Regulation of Cell Death by Mitochondrial Transport Systems of Calcium and Bcl-2 Proteins


J. Heyrovský Institute of Physical Chemistry, Czech Academy of Sciences, 182 23 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Membranes 2020, 10(10), 299; https://doi.org/10.3390/membranes10100299
Submission received: 27 August 2020
/
Revised: 16 October 2020
/
Accepted: 19 October 2020
/
Published: 21 October 2020
(This article belongs to the Special Issue Membrane Channels and Transporters)
Abstract
:Mitochondria represent the fundamental system for cellular energy metabolism, by not only supplying energy in the form of ATP, but also by affecting physiology and cell death via the regulation of calcium homeostasis and the activity of Bcl-2 proteins. A lot of research has recently been devoted to understanding the interplay between Bcl-2 proteins, the regulation of these interactions within the cell, and how these interactions lead to the changes in calcium homeostasis. However, the role of Bcl-2 proteins in the mediation of mitochondrial calcium homeostasis, and therefore the induction of cell death pathways, remain underestimated and are still not well understood. In this review, we first summarize our knowledge about calcium transport systems in mitochondria, which, when miss-regulated, can induce necrosis. We continue by reviewing and analyzing the functions of Bcl-2 proteins in apoptosis. Finally, we link these two regulatory mechanisms together, exploring the interactions between the mitochondrial Ca2+ transport systems and Bcl-2 proteins, both capable of inducing cell death, with the potential to determine the cell death pathway—either the apoptotic or the necrotic one.
Keywords:
mitochondria; calcium transport; Bcl-2 proteins; apoptosis; necrosis; Bax; MCU; VDAC; mPTP
1. Introduction
Calcium (Ca2+) is a divalent cation and a universal second messenger that regulates the most important functions and facets of all eukaryotic cells, including gene expression, proliferation, regulation of bioenergetics, contraction of muscles, mediation of fertilization, and many other cellular functions [1,2,3,4,5]. Regulation of free intracellular concentration of Ca2+ is an important mechanism for intracellular signaling, and it is a key component in the mediation of many cell functions and biochemical reactions, being crucial for signal transduction in cells [2,6,7,8,9,10]. On top of all that, intra-mitochondrial Ca2+ regulates a cascade of physiological and pathophysiological processes in cells [10,11,12,13,14,15].
The organelles responsible for Ca2+ homeostasis are undoubtedly the mitochondria, which are essential for cellular bioenergetics by storing energy in the form of ATP and by playing a major role in Ca2+ signaling [5,12,16,17]. Ca2+ uptake by mitochondria not only participates in the regulation of cytosolic Ca2+ concentration ([Ca2+]), but also stimulates mitochondrial respiration and ATP production [18,19]. These properties make these organelles the major cellular components in the regulation of the fate of a cell [9,12,17,20,21].
Localization of mitochondria inside the cell can vary significantly: from the periphery of the cell, around the nucleus, but also close to the plasma membrane or the endoplasmic/sarcoplasmic reticulum (ER/SR) [22,23]. These different localizations determine the Ca2+-buffering capacity of each individual mitochondria, as well as the mitochondrial network [12]. Upon contact of the mitochondria, or more specifically the outer mitochondrial membrane (OMM) with other organelles, membrane contact sites are formed [24]. These inter-organelle associations have various functions. For instance, those formed between the mitochondria and ER/SR (mitochondria-associated membranes, MAMs) determine Ca2+-uptake from the cytoplasm to the mitochondria, and therefore play an essential role in the Ca2+ signaling pathways [23,25,26]. It is established that such associations contain microdomains with high Ca2+ concentrations that determine the mediation of Ca2+ transport between the mitochondria and the ER/SR [24]. Moreover, mitochondrial associations with the plasma membrane are engaged in the mediation of Ca2+ transport from the extracellular environment [27].
Mitochondria are the power generators of cells. They produce ATP in the citric acid cycle (the tricarboxylic acid (TCA) or the Krebs cycle (see Box 1 for more information). Production of ATP involves activation of the Ca2+-dependent dehydrogenases in the citric acid cycle, F0F1-ATP-synthase and metabolite transporters; all of them being supplied by basal oscillating increases in the concentration of Ca2+ in the mitochondrial matrix [6,28,29,30]. In addition to these normal physiological oscillations, large Ca2+ spikes in mitochondria can cause an opening of the mitochondrial permeability transition pore (mPTP) [9,28,29]. In turn, this induces a collapse of the mitochondrial membrane potential, termination of oxidative phosphorylation processes, osmotic changes, mitochondrial swelling, and inner membrane remodeling. All of these processes culminate by mitochondrial outer membrane permeabilization (MOMP) and the release of cytochrome c; being both an inducer of apoptosis and modulator of other proapoptotic factors [2,12,31,32]. Whereas mostly associated with programmed cell death, a number of compounds trigger changes in Ca2+ homeostasis and mPTP-induced apoptosis [2,9,29].
Box 1. The generation of ATP by mitochondria.
Mitochondria are the power generators within all eukaryotic cells. They release their energy in the form of ATP by the oxidation of
sugars. Electrons supplied by NADH are transferred to oxygen by a series of protein complexes in the inner mitochondrial membrane. By pumping protons across the membrane, these complexes create a transmembrane electrochemical gradient (ΔΨ, ~−180 mV). This reverse current of protons into the mitochondrial matrix occurs through a proton channel formed by ATP synthase, and it is used to store energy in the form of ATP.
The ability of mitochondria to uptake and retain Ca2+ had already been described in the early 1960s using isolated mitochondria [33,34]. During the same years, the chemiosmotic theory, as proposed by Mitchell [35], revealed the thermodynamic basis of the process.
Mitochondria are able to rapidly accumulate and transiently store Ca2+ for later quick release, making these organelles important cytosolic depots or buffers for Ca2+ regarding mediation of the cell’s physiological and pathological processes, including from cell survival to cell death [3,7,9,12,21,36,37]. Regulated elevations of Ca2+ levels in the mitochondrial matrix are necessary for the regulation of Ca2+-dependent mitochondrial enzyme activity, which sequentially mediates the metabolic balance and function of the mitochondrial electron transport chain, as well as the production of mitochondria-generated reactive oxygen species (ROS) [12,38,39,40]. Undoubtedly, the precise regulation of mitochondrial Ca2+ uptake and release is necessary for proper cellular functioning and regulation of mitochondrial bioenergetics. The normal level of intra-mitochondrial Ca2+ is essential for the correct functioning of mitochondria, whereas Ca2+ overload is typical for a wide range of mitochondrial dysfunctions and pathophysiological processes [14,15,37,39,41]. Homeostasis of Ca2+ in the mitochondria is determined by the delicate balance of mitochondrial Ca2+ transport systems in both the inner (IMM) and outer mitochondrial membrane (OMM) (Figure 1). Ca2+ influx and efflux systems are composed of different components, including: channels, pumps, antiporters, or Ca2+ binding proteins that cooperate to maintain intra-mitochondrial Ca2+ homeostasis [10,14,38,39,42].
2. Calcium Transport Systems in Mitochondria
2.1. Calcium Influx and Efflux through OMM
When Ca2+ enters the mitochondrial matrix from the cytoplasm, it first encounters the OMM. This membrane is highly permeable to cations, anions, and molecules with molecular weights < 5 kDa due to the presence of large conductance channels. These channels, formed by voltage-dependent anion channel proteins (VDACs), allow for the exchange of Ca2+ and small molecules by concentration gradients [2,43,44,45,46,47]. They not only regulate transport of Ca2+ from the cytoplasm into the intermembrane space (IMS), but are additionally engaged in the mediation of cellular metabolism by transporting ATP and other small metabolites across the OMM [12,48]. Importantly, the permeability of VDACs is precisely controlled and regulated, particularly by ATP and a variety of cellular regulatory factors.
VDAC (Figure 1) was the first channel that has been reconstituted and characterized in detail at the single-channel level [44,49]. It has been proposed to work as the principal metabolite transport system across the OMM, and had also been proposed to serve as the interconnection point between the OMM and IMM [50]. Later, three different VDAC isoforms were identified: VDAC1, VDAC2, and VDAC3 [2,44,47,51].
VDAC1 is highly expressed in most cells, and seems to be the most prevalent and most extensively characterized; it is also considered as the main transport channel for Ca2+ [2,52]. VDAC1 is the gatekeeper for the passage of ions and metabolites, and is crucial for the regulation of apoptosis, thanks to its interactions with pro- and anti-apoptotic proteins [23,53]. Activity of VDAC1 is critical for the mitochondrial metabolic pathways balance, as well as for cell survival [53]. Imaging of VDAC1 by stimulated emission depletion nanoscopy revealed the organization of VDAC proteins into clusters in H9C2 cells, which has also been studied in VDAC transfected U2OS cells [54,55]. VDAC1 consists of 19 transmembrane β-strands that are organized into the membrane-incorporated β-barrel and a amphipathic 26-residue-long N-terminal domain, which can translocate from the pore interior to the channel surface [56]. This behavior is crucial for controlling the gating of the channel as well as its interactions with apoptotic proteins [56,57]. Whereas isoforms of VDAC1 and VDAC2 self-assemble into structures resembling a pore, VDAC3 forms smaller conductance channels that are able to modulate the physiological functions of various proteins [58]. As demonstrated by Checchetto et al. [58], VDAC3 isoforms demonstrate different electrophysiological properties compared with those of VDAC1 and VDAC2. In the context of their structural/functional characteristics, VDAC1, VDAC2, and VDAC3 have some similarities; at the same time, they exhibit different physiological functions regarding their interaction with cytosolic proteins and other mitochondrial proteins [2,47,57,59]. Furthermore, only limited information is available regarding the potential functions of VDAC2 and VDAC3 for the influx of Ca2+ [43,59,60].
2.2. Calcium Influx through IMM
Compared to the OMM, the IMM exhibits a fundamentally higher selectivity for anions and cations thanks to the presence of highly-specific and different protein machinery in the IMM. The key transporters that determine Ca2+ uptake by mitochondria through the IMM until recently were unclear. It is now believed that the transport of Ca2+ through the IMM is accomplished by a group of mitochondrial Ca2+ uptake transporters. Basically, three main mechanisms of Ca2+ influx have been proposed (Figure 1): (1) a mechanism that requires an electrogenic mitochondrial Ca2+ uniporter multi-protein complex (MCU complex); (2) a so-called rapid mode (RaM); (3) a mechanism requiring the mitochondrial ryanodine receptor (mRyR) [7,10,12,13,38]; and (4) additionally, leucine zipper-EF-hand containing transmembrane protein (LETM1) could represent another Ca2+ influx mechanism, but its role is still under discussion [61,62,63,64].
2.2.1. Calcium Influx by Mitochondrial Ca2+ Uniporter (MCU) Multi-Protein Complex
The molecular identity of this Ca2+ transport pathway had been unclear for several decades. However, in 2011, the CCDC109a gene, a pore-forming component of the MCU channel, mediating Ca2+ influx into mitochondria was discovered [65,66]. The protein encoded by the CCDC109a gene is responsible for Ruthenium Red-sensitive mitochondrial Ca2+ uptake. Currently, accumulation of Ca2+ through the MCU multi-protein complex is the most widely characterized and commonly accepted pathway of Ca2+ influx into mitochondria; and it is considered as the major pathway of the mitochondrial Ca2+ influx. It is determined by a large electrochemical gradient (~−180 mV) across the IMM, and may be inhibited by Ruthenium Red and lanthanides [7,13,42,67,68,69]. The complex consists of several subunits, including transmembrane core components and regulatory subunits that are associated with the membrane. The core components of the MCU multi-protein complex (see Box 2 for details) are comprised of: (a) core protein components: Mitochondrial Ca2+ Uniporter (MCU), a MCU dominant negative beta subunit (MCUb), and Essential MCU REgulator (EMRE); and (b) membrane associated regulatory components: mitochondrial Ca2+ uptake protein 1–3 (MICU1–3) and Mitochondrial Ca2+ Uniporter Regulator 1 (MCUR1) [12,13,23,42,47,68,69,70,71,72]. Solute Carrier 25A23 (SLC25A23)) was initially identified as an essential component of MCU, however, it is currently under debate whether SLC25A23 is an component of MCU or whether it influences MCU indirectly [13,42,73]. Importantly, the MCU complex can be found in multiple states.
Box 2. Structure of the MCU multi-protein complex.
Core components
MCU (mitochondrial Ca2+ uniporter, previously known as CCDC109a, 40 kDa) is a key core component of the complex. It is encoded by a highly conservative MCU gene and is present in virtually all eukaryotic organisms [10,13,47]. MCU can be found in multiple states, and it consists of two coiled-coil domains (CC) and two transmembrane domains connected via a short loop (9 amino acid residues) containing a highly conserved DIME motif [42,65,66].
MCUb (MCU dominant negative beta subunit, formerly known as CCDC109b, 40 kDa) is a core component of the MCU multi-protein complex encoded by the MCUb gene, and is present in all vertebrates [71,72,74]. It exhibits a 50% homology with MCU; however, MCU and MCUb demonstrate diverse expression profiles in different tissues. Importantly, MCUb significantly impairs Ca2+ permeation through MCU [42,69,70].
EMRE (essential MCU regulatory element, 10–12 kDa) is the last core component identified in the complex. It contains a single transmembrane segment, and crucially regulates MCU activity as has been shown using EMRE knockout cells, which inhibited mitochondrial Ca2+ influx [75]. EMRE is assumed to be involved in the formation of interactions between the core and the regulatory subunits, despite the fact that such ensembles of regulatory components do not require the presence of EMRE [13,42,47,75].
Membrane-associated regulatory components
MICU1 (mitochondrial Ca2+ uptake protein 1, known as CBARA1/EFHA3, 54kDa) known as CBARA1/EFHA3, is a membrane-associated and water-soluble component localized in the inter-membrane space; it is considered as central for the activation of MCU. In the resting state (i.e., at low intracellular concentrations of Ca2+), MICU1 blocks access of Ca2+ to the MCU channel [75,76,77,78]. It also acts as a cooperative activator of MCU and it stimulates MCU Ca2+-transport conductivity [76].
MICU2 (mitochondrial Ca2+ uptake protein 2, known as EFHA1, 50 kDa) and MICU3 (mitochondrial Ca2+ uptake protein 3, known as EFHA2, 60 kDa) display the EF-hand domains in the protein structure, and were identified as MiCU1 paralogs with 41% and 34% identity to the MICU1, respectively [78,79,80]. MiCU2 forms heterodimers with MiCU1 through disulfide bonds, and acts as a Ca2+ sensor, protecting the mitochondria against Ca2+ overload, and it also acts as the regulator of several cell functions [76,81,82].
MCUR1 (mitochondrial Ca2+ uniporter regulator 1, known as CCDC90A, 40 kDa) is composed of 2 transmembrane domains and 1 specific coiled-coil region, and it belongs to yet another regulatory component of the MCU complex [83,84]. MCUR1 knockdown prevents Ca2+ entry into the mitochondria; whereas, its overexpression promotes mitochondrial Ca2+ uptake [82,84]. MCUR1 interacts with EMRE and MCU-pore via its coiled-coil domains, which stabilize all components of the MCU complex [85]. It is involved in the assembly of the mitochondrial respiratory chain, and represents a cytochrome c oxidase assembly factor; possibly also regulating the mitochondrial membrane potential [86].
SLC25A23 (solute carrier 25A23, 48–54 kDa) was initially identified in the IMM as a protein with the EF-hand domain, and has been proposed as a component of MCU multi-protein complex [2,86,87]. SLC25A23 may also function as an ATP-Mg/Pi exchanger, promoting the influx of adenine nucleotides into the matrix of mitochondria and the efflux of inorganic phosphate. Of note, SLC24A23 functions in a Ca2+ dependent manner [73,88]. Mutations and modifications of the EF-hand domains in this carrier decrease Ca2+ influx into mitochondria; however, it still remains unclear whether SLC25A23 influences the uniporter complex directly or whether it affects the mitochondrial bioenergetics [13,42,87]. Further studies are necessary to understand the exact mechanism by which SLC25A23 regulates mitochondrial Ca2+ influx.
2.2.2. Rapid Mode Mechanism (RaM) of Ca2+ Uptake
The RaM (RApid Mode of Ca2+ uptake) mechanism is able to accumulate Ca2+ up to a hundred times faster compared with the MCU multi-protein complex (no molecular structure responsible for this mechanism has yet been identified) [89,90]. It is transiently activated by low calcium concentrations (50–100 nM) and by high concentrations of Ruthenium Red [13,90,91]. This behavior contrasts sharply with MCU, which is activated by Ca2+ concentrations higher than 500 nM. RaM promotes mitochondria to rapidly sequester Ca2+ at the beginning of each cytosolic Ca2+ pulse, and rapidly recovers between pulses, allowing mitochondria to respond to repetitive Ca2+ oscillations [13,91]. It is still speculated that RaM is just an additional state of the MCU multi-protein complex because of their similarity as well as the absence of RaM in MCU knockout mitochondria [65,90,91]. At present, the progress of research targeted on explaining the role of RaM in Ca2+ influx at the molecular level is very limited.
2.2.3. The Mechanism of Ca2+ Uptake Requiring Mitochondrial Ryanodine Receptor (mRyR)
mRyR (mitochondrial ryanodine receptor, 600 kDa) is the ryanodine-sensitive mitochondrial Ca2+ uptake mechanism, capable of Ca2+ transport, which was detected in the IMM of isolated heart mitochondria in 2001 by Beutner at al. [92]. This group confirmed the presence of the ryanodine receptor in the IMM using [3H]ryanodine binding, RyR antibody conjugated immunogold particles, and Western blot analysis [92]. It could serve as an alternative mechanism for Ca2+ accumulation in mitochondria as well as a regulator of Ca2+ efflux under mitochondrial Ca2+ overload and pathological conditions [92,93,94]. Interestingly, the single channel activity of mRyR was confirmed on recombinant mRyR proteins reconstituted in supported lipid bilayers prepared from IMM vesicles [95]. This study elucidates pharmacological and electrophysiological features of mRyR in the model of IMM merged to lipid bilayers, where a mitochondrial transporter with gating properties similar to those of RyR in ER/SR was demonstrated [95].
2.2.4. The Mechanism of Ca2+ Uptake Including LETM1
LETM1 (leucine zipper- EF-hand containing transmembrane protein, 70 kDa) is an integral mitochondrial inner membrane protein, usually co-localized with a mitochondrial matrix protein HSP60 [62,96]. The N-terminus of this protein is linked to the IMM via a transmembrane domain consisting of 3 proline residues, whereas the C-terminus extends to the mitochondrial matrix [62,63,97]. It was also demonstrated previously that LETM1 is an endogenous protein in HeLa cells, with a molecular weight of 83 kDa, and it has been assumed that it is initially produced as a cytosolic precursor with a presequence [62,96,98,99]. LETM1 is a transporter protein shown to exhibit Ca2+/H⁺ exchange activity, acting as a crucial component in the regulation of Ca2+ homeostasis [62,96,100,101,102,103,104]. Later it was proposed as an inner mitochondrial membrane Ca2+/H+ antiporter [103] that is able to transport Ca2+ bidirectionally across the membrane. In addition, experimental work indicated the important role of LETM1 in maintaining K+ homeostasis, and this has led to the suggestion that LETM1 works as an H+/K+ exchanger with an electroneutral activity (1H+/1K+) [105]. Of note, this exchanger shares a key role with MCU to catalyze Ruthenium Red-sensitive transport of Ca2+ into mitochondria [103]. It would likely serve as an alternative mechanism for Ca2+ accumulation in mitochondria, as well as a regulator of Ca2+ efflux under mitochondrial Ca2+ overload [61,101,106]. In summary, although the importance of LETM1 for cellular functioning is clear, the molecular characteristics and details of LETM1 organization still remain unclear.
2.3. Calcium Efflux through IMM
In order to maintain the intra-mitochondrial Ca2+ homeostasis under physiological and pathological conditions, the balance between Ca2+ influx and efflux into/from mitochondria has to be maintained. The functional and molecular characterization of the mitochondrial Ca2+ efflux system already had started in the 1970s, when Na+-dependent Ca2+ efflux from mitochondria was described in isolated rat heart mitochondria [107], and two different mechanisms were proposed: (1) Na+-dependent (Na+/Ca2+/Li+ exchange, NCLX) and (2) Na+-independent (H+/Ca2+ exchange, HCX) mechanisms. It was reported that the Na+/Ca2+ exchange takes place in excitable tissues (i.e., brain, heart), whereas H+/Ca2+ exchange is typical for non-excitable tissues (i.e., liver). However, both systems provide slow Ca2+ release in comparison to the rate of Ca2+ influx through the MCU [108,109]. Later, (3) the mitochondrial permeability transition pore complex (mPTPC) was identified as an important Ca2+ efflux mechanism [3,110]. Besides this, LETM1 (4) has been proposed as an additional Ca2+ efflux system [12,103,111,112] (Figure 1).
2.3.1. The Mechanism of Ca2+ efflux by NCLX
NCLX (Na+/Ca2+/Li+ exchanger systems): Mitochondrial Na+/Ca2+ (NCX) exchange was discovered by Carafoli et al. in 1974 [107]. However, the molecular composition of the Na+-dependent Ca2+ efflux system was resolved relatively recently [113], and interestingly, seems to function as a transporter of Li+ ions as well, being a member of the family of Na+/Ca2+ exchangers [73,113,114,115]. The ability of NCLX to conduct both Na+/Ca2+ and Li+/Ca2+ transport is a unique feature of the mitochondrial carrier [73,115,116]. In fact, it can transport either Li+ or Na+ in exchange for Ca2+. NCLX is the only known member of the Na+/Ca2+ exchanger superfamily that can also transport Li+ [73,115]. Na+/Ca2+ exchangers are characterized as transporters with a low affinity and high capacity; thus, they could be most effective in regulating of Ca2+ homeostasis during transient Ca2+ fluxes commonly expressed in excitable cells [116,117].
NCLX mechanism predominates in the mitochondria of cardiomyocytes, neurons, cells of the skeletal muscle, parotid gland, adrenal cortex, and brown fat [115,118,119] and to a lesser extent also being present in lung mitochondria and mitochondria of the kidney and liver [2,67,120]. NCLX can be inhibited by benzodiazepines and CGP37157 inhibitor of the mitochondrial Na+/Ca2+ exchanger [121,122]. Of note, under conditions when mitochondria are depolarized, all types of Ca2+ exchangers can act in the reverse mode, pumping Ca2+ into the mitochondria [13,123].
2.3.2. The Mechanism of Ca2+ Efflux by HCX
HCX (H+/Ca2+ exchanger): Na+-independent Ca2+ efflux (HCX) is prevalent in mitochondria of non-excitable cells (i.e., liver, kidney, lung, smooth muscles), in contrast to the NCLX mechanism [16,67,94,119,124,125]. The molecular composition of the HCX is still unclear and the literature on this complex sparse; however, it is assumed to be electroneutral with the stoichiometry of 2 molecules of H+ per 1 molecule of Ca2+ [125,126]. The rate of Ca2+ efflux through HCX decreases with an increase in the pH gradient [124,126].
2.3.3. The Mechanism of Ca2+ Efflux by LETM1
LETM1 (leucine zipper- EF-hand containing transmembrane protein, 70 kDa): In comparison to NCX, NCLX, or HCX, Ca2+ efflux via LETM1 does not represent the major pathway, but it could serve as an alternative mechanism for the release of Ca2+ [2,12,103,111,112,119]. Moreover, the activity of this protein might be essential for maintenance of the tubular shape of mitochondria and for cristae organization [9,96]. In addition, LETM1 can work as a Ca2+/H+ antiporter (see Section 2.2.4—The Mechanism of Ca2+ Uptake Including LETM1) [127].
2.3.4. The Mechanism of Ca2+ Efflux by mPTP/mPTPC
mPTP/mPTPC (mitochondrial permeability transition pore or mPTP complex): mPTP or mPTPC is considered as the main transport system for Ca2+ efflux from mitochondria under pathophysiological conditions [2,3,12,31,32,128,129,130,131,132,133,134]. Although the mPTPC was initially described in swelling experiments using the fraction of isolated mitochondria and characterized as a non-selective channel that transports ionic and nonionic molecules as early as 1979 [110], the transport mechanism of this channel actually remains poorly understood.
It is commonly believed that mPTPC is a multi-protein system in the OMM and IMM. Originally, only regulatory components were identified. The first unambiguously established component was CypD (Figure 1), which still remains the only protein whose involvement in mPTPC pore formation and activity regulation is undisputed [135,136,137,138,139]. CypD can stimulate structural rearrangements in the proteins responsible for the formation of mPTPC pore channel, preventing mPTP-mediated necrosis [13,136]. Most of the studies on the role of CypD in the regulation of mPTP relied on pharmacological cyclosporin A or transient siRNA inhibition of CypD, as well as on the results obtained on models of the knockout mouse, which demonstrated its interconnection with mPTPC [135,137,138,140].
Adenine nucleotide translocase (ANT) was initially believed to represent the main regulatory component of mPTPC [141]. Recent studies characterized ANT as a pore-forming component and proposed a “multi-pore model” with two separate pore-forming molecular components: one of which is ANT and the other depends on CypD [142]. It is also possible that CypD and ANT function in a “dual regulatory model”, where mPTPC is regulated by both ANT and CypD [142]. Moreover, it is currently believed that ANTs are multifunctional proteins, which represent not only the pore-forming component of the mPTPC but may also be crucial for mitochondrial uncoupling and for the stimulation of mitophagy [143].
Furthermore, F0F1 ATP Synthase and the phosphate carrier (PiC) are considered as the core pore-forming components of mPTPC [130,144,145,146,147,148]. FoF1 ATP Synthase forms the channel in mPTC and transports molecules through the 2 ATP synthase monomers or through the ring of the c-subunit, which overlaps with the IMM and the pore forming component [148,149]. However, it should be noted that classification of the last named component (PiC) is more complicated, since in the context of its ability to activate mPTP opening it can be considered as the pore forming component [150]. At the same time, following patch clamping of the PiC displayed too low of a conductance to assume that it functions as the core pore-forming constituent of the mPTPC. Undoubtedly, the precise nature and molecular organization of the pore-forming part of mPTPC remain controversial [130,148,149,151,152,153,154,155,156,157,158,159,160,161]. m-AAA protease Spastic Paraplegia 7 (SPG7) was previously thought to be a core component of the mPTP that is able to interact with CypD and with VDAC1 at the OMM/IMM contact sites [136]. However, recent results demonstrate that SPG7 is not a core component of the mPTP, but could regulate the mPTP activity by decreasing Ca2+ levels in mitochondrial matrix through modulation of MCUR1 and MCU assembly [146].
The efflux of Ca2+ occurs through a transient or low conductance opening of mPTP, most likely by lower oligomeric states of mPTP [13,131,162,163,164,165,166,167,168]. The evidence for transient opening of mPTP for Ca2+ was demonstrated by the early studies on the inhibition of Ca2+ release by Cyclosporin A in isolated adult rat ventricular cardiomyocytes [133]. Transient opening or low conductance opening of the mPTP represent a Ca2+ efflux mechanism, and various studies have confirmed the essential role of mPTP in the release of Ca2+ [13,131,164,165,166,167,168]. mPTP is a nonspecific channel, used by cells in signal transduction and the transfer of molecules between the mitochondrial matrix and cytoplasm. In particular it maintains Ca2+ homeostasis, regulates oxidative stress signals, and mediates cell death [128,131,169,170]. Regarding the multi-conductance function of mPTPC, it likely can be assumed that mPTPC is partially oligomerized into a complex with multiple subunits [132,170,171]. The first studies using different sized polyethylene glycols identified solutes of up to 1500 Da that could be transported through the pore that matches the modeled pore size of 1.4 nm [110]. Importantly, mPTP is able to reversibly open upon an increase in ADP concentration, as well as during restoration of the Mg2+/Ca2+ ratio [110], reestablishing mitochondrial membrane potential, and allowing for mPTP to have either a sustained or transient opening [132,172]. The different regimes of mPTP opening determine the selectivity in signaling.
The opening of mPTP is directly regulated by the concentration of free Ca2+, and triggered by mitochondrial Ca2+ overload, allowing for rapid Ca2+ release from mitochondria [3,13,32,70,132,156,173]. Obviously, Ca2+ is the most important regulator and inductor of mPTP opening, regarding its numerous indirect roles in the regulation and modulation of the mPTP [130,131,132,174]. The functional dualism of Ca2+ is an important factor of mPTP mediation. At physiological levels of Ca2+ it can activate transient opening of the pore; whereas at Ca2+ overload it can induce pathological changes, resulting in sustained mPTP opening and subsequent mitochondrial and cellular dysfunction [128,132,134,173,175].
Activation of mPTP could also be mediated at different levels through regulation by kinases, as well as posttranslational modification of CypD [176]. It has been shown, that mPTP could be stimulated by Ca2+ in combination with an increase in the concentration of ROS and phosphate; additionally, that it could be inhibited by divalent cations (such as Mg2+, Mn2+), adenine nucleotides, low pH, or CypD inhibitors (such as CsA and sanglifehrin A) [177]. Importantly, modifications and loss of CypD induce a significant increase in the threshold concentration of Ca2+ required for pore opening [13,136].
Hypothetically, VDAC could also mediate mPTPC activity; however, genetic analysis did not prove to be any essential function of this protein in mPTP-mediated cell death [53,175]. Electrophysiological and biochemical studies supported the molecular model of mPTPC with the VDAC on the OMM, ANT on the IMM, and CypD in the matrix [178,179,180]. In brief, the following facts speak for involvement of VDAC1 in mPTP opening and function: overexpression of microRNA-7 prevents opening of mPTP by downregulating VDAC1 [181]; the loss of mitochondrial fission factor Mff inhibits mPTP opening via blocking of VDAC1 oligomerization and separation of HKII, which leads to the inhibition of mPTP opening [1,182]. On the other hand, additional studies have provided opposing results, indicating that the closed state of VDAC stimulates Ca2+ permeability, and therefore forces mPTP opening [183,184].
3. The Family of Bcl-2 Proteins
The Bcl-2 (B-cell lymphoma-2) family of proteins represents a collection of pro- and anti-apoptotic proteins, functioning in a delicate balance during homeostasis (Figure 2). The main role of this group of proteins is the regulation of caspase activity and the execution of cells as a consequence of different intracellular and extracellular stimuli, and thereby dictating the fate of a cell [185,186,187,188,189]. They work as regulators of cell cycle progression, or autophagy, as well as the mediators of Ca2+ concentration, unfolded protein response and metabolism of the lipids and carbohydrates [169,185,190,191]. Importantly, Bcl-2 proteins are crucial regulators of Ca2+ homeostasis and dynamics due to their ability to directly target and regulate the intracellular Ca2+-transport systems, in particular IP3 receptors and mitochondrial Ca2+ transport systems [192,193,194].
Members of the Bcl-2 group of proteins arose during metazoan evolution [195,196], and they are characterized by the existence of short conserved sequence regions, the so-called Bcl-2 homology (BH) domains [197]. Up to now, four different BH domains have been described, and are numbered according to their date of discovery [198]. The first protein to be discovered was the Bcl-2 protein, first observed at the chromosome translocation breakpoints of leukemic cells and follicular lymphoma [199,200,201]. Bcl-2 members are typically α-helical globular proteins composed of 9 helices, although most BH3-only proteins are largely unstructured [202]. All proteins from this family share a common BH3 amino acid sequence, which is present in both anti-apoptotic and pro-apoptotic Bcl-2 homologs, as well as in a separate group of BH3-only proteins. BH3-containing helices regulate various of their physical interactions, which result either in cell survival or in one of the types of cell death. These properties have recently led to the development of therapeutic BH3 mimetics [188,189,191,203,204,205].
The Bcl-2 family comprises numerous members, categorized into 3 groups according to the total number of BH domains and their role in apoptosis (Figure 2): (1) pro-survival (i.e., anti-apoptotic) Bcl-2 proteins; (2) pro-cell death (pro-apoptotic) Bcl-2 proteins; and (3) BH3-only proteins [3,169,187,206,207,208,209,210,211].
3.1. Anti-Apoptotic Bcl-2 Proteins
The subfamily of pro-survival Bcl-2 proteins includes proteins with anti-apoptotic activity: Bcl-2 itself, Bcl-XL (Bcl-extra long), Mcl-1 (myeloid cell leukemia-1), Bcl-w (Bcl-2-like protein 2), and BFL-1/A1 (Table 1). This subfamily contains BH1, BH2, and BH3, and in some cases also a BH4 domain [211,212,213,214]. These members are generally integrated within the mitochondrial membrane, but are also present in other organelles (such as the endoplasmic reticulum). Furthermore, they are found in many cell types and tissues, where co-expression of multiple pro-survival proteins often occur [214,215]. Typically, they are anchored in the OMM by their α9 helix, and exposed to the cytoplasm by their globular Bcl-2 fold [202]. The lead member Bcl-2 (around 26 kDa) and its homologue Bcl-XL (around 27 kDa) are characterized by the presence of all four BH domains, and exert their anti-apoptotic role mainly through the involvement of BH1, BH2, and BH3 homology domains that mediate the interactions with the pro-apoptotic Bcl-2 members, preventing MOMP and subsequent apoptosis [188,211,216]. Another member, Mcl-1 (around 37 kDa), exhibits several unique features. First, it undergoes rapid degradation in response to different stress stimuli that may shift the threshold for induction of apoptosis. Second, it displays its own binding-affinity profile [217,218]. Mcl-1 sequesters both BIM and tBID activators, plus the putative PUMA and NOXA activators, as well as pore-forming proteins (Bak and Bax) [218,219,220].
3.2. Pro-Apoptotic Bcl-2 Proteins
The subfamily of pro-apoptotic Bcl-2 proteins contains: Bax (Bcl-2-associated X protein), Bak (Bcl-2-antagonist/killer-1) directly promoting MOMP, and Bok (Bcl-2 related ovarian killer), which is crucial for embryonic development; however, its role in cell death is as yet unclear. Proteins from this subfamily contain all four BH domains (Figure 2) [221,222,223]. Corresponding to the anti-apoptotic members of the Bcl-2 group of proteins, various pro-apoptotic proteins are present in cells and are involved in the regulation of apoptosis [215]. Bax (around 21 kDa) was the first identified pro-apoptotic member and an inhibitory binding partner of Bcl-2 [224].
Both Bax and Bak (around 28 kDa) are directed at the OMM by a C-terminal tail anchor [225]. It is proposed that Bak, like other C-terminal tail anchor proteins, is constitutively bound to its target—the mitochondrial membrane. In contrast, it has been shown that Bax is not normally bound to the mitochondrial membrane [226,227]. Bax is predominantly a monomer within the cytosol of nonapoptotic cells, with its C-terminal tail anchor sequence folded back and hidden within a hydrophobic groove on the protein’s surface [228]. However, recent results have shown that Bax is constitutively targeted to mitochondria during the apoptotic process and translocated back to the cytosol during normal cellular functioning [227]. It is now believed that Bax translocation between cytosol and mitochondria is a constitutive and dynamic process, and that Bax is in a dynamic balance between the cytosol and the membrane [215,227].
The pro-apoptotic function of Bax and Bak is activated in response to a range of cell death factors, causing them to undergo conformational changes, membrane-insertion, oligomerization, and the formation of a channel in the OMM [188]. Permeabilization of the mitochondrial membrane leads to the release of cytochrome c, caspase activation, and cell death [188]. Another member of this subfamily Bok (around 23 kDa) is characterized as the homolog of Bax and Bak, and it is involved in the regulation of apoptotic processes. It was demonstrated that mice with the Bok deficiency have a normal phenotype; however, enforced Bok expression could induce apoptosis in a wide range of cell types [215,229,230]. Moreover, it is still unclear whether Bok involvement in the regulation of apoptosis requires Bax or Bak [214,231].
3.3. BH3-Only Bcl-2 Proteins
The subfamily of BH3-only Bcl-2 proteins represents a heterogeneous group, which with the exception of BID contains proteins with only one highly conserved BH3 domain: BIM (Bcl-2-like 11), BID (BH3 interacting domain death agonist), PUMA (p53-upregulated modulator of apoptosis), NOXA (phorbol-12-myristate13-acetate-induced protein 1), BAD (Bcl-2-associated agonist of cell death), BIK (Bcl-2 interacting killer), HRK (Harakiri)/HRK/DP5 (Bcl-2 interacting protein death protein 5), and BMF (Bcl-2 modifying factor) (Figure 2) [3,209,210,212,231].
Upregulation of BH3-only proteins can occur at transcriptional/post-translational levels in response to stress, and this may result in cell death [215]. For example, PUMA (ca. 22 kDa) and NOXA (ca 11 kDa) are transcriptional targets of the p53 tumor suppressor, and their expression is increased in response to cytotoxic stimuli that activate p53 [232]. BID (ca 22 kDa) bears a BH3-motif with a folded α-helical structure [233,234]. During apoptosis, BID is cleaved by caspase-8 or other compounds including other caspases, granzyme B, calpains, or cathepsins [234]. Protease-cleaved Bid migrates to the mitochondria where it induces MOMP, which is dependent on the presence of Bax and/or Bak. Thus, BID acts as a sentinel for protease-mediated death signals [197,235].
3.4. Interactions between Bcl-2 Proteins
Various competing models describe the interactions of Bcl-2 proteins during the mediation of MOMP [187,210,236,237]. In all models, the BH3 domain is essential for proper functioning of the proteins, to a large extent controlled by mutual interactions of the proteins in intracellular membranes [189,238,239,240]. The most accepted model for the regulation of apoptosis by Bcl-2 proteins suggests heterodimeric interactions between Bcl-2 proteins playing an important role in the activation of Bax and Bak [3,241,242]. A specific subset of BH3-only proteins named “activators” (including BID, BIM, PUMA, and NOXA) directly bind and activate Bax and Bak, or inhibit pro-survival proteins; thereby inducing a series of their conformational changes, subsequent homo- and hetero-oligomerization within the OMM, and finally MOMP [3,189,218,240,241,242,243,244]. In turn, MOMP allows for the release of mitochondrial apoptogenic factors into the cytosol (including cytochrome c), SMAC (second mitochondria-derived activator of caspase), and DIABLO (direct IAP binding protein with low pI), HtrA2 (high temperature requirement protein A2), as well as others, which then initiate apoptosome formation and the induction of the caspase cascade [17,218,236,242].
Other members of the BH3-only proteins group most likely act as “sensitizers”. They are not the direct activators of Bax or Bak; however, they are able to bind Bcl-2 proteins with pro-survival functions [3,240]. The BH3 activator cBID sequestered by Bcl-XL complexes only changes from an inactive to an active form while bound to a Bcl-XL complex (and when Bad is also bound). Bcl-XL complexes enable Bad to function as a non-competitive inhibitor of Bcl-XL and allosterically activate BID, dramatically enhancing the pro-apoptotic potency of BAD [185,189,241]. Both the activators and sensitizers could be activated via specific mechanisms, leading to the transfer of the apoptotic stimuli to Bax and Bak, as well as to the pro-survival Bcl-2 proteins [3,242].
Interestingly, Bcl-2 proteins have a significant impact on the dynamic processes related to mitochondrial fusion and fission in cells [218,245,246,247,248,249]. The mitochondrial apoptotic pathway is based on cooperation between Bcl-2 proteins and other proteins found in mitochondria, and it is intimately related with membrane fission, thereby linking mitochondrial dynamics to apoptosis [24,247,250,251,252]. These processes are closely related to the functioning of the Ca2+ transport systems. For instance, it was demonstrated that Bax, Bak, and Bcl-XL induce changes in mitochondrial dynamics by regulating the activity of the proteins responsible for fusion and fission [246,248,249,253]. More specifically, it has been reported that Bax and Bak are involved in the control of mitochondrial fusion in different cell models (primary mouse neurons, mouse embryonic fibroblasts, and human colon carcinoma cells) through binding to the mitofusins—Mfn1 and Mfn2 [249]. It has also been shown that Bax or Bak are required in healthy cells for normal fusion of mitochondria into elongated tubules. Nevertheless, despite these interesting findings, the exact molecular mechanisms underlying the (de)activation of the mitochondrial dynamic machinery by Bcl-2 proteins still remain to be elucidated [245].
4. Regulation of Mitochondrial Ca2+ Transport Systems by Bcl-2 Proteins
Apart from the ability to influence mitochondrial permeability directly, Bcl-2 proteins are also involved in the mediation of apoptotic signaling by regulating Ca2+ homeostasis [29,185,193,254]. This regulation can occur through direct interactions between the intracellular Ca2+-transport systems and anti-apoptotic Bcl-2 members; in particular Bcl-2, Bcl-XL, Mcl-1, Bax, and Bak [255,256,257,258,259,260,261]. These members target crucial intracellular Ca2+-transport systems such as Ca2+-efflux systems of ER/SR (inositol 1,4,5-trisphosphate receptors (IP3Rs) and ryanodine receptors (RyRs)) and the Ca2+-efflux system of mitochondria (involving VDAC, which provides Ca2+ flux trough OMM, and mPTP, which represents the main Ca2+ efflux system from mitochondria under pathophysiological conditions) [193,194,211,262].
In general, the Bcl-2 proteins are major regulators of a wide range of mitochondrial functioning processes. Those involving regulation of mitochondrial dynamics and morphology have already been mentioned in chapter 3.4. Other processes are responsible for Ca2+ homeostasis and include regulation of the metabolism and functioning of the electron transport chain, permeability transition, and mPTP opening [185,193,194,254,255,263]. A substantial part of these processes is summarized in Table 1 for clarity.
4.1. Bcl-2 Proteins and Ca2+ Influx through VDAC
VDAC1 is a key component in the apoptotic pathway mediated by mitochondria, and is a target for pro- and anti-apoptotic Bcl-2 proteins [263,265,266]. The molecular composition of VDAC1 determines its mobility, which is necessary for controlling the channel gating and the interactions with pro- and anti-apoptotic proteins [56,256,263,266,267].
It was demonstrated that Bcl-2 proteins are able to directly target VDAC, and in turn modulate its activity [255]. These interactions mediate changes in a large number of mitochondrial processes, and may also regulate Ca2+ transport in mitochondria [268,269]. VDAC1 is considered as a key protein in the mPTP-mediated apoptosis by associating with pro- and anti-apoptotic members of the Bcl-2 family and hexokinase, and promotes the release of apoptotic proteins [268]. It has been shown that when the VDAC1 channel closes, it becomes impermeable to ATP and metabolites, but the permeability for Ca2+ increases [270]. This confirms the critical role of VDAC1 in mitochondrial Ca2+ homeostasis and the induction of cell death. On top of that, VDAC1 regulates translocation of pro-survival Ca2+ signals from the cytosol and ER/SR, as well as the ATP transport and metabolite exchange between the cytosol and mitochondria [193]. Furthermore, pro-survival proteins (e.g., hexokinase-I) are required for the correct functioning of VDAC1 and Ca2+ signal transduction to mitochondria (also see Section 2.1) [271,272,273].
Considering all these VDAC1 roles in Ca2+-determined cell functioning and death it can be expected that Mcl-1, Bcl-XL, and Bcl-2 are able to emulate the interactions between VDAC1 and Ca2+ transport in mitochondria (Table 1). For instance, it has been shown that Bcl-2 and Bcl-XL bind directly to the N-terminus of VDAC1, thereby inhibiting mitochondrial Ca2+ uptake and protecting cells from extreme mitochondrial uptake of Ca2+ [256,266]. Similarly, it was shown that BH4 oligopeptides of Bcl-2 and Bcl-XL inhibit VDAC1 activity on liposomes and apoptotic depolarization of isolated mitochondria [254]. Turning to the mechanism by which this could happen, it has been suggested that Bcl-XL promotes the open configuration of VDAC1 [274] and the BH4 domain of Bcl-XL selectively targets one to two VDAC1 molecules [260,269,275,276]. Despite these findings, a study that comparing wild-type MEF cells versus Bcl-XL-gene deficient MEF cells demonstrated that the presence of Bcl-XL promoted the accumulation of Ca2+ in the matrix by increasing Ca2+ transfer across the OMM [269]. As was demonstrated by electrophysiological studies, Bcl-XL could stimulate either the increase [274] or the decrease [276] in VDAC1 conductance. Interestingly, structural-functional studies of Bcl-XL demonstrate that induction of Ca2+ influx into mitochondria by Bcl-XL is related to the formation of Bcl-XL dimers [277]. Formation of these dimers is facilitated by the recruitment of Bcl-XL to the membrane, and is inhibited by BH3 mimetics [277]. From this point of view, Bcl-XL is able to enhance the activity of enzymes involved in the Krebs cycle, and can therefore stimulate mitochondrial bioenergetic processes in healthy nonapoptotic cells. This means that Bcl-XL could function as a direct anti-apoptotic protein capable of enhancing transduction of the Ca2+ signals to the mitochondria from the cytosol and ER/SR, and mediate Ca2+ oscillations that are crucial for both ATP synthesis and the bioenergetic balance [193,278].
Another key anti-apoptotic member of the Bcl-2 family that is capable of direct binding to VDAC channels is Mcl-1 [257]. It was reported that Mcl-1 associates with high affinity to VDAC1 and VDAC3 isoforms, but not to VDAC2 [257]. In A549 cells, reduction of Mcl-1 expression levels lowered Ca2+ uptake into the mitochondrial matrix and inhibited generation of ROS. It has been demonstrated that in A549, H1299, and H460 cells Mcl-1 knockdown and VDAC derived peptides decreased cell migration, but that cell proliferation remained unchanged. Of note, cell migration could be restored in Mcl-1 knockdown cells by increasing the levels of ROS (following the experimental model in which migration is activated by the production of ROS). These data suggest that an interaction between Mcl-1 and VDAC promotes cell migration by a mechanism that involves Ca2+-dependent ROS production [257].
Furthermore, there is genetic and pharmacologic evidence that the BID and BAD pro-apoptotic factors also participate in the influx of Ca2+ by close cooperation with Bcl-XL and Bcl-2 [194] (Table 1). More specifically, the mitochondrion-localized isoform of Bcl-XL could induce cell migration by interacting with VDAC1 to mediate Ca2+ transport to mitochondria from the ER/SR and other Ca2+ depots in a BH3-dependent manner.
4.2. Bcl-2 Proteins and Na+/Ca2+ Exchangers
Although there is no evidence for direct interaction between a member of the Bcl-2 family and the Na+/Ca2+ exchangers family, it was shown already in 2001 that Bcl-2 could inhibit Ca2+ transport via the Na+/Ca2+ exchanger [264] (Table 1). More specifically, Bcl-2 overexpression in transgenic mice induced reduction of Na+/Ca2+ exchange and promoted the resistance to the permeability transition. This behavior was accompanied by increased Ca2+ concentration and the induction of transport proteins that could regulate the apoptotic pathway [264]. Despite these reports supporting the regulation of Na+/Ca2+ exchangers by Bcl-2 proteins, this research needs to be continued.
4.3. Bcl-2 Proteins and mPTP (See Also Section Calcium Efflux through mPTPC)
Direct protein–protein interactions between Bcl-2 proteins and proteins involved in mPTPC formation (ANT, ATP synthase, and VDAC) have been documented [32,255,261] (Table 1). In particular, the pro-apoptotic Bcl-2 proteins Bax and Bak have been reported to interact directly with mPTPC [252,258]. Both Bax and Bak participate in mPTPC mediation because of their ability to stimulate and increase MOMP, which allows mPTP opening followed by mitochondrial swelling [255,258,261]. It was also demonstrated that Bcl-2 could positively modulate ANT activity, whereas Bax inhibits ANT function by destabilizing the interactions with Bcl-2 [279]. The following findings that support a close relationship between Bcl-2 and mPTPC were observed in cells/mice deficient in Bax and Bak: (1) mPTP-induced cell death and related mitochondrial damage is inhibited in cardiac cells after a myocardial infarction [259]; (2) mitochondria are resistant to the opening of mPTP; (3) reconstitution of Bax in such cells restored both the mPTP opening [258,259] and outer mitochondrial membrane permeability [258]; and (4) Bax/Bak deficiency is protective regarding the conditions of ischemia-reperfusion injury [258,259]. According to expectations, Bcl-2 members with anti-apoptotic activity could also influence mPTPC by functioning through the interactions with regulatory components [274,280,281]. Further investigations are needed to clarify the molecular mechanism of mPTP regulation by Bax and Bak; nevertheless, recent studies demonstrate the importance of mitochondrial lipid composition for their ability to permeabilize the mitochondrial membrane [282].
In summary, it is evident that Bcl-2 proteins regulate the activity of the proteins and multiprotein complexes of mitochondrial Ca2+-transport systems, including VDAC, Na+/Ca2+ exchangers, and mPTPC. These interconnections influence regulation of Ca2+ homeostasis in mitochondria and in the cell, and mediate mitochondrial cell death pathways.
5. Regulation of Cell Death by Mitochondrial Transport of Calcium and Bcl-2 Proteins
The Nomenclature Committee on Cell Death 2018 classified cell death types regarding their biochemical characteristics, and following this protocol there are 12 different types of regulated cell death (RCD) (see Box 3 for more details) [283]. Additionally, programmed cell death (PCD) that takes place during physiological development and tissue turnover is classified as a physiological form of RCD [283].
Box 3. The cell death.
The 12 different types of the regulated cell death have so far been classified: necroptosis, ferroptosis, pyroptosis, parthanatos, entotic cell death, NETotic cell death, lysosome-dependent cell death (LDCD), autophagy-dependent cell death (ADCD), immunogenic cell death (ICD), intrinsic apoptosis, extrinsic apoptosis, and mitochondrial permeability transition (MPT)-driven (mediated) necrosis.
The extrinsic (i.e., the death receptor pathway) and/or intrinsic apoptotic pathway (i.e., mitochondrial, MOMP-mediated pathway) occur in response to developmental, homeostatic, or internal damage signals [284,285,286,287]. These types of cell death converge in a series of biochemical events including: activation of caspases, cell shrinkage, and plasma membrane blebbing [17,288,289,290].
Necrosis had previously been considered as unregulated and accidental cell death, a toxic process, where the cell is a passive victim that follows an energy-independent mode of death [287,288,291]. Regarding the novel classification, now it is more correct to classify separately MPT-mediated necrosis, which is often caused by intracellular factors, including oxidative stress and Ca2+ overload, leading to cell swelling, and eventually to the rupture of the plasma membrane and the typical necrotic morphological changes [283,289,290,291].
mPTP-mediated necrosis and mitochondrial-mediated apoptosis are two types of RCD that are mediated through the interconnection of the mitochondrial transport of Calcium and Bcl-2 proteins (Figure 3) [3,214]. Although both of the biochemical mechanisms and morphological changes of apoptosis and necrosis vary significantly, both mechanisms can be characterized as the “apoptosis-necrosis continuum” with a shared biochemical network [292]. Bcl-2 proteins are crucially involved in these processes by working as the key regulators of the MOMP-mediated pathway of apoptosis. This includes MOMP, primarily induced by direct binding interactions between Bcl-2 proteins (Figure 3), leading to irreversible release of intermembrane space proteins, and subsequent caspase activation, which culminates in biochemical and morphological apoptotic changes, and apoptosis itself [218,242,289,293].
As already outlined in Chapter 4, Bcl-2 proteins also interact with mPTP, which represents the key component in mPTP-mediated necrosis. Thus, the interactions between Bcl-2 proteins and mitochondrial Ca2+ transport systems might be crucial for determination of the cell death pathway—either the apoptotic or necrotic one [3,292,294] (Figure 3). Of note, mPTP opening originally was solely associated with early apoptosis; however, it was recently shown that MOMP, mediated by mPTP, is linked more closely to the late events of apoptosis and mPTP-mediated necrosis [3,174,295].
The crucial event criterion of mPTP-mediated necrosis is that Ca2+-induced formation of mPTPC takes place in the IMM [175,296]. This process is relatively quick (usually taking minutes) and is a result of an elevated Ca2+ concentration in the mitochondrial matrix [259]. Bcl-2 proteins are crucially involved in the mediation of mPTP opening, which rapidly dissipates the proton gradient across the IMM required for ATP synthesis [3,169,174,258]. Similarly, it has been demonstrated that the pro-apoptotic Bax and Bak function as the mPTPC components in the OMM and could regulate the necrotic cell death [258]. More specifically, loss of Bax/Bak decreased OMM permeability without changing IMM MPTPC function, leading to resistance to mitochondrial Ca2+-overload and mPTP-mediated necrosis. Furthermore, it has been shown that membrane reconstitution of mutants of Bax with their propensity to oligomerize and form disabled membrane pores, yet still capable of enhancing OMM permeability, allowed mPTPC-dependent swelling of mitochondria and restored necrotic cell death [258]. It has also been suggested that Bax and Bak are able to mediate mPTP opening through interactions between OMM and IMM protein contact sites [156].
Interestingly, anti-apoptotic Bcl-2 members can also mediate the activity of mPTP by direct interactions with regulatory components [169]. For instance, Bcl-XL was once thought to be present exclusively on the OMM, but currently it is widely accepted as a F0F1-ATP synthase regulator in the IMM [281,297]. Moreover, pharmacological inhibition of Bcl-2 and/or BCL-XL may induce cell sensitivity to mPTP-mediated necrosis [298]. BID has been characterized as the BH3 protein, which could cooperate with the mPTPC components and regulate necrosis [299]. Altogether, both mPTP-mediated necrosis and MOMP-induced apoptosis can be regulated by Bcl-2 proteins, and triggered by mitochondrial Ca2+ overload; causing damage of IMM integrity with catastrophic consequences, particularly bioenergetic collapse by dissipation of the mitochondrial membrane potential (Figure 3) [3,32,283,293].
6. Conclusions
In this paper, we have tried to outline a possible functional link between Bcl-2 proteins and calcium transport systems in mitochondria. The ability of Bcl-2 to control Ca2+ at multiple levels plays an undisputed role in mediation of Ca2+ homeostasis and cell death pathways. MOMP-mediated apoptosis and mPTP-mediated necrosis are determined by Ca2+ homeostasis in cells, and particularly in the mitochondria. In this respect, the opening of mPTP and MOMP are interconnected functionally through mitochondrial Ca2+ transport systems, and both lead to cell death—either MOMP-mediated apoptosis or mPTP-mediated necrosis.
Furthermore, there exists a regulatory dualism between Bcl-2 proteins and Ca2+ transport systems in mitochondria: Bcl-2 proteins not only mediate the activity of mitochondrial Ca2+ transport systems, but at the same time the changes in Ca2+ transport are important for the induction of mitochondrial cell death pathways via Bcl-2 proteins. Great efforts have been made to understand the interactions of Bcl-2 proteins and their ability to mediate cell fate by regulation of Ca2+ homeostasis. However, the regulatory mechanisms of these interactions are still an open issue, which requires additional research. Undoubtedly, a detailed understanding of mitochondrial processes is very important for the analysis of the functions and dysfunctions in living cells, from the viewpoint of mitochondrial biochemistry as well as from the viewpoint of applications in mitochondrial biomedicine used for the diagnosis and treatment of various complex diseases at the subcellular and molecular levels.
Author Contributions
Conceptualization, N.N. and R.Š.; writing—original draft preparation, N.N. and R.Š.; writing—review and editing, N.N. and R.Š.; funding acquisition, R.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by GAČR grant number 18-04871S.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ATP | adenosine triphosphate |
| Bcl-2 | B-cell lymphoma-2 |
| TCA | tricarboxylic acid |
| MOMP | mitochondrial outer membrane permeabilization |
| NADH | nicotinamide adenine dinucleotide |
| IMM | inner mitochondrial membrane |
| IMS | intermembrane space |
| OMM | outer mitochondrial membrane |
| MCU | mitochondrial Ca2+ uniporter |
| CC-domains | coiled-coil domains |
| mRyR | mitochondrial ryanodine receptor |
| RaM | rapid mode of Ca2+ uptake |
| LETM1 | leucine zipper- EF-hand containing transmembrane protein |
| ETC | electron transport chain |
| NCX | Na+/Ca2+ exchanger |
| NCLX | Na+/Ca2+/Li+ exchanger |
| HCX | H+/Ca2+ exchanger |
| mPTPC | mitochondrial permeability transition pore complex |
| mPTP | mitochondrial permeability transition pore |
| ANT | adenine nucleotide translocator |
| CypD | cyclophilin D |
| PiC | phosphate carrier |
| VDAC | voltage-dependent anion channel |
| MCUb | mitochondrial Ca2+ uniporter dominant negative beta subunit |
| EMRE | essential MCU regulatory element |
| MICU | mitochondrial Ca2+ uptake protein |
| MCUR1 | mitochondrial Ca2+ uniporter regulator 1 |
| RaM | RApid mode of Ca2+ uptake |
| SLC25A23 | solute carrier 25A23 |
| mRyR | mitochondrial ryanodine receptor |
| PiC | phosphate carrier |
| SPG7 | m-AAA protease Spastic Paraplegia 7 |
| Mff | mitochondrial fission factor |
| BH domains | Bcl-2 homology domains |
| Bcl-XL | Bcl-extra long |
| Mcl-1 | myeloid cell leukemia-1 |
| Bcl-w | Bcl-2-like protein 2 |
| BFL-1/A1 | Bcl-2-related protein A1 |
| BIM | Bcl-2-like 11 |
| BID | BH3 interacting domain death agonist |
| PUMA | p53-upregulated modulator of apoptosis |
| NOXA | phorbol-12-myristate13-acetate-induced protein 1 |
| BAD | Bcl-2-associated agonist of cell death |
| BIK | Bcl-2 interacting killer |
| HRK | Harakiri protein |
| HRK/DP5 | Harakiri protein/Bcl-2 interacting protein death protein 5 |
| BMF | Bcl-2 modifying factor |
| SMAC | second mitochondria-derived activator of caspase |
| DIABLO | direct IAP binding protein with low pI |
| HtrA2 | high temperature requirement protein A2 |
| Mfn | mitofusin |
| ROS | reactive oxygen species |
| RCD | regulated cell death |
| PCD | programmed cell death |
| ADCD | autophagy-dependent cell death |
| ICD | immunogenic cell death |
| LDCD | lysosome-dependent cell death |
| ER | endoplasmic reticulum |
| SR | sarcoplasmic reticulum |
References
- Carafoli, E.; Krebs, J. Why calcium? how calcium became the best communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Membrane Dynamics and Calcium Signaling. In Advances in Experimental Medicine and Biology; Krebs, J. (Ed.) Springer International Publishing: Cham, Switzerland, 2017; Volume 981, ISBN 978-3-319-55857-8. [Google Scholar]
- Del Re, D.P.; Amgalan, D.; Linkermann, A.; Liu, Q.; Kitsis, R.N. Fundamental mechanisms of regulated cell death and implications for heart disease. Physiol. Rev. 2019, 99, 1765–1817. [Google Scholar] [CrossRef] [PubMed]
- Eisner, D.A.; Caldwell, J.L.; Kistamás, K.; Trafford, A.W. Calcium and excitation-contraction coupling in the heart. Circ. Res. 2017, 121, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Brini, M.; Calì, T.; Ottolini, D.; Carafoli, E. Neuronal calcium signaling: Function and dysfunction. Cell Mol. Life Sci. 2014, 71, 2787–2814. [Google Scholar] [CrossRef] [PubMed]
- Duchen, M.R. Ca2+-Dependent changes in the mitochondrial energetics in single dissociated mouse sensory neurons. Biochem. J. 1992, 283, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elustondo, P.A.; Nichols, M.; Robertson, G.S.; Pavlov, E.V. Mitochondrial Ca2+ uptake pathways. J. Bioenergy Biomembr. 2017, 49, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Duchen, M.R. Mitochondria in health and disease: Perspectives on a new mitochondrial biology. Mol. Asp. Med. 2004, 25, 365–451. [Google Scholar] [CrossRef]
- Mitochondrial Dynamics in Cardiovascular Medicine. In Advances in Experimental Medicine and Biology; Santulli, G. (Ed.) Springer International Publishing: Cham, Switzerland, 2017; Volume 982, ISBN 978-3-319-55329-0. [Google Scholar]
- Bravo-Sagua, R.; Parra, V.; Lopez-Crisosto, C.; Diaz, P.; Quest, A.F.G.; Lavandero, S. Calcium transport and signaling in mitochondria. Compr. Physiol. 2017, 7, 623–634. [Google Scholar] [CrossRef]
- Giorgio, V.; Guo, L.; Bassot, C.; Petronilli, V.; Bernardi, P. Calcium and regulation of the mitochondrial permeability transition. Cell Calcium 2018, 70, 56–63. [Google Scholar] [CrossRef]
- Delierneux, C.; Kouba, S.; Shanmughapriya, S.; Potier-Cartereau, M.; Trebak, M.; Hempel, N. Mitochondrial calcium regulation of Redox signaling in cancer. Cells 2020, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Belosludtsev, K.N.; Dubinin, M.V.; Belosludtseva, N.V.; Mironova, G.D. Mitochondrial Ca2+ transport: Mechanisms, molecular structures, and orle in cells. Biochemistry 2019, 84, 593–607. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Schulz, R.; Girao, H.; Kwak, B.R.; De Stefani, D.; Rizzuto, R.; Bernardi, P.; Di Lisa, F. Mitochondrial ion channels as targets for cardioprotection. J. Cell. Mol. Med. 2020, 24, 7102–7114. [Google Scholar] [CrossRef] [PubMed]
- Glaser, T.; Arnaud Sampaio, V.F.; Lameu, C.; Ulrich, H. Calcium signalling: A common target in neurological disorders and neurogenesis. Semin. Cell Dev. Biol. 2019, 95, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Trebak, M.; Earley, S. Mitochondria structure and position in the local control of calcium signals in smooth muscle cells. In Signal Transduction and Smooth Muscle; McCarron, J.G., Saunter, C., Wilson, C., Girkin, J.M., Chalmers, S., Eds.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781498774222. [Google Scholar]
- Bock, F.J.; Tait, S.W.G. Mitochondria as multifaceted regulators of cell death. Nat. Rev. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar] [CrossRef]
- Wacquier, B.; Combettes, L.; Dupont, G. Dual dynamics of mitochondrial permeability transition pore opening. Sci. Rep. 2020, 10, 3924. [Google Scholar] [CrossRef] [Green Version]
- Denton, R.M. Regulation of mitochondrial dehydrogenases by calcium ions. Biochim. Biophys. Acta Bioenergy 2009, 1787, 1309–1316. [Google Scholar] [CrossRef] [Green Version]
- Picard, M.; Wallace, D.C.; Burelle, Y. The rise of mitochondria in medicine. Mitochondrion 2016, 30, 105–116. [Google Scholar] [CrossRef]
- Duchen, M.R. Mitochondria and calcium: From cell signalling to cell death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef]
- Park, M.K. Perinuclear, perigranular and sub-plasmalemmal mitochondria have distinct functions in the regulation of cellular calcium transport. EMBO J. 2001, 20, 1863–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgi, C.; Marchi, S.; Pinton, P. The machineries, regulation and cellular functions of mitochondrial calcium. Nat. Rev. Mol. Cell Biol. 2018, 19, 713–730. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The cell biology of mitochondrial membrane dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef] [PubMed]
- Morciano, G.; Marchi, S.; Morganti, C.; Sbano, L.; Bittremieux, M.; Kerkhofs, M.; Corricelli, M.; Danese, A.; Karkucinska-Wieckowska, A.; Wieckowski, M.R.; et al. Role of Mitochondria-associated ER membranes in Calcium regulation in cancer-specific settings. Neoplasia 2018, 20, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Cruz, M.S.; Simmen, T. Cancer: Untethering Mitochondria from the endoplasmic reticulum? Front. Oncol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Singaravelu, K.; Nelson, C.; Bakowski, D.; de Brito, O.M.; Ng, S.-W.; Di Capite, J.; Powell, T.; Scorrano, L.; Parekh, A.B. Mitofusin 2 regulates STIM1 migration from the Ca2+ store to the plasma membrane in cells with depolarized Mitochondria. J. Biol. Chem. 2011, 286, 12189–12201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouaville, L.S.; Pinton, P.; Bastianutto, C.; Rutter, G.A.; Rizzuto, R. Regulation of mitochondrial ATP synthesis by calcium: Evidence for a long-term metabolic priming. Proc. Natl. Acad. Sci. USA 1999, 96, 13807–13812. [Google Scholar] [CrossRef] [Green Version]
- Carpio, M.A.; Katz, S.G. Methods to probe Calcium regulation by BCL-2 family members. Methods Mol. Biol. 2019, 1877, 173–183. [Google Scholar] [CrossRef]
- Rong, Y.; Distelhorst, C.W. Bcl-2 protein family members: Versatile regulators of Calcium signaling in cell survival and apoptosis. Annu. Rev. Physiol. 2008, 70, 73–91. [Google Scholar] [CrossRef]
- Briston, T.; Roberts, M.; Lewis, S.; Powney, B.; Staddon, J.M.; Szabadkai, G.; Duchen, M.R. Mitochondrial permeability transition pore: Sensitivity to opening and mechanistic dependence on substrate availability. Sci. Rep. 2017, 7, 10492. [Google Scholar] [CrossRef]
- Bonora, M.; Patergnani, S.; Ramaccini, D.; Morciano, G.; Pedriali, G.; Kahsay, A.E.; Bouhamida, E.; Giorgi, C.; Wieckowski, M.R.; Pinton, P. Physiopathology of the permeability transition pore: Molecular mechanisms in human pathology. Biomolecules 2020, 10, 998. [Google Scholar] [CrossRef]
- Vasington, F.D.; Murphy, J.V. Ca ion uptake by rat kidney mitochondria and its dependence on respiration and phosphorylation. J. Biol. Chem. 1962, 237, 2670–2677. [Google Scholar]
- DeLuca, H.F.; Engstrom, G.W. Calcium uptake by rat kidney mitochondria. Proc. Natl. Acad. Sci. USA 1961, 47, 1744–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P.; Moyle, J. Chemiosmotic hypothesis of oxidative phosphorylation. Nature 1967, 213, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Ludtmann, M.H.R.; Abramov, A.Y. Mitochondrial calcium imbalance in Parkinson’s disease. Neurosci. Lett. 2018, 663, 86–90. [Google Scholar] [CrossRef]
- Bhosale, G.; Sharpe, J.A.; Sundier, S.Y.; Duchen, M.R. Calcium signaling as a mediator of cell energy demand and a trigger to cell death. Ann. N. Y. Acad. Sci. 2015, 1350, 107–116. [Google Scholar] [CrossRef]
- Rossi, A.; Pizzo, P.; Filadi, R. Calcium, mitochondria and cell metabolism: A functional triangle in bioenergetics. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1068–1078. [Google Scholar] [CrossRef]
- Bertero, E.; Maack, C. Calcium signaling and reactive oxygen species in mitochondria. Circ. Res. 2018, 122, 1460–1478. [Google Scholar] [CrossRef]
- Missiroli, S.; Perrone, M.; Genovese, I.; Pinton, P.; Giorgi, C. Cancer metabolism and mitochondria: Finding novel mechanisms to fight tumours. EBioMedicine 2020, 59, 102943. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, J.R.; Mongue-Din, H.; Eaton, P.; Shah, A.M. Redox signaling in cardiac physiology and pathology. Circ. Res. 2012, 111, 1091–1106. [Google Scholar] [CrossRef] [PubMed]
- Mammucari, C.; Raffaello, A.; Vecellio Reane, D.; Gherardi, G.; De Mario, A.; Rizzuto, R. Mitochondrial calcium uptake in organ physiology: From molecular mechanism to animal models. Pflug. Arch. 2018, 470, 1165–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Hail, D.; Shoshan-Barmatz, V. VDAC1-interacting anion transport inhibitors inhibit VDAC1 oligomerization and apoptosis. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 1612–1623. [Google Scholar] [CrossRef]
- Schein, S.J.; Colombini, M.; Finkelstein, A. Reconstitution in planar lipid bilayers of a voltage-dependent anion-selective channel obtained from paramecium mitochondria. J. Membr. Biol. 1976, 30, 99–120. [Google Scholar] [CrossRef] [PubMed]
- Mazure, N.M. VDAC in cancer. Biochim. Biophys. Acta Bioenergy 2017, 1858, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Wagner, R. Mitochondrial outer membrane channels: Emerging diversity in transport processes. BioEssays 2018, 40, 1800013. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Queralt-Martín, M.; Rosencrans, W.M.; Bezrukov, S.M. Targeting the multiple physiologic roles of VDAC with steroids and hydrophobic drugs. Front. Physiol. 2020, 11, 446. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Gincel, D. The voltage-dependent anion channel: Characterization, modulation, and role in mitochondrial function in cell life and death. Cell Biochem. Biophys. 2003, 39, 279–292. [Google Scholar] [CrossRef]
- Colombini, M.; Mannella, C.A. VDAC, the early days. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1438–1443. [Google Scholar] [CrossRef] [Green Version]
- Colombini, M. A candidate for the permeability pathway of the outer mitochondrial membrane. Nature 1979, 279, 643–645. [Google Scholar] [CrossRef]
- Kusano, T.; Tateda, C.; Berberich, T.; Takahashi, Y. Voltage-dependent anion channels: Their roles in plant defense and cell death. Plant Cell Rep. 2009, 28, 1301–1308. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Mizrachi, D. VDAC1: From structure to cancer therapy. Front. Oncol. 2012, 2, 164. [Google Scholar] [CrossRef] [Green Version]
- Camara, A.K.S.; Zhou, Y.; Wen, P.C.; Tajkhorshid, E.; Kwok, W.M. Mitochondrial VDAC1: A key gatekeeper as potential therapeutic target. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Anion channels of mitochondria. In Handbook of Experimental Pharmacology; Ponnalagu, D.; Singh, H. (Eds.) Springer: Berlin/Heidelberg, Germany, 2016; Volume 240, pp. 71–101. [Google Scholar]
- Neumann, D.; Bückers, J.; Kastrup, L.; Hell, S.W.; Jakobs, S. Two-color STED microscopy reveals different degrees of colocalization between hexokinase-I and the three human VDAC isoforms. PMC Biophys. 2010, 3, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geula, S.; Ben-Hail, D.; Shoshan-Barmatz, V. Structure-based analysis of VDAC1: N-terminus location, translocation, channel gating and association with anti-apoptotic proteins. Biochem. J. 2012, 444, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, E.H.Y.; Sheiko, T.V.; Fisher, J.K.; Craigen, W.J.; Korsmeyer, S.J. VDAC2 inhibits BAK activation and mitochondrial apoptosis. Science 2003, 301, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Checchetto, V.; Reina, S.; Magrì, A.; Szabo, I.; De Pinto, V. Recombinant human voltage dependent anion selective channel Isoform 3 (hVDAC3) forms pores with a very small conductance. Cell. Physiol. Biochem. 2014, 34, 842–853. [Google Scholar] [CrossRef] [PubMed]
- De Pinto, V.; Guarino, F.; Guarnera, A.; Messina, A.; Reina, S.; Tomasello, F.M.; Palermo, V.; Mazzoni, C. Characterization of human VDAC isoforms: A peculiar function for VDAC3? Biochim. Biophys. Acta 2010, 1797, 1268–1275. [Google Scholar] [CrossRef] [Green Version]
- Lemasters, J.J.; Holmuhamedov, E.L.; Czerny, C.; Zhong, Z.; Maldonado, E.N. Regulation of mitochondrial function by voltage dependent anion channels in ethanol metabolism and the Warburg effect. Biochim. Biophys. Acta 2012, 1818, 1536–1544. [Google Scholar] [CrossRef] [Green Version]
- Austin, S.; Nowikovsky, K. LETM1: Essential for mitochondrial biology and cation homeostasis? Trends Biochem. Sci. 2019, 44, 648–658. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tran, Q.; Shrestha, R.; Piao, L.; Park, S.; Park, J.; Park, J. LETM1 is required for mitochondrial homeostasis and cellular viability (review). Mol. Med. Rep. 2019, 19, 3367–3375. [Google Scholar] [CrossRef]
- Shao, J.; Fu, Z.; Ji, Y.; Guan, X.; Guo, S.; Ding, Z.; Yang, X.; Cong, Y.; Shen, Y. Leucine zipper-EF-hand containing transmembrane protein 1 (LETM1) forms a Ca(2+)/H(+) antiporter. Sci. Rep. 2016, 6, 34174. [Google Scholar] [CrossRef] [Green Version]
- Waldeck-Weiermair, M.; Jean-Quartier, C.; Rost, R.; Khan, M.J.; Vishnu, N.; Bondarenko, A.I.; Imamura, H.; Malli, R.; Graier, W.F. Leucine zipper EF hand-containing Transmembrane Protein 1 (Letm1) and uncoupling proteins 2 and 3 (UCP2/3) contribute to two distinct mitochondrial Ca2+ uptake pathways. J. Biol. Chem. 2011, 286, 28444–28455. [Google Scholar] [CrossRef] [Green Version]
- De Stefani, D.; Raffaello, A.; Teardo, E.; Szabo, I.; Rizzuto, R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 2011, 476, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Baughman, J.M.; Perocchi, F.; Girgis, H.S.; Plovanich, M.; Belcher-Timme, C.A.; Sancak, Y.; Bao, X.R.; Strittmatter, L.; Goldberger, O.; Bogorad, R.L.; et al. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 2011, 476, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunter, T.E.; Pfeiffer, D.R. Mechanisms by which mitochondria transport calcium. Am. J. Physiol. 1990, 258, C755–C786. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Jhun, B.S.; Hurst, S.; O-Uchi, J.; Csordás, G.; Sheu, S.S. The mitochondrial Ca2+ uniporter: Structure, function, and pharmacology. Handb. Exp. Pharmacol. 2017, 240, 129–156. [Google Scholar] [CrossRef] [Green Version]
- Pallafacchina, G.; Zanin, S.; Rizzuto, R. Recent advances in the molecular mechanism of mitochondrial calcium uptake. F1000Research 2018, 7, 1858. [Google Scholar] [CrossRef]
- Granatiero, V.; De Stefani, D.; Rizzuto, R. Mitochondrial calcium handling in physiology and disease. Adv. Exp. Med. Biol. 2017, 982, 25–47. [Google Scholar] [CrossRef]
- Sancak, Y.; Markhard, A.L.; Kitami, T.; Kovacs-Bogdan, E.; Kamer, K.J.; Udeshi, N.D.; Carr, S.A.; Chaudhuri, D.; Clapham, D.E.; Li, A.A.; et al. EMRE is an essential component of the mitochondrial calcium uniporter complex. Science 2013, 342, 1379–1382. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Yang, J.; Fu, L.; Wang, M.; Wang, X. Progress in understanding mitochondrial calcium uniporter complex-mediated calcium signalling: A potential target for cancer treatment. Br. J. Pharmacol. 2019, 176, 1190–1205. [Google Scholar] [CrossRef]
- Pathak, T.; Trebak, M. Mitochondrial Ca2+ signaling. Pharmacol. Ther. 2018, 192, 112–123. [Google Scholar] [CrossRef]
- Raffaello, A.; De Stefani, D.; Sabbadin, D.; Teardo, E.; Merli, G.; Picard, A.; Checchetto, V.; Moro, S.; Szabo, I.; Rizzuto, R. The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit. EMBO J. 2013, 32, 2362–2376. [Google Scholar] [CrossRef] [Green Version]
- Patron, M.; Checchetto, V.; Raffaello, A.; Teardo, E.; Vecellio Reane, D.; Mantoan, M.; Granatiero, V.; Szabo, I.; De Stefani, D.; Rizzuto, R. MICU1 and MICU2 finely tune the mitochondrial Ca2+ uniporter by exerting opposite effects on MCU activity. Mol. Cell 2014, 53, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csordás, G.; Golenár, T.; Seifert, E.L.; Kamer, K.J.; Sancak, Y.; Perocchi, F.; Moffat, C.; Weaver, D.; de la Fuente Perez, S.; Bogorad, R.; et al. MICU1 controls both the threshold and cooperative activation of the mitochondrial Ca2+ uniporter. Cell Metab. 2013, 17, 976–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yang, X.; Li, S.; Wang, Z.; Liu, Y.; Feng, J.; Zhu, Y.; Shen, Y. Structural and mechanistic insights into MICU1 regulation of mitochondrial calcium uptake. EMBO J. 2014, 33, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Vais, H.; Payne, R.; Paudel, U.; Li, C.; Foskett, J.K. Coupled transmembrane mechanisms control MCU-mediated mitochondrial Ca2+ uptake. Proc. Natl. Acad. Sci. USA 2020, 117, 21731–21739. [Google Scholar] [CrossRef]
- Paillard, M.; Csordás, G.; Szanda, G.; Golenár, T.; Debattisti, V.; Bartok, A.; Wang, N.; Moffat, C.; Seifert, E.L.; Spät, A.; et al. Tissue-specific mitochondrial decoding of cytoplasmic Ca2+ signals is controlled by the stoichiometry of MICU1/2 and MCU. Cell Rep. 2017, 18, 2291–2300. [Google Scholar] [CrossRef] [Green Version]
- Plovanich, M.; Bogorad, R.L.; Sancak, Y.; Kamer, K.J.; Strittmatter, L.; Li, A.A.; Girgis, H.S.; Kuchimanchi, S.; De Groot, J.; Speciner, L.; et al. MICU2, a paralog of MICU1, resides within the mitochondrial uniporter complex to regulate calcium handling. PLoS ONE 2013, 8, e55785. [Google Scholar] [CrossRef] [Green Version]
- Kamer, K.J.; Grabarek, Z.; Mootha, V.K. High-affinity cooperative Ca2+ binding by MICU 1– MICU 2 serves as an on–off switch for the uniporter. EMBO Rep. 2017, 18, 1397–1411. [Google Scholar] [CrossRef]
- Payne, R.; Hoff, H.; Roskowski, A.; Foskett, J.K. MICU2 restricts spatial crosstalk between InsP 3 R and MCU channels by regulating threshold and gain of MICU1-mediated inhibition and activation of MCU. Cell Rep. 2017, 21, 3141–3154. [Google Scholar] [CrossRef] [Green Version]
- Mallilankaraman, K.; Cardenas, C.; Doonan, P.J.; Chandramoorthy, H.C.; Irrinki, K.M.; Golenar, T.; Csordas, G.; Madireddi, P.; Yang, J.; Muller, M.; et al. MCUR1 is an essential component of mitochondrial Ca2+ uptake that regulates cellular metabolism. Nat. Cell Biol. 2012, 14, 1336–1343. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Wang, J.; Zhang, H.; Yuan, P.; Zhu, J.; Wu, Y.; Huang, Q.; Guo, X.; Zhang, J.; Ji, L.; et al. MCUR1-mediated mitochondrial calcium signaling facilitates cell survival of hepatocellular carcinoma via reactive oxygen species-dependent P53 degradation. Antioxid. Redox Signal. 2018, 28, 1120–1136. [Google Scholar] [CrossRef]
- Tomar, D.; Dong, Z.; Shanmughapriya, S.; Koch, D.A.; Thomas, T.; Hoffman, N.E.; Timbalia, S.A.; Goldman, S.J.; Breves, S.L.; Corbally, D.P.; et al. MCUR1 is a scaffold factor for the MCU complex function and promotes mitochondrial bioenergetics. Cell Rep. 2016, 15, 1673–1685. [Google Scholar] [CrossRef]
- Paupe, V.; Prudent, J.; Dassa, E.P.; Rendon, O.Z.; Shoubridge, E.A. CCDC90A (MCUR1) is a cytochrome c oxidase assembly factor and not a regulator of the mitochondrial calcium uniporter. Cell Metab. 2015, 21, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassi, M.T.; Manzoni, M.; Bresciani, R.; Pizzo, M.T.; Della Monica, A.; Barlati, S.; Monti, E.; Borsani, G. Cellular expression and alternative splicing of SLC25A23, a member of the mitochondrial Ca2+-dependent solute carrier gene family. Gene 2005, 345, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Harborne, S.P.D.; King, M.S.; Crichton, P.G.; Kunji, E.R.S. Calcium regulation of the human mitochondrial ATP-Mg/Pi carrier SLC25A24 uses a locking pin mechanism. Sci. Rep. 2017, 7, 45383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunter, T.E.; Gunter, K.K. Uptake of calcium by mitochondria: Transport and possible function. IUBMB Life 2001, 52, 197–204. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, D.; He, X.; Huang, Y.; Shao, H. Transport of calcium ions into mitochondria. Curr. Genom. 2016, 17, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Sparagna, G.C.; Gunter, K.K.; Sheu, S.-S.; Gunter, T.E. Mitochondrial Calcium uptake from physiological-type pulses of Calcium. J. Biol. Chem. 1995, 270, 27510–27515. [Google Scholar] [CrossRef] [Green Version]
- Beutner, G.; Sharma, V.K.; Giovannucci, D.R.; Yule, D.I.; Sheu, S.S. Identification of a Ryanodine receptor in Rat Heart mitochondria. J. Biol. Chem. 2001, 276, 21482–21488. [Google Scholar] [CrossRef] [Green Version]
- Beutner, G.; Sharma, V.K.; Lin, L.; Ryu, S.Y.; Dirksen, R.T.; Sheu, S.S. Type 1 Ryanodine receptor in cardiac mitochondria: Transducer of excitation–metabolism coupling. Biochim. Biophys. Acta Biomembr. 2005, 1717, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Babich, L.G.; Shlykov, S.G.; Kosterin, S.O. Ca ion transport in smooth muscle mitochondria. Ukr. Biochem. J. 2014, 86, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Altschafl, B.A.; Beutner, G.; Sharma, V.K.; Sheu, S.S.; Valdivia, H.H. The mitochondrial ryanodine receptor in rat heart: A pharmaco-kinetic profile. Biochim. Biophys. Acta Biomembr. 2007, 1768, 1784–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamai, S.; Iida, H.; Yokota, S.; Sayano, T.; Kiguchiya, S.; Ishihara, N.; Hayashi, J.-I.; Mihara, K.; Oka, T. Characterization of the mitochondrial protein LETM1, which maintains the mitochondrial tubular shapes and interacts with the AAA-ATPase BCS1L. J. Cell Sci. 2008, 121, 2588–2600. [Google Scholar] [CrossRef] [Green Version]
- Endele, S.; Fuhry, M.; Pak, S.J.; Zabel, B.U.; Winterpacht, A. LETM1, a novel gene encoding a putative EF-hand Ca(2+)-binding protein, flanks the Wolf-Hirschhorn syndrome (WHS) critical region and is deleted in most WHS patients. Genomics 1999, 60, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; van der Bliek, A.M. Inverse correlation between expression of the Wolfs Hirschhorn candidate gene Letm1 and mitochondrial volume in C. elegans and in mammalian cells. Hum. Mol. Genet. 2007, 16, 2061–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlickum, S.; Moghekar, A.; Simpson, J.C.; Steglich, C.; O’Brien, R.J.; Winterpacht, A.; Endele, S.U. LETM1, a gene deleted in Wolf-Hirschhorn syndrome, encodes an evolutionarily conserved mitochondrial protein. Genomics 2004, 83, 254–261. [Google Scholar] [CrossRef]
- Lin, Q.T.; Stathopulos, P.B. Molecular mechanisms of leucine zipper EF-Hand containing transmembrane Protein-1 function in health and disease. Int. J. Mol. Sci. 2019, 20, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doonan, P.J.; Chandramoorthy, H.C.; Hoffman, N.E.; Zhang, X.; Cardenas, C.; Shanmughapriya, S.; Rajan, S.; Vallem, S.; Chen, X.; Foskett, J.K.; et al. LETM1-dependent mitochondrial Ca2+ flux modulates cellular bioenergetics and proliferation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 4936–4949. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Zhao, L.; Clish, C.B.; Clapham, D.E. Letm1, the mitochondrial Ca2+/H+ antiporter, is essential for normal glucose metabolism and alters brain function in Wolf-Hirschhorn syndrome. Proc. Natl. Acad. Sci. USA 2013, 110, E2249–E2254. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Zhao, L.; Clapham, D.E. Genome-wide RNAi screen identifies Letm1 as a mitochondrial Ca2+/H+ antiporter. Science 2009, 326, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Matsushita, S.; Kato, Y.; Watanabe, H.; Matsui, A.; Oka, T.; Matsuura, T. In vitro synthesis of the human calcium transporter Letm1 within cell-sized liposomes and investigation of its lipid dependency. J. Biosci. Bioeng. 2019, 127, 544–548. [Google Scholar] [CrossRef]
- Nowikovsky, K.; Bernardi, P. LETM1 in mitochondrial cation transport. Front. Physiol. 2014, 5, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marchi, U.; Santo-Domingo, J.; Castelbou, C.; Sekler, I.; Wiederkehr, A.; Demaurex, N. NCLX protein, but not LETM1, mediates mitochondrial Ca2+ extrusion, thereby limiting Ca2+-induced NAD(P)H production and modulating matrix redox state. J. Biol. Chem. 2014, 289, 20377–20385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carafoli, E.; Tiozzo, R.; Lugli, G.; Crovetti, F.; Kratzing, C. The release of calcium from heart mitochondria by sodium. J. Mol. Cell. Cardiol. 1974, 6, 361–371. [Google Scholar] [CrossRef]
- Wingrove, D.E.; Gunter, T.E. Kinetics of mitochondrial calcium transport. I. Characteristics of the sodium-independent calcium efflux mechanism of liver mitochondria. J. Biol. Chem. 1986, 261, 15159–15165. [Google Scholar]
- Wingrove, D.E.; Gunter, T.E. Kinetics of mitochondrial calcium transport. II. A kinetic description of the sodium-dependent calcium efflux mechanism of liver mitochondria and inhibition by ruthenium red and by tetraphenylphosphonium. J. Biol. Chem. 1986, 261, 15166–15171. [Google Scholar]
- Hunter, D.R.; Haworth, R.A.; Hunter, D.R.; Haworth, R.A. The Ca2+-induced membrane transition in mitochondria. Arch. Biochem. Biophys. 1979, 195, 468–477. [Google Scholar] [CrossRef]
- Tsai, M.F.; Jiang, D.; Zhao, L.; Clapham, D.; Miller, C. Functional reconstitution of the mitochondrial Ca2+/H+ antiporter Letm1. J. Gen. Physiol. 2014, 143, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Nowikovsky, K.; Pozzan, T.; Rizzuto, R.; Scorrano, L.; Bernardi, P. The pathophysiology of LETM1. J. Gen. Physiol. 2012, 139, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Palty, R.; Silverman, W.F.; Hershfinkel, M.; Caporale, T.; Sensi, S.L.; Parnis, J.; Nolte, C.; Fishman, D.; Shoshan-Barmatz, V.; Herrmann, S.; et al. NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc. Natl. Acad. Sci. USA 2010, 107, 436–441. [Google Scholar] [CrossRef] [Green Version]
- Luongo, T.S.; Lambert, J.P.; Gross, P.; Nwokedi, M.; Lombardi, A.A.; Shanmughapriya, S.; Carpenter, A.C.; Kolmetzky, D.; Gao, E.; van Berlo, J.H.; et al. The mitochondrial Na(+)/Ca(2+) exchanger is essential for Ca(2+) homeostasis and viability. Nature 2017, 545, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Kostic, M.; Sekler, I. Functional properties and mode of regulation of the mitochondrial Na+/Ca2+ exchanger, NCLX. Semin. Cell Dev. Biol. 2019, 94, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Palty, R.; Ohana, E.; Hershfinkel, M.; Volokita, M.; Elgazar, V.; Beharier, O.; Silverman, W.F.; Argaman, M.; Sekler, I. Lithium-calcium exchange is mediated by a distinct potassium-independent sodium-calcium exchanger. J. Biol. Chem. 2004, 279, 25234–25240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.-Z.; Prinsen, C.F.M.; Clark, R.B.; Giles, W.R.; Schnetkamp, P.P.M. Na+-Ca2+-K+ currents measured in insect cells transfected with the retinal cone or Rod Na+-Ca2+-K+ exchanger cDNA. Biophys. J. 2000, 79, 1945–1953. [Google Scholar] [CrossRef] [Green Version]
- Gunter, T.E.; Yule, D.I.; Gunter, K.K.; Eliseev, R.A.; Salter, J.D. Calcium and mitochondria. FEBS Lett. 2004, 567, 96–102. [Google Scholar] [CrossRef]
- Takeuchi, A.; Kim, B.; Matsuoka, S. The destiny of Ca2+ released by mitochondria. J. Physiol. Sci. 2015, 65, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Haworth, R.A.; Hunter, D.R.; Berkoff, H.A. Na+ releases Ca2+ from liver, kidney and lung mitochondria. FEBS Lett. 1980, 110, 216–218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lipton, P. Cytosolic Ca2+ changes during in vitro ischemia in rat hippocampal slices: Major roles for glutamate and Na+-dependent Ca2+ release from mitochondria. J. Neurosci. 1999, 19, 3307–3315. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Takeuchi, A.; Matsuoka, S. Membrane current evoked by mitochondrial Na+–Ca2+ exchange in mouse heart. J. Physiol. Sci. 2020, 70, 24. [Google Scholar] [CrossRef]
- Samanta, K.; Mirams, G.R.; Parekh, A.B. Sequential forward and reverse transport of the Na+ Ca2+ exchanger generates Ca2+ oscillations within mitochondria. Nat. Commun. 2018, 9, 156. [Google Scholar] [CrossRef]
- Kolomiets, O.V.; Danylovych, Y.V.; Danylovych, H.V.; Kosterin, S.O. Ca(2+)/H(+)-exchange in myometrium mitochondria. Ukr. Biochem. J. 2014, 86, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Kandaurova, N.V. Ca2+-Induced Changes of Membrane Potential of Myometrium Mitochondria. Ph.D. Thesis, Palladin Institute of Biochemistry, National Academy of Sciences, Kiev, Ukraine, 2011. [Google Scholar]
- Gunter, K.K.; Zuscik, M.J.; Gunter, T.E. The Na(+)-independent Ca2+ efflux mechanism of liver mitochondria is not a passive Ca2+/2H+ exchanger. J. Biol. Chem. 1991, 266, 21640–21648. [Google Scholar]
- Huang, E.; Qu, D.; Huang, T.; Rizzi, N.; Boonying, W.; Krolak, D.; Ciana, P.; Woulfe, J.; Klein, C.; Slack, R.S.; et al. PINK1-mediated phosphorylation of LETM1 regulates mitochondrial calcium transport and protects neurons against mitochondrial stress. Nat. Commun. 2017, 8, 1399. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.J.; Quintanilla, R.A. Development or disease: Duality of the mitochondrial permeability transition pore. Dev. Biol. 2017, 426, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Britti, E.; Delaspre, F.; Tamarit, J.; Ros, J. Mitochondrial calcium signalling and neurodegenerative diseases. Neuronal Signal. 2018, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasutto, L.; Azzolini, M.; Szabò, I.; Zoratti, M. The mitochondrial permeability transition pore in AD 2016: An update. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2515–2530. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, J.; Wu, R.; Bai, J.; Hou, Y.; Zeng, Y.; Zhang, Y.; Wang, X.; Wang, Z.; Meng, X. Mitochondrial MPTP: A novel target of ethnomedicine for stroke treatment by apoptosis inhibition. Front. Pharmacol. 2020, 11, 352. [Google Scholar] [CrossRef]
- Hurst, S.; Hoek, J.; Sheu, S.S. Mitochondrial Ca2+ and regulation of the permeability transition pore. J. Bioenergy Biomembr. 2017, 49, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Altschuld, R.A.; Hohl, C.M.; Castillo, L.C.; Garleb, A.A.; Starling, R.C.; Brierley, G.P. Cyclosporin inhibits mitochondrial calcium efflux in isolated adult rat ventricular cardiomyocytes. Am. J. Physiol. Circ. Physiol. 1992, 262, H1699–H1704. [Google Scholar] [CrossRef]
- Mnatsakanyan, N.; Beutner, G.; Porter, G.A.; Alavian, K.N.; Jonas, E.A. Physiological roles of the mitochondrial permeability transition pore. J. Bioenergy Biomembr. 2017, 49, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Basso, E.; Fante, L.; Fowlkes, J.; Petronilli, V.; Forte, M.A.; Bernardi, P. Properties of the permeability transition pore in mitochondria devoid of Cyclophilin, D. J. Biol. Chem. 2005, 280, 18558–18561. [Google Scholar] [CrossRef] [Green Version]
- Shanmughapriya, S.; Rajan, S.; Hoffman, N.E.; Higgins, A.M.; Tomar, D.; Nemani, N.; Hines, K.J.; Smith, D.J.; Eguchi, A.; Vallem, S.; et al. SPG7 is an essential and conserved component of the mitochondrial permeability transition pore. Mol. Cell 2015, 60, 47–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baines, C.P.; Kaiser, R.A.; Purcell, N.H.; Blair, N.S.; Osinska, H.; Hambleton, M.A.; Brunskill, E.W.; Sayen, M.R.; Gottlieb, R.A.; Dorn, G.W.; et al. Loss of Cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature 2005, 434, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Shimizu, S.; Watanabe, T.; Yamaguchi, O.; Otsu, K.; Yamagata, H.; Inohara, H.; Kubo, T.; Tsujimoto, Y. Cyclophilin D-Dependent mitochondrial permeability transition regulates some necrotic but not apoptotic cell death. Nature 2005, 434, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Schinzel, A.C.; Takeuchi, O.; Huang, Z.; Fisher, J.K.; Zhou, Z.; Rubens, J.; Hetz, C.; Danial, N.N.; Moskowitz, M.A.; Korsmeyer, S.J. Cyclophilin D is a component of mitochondrial permeability transition and mediates neuronal cell death after focal cerebral ischemia. Proc. Natl. Acad. Sci. USA 2005, 102, 12005–12010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.R.; Baetz, D.; Ovize, M. Cyclophilin D and myocardial ischemia–reperfusion injury: A fresh perspective. J. Mol. Cell. Cardiol. 2015, 78, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Kokoszka, J.E.; Waymire, K.G.; Levy, S.E.; Sligh, J.E.; Cai, J.; Jones, D.P.; MacGregor, G.R.; Wallace, D.C. The ADP/ATP translocator is not essential for the mitochondrial permeability transition pore. Nature 2004, 427, 461–465. [Google Scholar] [CrossRef]
- Karch, J.; Bround, M.J.; Khalil, H.; Sargent, M.A.; Latchman, N.; Terada, N.; Peixoto, P.M.; Molkentin, J.D. Inhibition of mitochondrial permeability transition by deletion of the ANT family and CypD. Sci. Adv. 2019, 5, eaaw4597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bround, M.J.; Bers, D.M.; Molkentin, J.D. A 20/20 view of ANT function in mitochondrial biology and necrotic cell death. J. Mol. Cell. Cardiol. 2020, 144, A3–A13. [Google Scholar] [CrossRef]
- Kwong, J.Q.; Davis, J.; Baines, C.P.; Sargent, M.A.; Karch, J.; Wang, X.; Huang, T.; Molkentin, J.D. Genetic deletion of the mitochondrial phosphate carrier desensitizes the mitochondrial permeability transition pore and causes cardiomyopathy. Cell Death Differ. 2014, 21, 1209–1217. [Google Scholar] [CrossRef] [Green Version]
- Varanyuwatana, P.; Halestrap, A.P. The roles of phosphate and the phosphate carrier in the mitochondrial permeability transition pore. Mitochondrion 2012, 12, 120–125. [Google Scholar] [CrossRef]
- Hurst, S.; Baggett, A.; Csordas, G.; Sheu, S.-S. SPG7 targets the m-AAA protease complex to process MCU for uniporter assembly, Ca2+ influx, and regulation of mitochondrial permeability transition pore opening. J. Biol. Chem. 2019, 294, 10807–10818. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.W.C.; Varanyuwatana, P.; Halestrap, A.P. The mitochondrial phosphate carrier interacts with Cyclophilin D and may play a key role in the permeability transition. J. Biol. Chem. 2008, 283, 26312–26323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgio, V.; von Stockum, S.; Antoniel, M.; Fabbro, A.; Fogolari, F.; Forte, M.; Glick, G.D.; Petronilli, V.; Zoratti, M.; Szabo, I.; et al. Dimers of mitochondrial ATP synthase form the permeability transition pore. Proc. Natl. Acad. Sci. USA 2013, 110, 5887–5892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonora, M.; Bononi, A.; De Marchi, E.; Giorgi, C.; Lebiedzinska, M.; Marchi, S.; Patergnani, S.; Rimessi, A.; Suski, J.M.; Wojtala, A.; et al. Role of the c subunit of the F O ATP synthase in mitochondrial permeability transition. Cell Cycle 2013, 12, 674–683. [Google Scholar] [CrossRef] [Green Version]
- Crompton, M.; Costi, A. Kinetic evidence for a heart mitochondrial pore activated by Ca2+, inorganic phosphate and oxidative stress. A potential mechanism for mitochondrial dysfunction during cellular Ca2+ overload. Eur. J. Biochem. 1988, 178, 489–501. [Google Scholar] [CrossRef]
- Carraro, M.; Giorgio, V.; Šileikytė, J.; Sartori, G.; Forte, M.; Lippe, G.; Zoratti, M.; Szabò, I.; Bernardi, P. Channel formation by yeast F-ATP synthase and the role of dimerization in the mitochondrial permeability transition. J. Biol. Chem. 2014, 289, 15980–15985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Marinelli, F.; Nief, C.; Faraldo-Gómez, J.D. Atomistic simulations indicate the c-subunit ring of the F1Fo ATP synthase is not the mitochondrial permeability transition pore. Elife 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Ford, H.C.; Carroll, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Persistence of the mitochondrial permeability transition in the absence of subunit c of human ATP synthase. Proc. Natl. Acad. Sci. USA 2017, 114, 3409–3414. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Carroll, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Permeability transition in human mitochondria persists in the absence of peripheral stalk subunits of ATP synthase. Proc. Natl. Acad. Sci. USA 2017, 114, 9086–9091. [Google Scholar] [CrossRef] [Green Version]
- Bonora, M.; Wieckowski, M.R.; Chinopoulos, C.; Kepp, O.; Kroemer, G.; Galluzzi, L.; Pinton, P. Molecular mechanisms of cell death: Central implication of ATP synthase in mitochondrial permeability transition. Oncogene 2015, 34, 1475–1486. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Richardson, A.P. The mitochondrial permeability transition: A current perspective on its identity and role in ischaemia/reperfusion injury. J. Mol. Cell. Cardiol. 2015, 78, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, P. The mitochondrial permeability transition pore: A mystery solved? Front. Physiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, E.A.; Porter, G.A.; Beutner, G.; Mnatsakanyan, N.; Alavian, K.N. Cell death disguised: The mitochondrial permeability transition pore as the c-subunit of the F1FO ATP synthase. Pharmacol. Res. 2015, 99, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elustondo, P.A.; Nichols, M.; Negoda, A.; Thirumaran, A.; Zakharian, E.; Robertson, G.S.; Pavlov, E. V Mitochondrial permeability transition pore induction is linked to formation of the complex of ATPase C-subunit, polyhydroxybutyrate and inorganic polyphosphate. Cell Death Discov. 2016, 2, 16070. [Google Scholar] [CrossRef]
- Alavian, K.N.; Beutner, G.; Lazrove, E.; Sacchetti, S.; Park, H.-A.; Licznerski, P.; Li, H.; Nabili, P.; Hockensmith, K.; Graham, M.; et al. An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore. Proc. Natl. Acad. Sci. USA 2014, 111, 10580–10585. [Google Scholar] [CrossRef] [Green Version]
- Chinopoulos, C. Mitochondrial permeability transition pore: Back to the drawing board. Neurochem. Int. 2018, 117, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Ichas, F.; Jouaville, L.S.; Mazat, J.P. Mitochondria are excitable organelles capable of generating and conveying electrical and calcium signals. Cell 1997, 89, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Kwong, J.Q.; Molkentin, J.D.; Bers, D.M. Individual cardiac mitochondria undergo rare transient permeability transition pore openings. Circ. Res. 2016, 118, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Ichas, F.; Mazat, J.-P. From calcium signaling to cell death: Two conformations for the mitochondrial permeability transition pore. Switching from low- to high-conductance state. Biochim. Biophys. Acta Bioenergy 1998, 1366, 33–50. [Google Scholar] [CrossRef] [Green Version]
- Gainutdinov, T.; Molkentin, J.D.; Siemen, D.; Ziemer, M.; Debska-Vielhaber, G.; Vielhaber, S.; Gizatullina, Z.; Orynbayeva, Z.; Gellerich, F.N. Knockout of cyclophilin D in Ppif−/− mice increases stability of brain mitochondria against Ca2+ stress. Arch. Biochem. Biophys. 2015, 579, 40–46. [Google Scholar] [CrossRef]
- Bernardi, P.; von Stockum, S. The permeability transition pore as a Ca2+ release channel: New answers to an old question. Cell Calcium 2012, 52, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korge, P.; Yang, L.; Yang, J.-H.; Wang, Y.; Qu, Z.; Weiss, J.N. Protective role of transient pore openings in calcium handling by cardiac mitochondria. J. Biol. Chem. 2011, 286, 34851–34857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrod, J.W.; Wong, R.; Mishra, S.; Vagnozzi, R.J.; Sakthievel, B.; Goonasekera, S.A.; Karch, J.; Gabel, S.; Farber, J.; Force, T.; et al. Cyclophilin D controls mitochondrial pore–dependent Ca2+ exchange, metabolic flexibility, and propensity for heart failure in mice. J. Clin. Investig. 2010, 120, 3680–3687. [Google Scholar] [CrossRef]
- Lamb, H.M. Double agents of cell death: Novel emerging functions of apoptotic regulators. FEBS J. 2020, 287, 2647–2663. [Google Scholar] [CrossRef] [Green Version]
- Baines, C.P.; Gutiérrez-Aguilar, M. The still uncertain identity of the channel-forming unit(s) of the mitochondrial permeability transition pore. Cell Calcium 2018, 73, 121–130. [Google Scholar] [CrossRef]
- Zorow, D.B.; Kinnally, K.W.; Perini, S.; Tedeschi, H. Multiple conductance levels in rat heart inner mitochondrial membranes studied by patch clamping. Biochim. Biophys. Acta Biomembr. 1992, 1105, 263–270. [Google Scholar] [CrossRef]
- Petronilli, V.; Miotto, G.; Canton, M.; Brini, M.; Colonna, R.; Bernardi, P.; Di Lisa, F. Transient and long-lasting openings of the mitochondrial permeability transition pore can be monitored directly in intact cells by changes in mitochondrial calcein fluorescence. Biophys. J. 1999, 76, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Cui, S.; Zhang, Y.; Ren, J. Mitochondrial Ca2+ regulation in the etiology of heart failure: Physiological and pathophysiological implications. Acta Pharmacol. Sin. 2020, 10, 1301–1309. [Google Scholar] [CrossRef]
- Kinnally, K.W.; Peixoto, P.M.; Ryu, S.-Y.; Dejean, L.M. Is mPTP the gatekeeper for necrosis, apoptosis, or both? Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 616–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, P.; Rasola, A.; Forte, M.; Lippe, G. The mitochondrial permeability transition pore: Channel formation by F-ATP synthase, integration in signal transduction, and role in pathophysiology. Physiol. Rev. 2015, 95, 1111–1155. [Google Scholar] [CrossRef]
- Elrod, J.W.; Molkentin, J.D. Physiologic functions of Cyclophilin D and the mitochondrial permeability transition pore. Circ. J. 2013, 77, 1111–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halestrap, A. Mitochondrial permeability transition pore opening during myocardial reperfusion—A target for cardioprotection. Cardiovasc. Res. 2004, 61, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Szabó, I.; Zoratti, M. The mitochondrial permeability transition pore may comprise VDAC molecules. FEBS Lett. 1993, 330, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Crompton, M.; Virji, S.; Ward, J.M. Cyclophilin-D binds strongly to complexes of the voltage-dependent anion channel and the adenine nucleotide translocase to form the permeability transition pore. Eur. J. Biochem. 1998, 258, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Shi, Y.; Tian, C.; Jiang, C.; Jin, H.; Chen, J.; Almasan, A.; Tang, H.; Chen, Q. Essential role of the voltage-dependent anion channel (VDAC) in mitochondrial permeability transition pore opening and cytochrome c release induced by arsenic trioxide. Oncogene 2004, 23, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, A.D.; Choi, D.C.; Kabaria, S.; Tran, A.; Junn, E. MicroRNA-7 regulates the function of mitochondrial permeability transition pore by targeting VDAC1 expression. J. Biol. Chem. 2016, 291, 6483–6493. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Hu, S.; Jin, Q.; Shi, C.; Zhang, Y.; Zhu, P.; Ma, Q.; Tian, F.; Chen, Y. Mff-dependent mitochondrial fission contributes to the pathogenesis of cardiac microvasculature ischemia/reperfusion injury via induction of mROS-mediated cardiolipin oxidation and HK2/VDAC1 disassociation-involved mPTP opening. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Colombini, M. VDAC closure increases calcium ion flux. Biochim. Biophys. Acta Biomembr. 2007, 1768, 2510–2515. [Google Scholar] [CrossRef] [Green Version]
- Tikunov, A.; Johnson, C.B.; Pediaditakis, P.; Markevich, N.; Macdonald, J.M.; Lemasters, J.J.; Holmuhamedov, E. Closure of VDAC causes oxidative stress and accelerates the Ca2+-induced mitochondrial permeability transition in rat liver mitochondria. Arch. Biochem. Biophys. 2010, 495, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Glab, J.A.; Cao, Z.; Puthalakath, H. Bcl-2 family proteins, beyond the veil. Int. Rev. Cell Mol. Biol. 2020, 351, 1–22. [Google Scholar]
- Insoluble Proteins. In Methods in Molecular Biology; García-Fruitós, E. (Ed.) Springer: New York, NY, USA, 2015; Volume 1258, ISBN 978-1-4939-2204-8. [Google Scholar]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Hardwick, J.M.; Soane, L. Multiple functions of BCL-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, M.L.; Gama, V. A connection in life and death: The BCL-2 family coordinates mitochondrial network dynamics and stem cell fate. Int. Rev. Cell Mol. Biol. 2020, 353, 255–284. [Google Scholar] [PubMed]
- Adams, J.M. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef]
- Choudhury, S. A comparative analysis of BCL-2 family. Bioinformation 2019, 15, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, H.; Wagner, L.E.; Tanimura, A.; Vandermarliere, E.; Luyten, T.; Welkenhuyzen, K.; Alzayady, K.J.; Wang, L.; Hamada, K.; Mikoshiba, K.; et al. Bcl-2 and IP3 compete for the ligand-binding domain of IP3Rs modulating Ca2+ signaling output. Cell. Mol. Life Sci. 2019, 76, 3843–3859. [Google Scholar] [CrossRef]
- Vervliet, T.; Parys, J.B.; Bultynck, G. Bcl-2 proteins and calcium signaling: Complexity beneath the surface. Oncogene 2016, 35, 5079–5092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouqué, A.; Lepvrier, E.; Debure, L.; Gouriou, Y.; Malleter, M.; Delcroix, V.; Ovize, M.; Ducret, T.; Li, C.; Hammadi, M.; et al. The apoptotic members CD95, BclxL, and Bcl-2 cooperate to promote cell migration by inducing Ca2+ flux from the endoplasmic reticulum to mitochondria. Cell Death Differ. 2016, 23, 1702–1716. [Google Scholar] [CrossRef] [Green Version]
- Lanave, C.; Santamaria, M.; Saccone, C. Comparative genomics: The evolutionary history of the Bcl-2 family. Gene 2004, 333, 71–79. [Google Scholar] [CrossRef]
- Zmasek, C.M.; Godzik, A. Evolution of the animal apoptosis network. Cold Spring Harb. Perspect. Biol. 2013, 5, a008649. [Google Scholar] [CrossRef] [Green Version]
- Kvansakul, M.; Caria, S.; Hinds, M. The Bcl-2 Family in host-virus interactions. Viruses 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aouacheria, A.; Rech de Laval, V.; Combet, C.; Hardwick, J.M. Evolution of Bcl-2 homology motifs: Homology versus homoplasy. Trends Cell Biol. 2013, 23, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Finger, L.; Yunis, J.; Nowell, P.; Croce, C. Cloning of the chromosome breakpoint of neoplastic B cells with the t(14;18) chromosome translocation. Science 1984, 226, 1097–1099. [Google Scholar] [CrossRef]
- Cleary, M.L.; Smith, S.D.; Sklar, J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t(14;18) translocation. Cell 1986, 47, 19–28. [Google Scholar] [CrossRef]
- Vaux, D.L.; Cory, S.; Adams, J.M. Bcl-2 gene promotes haemopoietic cell survival and cooperates with c-myc to immortalize pre-B cells. Nature 1988, 335, 440–442. [Google Scholar] [CrossRef]
- Birkinshaw, R.W.; Czabotar, P.E. The BCL-2 family of proteins and mitochondrial outer membrane permeabilisation. Semin. Cell Dev. Biol. 2017, 72, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Huska, J.D.; Lamb, H.M.; Hardwick, J.M. Overview of BCL-2 family proteins and therapeutic potentials. Methods Mol. Biol. 2019, 1877, 1–21. [Google Scholar]
- Delbridge, A.R.D.; Strasser, A. The BCL-2 protein family, BH3-mimetics and cancer therapy. Cell Death Differ. 2015, 22, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.M. BAX and BAK become killers without a BH3 trigger. Cell Res. 2019, 29, 967–968. [Google Scholar] [CrossRef]
- Vervloessem, T.; Kerkhofs, M.; La Rovere, R.M.; Sneyers, F.; Parys, J.B.; Bultynck, G. Bcl-2 inhibitors as anti-cancer therapeutics: The impact of and on calcium signaling. Cell Calcium 2018, 70, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Gavathiotis, E. (Ed.) BCL-2 Family Proteins-Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1877, ISBN 978-1-4939-8860-0. [Google Scholar]
- Chipuk, J.E.; Moldoveanu, T.; Llambi, F.; Parsons, M.J.; Green, D.R. The BCL-2 family reunion. Mol. Cell 2010, 37, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.; Lee, S.-H.; Meng, X.W.; Vincelette, N.D.; Knorr, K.L.B.; Ding, H.; Nowakowski, G.S.; Dai, H.; Kaufmann, S.H. Emerging understanding of Bcl-2 biology: Implications for neoplastic progression and treatment. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 1658–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Voss, A.K.; Strasser, A. The essentials of developmental apoptosis. F1000Research 2020, 9, 148. [Google Scholar] [CrossRef]
- Suvarna, V.; Singh, V.; Murahari, M. Current overview on the clinical update of Bcl-2 anti-apoptotic inhibitors for cancer therapy. Eur. J. Pharmacol. 2019, 862, 172655. [Google Scholar] [CrossRef]
- Luna-Vargas, M.P.A.; Chipuk, J.E. Physiological and pharmacological control of BAK, BAX, and beyond. Trends Cell Biol. 2016, 26, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.J.; Tait, S.W.G. Targeting BCL-2 regulated apoptosis in cancer. Open Biol. 2018, 8, 180002. [Google Scholar] [CrossRef]
- Gabellini, C.; Trisciuoglio, D.; Del Bufalo, D. Non-canonical roles of Bcl-2 and Bcl-xL proteins: Relevance of BH4 domain. Carcinogenesis 2017, 38, 579–587. [Google Scholar] [CrossRef]
- Senichkin, V.V.; Kopeina, G.S.; Prokhorova, E.A.; Zamaraev, A.V.; Lavrik, I.N.; Zhivotovsky, B. Modulation of Mcl-1 transcription by serum deprivation sensitizes cancer cells to cisplatin. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 557–566. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certo, M.; Moore, V.D.G.; Nishino, M.; Wei, G.; Korsmeyer, S.; Armstrong, S.A.; Letai, A. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 2006, 9, 351–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, B.; Liang, C.; Jung, J.U.; Oh, B.H. Evidence that inhibition of BAX activation by BCL-2 involves its tight and preferential interaction with the BH3 domain of BAX. Cell Res. 2011, 21, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Llambi, F.; Wang, Y.-M.; Victor, B.; Yang, M.; Schneider, D.M.; Gingras, S.; Parsons, M.J.; Zheng, J.H.; Brown, S.A.; Pelletier, S.; et al. BOK is a non-canonical BCL-2 family effector of apoptosis regulated by ER-associated degradation. Cell 2016, 165, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, F.F.S.; Vanyai, H.K.; Cowan, A.D.; Delbridge, A.R.D.; Whitehead, L.; Grabow, S.; Czabotar, P.E.; Voss, A.K.; Strasser, A. Embryogenesis and adult life in the absence of intrinsic apoptosis effectors BAX, BAK, and BOK. Cell 2018, 173, 1217–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhaili, S.H.; Karimian, H.; Stellato, M.; Lee, T.H.; Aguilar, M.I. Mitochondrial outer membrane permeabilization: A focus on the role of mitochondrial membrane structural organization. Biophys. Rev. 2017, 9, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Oltval, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programed cell death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Youle, R.J. Bax in murine thymus is a soluble monomeric protein that displays differential detergent-induced conformations. J. Biol. Chem. 1998, 273, 10777–10783. [Google Scholar] [CrossRef] [Green Version]
- Schellenberg, B.; Wang, P.; Keeble, J.A.; Rodriguez-Enriquez, R.; Walker, S.; Owens, T.W.; Foster, F.; Tanianis-Hughes, J.; Brennan, K.; Streuli, C.H.; et al. Bax exists in a dynamic equilibrium between the cytosol and mitochondria to control apoptotic priming. Mol. Cell 2013, 49, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Youle, R.J.; Tjandra, N. Structure of bax. Cell 2000, 103, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Ke, F.; Voss, A.; Kerr, J.B.; O’Reilly, L.A.; Tai, L.; Echeverry, N.; Bouillet, P.; Strasser, A.; Kaufmann, T. BCL-2 family member BOK is widely expressed but its loss has only minimal impact in mice. Cell Death Differ. 2012, 19, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Echeverry, N.; Bachmann, D.; Ke, F.; Strasser, A.; Simon, H.U.; Kaufmann, T. Intracellular localization of the BCL-2 family member BOK and functional implications. Cell Death Differ. 2013, 20, 785–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, S.; Low, I.C.C.; Pervaiz, S. Regulation of mitochondrial metabolism: Yet another facet in the biology of the oncoprotein Bcl-2. Biochem. J. 2011, 435, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Villunger, A. p53- and drug-induced apoptotic responses mediated by BH3-only proteins puma and noxa. Science 2003, 302, 1036–1038. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.J.; Li, H.; Salvesen, G.S.; Yuan, J.; Wagner, G. Solution structure of BID, an intracellular amplifier of apoptotic signaling. Cell 1999, 96, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Billen, L.P.; Shamas-Din, A.; Andrews, D.W. Bid: A Bax-like BH3 protein. Oncogene 2008, 27, S93–S104. [Google Scholar] [CrossRef] [Green Version]
- Edlich, F. BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem. Biophys. Res. Commun. 2018, 500, 26–34. [Google Scholar] [CrossRef]
- Shamas-Din, A.; Kale, J.; Leber, B.; Andrews, D.W. Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008714. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef]
- Moldoveanu, T.; Grace, C.R.; Llambi, F.; Nourse, A.; Fitzgerald, P.; Gehring, K.; Kriwacki, R.W.; Green, D.R. BID-induced structural changes in BAK promote apoptosis. Nat. Struct. Mol. Biol. 2013, 20, 589–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czabotar, P.E.; Westphal, D.; Dewson, G.; Ma, S.; Hockings, C.; Fairlie, W.D.; Lee, E.F.; Yao, S.; Robin, A.Y.; Smith, B.J.; et al. Bax crystal structures reveal how BH3 domains activate Bax and nucleate its oligomerization to induce apoptosis. Cell 2013, 152, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.H.; Viacava Follis, A.; Kriwacki, R.W.; Moldoveanu, T. Discoveries and controversies in BCL-2 protein-mediated apoptosis. FEBS J. 2016, 283, 2690–2700. [Google Scholar] [CrossRef] [PubMed]
- Bogner, C.; Kale, J.; Pogmore, J.; Chi, X.; Shamas-Din, A.; Fradin, C.; Leber, B.; Andrews, D.W. Allosteric regulation of BH3 proteins in Bcl-xL complexes enables switch-like activation of bax. Mol. Cell 2020, 77, 901–912.e9. [Google Scholar] [CrossRef] [PubMed]
- Kalkavan, H.; Green, D.R. MOMP, cell suicide as a BCL-2 family business. Cell Death Differ. 2018, 25, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Tu, H.C.; Ren, D.; Takeuchi, O.; Jeffers, J.R.; Zambetti, G.P.; Hsieh, J.J.-D.; Cheng, E.H.Y. Stepwise activation of BAX and BAK by tBID, BIM, and PUMA initiates mitochondrial apoptosis. Mol. Cell 2009, 36, 487–499. [Google Scholar] [CrossRef] [Green Version]
- Lovell, J.F.; Billen, L.P.; Bindner, S.; Shamas-Din, A.; Fradin, C.; Leber, B.; Andrews, D.W. Membrane binding by tBid initiates an ordered series of events culminating in membrane permeabilization by bax. Cell 2008, 135, 1074–1084. [Google Scholar] [CrossRef] [Green Version]
- Gross, A.; Katz, S.G. Non-apoptotic functions of BCL-2 family proteins. Cell Death Differ. 2017, 24, 1348–1358. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Jones, A.F.; Sanger, R.H.; Collis, L.P.; Flannery, R.; McNay, E.C.; Yu, T.; Schwarzenbacher, R.; Bossy, B.; et al. Bcl-xL induces Drp1-dependent synapse formation in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 2169–2174. [Google Scholar] [CrossRef] [Green Version]
- Autret, A.; Martin, S.J. Bcl-2 family proteins and mitochondrial fission/fusion dynamics. Cell. Mol. Life Sci. 2010, 67, 1599–1606. [Google Scholar] [CrossRef]
- Karbowski, M.; Norris, K.L.; Cleland, M.M.; Jeong, S.Y.; Youle, R.J. Role of Bax and Bak in mitochondrial morphogenesis. Nature 2006, 443, 658–662. [Google Scholar] [CrossRef]
- Cleland, M.M.; Norris, K.L.; Karbowski, M.; Wang, C.; Suen, D.F.; Jiao, S.; George, N.M.; Luo, X.; Li, Z.; Youle, R.J. Bcl-2 family interaction with the mitochondrial morphogenesis machinery. Cell Death Differ. 2011, 18, 235–247. [Google Scholar] [CrossRef]
- Morciano, G.; Giorgi, C.; Balestra, D.; Marchi, S.; Perrone, D.; Pinotti, M.; Pinton, P. Mcl-1 involvement in mitochondrial dynamics is associated with apoptotic cell death. Mol. Biol. Cell 2016, 27, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Cereghetti, G.M.; Stangherlin, A.; de Brito, O.M.; Chang, C.R.; Blackstone, C.; Bernardi, P.; Scorrano, L. Dephosphorylation by calcineurin regulates translocation of Drp1 to mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 15803–15808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morciano, G.; Pedriali, G.; Sbano, L.; Iannitti, T.; Giorgi, C.; Pinton, P. Intersection of mitochondrial fission and fusion machinery with apoptotic pathways: Role of Mcl-1. Biol. Cell 2016, 108, 279–293. [Google Scholar] [CrossRef]
- Brooks, C.; Wei, Q.; Feng, L.; Dong, G.; Tao, Y.; Mei, L.; Xie, Z.-J.; Dong, Z. Bak regulates mitochondrial morphology and pathology during apoptosis by interacting with mitofusins. Proc. Natl. Acad. Sci. USA 2007, 104, 11649–11654. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, S.; Konishi, A.; Kodama, T.; Tsujimoto, Y. BH4 domain of antiapoptotic Bcl-2 family members closes voltage-dependent anion channel and inhibits apoptotic mitochondrial changes and cell death. Proc. Natl. Acad. Sci. USA 2000, 97, 3100–3105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, S.; Narita, M.; Tsujimoto, Y.; Tsujimoto, Y. Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC. Nature 1999, 399, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Arbel, N.; Ben-Hail, D.; Shoshan-Barmatz, V. Mediation of the antiapoptotic activity of Bcl-xL protein upon interaction with VDAC1 protein. J. Biol. Chem. 2012, 287, 23152–23161. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Shah, K.; Bradbury, N.A.; Li, C.; White, C. Mcl-1 promotes lung cancer cell migration by directly interacting with VDAC to increase mitochondrial Ca2+ uptake and reactive oxygen species generation. Cell Death Dis. 2014, 5, e1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karch, J.; Kwong, J.Q.; Burr, A.R.; Sargent, M.A.; Elrod, J.W.; Peixoto, P.M.; Martinez-Caballero, S.; Osinska, H.; Cheng, E.H.-Y.; Robbins, J.; et al. Bax and Bak function as the outer membrane component of the mitochondrial permeability pore in regulating necrotic cell death in mice. Elife 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Whelan, R.S.; Konstantinidis, K.; Wei, A.-C.; Chen, Y.; Reyna, D.E.; Jha, S.; Yang, Y.; Calvert, J.W.; Lindsten, T.; Thompson, C.B.; et al. Bax regulates primary necrosis through mitochondrial dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, 6566–6571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, G.; Decrock, E.; Arbel, N.; van Vliet, A.R.; La Rovere, R.M.; De Smedt, H.; Parys, J.B.; Agostinis, P.; Leybaert, L.; Shoshan-Barmatz, V.; et al. The BH4 domain of anti-apoptotic Bcl-XL, but not that of the related Bcl-2, limits the voltage-dependent anion channel 1 (VDAC1)-mediated transfer of pro-apoptotic Ca2+ signals to mitochondria. J. Biol. Chem. 2015, 290, 9150–9161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzo, I. Bax and adenine nucleotide translocator cooperate in the mitochondrial control of apoptosis. Science 1998, 281, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, H.; Vervliet, T.; Monaco, G.; Terry, L.E.; Rosa, N.; Baker, M.R.; Parys, J.B.; Serysheva, I.I.; Yule, D.I.; Bultynck, G. Bcl-2-protein family as modulators of IP3 receptors and other organellar Ca2+ channels. Cold Spring Harb. Perspect. Biol. 2020, 12. [Google Scholar] [CrossRef]
- Arbel, N.; Shoshan-Barmatz, V. Voltage-dependent anion channel 1-based peptides interact with Bcl-2 to prevent antiapoptotic activity. J. Biol. Chem. 2010, 285, 6053–6062. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Yu, Y.; Chua, B.H.L.; Ho, Y.S.; Kuo, T.H. Regulation of sodium–calcium exchange and mitochondrial energetics by Bcl-2 in the heart of transgenic mice. J. Mol. Cell. Cardiol. 2001, 33, 2135–2144. [Google Scholar] [CrossRef]
- Shteinfer-Kuzmine, A.; Argueti, S.; Gupta, R.; Shvil, N.; Abu-Hamad, S.; Gropper, Y.; Hoeber, J.; Magrì, A.; Messina, A.; Kozlova, E.N.; et al. A VDAC1-derived N-terminal peptide inhibits mutant SOD1-VDAC1 interactions and toxicity in the SOD1 model of ALS. Front. Cell. Neurosci. 2019, 13. [Google Scholar] [CrossRef] [Green Version]
- Abu-Hamad, S.; Arbel, N.; Calo, D.; Arzoine, L.; Israelson, A.; Keinan, N.; Ben-Romano, R.; Friedman, O.; Shoshan-Barmatz, V. The VDAC1 N-terminus is essential both for apoptosis and the protective effect of anti-apoptotic proteins. J. Cell Sci. 2009, 122, 1906–1916. [Google Scholar] [CrossRef] [Green Version]
- Shoshan-Barmatz, V.; Ben-Hail, D.; Admoni, L.; Krelin, Y.; Tripathi, S.S. The mitochondrial voltage-dependent anion channel 1 in tumor cells. Biochim. Biophys. Acta Biomembr. 2015, 1848, 2547–2575. [Google Scholar] [CrossRef] [Green Version]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A. VDAC1 functions in Ca2+ homeostasis and cell life and death in health and disease. Cell Calcium 2018, 69, 81–100. [Google Scholar] [CrossRef]
- Huang, H.; Hu, X.; Eno, C.O.; Zhao, G.; Li, C.; White, C. An interaction between Bcl-x L and the voltage-dependent anion channel (VDAC) promotes mitochondrial Ca2+ uptake. J. Biol. Chem. 2013, 288, 19870–19881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, E.; Grigoriev, S.M.; Dejean, L.M.; Zweihorn, C.L.; Mannella, C.A.; Kinnally, K.W. The mitochondrial channel VDAC has a cation-selective open state. Biochim. Biophys. Acta Bioenerg. 2005, 1710, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Abu-Hamad, S.; Zaid, H.; Israelson, A.; Nahon, E.; Shoshan-Barmatz, V. Hexokinase-I protection against apoptotic cell death is mediated via interaction with the voltage-dependent anion channel-1. J. Biol. Chem. 2008, 283, 13482–13490. [Google Scholar] [CrossRef] [Green Version]
- Shoshan-Barmatz, V.; Keinan, N.; Zaid, H. Uncovering the role of VDAC in the regulation of cell life and death. J. Bioenergy Biomembr. 2008, 40, 183–191. [Google Scholar] [CrossRef]
- Zaid, H.; Abu-Hamad, S.; Israelson, A.; Nathan, I.; Shoshan-Barmatz, V. The voltage-dependent anion channel-1 modulates apoptotic cell death. Cell Death Differ. 2005, 12, 751–760. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Li, X.X.; Gottleib, E.; Hill, R.B.; Thompson, C.B.; Colombini, M. Bcl-x l promotes the open configuration of the voltage-dependent anion channel and metabolite passage through the outer mitochondrial membrane. J. Biol. Chem. 2001, 276, 19414–19419. [Google Scholar] [CrossRef] [Green Version]
- Malia, T.J.; Wagner, G. NMR structural investigation of the mitochondrial outer membrane protein VDAC and its interaction with antiapoptotic Bcl-x L †. Biochemistry 2007, 46, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, S.; Shinohara, Y.; Tsujimoto, Y. Bax and Bcl-xL independently regulate apoptotic changes of yeast mitochondria that require VDAC but not adenine nucleotide translocator. Oncogene 2000, 19, 4309–4318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, S.; Choi, M.; Nguyen, Q.T.; Ye, H.; Liu, W.; Toh, H.T.; Kang, C.; Kamariah, N.; Li, C.; Huang, H.; et al. Structural transition in Bcl-xL and its potential association with mitochondrial calcium ion transport. Sci. Rep. 2015, 5, 10609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzuto, R.; De Stefani, D.; Raffaello, A.; Mammucari, C. Mitochondria as sensors and regulators of calcium signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Belzacq, A.-S.; Vieira, H.L.A.; Verrier, F.; Vandecasteele, G.; Cohen, I.; Prévost, M.-C.; Larquet, E.; Pariselli, F.; Petit, P.X.; Kahn, A.; et al. Bcl-2 and Bax modulate adenine nucleotide translocase activity. Cancer Res. 2003, 63, 541–546. [Google Scholar]
- Heiden, M.G.V.; Chandel, N.S.; Schumacker, P.T.; Thompson, C.B. Bcl-xL prevents cell death following growth factor withdrawal by facilitating mitochondrial ATP/ADP exchange. Mol. Cell 1999, 3, 159–167. [Google Scholar] [CrossRef]
- Chen, Y.; Aon, M.A.; Hsu, Y.-T.; Soane, L.; Teng, X.; McCaffery, J.M.; Cheng, W.-C.; Qi, B.; Li, H.; Alavian, K.N.; et al. Bcl-xL regulates mitochondrial energetics by stabilizing the inner membrane potential. J. Cell Biol. 2011, 195, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Lidman, M.; Pokorná, Š.; Dingeldein, A.P.G.; Sparrman, T.; Wallgren, M.; Šachl, R.; Hof, M.; Gröbner, G. The oxidized phospholipid PazePC promotes permeabilization of mitochondrial membranes by Bax. Biochim. Biophys. Acta Biomembr. 2016, 1858, 1288–1297. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Ellis, H. Genetic control of programmed cell death in the nematode C. elegans. Cell 1986, 44, 817–829. [Google Scholar] [CrossRef]
- Garrido, C.; Kroemer, G. Life’s smile, death’s grin: Vital functions of apoptosis-executing proteins. Curr. Opin. Cell Biol. 2004, 16, 639–646. [Google Scholar] [CrossRef]
- Galluzzi, L.; Joza, N.; Tasdemir, E.; Maiuri, M.C.; Hengartner, M.; Abrams, J.M.; Tavernarakis, N.; Penninger, J.; Madeo, F.; Kroemer, G. No death without life: Vital functions of apoptotic effectors. Cell Death Differ. 2008, 15, 1113–1123. [Google Scholar] [CrossRef] [Green Version]
- Guicciardi, M.E.; Malhi, H.; Mott, J.L.; Gores, G.J. (Eds.) Apoptosis and necrosis in the liver. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Fink, S.L.; Cookson, B.T. Apoptosis, pyroptosis, and necrosis: Mechanistic description of dead and dying eukaryotic cells. Infect. Immun. 2005, 73, 1907–1916. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Kang, J.; Fu, C. The independence of and associations among apoptosis, autophagy, and necrosis. Signal Transduct. Target. Ther. 2018, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikoletopoulou, V.; Markaki, M.; Palikaras, K.; Tavernarakis, N. Crosstalk between apoptosis, necrosis and autophagy. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 3448–3459. [Google Scholar] [CrossRef] [Green Version]
- Kroemer, G.; El-Deiry, W.S.; Golstein, P.; Peter, M.E.; Vaux, D.; Vandenabeele, P.; Zhivotovsky, B.; Blagosklonny, M.V.; Malorni, W.; Knight, R.A.; et al. Classification of cell death: Recommendations of the nomenclature committee on cell death. Cell Death Differ. 2005, 12, 1463–1467. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, C.J. The apoptosis-necrosis continuum: Insights from genetically altered mice. Vet. Pathol. 2003, 40, 481–495. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, G.; Konstantinidis, K.; Kitsis, R.N. Programmed necrosis, not apoptosis, in the heart. Circ. Res. 2011, 108, 1017–1036. [Google Scholar] [CrossRef] [Green Version]
- Izzo, V.; Bravo-San Pedro, J.M.; Sica, V.; Kroemer, G.; Galluzzi, L. Mitochondrial permeability transition: New findings and persisting uncertainties. Trends Cell Biol. 2016, 26, 655–667. [Google Scholar] [CrossRef]
- Alavian, K.N.; Li, H.; Collis, L.; Bonanni, L.; Zeng, L.; Sacchetti, S.; Lazrove, E.; Nabili, P.; Flaherty, B.; Graham, M.; et al. Bcl-xL regulates metabolic efficiency of neurons through interaction with the mitochondrial F1FO ATP synthase. Nat. Cell Biol. 2011, 13, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, H.; Xu, A.; Ross, T.; Bowler, E.; Hu, Y.; Lesnefsky, E.J. Inhibition of Bcl-2 sensitizes mitochondrial permeability transition pore (MPTP) opening in ischemia-damaged mitochondria. PLoS ONE 2015, 10, e0118834. [Google Scholar] [CrossRef] [Green Version]
- Zamzami, N.; Hamel, C.E.L.; Maisse, C.; Brenner, C.; Muñoz-Pinedo, C.; Belzacq, A.-S.; Costantini, P.; Vieira, H.; Loeffler, M.; Molle, G.; et al. Bid acts on the permeability transition pore complex to induce apoptosis. Oncogene 2000, 19, 6342–6350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic presentation of Ca2+ transport systems in mitochondria. (1) Ca2+ influx and efflux through the outer mitochondrial membrane (OMM) driven via the voltage-dependent anion channel (VDAC). (2) Ca2+ influx through the inner mitochondrial membrane (IMM) driven by three major transport systems: (i) mitochondrial Ca2+ uniporter (MCU), (ii) mitochondrial ryanodine receptor (mRyR), (iii) rapid mode of Ca2+ uptake (RaM) and one mitochondrial system under debate: (iv) leucine zipper- EF-hand containing transmembrane protein (LETM1). Ca2+ influx through the MCU is established by the electrochemical gradient created by the electron transport chain (ETC). (3) Ca2+ efflux through the IMM driven by three other major transport systems: (i) Na+/Ca2+/Li+ (NCLX) exchanger, (ii) H+/Ca2+ exchanger (HCX), (iii) mitochondrial permeability transition pore complex (mPTPC) and one mitochondrial system under debate: (iv) leucine zipper- EF-hand containing transmembrane protein (LETM1). (4) The core constituents of mPTPC include: the adenine nucleotide translocase (ANT), matrix cyclophilin D (CypD) and phosphate carrier (PiC), which serve as pore regulators, and the pro-apoptotic proteins Bax and Bak, which can induce mitochondrial swelling and rupture during the mPTP opening. ATP-synthase is the key IMM-pore forming unit of mPTPC.
Figure 1.
Schematic presentation of Ca2+ transport systems in mitochondria. (1) Ca2+ influx and efflux through the outer mitochondrial membrane (OMM) driven via the voltage-dependent anion channel (VDAC). (2) Ca2+ influx through the inner mitochondrial membrane (IMM) driven by three major transport systems: (i) mitochondrial Ca2+ uniporter (MCU), (ii) mitochondrial ryanodine receptor (mRyR), (iii) rapid mode of Ca2+ uptake (RaM) and one mitochondrial system under debate: (iv) leucine zipper- EF-hand containing transmembrane protein (LETM1). Ca2+ influx through the MCU is established by the electrochemical gradient created by the electron transport chain (ETC). (3) Ca2+ efflux through the IMM driven by three other major transport systems: (i) Na+/Ca2+/Li+ (NCLX) exchanger, (ii) H+/Ca2+ exchanger (HCX), (iii) mitochondrial permeability transition pore complex (mPTPC) and one mitochondrial system under debate: (iv) leucine zipper- EF-hand containing transmembrane protein (LETM1). (4) The core constituents of mPTPC include: the adenine nucleotide translocase (ANT), matrix cyclophilin D (CypD) and phosphate carrier (PiC), which serve as pore regulators, and the pro-apoptotic proteins Bax and Bak, which can induce mitochondrial swelling and rupture during the mPTP opening. ATP-synthase is the key IMM-pore forming unit of mPTPC.

Figure 2.
Classification of Bcl-2 of proteins based on their apoptotic activity. (A) anti-apoptotic; (B) pro-apoptotic; and (C) apoptotic mediators of the Bcl-2 family proteins containing one BH-3 domain.
Figure 2.
Classification of Bcl-2 of proteins based on their apoptotic activity. (A) anti-apoptotic; (B) pro-apoptotic; and (C) apoptotic mediators of the Bcl-2 family proteins containing one BH-3 domain.

Figure 3.
Bcl-2 proteins and mitochondrial Ca2+ transport systems determine the necrotic or apoptotic cell death pathways. mPTP-mediated necrosis and MOMP-mediated apoptosis are initiated by various extracellular and intracellular cell death stimuli. Central events for cell death rely on interactions between “activators” and “sensitizers” BH3-only proteins (these proteins are marked by dark blue). Activators (BID, BIM, PUMA, NOXA) bind and activate Bax and Bak and induce a series of their conformational changes and subsequent oligomerization within the OMM, finally resulting in MOMP. Sensitizers (BAD, BIK, BMF, HRK) bind the pro-survival Bcl-2 proteins (Bcl-2, Bcl-XL, Mcl-1) and release them to activate Bax and Bak. MOMP results in the release of pro-apoptogenic factors, formation of an apoptosome, and activation of the cascade that leads to MOMP-mediated apoptosis (these events are marked by orange). In contrast, the central event for mPTP-mediated necrosis is the Ca2+-induced formation of mPTPC in the IMM and its mediation by the Bcl-2 members (Bax, Bak, Bcl-XL, Bcl-2, BID). mPTP opening rapidly dissipates the proton gradient across the IMM and induce mPTP-mediated necrosis (all of these events are marked by green). Thus, a possible functional link between Bcl-2 proteins and Ca2+-transport systems and between MOMP-mediated apoptosis and mPTP-mediated necrosis may exist. Intracellular or extracellular cell death stimuli may lead to the other types of RCD that are not directly related to the mediation of Ca2+ homeostasis by Bcl-2 members (including extrinsic apoptosis, necroptosis, ferroptosis, pyroptosis, parthanatos, entotic cell death, NETotic cell death, LDCD, ADCD, ICD) (these events are marked by light blue).
Figure 3.
Bcl-2 proteins and mitochondrial Ca2+ transport systems determine the necrotic or apoptotic cell death pathways. mPTP-mediated necrosis and MOMP-mediated apoptosis are initiated by various extracellular and intracellular cell death stimuli. Central events for cell death rely on interactions between “activators” and “sensitizers” BH3-only proteins (these proteins are marked by dark blue). Activators (BID, BIM, PUMA, NOXA) bind and activate Bax and Bak and induce a series of their conformational changes and subsequent oligomerization within the OMM, finally resulting in MOMP. Sensitizers (BAD, BIK, BMF, HRK) bind the pro-survival Bcl-2 proteins (Bcl-2, Bcl-XL, Mcl-1) and release them to activate Bax and Bak. MOMP results in the release of pro-apoptogenic factors, formation of an apoptosome, and activation of the cascade that leads to MOMP-mediated apoptosis (these events are marked by orange). In contrast, the central event for mPTP-mediated necrosis is the Ca2+-induced formation of mPTPC in the IMM and its mediation by the Bcl-2 members (Bax, Bak, Bcl-XL, Bcl-2, BID). mPTP opening rapidly dissipates the proton gradient across the IMM and induce mPTP-mediated necrosis (all of these events are marked by green). Thus, a possible functional link between Bcl-2 proteins and Ca2+-transport systems and between MOMP-mediated apoptosis and mPTP-mediated necrosis may exist. Intracellular or extracellular cell death stimuli may lead to the other types of RCD that are not directly related to the mediation of Ca2+ homeostasis by Bcl-2 members (including extrinsic apoptosis, necroptosis, ferroptosis, pyroptosis, parthanatos, entotic cell death, NETotic cell death, LDCD, ADCD, ICD) (these events are marked by light blue).


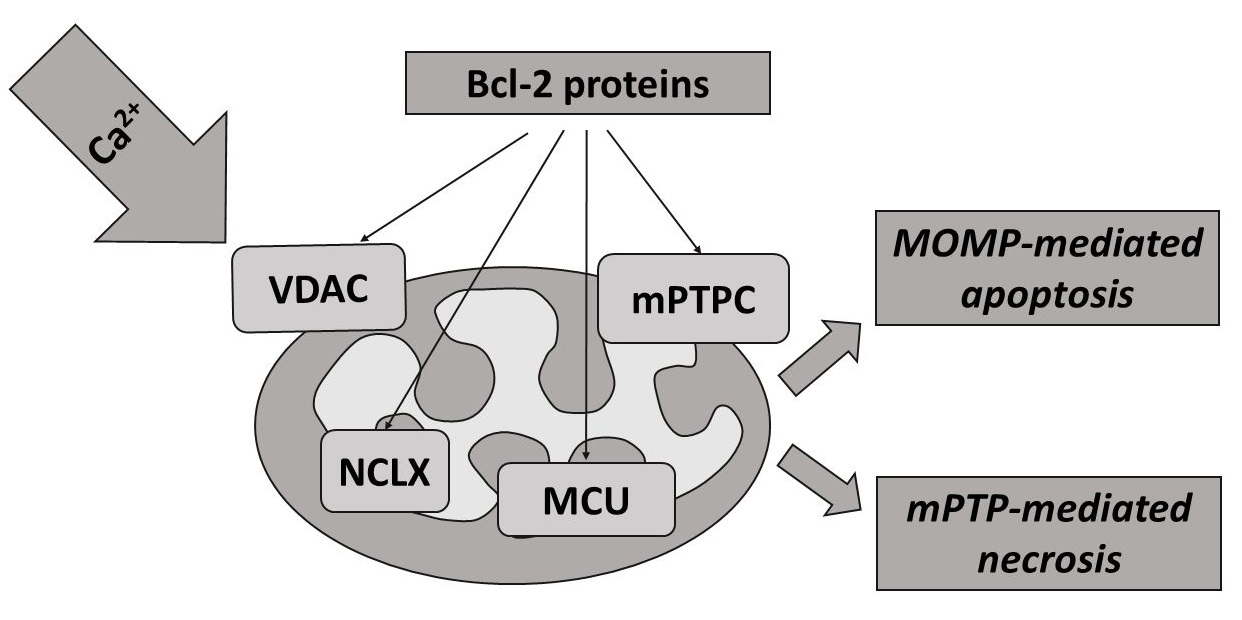{kind=link}
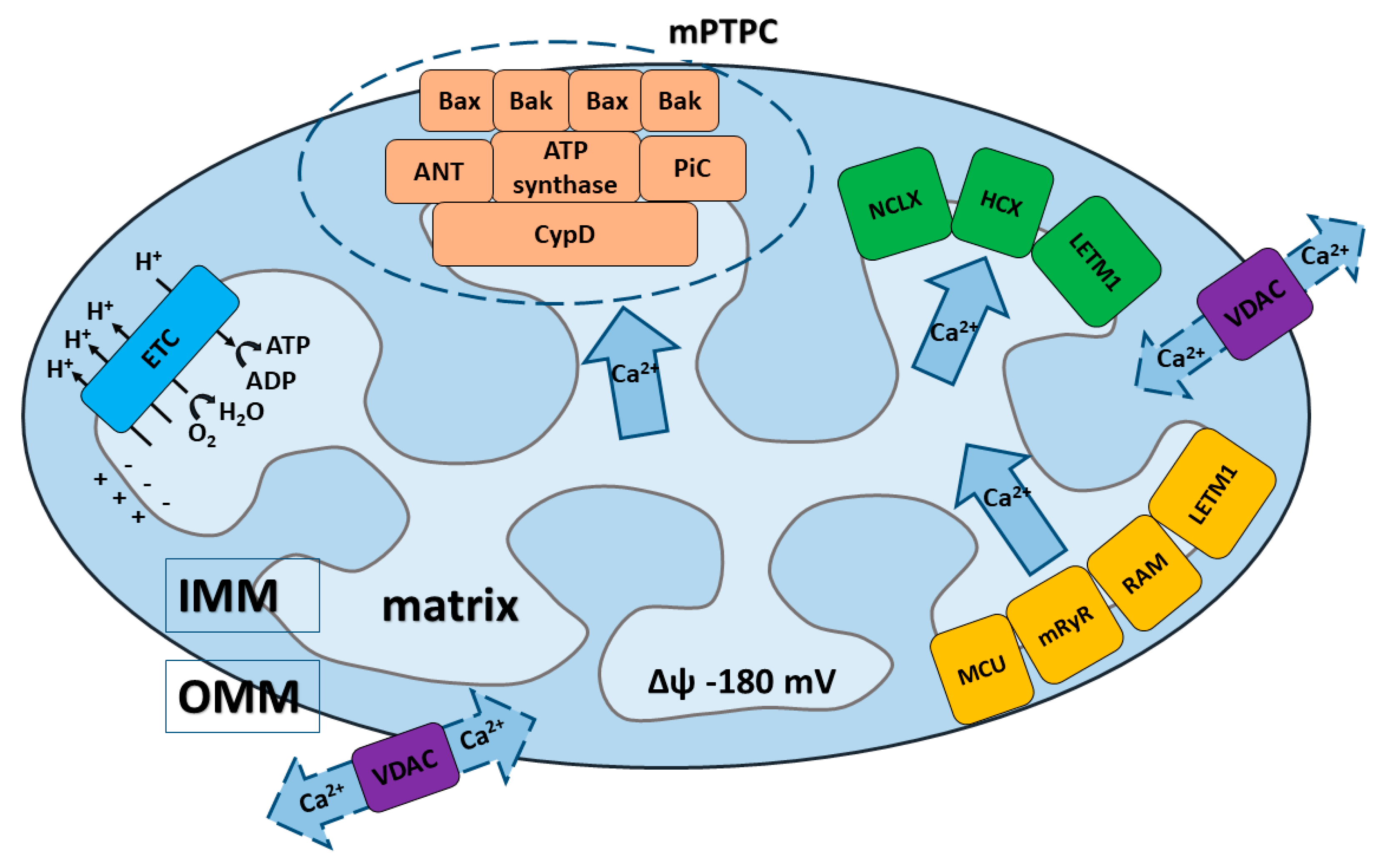{kind=link}
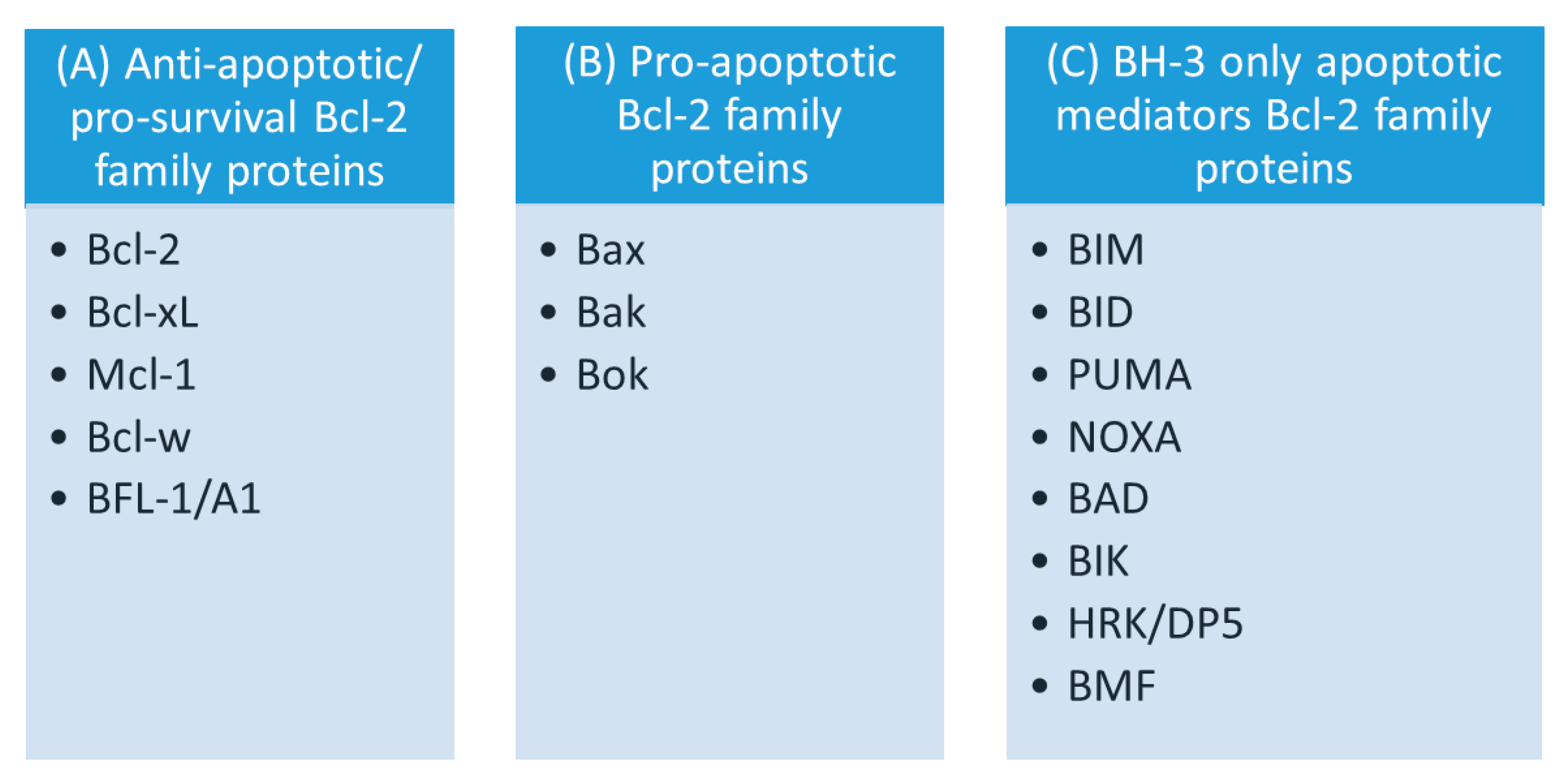{kind=link}
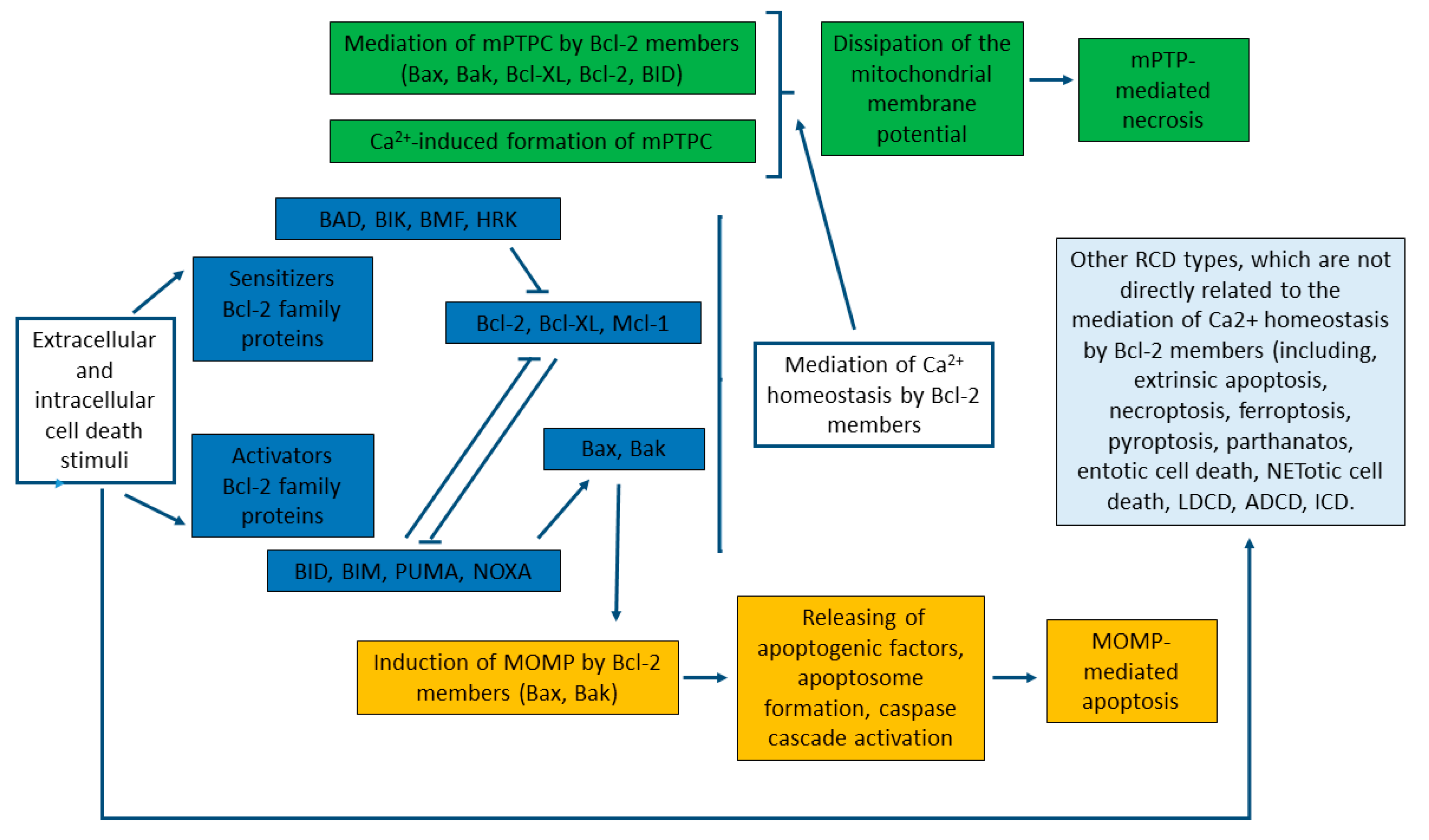{kind=link}
Table 1.
The interactions between mitochondrial Ca2+ transport systems and Bcl-2 proteins.
| Direction of Ca2+ Transport | Mitochondrial Membrane Through Which Ca2+ Transport is Realized | The System Responsible for Ca2+ Transport | Characteristic Features of the Transport System | Regulation of Mitochondrial Ca2+ Transport Systems by Bcl-2 Proteins |
|---|---|---|---|---|
| Ca2+ influx | OMM | VDAC | The main transport system for metabolites, cations, and anions across OMM. It serves as a contact point between the OMM and IMM [42]; 3 different VDAC isoforms have been identified [45,48,53,60] | Ca2+ influx via VDAC is regulated by Bcl-2, Bcl-XL and Mcl-1 [29,254,255,256,257,260,263] |
| IMM | MCU | MCU is the major pathway of the mitochondrial Ca2+ uptake [9,17,60,63,64,65]. MCU consists of several subunits (see Box 2 for details [65,66,68,69,70] | Activity of MCU is mediated by Bcl-2 proteins (BID, BAD, Bcl-XL) [194]. | |
| RaM | RaM accumulates Ca2+ with the kinetics hundreds of times faster than MCU. It could also represent a different form or substrate of MCU [89,91] | |||
| mRyR (mitochondrial ryanodine receptor) | The isoform of RyR1. Proposed as a Ca2+-influx system, involved in the regulation of Ca2+ efflux under mitochondrial Ca2+ overload [92,93] | |||
| LETM1 | Proposed as Ca2+/H+ antiporter, also involved in the K+/H+ exchange. Shares a key role with MCU in catalyzing Ca2+ uptake into mitochondria [62,96,101,103] | |||
| Ca2+ efflux | OMM | VDAC | See above | See above |
| mPTP pore complex | ANT, PiC and CypD serve as mPTP regulators in the IMM. The F1F0ATP synthase has been suggested as pore-forming unit in the IMM [110,135,138,141,144,148] | Activity of mPTPC is mediated by the ensemble of Bcl-2 proteins (Bax/Bak, BID, BAD, Bcl-XL) [255,258,259,261] | ||
| IMM | (NCLX) Na+/Ca2+/Li+ exchanger | Na+-dependent Ca2+ transport also includes the transport of Li+. It is typical for excitable tissues (brain, heart) [113,114]. | Bcl-2 protein can modulate the activity of NCLX [264]. | |
| HCX (H+/Ca2+ exchanger) | Na+-independent Ca2+ efflux is dominant in the non-excitable tissues (liver, kidney, lung, smooth muscle) [119,126] | |||
| LETM1 | Proposed as an alternative mechanism for the regulation of Ca2+ release and functions as a Ca2+/H+ antiporter during specific conditions [62,96,101,102,103] | |||
| mPTP pore complex | See above | See above |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Naumova, N.; Šachl, R. Regulation of Cell Death by Mitochondrial Transport Systems of Calcium and Bcl-2 Proteins. Membranes 2020, 10, 299. https://doi.org/10.3390/membranes10100299
AMA Style
Naumova N, Šachl R. Regulation of Cell Death by Mitochondrial Transport Systems of Calcium and Bcl-2 Proteins. Membranes. 2020; 10(10):299. https://doi.org/10.3390/membranes10100299
Chicago/Turabian StyleNaumova, Natalia, and Radek Šachl. 2020. "Regulation of Cell Death by Mitochondrial Transport Systems of Calcium and Bcl-2 Proteins" Membranes 10, no. 10: 299. https://doi.org/10.3390/membranes10100299
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.