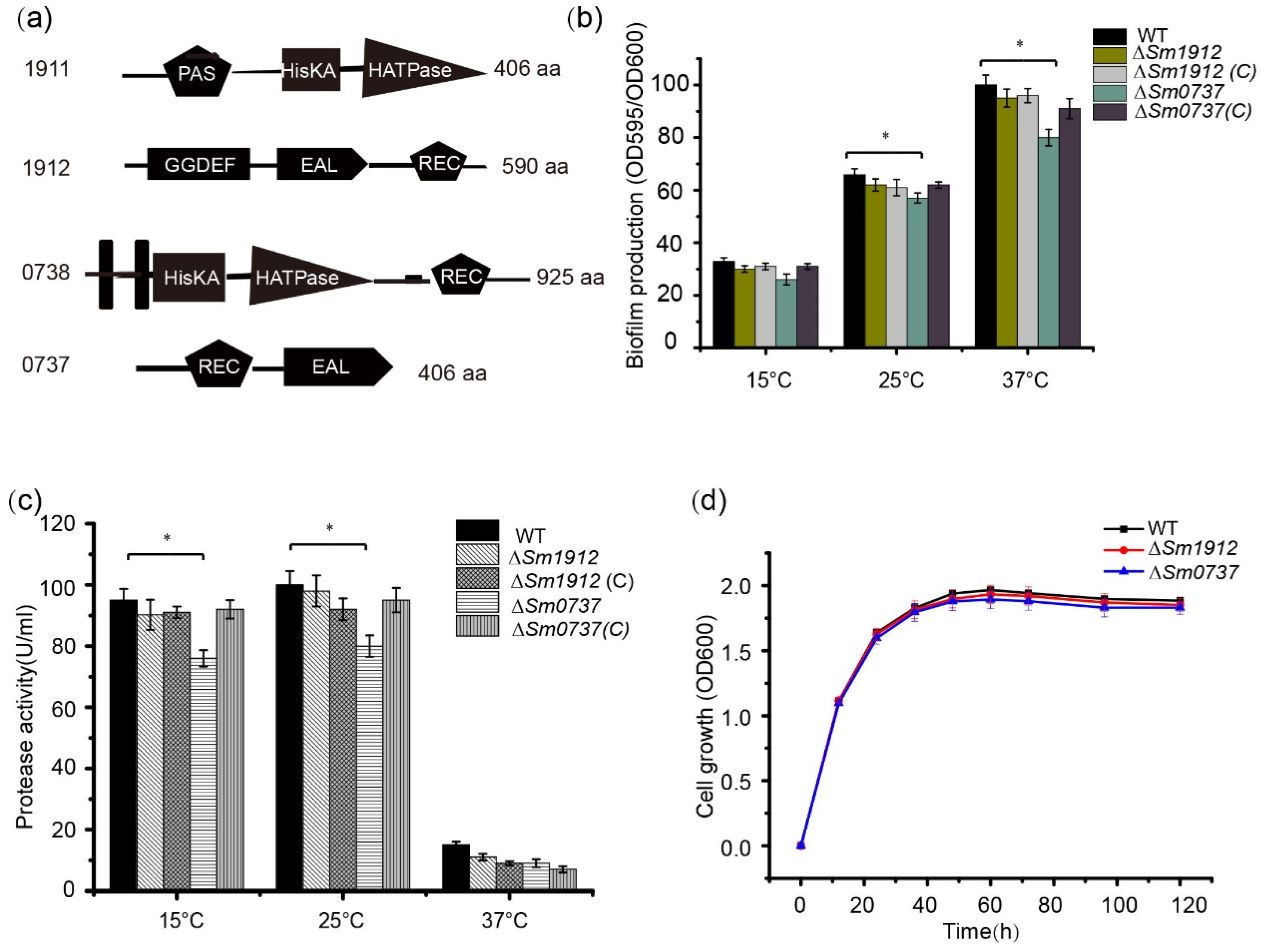
3.6. S. maltophilia Encodes Two Additional TCSs Related to c-di-GMP Metabolism
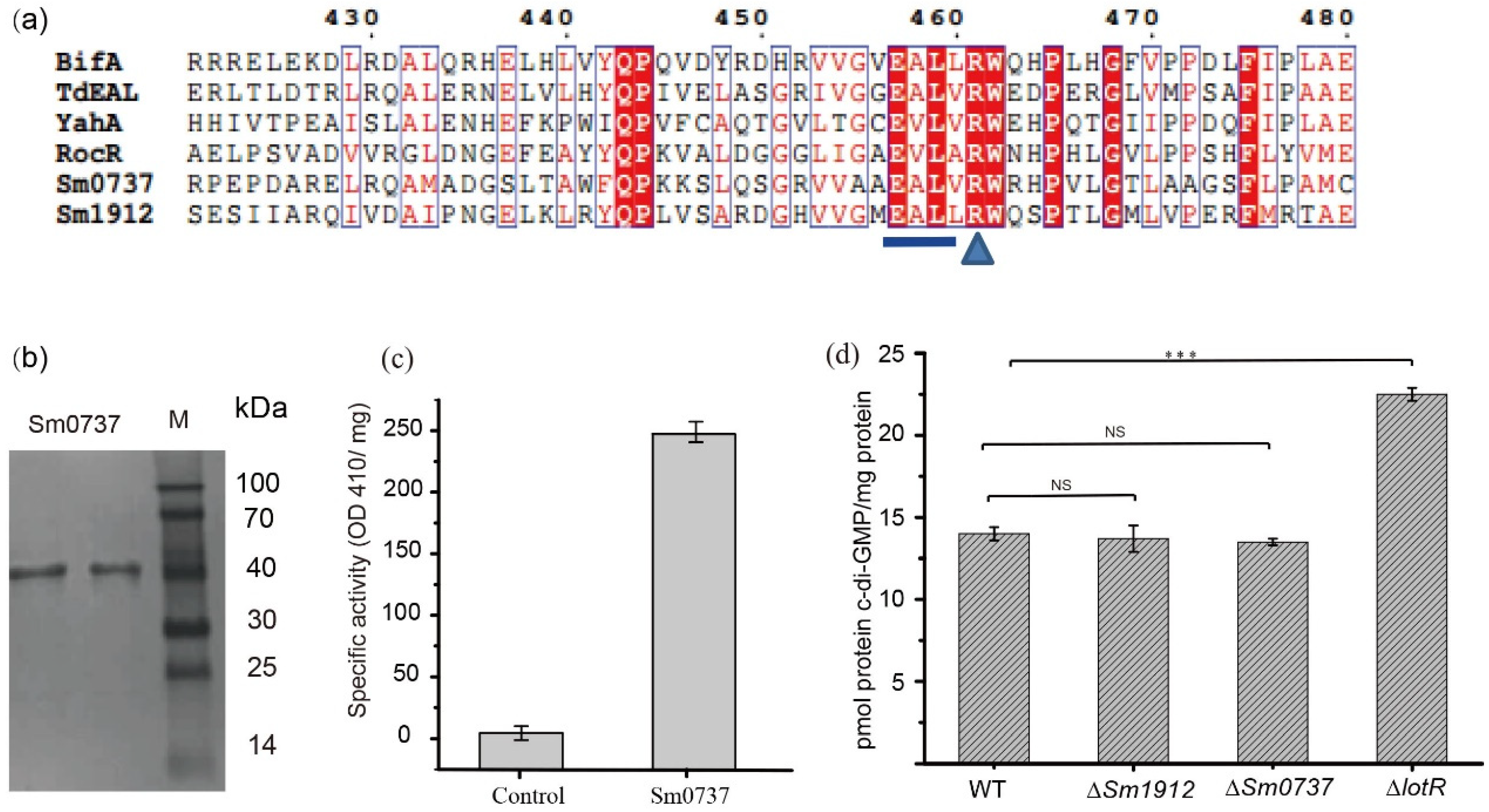
3.7. The Effect of Sm0737 and Sm1912 on Physiological Functions
3.8. Enzymatic Activity Was Not a Vital Contributor to c-di-GMP Levels
3.9. Detection of Intracellular c-di-GMP Levels in WT and Various Deletion Strains
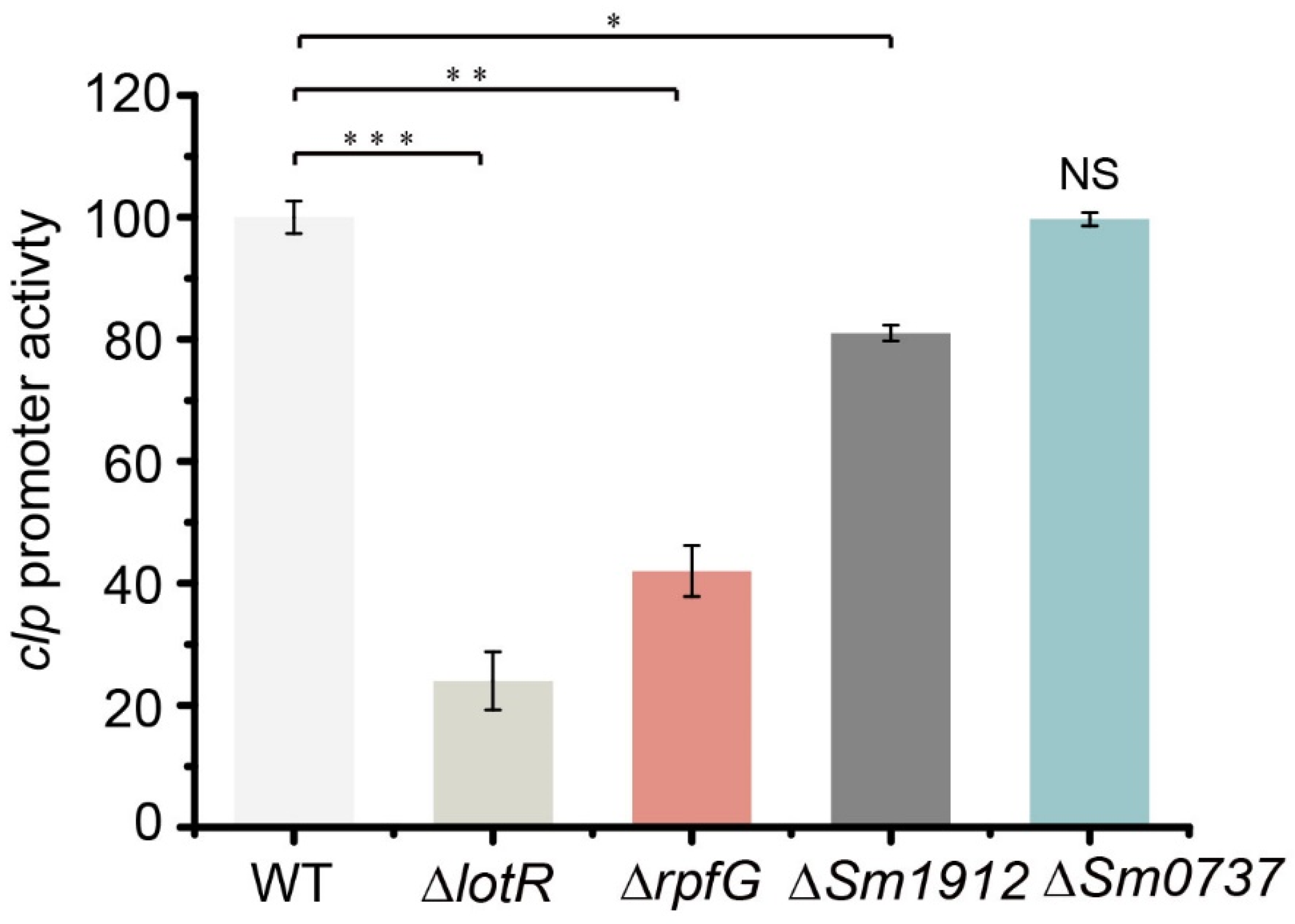
3.10. TCSs Regulated Biofilm Formation in Multiple Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Adamek, M.; Overhage, J.; Bathe, S.; Winter, J.; Fischer, R.; Schwartz, T. Genotyping of environmental and clinical Stenotrophomonas maltophilia isolates and their pathogenic potential. PLoS ONE 2011, 6, e27615. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Wang, K.; Chen, D.; Huang, X.; He, M.; Yin, Z. Stenotrophomonas maltophilia, an emerging opportunist pathogen for cultured channel catfish, Ictalurus punctatus, in China. Aquaculture 2010, 308, 132–135. [Google Scholar] [CrossRef]
- Amin, R.; Waters, V. Antibiotic treatment for Stenotrophomonas maltophilia in people with cystic fibrosis. Cochrane Database Syst. Rev. 2016, 7, CD009249. [Google Scholar] [CrossRef]
- Chemao-Elfihri, M.W.; Manni, A.; Laamarti, M.; Kartti, S.; Essabbar, A.; Alouane, T.; Temsamani, L.; Eljamali, J.E.; Ouadghiri, M.; El Hajjami, N.; et al. Draft genome sequence of Stenotrophomonas maltophilia MDMC339, isolated from soil of merzouga desert in Morocco. Microbiol. Resour. Announc. 2020, 9, e00634-20. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Chen, H.; Song, Z.; Guo, H.; Yuan, Y.; Yue, T. Antibacterial activity of essential oils against Stenotrophomonas maltophilia and the effect of citral on cell membrane. LWT 2020, 117, 108667. [Google Scholar] [CrossRef]
- Rayvathy Balasubramanian, P.-E.F. Thangam Menon. Culture-negative endocarditis caused by Stenotrophomonas maltophilia: A report of two cases. Future Microbiol. 2022, 17, 819–822. [Google Scholar] [CrossRef]
- Nayyar, C.; Thakur, P.; Tak, V.; Saigal, K. Stenotrophomonas maltophilia: An emerging pathogen in paediatric population. J. Clin. Diagn. Res. 2017, 11, DC08–DC11. [Google Scholar] [CrossRef]
- Alsuhaibani, M.; Aljarbou, A.; Althawadi, S.; Alsweed, A.; Al-Hajjar, S. Stenotrophomonas maltophilia bacteremia in children: Risk factors and mortality rate. Antimicrob. Resist. Infect. Control 2021, 10, 19. [Google Scholar] [CrossRef]
- Morte, E.; Tolmos, M.; Halperin, A.; Morosini, M.I.; Canton, R.; Hermida, J.M. Early prosthetic endocarditis caused by Stenotrophomonas maltophilia. Med. Mal. Infect. 2018, 48, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Flores-Trevino, S.; Bocanegra-Ibarias, P.; Camacho-Ortiz, A.; Morfin-Otero, R.; Salazar-Sesatty, H.A.; Garza-Gonzalez, E. Stenotrophomonas maltophilia biofilm: Its role in infectious diseases. Expert Rev. Anti Infect. Ther. 2019, 17, 877–893. [Google Scholar] [CrossRef] [PubMed]
- Millar, B.C.; McCaughan, J.; Rendall, J.C.; Moore, J.E. Infection dynamics of Stenotrophomonas maltophilia in patients with cystic fibrosis. J. Infect. 2023, 86, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Munsch-Alatossava, P.; Alatossava, T. Phenotypic characterization of raw milk-associated psychrotrophic bacteria. Microbiol. Res. 2006, 161, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, J.B.; Kim, M.; Roh, E.; Jung, K.; Choi, M.; Oh, C.; Choi, J.; Yun, J.; Heu, S. Microbiota on spoiled vegetables and their characterization. J. Food Prot. 2013, 76, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, J.; Yuan, Y.; Yue, T. Diversity and characterization of spoilage-associated psychrotrophs in food in cold chain. Int. J. Food Microbiol. 2019, 290, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.B.; Simoes, L.C.; Simoes, M. The effects of emerging environmental contaminants on Stenotrophomonas maltophilia isolated from drinking water in planktonic and sessile states. Sci. Total Environ. 2018, 643, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Mohagheghzadeh, N.; Hashemizadeh, Z.; Khashei, R.; Kholdi, S.; Mohebi, S.; Motamedifar, M. High occurrence of antibiotic resistance and biofilm-formation among Stenotrophomonas maltophilia isolated from a tertiary hospital in Southwest of Iran. Gene Rep. 2020, 21, 100827. [Google Scholar] [CrossRef]
- Molloy, K.; Smith, S.G.; Cagney, G.; Dillon, E.T.; Greene, C.M.; McElvaney, N.G. Characterisation of the major extracellular proteases of Stenotrophomonas maltophilia and their effects on pulmonary antiproteases. Pathogens 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, M.; Iacobino, A.; Prosseda, G.; Fiscarelli, E.; Zarrilli, R.; De Carolis, E.; Petrucca, A.; Nencioni, L.; Colonna, B.; Casalino, M. Stenotrophomonas maltophilia strains from cystic fibrosis patients: Genomic variability and molecular characterization of some virulence determinants. Int. J. Med. Microbiol. 2011, 301, 34–43. [Google Scholar] [CrossRef]
- Ashley, L.; DuMont, N.P.C. Stenotrophomonas maltophilia serine protease stmpr1 induces matrilysis anoikis and protease activated. Infect. Immun. 2017, 85, e00544-17. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; de Sousa, J.M.V.B.; Villa, T.G.; Barros-Velazquez, J. Characterization of biogenic amine-producing Stenotrophomonas maltophilia strains isolated from white muscle of fresh and frozen albacore tuna. Int. J. Food Microbiol. 2000, 57, 19–31. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, G.; Spedicato, I.; D’Antonio, D.; Robuffo, I.; Piccolomini, R. Biofilm formation by Stenotrophomonas maltophilia: Modulation by quinolones, trimethoprim-sulfamethoxazole, and ceftazidime. Antimicrob. Agents Chemother. 2003, 48, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Sameni, F.; Hajikhani, B.; Hashemi, A.; Owlia, P.; Niakan, M.; Dadashi, M. The relationship between the biofilm genes and antibiotic resistance in Stenotrophomonas maltophilia. Int. J. Microbiol. 2023, 2023, 8873948. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.P.; Somers, E.B.; Wong, A.C. Differential biofilm formation and motility associated with lipopolysaccharide/exopolysaccharide-coupled biosynthetic genes in Stenotrophomonas maltophilia. J. Bacteriol. 2006, 188, 3116–3120. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, S.; Nair, H.A.; Lee, K.W.; Ong, J.; Goh, J.Q.; Kjelleberg, S.; Rice, S.A. Pseudomonas aeruginosa PAO1 exopolysaccharides are important for mixed species biofilm community development and stress tolerance. Front. Microbiol. 2015, 6, 851. [Google Scholar] [CrossRef] [PubMed]
- Nagar, E.; Zilberman, S.; Sendersky, E.; Simkovsky, R.; Shimoni, E.; Gershtein, D.; Herzberg, M.; Golden, S.S.; Schwarz, R. Type 4 pili are dispensable for biofilm development in the cyanobacterium Synechococcus elongatus. Environ. Microbiol. 2017, 19, 2862–2872. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.; Liu, H.; Zhang, L.; Guo, Y.; Yu, S.; Wozniak, D.J.; Ma, L.Z. The exopolysaccharide Psl-eDNA interaction enables the formation of a biofilm skeleton in Pseudomonas aeruginosa. Environ. Microbiol. Rep. 2015, 7, 330–340. [Google Scholar] [CrossRef]
- Ming, D.; Wang, D.; Cao, F.; Xiang, H.; Mu, D.; Cao, J.; Li, B.; Zhong, L.; Dong, X.; Zhong, X.; et al. Kaempferol inhibits the primary attachment phase of biofilm formation in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2263. [Google Scholar] [CrossRef]
- Khong, N.Z.; Zeng, Y.; Lai, S.K.; Koh, C.G.; Liang, Z.X.; Chiam, K.H.; Li, H.Y. Dynamic swimming pattern of Pseudomonas aeruginosa near a vertical wall during initial attachment stages of biofilm formation. Sci. Rep. 2021, 11, 1952. [Google Scholar] [CrossRef]
- Kampf, J.; Gerwig, J.; Kruse, K.; Cleverley, R.; Dormeyer, M.; Grunberger, A.; Kohlheyer, D.; Commichau, F.M.; Lewis, R.J.; Stulke, J. Selective pressure for biofilm formation in Bacillus subtilis: Differential effect of mutations in the master regulator SinR on bistability. mBio 2018, 9, e01464-18. [Google Scholar] [CrossRef]
- Obeng, N.; Czerwinski, A.; Schutz, D.; Michels, J.; Leipert, J.; Bansept, F.; Garcia Garcia, M.J.; Schultheiss, T.; Kemlein, M.; Fuss, J.; et al. Bacterial c-di-GMP has a key role in establishing host-microbe symbiosis. Nat. Microbiol. 2023, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Song, L.; Liu, X.; Shen, X.; Li, X. Bacterial second messenger c-di-GMP: Emerging functions in stress resistance. Microbiol. Res. 2023, 268, 127302. [Google Scholar] [CrossRef] [PubMed]
- Ryjenkov, D.A.; Tarutina, M.; Moskvin, O.V.; Gomelsky, M. Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: Insights into biochemistry of the GGDEF protein domain. J. Bacteriol. 2005, 187, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.J.; Ryjenkov, D.A.; Gomelsky, M. The ubiquitous protein domain EAL is a cyclic diguanylate-specific phosphodiesterase: Enzymatically active and inactive EAL domains. J. Bacteriol. 2005, 187, 4774–4781. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Fouhy, Y.; Lucey, J.F.; Crossman, L.C.; Spiro, S.; He, Y.W.; Zhang, L.H.; Heeb, S.; Camara, M.; Williams, P.; et al. Cell-cell signaling in Xanthomonas campestris involves an HD-GYP domain protein that functions in cyclic di-GMP turnover. Proc. Natl. Acad. Sci. USA 2006, 103, 6712–6717. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ji, F.; Guo, J.; Wang, Y.; Li, Y.; Wang, J.; An, L.; Bao, Y. LotS/LotR/Clp, a novel signal pathway responding to temperature, modulating protease expression via c-di-GMP mediated manner in Stenotrophomonas maltophilia FF11. Microbiol. Res. 2018, 214, 60–73. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, J.; An, L.; Bao, Y. Integration of DSF and temperature signals for RpfC/RpfG Two-component system modulating protease production in Stenotrophomonas maltophilia FF11. Curr. Microbiol. 2022, 79, 54. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Huang, Y.W.; Hu, R.M.; Chiang, K.H.; Yang, T.C. The role of AmpR in regulation of L1 and L2 beta-lactamases in Stenotrophomonas maltophilia. Res. Microbiol. 2009, 160, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, A.; Avison, M.B. Induction of L1 and L2 beta-lactamase production in Stenotrophomonas maltophilia is dependent on an AmpR-type regulator. Antimicrob. Agents Chemother. 2008, 52, 1525–1528. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Musken, M.; Di Fiore, S.; Romling, U.; Haussler, S. A 96-well-plate–based optical method for the quantitative and qualitative evaluation of Pseudomonas aeruginosa biofilm formation and its application to susceptibility testing. Nat. Protoc. 2010, 5, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Yao, X.; Duan, M.; Luo, Y.; Liu, B.; Qi, P.; Sun, M.; Ruan, L. Two overlapping two-component systems in Xanthomonas oryzae pv. oryzae contribute to full fitness in rice by regulating virulence factors expression. Sci. Rep. 2016, 6, 22768. [Google Scholar] [CrossRef] [PubMed]
- He, Y.W.; Boon, C.; Zhou, L.; Zhang, L.H. Co-regulation of Xanthomonas campestris virulence by quorum sensing and a novel two-component regulatory system RavS/RavR. Mol. Microbiol. 2009, 71, 1464–1476. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Wang, X.; Chen, H.; Yuan, Y.; Yue, T. Distribution of cold-resistant bacteria in quick-frozen dumpling and its inhibition by different antibacterial agents. J. Food Process Preserv. 2020, 44, e14710. [Google Scholar] [CrossRef]
- Naila, A.; Flint, S.; Fletcher, G.; Bremer, P.; Meerdink, G. Control of biogenic amines in food--existing and emerging approaches. J. Food Sci. 2010, 75, R139–R150. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Xie, J. Assessment of spoilage potential and amino acids deamination & decarboxylation activities of Shewanella putrefaciens in bigeye tuna (Thunnus obesus). LWT 2022, 156, 113016. [Google Scholar] [CrossRef]
- Zhuang, S.; Liu, X.; Li, Y.; Zhang, L.; Hong, H.; Liu, J.; Luo, Y. Biochemical changes and amino acid deamination & decarboxylation activities of spoilage microbiota in chill-stored grass carp (Ctenopharyngodon idella) fillets. Food Chem. 2021, 336, 127683. [Google Scholar] [CrossRef]
- Yang, S.-P.; Xie, J.; Cheng, Y.; Zhang, Z.; Zhao, Y.; Qian, Y.-F. Response of Shewanella putrefaciens to low temperature regulated by membrane fluidity and fatty acid metabolism. LWT 2020, 117, 108638. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, L.; Lu, H.; Zhu, J.; Kumar, V.; Liu, X. Transcriptomic analysis of the food spoilers Pseudomonas fluorescens reveals the antibiofilm of carvacrol by interference with intracellular signaling processes. Food Control 2021, 127, 108115. [Google Scholar] [CrossRef]
- Cen, C.; Wang, X.; Li, H.; Chen, J.; Wang, Y. An inhibitor of the adaptability of Pseudomonas fluorescens in a high-salt environment: Phenomenon and mechanism of inhibition. Int. J. Food Microbiol. 2024, 412, 110553. [Google Scholar] [CrossRef]
- Sondermann, H.; Shikuma, N.J.; Yildiz, F.H. You’ve come a long way: c-di-GMP signaling. Curr. Opin. Microbiol. 2012, 15, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Yan, J.; Ding, Z.; Xie, J. The HD-GYP domain protein of Shewanella putrefaciens YZ08 regulates biofilm formation and spoilage activities. Food Res. Int. 2022, 157, 111466. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, C.; Jiang, W.; Wang, L.; Li, C.; Wang, Y.; Dow, J.M.; Sun, W. The HD-GYP domain protein RpfG of Xanthomonas oryzae pv. oryzicola regulates synthesis of extracellular polysaccharides that contribute to biofilm formation and virulence on rice. PLoS ONE 2013, 8, e59428. [Google Scholar] [CrossRef] [PubMed]
- Rott, P.; Fleites, L.A.; Mensi, I.; Sheppard, L.; Daugrois, J.H.; Dow, J.M.; Gabriel, D.W. The RpfCG two-component system negatively regulates the colonization of sugar cane stalks by Xanthomonas albilineans. Microbiology 2013, 159 Pt 6, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Q.; Wu, Y.; Cai, Z.; Qian, W. BDSF Is a degradation-prone quorum-sensing signal detected by the histidine kinase RpfC of Xanthomonas campestris pv. campestris. Appl. Environ. Microbiol. 2022, 88, e0003122. [Google Scholar] [CrossRef]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J. Biol. Chem. 2005, 280, 30829–30837. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Description | Reference or Source |
|---|---|---|
| Strains | ||
| Stenotrophomonas moltophilia FF11 | Wild-type(FF11) | Lab collection |
| ∆lotR or ∆lotS mutant | lotR or lotS gene deletion mutant derived from FF11, Cmr | Previous study |
| ∆rpfG or ∆rpfC mutant | rpfG or rpfC gene deletion mutant derived from FF11, Cmr | Previous study |
| ∆Sm1912 mutant | Sm1912 gene deletion mutant derived from FF11, Cmr | This study |
| ∆Sm0737 mutant | Sm0737 gene deletion mutant derived from FF11, Cmr | This study |
| ∆lotR (C) or ∆lotS (C) | Complement of lotR in ∆lotR mutant, Te | This study |
| ∆rpfG (C) or ∆rpfC (C) | Complement of rpfG in ∆rpfG mutant, Te | This study |
| ∆Sm1912 (C) | Complement of Sm1912 in ∆Sm1912 mutant, Te | This study |
| ∆Sm0737 (C) | Complement of Sm0737 in ∆Sm0737 mutant, Te | This study |
| E. coli | ||
| JM109 | RecA1, endA1, gyrA96, thi, supE44, relA1, Δ(lac-proAB)/F′ traD36, lacI q, lacZ | Novagen |
| S17-1 λ pir | λ pir lysogen of S17-1 [thi pro hsdR2 hsdM+ recA RP4 2-Tc::Mu-Km::Tn7 (Tpr Smr)]; permissive host able to transfer suicide plasmids requiring the pir protein by conjugation to recipient cells | Novagen |
| Plasmids | ||
| pEX18Tc | Low copy number plasmid; For gene knockout; Tc | HonorGene |
| pLAFR3 | Broad host range cloning vector, RK2 replicon, Mob+, Tcr | Bio Sci |
| Primers | Sequence (5′ to 3′) |
|---|---|
| 1912 up F | CGCGGATCCCTGGAAACCAGCAACCGCGAG |
| 1912 up R | GGGGTGGTGCTGCCGCGACCACGCAGGTAGGAC |
| 1912 cmr F | GGTCGCGGCAGCACCACCCCGTCAGTAGCTGAA |
| 1912 cmr R | ATGTGCTTGCTTACGCCCCGCCCTGCCACTCAT |
| 1912 down F | CGGGGCGTAAGCAAGCACATCCACCAGGCGTTC |
| 1912 down R | CCCAAGCTTTGCCGTCGGAGTTGAACCAGGTC |
| 0737 up F | CCGGAATTCATGACCCAGCGTGTCCTGATCCT |
| 0737 up R | GGGGTGGTGCCGCAACAGCTCGCCTTCCAG |
| 0737 cmr F | AGCTGTTGCGGCACCACCCCGTCAGTAGCTGAA |
| 0737 cmr R | GCGTTTCATCTTACGCCCCGCCCTGCCACTCAT |
| 0737 down F | CGGGGCGTAACTTCCGGGTGCCGGTTTCCATC |
| 0737 down R | CGCGGATCCACGGCCCGTGGCGTTTCATC |
| pLAFR3-0737 F | CATGCCATGGTGACCCAGCGTGTCCTGATC |
| pLAFR3-0737 R | CCGCTCGAGTCAACGCAGGACGGCCCGTG |
| pL6rmlA-gusA F | CCGGAATTCTACTGGGTGGTCTTGAAGGTGTCG |
| pL6rmlA-gusA R | CGGGGTACCTGTGGGCACTACCTGTTCTCCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.; Liao, M.; Wang, Q. Co-regulation of Thermosensor Pathogenic Factors by C-di-GMP-Related Two-Component Systems and a cAMP Receptor-like Protein (Clp) in Stenotrophomonas maltophilia. Foods 2024, 13, 1201. https://doi.org/10.3390/foods13081201
Ding J, Liao M, Wang Q. Co-regulation of Thermosensor Pathogenic Factors by C-di-GMP-Related Two-Component Systems and a cAMP Receptor-like Protein (Clp) in Stenotrophomonas maltophilia. Foods. 2024; 13(8):1201. https://doi.org/10.3390/foods13081201
Chicago/Turabian StyleDing, Jieqiong, Minghong Liao, and Qingling Wang. 2024. "Co-regulation of Thermosensor Pathogenic Factors by C-di-GMP-Related Two-Component Systems and a cAMP Receptor-like Protein (Clp) in Stenotrophomonas maltophilia" Foods 13, no. 8: 1201. https://doi.org/10.3390/foods13081201