
Pathological Contribution of Extracellular Vesicles and Their MicroRNAs to Progression of Chronic Liver Disease


1
Institute of Systems Biology, College of Natural Science, Pusan National University, Pusan 46241, Korea
2
Department of Integrated Biological Science, College of Natural Science, Pusan National University, Pusan 46241, Korea
3
Department of Biological Sciences, College of Natural Science, Pusan National University, Pusan 46241, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biology 2022, 11(5), 637; https://doi.org/10.3390/biology11050637
Submission received: 30 March 2022
/
Revised: 16 April 2022
/
Accepted: 20 April 2022
/
Published: 21 April 2022
(This article belongs to the Special Issue Extracellular Vesicles in Inflammation and Inflammatory Diseases Edition 2.0)
Abstract
:Simple Summary
Extracellular vesicles (EVs) are membrane-enclosed vesicles secreted from most types of cells. EVs encapsulate many diverse bioactive cargoes, such as proteins and nucleic acid, of parental cells and delivers them to recipient cells. Upon injury, the contents altered by cellular stress are delivered into target cells and affect their physiological properties, spreading the disease microenvironment to exacerbate disease progression. Therefore, EVs are emerging as good resources for studying the pathophysiological mechanisms of diseases because they reflect the characteristics of donor cells and play a central role in intercellular communication. Chronic liver disease affects millions of people worldwide and has a high mortality rate. In chronic liver disease, the production and secretion of EVs are significantly elevated, and increased and altered cargoes are packed into EVs, enhancing inflammation, fibrosis, and angiogenesis. Herein, we review EVs released under specific chronic liver disease and explain how EVs are involved in intercellular communication to aggravate liver disease.
Abstract
Extracellular vesicles (EVs) are membrane-bound endogenous nanoparticles released by the majority of cells into the extracellular space. Because EVs carry various cargo (protein, lipid, and nucleic acids), they transfer bioinformation that reflects the state of donor cells to recipient cells both in healthy and pathologic conditions, such as liver disease. Chronic liver disease (CLD) affects numerous people worldwide and has a high mortality rate. EVs released from damaged hepatic cells are involved in CLD progression by impacting intercellular communication between EV-producing and EV-receiving cells, thereby inducing a disease-favorable microenvironment. In patients with CLD, as well as in the animal models of CLD, the levels of released EVs are elevated. Furthermore, these EVs contain high levels of factors that accelerate disease progression. Therefore, it is important to understand the diverse roles of EVs and their cargoes to treat CLD. Herein, we briefly explain the biogenesis and types of EVs and summarize current findings presenting the role of EVs in the pathogenesis of CLD. As the role of microRNAs (miRNAs) within EVs in liver disease is well documented, the effects of miRNAs detected in EVs on CLD are reviewed. In addition, we discuss the therapeutic potential of EVs to treat CLD.
1. Introduction
The liver is damaged by several deleterious factors, such as excessive alcohol consumption, a high-calorie diet, viral infection, and genetic disorders [1,2,3]. When the liver is severely and/or chronically injured, massive hepatocyte death occurs, which is accompanied by inflammation and fibrosis [4,5]. Dying hepatocytes release various cytokines and extracellular vesicles (EVs) carrying various cargos, which affect the pathogenesis of liver diseases [6,7,8]. Among these pathological contributors, EVs play an essential role in sharing cellular information and transmitting diverse pathophysiological conditions in the liver. EVs that are nano- to micro-sized vesicles enclosed by a membrane are released from most cell types into biological fluids, such as blood, saliva, urine, and amniotic fluid [9,10]. EVs include apoptotic bodies, microvesicles, and exosomes, and they are distinguished by their biogenesis, release pathway, size, content, and function [11,12]. EVs deliver bioinformation from donor cells to adjacent or distant target cells to mediate intercellular communications. EV-carried cargoes reflect the pathophysiological state of donor cells by delivering lipids, proteins, nucleic acids, and cellular organelles of donor cells. In the liver, all types of hepatic cells produce and release EVs to assist liver homeostasis [13]. Conde-Vancells et al. [14] found that EVs and their cargoes from hepatocytes are involved in maintaining hepatic functions, such as detoxification and lipid, carbohydrate, and amino acid metabolism. In addition, Nojima et al. [15] reported that normal hepatocyte-derived EVs improved hepatocyte survival and promoted cell proliferation through intercellular communication with adjacent hepatocytes. However, injured hepatocytes increase the amount of secreted EVs loading pro-inflammatory, angiogenic, and fibrotic substances, which influences the activity of target cells, eventually promoting disease progression [16,17]. Therefore, EVs released under disease conditions can be employed as diagnostic markers for specific diseases and as treatment targets for chronic liver disease (CLD). In this review, we summarize EVs released under specific CLD conditions and discuss how EVs are involved in intercellular communication to aggravate CLD progression, focusing on miRNAs in EVs.
2. Extracellular Vesicles
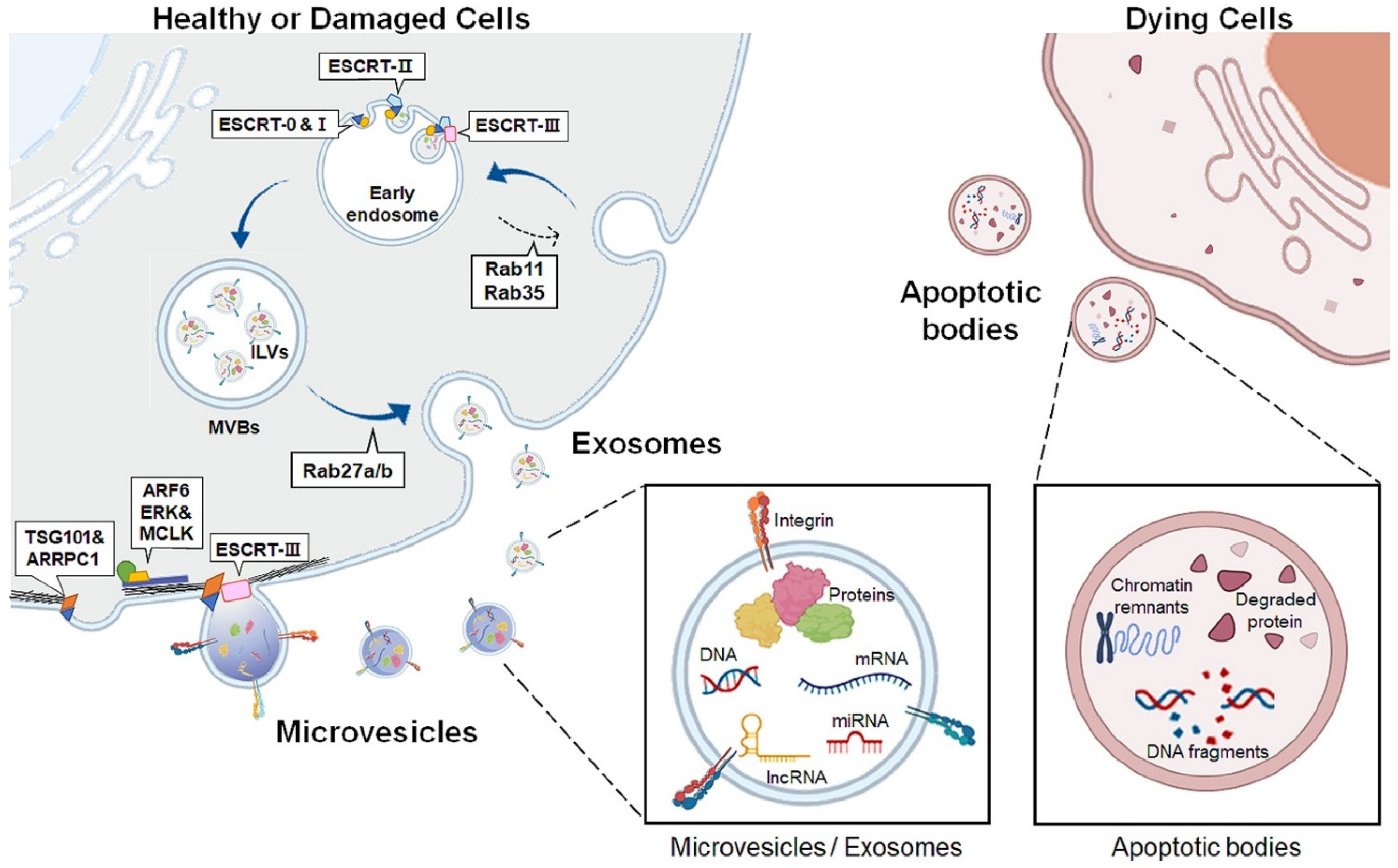
EVs are lipid bilayer membrane-enclosed vesicles released outside the cells. EVs are classified into three subtypes: apoptotic bodies, exosomes, and microvesicles, according to their biogenesis and secretion routes into the extracellular space (Table 1 and Figure 1) [12,18]. Apoptotic bodies, the largest EVs ranging from 1–4 μm in diameter, are generated only during programmed cell death and are considered a hallmark of apoptosis [19,20,21]. In the final phase of apoptosis, dying cells are split into distinct subcellular fragments, termed apoptotic bodies or apoptosomes. These vesicles contain various residual byproducts of dying cells, such as chromatin remnants, degraded proteins, DNA fragments, and intact organelles [22]. Some apoptotic bodies deliver apoptotic contents to phagocytes, such as macrophages or dendritic cells to simulate the immune system, whereas other apoptotic bodies are quickly removed by macrophages or surrounding cells without inducing inflammation [23,24,25].
The exosome sizes range from 30 to 100 nm in diameter [26,27,28]. During exosome biogenesis, early endosomes are produced by inward invagination of the plasma membrane [29]. Subsequently, early endosomal membranes protrude inward to generate intraluminal vesicles (ILVs), finally forming multivesicular bodies (MVBs) [30]. Various proteins including the endosomal sorting complex required for transport (ESCRT) are involved in exosome generation [31,32,33]. ESCRT protein complexes are categorized into four types: ESCRT-0, -I, -II, and -III [34]. The ESCRT-0 complex recognizes and selects ubiquitinated proteins and recruits ESCRT-I to the endosomal membrane [35,36]. ESCRT-I gathers ESCRT-II to form the ESCRT-I/II complex, which promotes the deformation of the endosomal membrane and forms initial buds containing sorted cargoes [37,38]. ESCRT-III, which is responsible for cutting the neck of the bud, is incorporated into the complex, and forms an ILV in MVB lumen [39,40,41]. MVBs are then fused with the plasma membrane to secrete ILVs into the extracellular milieu, and the secreted ILVs are now termed exosomes [42]. Exosome biogenesis is also mediated by an ESCRT-independent pathway. Rab11 and Rab35, small GTPases, induce MVB formation and resend membrane components from endosomes to the plasma membrane [43,44,45]. Rab27a/b is involved in exosome secretion by controlling MVB docking and fusion with the plasma membrane [46].
Microvesicles are larger than exosomes, ranging from 100 to 1000 nm in diameter, and are known as ectosomes or microparticles [47]. Unlike exosomes, microvesicles are generated by budding and pinching outward, directly from the plasma membrane [48,49]. Although the molecular mechanism underlying microvesicle formation is less understood than that of exosome formation, recent studies have suggested that common molecular players involved in exosome formation, such as the ESCRT complex components, participate in microvesicle biogenesis. Tumor susceptibility gene 101 protein (TSG101), a subunit of ESCRT-I, interacts with arrestin domain-containing 1, causing protrusion of the plasma membrane into the extracellular space [50]. The ESCRT-III protein drives membrane curvature and fission of microvesicles [51]. Small GTPases also impact microvesicle formation by promoting actin-myosin rearrangement for microvesicle evagination. ADP-ribosylation factor 6, a member of the rho family, activates phospholipase D to recruit extracellular signal-regulated kinase (ERK) to the plasma membrane [52]. ERK phosphorylates myosin light-chain kinase, which induces contraction of the cytoskeleton, thereby stimulating microvesicle shedding [52].
Accumulating evidence has shown that microvesicles and exosomes have similar biological properties in that they encapsulate many diverse bioactive cargoes, such as cytoskeletal proteins, transporters, adhesion proteins, biogenesis-related proteins, secreted proteins, and metabolites [53,54,55]. Microvesicles and exosomes also deliver genetic material to recipient cells by transferring messenger RNA, microRNA (miRNA), mitochondrial DNA (mtDNA), and long noncoding RNA (lncRNA) [56]. Hence, they act as messengers of cell-to-cell communication by shuttling bioactive substances and genetic materials [57,58]. In addition, their cargo content differs depending on the origin and status of the parental cells [59,60]. Upon injury, contents altered by cellular stress are delivered into target cells and impact their physiological properties, spreading the disease microenvironment to exacerbate disease progression [61,62]. Therefore, microvesicles and exosomes among EVs are emerging as good resources for studying the pathophysiological mechanisms of diseases because they reflect the characteristics of donor cells and play a central role in intercellular communication.
3. EVs Produced in Chronic Liver Disease
Cells release or receive EVs and use EVs as a tool for intercellular communication in both healthy and damaged livers [61]. Hence, EVs allow cells to influence the surrounding cells and alter the liver microenvironment [13]. Alterations in EVs include the amount released and their cargo composition. In damaged livers, secreted levels of EVs containing various harmful factors increase, and EVs deliver pathological cargo to target cells, thereby contributing to CLD progression [63,64]. Hence, EVs have been suggested as attractive biomarkers for diagnosing CLD and evaluating its severity. In this section, we summarize altered EVs according to the types of CLD, nonalcoholic fatty liver disease (NAFLD), alcoholic liver disease (ALD), viral hepatitis, cholestatic liver disease, and liver cancer, and briefly describe their pathogenic roles based on the results reported to date (Table 2). We mainly discuss microvesicles and exosomes because studies on apoptotic bodies in liver disease are lacking. Therefore, EVs are used as umbrella terms for exosomes and microvesicles.
3.1. EVs in NAFLD
NAFLD is the most common chronic liver disease worldwide, with a prevalence of 25.2% [65]. NAFLD is a metabolic disease that occurs in people who consume little to no alcohol and includes simple steatosis, steatohepatitis (NASH), cirrhosis, and even liver cancer [66]. Lipotoxicity causes excessive hepatocyte death, which is a leading cause of NAFLD pathogenesis [67,68,69]. The damaged hepatocytes release several EVs. Srinivas et al. [70] found that the amount of circulating EVs derived from lipotoxic stressed hepatocytes was elevated in both NAFLD patients and animal models. It was also reported that lipotoxicity promoted caspase cascades triggered by the ligand-independent activation of the death 5 receptor, and activated Rho-associated coiled-coil containing protein kinase 1 (ROCK1) signaling, increasing EV secretion from hepatocytes [71]. A damage-regulated autophagy modulator (DRAM) was shown to increase EV release in patients with NAFLD by recruiting stomatin, which induces lysosomal membrane permeabilization, and DRAM deficiency reduced exosome release in HFD-fed mice [72]. Mixed lineage kinase 3 (MLK3) is also involved in increasing EV secretion from lipotoxicity-injured hepatocytes in NASH animal models [73,74]. Endoplasmic reticulum (ER) stress is a key regulator of EV release in hepatocytes exposed to lipotoxic stress. Kakazu et al. [75] demonstrated that palmitate activates inositol-requiring enzyme 1A (IRE1A), an orchestrator of ER stress, to control the de novo biogenesis of ceramide, which in turn increases EV generation in damaged hepatocytes. These findings are supported by Dasgupta et al. [76] who reported that hepatocyte-specific Ire1a-deficiency or Ire1a suppression by an inhibitor significantly reduced EV production in the livers of mice fed high fat, high fructose, and high cholesterol diets.
EVs released from hepatocytes damaged by lipotoxicity carry various biomolecules that aggravate the pathogenesis and progression of NAFLD. MtDNA enriched in EVs is transferred into macrophages and enhances inflammation through the activation of toll-like receptor 9 (TLR9) [77]. Protein cargo within EVs also exacerbates NAFLD progression [78]. C-X-C motif ligand 10, a chemotactic factor for macrophages, in EVs induces hepatic macrophage infiltration in mice with diet-induced NASH [73]. Integrin β1 in EVs increases macrophage recruitment by facilitating monocyte adhesion to liver sinusoidal endothelial cells (LSEC) [79]. TNF-related apoptosis-inducing ligand -bearing EVs stimulated macrophage activation by inducing the production of the pro-inflammatory cytokines interleukin (IL)-1β and IL-6 [71]. Microvesicles from damaged hepatocytes delivered Vanin-1 (VNN1) to endothelial cells to promote angiogenesis, contributing to the progression of inflammation and fibrosis in NASH [80]. Povero et al. [81] also conducted protein profiling in EVs from healthy controls and NASH patients and found that proteins, such as IL-1β and intercellular cell adhesion molecule 2, retained in EVs from NASH patients were related to NASH progression. Furthermore, EVs derived from extrahepatic sources like adipocytes contribute to NAFLD progression. Adipocyte-released EVs upregulated pro-inflammatory cytokines, such as IL-6 and monocyte chemoattractant protein-1, in hepatocytes, and interfered with insulin and gluconeogenesis [82]. Obese adipose tissue-derived EVs were shuttled to HSCs and increased the expression of fibrotic markers, such as issue inhibitor matrix metalloproteinase (TIMP)-1, TIMP-4 and integrin ανβ-5, by dysregulation of TGF-β signaling [83]. Taken together, the increased production of EVs from lipotixicity-damaged hepatocytes and adipocytes is associated with NASH progression through the promotion of inflammation and fibrosis.
3.2. EVs in ALD
ALD is caused by excessive alcohol consumption, unlike NAFLD, and includes steatosis, alcoholic steatohepatitis, cirrhosis, and liver cancer [84]. Ethanol generates large amounts of EVs by increasing ceramide production or ESCR protein expression [85]. Moreover, alcohol obstructs autolysosomal degradation of MVB and elevates EVs release. Upregulated miR-155 by alcohol reduced the expression of multiple target genes regulating autolysosomal degradation, such as the mammalian target of rapamycin, lysosomal-associated membrane protein 1 (LAMP1), and LAMP2, and suppressed the merging of autophagosomes and MVB, enhancing EVs production in hepatocytes and macrophages [86]. Similar to NAFLD patients, patients with ALD also have higher amounts of circulating EVs in the serum than healthy people [87,88]. In alcohol-treated mice, the level of circulating EVs increased in a caspase-dependent manner, similar to the elevation of EV secretion in NAFLD [89]. These EVs are shuttled to neighboring hepatocytes or non-parenchymal cells, and contribute to ALD pathogenesis. Cytochrome P450-enriched EVs released from acetaldehyde-injured hepatocytes have been reported to trigger apoptotic signaling in undamaged target hepatocytes by increasing phospho-c-Jun N-terminal protein kinase, Bax, and active caspase-3 [90]. Alcohol-damaged hepatocytes release cluster differentiation 40 ligand (CD40L)-containing EVs, which promote macrophage activation [89]. EVs released from ethanol-damaged hepatocytes contain higher amounts of damage-associated molecular patterns (DAMP), and activate macrophages via the TLR9 pathway [91]. Ethanol-induced DRAM1 also elevated the secretion of pyruvate kinase M2 (PMK2)-enriched EVs from hepatocytes and activated macrophages, and exacerbated ALD progression [92]. Ma et al. [93] reported that apoptosis signal–regulating kinase 1/p38MAPKα signaling activated by alcohol caused oxidative stress in hepatocytes and led them to undergo apoptosis, enhancing production of mtDNA-enriched EVs from hepatocytes in an ALD mice model. It has been shown that the amount of mtDNA-enriched EVs in serum is elevated in chronically damaged liver by ethanol, promoting inflammation and hepatocyte damage [94]. Ethanol exposure also increased hepatocyte-originated exosomes having plentiful mitochondrial double strand RNA, and these exosomes enhanced IL-1β production in Kupffer cells by activating TLR3. The TLR3-dependent increase of IL-1β upregulated IL-17A in γδ T cells and CD4+ T cells, in the early and late stage of ALD, respectively [95].
In addition, crosstalk between alcohol-damaged hepatocytes and HSCs orchestrates the progression of ALD. EVs from alcohol-fed mice upregulated the expression of fibrotic markers, such as alpha-smooth muscle actin (α-SMA) and type 1 alpha 1 collagen (Col1α1) in primary HSCs from mice. Furthermore, EVs with mtDNA elevate IL-1β and IL-17 levels in hepatic macrophages in a TLR9-dependent manner, subsequently promoting the transdifferentiation of quiescent HSCs into activated HSCs [91]. Activated HSCs also released EVs to promote the activation of quiescent HSCs. EVs from activated HSCs by ethanol have a reduced level of Twist family basic helix-loop-helix transcription factor 1 (Twist1) compared to EVs from quiescent HSCs [96]. Low amounts of exosomal Twist1 inhibited expression of miR-214 targeting connective tissue growth factor (CTGF) in recipient HSCs with inactivation, promoting HSCs activation [96]. Exosomes from activated HSCs contained glycolysis-related proteins such as glucose transporter 1 and PKM2, which induced HSC activation by metabolic conversion [97].
3.3. EVs in Cholestatic Liver
When the liver is damaged, cholangiocytes respond to sustained pro-inflammatory stimuli and show dysregulated proliferation, commonly known as the ductular reaction [98]. Similar to hepatocytes, cholangiocytes are injured in cholestatic liver disease because of the progressive destruction of the bile ducts, the accumulation of bile acids, and the self-perpetuation of inflammation [99,100]. Activated cholangiocytes release EVs with abundant lncRNA H19, which has been shown to be correlated with the severity of cholestatic liver damage in both CLD patients and animal experimental models [101]. The exosomal lncRNA H19 interrupts bile acid homeostasis by blocking the small heterodimeric partner in hepatocytes and promotes HSC activation in the cholestatic liver [102]. It has also been reported that activated cholangiocytes secrete EVs carrying pro-inflammatory cargoes. EVs derived from cholangiocytes were rapidly taken up by Kupffer cells, and the delivered lncRNA H19 upregulated the levels of pro-inflammatory factors, including IL-6 and chemokine (C-C motif) ligand 2 in Kupffer cells [103]. Cholangiocyte-originated EVs delivered DAMP and S100 calcium binding protein A11 (S100A11) to the macrophage to activate it, promoting inflammation [104]. Although the role of EVs in cholestatic liver disease is relatively less known than that of other types of CLD, it is apparent that EVs isolated from bile are closely associated with the pathophysiology of cholangiopathies. Therefore, further studies are needed to explore the various pathogenic cargoes within EVs implicated in the pathogenesis and progression of cholestatic liver diseases.
3.4. Viral Hepatitis
Viral hepatitis refers to the inflammatory condition of the liver due to viral infection and is classified into five main types: hepatitis A, B, C, D, and E [105]. Among these, hepatitis B (HBV) and C (HCV) viruses cause chronic liver disease, with an estimated 296 and 130–170 million people suffering from chronic HBV and HCV, respectively, worldwide [106,107]. Similar to other CLD, EVs play an important role in multiple events in the pathogenesis of viral hepatitis. Hepatitis viruses utilize the EV formation machinery as a means of viral spread. Various viral particles use endosomal cellular complexes that are involved in the production of ILV and MVBs [108,109]. In addition, the hepatitis A virus structural protein Px was reported to increase exosome-like vesicles containing virions and viral proteins in Huh-7 cells, a human hepatoma cell line, by interacting with ALG-2-interacting protein X involved in EV biogenesis [110]. Several studies have demonstrated that virus-infected hepatocytes release large amounts of viral RNA-carrying EVs, which cause hepatic inflammation and liver fibrosis. Exosomes released from HBV-transfected hepatocytes induced natural killer group 2, member D ligand in macrophages in a myeloid differentiation factor 88-dependent manner and enhanced hepatic inflammation [111]. HBV-RNA-enriched EVs from hepatocytes upregulate type 1 interferon in dendritic cells [112]. EVs released from hepatitis virus-infected cells help the hepatitis virus to escape from immune reactions by interrupting the function of the immune cells. When EVs with HBV RNA or DNA were transferred to natural killer cells isolated from healthy people, the expression of inflammatory cytokines, such as IFN-γ and tumor necrosis factor (TNF)-α, in these cells decreased [113]. HCV-infected hepatocytes released EVs containing increased levels of galectin-9 led to the suppression of T cell activation by elevating programmed cell death-ligand-1 in monocytes [114]. In addition, EVs from hepatocytes transfected with HCV mediated polarization of monocytes into the M2 phenotype, and induced HSC activation and liver fibrosis [115]. Nonparenchymal cells infected with virus also participated in hepatitis progression. HCV particles were taken up by LSECs, and enhanced type 1 and 3 interferon in these cells [116]. These findings suggest that EVs are involved in viral expansion and the immune responses caused by viral hepatitis.
3.5. EVs in Liver Cancer
Liver cancer is the fourth leading cause of cancer-related deaths worldwide [117]. Hepatocellular carcinoma (HCC) is the most common primary liver cancer, and together with cholangiocarcinoma (CCA), comprise approximately 12–15% of liver cancer cases [118]. EVs produced in the cancer microenvironment also mediate liver cancer progression by influencing cell proliferation, invasion, and metastasis. Proteome and transcriptome analyses of the contents within HCC-derived exosomes showed that they retained numerous oncogenic factors, such as caveolin, RAS related, S100A4, and S100A11, which promoted migration and invasion of immortalized hepatocytes by activating PI3K/AKT and MAPK signaling and increasing the secretion of active matrix metalloproteinases (MMP)-2 and MMP9 [119]. S100A protein family mediates cell migration, invasion, and metastasis. S100A11 protein enhanced HCC tumorigenesis and angiogenesis by inducing vascular tube formation and was found to be highly correlated with the poor survival of HCC patients [120]. In line with these findings, protein profiling of cancer-derived exosomes identified 129 proteins implicated in HCC progression. Among these, adenylyl cyclase-associated protein 1 was remarkably enriched in exosomes released from HCC cells with high metastatic potential [121].
In addition, both levels of 14-3-3ζ mRNA and protein in exosomes were significantly higher in HCC patients with poor survival. 14-3-3ζ expression abolished the antitumor effect of tumor-infiltrating T lymphocytes and evaded the immune response through T cell exhaustion [122]. Xie et al. [123] demonstrated that angiopoietin-2-containing exosomes were delivered into endothelial cells and induced angiogenesis by increasing AKT/eNOs and AKT/β-catenin signaling. Although lncRNAs are present in low amounts in EVs, TUC339, a 1198-base paired lncRNA, is elevated in EVs obtained from HCC cells and promotes HCC cell proliferation and adhesion [124,125]. Conigliaro et al. [126] found that CD90-positive liver cancer cells delivered the lncRNA H19 within exosomes to endothelial cells and stimulated angiogenesis. Increased exosomal SMAD family member 3 (SMAD3) in patients with HCC promotes lung metastases by enhancing circulating primary tumor adhesion [127]. HCC cells-derived exosomes transferred lysyl oxidase-like 4 between HCC cells and modulated cell-matrix adhesion and tumor invasion via activating the focal adhesion kinase/Src signaling pathway [128].

{kind=link}
Table 2.
Summary of EVs and their cargoes associated with progression of CLD.
| CLD | EVs Source | Biological Fluids | Cargo | Target | Effects | Ref. |
|---|---|---|---|---|---|---|
| NAFLD | Hepatocytes | plasma | mtDNA | Macrophage | Increased inflammation | [77] |
| Hepatocytes | Cell culture medium, serum | CXCL10 | Macrophage | Increased hepatic macrophage infiltration | [73] | |
| Hepatocytes | Cell culture medium, serum | Integrin β1 | Monocyte | Increased adhesion to LSEC | [79] | |
| Hepatocytes | Cell culture medium, serum | TRAIL | Macrophage | Increased inflammation | [71] | |
| Hepatocytes | Cell culture medium, plasma | VNN1 | LSECs | Promoted angiogenesis | [80] | |
| NASH patients | Serum | IL-1β, ICAM2 | - | Related with NASH progression | [81] | |
| Adipocyte | Cell culture medium | IL-6, MCP-1 | Hepatocytes | Interfered with insulin and gluconeogenesis | [82] | |
| Adipocyte | Cell culture medium | - | HSCs | Increased fibrotic marker expression | [83] | |
| ALD | Hepatocytes | Cell culture medium, plasma | CYP2E1 | Hepatocytes | Increased hepatocyte apoptosis | [90] |
| Hepatocytes | Cell culture medium, serum | CD40L | Macrophage | Induced macrophage activation | [89] | |
| Hepatocytes | Cell culture medium | DAMP | Macrophage | Increased inflammation | [91] | |
| Hepatocytes | mtDNA | Macrophage | Enhanced HSCs activation Increased fibrotic marker expression | |||
| EtOH-fed mice | Serum | - | HSCs | |||
| Hepatocytes | Serum | PKM2 | Macrophage | Increased inflammation | [92] | |
| Hepatocytes | Serum | mtDNA | Neutrophil Hepatocytes | Increased inflammation and hepatocyte injury | [93,94] | |
| Hepatocytes | Cell culture medium | mtdsRNA | Kupffer cells | Increased inflammation | [95] | |
| Activated HSCs | Cell culture medium | Twist1 | HSCs | Enhanced HSCs activation | [96] | |
| Activated HSCs | Cell culture medium | GLUT1, PKM2 | HSCs | Enhanced HSCs activation | [97] | |
| Cholestatic liver | Cholangiocyte | Cell culture medium, serum | lncRNA H19 | Hepatocytes HSCs | Interrupted bile homeostasis Promoted HSCs activation | [102] |
| Cholangiocyte | Cell culture medium | lncRNA H19 | Kupffer cells | Upregulated pro-inflammatory cytokines | [103] | |
| Cholangiocyte | Cell culture medium | DAMP, S100A11 | Macrophage | Increased inflammation | [104] | |
| Viral hepatitis | Hepatocytes | Cell culture medium | - | Macrophage | Increased inflammation | [111] |
| Hepatocytes | Cell culture medium | HBV-RNA | Dendritic cells | Increased inflammation | [112] | |
| Hepatocytes | Serum | HBV-RNA/DNA | Natural Killer cells | Help hepatitis virus to escape from immune reaction | [113] | |
| Hepatocytes | Cell culture medium | - | Monocyte | Suppression of T-cell activation | [114] | |
| Hepatocytes | Cell culture medium | - | Monocyte | Enhanced HSCs activation | [115] | |
| LSEC | Cell culture medium | - | Hepatocytes | Controlled HCV replication | [116] | |
| Liver cancer | HCC cells | Cell culture medium | Caveolin, RRAS, S100A4, S100A11 | Immortalized hepatocytes | Stimulated migration and invasion of immortalized hepatocytes | [119] |
| HCC cells | Cell culture medium | CAP1 | - | Related with high metastatic potential | [121] | |
| HCC cells | Cell culture medium, Serum | 14-3-3ζ | T-lymphocytes | Inhibited anti-tumor effects of T-lymphocytes | [122] | |
| HCC cells | Cell culture medium, Serum | ANGPT2 | Endothelial cells | Promoted angiogenesis | [123] | |
| HCC cells | Cell culture medium | TUC339 | HCC cells | Elevated HCC cell proliferation and adhesion. | [125] | |
| CD90-positive liver cancer cells | Cell culture medium | lncRNA H19 | Endothelial cells | Promoted angiogenesis | [126] | |
| HCC cells | Cell culture medium | SMAD3 | Detached HCC cells | Promotes lung metastases by enhancing circulating primary tumor adhesion | [127] | |
| HCC cells | Cell culture medium | LOXL4 | HCC cells | Promoted invasion | [128] |
CLD, chronic liver disease; NAFLD, nonalcoholic liver disease; mtDNA, mitochondrial DNA; CXCL10, C-X-C motif ligand 10; LSEC, liver sinusoidal endothelial cells; TRAIL, TNF-related apoptosis-inducing ligand; VNN1,vanin-1; IL-1β, interleukin-1β; ICAM2, intercellular cell adhesion molecule 2; ALD, alcoholic liver disease; CYP2E1, cytochrome P450 2E1; CD40L, cluster differentiation 40 ligand; DAMP, damage-associated molecular patterns; mtDNA, mitochondrial DNA; PKM2, pyruvate kinase M2; mtdsRNA, mitochondrial double strand RNA; Twist1, Twist family basic helix-loop-helix transcription factor 1; HSCs, hepatic stellate cells; GLUT1, glucose transporter 1; EtOH, ethanol; lncRNA, long noncoding RNA; S100A11, S100 calcium binding protein A11; HBV, hepatitis B virus; LSEC, liver sinusoidal endothelial cell; HCC, hepatocellular carcinoma; RRAS, Ras related; S100A4, S100 calcium binding protein A4; CAP1, adenylyl cyclase-associated protein 1; ANGPT2, angiopoietin-2; SMAD3, SMAD family member 3; LOXL4, lysyl oxidase-like 4.
4. Contribution of miRNAs in EVs to CLD
A growing body of evidence has revealed the contribution of various cargoes in EVs to CLD [129,130]. Among these, miRNAs are one of the most well-studied biomolecules within EVs [131]. MiRNAs are the evolutionarily conserved ncRNA of 19–24 nucleotides in length and function as post-transcriptional regulators of gene expression [132,133]. MiRNAs, transferred into recipient cells by EVs, impact cell behaviors by binding target sequences of mRNAs to interfere with the translational machinery [129,130]. In patients with CLD, elevated EVs carry multiple pathological miRNAs, resulting in the exacerbation of liver damage [134,135]. In addition, disease-specific alterations of miRNAs within EVs are in the spotlight as precise diagnostic biomarkers for distinguishing different types and progression stages of CLD [134,135]. Hence, in this section, we review the roles of EV-mediated miRNA in intercellular communications following each CLD and further explain the potential of miRNA as a disease-specific diagnostic marker (Table 3).
4.1. EV-Derived miRNAs in NAFLD
Along with the increase in circulating EVs released from damaged hepatocytes in patients with NAFLD and animal models mimicking NAFLD, the actions of miRNAs in EVs have recently been emphasized in the pathogenesis and progression of NAFLD [78,136]. The analysis of miRNAs in EVs isolated from patients or experimental animal models revealed a significant difference in the expression of various miRNAs. Newman et al. [136] analyzed serum EVs isolated from patients with NAFLD and found that the levels of miR-122, miR-192, and miR-128-3p were significantly higher in liver-specific asialoglycoprotein receptor 1 (ASGPR1)-positive EVs than in healthy controls, suggesting the correlation of miRNA levels with NAFLD severity. In line with these findings, Povero et al. [78] reported that miR-122 and miR-192 in liver-specific ASGPR1-positive EVs were upregulated in choline-deficient, L-amino acid-defined diet (NAFLD inducing diet) -fed mice compared to control diet-fed mice. In a rat model of NAFLD caused by high-fat and high-cholesterol diet-fed rats, miR-192-5p in EVs from injured hepatocytes was significantly increased compared to the control group [137]. MiR-192-5p-enriched EVs promoted M1 polarization by regulating the Rictor-Akt-FoxO1 axis and led to the activation of pro-inflammatory macrophages [137]. MiR-122 was also significantly higher in circulating EVs from both NAFLD patients and rodent models and inhibited AMP-activated protein kinase (AMPK) signaling [138]. Because AMPK is a major regulator of fatty acid and cholesterol metabolism, the effect of miR-122 on AMPK signaling promotes NAFLD progression by interfering with lipid metabolism [138,139]. Cholesterol exposure increased production of miR-122-5p-contained exosomes in hepatocytes, and exsomal miR-122-5p promoted hepatic inflammation by M1 polarization of the macrophage [140]. In addition, EVs from lipotoxicity-stressed hepatocytes delivered miR-128-3p to HSCs to downregulate peroxisome proliferator-activated receptor gamma and activate HSCs, promoting liver fibrosis [141]. Exosomal miR-1297 and miR-27a from damaged hepatocytes were transferred to and activated HSCs. MiR-1297-enriched exosomes activated HSCs through the phosphatase and tensin homolog (PTEN)/phosphoinositide 3-kinase/protein kinase B signaling pathway, accelerating the progression of NALFD [142]. Exosomal miR-27a downregulated PTEN-induced expression of putative protein kinase 1(PINK1) in HSCs [143]. Reduced RINK1 inhibited mitophagy inducing HSC apoptosis, and resulted in the proliferation and activation of HSCs [143,144].
4.2. EV-Derived miRNAs in ALD
Alcohol-damaged livers also produced significantly increased amounts of EVs containing miRNAs [88,145]. Momen-Heravi et al. [88] reported that several miRNAs, including miR-130a, miR-30a, miR-192, miR-1246, and miR-744, were significantly higher in circulating EVs from chronic EtOH-fed mice than in control mice. EVs isolated from the ALD mouse model showed the enrichment of miR-29a and miR-340, and these miRNAs were also identified in EVs from the blood of patients with ALD [145]. The increase in specific miRNAs in EVs contributes to the progression of ALD by influencing various pathways. It has been reported that alcohol elevates EV production in monocytes, a type of leukocyte, and transfers miR-27a into adjacent naive monocytes to trigger the polarization of monocytes to M2 macrophages, promoting liver fibrosis [146,147]. Alcohol causes hepatocytes to secrete miR-122-rich exosomes, which are taken up by monocytes. Delivered miR-122 blocks the heme oxygenase 1 pathway in monocytes and increases their sensitivity to lipopolysaccharide and pro-inflammatory cytokines, such as IL-1β and TNFα, contributing to the pathogenic progression of alcoholic hepatitis [148]. Additionally, exosomal miRNAs isolated from activated HSCs have been shown to induce hepatic fibrogenesis. The miR-17-92 cluster, which includes miR-17a, miR-19a, miR-19b, miR-20a, and miR-92, is known to have various functions in cellular pathways, such as inflammation, proliferation, apoptosis, and necrosis [149,150]. MiR-92 is upregulated in EVs isolated from the conditioned media of activated HSCs and the serum of alcohol-injured mice [149]. It has been reported that miR-92 regulates TGF-β by binding to the 3′ untranslated region of SMAD7, although the direct effect of EV-delivered miR-92 on HSC activation has not been studied [149,151]. To assure the impact of miR-92 on ALD to apply miR-92 for diagnostic markers, further study is required.
4.3. EV-Derived miRNAs in Viral Hepatitis
Several miRNAs have been characterized in viral hepatitis, including the HBV and HCV [152]. In HBV-infected hepatocytes, miR-3 is produced and secreted extracellularly via EVs [153]. Increased miR-3 expression by HBV infection reduces suppressor of cytokine signaling (SOCS)5 expression and activates the JAK/STAT signaling pathway in macrophages to promote M1 polarization, eventually stimulating the secretion of IL-6, a pro-inflammatory cytokine that affects innate immune responses. EVs originated from HCV-infected cells also contribute to fibrosis [153]. Devhare et al. [154] showed that miR-19a contained in EVs from HCV-infected hepatocytes targeted SOCS3 to activate the STAT3-TGF-β pathway, and resulted in HSC activation. MiR-192 from an HCV-infected hepatocyte line directly influences HSC activation by upregulating TGF-β [155]. In addition, diagnostic studies of circulating miRNAs in patients have been performed. Jin et al. [156] profiled circulating miRNAs in the blood of patients with chronic hepatitis B (CHB). Among the 53 miRNAs, 22 were differentially expressed in CHB patients compared with that in healthy controls. In particular, CHB patients with high levels of HBV in the serum showed a significant increase in miR-122-5p levels compared to that in patients with low levels of HBV [156]. Meanwhile, the levels of miR-1246, miR-150-5p, miR-5787 and miR-8069 in plasma EVs were significantly lower in HBV patients than in healthy controls [157]. These findings indicate that deregulated miRNAs are involved and have potential as novel diagnostic tools for viral hepatitis.
4.4. EV-Derived miRNAs in Liver Cancer
MiRNAs providing a favorable microenvironment for the proliferation of cancer cells are upregulated in EVs from HCC patients and animal models [158,159]. The levels of pro-proliferative and pro-migratory miR-222, miR-221, and miR-18a were higher, and the expression of pro-apoptotic and anti-growth miR-101, miR-122, and miR-195 was lower in the serum of patients with HCC than in those with cirrhosis [158]. Yang et al. [160] demonstrated that increased miR-3129 in plasma EVs from HCC patients promotes the proliferation of cancer cells and epithelial to mesenchymal transition by targeting thioredoxin-interacting protein, an important inhibitor of glucose uptake and cell proliferation. MiR-378b in EVs derived from HCC cells promoted angiogenesis by targeting TGF-β receptor III in endothelial cells [161]. MiRNA-584-5p in EVs from HCC cells also induced angiogenesis by inhibiting phosphoenolpyruvate carboxykinase 1-mediating nuclear factor erythroid-2-related factor 2 activation [162]. Highly metastatic HCC cells secrete EVs carrying miR-1247-3p, which directly targets β-1,4-Galactosyltransferase 3 and activates β1-integrin-NF-κB signaling in fibroblasts [163]. Activated fibroblasts secrete pro-inflammatory cytokines promoting cancer progression [163]. The MiRNA profiling of exosomes from CCA, the second most common primary malignant liver cancer with aberrant proliferation of cholangiocytes, showed that miR-205-5p was highly upregulated in CCA-derived exosomes compared to that in normal cholangiocytes, and its expression level paralleled the capacities of invasion and migration of CCA cells [164]. Ni et al. revealed that exosomal miR-23a-3p from CCA cells promoted tumor growth and metastasis by downregulating dynamin-3 [165].
In addition to their roles as influencers of liver cancer progression, exosomal miRNAs have been suggested as biomarkers for predicting the recurrence of HCC [166,167]. Exosomal miR-718, which suppresses HCC cell proliferation by targeting homeobox B8, is related to cancer prognosis. Sugimachi et al. [168] found that miR-718 expression was higher in circulating EV from HCC patients who underwent liver transplantation without recurrence, whereas its expression was lower in EVs from patients with HCC recurrence, suggesting that the level of exosomal miR-718 is related to HCC aggressiveness. A recent study analyzing clinicopathological data presents the significant upregulation of exosomal miR-92a-3p in patients with liver cancer having tumors larger than 5cm in diameter [169]. Li et al. [170] also revealed that the expression of miR-191, miR-486-3p, miR-1274b, miR-16, and miR-484 was significantly increased in circulating EVs isolated from the bile of CCA patients compared with those isolated from bile of patients with bile leak syndrome, benign biliary obstruction, or primary sclerosing cholangitis.
Based on the disease-specific expression and stability of miRNAs in EVs and the easier detection with non-invasive techniques, they are considered diagnostic markers for specific diseases [171,172]. Hence, numerous preclinical studies on miRNAs in EVs have been conducted [129,130,173]. However, most studies have focused on profiling miRNAs in EVs from specific liver diseases using meta-analysis without providing the detailed roles and mechanisms explaining their roles. Using miRNAs in EVs as markers for the diagnosis or prognosis of disease is still in an early stage because of the lack of clinical evidence for miRNAs in EVs. Therefore, further research to elucidate their roles and relevant mechanisms in liver cancer is necessary to establish them as reliable and novel diagnostic markers for liver diseases.
Table 3.
Summary of EV-associated miRNAs in progression of CLD.
| CLD | EVs Source | Biological Fluids | miRNA | Target | Effects | Ref. |
|---|---|---|---|---|---|---|
| NAFLD | NAFLD patients | Serum | Upregulated miR-122 miR-192 miR-128-3p | - | Related with NAFLD progression | [136] |
| NAFLD mice model | Plasma | Upregulated miR-122 miR-192 | - | Related with NAFLD progression | [78] | |
| Hepatocytes | Cell culture medium, Serum | miR-192-5p | Macrophages | Triggered M1 polarization | [137] | |
| Hepatocytes | Serum | miR-122 | Hepatocytes | Dysregulated lipid metabolism | [138] | |
| Hepatocytes | Cell culture medium | miR-122-5p | Macrophages | Triggered M1 polarization | [140] | |
| Hepatocytes | Cell culture medium | miR-128-3p | HSCs | Enhanced HSCs activation | [141] | |
| Hepatocytes | Cell culture medium, serum | miR-1297 miR-27a | HSCs | Enhanced HSCs activation | [142,143] | |
| ALD | EtOH-fed mice | Serum or Plasma | Upregulated miR-130a miR-30a miR-192 miR-1246 miR-744 | - | Related with ALD progression | [88] |
| Hepatocytes-isolated from EtOH-fed mice/ ALD patients | Cell culture medium/ Serum | Upregulated miR-29a miR-340 | - | Related with ALD progression | [145] | |
| Monocytes | Cell culture medium | miR-27a | Naive monocytes | Promoted liver fibrosis | [146,147] | |
| Hepatocytes | Cell culture medium, Serum | miR-122 | Monocytes | Increased inflammation | [148] | |
| Activated HSCs | Cell culture medium, Serum | miR-92 | HSCs | Enhanced HSCs activation | [149] | |
| Viral hepatitis | HBV-infected hepatocytes | Cell culture medium | miR-3 | Macrophages | Increased inflammation | [153] |
| HCV-infected hepatocytes | Cell culture medium | miR-19a | HSCs | Enhanced HSCs activation | [154] | |
| HCV-infected hepatocytes line | Cell culture medium | miR-192 | HSCs | Enhanced HSCs activation | [155] | |
| Chronic hepatitis B patients | Serum | miR-122-5p | - | Related with level of HBV | [156] | |
| Chronic hepatitis B patients | plasma | Downregulated miR-1246 miR-150-5p miR-5787 miR-8069 | - | Related with level of HBV | [157] | |
| Liver cancer | HCC patients | Serum | Upregulated miR-222 miR-221 miR-18a | - | Elevated proliferation and migration of HCC cells | [158] |
| Serum | Downregulated miR-101 miR-122 miR-195 | - | Inhibited growth and apoptosis of HCC cells | |||
| HCC cells | plasma | miR-3129 | HCC cells | Elevated HCC cells proliferation. | [160] | |
| HCC cells | Cell culture medium | miR-378b | Promoted angiogenesis | [161] | ||
| HCC cells | Serum | miR-584-5p | Promoted angiogenesis | [162] | ||
| HCC cells | Cell culture medium | miR-1247-3p | Fibroblasts | Increased inflammation | [163] | |
| CCA cells | Cell culture medium | miR-205-5p | CCA cells | Increased invasion and migration | [164] | |
| CCA cells | Cell culture medium | miR-23a-3p | Promoted tumor growth and metastasis | [165] | ||
| HCC patients | Serum | Downregulated miR-718 | - | Biomarker for predicting recurrence of HCC | [168] | |
| Liver cancer fibroblast /Liver cancer tissue | Cell culture medium | miR-92a-3p | Related with progress of HCC | [169] | ||
| Bile from CCA patients | Bile | Upregulated miR-191 miR-486-3p miR-1274b miR-16 miR-484 | - | Biomarker for CCA | [170] |
CLD, chronic liver disease; miRNA, microRNA; NAFLD, nonalcoholic liver disease; HSCs, hepatic stellate cells; ALD, alcoholic liver disease; EtOH, ethanol; HBV, hepatitis B virus; HCV, hepatitis C virus; HCC, hepatocellular carcinoma; CCA, cholangiocarcinoma.
5. Possibility of EVs as Therapeutics for Liver Disease
As mentioned above, the amount of EVs released which carry pathological molecules was significantly increased in most CLD cases, and EVs, including their cargoes, participate in the onset and progression of CLD by affecting cell-cell communication. Hence, the inhibition of pathological EV generation and secretion and of the uptake of altered cargo by target cells has been proposed as a strategy to alleviate CLD. First, it is possible to interfere with the action of EVs in CLD by reducing EV production and/or secretion. A locked nucleic acid (LNA) is modified oligonucleotide as an anti-miRNA that has high and stable affinity for the target [174]. Momen-Heravi et al. [175] reported that LNA-miR-132 reduced EV secretion and liver fibrosis. Pharmacological inhibitors of MLK3 lowered the levels of pro-inflammatory cytokines-contained EVs from injured hepatocytes in a NASH mouse model [73]. The exosome biogenesis inhibitor GW4689 suppressed the formation of ILVs and the delivery of intracellular RNA from hepatocytes into HSCs [85]. GW4689 reduces CTGF-contained EVs from activated HSCs and inhibits the activation of quiescent HSCs [176]. GW4689 also inhibited exosome production from white adipose tissue and alleviated the liver damage caused by a high-fat diet [177]. Fasudil, an inhibitor of ROCK1, has been shown to reduce circulating EVs and attenuate liver injury, inflammation, and fibrosis in a NASH mouse model [71]. It also reduced the release of EVs containing CD40L in damaged hepatocytes and decreased macrophage activation and subsequent hepatic injury [89]. Another strategy is to interrupt the actions of pathological cargo loaded within the EVs. Hepatocytes stressed by lipotoxicity release sphingosine 1-phosphate (S1P)-abundant EVs and recruit macrophages. Suppression of sphingosine kinases (SK) 1 and 2, which play an important role in S1P enrichment, eliminates chemoattraction to macrophages [178]. Administration of the S1P antagonist FTY720 ameliorated liver damage, inflammation, and fibrosis in a NASH mouse model [179]. A TLR9 antagonist abrogated the action of mtDNA in EVs from HFD-fed mice to inhibit pro-inflammatory responses in macrophages in a murine NASH model [77]. Wang et al. [180] found that the ER stress inhibitor,4-phenybutric acid reduced exosomal miR-122 and attenuated liver injury in a mouse model of ALD. In addition, blocking the binding of EV to recipient cells restrains the effect of EVs on CLD. Wang et al. [181] demonstrated that pharmacological inhibitors blocking the link between fibronectin and integrin α5β1 reduced the binding of endothelial cell-derived exosomes to the HSC surface, and exosomal SK1 and S1P rarely promoted HSC activation and migration. Neutralization of VNN1 within EVs released from lipotoxicity-damaged hepatocytes abolished EV uptake by endothelial cells and hardly brought pro-angiogenic effects to these cells [80].
In addition, EVs could be used as delivery vehicles carrying small interfering RNAs or chemotherapeutic reagents. Because they are natural biocarriers derived from endogenous cells, EVs have many advantages, such as high in vivo stability and low cytotoxicity and immunogenicity [182,183,184]. Given that approximately 60% of intravenously injected EVs are delivered to the liver, a strategy using EVs as a delivery tool might be effective in treating liver disease [185]. It was shown that HEK293T cell-derived exosomes loading artificially RBP-J hairpin-decoy oligodeoxynucleotide (ODN) were transferred into hepatic macrophages and reduced Notch activation in macrophages, attenuating liver fibrosis [186]. Momen-Heravi et al. [187] reported that B cell-released EVs engineered with carrying miR-155 inhibitor suppressed LPS-induced TNF-α production in macrophages in vitro. In line with these findings, Zhang et al. [188] demonstrated that red blood cell-released EVs (RBC-EVs) having miR-155 antisense oligonucleotides (ASO) were phagocytosed by hepatic macrophage in a C1q-dependent manner, and miR-155 ASO alleviated acute liver failure by inducing the M2 polarization of macrophages. They also showed that RBC-EVs engineered to carry doxorubicin or sorafenib effectively delivered these therapeutic drugs into the liver, and inhibited orthotopic cancer growth. Ji et al. [189] encapsulated fibroblast-originated EVs with Clodronate to escape from non-specific phagocytosis by Kupffer cells, and then loaded nintedanib (NIN), an anti-fibrosis agent, in these EVs. These modified EVs effectively ameliorated liver fibrosis by inhibiting the proliferation of hepatic fibroblasts. These data suggest EVs as the safe and effective drug delivery tool for CLD.
However, there are many points that need to be considered, although the regulation of EV biology has recently emerged as a new strategy to treat CLD. If engineered EVs are unexpectedly delivered to cells other than the target cells, it could damage both the targeted and non-targeted cells. The inhibition of EV biogenesis, the uptake by target cells, and the action of EV contents might interfere with other vital processes, such as autophagy, cell differentiation, and cytoskeleton reorganization [190]. For example, GW4689 could disturb starvation-induced autophagy by inhibiting neutral sphingomyelinase 2 that facilitates autophagic flux by increasing Golgi-localized ceramide [191]. Imipramine, which inhibits EV release by suppressing ceramide generation causes many adverse effects, such as nightmares, nausea, drowsiness, low blood pressure, and dizziness [192]. Therefore, it is necessary to enhance the safety, efficacy, and targeting of EV inhibitors to apply them in treating liver diseases.
6. Conclusions
EVs are generated in all types of liver cells and provide bioinformation that reflects the characteristics of the parental cells [9,10]. Accumulating evidence indicates that they mediate intercellular communication between liver cells in hepatic pathobiology [6,7,8]. EVs derived from damaged cells are delivered into recipient cells and spread the pathological condition of EV-producing cells to EV-receiving cells, leading to alternation of the liver microenvironment [13]. In CLD, the production and secretion of EVs are significantly elevated, and their cargo composition is altered. Higher amounts of bioactive substances, including proteins and genetic materials, are packed into EVs to enhance inflammation, fibrosis, and angiogenesis [16,17]. MiRNAs are the most well-studied cargo groups within EVs. MiRNAs in EVs play important roles in CLD by regulating gene expression in target cells and accelerating disease progression [134,135]. Therefore, EVs and their cargo have attracted interest as potential biomarkers and important targets for the development of therapeutics for CLD [64]. However, several challenges remain to be overcome in EV applications in CLD, despite the great potential of EVs. The detailed mechanism underlying the characteristic changes in EVs in response to liver damage has not yet been fully elucidated; for example, how EV biosynthesis is increased and pathogenic cargoes are sorted and loaded in EVs is not known. In addition, studies on EV contents involved in CLD progression are limited to miRNAs, and research on other cargoes, including proteins and lncRNAs, is relatively insufficient. Further investigations to solve these challenges will open the door to understanding the role of EVs in the development and progression of CLD and provide promising diagnostic tools and therapeutic strategies for CLD.
Author Contributions
C.L. and J.H. contributed to this paper with literature review and drafting the manuscript; Y.J. contributed to this paper with conception, review, drafting and editing the manuscript, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Y.J and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) to C.L. (No. 2022R1C1C2008830).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Figure 1 was partly created with Biorender.com (last accessed on 29 March 2022).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Pinzani, M.; Rombouts, K. Liver fibrosis: From the bench to clinical targets. Dig. Liver Dis. 2004, 36, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Suresh, D.; Srinivas, A.N.; Kumar, D.P. Etiology of hepatocellular carcinoma: Special focus on fatty liver disease. Front. Oncol. 2020, 10, 601710. [Google Scholar] [CrossRef]
- Karlsen, T.H.; Lammert, F.; Thompson, R.J. Genetics of liver disease: From pathophysiology to clinical practice. J. Hepatol. 2015, 62, S6–S14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef]
- Yang, D.; Liu, J. Targeting extracellular vesicles-mediated hepatic inflammation as a therapeutic strategy in liver diseases. Liver Int. 2020, 40, 2064–2073. [Google Scholar] [CrossRef]
- Jung, Y.; Witek, R.P.; Syn, W.K.; Choi, S.S.; Omenetti, A.; Premont, R.; Guy, C.D.; Diehl, A.M. Signals from dying hepatocytes trigger growth of liver progenitors. Gut 2010, 59, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Maji, S.; Matsuda, A.; Yan, I.K.; Parasramka, M.; Patel, T. Extracellular vesicles in liver diseases. Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 312, G194–G200. [Google Scholar] [CrossRef]
- Magni, F.; Van Der Burgt, Y.E.; Chinello, C.; Mainini, V.; Gianazza, E.; Squeo, V.; Deelder, A.M.; Kienle, M.G. Biomarkers discovery by peptide and protein profiling in biological fluids based on functionalized magnetic beads purification and mass spectrometry. Blood Transfus. 2010, 8 (Suppl. 3), s92. [Google Scholar] [CrossRef]
- Pulliero, A.; Pergoli, L.; La Maestra, S.; Micale, R.; Camoirano, A.; Bollati, V.; Izzotti, A.; De Flora, S. Extracellular vesicles in biological fluids. A biomarker of exposure to cigarette smoke and treatment with chemopreventive drugs. J. Prev. Med. Hyg. 2019, 60, E327–E336. [Google Scholar] [CrossRef]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular vesicles: Composition, biological relevance, and methods of study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, S.; Kim, J.; Jung, Y. Liver-derived exosomes and their implications in liver pathobiology. Int. J. Mol. Sci. 2018, 19, 3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde-Vancells, J.; Rodriguez-Suarez, E.; Embade, N.; Gil, D.; Matthiesen, R.; Valle, M.; Elortza, F.; Lu, S.C.; Mato, J.M.; Falcon-Perez, J.M. Characterization and comprehensive proteome profiling of exosomes secreted by hepatocytes. J. Proteome Res. 2008, 7, 5157–5166. [Google Scholar] [CrossRef] [Green Version]
- Nojima, H.; Freeman, C.M.; Schuster, R.M.; Japtok, L.; Kleuser, B.; Edwards, M.J.; Gulbins, E.; Lentsch, A.B. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J. Hepatol. 2016, 64, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Azparren-Angulo, M.; Royo, F.; Gonzalez, E.; Liebana, M.; Brotons, B.; Berganza, J.; Goñi-de-Cerio, F.; Manicardi, N.; Abad-Jordà, L.; Gracia-Sancho, J.; et al. Extracellular vesicles in hepatology: Physiological role, involvement in pathogenesis, and therapeutic opportunities. Pharmacol. Ther. 2021, 218, 107683. [Google Scholar] [CrossRef]
- Garcia-Martinez, I.; Alen, R.; Rada, P.; Valverde, A.M. Insights into extracellular vesicles as biomarker of NAFLD pathogenesis. Front. Med. 2020, 7, 395. [Google Scholar] [CrossRef]
- Turchinovich, A.; Drapkina, O.; Tonevitsky, A. Transcriptome of extracellular vesicles: State-of-the-Art. Front. Immunol. 2019, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Hristov, M.; Erl, W.; Linder, S.; Weber, P.C. Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood 2004, 104, 2761–2766. [Google Scholar] [CrossRef] [PubMed]
- Battistelli, M.; Falcieri, E. Apoptotic bodies: Particular extracellular vesicles involved in intercellular communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Turiák, L.; Misják, P.; Szabó, T.G.; Aradi, B.; Pálóczi, K.; Ozohanics, O.; Drahos, L.; Kittel, A.; Falus, A.; Buzás, E.I.; et al. Proteomic characterization of thymocyte-derived microvesicles and apoptotic bodies in BALB/c mice. J. Proteom. 2011, 74, 2025–2033. [Google Scholar] [CrossRef]
- Lleo, A.; Zhang, W.; McDonald, W.H.; Seeley, E.H.; Leung, P.S.; Coppel, R.L.; Ansari, A.A.; Adams, D.H.; Afford, S.; Invernizzi, P.; et al. Shotgun proteomics: Identification of unique protein profiles of apoptotic bodies from biliary epithelial cells. Hepatology 2014, 60, 1314–1323. [Google Scholar] [CrossRef] [Green Version]
- van der Pol, E.; Hoekstra, A.G.; Sturk, A.; Otto, C.; van Leeuwen, T.G.; Nieuwland, R. Optical and non-optical methods for detection and characterization of microparticles and exosomes. J. Thromb. Haemost. 2010, 8, 2596–2607. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Jeyaraj, M.; Qasim, M.; Kim, J.H. Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Pertierra, E.; Oliveira-Rodríguez, M.; Rivas, M.; Oliva, P.; Villafani, J.; Navarro, A.; Blanco-López, M.C.; Cernuda-Morollón, E. Characterization of plasma-derived extracellular vesicles isolated by different methods: A comparison study. Bioengineering 2019, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Teng, F.; Fussenegger, M. Shedding light on extracellular vesicle biogenesis and bioengineering. Adv. Sci. 2020, 8, 2003505. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Williams, R.L.; Urbé, S. The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell Biol. 2007, 8, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Hope, L.W.; Brasch, M.; Reinhard, C.; Cohen, S.N. TSG101 interaction with HRS mediates endosomal trafficking and receptor down-regulation. Proc. Natl. Acad. Sci. USA 2003, 100, 7626–7631. [Google Scholar] [CrossRef] [Green Version]
- Bilodeau, P.S.; Winistorfer, S.C.; Kearney, W.R.; Robertson, A.D.; Piper, R.C. Vps27-Hse1 and ESCRT-I complexes cooperate to increase efficiency of sorting ubiquitinated proteins at the endosome. J. Cell Biol. 2003, 163, 237–243. [Google Scholar] [CrossRef]
- Teo, H.; Gill, D.J.; Sun, J.; Perisic, O.; Veprintsev, D.B.; Vallis, Y.; Emr, S.D.; Williams, R.L. ESCRT-I core and ESCRT-II GLUE domain structures reveal role for GLUE in linking to ESCRT-I and membranes. Cell 2006, 125, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Wollert, T.; Hurley, J.H. Molecular mechanism of multivesicular body biogenesis by ESCRT complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Saksena, S.; Wahlman, J.; Teis, D.; Johnson, A.E.; Emr, S.D. Functional reconstitution of ESCRT-III assembly and disassembly. Cell 2009, 136, 97–109. [Google Scholar] [CrossRef] [Green Version]
- Wollert, T.; Wunder, C.; Lippincott-Schwartz, J.; Hurley, J.H. Membrane scission by the ESCRT-III complex. Nature 2009, 458, 172–177. [Google Scholar] [CrossRef]
- Hurley, J.H.; Hanson, P.I. Membrane budding and scission by the ESCRT machinery: It’s all in the neck. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Larios, J.; Mercier, V.; Roux, A.; Gruenberg, J. ALIX- and ESCRT-III-dependent sorting of tetraspanins to exosomes. J. Cell Biol. 2020, 219, e201904113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eva, R.; Dassie, E.; Caswell, P.T.; Dick, G.; Ffrench-Constant, C.; Norman, J.C.; Fawcett, J.W. Rab11 and its effector Rab coupling protein contribute to the trafficking of beta 1 integrins during axon growth in adult dorsal root ganglion neurons and PC12 cells. J. Neurosci. 2010, 30, 11654–11669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramel, D.; Wang, X.; Laflamme, C.; Montell, D.J.; Emery, G. Rab11 regulates cell-cell communication during collective cell movements. Nat. Cell Biol. 2013, 15, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Prekeris, R.; Gould, G.W. Role of endosomal Rab GTPases in cytokinesis. Eur. J. Cell Biol. 2007, 86, 25–35. [Google Scholar] [CrossRef]
- Ostrowski, M.; Carmo, N.B.; Krumeich, S.; Fanget, I.; Raposo, G.; Savina, A.; Moita, C.F.; Schauer, K.; Hume, A.N.; Freitas, R.P.; et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 2010, 12, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Bebelman, M.P.; Smit, M.J.; Pegtel, D.M.; Baglio, S.R. Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 2018, 188, 1–11. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, J.F.; Hu, R.; Oh, R.S.; Cohen, S.N.; Lu, Q. Formation and release of arrestin domain-containing protein 1-mediated microvesicles (ARMMs) at plasma membrane by recruitment of TSG101 protein. Proc. Natl. Acad. Sci. USA 2012, 109, 4146–4151. [Google Scholar] [CrossRef] [Green Version]
- Chiaruttini, N.; Redondo-Morata, L.; Colom, A.; Humbert, F.; Lenz, M.; Scheuring, S.; Roux, A. Relaxation of loaded ESCRT-III spiral springs drives membrane deformation. Cell 2015, 163, 866–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan-Chari, V.; Clancy, J.; Plou, C.; Romao, M.; Chavrier, P.; Raposo, G.; D’Souza-Schorey, C. ARF6-regulated shedding of tumor cell-derived plasma membrane microvesicles. Curr. Biol. 2009, 19, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbar, N.; Azzimato, V.; Choudhury, R.P.; Aouadi, M. Extracellular vesicles in metabolic disease. Diabetologia 2019, 62, 2179–2187. [Google Scholar] [CrossRef] [Green Version]
- Mir, B.; Goettsch, C. Extracellular vesicles as delivery vehicles of specific cellular cargo. Cells 2020, 9, 1601. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhu, H.; Wang, H. Extracellular vesicles in non-alcoholic fatty liver disease and alcoholic liver disease. Front. Physiol. 2021, 12, 707429. [Google Scholar] [CrossRef]
- Stahl, P.D.; Raposo, G. Extracellular vesicles: Exosomes and microvesicles, integrators of homeostasis. Physiology 2019, 34, 169–177. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Lee, E.G.; Danning, C.L.; Illei, G.G.; McInnes, I.B.; Boumpas, D.T. CD40 ligand-activated human monocytes amplify glomerular inflammatory responses through soluble and cell-to-cell contact-dependent mechanisms. J. Immunol. 1999, 163, 2168–2175. [Google Scholar]
- Harvey, S.; Martínez-Moreno, C.G.; Luna, M.; Arámburo, C. Autocrine/paracrine roles of extrapituitary growth hormone and prolactin in health and disease: An overview. Gen. Comp. Endocrinol. 2015, 220, 103–111. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Hirsova, P.; Ibrahim, S.H.; Verma, V.K.; Morton, L.A.; Shah, V.H.; LaRusso, N.F.; Gores, G.J.; Malhi, H. Extracellular vesicles in liver pathobiology: Small particles with big impact. Hepatology 2016, 64, 2219–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.B.; Bell, C.R.; Bibb, K.E.; Gu, L.; Coats, M.T.; Matthews, Q.L. Pathogens and their effect on exosome biogenesis and composition. Biomedicines 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostallari, E.; Valainathan, S.; Biquard, L.; Shah, V.H.; Rautou, P.E. Role of extracellular vesicles in liver diseases and their therapeutic potential. Adv. Drug Deliv. Rev. 2021, 175, 113816. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Momen-Heravi, F. Extracellular vesicles in liver disease and potential as biomarkers and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Matteoni, C.A.; Younossi, Z.M.; Gramlich, T.; Boparai, N.; Liu, Y.C.; McCullough, A.J. Nonalcoholic fatty liver disease: A spectrum of clinical and pathological severity. Gastroenterology 1999, 116, 1413–1419. [Google Scholar] [CrossRef]
- Zámbó, V.; Simon-Szabó, L.; Szelényi, P.; Kereszturi, E.; Bánhegyi, G.; Csala, M. Lipotoxicity in the liver. World J. Hepatol. 2013, 5, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Gambino, R.; Cassader, M.; Pagano, G. Meta-analysis: Natural history of non-alcoholic fatty liver disease (NAFLD) and diagnostic accuracy of non-invasive tests for liver disease severity. Ann. Med. 2011, 43, 617–649. [Google Scholar] [CrossRef]
- Srinivas, A.N.; Suresh, D.; Santhekadur, P.K.; Suvarna, D.; Kumar, D.P. Extracellular vesicles as inflammatory drivers in NAFLD. Front. Immunol. 2020, 11, 627424. [Google Scholar] [CrossRef]
- Hirsova, P.; Ibrahim, S.H.; Krishnan, A.; Verma, V.K.; Bronk, S.F.; Werneburg, N.W.; Charlton, M.R.; Shah, V.H.; Malhi, H.; Gores, G.J. Lipid-induced signaling causes release of inflammatory extracellular vesicles from hepatocytes. Gastroenterology 2016, 150, 956–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Tan, J.; Wang, M.; Wang, Y.; Dong, M.; Ma, X.; Sun, B.; Liu, S.; Zhao, Z.; Chen, L.; et al. Lipid-induced DRAM recruits STOM to lysosomes and induces LMP to promote exosome release from hepatocytes in NAFLD. Sci. Adv. 2021, 7, eabh1541. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.H.; Hirsova, P.; Tomita, K.; Bronk, S.F.; Werneburg, N.W.; Harrison, S.A.; Goodfellow, V.S.; Malhi, H.; Gores, G.J. Mixed lineage kinase 3 mediates release of C-X-C motif ligand 10-bearing chemotactic extracellular vesicles from lipotoxic hepatocytes. Hepatology 2016, 63, 731–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.H.; Gores, G.J.; Hirsova, P.; Kirby, M.; Miles, L.; Jaeschke, A.; Kohli, R. Mixed lineage kinase 3 deficient mice are protected against the high fat high carbohydrate diet-induced steatohepatitis. Liver Int. 2014, 34, 427–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakazu, E.; Mauer, A.S.; Yin, M.; Malhi, H. Hepatocytes release ceramide-enriched pro-inflammatory extracellular vesicles in an IRE1α-dependent manner. J. Lipid Res. 2016, 57, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, D.; Nakao, Y.; Mauer, A.S.; Thompson, J.M.; Sehrawat, T.S.; Liao, C.Y.; Krishnan, A.; Lucien, F.; Guo, Q.; Liu, M.; et al. IRE1A Stimulates hepatocyte-derived extracellular vesicles that promote inflammation in mice with steatohepatitis. Gastroenterology 2020, 159, 1487–1503.e17. [Google Scholar] [CrossRef]
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, M.J.; Coffman, R.L.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J. Clin. Investig. 2016, 126, 859–864. [Google Scholar] [CrossRef] [Green Version]
- Povero, D.; Eguchi, A.; Li, H.; Johnson, C.D.; Papouchado, B.G.; Wree, A.; Messer, K.; Feldstein, A.E. Circulating extracellular vesicles with specific proteome and liver microRNAs are potential biomarkers for liver injury in experimental fatty liver disease. PLoS ONE 2014, 9, e113651. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Furuta, K.; Lucien, F.; Gutierrez Sanchez, L.H.; Hirsova, P.; Krishnan, A.; Kabashima, A.; Pavelko, K.D.; Madden, B.; Alhuwaish, H.; et al. Integrin β(1)-enriched extracellular vesicles mediate monocyte adhesion and promote liver inflammation in murine NASH. J. Hepatol. 2019, 71, 1193–1205. [Google Scholar] [CrossRef]
- Povero, D.; Eguchi, A.; Niesman, I.R.; Andronikou, N.; de Mollerat du Jeu, X.; Mulya, A.; Berk, M.; Lazic, M.; Thapaliya, S.; Parola, M.; et al. Lipid-induced toxicity stimulates hepatocytes to release angiogenic microparticles that require Vanin-1 for uptake by endothelial cells. Sci. Signal. 2013, 6, ra88. [Google Scholar] [CrossRef] [Green Version]
- Povero, D.; Yamashita, H.; Ren, W.; Subramanian, M.G.; Myers, R.P.; Eguchi, A.; Simonetto, D.A.; Goodman, Z.D.; Harrison, S.A.; Sanyal, A.J.; et al. Characterization and proteome of circulating extracellular vesicles as potential biomarkers for NASH. Hepatol. Commun. 2020, 4, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Kranendonk, M.E.; Visseren, F.L.; van Herwaarden, J.A.; Nolte-‘t Hoen, E.N.; de Jager, W.; Wauben, M.H.; Kalkhoven, E. Effect of extracellular vesicles of human adipose tissue on insulin signaling in liver and muscle cells. Obesity 2014, 22, 2216–2223. [Google Scholar] [CrossRef] [PubMed]
- Koeck, E.S.; Iordanskaia, T.; Sevilla, S.; Ferrante, S.C.; Hubal, M.J.; Freishtat, R.J.; Nadler, E.P. Adipocyte exosomes induce transforming growth factor beta pathway dysregulation in hepatocytes: A novel paradigm for obesity-related liver disease. J. Surg. Res. 2014, 192, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological aspects of alcohol metabolism in the liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Kemper, S.; Brigstock, D.R. Pathways of production and delivery of hepatocyte exosomes. J. Cell Commun. Signal. 2018, 12, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Babuta, M.; Furi, I.; Bala, S.; Bukong, T.N.; Lowe, P.; Catalano, D.; Calenda, C.; Kodys, K.; Szabo, G. Dysregulated autophagy and lysosome function are linked to exosome production by micro-RNA 155 in alcoholic liver disease. Hepatology 2019, 70, 2123–2141. [Google Scholar] [CrossRef]
- Sehrawat, T.S.; Arab, J.P.; Liu, M.; Amrollahi, P.; Wan, M.; Fan, J.; Nakao, Y.; Pose, E.; Navarro-Corcuera, A.; Dasgupta, D.; et al. Circulating extracellular vesicles carrying sphingolipid cargo for the diagnosis and dynamic risk profiling of alcoholic hepatitis. Hepatology 2021, 73, 571–585. [Google Scholar] [CrossRef]
- Momen-Heravi, F.; Saha, B.; Kodys, K.; Catalano, D.; Satishchandran, A.; Szabo, G. Increased number of circulating exosomes and their microRNA cargos are potential novel biomarkers in alcoholic hepatitis. J. Transl. Med. 2015, 13, 261. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.K.; Li, H.; Wang, R.; Hirsova, P.; Mushref, M.; Liu, Y.; Cao, S.; Contreras, P.C.; Malhi, H.; Kamath, P.S.; et al. Alcohol stimulates macrophage activation through caspase-dependent hepatocyte derived release of CD40L containing extracellular vesicles. J. Hepatol. 2016, 64, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.E.; Mezey, E.; Hardwick, J.P.; Salem, N., Jr.; Clemens, D.L.; Song, B.J. Increased ethanol-inducible cytochrome P450-2E1 and cytochrome P450 isoforms in exosomes of alcohol-exposed rodents and patients with alcoholism through oxidative and endoplasmic reticulum stress. Hepatol. Commun. 2017, 1, 675–690. [Google Scholar] [CrossRef]
- Eguchi, A.; Yan, R.; Pan, S.Q.; Wu, R.; Kim, J.; Chen, Y.; Ansong, C.; Smith, R.D.; Tempaku, M.; Ohno-Machado, L.; et al. Comprehensive characterization of hepatocyte-derived extracellular vesicles identifies direct miRNA-based regulation of hepatic stellate cells and DAMP-based hepatic macrophage IL-1β and IL-17 upregulation in alcoholic hepatitis mice. J. Mol. Med. 2020, 98, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Zhang, J.; Wang, M.; Wang, Y.; Dong, M.; Ma, X.; Sun, B.; Liu, S.; Zhao, Z.; Chen, L.; et al. DRAM1 increases the secretion of PKM2-enriched EVs from hepatocytes to promote macrophage activation and disease progression in ALD. Mol. Ther.-Nucleic Acids 2022, 27, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cao, H.; Rodrigues, R.M.; Xu, M.; Ren, T.; He, Y.; Hwang, S.; Feng, D.; Ren, R.; Yang, P.; et al. Chronic-plus-binge alcohol intake induces production of proinflammatory mtDNA-enriched extracellular vesicles and steatohepatitis via ASK1/p38MAPKα-dependent mechanisms. JCI Insight 2020, 5, e136496. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xu, M.J.; Koritzinsky, E.H.; Zhou, Z.; Wang, W.; Cao, H.; Yuen, P.S.; Ross, R.A.; Star, R.A.; Liangpunsakul, S.; et al. Mitochondrial DNA-enriched microparticles promote acute-on-chronic alcoholic neutrophilia and hepatotoxicity. JCI Insight 2017, 2, e92634. [Google Scholar] [CrossRef]
- Lee, J.H.; Shim, Y.R.; Seo, W.; Kim, M.H.; Choi, W.M.; Kim, H.H.; Kim, Y.E.; Yang, K.; Ryu, T.; Jeong, J.M.; et al. Mitochondrial double-stranded RNA in exosome promotes interleukin-17 production through toll-like receptor 3 in alcohol-associated liver injury. Hepatology 2020, 72, 609–625. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Kemper, S.; Charrier, A.; Brigstock, D.R. Suppression of fibrogenic signaling in hepatic stellate cells by Twist1-dependent microRNA-214 expression: Role of exosomes in horizontal transfer of Twist1. Am. J. Physiol.-Gastrointest. Liver Physiol. 2015, 309, G491–G499. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Xia, T.; Du, Y.; Liu, J.; Xie, Y.; Zhang, Y.; Guan, F.; Wu, J.; Wang, X.; Shi, C. Exosomes from activated hepatic stellate cells contain GLUT1 and PKM2: A role for exosomes in metabolic switch of liver nonparenchymal cells. FASEB J. 2019, 33, 8530–8542. [Google Scholar] [CrossRef]
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular reaction in liver diseases: Pathological mechanisms and translational significances. Hepatology 2019, 69, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Hirschfield, G.M.; Heathcote, E.J.; Gershwin, M.E. Pathogenesis of cholestatic liver disease and therapeutic approaches. Gastroenterology 2010, 139, 1481–1496. [Google Scholar] [CrossRef]
- Santiago, P.; Scheinberg, A.R.; Levy, C. Cholestatic liver diseases: New targets, new therapies. Ther. Adv. Gastroenterol. 2018, 11, 1756284818787400. [Google Scholar] [CrossRef]
- Li, X.; Liu, R.; Huang, Z.; Gurley, E.C.; Wang, X.; Wang, J.; He, H.; Yang, H.; Lai, G.; Zhang, L.; et al. Cholangiocyte-derived exosomal long noncoding RNA H19 promotes cholestatic liver injury in mouse and humans. Hepatology 2018, 68, 599–615. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, X.; Zhu, W.; Wang, Y.; Zhao, D.; Wang, X.; Gurley, E.C.; Liang, G.; Chen, W.; Lai, G.; et al. Cholangiocyte-derived exosomal long noncoding RNA H19 promotes hepatic stellate cell activation and cholestatic liver fibrosis. Hepatology 2019, 70, 1317–1335. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, R.; Wang, Y.; Zhu, W.; Zhao, D.; Wang, X.; Yang, H.; Gurley, E.C.; Chen, W.; Hylemon, P.B.; et al. Cholangiocyte-derived exosomal lncRNA H19 promotes macrophage activation and hepatic inflammation under cholestatic conditions. Cells 2020, 9, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsumi, T.; Guicciardi, M.E.; Azad, A.; Bronk, S.F.; Krishnan, A.; Gores, G.J. Activated cholangiocytes release macrophage-polarizing extracellular vesicles bearing the DAMP S100A11. Am. J. Physiol.-Cell Physiol. 2019, 317, C788–C799. [Google Scholar] [CrossRef]
- Zhou, H.; Yan, Z.H.; Yuan, Y.; Xing, C.; Jiang, N. The role of exosomes in viral hepatitis and its associated liver diseases. Front. Med. 2021, 8, 782485. [Google Scholar] [CrossRef]
- Sonderup, M.W.; Spearman, C.W. Global disparities in hepatitis B elimination-A focus on Africa. Viruses 2022, 14, 82. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Elbedewy, T.A.; El-Serafy, M.; El-Toukhy, N.; Ahmed, W.; Ali El Din, Z. Hepatitis C virus: A global view. World J. Hepatol. 2015, 7, 2676–2680. [Google Scholar] [CrossRef]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the ORF3 protein of hepatitis E virus (HEV) can capture HEV particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef]
- Lim, H.K.; Jeffrey, G.P.; Ramm, G.A.; Soekmadji, C. Pathogenesis of viral hepatitis-induced chronic liver disease: Role of extracellular vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 587628. [Google Scholar] [CrossRef]
- Jiang, W.; Ma, P.; Deng, L.; Liu, Z.; Wang, X.; Liu, X.; Long, G. Hepatitis A virus structural protein pX interacts with ALIX and promotes the secretion of virions and foreign proteins through exosome-like vesicles. J. Extracell. Vesicles 2020, 9, 1716513. [Google Scholar] [CrossRef]
- Kouwaki, T.; Fukushima, Y.; Daito, T.; Sanada, T.; Yamamoto, N.; Mifsud, E.J.; Leong, C.R.; Tsukiyama-Kohara, K.; Kohara, M.; Matsumoto, M.; et al. Extracellular vesicles including exosomes regulate innate immune responses to hepatitis B virus infection. Front. Immunol. 2016, 7, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreux, M.; Garaigorta, U.; Boyd, B.; Décembre, E.; Chung, J.; Whitten-Bauer, C.; Wieland, S.; Chisari, F.V. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012, 12, 558–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Han, Q.; Hou, Z.; Zhang, C.; Tian, Z.; Zhang, J. Exosomes mediate hepatitis B virus (HBV) transmission and NK-cell dysfunction. Cell. Mol. Immunol. 2017, 14, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Mengshol, J.A.; Golden-Mason, L.; Arikawa, T.; Smith, M.; Niki, T.; McWilliams, R.; Randall, J.A.; McMahan, R.; Zimmerman, M.A.; Rangachari, M.; et al. A crucial role for Kupffer cell-derived galectin-9 in regulation of T cell immunity in hepatitis C infection. PLoS ONE 2010, 5, e9504. [Google Scholar] [CrossRef]
- Saha, B.; Kodys, K.; Szabo, G. Hepatitis C virus-induced monocyte differentiation into polarized M2 macrophages promotes stellate cell activation via TGF-β. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 302–316.e8. [Google Scholar] [CrossRef] [Green Version]
- Giugliano, S.; Kriss, M.; Golden-Mason, L.; Dobrinskikh, E.; Stone, A.E.; Soto-Gutierrez, A.; Mitchell, A.; Khetani, S.R.; Yamane, D.; Stoddard, M.; et al. Hepatitis C virus infection induces autocrine interferon signaling by human liver endothelial cells and release of exosomes, which inhibits viral replication. Gastroenterology 2015, 148, 392–402.e13. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, A. Hepatocellular carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, P.; Henshaw, C.; Youlden, D.R.; Clark, P.J.; Aitken, J.F.; Baade, P.D. Global trends in incidence rates of primary adult liver cancers: A systematic review and meta-analysis. Front. Oncol. 2020, 10, 171. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Qin, H.; Poon, T.C.; Sze, S.C.; Ding, X.; Co, N.N.; Ngai, S.M.; Chan, T.F.; Wong, N. Hepatocellular carcinoma-derived exosomes promote motility of immortalized hepatocyte through transfer of oncogenic proteins and RNAs. Carcinogenesis 2015, 36, 1008–1018. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Chen, C.K.; Ho, C.M.; Lee, S.S.; Chang, C.Y.; Chen, K.J.; Jou, Y.S. EIF3C-enhanced exosome secretion promotes angiogenesis and tumorigenesis of human hepatocellular carcinoma. Oncotarget 2018, 9, 13193–13205. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Chen, G.; Lin, X.; Xing, X.; Cai, Z.; Liu, X.; Liu, J. Role of exosomes in hepatocellular carcinoma cell mobility alteration. Oncol. Lett. 2017, 14, 8122–8131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Shen, H.; Zhangyuan, G.; Huang, R.; Zhang, W.; He, Q.; Jin, K.; Zhuo, H.; Zhang, Z.; Wang, J.; et al. 14-3-3ζ delivered by hepatocellular carcinoma-derived exosomes impaired anti-tumor function of tumor-infiltrating T lymphocytes. Cell Death Dis. 2018, 9, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.Y.; Wei, J.X.; Lv, L.H.; Han, Q.F.; Yang, W.B.; Li, G.L.; Wang, P.X.; Wu, S.B.; Duan, J.X.; Zhuo, W.F.; et al. Angiopoietin-2 induces angiogenesis via exosomes in human hepatocellular carcinoma. Cell Commun. Signal. 2020, 18, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef]
- Kogure, T.; Yan, I.K.; Lin, W.L.; Patel, T. Extracellular vesicle-mediated transfer of a novel long noncoding RNA TUC339: A mechanism of intercellular signaling in human hepatocellular cancer. Genes Cancer 2013, 4, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Conigliaro, A.; Costa, V.; Lo Dico, A.; Saieva, L.; Buccheri, S.; Dieli, F.; Manno, M.; Raccosta, S.; Mancone, C.; Tripodi, M.; et al. CD90+ liver cancer cells modulate endothelial cell phenotype through the release of exosomes containing H19 lncRNA. Mol. Cancer 2015, 14, 155. [Google Scholar] [CrossRef]
- Fu, Q.; Zhang, Q.; Lou, Y.; Yang, J.; Nie, G.; Chen, Q.; Chen, Y.; Zhang, J.; Wang, J.; Wei, T.; et al. Primary tumor-derived exosomes facilitate metastasis by regulating adhesion of circulating tumor cells via SMAD3 in liver cancer. Oncogene 2018, 37, 6105–6118. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wang, Y.; Zhang, X.; Feng, M.; Ma, J.; Li, J.; Yang, X.; Fang, F.; Xia, Q.; Zhang, Z.; et al. Exosome-mediated secretion of LOXL4 promotes hepatocellular carcinoma cell invasion and metastasis. Mol. Cancer 2019, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Babuta, M.; Szabo, G. Extracellular vesicles in inflammation: Focus on the microRNA cargo of EVs in modulation of liver diseases. J. Leukoc. Biol. 2022, 111, 75–92. [Google Scholar] [CrossRef]
- Yang, J.; Li, C.; Zhang, L.; Wang, X. Extracellular vesicles as carriers of non-coding RNAs in liver diseases. Front. Pharmacol. 2018, 9, 415. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, Q.; Zhang, J.; Li, C.; Miao, Y.R.; Lei, Q.; Li, Q.; Guo, A.Y. EVmiRNA: A database of miRNA profiling in extracellular vesicles. Nucleic Acids Res. 2019, 47, D89–D93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaikkonen, M.U.; Lam, M.T.; Glass, C.K. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Meng, F.; Glaser, S.; Alpini, G. Exosomes in liver pathology. J. Hepatol. 2016, 65, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Yang, Y.M. Exosomal microRNAs as diagnostic and therapeutic biomarkers in non-malignant liver diseases. Arch. Pharm. Res. 2021, 44, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Newman, L.A.; Useckaite, Z.; Johnson, J.; Sorich, M.J.; Hopkins, A.M.; Rowland, A. Selective isolation of liver-derived extracellular vesicles redefines performance of miRNA biomarkers for non-alcoholic fatty liver disease. Biomedicines 2022, 10, 195. [Google Scholar] [CrossRef]
- Liu, X.L.; Pan, Q.; Cao, H.X.; Xin, F.Z.; Zhao, Z.H.; Yang, R.X.; Zeng, J.; Zhou, H.; Fan, J.G. Lipotoxic hepatocyte-derived exosomal MicroRNA 192-5p activates macrophages through Rictor/Akt/Forkhead box transcription factor O1 signaling in nonalcoholic fatty liver disease. Hepatology 2020, 72, 454–469. [Google Scholar] [CrossRef]
- Long, J.K.; Dai, W.; Zheng, Y.W.; Zhao, S.P. miR-122 promotes hepatic lipogenesis via inhibiting the LKB1/AMPK pathway by targeting Sirt1 in non-alcoholic fatty liver disease. Mol. Med. 2019, 25, 26. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, G.R.; Macaulay, S.L.; Febbraio, M.A.; Kemp, B.E. AMP-activated protein kinase—The fat controller of the energy railroad. Can. J. Physiol. Pharmacol. 2006, 84, 655–665. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhong, L.; Li, P.; He, K.; Qiu, C.; Zhao, L.; Gong, J. Cholesterol impairs hepatocyte lysosomal function causing M1 polarization of macrophages via exosomal miR-122-5p. Exp. Cell Res. 2020, 387, 111738. [Google Scholar] [CrossRef]
- Povero, D.; Panera, N.; Eguchi, A.; Johnson, C.D.; Papouchado, B.G.; de Araujo Horcel, L.; Pinatel, E.M.; Alisi, A.; Nobili, V.; Feldstein, A.E. Lipid-induced hepatocyte-derived extracellular vesicles regulate hepatic stellate cell via microRNAs targeting PPAR-γ. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 646–663.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Luo, S.Z.; Xu, Z.X.; Zhou, C.; Li, Z.H.; Zhou, X.Y.; Xu, M.Y. Lipotoxic hepatocyte-derived exosomal miR-1297 promotes hepatic stellate cell activation through the PTEN signaling pathway in metabolic-associated fatty liver disease. World J. Gastroenterol. 2021, 27, 1419–1434. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xu, Z.X.; Wu, J.C.; Luo, S.Z.; Xu, M.Y. Hepatocyte-derived exosomal miR-27a activateshepatic stellate cells through the inhibitionof PINK1-mediated mitophagy in MAFLD. Mol. Ther. Nucleic Acids 2021, 26, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Xie, X.L.; Wang, M.M.; Yin, J.; Tian, J.M.; Jiang, X.Y.; Zhang, D.; Han, J.; Bai, Y.; Cui, Z.J.; et al. The role of the apoptosis-related protein BCL-B in the regulation of mitophagy in hepatic stellate cells during the regression of liver fibrosis. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, A.; Lazaro, R.G.; Wang, J.; Kim, J.; Povero, D.; Willliams, B.; Ho, S.B.; Stärkel, P.; Schnabl, B.; Ohno-Machado, L.; et al. Extracellular vesicles released by hepatocytes from gastric infusion model of alcoholic liver disease contain a MicroRNA barcode that can be detected in blood. Hepatology 2017, 65, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Momen-Heravi, F.; Kodys, K.; Szabo, G. MicroRNA Cargo of Extracellular vesicles from alcohol-exposed monocytes signals naive monocytes to differentiate into M2 macrophages. J. Biol. Chem. 2016, 291, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Bruneau, J.C.; Kodys, K.; Szabo, G. Alcohol-induced miR-27a regulates differentiation and M2 macrophage polarization of normal human monocytes. J. Immunol. 2015, 194, 3079–3087. [Google Scholar] [CrossRef] [Green Version]
- Momen-Heravi, F.; Bala, S.; Kodys, K.; Szabo, G. Exosomes derived from alcohol-treated hepatocytes horizontally transfer liver specific miRNA-122 and sensitize monocytes to LPS. Sci. Rep. 2015, 5, 9991. [Google Scholar] [CrossRef] [Green Version]
- Brandon-Warner, E.; Feilen, N.A.; Culberson, C.R.; Field, C.O.; deLemos, A.S.; Russo, M.W.; Schrum, L.W. Processing of miR17-92 cluster in hepatic stellate cells promotes hepatic fibrogenesis during alcohol-induced injury. Alcohol. Clin. Exp. Res. 2016, 40, 1430–1442. [Google Scholar] [CrossRef] [Green Version]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef]
- Butz, H.; Rácz, K.; Hunyady, L.; Patócs, A. Crosstalk between TGF-β signaling and the microRNA machinery. Trends Pharmacol. Sci. 2012, 33, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, F.S.Y.; Teixeira, F.M.E.; Sato, M.N.; Oliveira, L. Delivery of microRNAs by extracellular vesicles in viral infections: Could the news be packaged? Cells 2019, 8, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Sun, L.; Mu, T.; Yi, J.; Ma, C.; Xie, H.; Liu, M.; Tang, H. An HBV-encoded miRNA activates innate immunity to restrict HBV replication. J. Mol. Cell Biol. 2020, 12, 263–276. [Google Scholar] [CrossRef]
- Devhare, P.B.; Sasaki, R.; Shrivastava, S.; Di Bisceglie, A.M.; Ray, R.; Ray, R.B. Exosome-mediated intercellular communication between hepatitis C virus-infected hepatocytes and hepatic stellate cells. J. Virol. 2017, 91, e02225-16. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, C.H.; Lee, S.W. Exosomal transmission of MicroRNA from HCV replicating cells stimulates transdifferentiation in hepatic stellate cells. Mol. Ther. Nucleic Acids 2019, 14, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.X.; Zhang, Y.H.; Jin, W.J.; Sun, X.Y.; Qiao, G.F.; Wei, Y.Y.; Sun, L.B.; Zhang, W.H.; Li, N. MicroRNA panels as disease biomarkers distinguishing hepatitis B virus infection caused hepatitis and liver cirrhosis. Sci. Rep. 2015, 5, 15026. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Huang, T.; Ren, T.; Liu, Q.; Zhou, Z.; Ge, L.; Chen, Z.; Liu, J.; Nie, H.; Ma, W.; et al. Identification of blood exosomal miRNA-1246, miRNA-150-5p, miRNA-5787 and miRNA-8069 as sensitive biomarkers for hepatitis B virus infection. Clin. Lab. 2022, 68, 2. [Google Scholar] [CrossRef]
- Sohn, W.; Kim, J.; Kang, S.H.; Yang, S.R.; Cho, J.Y.; Cho, H.C.; Shim, S.G.; Paik, Y.H. Serum exosomal microRNAs as novel biomarkers for hepatocellular carcinoma. Exp. Mol. Med. 2015, 47, e184. [Google Scholar] [CrossRef]
- Li, S.; Yao, J.; Xie, M.; Liu, Y.; Zheng, M. Exosomal miRNAs in hepatocellular carcinoma development and clinical responses. J. Hematol. Oncol. 2018, 11, 54. [Google Scholar] [CrossRef]
- Yang, Y.; Mao, F.; Guo, L.; Shi, J.; Wu, M.; Cheng, S.; Guo, W. Tumor cells derived-extracellular vesicles transfer miR-3129 to promote hepatocellular carcinoma metastasis by targeting TXNIP. Dig. Liver Dis. 2021, 53, 474–485. [Google Scholar] [CrossRef]
- Chen, W.; Huang, L.; Liang, J.; Ye, Y.; He, S.; Niu, J. Hepatocellular carcinoma cells-derived exosomal microRNA-378b enhances hepatocellular carcinoma angiogenesis. Life Sci. 2021, 273, 119184. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Pan, Q.; Zhang, Y. Hepatocellular carcinoma cell-derived extracellular vesicles encapsulated microRNA-584-5p facilitates angiogenesis through PCK1-mediated nuclear factor E2-related factor 2 signaling pathway. Int. J. Biochem. Cell Biol. 2020, 125, 105789. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat. Commun. 2018, 9, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitdumrongthum, S.; Metheetrairut, C.; Charoensawan, V.; Ounjai, P.; Janpipatkul, K.; Panvongsa, W.; Weerachayaphorn, J.; Piyachaturawat, P.; Chairoungdua, A. Dysregulated microRNA expression profiles in cholangiocarcinoma cell-derived exosomes. Life Sci. 2018, 210, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ni, Q.; Zhang, H.; Shi, X.; Li, X. Exosomal microRNA-23a-3p contributes to the progression of cholangiocarcinoma by interaction with Dynamin3. Bioengineered 2022, 13, 6208–6221. [Google Scholar] [CrossRef]
- Sorop, A.; Constantinescu, D.; Cojocaru, F.; Dinischiotu, A.; Cucu, D.; Dima, S.O. Exosomal microRNAs as biomarkers and therapeutic targets for hepatocellular carcinoma. Int. J. Mol. Sci. 2021, 22, 4997. [Google Scholar] [CrossRef]
- Li, C.; Xu, X. Biological functions and clinical applications of exosomal non-coding RNAs in hepatocellular carcinoma. Cell. Mol. Life Sci. 2019, 76, 4203–4219. [Google Scholar] [CrossRef]
- Sugimachi, K.; Matsumura, T.; Hirata, H.; Uchi, R.; Ueda, M.; Ueo, H.; Shinden, Y.; Iguchi, T.; Eguchi, H.; Shirabe, K.; et al. Identification of a bona fide microRNA biomarker in serum exosomes that predicts hepatocellular carcinoma recurrence after liver transplantation. Br. J. Cancer 2015, 112, 532–538. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, D.; Cui, M.; Qu, F.; Yu, J.; Tang, Z.; Wang, L. Exosome marker proteins of tumor-associated fibroblasts and exosome-derived miR-92a-3p act as potential biomarkers for liver cancer. Crit. Rev. Eukaryot. Gene Expr. 2022, 32, 49–57. [Google Scholar] [CrossRef]
- Li, L.; Masica, D.; Ishida, M.; Tomuleasa, C.; Umegaki, S.; Kalloo, A.N.; Georgiades, C.; Singh, V.K.; Khashab, M.; Amateau, S.; et al. Human bile contains microRNA-laden extracellular vesicles that can be used for cholangiocarcinoma diagnosis. Hepatology 2014, 60, 896–907. [Google Scholar] [CrossRef]
- Jiménez-Avalos, J.A.; Fernández-Macías, J.C.; González-Palomo, A.K. Circulating exosomal MicroRNAs: New non-invasive biomarkers of non-communicable disease. Mol. Biol. Rep. 2021, 48, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Nik Mohamed Kamal, N.; Shahidan, W.N.S. Non-exosomal and exosomal circulatory MicroRNAs: Which are more valid as biomarkers? Front. Pharmacol. 2019, 10, 1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devhare, P.B.; Ray, R.B. Extracellular vesicles: Novel mediator for cell to cell communications in liver pathogenesis. Mol. Asp. Med. 2018, 60, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Braasch, D.A.; Corey, D.R. Locked nucleic acid (LNA): Fine-tuning the recognition of DNA and RNA. Chem. Biol. 2001, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Momen-Heravi, F.; Catalano, D.; Talis, A.; Szabo, G.; Bala, S. Protective effect of LNA-anti-miR-132 therapy on liver fibrosis in mice. Mol. Ther. Nucleic Acids 2021, 25, 155–167. [Google Scholar] [CrossRef]
- Charrier, A.; Chen, R.; Chen, L.; Kemper, S.; Hattori, T.; Takigawa, M.; Brigstock, D.R. Exosomes mediate intercellular transfer of pro-fibrogenic connective tissue growth factor (CCN2) between hepatic stellate cells, the principal fibrotic cells in the liver. Surgery 2014, 156, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Tian, X.; Li, J.; Liu, D.; Ye, D.; Xie, Z.; Han, Y.; Zou, M.H. A high-fat diet attenuates AMPK α1 in adipocytes to induce exosome shedding and nonalcoholic fatty liver development in vivo. Diabetes 2021, 70, 577–588. [Google Scholar] [CrossRef]
- Liao, C.Y.; Song, M.J.; Gao, Y.; Mauer, A.S.; Revzin, A.; Malhi, H. Hepatocyte-derived lipotoxic extracellular vesicle sphingosine 1-phosphate induces macrophage chemotaxis. Front. Immunol. 2018, 9, 2980. [Google Scholar] [CrossRef] [Green Version]
- Mauer, A.S.; Hirsova, P.; Maiers, J.L.; Shah, V.H.; Malhi, H. Inhibition of sphingosine 1-phosphate signaling ameliorates murine nonalcoholic steatohepatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G300–G313. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Luan, J.; Lv, X. Inhibition of endoplasmic reticulum stress attenuated ethanol-induced exosomal miR-122 and acute liver injury in mice. Alcohol Alcohol. 2019, 54, 465–471. [Google Scholar] [CrossRef]
- Wang, R.; Ding, Q.; Yaqoob, U.; de Assuncao, T.M.; Verma, V.K.; Hirsova, P.; Cao, S.; Mukhopadhyay, D.; Huebert, R.C.; Shah, V.H. Exosome adherence and internalization by hepatic stellate cells triggers sphingosine 1-phosphate-dependent migration. J. Biol. Chem. 2015, 290, 30684–30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S.Y.; Sood, A.K.; Hua, S. Advances and challenges of liposome assisted drug delivery. Front. Pharmacol. 2015, 6, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrelli, D.A.; Yankson, K.; Shukla, N.; Vilanilam, G.; Ticer, T.; Wolfram, J. Extracellular vesicle therapeutics for liver disease. J. Control. Release 2018, 273, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Sadallah, S.; Eken, C.; Martin, P.J.; Schifferli, J.A. Microparticles (ectosomes) shed by stored human platelets downregulate macrophages and modify the development of dendritic cells. J. Immunol. 2011, 186, 6543–6552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiklander, O.P.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Li, W.N.; Li, X.X.; Yue, K.Y.; Duan, J.L.; Ruan, B.; Liu, J.J.; Song, P.; Yue, Z.S.; Tao, K.S.; et al. Exosome-mediated delivery of RBP-J decoy oligodeoxynucleotides ameliorates hepatic fibrosis in mice. Theranostics 2022, 12, 1816–1828. [Google Scholar] [CrossRef]
- Momen-Heravi, F.; Bala, S.; Bukong, T.; Szabo, G. Exosome-mediated delivery of functionally active miRNA-155 inhibitor to macrophages. Nanomedicine 2014, 10, 1517–1527. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Huang, X.; Xiu, H.; Sun, Y.; Chen, J.; Cheng, G.; Song, Z.; Peng, Y.; Shen, Y.; Wang, J.; et al. Extracellular vesicles: Natural liver-accumulating drug delivery vehicles for the treatment of liver diseases. J. Extracell. Vesicles 2020, 10, e12030. [Google Scholar] [CrossRef]
- Ji, K.; Fan, M.; Huang, D.; Sun, L.; Li, B.; Xu, R.; Zhang, J.; Shao, X.; Chen, Y. Clodronate-nintedanib-loaded exosome-liposome hybridization enhances the liver fibrosis therapy by inhibiting Kupffer cell activity. Biomater. Sci. 2022, 10, 702–713. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Back, M.J.; Ha, H.C.; Fu, Z.; Choi, J.M.; Piao, Y.; Won, J.H.; Jang, J.M.; Shin, I.C.; Kim, D.K. Activation of neutral sphingomyelinase 2 by starvation induces cell-protective autophagy via an increase in Golgi-localized ceramide. Cell Death Dis. 2018, 9, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalano, M.; O’Driscoll, L. Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. J. Extracell. Vesicles 2020, 9, 1703244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
A schematic description of EVs biogenesis and secretion. The schematic representation depicts the production of broad categories of extracellular vesicles (EVs) derived from living cells (left) and apoptotic cells (right). Exosomes originate from the endosomal system. Early endosomes are generated from the inward budding of the plasma membrane, and the early endosomal membrane invaginates inward to form intraluminal vesicles (ILVs), maturing into multivesicular bodies (MVBs). In this process, the endosomal sorting complex required for transport (ESCRT) machinery participates in ILV formation. ESCRT-0 recognizes and selects ubiquitinated proteins and recruits ESCRT-I to the endosomal membrane where ESCRT-I forms ESCRT-I/II complex. ESCRT-III, cutting the neck of the bud, is added to the ESCRT-I/II complex, and forms ILVs in MVB lumen. Next, Rab27a/b mediates the docking and fusion of MVBs with plasma membranes to secrete ILVs to the extracellular space, and the released ILVs are now termed exosomes. Alternatively, early endosomes can be recycled back to the plasma membrane by Rab11 and Rab35. Microvesicles are formed by the direct budding and pinching outward from the plasma membrane. Although the molecular mechanism explaining microvesicle formation is not fully elucidated, the ESCRT machinery and small GTPases are related to their biogenesis and release. Cytoskeletal remodeling promotes microvesicles protruding through ADP-ribosylation factor 6 (ARF6)-ERK-myosin light-chain kinase (MLCK) axis. Tumor susceptibility gene 101 protein (TSG101), one of the ESCRT-1 components, and ESCRT-III proteins also mediate microvesicle release. Both exosomes and microvesicles contain various cargoes such as proteins and genetic materials, DNA, mRNA, long noncoding RNA (lncRNA), and microRNA (miRNA), reflecting the microenvironment of donor cells. On the other hand, apoptotic bodies are released by membrane blebbing from cells undergoing apoptosis. Apoptotic bodies carry a variety of cellular components such as chromatin remnants, degraded proteins and DNA fragments. This figure was created with BioRender.com.
Figure 1.
A schematic description of EVs biogenesis and secretion. The schematic representation depicts the production of broad categories of extracellular vesicles (EVs) derived from living cells (left) and apoptotic cells (right). Exosomes originate from the endosomal system. Early endosomes are generated from the inward budding of the plasma membrane, and the early endosomal membrane invaginates inward to form intraluminal vesicles (ILVs), maturing into multivesicular bodies (MVBs). In this process, the endosomal sorting complex required for transport (ESCRT) machinery participates in ILV formation. ESCRT-0 recognizes and selects ubiquitinated proteins and recruits ESCRT-I to the endosomal membrane where ESCRT-I forms ESCRT-I/II complex. ESCRT-III, cutting the neck of the bud, is added to the ESCRT-I/II complex, and forms ILVs in MVB lumen. Next, Rab27a/b mediates the docking and fusion of MVBs with plasma membranes to secrete ILVs to the extracellular space, and the released ILVs are now termed exosomes. Alternatively, early endosomes can be recycled back to the plasma membrane by Rab11 and Rab35. Microvesicles are formed by the direct budding and pinching outward from the plasma membrane. Although the molecular mechanism explaining microvesicle formation is not fully elucidated, the ESCRT machinery and small GTPases are related to their biogenesis and release. Cytoskeletal remodeling promotes microvesicles protruding through ADP-ribosylation factor 6 (ARF6)-ERK-myosin light-chain kinase (MLCK) axis. Tumor susceptibility gene 101 protein (TSG101), one of the ESCRT-1 components, and ESCRT-III proteins also mediate microvesicle release. Both exosomes and microvesicles contain various cargoes such as proteins and genetic materials, DNA, mRNA, long noncoding RNA (lncRNA), and microRNA (miRNA), reflecting the microenvironment of donor cells. On the other hand, apoptotic bodies are released by membrane blebbing from cells undergoing apoptosis. Apoptotic bodies carry a variety of cellular components such as chromatin remnants, degraded proteins and DNA fragments. This figure was created with BioRender.com.

Table 1.
Summary of characteristics of apoptotic body, exosome and microvesicle.
| Apoptotic Body | Exosome | Microvesicle | |
|---|---|---|---|
| Size | 1–4 μm | 30–100 nm | 100–1000 nm |
| Biogenesis | Programmed cell death | Inward protrusion of early endosomal membrane | Budding and pinching outward directly from the plasma membrane |
| Contents | Chromatin remnants, Degraded proteins, DNA fragments, Intact organelles | Proteins, Metabolites, DNA, mRNA, miRNA, mtDNA, lncRNA | |
| Biological function | Simulating the immune system | Shuttling bioactive substances and genetic materials Reflecting characteristics of donor cells Acting as messengers of cell-to-cell communication | |
DNA, deoxyribonucleic acid; mRNA, messenger RNA; miRNA, microRNA; mtDNA, mitochondrial DNA; lncRNA, long noncoding RNA.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, C.; Han, J.; Jung, Y. Pathological Contribution of Extracellular Vesicles and Their MicroRNAs to Progression of Chronic Liver Disease. Biology 2022, 11, 637. https://doi.org/10.3390/biology11050637
AMA Style
Lee C, Han J, Jung Y. Pathological Contribution of Extracellular Vesicles and Their MicroRNAs to Progression of Chronic Liver Disease. Biology. 2022; 11(5):637. https://doi.org/10.3390/biology11050637
Chicago/Turabian StyleLee, Chanbin, Jinsol Han, and Youngmi Jung. 2022. "Pathological Contribution of Extracellular Vesicles and Their MicroRNAs to Progression of Chronic Liver Disease" Biology 11, no. 5: 637. https://doi.org/10.3390/biology11050637
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.