
KCa2 and KCa3.1 Channels in the Airways: A New Therapeutic Target
by
 ,
,
Razan Orfali
1,2,
Ali AlFaiz
2,
Mohammad Asikur Rahman
1,
Liz Lau
1,
Young-Woo Nam
1,* and
Miao Zhang
1,* 1
Department of Biomedical and Pharmaceutical Sciences, Chapman University School of Pharmacy, Irvine, CA 92618, USA
2
Biomedical Research Administration, Research Centre, King Fahad Medical City, Riyadh Second Health Cluster, Riyadh 12231, Saudi Arabia
*
Authors to whom correspondence should be addressed.
Biomedicines 2023, 11(7), 1780; https://doi.org/10.3390/biomedicines11071780
Submission received: 22 May 2023
/
Revised: 8 June 2023
/
Accepted: 13 June 2023
/
Published: 21 June 2023
(This article belongs to the Special Issue Molecular Mechanisms, Diagnoses, and Treatments of Respiratory Diseases)
Abstract
:K+ channels are involved in many critical functions in lung physiology. Recently, the family of Ca2+-activated K+ channels (KCa) has received more attention, and a massive amount of effort has been devoted to developing selective medications targeting these channels. Within the family of KCa channels, three small-conductance Ca2+-activated K+ (KCa2) channel subtypes, together with the intermediate-conductance KCa3.1 channel, are voltage-independent K+ channels, and they mediate Ca2+-induced membrane hyperpolarization. Many KCa2 channel members are involved in crucial roles in physiological and pathological systems throughout the body. In this article, different subtypes of KCa2 and KCa3.1 channels and their functions in respiratory diseases are discussed. Additionally, the pharmacology of the KCa2 and KCa3.1 channels and the link between these channels and respiratory ciliary regulations will be explained in more detail. In the future, specific modulators for small or intermediate Ca2+-activated K+ channels may offer a unique therapeutic opportunity to treat muco-obstructive lung diseases.

1. Introduction
The epithelial surface of the respiratory tract between the nose and the alveoli is constantly exposed to potentially harmful pathogens, particulates, and gaseous materials [1,2,3]. In response to these challenges, the human body utilizes a series of defense mechanisms to protect the airways, and the primary defense mechanism in the lung is mucociliary clearance (MCC) [1,4]. MCC is a process of specialized organelles called cilia that beat in metachronal waves to impel pathogens and particles trapped by the mucous layer out of the airways. Cilia within the mucociliary system present critical functions in human health; abnormalities in each compartment of the mucociliary system could compromise the mucus clearance process and lead to chronic lung disease [2,3]. Mucociliary dysfunction is commonly associated with chronic airway diseases, and it is one of the pathological observations in patients with cystic fibrosis, primary ciliary dyskinesia, chronic bronchitis, and asthma [5,6]. Airway diseases with associated mucociliary dysfunction remain largely unaddressed, despite the therapeutic progress in treating inflammatory lung diseases [5].
The lung’s lining is covered by a thin layer of fluid called airway surface liquid (ASL); it separates the airway epithelium’s luminal surface from the external environment. ASL is mainly composed of water, electrolytes, and mucins; it is essential for normal airway function, particularly for proper MCC [3,7,8]. ASL epithelia contain various cell types with distinct morphologies and functions. Of the cell population in the trachea, approximately 60% are ciliated cells; these cells also retain other important roles other than coordinating ciliated movements, such as regulating ion transfer [1,9].
There are detections of over 30 diverse K+ channels in the airway epithelia, and these K+ channels maintain the electrochemical gradient and support lung ion and fluid homeostases [1,10,11,12]. A large portion of airway chloride secretion occurs through the apically located bicarbonate and chloride channels [10]. K+ channels are involved in many vital functions in lung physiology, such as oxygen sensing, inflammatory responses, enhancing Cl− transport, and respiratory epithelia repair [9,13]. The basolateral K+ channel has known regulation effects on Na+ absorption; reduced Na+ absorption in the lung shows improvement in muco-obstructive disease. A large portion of airway chloride secretion occurs through the apically located bicarbonate and chloride channels, significantly influenced by some Ca2+-activated K+ channels (KCa) that are located apically in the lung [10]. Hence, the specific K+ group KCa also regulates MCC and ASL volumes [1]. The small-conductance KCa2 channels and intermediate-conductance KCa3.1 channels are voltage-independent and activated solely by the elevation of the intracellular Ca2+ concentration. In this context, we will discuss current knowledge of the functional roles of KCa2 and KCa3.1 channels in the respiratory tract, focusing on their physiological roles in respiratory diseases.
2. Introduction to KCa Channels
There are several kinds of K+ channels present in the respiratory epithelium lining airways, and the most indispensable K+ channels in airway epithelial cells are the Ca2+-activated K+ channels. They serve as the cell crossroad where Ca2+ influx, other ion outfluxes, and membrane potential, all processes governed by KCa channels, integrate to modulate an extensive array of cellular processes [14]. KCa channels are subdivided into three major groups, according to their single-channel conductance: large conductance (150–300 pS) K+ channels (BK or KCa1.1), small conductance (2–20 pS) K+ channels (SK or KCa2), and intermediate conductance (20–60 pS) K+ channels (IK or KCa3.1) [15,16,17]. Each group has specific distinct biophysical and pharmacological properties [18]. KCa2.x and KCa3.1 channels are voltage-independent and activated exclusively by intracellular Ca2+ via the calmodulin (CaM) that is typically bound to these channels and serves as their Ca2+ sensor [19]. KCa2x and KCa3.1 channels, before their cloning, were referred to as small-conductance (SK) or intermediate-conductance (IK) Ca2+-activated K+ channels, based on their singular conductance of ~10 pS or ∼40 pS in symmetrical solutions to differentiate them from the large-conductance potassium (BK) channel [19,20].
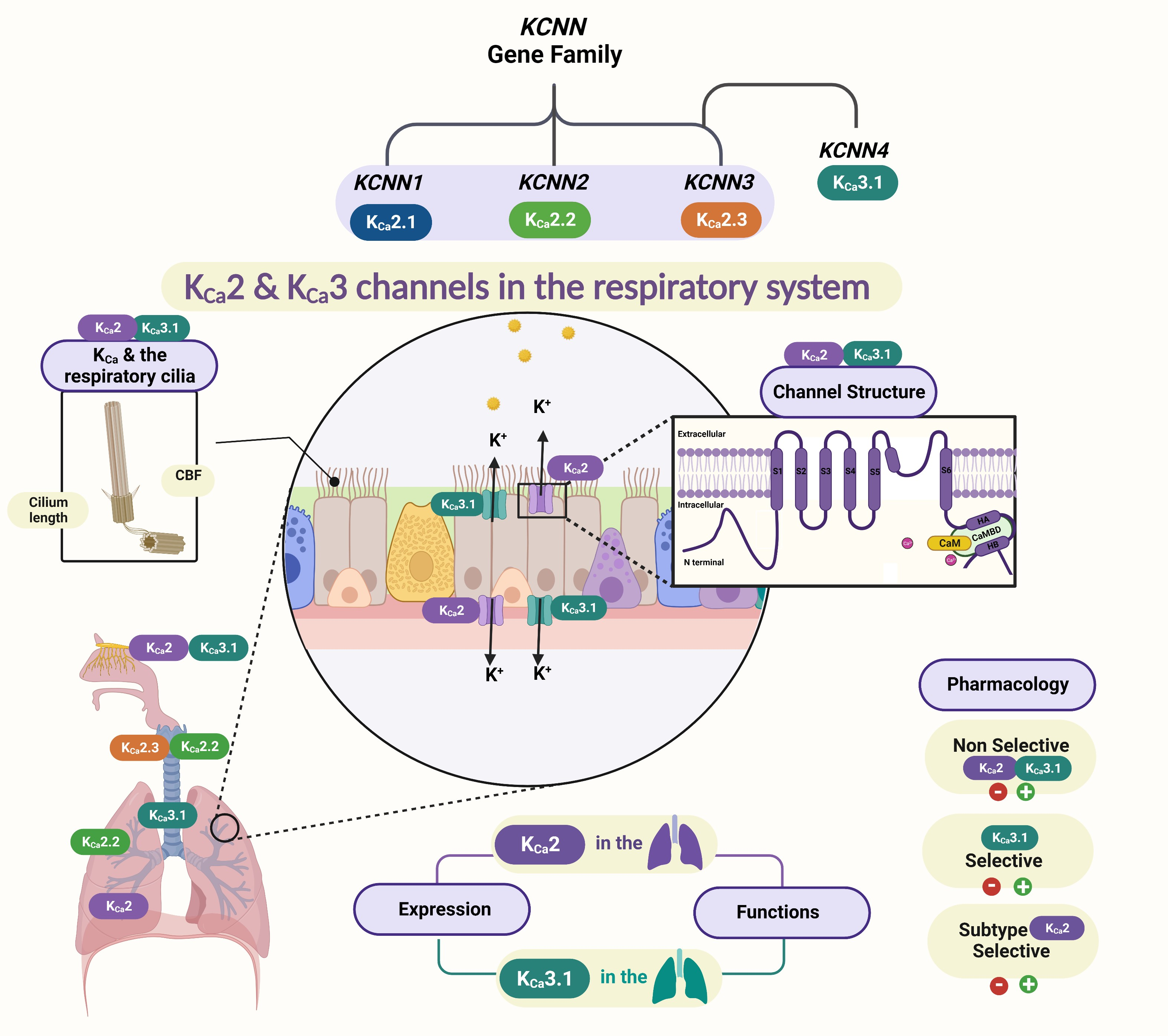
Four mammalian KCNN channel subtypes are encoded by the KCNN genes, including KCNN1 for KCa2.1, KCNN2 for KCa2.2, KCNN3 for KCa2.3 [21], and KCNN4 for KCa3.1 [22], respectively [23] (Table 1).
KCa2 and KCa3.1 Channel Structures
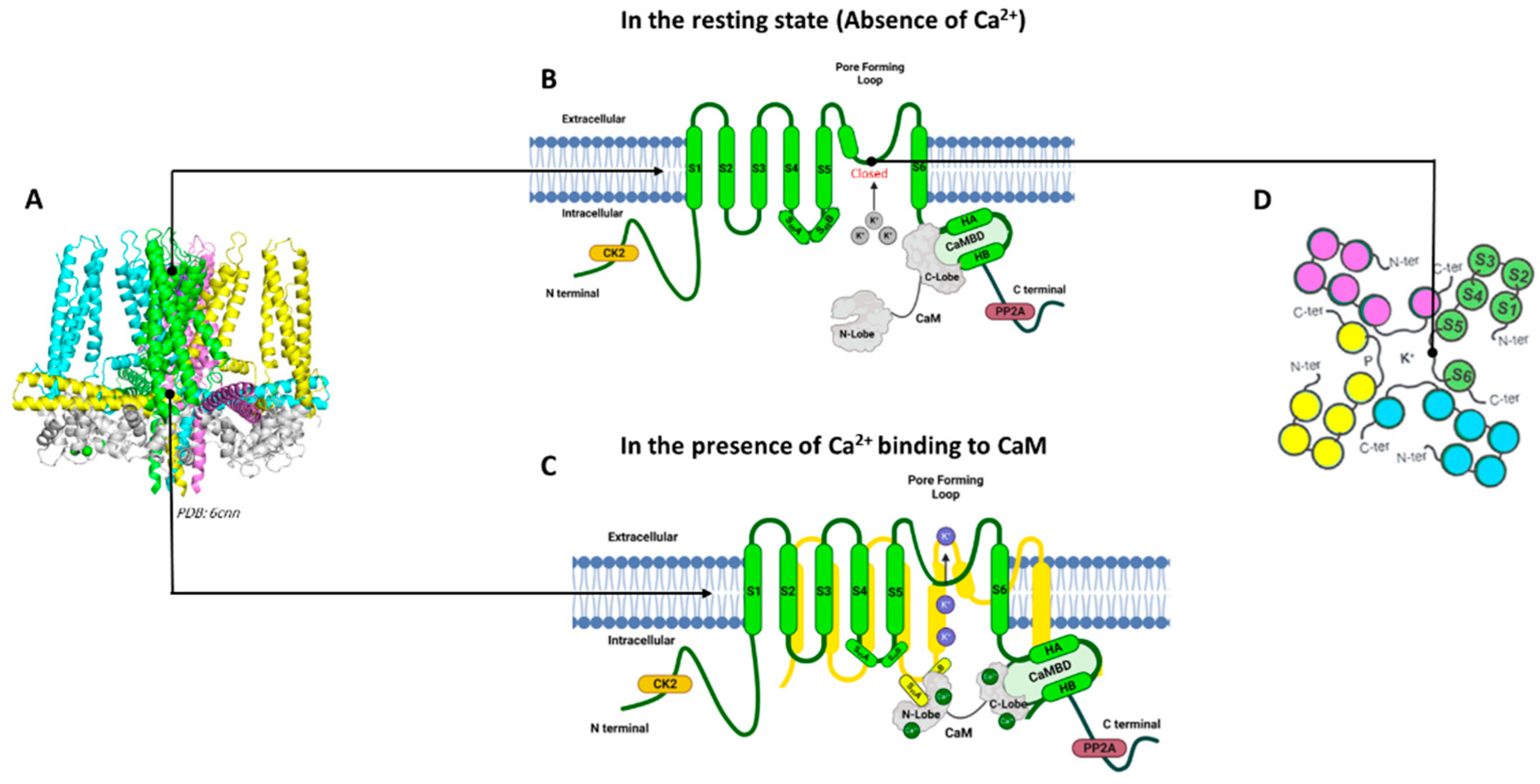
KCa2 and KCa3.1 channels are assembled as homotetramers of four α-subunits; each subunit is composed of six transmembrane α-helical domains denoted as S1–S6 (Figure 1). The selectivity filter within the channel pore between the S5 and S6 transmembrane domains is responsible for the selective permeability of the K+ ions [30,34]. The KCa2/KCa3.1 channel subtypes are highly homologous in their six transmembrane domains, but the amino acid sequences and lengths at their cytoplasmic N- and C-termini differ among the subtypes (Table 1) [37].
Among the four KCa2/KCa3.1 channel subtypes, the full-length cryogenic electron microscopy (cryo-EM) structure is only available for the KCa3.1 channel determined in the absence and presence of Ca2+, providing insight into the Ca2+/CaM gating mechanism for these channels [36]. The calmodulin-binding domain consists of two α-helices, HA and HB, whereas the S4–S5 linker includes two α-helices, S45A and S45B. The HA and HB helices from one channel subunit, the S4–S5 linker from a neighboring channel subunit, and calmodulin closely interact with each other (Figure 1). When Ca2+ is absent, the C-lobe of CaM binds to the HA/HB helices in the proximal channel C-terminus, the N-lobe of CaM is highly flexible, and the channel pore is closed (Figure 1B). In the presence of Ca2+, the N-lobe of CaM becomes well-structured and interacts with the linker between the S4 and S5 transmembrane domains (S4–S5 linker) of a neighboring α-subunit. The interaction between the Ca2+-bound CaM N-lobe and the S4–S5 linker causes the movement of the S6 transmembrane domain and the opening of the channel pore (Figure 1C) [33,38].
KCa2 channels are activated by Ca2+, with EC50 values ranging from 300 to 750 nM, whereas KCa3.1 channels exhibit apparent Ca2+ sensitivities of 100–400 nM [29,34,39]. KCa2 and KCa3.1 channels, therefore, play a critical role in the physiologies of various tissues and disease states [22,40]. The advances in understanding the KCa3.1 structure [36] (the cryo-electron microscopy of the human homotetrameric KCNN4 channel) and the resulting improvements in other KCa2 subtypes modeling [29,30,34] have yet to be used, not only for drug discovery but also for understanding the pathophysiological diseases.
3. KCa Channels in the Respiratory System
The involvement of K+ channels has been proposed in respiratory conditions such as asthma, chronic obstructive pulmonary diseases (COPD), and cystic fibrosis (CF) [1,12]. In airway epithelial cells, both Cl− and K+ transports rely, to some extent, on Ca2+-dependent channel activity (e.g., KCa channels) [1]. KCa channels are important in regulating Cl− secretion, MCC, and ASL volumes. KCa3.1 and KCa2 channel subtypes located in the airway epithelia, such as KCa2.1 [41] and KCa2.3 [42], maintain the electrochemical gradient and thus support lung ion and fluid homeostasis [1]. Table 2 summarizes the KCNN gene family, tissue distribution, physiological roles, and their roles in the lungs.
3.1. KCa Channels and the Respiratory Cilia
The KCa2 and KCa3.1 channels are tetramers, and each subunit comprises six transmembrane alpha-helical domains (six TMD), indicated as S1–S6 in each channel subunit. The selectivity of potassium ions across these channels is based on the pore-forming P-loop between the transmembrane S5 and S6 domains. KCa2/KCa3.1 are more sensitive to Ca2+ due to calmodulin CaM acting as a Ca2+ sensor (Figure 1) [26,47]. CaM is present in all eukaryotic cells, facilitating various cellular signaling processes, such as the modulation of ion channel actions, regulation of enzymatic activities, and gene expression [14,48]. The ciliary beat of the airway epithelium is believed to be regulated by the level of intracellular Ca2+ [49]. The association with calmodulin in the regulation of ciliary beats has been reported as the most important intraciliary Ca2+ binding protein [49,50]. Moreover, the activation of KCa2 channels in non-excitable cells, such as epithelial cells, increases Ca2+ entry through non-voltage-gated Ca2+ channels, thereby increasing intracellular Ca2+ concentration [51]. This elevation of intracellular Ca2+ is one of the primary regulators of ciliary movement [52]. Thus, KCa2 and KCa3.1 channels will regulate respiratory ciliary activities as part of a complex signaling network.
3.1.1. KCa Channels and Ciliary Beat Frequency
In vitro measurements of the changes in the CBF of human respiratory cells indicate that Ca2+ ionophore speeds the CBF of human respiratory cells mediated through a calmodulin-sensitive system [53]. Airway epithelial cells contain 100 nM of free Ca2+ in their cytoplasm, but ciliated cells bear a higher concentration at baseline than club cells [54]. This supports the idea that KCa2 channels may be active during normal conditions in specific airway cells, as these channels show a high sensitivity to Ca2+ (Table 2). Significantly, in CF mouse airways, a previous study by Vega et al. [5] determined that KCNN4-silencing enhanced MCC when Na+ absorption was decreased. Additionally, CBF was also increased by KCa3.1 inhibition. An explanation is that KCa3.1 inhibition reduces Na+ absorption in CF, thereby increasing CBF speeds by hyperpolarizing the apical membrane [5,55].
3.1.2. KCa2 Channels and Cilium Length
Muco-obstructive lung disease is considered the primary cause of morbidity and is responsible for 80% of mortality [55]. The presence of KCa2 channels in a human bron-chial epithelial cell, and structural similarities in the groups of KCa2 and KCa3.1, pro-vides a new direction in the investigating the expression and function of KCa2 channel subtypes in the ciliated human lung epithelial cells. Optimal MCC requires mucus, cilia, and a thin layer of ASL to facilitate ciliary beating [2]. Maintaining a normal range of respiratory cilia length (4 to 7 μm, depending on the airway region) is critical for adequate mucociliary clearance [56]. A qualitative difference exists between short and longer cilia waveform shapes [57], and various acquired lung disorders are marked by abnormalities in both cilia structure and function [56]. Our previous work determined the critical role of KCa2.3 channels in regulating the primary cilia in endothelial cells [58]. Taking advantage of the previous results could help to connect KCa2 channels and respiratory cilia, two crucial components in the Ca2+ signaling network of airway epithelial and smooth muscle cells, with potential implications in the pathogenesis of airway diseases.
4. Expression and Physiological Functions of KCa2 and KCa3.1 Channels in the Airways
Many human cells express KCa channels that have the exceptional ability to trans-late changes in the level of the intracellular second messenger, Ca2+, to changes in membrane K+ conductance and, thus, resting potential membrane. While KCa channel subtypes are all regulated by intracellular Ca2+, they are otherwise quite distinct entities, differing in tissue distribution and functions [59]. KCa2 channel subtypes, for example, are widely expressed in the nervous system, where they are involved in regulating the firing frequency of various neurons. On the other hand, the KCa3.1 channel subtype is expressed in peripheral cells, including the erythrocytes and lymphocytes, and has been determined in numerous cancer cells where they have been implicated in growth control [60,61]. Here we demonstrate the expressions and physiological roles of KCa2 and KCa3.1 channels in the airways.
4.1. Expression and Functions of KCa2 in the Respiratory Epithelia
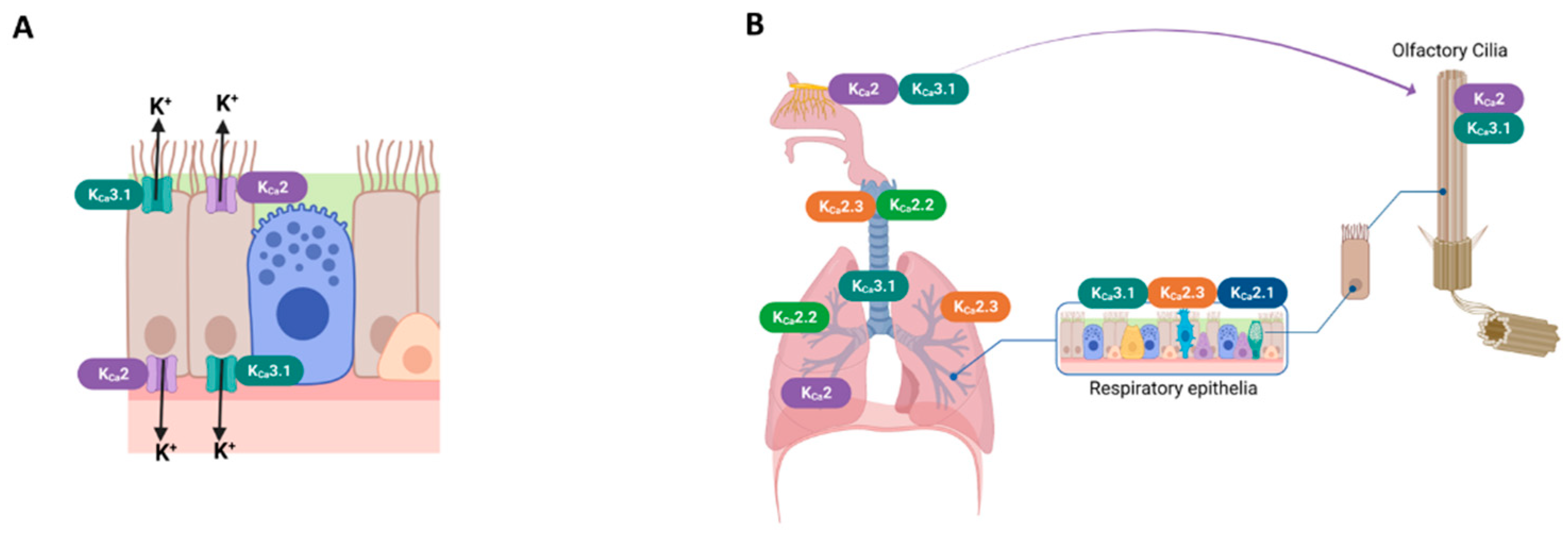
KCa2 channels are widely expressed in various tissues and play an important role in modulating excitable and non-excitable cells. The presence of KCa channel groups was confirmed at the apical and basolateral membranes of airway epithelial cells [1,62] (Figure 2A). The bronchial epithelium expresses KCa2.1 and KCa2.3 channel subtypes[35,41]. KCa2.2 and KCa2.3 mRNA were detected in the lungs and trachea [3,4]. KCa2.2 and KCa2.3 mRNA were detected in lungs and trachea [6]. KCa2.3 is the only subtype expressed in the pulmonary artery [5]. Figure 2-B shows the major expression sites of KCa2 and KCa3.1 channel subtypes in the airway.
Different ion channels seem to be present in motile cilia [63]. In the nasal cavity, olfactory receptor neurons (ORNs) are adapted to grow various long cilia; they are not motile but can move with the liquid stream of the nasal mucosa to sample odorants entering the nose. The presence of KCa channel groups in the cilia of ORNs was reported [64].
Figure 2.
Expression sites of KCa2 and KCa3.1 channels in the respiratory system. (A) Schematic drawing of ciliated airway epithelial cells of KCa2 and KCa3.1 channels. (B) KCa2 and KCa3.1 channels were expressed in airway smooth muscle [65], airway olfactory nerves [66] and olfactory cilia [64]; KCa2.2 and KCa2.3 subtypes presented in the lungs and the trachea [45]; and KCa3.1, KCa2.1, and KCa2.3 subtypes presented in the respiratory epithelia [41,42]. The pulmonary artery expressed the KCa2.3 subtype [42].
Figure 2.
Expression sites of KCa2 and KCa3.1 channels in the respiratory system. (A) Schematic drawing of ciliated airway epithelial cells of KCa2 and KCa3.1 channels. (B) KCa2 and KCa3.1 channels were expressed in airway smooth muscle [65], airway olfactory nerves [66] and olfactory cilia [64]; KCa2.2 and KCa2.3 subtypes presented in the lungs and the trachea [45]; and KCa3.1, KCa2.1, and KCa2.3 subtypes presented in the respiratory epithelia [41,42]. The pulmonary artery expressed the KCa2.3 subtype [42].

The involvement of K+ channels has been proposed in respiratory conditions such as asthma, chronic obstructive pulmonary diseases (COPD), and cystic fibrosis (CF) [1,12]. In airway epithelial cells, both Cl− and K+ transports rely, to some extent, on Ca2+-dependent channel activity (e.g., KCa channels) [1]. KCa channels are important in regulating Cl− secretion, MCC, and ASL volumes. KCa2 channel subtypes located in the airway epithelia, such as KCa2.1 [41] and KCa2.3 [42], maintain the electrochemical gradient and thus support lung ion and fluid homeostases [1].
In CF, the equilibrium between Na+ absorption and Cl− secretion throughout the airway epithelia is necessary to maintain adequate ASL volume and MCC. The Cl− secretion in the lungs involves several steps, starting from Cl− entry through a basolateral channel cotransporter, followed by its exit via apical Cl− channels, such as the cystic fibrosis transmembrane conductance regulator (CFTR) [3]. The dysfunction of CFTR channels in CF results in decreased Cl− and fluid secretions and increased Na+ absorption, leading to inefficient mucociliary clearance and mucus accumulation [67].
In COPD, KCa channel groups can also act as oxygen sensors for lung diseases, such as COPD associated with pulmonary hypertension [12]. In COPD, pulmonary hypertension is generally believed to be due to hypoxic pulmonary vasoconstriction [68]. KCa channels potentiated by the low partial pressure of oxygen (PO2) have been investigated in cerebral resistance myocytes [69]. When hypoxia occurs, KCa channels activate (preventing repolarization) and relax the pulmonary arteries [70,71].
KCa channels were proposed as new targets for bronchodilator therapy for chronic diseases such as asthma and COPD [72]. The mentioned COPD-related studies [69,70,71,72] examined KCa channel groups in general. Though one study suggested that human pulmonary artery and bronchial relaxations might be mediated by pharmacological activation of the KCa2.3 channel subtypes [42] (Table 2).
In anosmia, Odorant-induced K+ conductance is activated by Ca2+ [73], and the elevation of intracellular Ca2+ is often associated with odorant stimulation in some vertebrates and human olfactory neurons [51,74]. Olfactory Receptor Neurons (ORNs) are located in the nasal epithelia and exhibit spontaneous action potential firing. All KCa channel groups have been detected in olfactory cilia [52], and the electrophysiological of the whole-cell results confirmed that KCa channels participate in inhibitory chemo-transduction in the cilia [75]. According to these findings, an apical Ca2+ influx opens the KCa channels, causing membrane hyperpolarization in response to Ca2+ influx and thus triggering the inhibition [74].
Moreover, a Ca2+ channel blocker, nifedipine, was tested on odorants that induce an inhibitory current in olfactory neurons [74]. This drug effectively abolished the outward current and stimulated the cells with an odorant solution free of nifedipine, and the response was restored [74]. KCa2 channel subtypes which are completely Ca2+-dependent and voltage-independent may play a critical role in treating certain diseases, given the drugs that could target specific ion channels. For example, anosmia could be treated by targeting the KCa2 channel subtypes in olfactory cilia and testing their allosteric modulators [52]. However, the pharmacology of these channels in olfactory neurons has not been fully characterized.
4.2. Expression and Functions of KCa3.1 in the Respiratory Epithelia
The expression of Ca2+-activated potassium (KCa) channels often correlate positively with cell proliferation. As an example, the expression of KCa3.1 increases 4-fold upon T-lymphocyte activation, and this channel is inhibited with the specific inhibitor that inhibits T-lymphocyte proliferation [76]. This is because the KCa3.1 channel contributes to electrochemical gradients for Ca2+ influx, which is critical for the proliferation of the T cells [77]. KCa3.1 is also broadly expressed in other cells of the immune system, such as B cells, macrophages, microglia, and mast cells [78]. The major function of KCa3.1 in immune cells is to hyperpolarize the cell membrane and create the driving force for calcium entry, which is necessary for proliferation, activation, and cytokine production [79]. Previous findings [80] suggest that antigen sensitization up-regulates KCa3.1 expression, which may contribute to enhancing cell migration in response to lymphatic chemokines, particularly in the immunogenic lung dendritic cells subset. Therefore, targeting KCa3.1 crucial for controlling allergic airway inflammation [81] (Table 2 and Figure 2).
KCa channels have been found to be involved in regulating smooth muscle responses to both contractile and relaxant agonists that elevate intracellular Ca2+ [82]. Phenotypic modulation of smooth muscle cells is accompanied by changes in KCa3.1 channel expression characterizing “proliferative” cells [83]. KCa3.1 channels regulate the proliferative responses of vascular smooth muscle cells, fibroblasts, endothelial cells, and T lymphocytes, as well as a some transformed cell types [61,84]. KCa3.1 function is increased by protein kinase A (PKA) [85] and nucleoside diphoshate kinase B (NDPK-B) and inhibited by the histidine phosphatase PHPT1 [86,87]. Since NDPK-B and PHPT1 directly phosphorylate or dephosphorylate KCa3.1 on histidine in the C-terminus, KCa3.1 modulation in mammals is one of the rare examples of histidine kinase/phosphatase regulating a biological process [86].
In allergic lung diseases, KCa3.1 channels regulate Ca2+ entry into cells and thereby modulate Ca2+-signaling processes. The entry of positively charged Ca2+ into the cells depolarizes the membrane, which limits its own ability to enter the cell through some types of Ca2+ channels that are closed at more positive membrane potentials. KCa3.1 activation by elevated intracellular Ca2+ maintains a negative membrane potential, which helps to sustain Ca2+ entry into the cell. KCa3.1-mediated elevation of intracellular Ca2+ is necessary for the production of inflammatory chemokines and cytokines by T cells, mast cells, and macrophages [79,88]. Indeed, proliferation is accompanied by the transcriptional up-regulation of functional KCa3.1 expression and can be inhibited by KCa3.1 inhibitors [86]. It has been reported that the use of KCa3.1 blockers can provide a potential therapeutic target for mast cell-mediated diseases such as asthma [88]. Moreover, blocking KCa3.1 may offer a novel approach to treating idiopathic pulmonary fibrosis [89].
In muco-obstructive hyper tension, the inhibition of the KCa3.1 channel [5] and Kcnn4 silencing in ion transport and MCC in an animal model of CF/COPD-like muco-obstructive lung disease determined that Kcnn4 silencing enhances airway disease [5]. The effectiveness of the mucociliary clearance depends mainly on hydration. Water availability in the airways is controlled by transepithelial ion transport. Apical Cl− secretion and Na+ absorption play major roles in ASL volume homeostasis [90]. The decline in Na+ absorption is of potential benefit in muco-obstructive disorders, such as cystic fibroses. It was described earlier in the case of the kidney and intestine, where the inhibition of basolateral K+ channels decreased Na+ absorption [5,57], thus supporting the role of K+ channels on epithelial Na+ homeostasis.
In pulmonary artery hypertension, elevated pulmonary artery pressure occurs in several diseases, such as asthma, end-stage chronic obstructive pulmonary disease (COPD), and lung fibrosis [66,91,92]. In order to diagnose pulmonary artery hypertension, hemodynamic measurements are taken via right heart catheterization or echocardiography; the condition is defined as a mean pulmonary artery pressure above 25 mmHg at rest or greater than 30 mmHg during normal physical activity [92]. Studies suggest that pharmacologically activating KCa3.1 channels mediates human pulmonary artery and bronchial relaxations [42]
5. Pharmacological KCa2 and KCa3.1 Channel Modulators in Respiratory Diseases
The KCa2.3 and KCa3.1 potassium channels are characterized by their voltage independence, and thus, they are activated by intracellular Ca2+. Due to the distinct distribution of the channel subtypes in the mammalian cells and their involvement in the generation of afterhyperpolarization currents, there has been considerable interest in developing subtype-selective pharmacological tools to study these channels [93,94]. Additionally, KCa2.3 and KCa3.1 channels comprise attractive new targets for several diseases that currently have no effective therapies. The pharmacology of KCa channels developed relatively rapidly after the cloning of the KCa2 and KCa3.1 channels, as the field now has a wide range of peptides, small-molecule inhibitors, and positive- and negative-gating modulators with differential subtype selectivity available [44].
The KCa3.1 and KCa2 channels have relatively well-developed pharmacological tools. The field now has a wide range of peptides, small-molecule inhibitors, and positive- and negative-gating modulators with differential subtype selectivity available [93]. Table 3 shows the small molecule positive and negative modulators with differential KCa2 subtype selectivity [44]. For treating CF and other mucociliary diseases, KCa3.1 inhibitors are needed [5]. Senicapoc [95] and TRAM-34 [96] inhibit KCa3.1 channels with IC50 values of ~11 nM, and ~20 nM, respectively, and they are highly selective for KCa3.1 channels over KCa2 channel subtypes [33]. The selective negative modulator for the KCa2 channel AP14145 is equipotent in inhibiting KCa2.2 and KCa2.3 but is ineffective on KCa3.1 channels [97].
For treating anosmia, COPD and its related pulmonary hypertension, KCa2-positive modulators may be beneficial [42,46,62]. NS309 is a potent, non-selective activator of human KCa3.1 and KCa2 channels [98]. The KCa2.2 and KCa2.3 channels are potently and selectively activated by CyPPA [38], and their derivatives are chemically modified to create more efficient and selective positive modulators [99]. However, further investigations are needed to determine their effectiveness [33].

{kind=link}
{kind=link}
{kind=link}
Table 3.
Small-molecule positive and negative modulators of KCa2 and KCa3.1 channels.
| Nonselective KCa2/KCa3.1 | KCa2 Selective | KCa3.1 Selective | Subtype KCa2 Selective | |
|---|---|---|---|---|
| Positive modulators | NS309 [98] SKA-31 [100] 1-EBIO [101] Riluzole [102] | SKA-111 [44] SKA-121 [103] | KCa2.2/KCa2.3 selective | |
| CyPPA [38] NS13001 [104] Compound 2q * [99] | ||||
| KCa2.1 selective | ||||
| CM-TPMF [102] | ||||
| Negative modulators | RA-2 [103] | NS5893 [104] AP14145 [97] | Senicapoc [11,95] TRAM-34 [96] | KCa2.1 selective |
| Bu-TPMF [102] |
6. Conclusions and Perspectives
In recent years, remarkable progress has been made in understanding the physiological and pathophysiological roles of KCa channels. The advances in understanding the KCa3.1 structure and the resulting improvements in other KCa2 subtypes modeling have yet to be used, not only for drug discovery but also for understanding the pathophysiological diseases, particularly airway diseases, and developing more subtype-selective biophysical and pharmacological tools. Over the past few years, researchers have studied KCa3.1 channel expression and its physiological role in airway diseases. There are, however, few studies on KCa2 channels in the respiratory system. Evidence now suggests that KCa2 channels are present in the respiratory system and play an important role in airway disorders, such as asthma, chronic obstructive pulmonary disease, cystic fibrosis, and other muco-obstructive diseases. Nevertheless, further studies are necessary to unveil the exact cell distribution, subcellular localization, and protein interactions of KCa2 channels in the airways. Additional research is required to further establish and validate KCa2 and KCa3.1 channels as ion channels in airway diseases, their clinical relevance, and the development of more potent and subtype selective KCa2 channel modulators.
Author Contributions
R.O. drafted the manuscript; A.A., M.A.R. and L.L. edited and revised the manuscript; Y.-W.N. and M.Z. approved the final version of the manuscript. All authors contributed to the manuscript and the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a 23AIREA1039423 grant from the American Heart Association, a YI-SCA grant from the National Ataxia Foundation, and a 4R33NS101182-03 grant from NIH awarded to M.Z. And grant 023-012 from King Fahad Medical City Research Center awarded to R.O.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank King Fahad Medical City Research Center and Chapman University Writing Center for revising the manuscript. We thank Fan Feng and Reem Orfali for their technical support. Figures were created with BioRender and published with permission.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ASL | Airway surface liquid |
| Ca2+ | Calcium |
| CaM | Calmodulin |
| Cl− | Chloride |
| COPD | Chronic obstructive pulmonary disease |
| CBF | Cilia beating frequency |
| KCa | Ca2+-activated K+ channels |
| CF | Cystic fibrosis |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| KCa3.1 | Intermediate-conductance Ca2+-activated K+ channels |
| BK | Large-conductance Ca+2-activated K+ channels |
| MCC | Mucociliary clearance |
| ORNs | Olfactory receptor neurons |
| K+ | Potassium |
| KCa2 | Small-conductance Ca2+-activated K+ channels |
| Na+ | Sodium |
| TMs | Transmembrane helices |
| WT | Wild type |
References
- Bartoszewski, R.; Matalon, S.; Collawn, J.F. Ion channels of the lung and their role in disease pathogenesis. Am. J. Physiol. Cell. Mol. Physiol. 2017, 313, L859–L872. [Google Scholar] [CrossRef] [PubMed]
- Mall, M.A. Role of cilia, mucus, and airway surface liquid in mucociliary dysfunction: Lessons from mouse models. J. Aerosol. Med. Pulm. Drug Deliv. 2008, 21, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkholm, M.; Mortensen, J. Mucociliary clearance: Pathophysiological aspects. Clin. Physiol. Funct. Imaging 2014, 34, 171–177. [Google Scholar] [CrossRef]
- Bhowmik, A.; Chahal, K.; Austin, G.; Chakravorty, I. Improving mucociliary clearance in chronic obstructive pulmonary disease. Respir. Med. 2009, 103, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Vega, G.; Guequén, A.; Philp, A.R.; Gianotti, A.; Arzola, L.; Villalón, M.; Zegarra-Moran, O.; Galietta, L.J.; Mall, M.A.; Flores, C.A. Lack of Kcnn4 improves mucociliary clearance in muco-obstructive lung disease. JCI Insight 2020, 5, 140076. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.G.; Livraghi-Butrico, A.; Fletcher, A.A.; McElwee, M.M.; Evans, S.E.; Boerner, R.M.; Alexander, S.N.; Bellinghausen, L.K.; Song, A.S.; Petrova, Y.M.; et al. Muc5b is required for airway defence. Nature 2014, 505, 412–416. [Google Scholar] [CrossRef] [Green Version]
- Spina, D. Epithelium smooth muscle regulation and interactions. Am. J. Respir. Crit. Care Med. 1998, 158, S141–S145. [Google Scholar] [CrossRef]
- Callaghan, P.J.; Ferrick, B.; Rybakovsky, E.; Thomas, S.; Mullin, J.M. Epithelial barrier function properties of the 16HBE14o- human bronchial epithelial cell culture model. Biosci. Rep. 2020, 40, BSR20201532. [Google Scholar] [CrossRef]
- Hollenhorst, M.I.; Richter, K.; Fronius, M. Ion transport by pulmonary epithelia. J. Biomed. Biotechnol. 2011, 2011, 174306. [Google Scholar] [CrossRef] [Green Version]
- Bernard, K.; Bogliolo, S.; Soriani, O.; Ehrenfeld, J. Modulation of calcium-dependent chloride secretion by basolateral SK4-like channels in a human bronchial cell line. J. Membr. Biol. 2003, 196, 15–31. [Google Scholar] [CrossRef]
- Staal, R.G.W.; Weinstein, J.R.; Nattini, M.; Cajina, M.; Chandresana, G.; Möller, T. Senicapoc: Repurposing a Drug to Target Microglia KCa3.1 in Stroke. Neurochem. Res. 2017, 42, 2639–2645. [Google Scholar] [CrossRef]
- Bardou, O.; Trinh, N.T.N.; Brochiero, E. Molecular diversity and function of K+ channels in airway and alveolar epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L145–L155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KCa3.1 Channel Blockade Attenuates Microvascular Remodelling in a Large Animal Model of Bleomycin-Induced Pulmonary Fibrosis|Scientific Reports n.d. Available online: https://www.nature.com/articles/s41598-019-56412-z (accessed on 15 March 2023).
- Zühlke, R.D.; Pitt, G.S.; Deisseroth, K.; Tsien, R.W.; Reuter, H. Calmodulin supports both inactivation and facilitation of L-type calcium channels. Nature 1999, 399, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.-W.; Orfali, R.; Liu, T.; Yu, K.; Cui, M.; Wulff, H.; Zhang, M. Structural insights into the potency of SK channel positive modulators. Sci. Rep. 2017, 7, 17178. [Google Scholar] [CrossRef] [Green Version]
- [PDF] Unstructured to Structured Transition of an Intrinsically Disordered Protein Peptide in Coupling Ca2+-Sensing and SK Channel Activation|Semantic Scholar n.d. Available online: https://www.semanticscholar.org/paper/Unstructured-to-structured-transition-of-an-protein-Zhang-Pascal/9f5cff1624625aaaeb67cab5639813dba6db10b4 (accessed on 15 March 2023).
- Catacuzzeno, L.; Fioretti, B.; Franciolini, F. Expression and Role of the Intermediate-Conductance Calcium-Activated Potassium Channel KCa3.1 in Glioblastoma. J. Signal. Transduct. 2012, 2012, 421564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, B.S.; Strøbaek, D.; Olesen, S.P.; Christophersen, P. The Ca2+-activated K+ channel of intermediate conductance: A molecular target for novel treatments? Curr. Drug. Targets 2001, 2, 401–422. [Google Scholar] [CrossRef]
- Kaczmarek, L.K.; Aldrich, R.W.; Chandy, K.G.; Grissmer, S.; Wei, A.D.; Wulff, H. International Union of Basic and Clinical Pharmacology. C. Nomenclature and Properties of Calcium-Activated and Sodium-Activated Potassium Channels. Pharmacol. Rev. 2017, 69, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Christophersen, P.; Wulff, H. Pharmacological gating modulation of small- and intermediate-conductance Ca2+ -activated K+ channels (KCa2.x and KCa3.1). Channels 2015, 9, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Padula, A.E.; Griffin, W.C.; Lopez, M.F.; Nimitvilai, S.; Cannady, R.; McGuier, N.S.; Chesler, E.J.; Miles, M.F.; Williams, R.W.; Randall, P.K.; et al. KCNN Genes that Encode Small-Conductance Ca2+-Activated K+ Channels Influence Alcohol and Drug Addiction. Neuropsychopharmacol 2015, 40, 1928–1939. [Google Scholar] [CrossRef] [PubMed]
- Begenisich, T.; Nakamoto, T.; Ovitt, C.E.; Nehrke, K.; Brugnara, C.; Alper, S.L.; Melvin, J.E. Physiological Roles of the Intermediate Conductance, Ca2+-activated Potassium Channel Kcnn4*. J. Biol. Chem. 2004, 279, 47681–47687. [Google Scholar] [CrossRef] [Green Version]
- Orfali, R.; Albanyan, N. Ca2+-Sensitive Potassium Channels. Molecules 2023, 28, 885. [Google Scholar] [CrossRef] [PubMed]
- Girault, A.; Haelters, J.-P.; Potier-Cartereau, M.; Chantôme, A.; Jaffrés, P.-A.; Bougnoux, P.; Joulin, V.; Vandier, C. Targeting SKCa channels in cancer: Potential new therapeutic approaches. Curr. Med. Chem. 2012, 19, 697–713. [Google Scholar] [CrossRef]
- Aldrich, R.; Chandy, K.G.; Grissmer, S.; Gutman, G.A.; Kaczmarek, L.K.; Wei, A.D.; Wulff, H. Calcium- and sodium-activated potassium channels (KCa, KNa) in GtoPdb v.2021.3. GtoPdb CITE 2021, 2021. [Google Scholar] [CrossRef]
- Xia, X.-M.; Fakler, B.; Rivard, A.; Wayman, G.; Johnson-Pais, T.; Keen, J.E.; Ishii, T.; Hirschberg, B.; Bond, C.T.; Lutsenko, S.; et al. Mechanism of calcium gating in small-conductance calcium-activated potassium channels. Nature 1998, 395, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Shmukler, B.E.; Bond, C.T.; Wilhelm, S.; Bruening-Wright, A.; Maylie, J.; Adelman, J.P.; Alper, S.L. Structure and complex transcription pattern of the mouse SK1 KCa channel gene, KCNN1. Biochim. Biophys. Acta 2001, 1518, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.M.; Kohli, V.; Adachi, R.; López, J.A.; Udden, M.M.; Sullivan, R. Calmodulin binding to the C-terminus of the small-conductance Ca2+-activated K+ channel hSK1 is affected by alternative splicing. Biochemistry 2001, 40, 3189–3195. [Google Scholar] [CrossRef]
- Nam, Y.-W.; Cui, M.; El-Sayed, N.S.; Orfali, R.; Nguyen, M.; Yang, G.; Rahman, M.A.; Lee, J.; Zhang, M. Subtype-selective positive modulation of KCa2 channels depends on the HA/HB helices. Br. J. Pharmacol. 2022, 179, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Nam, Y.-W.; Cui, M.; Orfali, R.; Viegas, A.; Nguyen, M.; Mohammed, E.H.M.; Zoghebi, K.A.; Rahighi, S.; Parang, K.; Zhang, M. Hydrophobic interactions between the HA helix and S4-S5 linker modulate apparent Ca2+ sensitivity of SK2 channels. Acta Physiol. 2021, 231, e13552. [Google Scholar] [CrossRef]
- El-Sayed, N.S.; Nam, Y.-W.; Egorova, P.A.; Nguyen, H.M.; Orfali, R.; Rahman, M.A.; Yang, G.; Wulff, H.; Bezprozvanny, I.; Parang, K.; et al. Structure-Activity Relationship Study of Subtype-Selective Positive Modulators of KCa2 Channels. J. Med. Chem. 2022, 65, 303–322. [Google Scholar] [CrossRef]
- Morales, P.; Garneau, L.; Klein, H.; Lavoie, M.-F.; Parent, L.; Sauvé, R. Contribution of the KCa3.1 channel–calmodulin interactions to the regulation of the KCa3.1 gating process. J. Gen. Physiol. 2013, 142, 37–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.-W.; Downey, M.; Rahman, M.A.; Cui, M.; Zhang, M. Channelopathy of small- and intermediate-conductance Ca2+-activated K+ channels. Acta Pharmacol. Sin. 2023, 44, 259–267. [Google Scholar] [CrossRef]
- Orfali, R.; Nam, Y.-W.; Nguyen, H.M.; Rahman, M.A.; Yang, G.; Cui, M.; Wulff, H.; Zhang, M. Channelopathy-causing mutations in the S45A/S45B and HA/HB helices of KCa2.3 and KCa3.1 channels alter their apparent Ca2+ sensitivity. Cell Calcium 2022, 102, 102538. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, A.S.; Benton, D.C.H.; Bahia, P.K.; Hosseini, R.; Shah, Y.A.; Haylett, D.G.; Moss, G.W.J. The SK3 Subunit of Small Conductance Ca2+-activated K+ Channels Interacts with Both SK1 and SK2 Subunits in a Heterologous Expression System. J. Biol. Chem. 2004, 279, 1003–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-H.; MacKinnon, R. Activation mechanism of a human SK-calmodulin channel complex elucidated by cryo-EM structures. Science 2018, 360, 508–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.M.; Silvia, C.; Hirschberg, B.; Bond, C.T.; Adelman, J.P.; Maylie, J. A human intermediate conductance calcium-activated potassium channel. Proc. Natl. Acad. Sci. USA 1997, 94, 11651–11656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hougaard, C.; Eriksen, B.L.; Jørgensen, S.; Johansen, T.H.; Dyhring, T.; Madsen, L.S.; Strøbaek, D.; Christophersen, P. Selective positive modulation of the SK3 and SK2 subtypes of small conductance Ca2+-activated K+ channels. Br. J. Pharmacol. 2007, 151, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfali, R. Genetic Mutations of KCa2.3 and KCa3.1 Channels Affect Ca2+ Sensitivity. Ph.D. Thesis, Pharmaceutical Sciences, Chapman University, Irvine, CA, USA, 2023. [Google Scholar] [CrossRef]
- Wulff, H.; Zhorov, B.S. K+ Channel Modulators for the Treatment of Neurological Disorders and Autoimmune Diseases. Chem. Rev. 2008, 108, 1744–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardou, O.; Trinh, N.T.N.; Brochiero, E. Canaux potassiques et physiologie de l’épithélium respiratoire. Med. Sci. 2009, 25, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Kroigaard, C.; Dalsgaard, T.; Nielsen, G.; Laursen, B.E.; Pilegaard, H.; Köhler, R.; Simonsen, U. Activation of endothelial and epithelial KCa2.3 calcium-activated potassium channels by NS309 relaxes human small pulmonary arteries and bronchioles. Br. J. Pharmacol. 2012, 167, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahm, A.; Wieder, T.; Gramlich, D.; Müller, M.E.; Wunsch, M.N.; El Tahry, F.A.; Heimberger, T.; Sandke, S.; Weis, T.; Most, P.; et al. Differential regulation of KCa2.1 (KCNN1) K+ channel expression by histone deacetylases in atrial fibrillation with concomitant heart failure. Physiol. Rep. 2021, 9, e14835. [Google Scholar] [CrossRef]
- Brown, B.M.; Shim, H.; Christophersen, P.; Wulff, H. Pharmacology of Small- and Intermediate-Conductance Calcium-Activated Potassium Channels. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 219–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.X.; Gorman, S.A.; Benson, B.; Singh, K.; Hieble, J.P.; Michel, M.C.; Tate, S.N.; Trezise, D.J. Small and intermediate conductance Ca2+-activated K+ channels confer distinctive patterns of distribution in human tissues and differential cellular localisation in the colon and corpus cavernosum. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Wulff, H.; Köhler, R. Endothelial Small- and Intermediate-Conductance KCa Channels: An Update on Their Pharmacology and Usefulness as Cardiovascular Targets. J. Cardiovasc. Pharmacol. 2013, 61, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, M.A.; Rivard, A.F.; Bächinger, H.P.; Adelman, J.P. Structure of the gating domain of a Ca2+-activated K+ channel complexed with Ca2+/calmodulin. Nature 2001, 410, 1120–1124. [Google Scholar] [CrossRef]
- Zhang, M.; Abrams, C.; Wang, L.; Gizzi, A.; He, L.; Lin, R.; Chen, Y.; Loll, P.J.; Pascal, J.M.; Zhang, J. Structural basis for calmodulin as a dynamic calcium sensor. Structure 2012, 20, 911–923. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.N.; Suzuki, H.; Ohkubo, J.-I.; Wakasugi, T.; Kitamura, T. Calmodulin Regulates Ciliary Beats in the Human Nasal Mucosa Through Adenylate/Guanylate Cyclases and Protein Kinases A/G. Int. Arch. Allergy Immunol. 2021, 182, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Valeyev, N.V.; Bates, D.G.; Heslop-Harrison, P.; Postlethwaite, I.; Kotov, N.V. Elucidating the mechanisms of cooperative calcium-calmodulin interactions: A structural systems biology approach. BMC Syst. Biol. 2008, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, D.; Okada, Y.; Teeter, J.H.; Lowry, L.D.; Cowart, B.; Brand, J.G. Human olfactory neurons respond to odor stimuli with an increase in cytoplasmic Ca2+. Biophys. J. 1993, 64, 1961–1966. [Google Scholar]
- Delgado, R.; Saavedra, M.V.; Schmachtenberg, O.; Sierralta, J.; Bacigalupo, J. Presence of Ca2+-dependent K+ channels in chemosensory cilia support a role in odor transduction. J. Neurophysiol. 2003, 90, 2022–2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Benedetto, G.; Magnus, C.J.; Gray, P.T.; Mehta, A. Calcium regulation of ciliary beat frequency in human respiratory epithelium in vitro. J. Physiol. 1991, 439, 103–113. [Google Scholar] [CrossRef]
- De Proost, I.; Pintelon, I.; Brouns, I.; Kroese, A.B.A.; Riccardi, D.; Kemp, P.J.; Timmermans, J.-P.; Adriaensen, D. Functional live cell imaging of the pulmonary neuroepithelial body microenvironment. Am. J. Respir. Cell Mol. Biol. 2008, 39, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohba, T.; Sawada, E.; Suzuki, Y.; Yamamura, H.; Ohya, S.; Tsuda, H.; Imaizumi, Y. Enhancement of Ca2+ influx and ciliary beating by membrane hyperpolarization due to ATP-sensitive K+ channel opening in mouse airway epithelial cells. J. Pharmacol. Exp. Ther. 2013, 347, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Tilley, A.E.; Walters, M.S.; Shaykhiev, R.; Crystal, R.G. Cilia Dysfunction in Lung Disease. Annu. Rev. Physiol. 2015, 77, 379–406. [Google Scholar] [CrossRef] [Green Version]
- Hebert, S.C.; Desir, G.; Giebisch, G.; Wang, W. Molecular diversity and regulation of renal potassium channels. Physiol. Rev. 2005, 85, 319–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.-W.; Pala, R.; El-Sayed, N.S.; Larin-Henriquez, D.; Amirrad, F.; Yang, G.; Rahman, M.A.; Orfali, R.; Downey, M.; Parang, K.; et al. Subtype-Selective Positive Modulation of KCa2.3 Channels Increases Cilia Length. ACS Chem. Biol. 2022, 17, 2344–2354. [Google Scholar] [CrossRef]
- Weaver, A.K.; Bomben, V.C.; Sontheimer, H. Expression and Function of Calcium-Activated Potassium Channels in Human Glioma Cells. Glia 2006, 54, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocker, M. Ca(2+)-activated K+ channels: Molecular determinants and function of the SK family. Nat. Rev. Neurosci. 2004, 5, 758–770. [Google Scholar] [CrossRef]
- Shepherd, M.C.; Duffy, S.M.; Harris, T.; Cruse, G.; Schuliga, M.; Brightling, C.E.; Neylon, C.B.; Bradding, P.; Stewart, A.G. KCa3.1 Ca2+Activated K+ Channels Regulate Human Airway Smooth Muscle Proliferation. Am. J. Respir. Cell Mol. Biol. 2007, 37, 525–531. [Google Scholar] [CrossRef]
- Kunzelmann, K.; Pavenstädt, H.; Beck, C.; Ünal, Ö.; Emmrich, P.; Arndt, H.J.; Greger, R. Characterization of potassium channels in respiratory cells. I. General properties. Pflugers Arch. 1989, 414, 291–296. [Google Scholar] [CrossRef]
- Pablo, J.L.; DeCaen, P.G.; Clapham, D.E. Progress in ciliary ion channel physiology. J. Gen. Physiol. 2017, 149, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Kawai, F. Ca2+-activated K+ currents regulate odor adaptation by modulating spike encoding of olfactory receptor cells. Biophys. J. 2002, 82, 2005–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, H.; Hall, I.P.; Washabau, R.J.; Takagi, K.; Kotlikoff, M.I. Beta-Adrenergic Agonists Regulate KCa Channels in Airway Smooth Muscle by cAMP-Dependent and -Independent Mechanisms. 1994. Available online: https://www.jci.org/articles/view/116969/scanned-page/371 (accessed on 17 March 2023).
- Fujita, A.; Takeuchi, T.; Hanai, J.; Hata, F. Expression of the small conductance Ca2+-activated K+ channel, SK3, in the olfactory ensheathing glial cells of rat brain. Cell Tissue Res. 2003, 313, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Collawn, J.F.; Matalon, S. CFTR and lung homeostasis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L917–L923. [Google Scholar] [CrossRef] [Green Version]
- Naeije, R.; Barberà, J.A. Pulmonary hypertension associated with COPD. Crit. Care 2001, 5, 286–289. [Google Scholar] [CrossRef] [PubMed]
- López-Barneo, J.; del Toro, R.; Levitsky, K.L.; Chiara, M.D.; Ortega-Sáenz, P. Regulation of oxygen sensing by ion channels. J. Appl. Physiol. 2004, 96, 1187–1195. [Google Scholar] [CrossRef]
- Franco-Obregón, A.; López-Barneo, J. Low PO2 inhibits calcium channel activity in arterial smooth muscle cells. Am. J. Physiol. 1996, 271, H2290–H2299. [Google Scholar] [CrossRef]
- Franco-Obregón, A.; Ureña, J.; López-Barneo, J. Oxygen-sensitive calcium channels in vascular smooth muscle and their possible role in hypoxic arterial relaxation. Proc. Natl. Acad. Sci. USA 1995, 92, 4715. [Google Scholar] [CrossRef] [Green Version]
- Kume, H.; Nishiyama, O.; Isoya, T.; Higashimoto, Y.; Tohda, Y.; Noda, Y. Involvement of Allosteric Effect and KCa Channels in Crosstalk between β2-Adrenergic and Muscarinic M2 Receptors in Airway Smooth Muscle. Int. J. Mol. Sci. 2018, 19, 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, B.; Labarca, P.; Bacigalupo, J. A ciliary K+ conductance sensitive to charibdotoxin underlies inhibitory responses in toad olfactory receptor neurons. FEBS Lett. 1995, 359, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Morales, B.; Madrid, R.; Bacigalupo, J. Calcium mediates the activation of the inhibitory current induced by odorants in toad olfactory receptor neurons. FEBS Lett. 1997, 402, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Sanhueza, M.; Schmachtenberg, O.; Bacigalupo, J. Excitation, inhibition, and suppression by odors in isolated toad and rat olfactory receptor neurons. Am. J. Physiol. Cell Physiol. 2000, 279, C31–C39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logsdon, N.J.; Kang, J.; Togo, J.A.; Christian, E.P.; Aiyar, J. A novel gene, hKCa4, encodes the calcium-activated potassium channel in human T lymphocytes. J. Biol. Chem. 1997, 272, 32723–32726. [Google Scholar] [CrossRef] [Green Version]
- Feske, S.; Wulff, H.; Skolnik, E.Y. Ion channels in innate and adaptive immunity. Annu. Rev. Immunol. 2015, 33, 291–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushal, V.; Koeberle, P.D.; Wang, Y.; Schlichter, L.C. The Ca2+-activated K+ channel KCNN4/KCa3.1 contributes to microglia activation and nitric oxide-dependent neurodegeneration. J. Neurosci. 2007, 27, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanshani, S.; Wulff, H.; Miller, M.J.; Rohm, H.; Neben, A.; Gutman, G.A.; Cahalan, M.D.; Chandy, K.G. Up-regulation of the IKCa1 Potassium Channel during T-cell Activation: Molecular Mechanism and Functional Consequences. J. Biol. Chem. 2000, 275, 37137–37149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Z.; Makinde, T.O.; Agrawal, D.K. Calcium-Activated Potassium Channel KCa3.1 in Lung Dendritic Cell Migration. Am. J. Respir. Cell Mol. Biol. 2011, 45, 962–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharp, D.L.; Wamhoff, B.R.; Turk, J.R.; Bowles, D.K. Upregulation of intermediate-conductance Ca2+-activated K+ channel (IKCa1) mediates phenotypic modulation of coronary smooth muscle. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2493–H2503. [Google Scholar] [CrossRef] [PubMed]
- Jaggar, J.H.; Wellman, G.C.; Heppner, T.J.; Porter, V.A.; Perez, G.J.; Gollasch, M.; Kleppisch, T.; Rubart, M.; Stevenson, A.S.; Lederer, W.J.; et al. Ca2+ channels, ryanodine receptors and Ca2+-activated K+ channels: A functional unit for regulating arterial tone. Acta Physiol. Scand. 1998, 164, 577–587. [Google Scholar] [CrossRef]
- Neylon, C.B.; Lang, R.J.; Fu, Y.; Bobik, A.; Reinhart, P.H. Molecular cloning and characterization of the intermediate-conductance Ca2+-activated K+ channel in vascular smooth muscle: Relationship between KCa channel diversity and smooth muscle cell function. Circ. Res. 1999, 85, e33–e43. [Google Scholar] [CrossRef] [Green Version]
- Peña, T.L.; Rane, S.G. The fibroblast intermediate conductance KCa channel, FIK, as a prototype for the cell growth regulatory function of the IK channel family. J. Membr. Biol. 1999, 172, 249–257. [Google Scholar] [CrossRef]
- Gerlach, A.C.; Gangopadhyay, N.N.; Devor, D.C. Kinase-dependent regulation of the intermediate conductance, calcium-dependent potassium channel, hIK1. J. Biol. Chem. 2000, 275, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Wulff, H.; Castle, N.A. Therapeutic potential of KCa3.1 blockers: An overview of recent advances, and promising trends. Expert Rev. Clin. Pharmacol. 2010, 3, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Zhdanova, O.; Di, L.; Li, Z.; Albaqumi, M.; Wulff, H.; Skolnik, E.Y. Protein histidine phosphatase 1 negatively regulates CD4 T cells by inhibiting the K+ channel KCa3.1. Proc. Natl. Acad. Sci. USA 2008, 105, 14442–14446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruse, G.; Duffy, S.M.; Brightling, C.E.; Bradding, P. Functional KCa3.1 K+ channels are required for human lung mast cell migration. Thorax 2006, 61, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Roach, K.M.; Duffy, S.M.; Coward, W.; Feghali-Bostwick, C.; Wulff, H.; Bradding, P. The K+ channel KCa3.1 as a novel target for idiopathic pulmonary fibrosis. PLoS ONE 2013, 8, e85244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarran, R. Regulation of airway surface liquid volume and mucus transport by active ion transport. Proc. Am. Thorac. Soc. 2004, 1, 42–46. [Google Scholar] [CrossRef]
- Parasuraman, S.; Walker, S.; Loudon, B.L.; Gollop, N.D.; Wilson, A.M.; Lowery, C.; Frenneaux, M.P. Assessment of pulmonary artery pressure by echocardiography—A comprehensive review. Int. J. Cardiol. Heart Vasc. 2016, 12, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Wulff, H.; Kolski-Andreaco, A.; Sankaranarayanan, A.; Sabatier, J.-M.; Shakkottai, V. Modulators of small- and intermediate-conductance calcium-activated potassium channels and their therapeutic indications. Curr. Med. Chem. 2007, 14, 1437–1457. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, U.S.; Strøbaek, D.; Christophersen, P.; Hougaard, C.; Jensen, M.L.; Nielsen, E.Ø.; Peters, D.; Teuber, L. Synthesis and structure-activity relationship studies of 2-(N-substituted)-aminobenzimidazoles as potent negative gating modulators ofsmall conductance Ca2+-activated K+ channels. J. Med. Chem. 2008, 51, 7625–7634. [Google Scholar] [CrossRef] [PubMed]
- Stocker, J.W.; De Franceschi, L.; McNaughton-Smith, G.A.; Corrocher, R.; Beuzard, Y.; Brugnara, C. ICA-17043, a novel Gardos channel blocker, prevents sickled red blood cell dehydration in vitro and in vivo in SAD mice. Blood 2003, 101, 2412–2418. [Google Scholar] [CrossRef]
- Agarwal, J.J.; Zhu, Y.; Zhang, Q.-Y.; Mongin, A.A.; Hough, L.B. TRAM-34, a putatively selective blocker of intermediate-conductance, calcium-activated potassium channels, inhibits cytochrome P450 activity. PLoS ONE 2013, 8, e63028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simó-Vicens, R.; Kirchhoff, J.E.; Dolce, B.; Abildgaard, L.; Speerschneider, T.; Sørensen, U.S.; Grunnet, M.; Diness, J.G.; Bentzen, B.H. A new negative allosteric modulator, AP14145, for the study of small conductance calcium-activated potassium (KCa2) channels. Br. J. Pharmacol. 2017, 174, 4396–4408. [Google Scholar] [CrossRef] [Green Version]
- Strøbaek, D.; Teuber, L.; Jørgensen, T.D.; Ahring, P.K.; Kjaer, K.; Hansen, R.S.; Olesen, S.P.; Christophersen, P.; Skaaning-Jensen, B. Activation of human IK and SK Ca2+ -activated K+ channels by NS309 (6,7-dichloro-1H-indole-2,3-dione 3-oxime). Biochim. Biophys. Acta 2004, 1665, 1–5. [Google Scholar] [CrossRef] [Green Version]
- John, C.M.; Khaddaj Mallat, R.; Mishra, R.C.; George, G.; Singh, V.; Turnbull, J.D.; Umeshappa, C.S.; Kendrick, D.J.; Kim, T.; Fauzi, F.M.; et al. SKA-31, an activator of Ca2+-activated K+ channels, improves cardiovascular function in aging. Pharmacol. Res. 2020, 151, 104539. [Google Scholar] [CrossRef] [PubMed]
- The IKCa and SKCa Channel Activator 1-EBIO (100 µM) Increased Outwards…. Available online: https://www.researchgate.net/figure/The-IKCa-and-SKCa-channel-activator-1-EBIO-100-M-increased-outwards-currents-in_fig5_235718670 (accessed on 3 April 2023).
- Dimitriadi, M.; Kye, M.J.; Kalloo, G.; Yersak, J.M.; Sahin, M.; Hart, A.C. The Neuroprotective Drug Riluzole Acts via Small Conductance Ca2+-Activated K+ Channels to Ameliorate Defects in Spinal Muscular Atrophy Models. J. Neurosci. 2013, 33, 6557–6562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasumu, A.W.; Hougaard, C.; Rode, F.; Jacobsen, T.A.; Sabatier, J.M.; Eriksen, B.L.; Strøbæk, D.; Liang, X.; Egorova, P.; Vorontsova, D.; et al. Selective positive modulator of calcium-activated potassium channels exerts beneficial effects in a mouse model of spinocerebellar ataxia type 2. Chem. Biol. 2012, 19, 1340–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hougaard, C.; Hammami, S.; Eriksen, B.L.; Sørensen, U.S.; Jensen, M.L.; Strøbæk, D.; Christophersen, P. Evidence for a common pharmacological interaction site on KCa2 channels providing both selective activation and selective inhibition of the human KCa2.1 subtype. Mol. Pharmacol. 2012, 81, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Oliván-Viguera, A.; Valero, M.S.; Coleman, N.; Brown, B.M.; Laría, C.; Murillo, M.D.; Gálvez, J.A.; Díaz-de-Villegas, M.D.; Wulff, H.; Badorrey, R.; et al. A Novel Pan-Negative-Gating Modulator of KCa2/3 Channels, Fluoro-Di-Benzoate, RA-2, Inhibits Endothelium-Derived Hyperpolarization–Type Relaxation in Coronary Artery and Produces Bradycardia In Vivo. Mol. Pharmacol. 2015, 87, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strøbæk, D.; Hougaard, C.; Johansen, T.H.; Sørensen, U.S.; Nielsen, E.Ø.; Nielsen, K.S.; Taylor, R.D.T.; Pedarzani, P.; Christophersen, P. Inhibitory Gating Modulation of Small Conductance Ca2+-Activated K+ Channels by the Synthetic Compound (R)-N-(Benzimidazol-2-yl)-1,2,3,4-tetrahydro-1-naphtylamine (NS8593) Reduces Afterhyperpolarizing Current in Hippocampal CA1 Neurons. Mol. Pharmacol. 2006, 70, 1771–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
KCa3.1 and KCa2 Channel Structures in the presence and absence of Ca2+. KCa2 and KCa 3.1 channels are assembled as homotetramers of four α-subunits. (A) Human KCa3.1 channel cryo-EM structure (PDB: 6cnn). For clarity, four-channel subunits are shown in different colors: green, blue, yellow, and purple, along with calmodulin (CaM) (gray). (B) Schematic representation of one channel subunit in the absence of Ca2+. (C) Schematic representation of one channel subunit in the presence of Ca2+. (D) Extracellular top view of the KCa3.1 and KCa2 channels. (A) was generated using Biorender.com. (B,C) were generated using Pymol (Schrödinger, LLC, New York, NY, USA).
Figure 1.
KCa3.1 and KCa2 Channel Structures in the presence and absence of Ca2+. KCa2 and KCa 3.1 channels are assembled as homotetramers of four α-subunits. (A) Human KCa3.1 channel cryo-EM structure (PDB: 6cnn). For clarity, four-channel subunits are shown in different colors: green, blue, yellow, and purple, along with calmodulin (CaM) (gray). (B) Schematic representation of one channel subunit in the absence of Ca2+. (C) Schematic representation of one channel subunit in the presence of Ca2+. (D) Extracellular top view of the KCa3.1 and KCa2 channels. (A) was generated using Biorender.com. (B,C) were generated using Pymol (Schrödinger, LLC, New York, NY, USA).

Table 1.
Apparent Ca2+ sensitivity, structural studies, amino acid sequences alignments and identities between KCa2/3 channel subtypes.
Table 1.
Apparent Ca2+ sensitivity, structural studies, amino acid sequences alignments and identities between KCa2/3 channel subtypes.
| KCa2/3 α Subunit | Amino Acids | Apparent Ca2+ Sensitivity (μM) | KCa2 Subtypes Structural Studies | Sequence Alignment among KCa2 and KCa3.1 Channels |
|---|---|---|---|---|
| KCa2.1 | 543 [24] | ~0.31 [25,26] | [27,28] | KCa2.1 and KCa3.1 share a 43.3% sequence identity [29] |
| KCa2.2 | 579 [25] | ~0.32 [25,30] | [15,26,29,31] | KCa2.2 and KCa3.1 share a 45% sequence identity [32] |
| KCa2.3 | 731 [25] | ~0.30 [33,34] | [34,35] | KCa2.3 and KCa3. share a 46.6% sequence identity [34] |
| KCa3.1 | 427 [25] | ~0.27 [33,34] | [36] |
Table 2.
The KCNN gene family. Human chromosomal location, tissue distribution, functional effects, and their roles in the lungs.
Table 2.
The KCNN gene family. Human chromosomal location, tissue distribution, functional effects, and their roles in the lungs.
| KCa2/3 α Subunit | Gene | Other Names | Human Chromosomal Location | Tissue Distribution | Physiological Roles | Role in the Lungs |
|---|---|---|---|---|---|---|
| KCa2.1 | KCNN1 | SK1 | 19p13.11 [25] | Brain [25] Heart [43] Lung [41] | The KCa2 channels underlie the medium AHP and regulate neuronal firing frequency [23,44]. | ND * |
| KCa2.2 | KCNN2 | SK2 | 5q22.3 [25] | Brain and heart Adrenal gland, lungs, prostate, bladder, and liver [25,45]. | ND * | |
| KCa2.3 | KCNN3 | SK3 | 1q21.3 [25] | Brain and heart Vascular endothelium, lungs, and bladder [25,44] | KCa2.3 and KCa3.1 mediate the endothelium-derived hyperpolarization response [33,46] | (+) KCa2.3 relaxes the pulmonary arteries and bronchi ** [32] |
| KCa3.1 | KCNN4 | SK4 IK | 19q13.31 [25] | Vascular endothelium T and B lymphocytes Microglia, placenta, colon, and red blood cells Lungs and bladder [25,44] | KCa3.1 channels regulate calcium signaling, cellular activation, and cell volume [23,44] | (−) KCa3.1 reduces Na+ absorption ***, (+) CBF, and MCC [5]. (+) KCa3.1 relaxes the pulmonary arteries and bronchi [42] |
* ND: not determined specifically in the respiratory system. ** (+): Activation. *** (−): Inhibition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Orfali, R.; AlFaiz, A.; Rahman, M.A.; Lau, L.; Nam, Y.-W.; Zhang, M. KCa2 and KCa3.1 Channels in the Airways: A New Therapeutic Target. Biomedicines 2023, 11, 1780. https://doi.org/10.3390/biomedicines11071780
AMA Style
Orfali R, AlFaiz A, Rahman MA, Lau L, Nam Y-W, Zhang M. KCa2 and KCa3.1 Channels in the Airways: A New Therapeutic Target. Biomedicines. 2023; 11(7):1780. https://doi.org/10.3390/biomedicines11071780
Chicago/Turabian StyleOrfali, Razan, Ali AlFaiz, Mohammad Asikur Rahman, Liz Lau, Young-Woo Nam, and Miao Zhang. 2023. "KCa2 and KCa3.1 Channels in the Airways: A New Therapeutic Target" Biomedicines 11, no. 7: 1780. https://doi.org/10.3390/biomedicines11071780
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.