
Different Transcutaneous Auricular Vagus Nerve Stimulation Parameters Modulate the Anti-Inflammatory Effects on Lipopolysaccharide-Induced Acute Inflammation in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
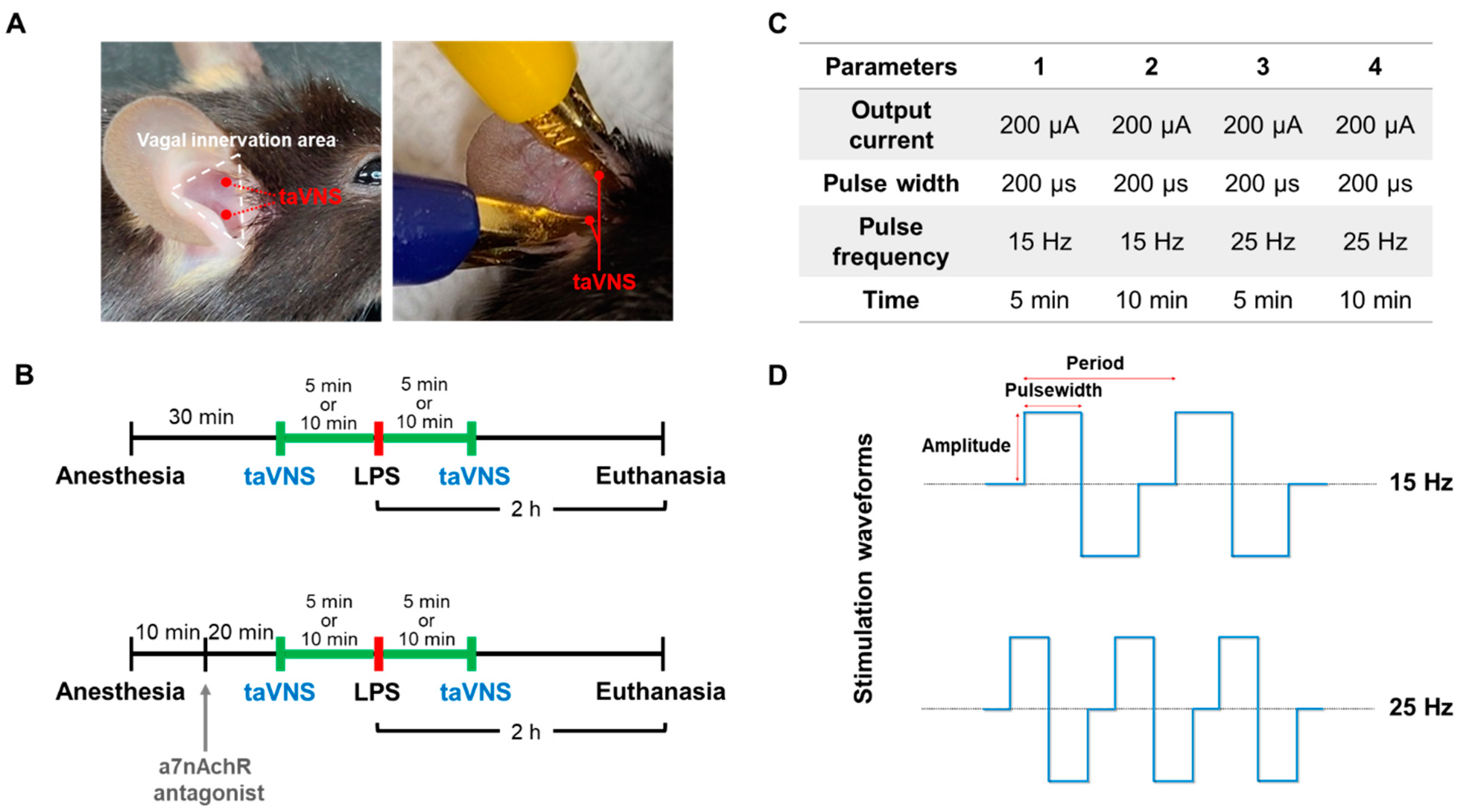
2. Materials and Methods
2.1. LPS-Induced Endotoxemia Mice Model
2.2. Transcutaneous Auricular VNS
2.3. Administration of α7nAChR Antagonist
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Western Blotting
2.6. Hematoxylin and Eosin Staining
2.7. Myeloperoxidase Staining
2.8. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
2.9. Cytokine Array
2.10. Statistical Analysis
3. Results
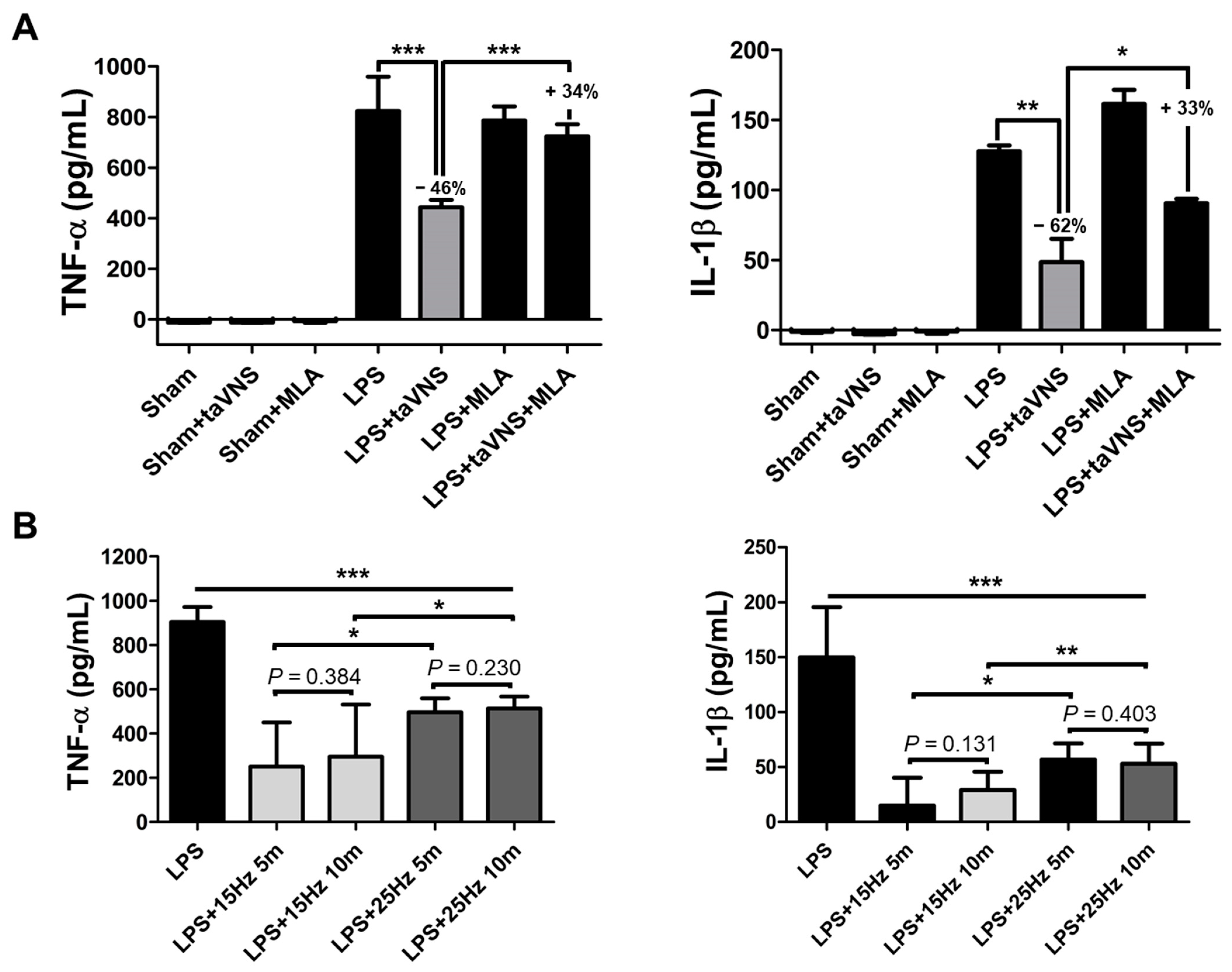
3.1. taVNS Reduced the Expression Levels of Pro-Inflammatory Cytokines in the Serum of the LPS-Induced Inflammation
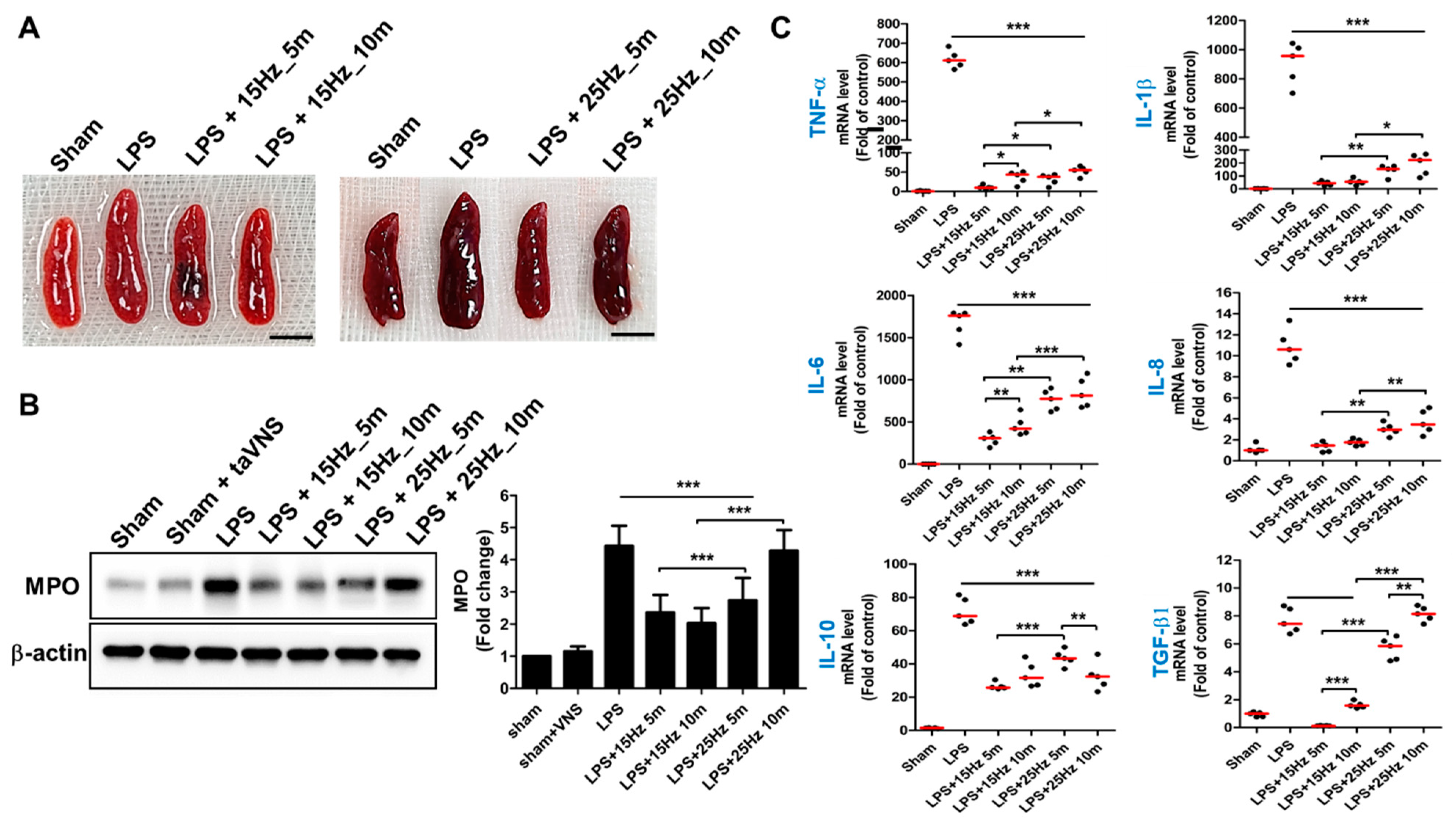
3.2. Anti-Inflammatory Effect of taVNS on Spleen Tissue of LPS-Induced Inflammation
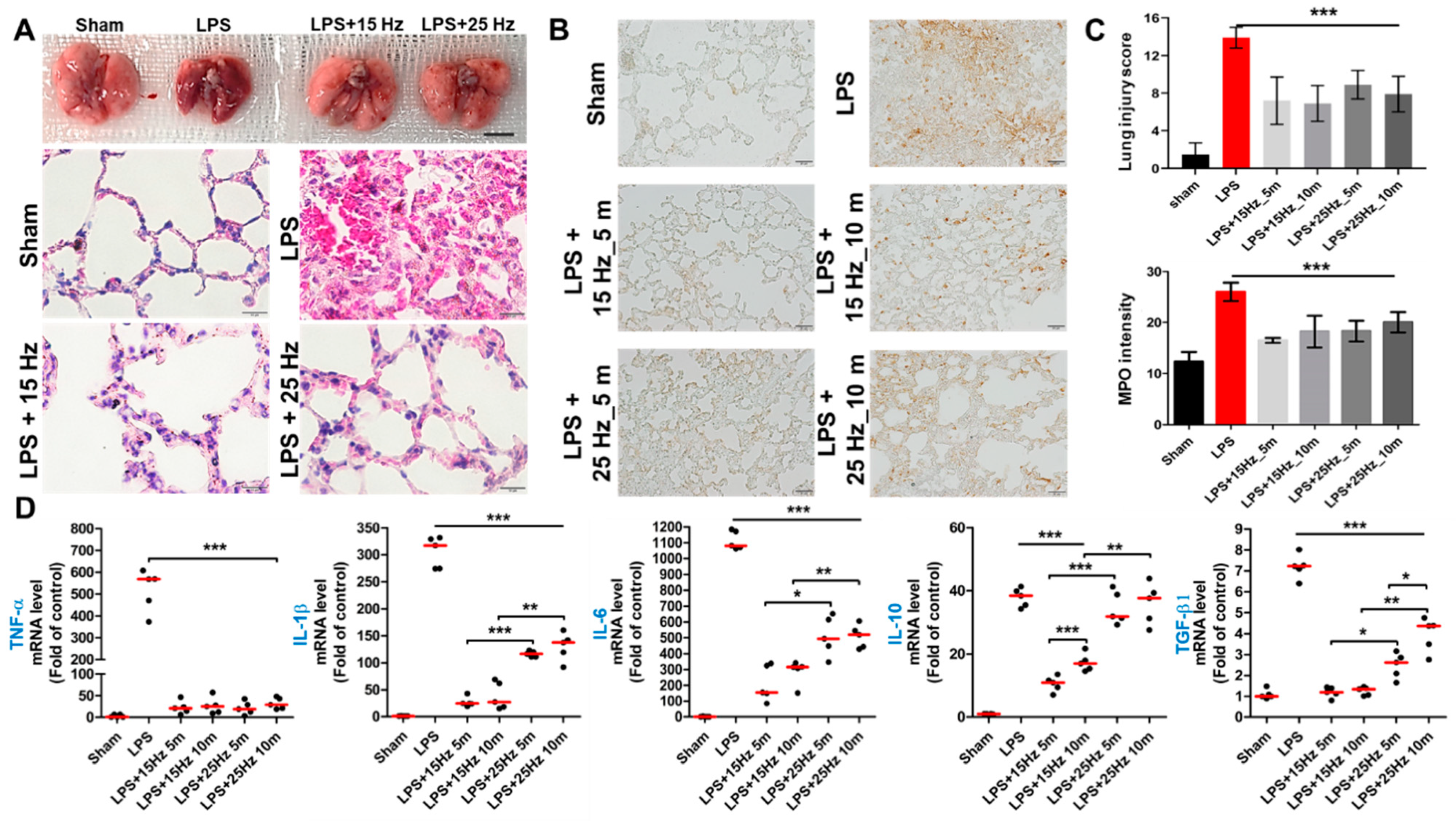
3.3. Anti-Inflammatory Effect of taVNS on Lung Injury of LPS-Induced Endotoxemia
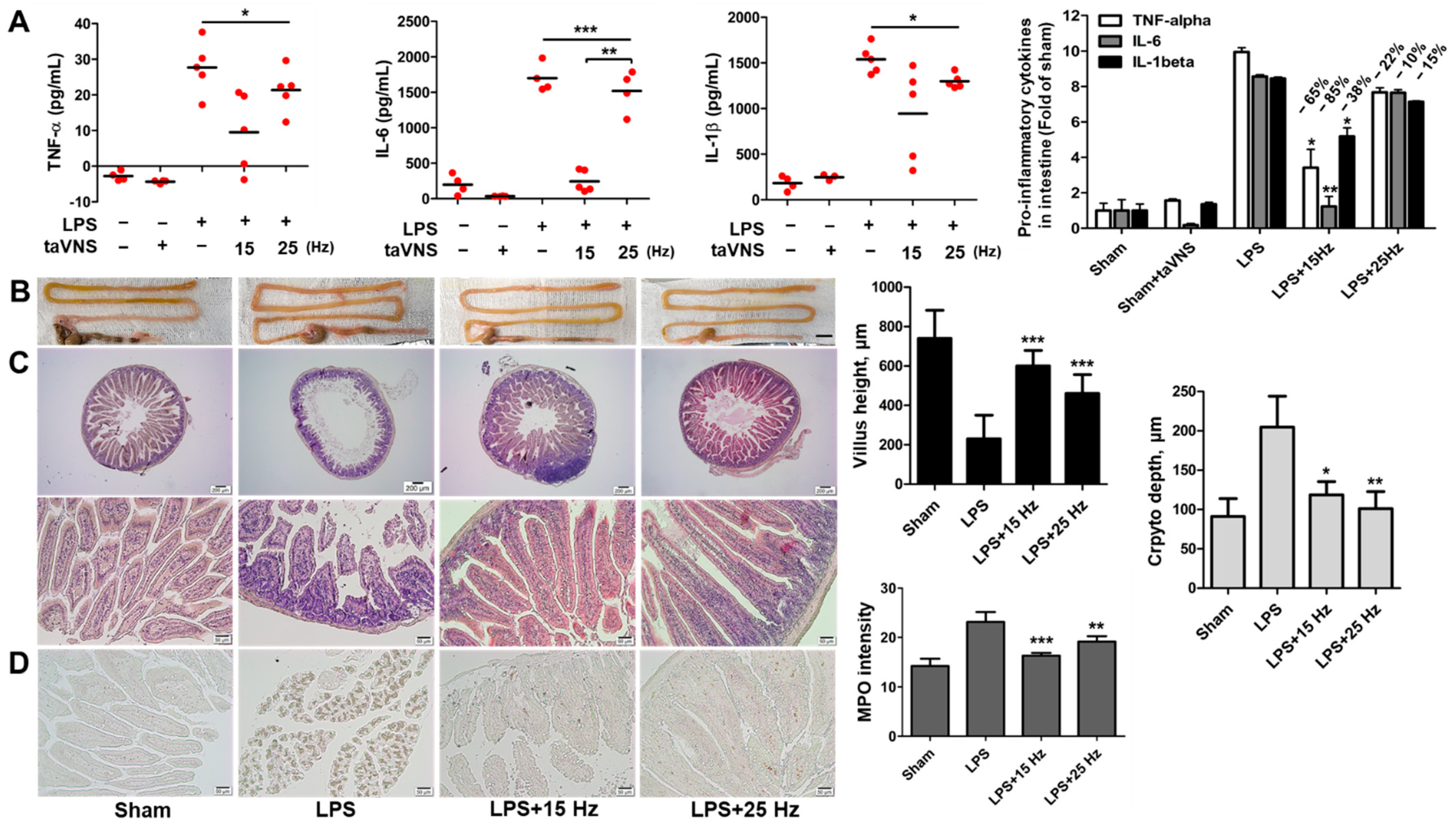
3.4. Anti-Inflammatory Effect of taVNS on Intestinal Inflammation Induced by LPS
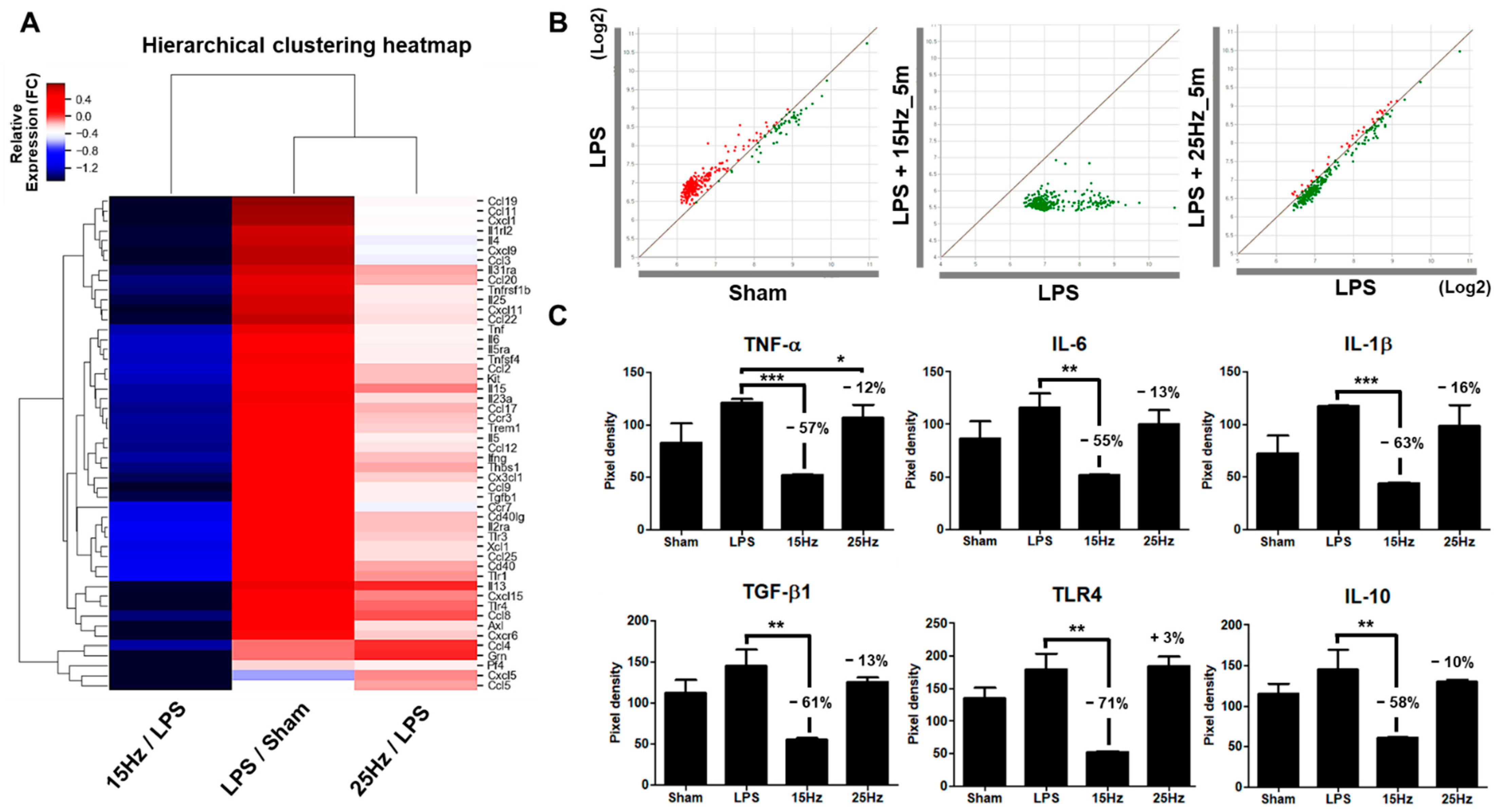
3.5. Electrical Frequency of the taVNS Parameter Regulated Inflammatory Cytokines in the Serum of LPS-Induced Endotoxemia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, H.K. Cranial nerve XI: The spinal accessory nerve. In Clinical Methods: The History, Physical, and Laboratory Examinations; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- Berthoud, H.R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Tracey, K.J. The vagus nerve and the inflammatory reflex—Linking immunity and metabolism. Nat. Rev. Endocrinol. 2012, 8, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Tracey, K.J. The cholinergic anti-inflammatory pathway. Brain Behav. Immun. 2005, 19, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Ballina, M.; Ochani, M.; Parrish, W.R.; Ochani, K.; Harris, Y.T.; Huston, J.M.; Chavan, S.; Tracey, K.J. Splenic nerve is required for cholinergic antiinflammatory pathway control of TNF in endotoxemia. Proc. Natl. Acad. Sci. USA 2008, 105, 11008–11013. [Google Scholar] [CrossRef] [Green Version]
- Koopman, F.A.; Chavan, S.S.; Miljko, S.; Grazio, S.; Sokolovic, S.; Schuurman, P.R.; Mehta, A.D.; Levine, Y.A.; Faltys, M.; Zitnik, R.; et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2016, 113, 8284–8289. [Google Scholar] [CrossRef] [Green Version]
- Yamakawa, K.; Matsumoto, N.; Imamura, Y.; Muroya, T.; Yamada, T.; Nakagawa, J.; Shimazaki, J.; Ogura, H.; Kuwagata, Y.; Shimazu, T. Electrical vagus nerve stimulation attenuates systemic inflammation and improves survival in a rat heatstroke model. PLoS ONE 2013, 8, e56728. [Google Scholar] [CrossRef]
- Johnson, R.L.; Wilson, C.G. A review of vagus nerve stimulation as a therapeutic intervention. J. Inflam. Res. 2018, 11, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.A.; Kouzani, A.; Lee, K.H.; Ross, E.K. Neurostimulation devices for the treatment of neurologic disorders. Mayo Clin. Proc. 2017, 92, 1427–1444. [Google Scholar] [CrossRef] [Green Version]
- Nemeroff, C.B.; Mayberg, H.S.; Krahl, S.E.; McNamara, J.; Frazer, A.; Henry, T.R.; George, M.S.; Charney, D.S.; Brannan, S.K. VNS therapy in treatment-resistant depression: Clinical evidence and putative neurobiological mechanisms. Neuropsychopharmacology 2006, 31, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Giordano, F.; Zicca, A.; Barba, C.; Guerrini, R.; Genitori, L. Vagus nerve stimulation: Surgical technique of implantation and revision and related morbidity. Epilepsia 2017, 58 (Suppl. 1), 85–90. [Google Scholar] [CrossRef] [PubMed]
- Carreno, F.R.; Frazer, A. Vagal nerve stimulation for treatment-resistant depression. Neurotherapeutics 2017, 14, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Van Leusden, J.W.; Sellaro, R.; Colzato, L.S. Transcutaneous vagal nerve stimulation (tVNS): A new neuromodulation tool in healthy humans? Front. Psychol. 2015, 6, 102. [Google Scholar] [CrossRef] [Green Version]
- Panebianco, M.; Rigby, A.; Weston, J.; Marson, A.G. Vagus nerve stimulation for partial seizures. Cochrane Database Syst. Rev. 2015, 4, CD002896. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): A review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the “cytokine storm” in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Kaniusas, E.; Szeles, J.C.; Kampusch, S.; Alfageme-Lopez, N.; Yucuma-Conde, D.; Li, X.; Mayol, J.; Neumayer, C.; Papa, M.; Panetsos, F. Non-invasive auricular vagus nerve stimulation as a potential treatment for COVID-19-originated acute respiratory distress syndrome. Front. Physiol. 2020, 11, 890. [Google Scholar] [CrossRef]
- Azabou, E.; Bao, G.; Bounab, R.; Heming, N.; Annane, D. Vagus nerve stimulation: A potential adjunct therapy for COVID-19. Front. Med. 2021, 8, 625836. [Google Scholar] [CrossRef]
- Bonaz, B.; Sinniger, V.; Pellissier, S. Targeting the cholinergic anti-inflammatory pathway with vagus nerve stimulation in patients with Covid-19? Bioelectron. Med. 2020, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Matteoli, G.; Gomez-Pinilla, P.J.; Nemethova, A.; Di Giovangiulio, M.; Cailotto, C.; van Bree, S.H.; Michel, K.; Tracey, K.J.; Schemann, M.; Boesmans, W.; et al. A distinct vagal anti-inflammatory pathway modulates intestinal muscularis resident macrophages independent of the spleen. Gut 2014, 63, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Huston, J.M.; Gallowitsch-Puerta, M.; Ochani, M.; Ochani, K.; Yuan, R.; Rosas-Ballina, M.; Ashok, M.; Goldstein, R.S.; Chavan, S.; Pavlov, V.A.; et al. Transcutaneous vagus nerve stimulation reduces serum high mobility group box 1 levels and improves survival in murine sepsis. Crit. Care Med. 2007, 35, 2762–2768. [Google Scholar] [PubMed]
- Hong, G.S.; Zillekens, A.; Schneiker, B.; Pantelis, D.; de Jonge, W.J.; Schaefer, N.; Kalff, J.C.; Wehner, S. Non-invasive transcutaneous auricular vagus nerve stimulation prevents postoperative ileus and endotoxemia in mice. Neurogastroenterol. Motil. 2019, 31, e13501. [Google Scholar] [CrossRef]
- Liu, J.; Li, C.; Peng, H.; Yu, K.; Tao, J.; Lin, R.; Chen, L. Electroacupuncture attenuates learning and memory impairment via activation of α7nAChR-mediated anti-inflammatory activity in focal cerebral ischemia/reperfusion injured rats. Exp. Ther. Med. 2017, 14, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Hirano, Y.; Aziz, M.; Yang, W.L.; Wang, Z.; Zhou, M.; Ochani, M.; Khader, A.; Wang, P. Neutralization of osteopontin attenuates neutrophil migration in sepsis-induced acute lung injury. Crit. Care 2015, 19, 53. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.M.; Rafikov, R.; Kumar, S.; Aggarwal, S.; Ham, P.B., III; Meadows, M.L.; Cherian-Shaw, M.; Kangath, A.; Sridhar, S.; Lucas, R.; et al. Endothelial nitric oxide synthase deficient mice are protected from lipopolysaccharide induced acute lung injury. PLoS ONE 2015, 10, e0119918. [Google Scholar]
- Tsaava, T.; Datta-Chaudhuri, T.; Addorisio, M.E.; Masi, E.B.; Silverman, H.A.; Newman, J.E.; Imperato, G.H.; Bouton, C.; Tracey, K.J.; Chavan, S.S.; et al. Specific vagus nerve stimulation parameters alter serum cytokine levels in the absence of inflammation. Bioelectron. Med. 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Wang, Y.; Cheng, B.; Xu, L.; Zhou, Y.; Wu, L. Protective effect of electrical stimulation of the vagus nerve in lipopolysaccharide-induced acute lung injury in rats. Mol. Med. Rep. 2021, 23, 365. [Google Scholar] [CrossRef]
- Bonaz, B.; Picq, C.; Sinniger, V.; Mayol, J.F.; Clarençon, D. Vagus nerve stimulation: From epilepsy to the cholinergic anti-inflammatory pathway. Neurogastroenterol. Motil. 2013, 25, 208–221. [Google Scholar] [CrossRef]
- Gorman, P.H.; Mortimer, J.T. The effect of stimulus parameters on the recruitment characteristics of direct nerve stimulation. IEEE Trans. Biomed. Eng. 1983, 30, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Yoo, P.B.; Liu, H.; Hincapie, J.G.; Ruble, S.B.; Hamann, J.J.; Grill, W.M. Modulation of heart rate by temporally patterned vagus nerve stimulation in the anesthetized dog. Physiol. Rep. 2016, 4, e12689. [Google Scholar] [CrossRef] [PubMed]
- Heck, C.; Helmers, S.L.; DeGiorgio, C.M. Vagus nerve stimulation therapy, epilepsy, and device parameters: Scientific basis and recommendations for use. Neurology 2002, 59 (Suppl. 4), S31–S37. [Google Scholar] [CrossRef] [PubMed]
- Groves, D.A.; Brown, V.J. Vagal nerve stimulation: A review of its applications and potential mechanisms that mediate its clinical effects. Neurosci. Biobehav. Rev. 2005, 29, 493–500. [Google Scholar] [CrossRef]
- Baratta, R.; Ichie, M.; Hwang, S.K.; Solomonow, M. Orderly stimulation of skeletal muscle motor units with tripolar nerve cuff electrode. IEEE Trans. Biomed. Eng. 1989, 36, 836–843. [Google Scholar] [CrossRef]
- Burke, D.; Mackenzie, R.A.; Skuse, N.F.; Lethlean, A.K. Cutaneous afferent activity in median and radial nerve fascicles: A microelectrode study. J. Neurol. Neurosurg. Psychiatry 1975, 38, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Piruzyan, M.; Shitanda, I.; Shimauchi, Y.; Okita, G.; Tsurekawa, Y.; Moriuchi, M.; Nakano, Y.; Teramoto, K.; Suico, M.A.; Shuto, T.; et al. A novel condition of mild electrical stimulation exerts immunosuppression via hydrogen peroxide production that controls multiple signaling pathway. PLoS ONE 2020, 15, e0234867. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Ahmed, U.; Jayaprakash, N.; Mughrabi, I.; Lin, Q.; Wu, Y.-C.; Abbas, A.; Gabalski, A.H.; Daytz, A.; Ashville, J.; et al. Manipulation of stimulus waveform and frequency permits targeted fiber activation during vagus nerve stimulation in 2 rodent species. bioRxiv 2021. [Google Scholar] [CrossRef]
- Musselman, E.D.; Pelot, N.A.; Grill, W.M. Empirically based guidelines for selecting vagus nerve stimulation parameters in epilepsy and heart failure. Cold Spring Harb. Perspect. Med. 2019, 9, a034264. [Google Scholar] [CrossRef]
- Martelli, D.; McKinley, M.J.; McAllen, R.M. The cholinergic anti-inflammatory pathway: A critical review. Auton. Neurosci. 2014, 182, 65–69. [Google Scholar] [CrossRef]
- Payne, S.C.; Furness, J.B.; Stebbing, M.J. Bioelectric neuromodulation for gastrointestinal disorders: Effectiveness and mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B. Parameters matter: Modulating cytokines using nerve stimulation. Bioelectron. Med. 2020, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, P.M.; Landgrebe, M.; Husser, O.; Resch, M.; Schecklmann, M.; Geisreiter, F.; Poeppl, T.B.; Prasser, S.J.; Hajak, G.; Langguth, B. Transcutaneous vagus nerve stimulation: Retrospective assessment of cardiac safety in a pilot study. Front. Psychiatry 2012, 3, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. The role of T lymphocytes in inflammation. Proc. Natl. Acad. Sci. USA 1994, 91, 2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soy, M.; Keser, G.; Atagündüz, P.; Tabak, F.; Atagündüz, I.; Kayhan, S. Cytokine storm in COVID-19: Pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef]
- González-Nicolás, M.Á.; González-Guerrero, C.; Pérez-Fernández, V.A.; Lázaro, A. Cilastatin: A potential treatment strategy against COVID-19 that may decrease viral replication and protect from the cytokine storm. Clin. Kidney J. 2020, 13, 903–905. [Google Scholar] [CrossRef]
- Langer-Gould, A.; Smith, J.B.; Gonzales, E.G.; Castillo, R.D.; Figueroa, J.G.; Ramanathan, A.; Li, B.H.; Gould, M.K. Early identification of COVID-19 cytokine storm and treatment with anakinra or tocilizumab. Int. J. Infect. Dis. 2020, 99, 291–297. [Google Scholar] [CrossRef]
- Yessayan, L.; Szamosfalvi, B.; Napolitano, L.; Singer, B.; Kurabayashi, K.; Song, Y.; Westover, A.; Humes, H.D. Treatment of cytokine storm in COVID-19 patients with immunomodulatory therapy. ASAIO J. 2020, 66, 1079–1083. [Google Scholar] [CrossRef]
- Saha, A.; Sharma, A.R.; Bhattacharya, M.; Sharma, G.; Lee, S.S.; Chakraborty, C. Tocilizumab: A therapeutic option for the treatment of cytokine storm syndrome in COVID-19. Arch. Med. Res. 2020, 51, 595–597. [Google Scholar] [CrossRef]
- Staats, P.; Giannakopoulos, G.; Blake, J.; Liebler, E.; Levy, R.M. The use of non-invasive vagus nerve stimulation to treat respiratory symptoms associated with COVID-19: A theoretical hypothesis and early clinical experience. Neuromodulation 2020, 23, 784–788. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Go, Y.-Y.; Ju, W.-M.; Lee, C.-M.; Chae, S.-W.; Song, J.-J. Different Transcutaneous Auricular Vagus Nerve Stimulation Parameters Modulate the Anti-Inflammatory Effects on Lipopolysaccharide-Induced Acute Inflammation in Mice. Biomedicines 2022, 10, 247. https://doi.org/10.3390/biomedicines10020247
Go Y-Y, Ju W-M, Lee C-M, Chae S-W, Song J-J. Different Transcutaneous Auricular Vagus Nerve Stimulation Parameters Modulate the Anti-Inflammatory Effects on Lipopolysaccharide-Induced Acute Inflammation in Mice. Biomedicines. 2022; 10(2):247. https://doi.org/10.3390/biomedicines10020247
Chicago/Turabian StyleGo, Yoon-Young, Won-Min Ju, Chan-Mi Lee, Sung-Won Chae, and Jae-Jun Song. 2022. "Different Transcutaneous Auricular Vagus Nerve Stimulation Parameters Modulate the Anti-Inflammatory Effects on Lipopolysaccharide-Induced Acute Inflammation in Mice" Biomedicines 10, no. 2: 247. https://doi.org/10.3390/biomedicines10020247