
Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mitochondria and ROS Production
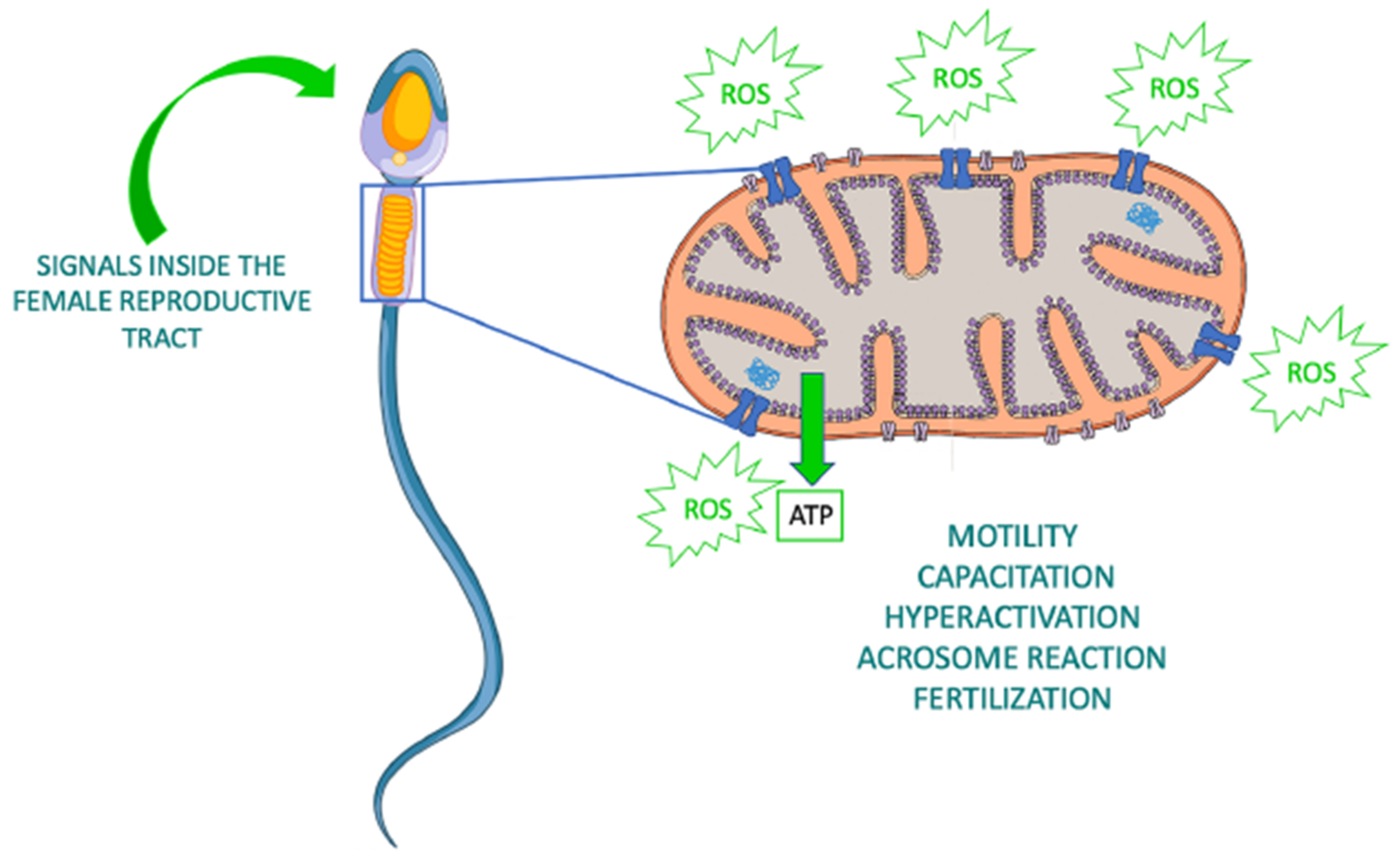
3. Roles of Sperm Mitochondria
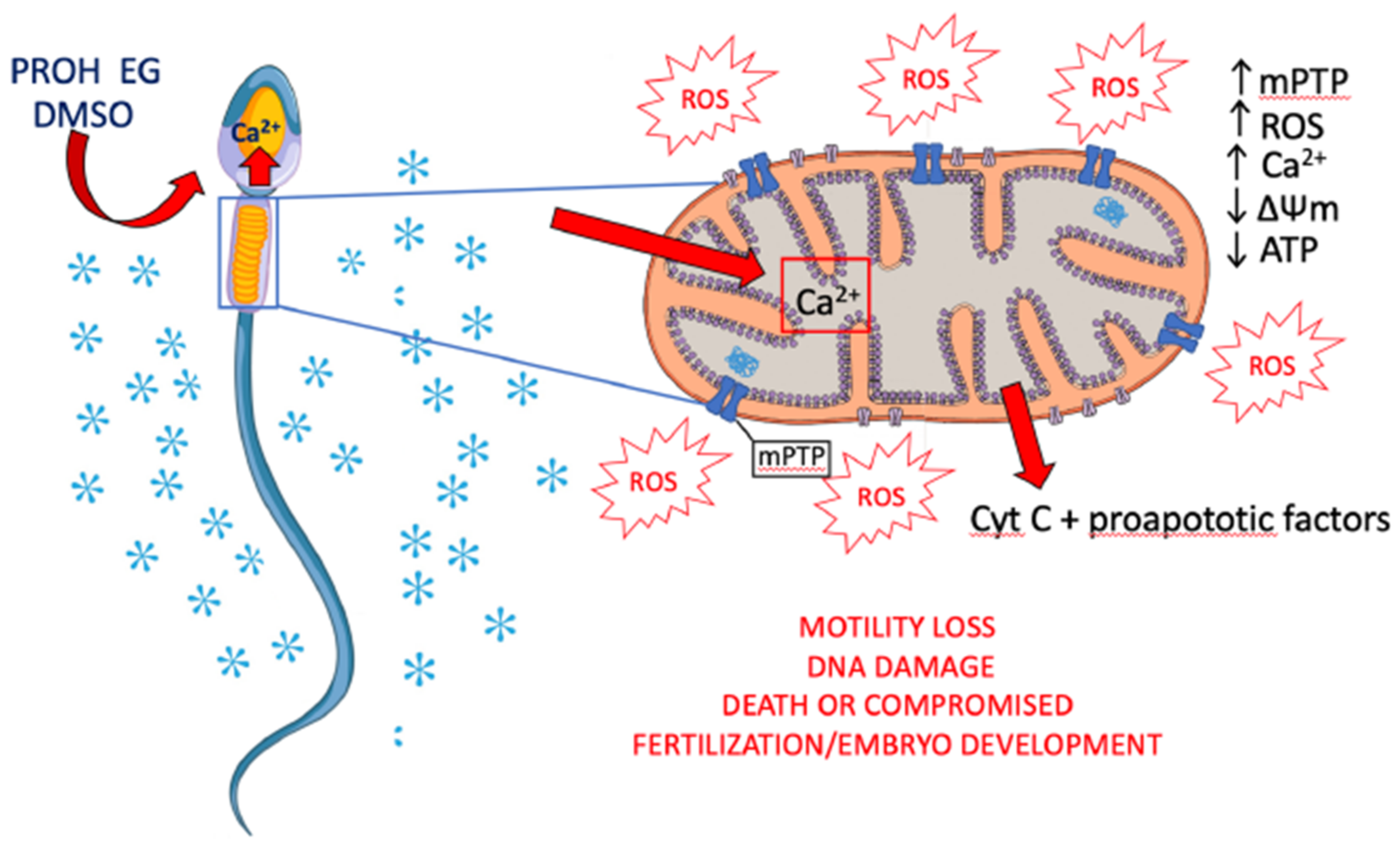
4. Effects of Sperm Cryopreservation on Mitochondria and Oxidative Stress
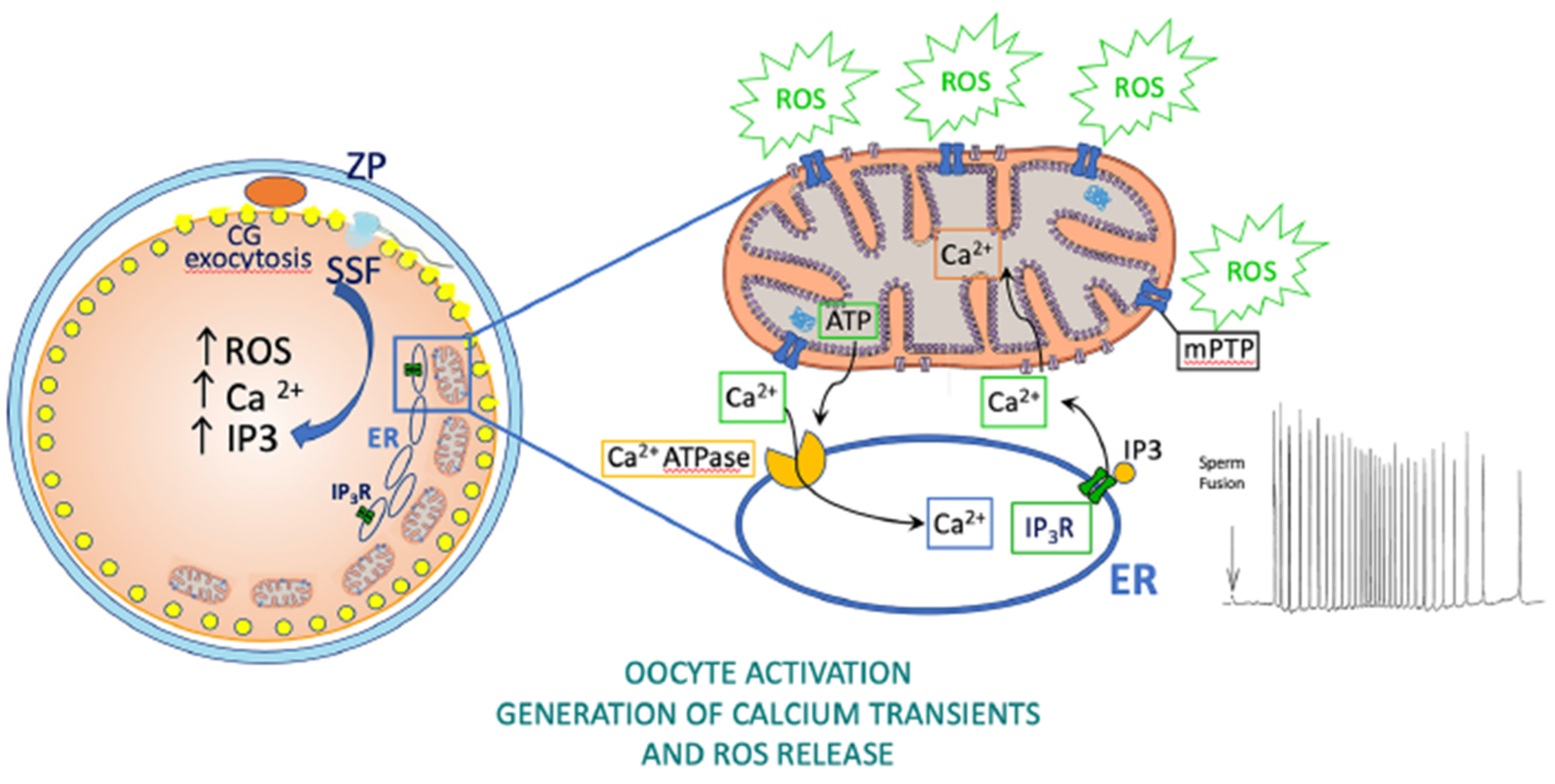
5. Roles of Oocyte Mitochondria
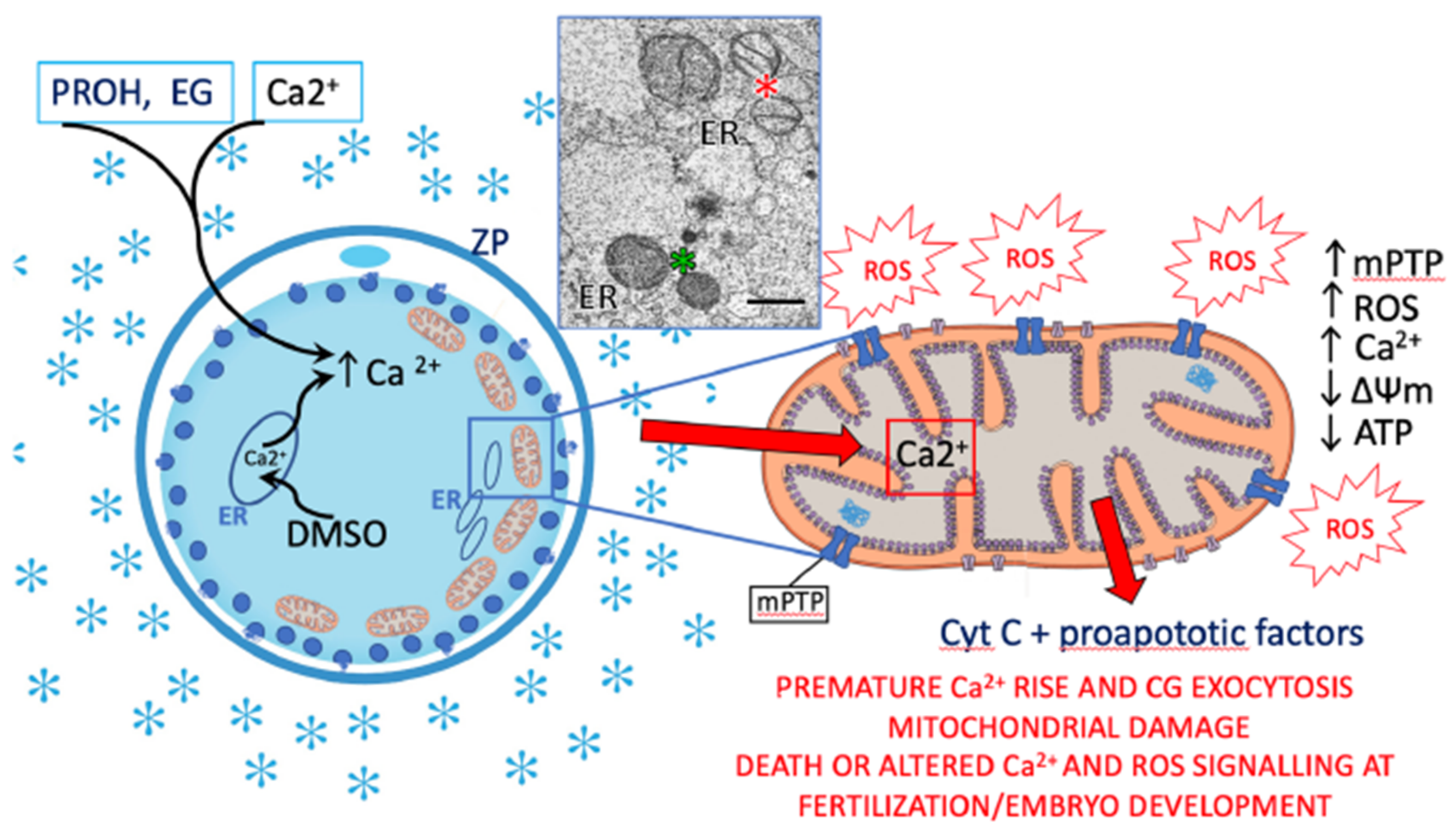
6. Effects of Oocyte Cryopreservation on Mitochondria, Oxidative Stress, Fertilization, and Embryo Development
7. Effects of Gonadal Tissue Cryopreservation on Mitochondria and Oxidative Stress
8. Concluding Remarks
Funding
Conflicts of Interest
References
- Spallanzani, L. Opuscoli di Fisica Animale e Vegetabile; Presso la Società Tipografica: Modena, Italy, 1776; Volume 2, p. 2. [Google Scholar]
- Polge, C.; Smith, A.; Parkes, A. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature 1949, 164, 666. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Zorov, D.B. Pros and cons of use of mitochondria-targeted antioxidants. Antioxidants 2019, 8, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [Green Version]
- Zinovkin, R.A.; Zamyatnin, A.A. Mitochondria-Targeted Drugs. Curr. Mol. Pharmacol. 2019, 12, 202–214. [Google Scholar] [CrossRef] [PubMed]
- de Lamirande, E.; Gagnon, C. Impact of reactive oxygen species on spermatozoa: A balancing act between beneficial and detrimental effects. Hum. Reprod. 1995, 10, 15–21. [Google Scholar] [CrossRef]
- Talevi, R.; Zagami, M.; Castaldo, M.; Gualtieri, R. Redox regulation of sperm surface thiols modulates adhesion to the fallopian tube epithelium. Biol. Reprod. 2007, 76, 728–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkov, A.A. The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. N. Y. Acad. Sci. USA 2008, 1147, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Gualtieri, R.; Mollo, V.; Duma, G.; Talevi, R. Redox control of surface protein sulphhydryls in bovine spermatozoa reversibly modulates sperm adhesion to the oviductal epithelium and capacitation. Reproduction 2009, 138, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Nasr-Esfahani, M.H.; Aitken, J.R.; Johnson, M.H. The origin of reactive oxygen species in mouse embryos cultured in vitro. Development 1991, 113, 551–560. [Google Scholar] [PubMed]
- Schomer, B.; Epel, D. Redox changes during fertilization and maturation of marine invertebrate eggs. Dev. Biol. 1998, 203, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Ward, Z.; Carroll, J.; Duchen, M.R. Regulation of redox metabolism in the mouse oocyte and embryo. Development 2007, 134, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumollard, R.; Duchen, M.; Carroll, J. The role of mitochondrial function in the oocyte and embryo. Curr. Top. Dev. Biol. 2007, 77, 21–49. [Google Scholar] [PubMed]
- Morado, S.A.; Cetica, P.D.; Beconi, M.T.; Dalvit, G.C. Reactive oxygen species in bovine oocyte maturation in vitro. Reprod. Fertil. Dev. 2009, 21, 608–614. [Google Scholar] [CrossRef]
- Lopes, A.S.; Lane, M.; Thompson, J.G. Oxygen consumption and ROS production are increased at the time of fertilization and cell cleavage in bovine zygotes. Hum. Reprod. 2010, 25, 2762–2773. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Ishibashi, S.; Iglesias-Gonzalez, J.; Chen, Y.; Love, N.R.; Amaya, E. Ca2+-induced mitochondrial ROS regulate the early embryonic cell cycle. Cell. Rep. 2018, 22, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. Oxidative stress: Introductory remarks. In Oxidative Stress, 1st ed.; Sies, H., Ed.; Academic Press: London, UK, 1985; pp. 1–8. [Google Scholar]
- Sies, H.; Jones, D.P. Oxidative stress. In Encyclopedia of Stress, 2nd ed.; Fink, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 45–48. [Google Scholar]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A. VDAC1 functions in Ca2+ homeostasis and cell life and death in health and disease. Cell Calcium. 2018, 69, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Boveris, A.; Chance, B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J. 1973, 134, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.J.; Brand, M.D. Superoxide production by NADH:ubiquinone oxidoreductase (complex I) depends on the pH gradient across the mitochondrial inner membrane. Biochem. J. 2004, 382, 511–517. [Google Scholar] [CrossRef]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39, BSR20191601. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J. An Update on Mitochondrial Reactive Oxygen Species Production. Antioxidants 2020, 9, 472. [Google Scholar] [CrossRef]
- Van Houten, B.; Woshner, V.; Santos, J.H. Role of mitochondrial DNA in toxic responses to oxidative stress. DNA Repair 2006, 5, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Gordeeva, A.V.; Zvyagilskaya, R.A.; Labas, Y.A. Cross-talk between reactive oxygen species and calcium in living cells. Biochemistry 2003, 68, 1077–1080. [Google Scholar]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonora, M.; Pinton, P. The mitochondrial permeability transition pore and cancer: Molecular mechanisms involved in cell death. Front. Oncol. 2014, 4, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell. Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Javadov, S.; Karmazyn, M. Mitochondrial permeability transition pore opening as an endpoint to initiate cell death and as a putative target for cardioprotection. Cell Physiol Biochem. 2007, 20, 1–22. [Google Scholar] [CrossRef]
- Hurst, S.; Hoek, J.; Sheu, S.S. Mitochondrial Ca2+ and regulation of the permeability transition pore. J. Bioenerg. Biomembr. 2017, 49, 27–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantrow, S.P.; Piantadosi, C.A. Release of cytochrome c from liver mitochondria during permeability transition. Biochem Biophys Res. Commun. 1997, 232, 669–671. [Google Scholar] [CrossRef]
- Kim, J.S.; He, L.; Lemasters, J.J. Mitochondrial permeability transition: A common pathway to necrosis and apoptosis. Biochem. Biophys. Res. Commun. 2003, 304, 463–470. [Google Scholar] [CrossRef]
- Susin, S.A.; Lorenzo, H.K.; Zamzami, N.; Marzo, I.; Snow, B.E.; Brothers, G.M.; Mangion, J.; Jacotot, E.; Costantini, P.; Loeffler, M.; et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 1999, 397, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.M.; Hershman, S.; Stabley, G.J.; Hoek, J.B.; Peterson, J.; Cahill, A. A Ca2+-induced mitochondrial permeability transition causes complete release of rat liver endonuclease G activity from its exclusive location within the mitochondrial intermembrane space. Identification of a novel endo-exonuclease activity residing within the mitochondrial matrix. Nucleic Acids. Res. 2003, 31, 1364–1373. [Google Scholar] [PubMed] [Green Version]
- Ankel-Simons, F.; Cummins, M.J. Misconceptions about mitochondria and mammalian fertilization: Implications for theories on human evolution. PNAS 1996, 93, 13859–13863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajender, S.; Rahul, P.; Ali Mahdi, A. Mitochondria, spermatogenesis and male infertility. Mitochondrion 2010, 10, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Cocuzza, M.; Abdelrazik, H.; Sharma, R. Oxidative stress measurement in patients with male or female factor infertility. In Handbook of Chemiluminescent Methods in Oxidative Stress Assessment; Popov, I., Lewin, G., Eds.; Transworld Research Network: Kerala, India, 2009; pp. 195–218. [Google Scholar]
- Durairajanayagam, D.; Singh, D.; Agarwal, A.; Henkel, R. Causes and consequences of sperm mitochondrial dysfunction. Andrologia 2020, 53, e13666. [Google Scholar] [CrossRef]
- Park, Y.J.; Pang, M.G. Mitochondrial functionality in male fertility: From spermatogenesis to fertilization. Antioxidants 2021, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Darr, C.R.; Cortopassi, G.A.; Datta, S.; Varner, D.D.; Meyers, S.A. Mitochondrial oxygen consumption is a unique indicator of stallion spermatozoal health and varies with cryopreservation media. Theriogenology. 2016, 86, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Losano, J.D.A.; Padín, J.F.; Méndez-López, I.; Angrimani, D.S.R.; García, A.G.; Barnabe, V.H.; Nichi, M. The stimulated glycolytic pathway is able to maintain ATP levels and kinetic patterns of bovine epididymal sperm subjected to mitochondrial uncoupling. Oxid. Med. Cell. Longev. 2017, 2017, 1682393. [Google Scholar] [CrossRef]
- Hu, C.H.; Zhuang, X.J.; Wei, Y.M.; Zhang, M.; Lu, S.S.; Lu, Y.Q.; Yang, X.G.; Lu, K.H. Comparison of mitochondrial function in boar and bull spermatozoa throughout cryopreservation based on JC-1 staining. Cryo Lett. 2017, 38, 75–79. [Google Scholar]
- Paoli, D.; Gallo, M.; Rizzo, F.; Baldi, E.; Francavilla, S.; Lenzi, A.; Lombardo, F.; Gandini, L. Mitochondrial membrane potential profile and its correlation with increasing sperm motility. Fertil Steril. 2011, 95, 2315–2319. [Google Scholar] [CrossRef]
- Ferramosca, A.; Provenzano, S.P.; Coppola, L.; Zara, V. Mitochondrial respiratory efficiency is positively correlated with human sperm motility. Urology 2012, 79, 809–814. [Google Scholar] [CrossRef]
- Kasai, T.; Ogawa, K.; Mizuno, K.; Nagai, S.; Uchida, Y.; Ohta, S.; Fujie, M.; Suzuki, K.; Hirata, S.; Hoshi, K. Relationship between sperm mitochondrial membrane potential, sperm motility, and fertility potential. Asian J. Androl. 2002, 4, 97–103. [Google Scholar]
- Sousa, A.P.; Amaral, A.; Baptista, M.; Tavares, R.; Caballero Campo, P.; Caballero Peregrin, P.; Freitas, A.; Paiva, A.; Almeida-Santos, T.; Ramalho-Santos, J. Not all sperm are equal: Functional mitochondria characterize a subpopulation of human sperm with better fertilization potential. PLoS ONE 2011, 6, e18112. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, P.; Ballot, C.; Jouy, N.; Thomas, P.; Marchetti, C. Influence of mitochondrial membrane potential of spermatozoa on in vitro fertilisation outcome. Andrologia 2012, 44, 136–141. [Google Scholar] [CrossRef]
- Malic Voncina, S.; Golob, B.; Ihan, A.; Kopitar, A.N.; Kolbezen, M.; Zorn, B. Sperm DNA fragmentation and mitochondrial membrane potential combined are better for predicting natural conception than standard sperm parameters. Fertil. Steril. 2016, 105, 637–644.e1. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yang, W.; Zou, P.; Jiang, F.; Zeng, Y.; Chen, Q.; Sun, L.; Yang, H.; Zhou, N.; Wang, X.; et al. Mitochondrial functionality modifies human sperm acrosin activity, acrosome reaction capability and chromatin integrity. Hum. Reprod. 2019, 34, 3–11. [Google Scholar] [CrossRef]
- Reynier, P.; Chrétien, M.F.; Savagner, F.; Larcher, G.; Rohmer, V.; Barrière, P.; Malthièry, Y. Long PCR analysis of human gamete mtDNA suggests defective mitochondrial maintenance in spermatozoa and supports the bottleneck theory for oocytes. Biochem. Biophys. Res. Commun. 1998, 252, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, S.; Deecaraman, M.; Kumar, R.; Shamsi, M.B.; Dada, R. Role of reactive oxygen species in the pathogenesis of mitochondrial DNA (mtDNA) mutations in male infertility. Indian J. Med. Res. 2009, 129, 127–137. [Google Scholar]
- Kumar, D.P.; Sangeetha, N. Mitochondrial DNA mutations and male infertility. Indian, J. Hum. Genet. 2009, 15, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May-Panloup, P.; Chretien, M.F.; Savagner, F.; Vasseur, C.; Jean, M.; Malthiery, Y.; Reynier, P. Increased sperm mitochondrial DNA content in male infertility. Hum. Reprod. 2003, 18, 550–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G.J.; Lewis, V. Mitochondrial DNA integrity and copy number in sperm from infertile men. Fert Steril. 2008, 90, 2238–2244. [Google Scholar] [CrossRef]
- Rosati, A.J.; Whitcomb, B.W.; Brandon, N.; Buck Louis, G.M.; Mumford, S.L.; Schisterman, E.F.; Pilsner, J.R. Sperm mitochondrial DNA biomarkers and couple fecundity. Hum. Reprod. 2020, 35, 2619–2625. [Google Scholar] [CrossRef] [PubMed]
- Tiegs, A.W.; Tao, X.; Landis, J.; Zhan, Y.; Franasiak, J.M.; Seli, E.; Wells, D.; Fragouli, E.; Scott, R.T., Jr. Sperm mitochondrial DNA copy number is not a predictor of intracytoplasmic sperm injection (ICSI) cycle outcomes. Reprod. Sci. 2020, 27, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Moraes, C.R.; Meyers, S. The sperm mitochondrion: Organelle of many functions. Anim. Reprod. Sci. 2018, 194, 71–80. [Google Scholar] [CrossRef]
- Buzadzic, B.; Vucetic, M.; Jankovic, A.; Stancic, A.; Korac, A.; Korac, B.; Otasevic, V. New insights into male (in)fertility: The importance of NO. Br. J. Pharmacol. 2015, 172, 1455–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Prabakaran, S.A.; Said, T.M. Prevention of oxidative stress injury to sperm. J. Androl. 2005, 26, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Abayasekara, D.R.; Aitken, R.J. Polyunsaturated fatty acids in male and female reproduction. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- Walters, J.L.H.; De Iuliis, G.N.; Nixon, B.; Bromfield, E.G. Oxidative stress in the male germline: A review of novel strategies to reduce 4-hydroxynonenal production. Antioxidants 2018, 7, 132. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Gibb, Z.; Mitchell, L.A.; Lambourne, S.R.; Connaughton, H.S.; De Iuliis, G.N. Sperm motility is lost in vitro as a consequence of mitochondrial free radical production and the generation of electrophilic aldehydes but can be significantly rescued by the presence of nucleophilic thiols. Biol. Reprod. 2012, 87, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J.; Whiting, S.; De Iuliis, G.N.; McClymont, S.; Mitchell, L.A.; Baker, M.A. Electrophilic aldehydes generated by sperm metabolism activate mitochondrial reactive oxygen species generation and apoptosis by targeting succinate dehydrogenase. J. Biol. Chem. 2012, 287, 33048–33060. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Abush, A.; Hauser, R.; Paz, G.; Kleiman, S.E.; Lehavi, O.; Yavetz, H.; Yogev, L. Thawed human sperm quality is influenced by the volume of the cryopreserved specimen. Fertil. Steril. 2014, 101, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Kopeika, J.; Thornhill, A.; Khalaf, Y. The effect of cryopreservation on the genome of gametes and embryos: Principles of cryobiology and critical appraisal of the evidence. Hum. Reprod. Update 2015, 21, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Najafi, A.; Asadi, E.; Moawad, A.R.; Mikaeili, S.; Amidi, F.; Adutwum, E.; Safa, M.; Sobhani, A.G. Supplementation of freezing and thawing media with brain-derived neurotrophic factor protects human sperm from freeze-thaw-induced damage. Fertil. Steril. 2016, 106, 1658–1665.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, D.; Pelloni, M.; Lenzi, A.; Lombardo, F. Cryopreservation of sperm: Effects on chromatin and strategies to prevent them. Adv. Exp. Med. Biol. 2019, 1166, 149–167. [Google Scholar] [PubMed]
- Kumar, A.; Prasad, J.K.; Srivastava, N.; Ghosh, S.K. Strategies to Minimize Various Stress-Related Freeze-Thaw Damages During Conventional Cryopreservation of Mammalian Spermatozoa. Biopreserv. Biobank 2019, 17, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Eva Mocé, A.J.F.; Graham James, K. Human sperm cryopreservation. EMJ Eur. Med J. 2016, 1, 86–91. [Google Scholar]
- Degl’Innocenti, S.; Filimberti, E.; Magini, A.; Krausz, C.; Lombardi, G.; Fino, M.G.; Rastrelli, G.; Maggi, M.; Baldi, E. Semen cryopreservation for men banking for oligospermia, cancers, and other pathologies: Prediction of post-thaw outcome using basal semen quality. Fertil. Steril. 2013, 100, 1555–1563.e1-3. [Google Scholar]
- Oehninger, S.; Duru, N.K.; Srisombut, C.; Morshedi, M. Assessment of sperm cryodamage and strategies to improve outcome. Mol. Cell. Endocrinol. 2000, 169, 3–10. [Google Scholar] [CrossRef]
- Kalthur, G.; Adiga, S.K.; Upadhya, D.; Rao, S.; Kumar, P. Effect of cryopreservation on sperm DNA integrity in patients with teratospermia. Fertil. Steril. 2008, 89, 1723–1727. [Google Scholar] [CrossRef]
- Brugnon, F.; Ouchchane, L.; Pons-Rejraji, H.; Artonne, C.; Farigoule, M.; Janny, L. Density gradient centrifugation prior to cryopreservation and hypotaurine supplementation improve post-thaw quality of sperm from infertile men with oligoasthenoteratozoospermia. Hum. Reprod. 2013, 28, 2045–2057. [Google Scholar] [CrossRef]
- Gualtieri, R.; Barbato, V.; Fiorentino, I.; Braun, S.; Rizos, D.; Longobardi, S.; Talevi, R. Treatment with zinc, d-aspartate, and coenzyme Q10 protects bull sperm against damage and improves their ability to support embryo development. Theriogenology 2014, 82, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; McPherson, N.O.; Fullston, T.; Spillane, M.; Sandeman, L.; Kang, W.X.; Zander-Fox, D.L. Oxidative stress in mouse sperm impairs embryo development, fetal growth and alters adiposity and glucose regulation in female offspring. PLoS ONE 2014, 9, e100832. [Google Scholar] [CrossRef] [Green Version]
- Barbato, V.; Talevi, R.; Braun, S.; Merolla, A.; Sudhakaran, S.; Longobardi, S.; Gualtieri, R. Supplementation of sperm media with zinc, D-aspartate and co-enzyme Q10 protects bull sperm against exogenous oxidative stress and improves their ability to support embryo development. Zygote 2017, 25, 168–175. [Google Scholar] [CrossRef]
- Ribas-Maynou, J.; Yeste, M.; Salas-Huetos, A. The relationship between sperm oxidative stress alterations and IVF/ICSI outcomes: A systematic review from nonhuman mammals. Biology 2020, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Ezzati, M.; Shanehbandi, D.; Hamdi, K.; Rahbar, S.; Pashaiasl, M. Influence of cryopreservation on structure and function of mammalian spermatozoa: An overview. Cell Tissue Bank 2020, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of apoptosis in sperm cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Salamon, S.; Maxwell, W.M.C. Frozen storage of ram semen I. Processing, freezing, thawing and fertility after cervical insemination. Anim. Reprod. Sci. 1995, 37, 185–249. [Google Scholar] [CrossRef]
- Ortega Ferrusola, C.; González Fernández, L.; Salazar Sandoval, C.; Macías García, B.; Rodríguez Martínez, H.; Tapia, J.A.; Peña, F.J. Inhibition of the mitochondrial permeability transition pore reduces "apoptosis like" changes during cryopreservation of stallion spermatozoa. Theriogenology 2010, 74, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Treulen, F.; Arias, M.E.; Aguila, L.; Uribe, P.; Felmer, R. Cryopreservation induces mitochondrial permeability transition in a bovine sperm model. Cryobiology 2018, 83, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhao, C.; Xiang, H.; Zhao, X.; Zhong, R. Melatonin inhibits formation of mitochondrial permeability transition pores and improves oxidative phosphorylation of frozen-thawed ram sperm. Front. Endocrinol. 2020, 10, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Zhao, C.; Xiang, H.; Jia, G.; Zhong, R. Melatonin improves cryopreservation of ram sperm by inhibiting mitochondrial permeability transition pore opening. Reprod. Domest. Anim. 2020, 55, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A mitochondrial targeting molecule involving mitochondrial protection and dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin protects ram spermatozoa from cryopreservation injuries in a dose-dependent manner. J. Pineal. Res. 2011, 50, 310–318. [Google Scholar] [CrossRef]
- Deng, S.L.; Sun, T.C.; Yu, K.; Wang, Z.P.; Zhang, B.L.; Zhang, Y.; Wang, X.X.; Lian, Z.X.; Liu, Y.X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef]
- Najafi, A.; Adutwum, E.; Yari, A.; Salehi, E.; Mikaeili, S.; Dashtestani, F.; Abolhassani, F.; Rashki, L.; Shiasi, S.; Asadi, E. Melatonin affects membrane integrity, intracellular reactive oxygen species, caspase3 activity and AKT phosphorylation in frozen thawed human sperm. Cell Tissue Res. 2018, 372, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Pariz, J.R.; Ranéa, C.; Monteiro, R.A.C.; Evenson, D.P.; Drevet, J.R.; Hallak, J. Melatonin and Caffeine Supplementation Used, Respectively, as Protective and Stimulating Agents in the Cryopreservation of Human Sperm Improves Survival, Viability, and Motility after Thawing compared to Traditional TEST-Yolk Buffer. Oxid. Med. Cell. Longev. 2019, 2019, 6472945. [Google Scholar] [CrossRef]
- Wang, J.Y.; Li, J.Q.; Xiao, Y.M.; Fu, B.; Qin, Z.H. Triphenylphosphonium (TPP)-based antioxidants: A new perspective on antioxidant design. ChemMedChem 2020, 15, 404–410. [Google Scholar] [CrossRef]
- Trnka, J.; Blaikie, F.H.; Smith, R.A.; Murphy, M.P. A mitochondria-targeted nitroxide is reduced to its hydroxylamine by ubiquinol in mitochondria. Free Radic. Biol. Med. 2008, 44, 1406–1419. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, Y.; Bai, H.; Liu, J.; Li, J.; Wu, B. Mitochondria-targeted antioxidant MitoTEMPO improves the post-thaw sperm quality. Cryobiology 2018, 80, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, X.; Li, J.; Xia, Q.; Gao, J.; Wu, B. Mito-Tempo alleviates cryodamage by regulating intracellular oxidative metabolism in spermatozoa from asthenozoospermic patients. Cryobiology 2019, 91, 18–22. [Google Scholar] [CrossRef]
- Ross, M.F.; Prime, T.A.; Abakumova, I.; James, A.M.; Porteous, C.M.; Smith, R.A.; Murphy, M.P. Rapid and extensive uptake and activation of hydrophobic triphenylphosphonium cations within cells. Biochem. J. 2008, 411, 633–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Bai, C.; Chen, Y.; Dai, J.; Xiang, Y.; Ji, X.; Huang, C.; Dong, Q. Inhibition of ROS production through mitochondria-targeted antioxidant and mitochondrial uncoupling increases post-thaw sperm viability in yellow catfish. Cryobiology 2014, 69.3, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, M.J.; Yu, T.H.; Cheng, Z.; Li, M.; Guo, Q.W. Mitochondria-targeted antioxidant Mitoquinone protects post-thaw human sperm against oxidative stress injury. Zhonghua Nan Ke Xue 2016, 22, 205–211. [Google Scholar]
- Szeto, H.H. First-in-class cardiolipin-protective compound as a therapeutic agent to restore mitochondrial bioenergetics. Br. J. Pharmacol. 2014, 171, 2029–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, V.E.; Bayir, H.A.; Belikova, N.A.; Kapralov, O.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Stoyanovsky, D.A.; Wipf, P.; Kochanek, P.M.; et al. Cytochrome c/cardiolipin relations in mitochondria: A kiss of death. Free Radic. Biol. Med. 2009, 46, 1439–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, M.; Phoon, C.K.; Schlame, M. Metabolism and function of mitochondrial cardiolipin. Prog. Lipid Res. 2014, 55, 1–16. [Google Scholar] [CrossRef]
- Lu, Y.W.; Claypool, S.M. Disorders of phospholipid metabolism: An emerging class of mitochondrial disease due to defects in nuclear genes. Front. Genet. 2015, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Xu, Y.; Erdjument-Bromage, H.; Donelian, A.; Phoon, C.K.L.; Terada, N.; Strathdee, D.; Neubert, T.A.; Schlame, M. Extramitochondrial cardiolipin suggests a novel function of mitochondria in spermatogenesis. J. Cell Biol. 2019, 218, 1491–1502. [Google Scholar] [CrossRef]
- Bai, H.; Zhang, Y.; Tian, S.; Hu, R.; Liang, Y.; Gao, J.; Wang, Y.; Wu, B. Elamipretide as a potential candidate for relieving cryodamage to human spermatozoa during cryopreservation. Cryobiology 2020, 95, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.; Costa, C.; Bassaizteguy, V.; Santos, M.; Cardozo, R.; Montes, J.; Settineri, R.; Nicolson, G.L. Incubation of human sperm with micelles made from glycerophospholipid mixtures increases sperm motility and resistance to oxidative stress. PLoS ONE 2018, 13, e0197897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, T.; Burdeos, G.C.; Itaya, M.; Nakagawa, K.; Miyazawa, T. Vitamin E: Regulatory redox interactions. IUBMB Life 2019, 71, 430–441. [Google Scholar] [CrossRef]
- Zhu, Z.; Fan, X.; Lv, Y.; Zhang, N.; Fan, C.; Zhang, P.; Zeng, W. Vitamin E analogue improves rabbit sperm quality during the process of cryopreservation through its antioxidative action. PLoS ONE 2015, 10, e0145383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, K.; Roberts, P.; Sanders, K.; Burton, P. Effect of antioxidant supplementation of cryopreservation medium on post-thaw integrity of human spermatozoa. Reprod. Biomed. Online 2009, 18, 184–189. [Google Scholar] [CrossRef]
- Kalthur, G.; Raj, S.; Thiyagarajan, A.; Kumar, S.; Kumar, P.; Adiga, S.K. Vitamin E supplementation in semen-freezing medium improves the motility and protects sperm from freeze-thaw-induced DNA damage. Fertil. Steril. 2011, 95, 1149–1151. [Google Scholar] [CrossRef] [PubMed]
- Safa, S.; Moghaddam, G.; Jozani, R.J.; Daghigh Kia, H.; Janmohammadi, H. Effect of vitamin E and selenium nanoparticles on post-thaw variables and oxidative status of rooster semen. Anim. Reprod. Sci. 2016, 174, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Moghbeli, M.; Kohram, H.; Zare-Shahaneh, A.; Zhandi, M.; Sharideh, H.; Sharafi, M. Effect of sperm concentration on characteristics and fertilization capacity of rooster sperm frozen in the presence of the antioxidants catalase and vitamin E. Theriogenology 2016, 86, 1393–1398. [Google Scholar] [CrossRef]
- Khellouf, A.; Benhenia, K.; Fatami, S.; Iguer-Ouada, M. The complementary effect of cholesterol and vitamin E preloaded in cyclodextrins on frozen bovine semen: Motility parameters, membrane integrity and lipid peroxidation. Cryo Lett. 2018, 39, 113–120. [Google Scholar]
- Satorre, M.M.; Breininger, E.; Cetica, P.D.; Córdoba, M. Relation between respiratory activity and sperm parameters in boar spermatozoa cryopreserved with alpha-tocopherol and selected by Sephadex. Reprod. Domest. Anim. 2018, 53, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Zerbinati, C.; Caponecchia, L.; Fiori, C.; Sebastianelli, A.; Salacone, P.; Ciacciarelli, M.; Iuliano, L. Alpha- and gamma-tocopherol levels in human semen and their potential functional implications. Andrologia 2020, 17, e13543. [Google Scholar] [CrossRef]
- Sánchez-Rubio, F.; Soria-Meneses, P.J.; Jurado-Campos, A.; Bartolomé-García, J.; Gómez-Rubio, V.; Soler, A.J.; Arroyo-Jimenez, M.M.; Santander-Ortega, M.J.; Plaza-Oliver, M.; Lozano, M.V.; et al. Nanotechnology in reproduction: Vitamin E nanoemulsions for reducing oxidative stress in sperm cells. Free Radic. Biol. Med. 2020, 160, 47–56. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Carnitines and male infertility. Reprod. Biomed. 2004, 8, 376–384. [Google Scholar] [CrossRef]
- Zhang, W.; Li, F.; Cao, H.; Li, C.; Du, C.; Yao, L.; Mao, H.; Lin, W. Protective effects of l-carnitine on astheno- and normozoospermic human semen samples during cryopreservation. Zygote 2016, 24, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Song, W.H.; Oman Ballard, J.W.; Yi, Y.J.; Sutovsky, P. Regulation of mitochondrial genome inheritance by autophagy and ubiquitin-proteasome system: Implications for health, fitness, and fertility. Biomed. Res. Int. 2014, 2014, 981867. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.F.; Murphy, M.P.; Turnbull, D.M. Do organellar genomes function as long-term redox damage sensors? Trends Genet. 2009, 25, 253–261. [Google Scholar] [CrossRef]
- Wai, T.; Teoli, D.; Shoubridge, E.A. The mitochondrial DNA genetic bottleneck results from replication of a subpopulation of genomes. Nat. Genet. 2008, 40, 1484–1488. [Google Scholar] [CrossRef] [PubMed]
- Wai, T.; Ao, A.; Zhang, X.; Cyr, D.; Dufort, D.; Shoubridge, E.A. The role of mitochondrial DNA copy number in mammalian fertility. Biol. Reprod. 2010, 83, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Reynier, P.; May-Panloup, P.; Chretien, M.-F.; Morgan, C.J.; Jean, M.; Savagner, F.; Barrière, P.; Malthièry, Y. Mitochondrial DNA content affects the fertilizability of human oocytes. Mol. Hum. Reprod. 2001, 7, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Eichenlaub-Ritter, U.; Wieczorek, M.; Luke, S.; Seidel, T. Age related changes in mitochondrial function and new approaches to study redox regulation in mammalian oocytes in response to age or maturation conditions. Mitochondrion 2011, 11, 783–796. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondria in early mammalian development. Semin. Cell. Dev. Biol. 2009, 20, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.; Burgstaller, J.; Sanchez-Nieto, J.M.; Rodríguez, T.A. The Mitochondria and the Regulation of Cell Fitness During Early Mammalian Development. Curr. Top. Dev. Biol. 2018, 128, 339–363. [Google Scholar] [PubMed]
- Trimarchi, J.R.; Liu, L.; Porterfield, D.M.; Smith, P.J.; Keefe, D.L. Oxidative phosphorylation-dependent and -independent oxygen consumption by individual preimplantation mouse embryos. Biol. Reprod. 2000, 62, 1866–1874. [Google Scholar] [CrossRef]
- Motta, P.; Nottola, S.; Makabe, S.; Heyn, R. Mitochondrial morphology in human fetal and adult female germ cells. Hum. Reprod. 2000, 15, 128–147. [Google Scholar] [CrossRef] [PubMed]
- Nagano, M.; Katagiri, S.; Takahashi, Y. ATP content and maturational/developmental ability of bovine oocytes with various cytoplasmic morphologies. Zygote 2006, 14, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Blerkom, J.; Davis, P. Mitochondrial signaling and fertilization. Mol. Hum. Reprod. 2007, 13, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Duran, H.E.; Simsek-Duran, F.; Oehninger, S.C.; Jones, H.W., Jr.; Castora, F.J. The association of reproductive senescence with mitochondrial quantity, function, and DNA integrity in human oocytes at different stages of maturation. Fertil. Steril. 2011, 96, 384–388. [Google Scholar] [CrossRef]
- Stojkovic, M.; Machado, S.A.; Stojkovic, P.; Zakhartchenko, V.; Hutzler, P.; Gonçalves, P.B.; Wolf, E. Mitochondrial distribution and adenosine triphosphate content of bovine oocytes before and after in vitro maturation: Correlation with morphological criteria and developmental capacity after in vitro fertilization and culture. Biol. Reprod. 2001, 64, 904–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Marangos, P.; Fitzharris, G.; Swann, K.; Duchen, M.; Carroll, J. Sperm-triggered [Ca2+] oscillations and Ca2+ homeostasis in the mouse egg have an absolute requirement for mitochondrial ATP production. Development 2004, 131, 3057–3067. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J.; Davis, P.; Mawhig, V.; Alexander, S. Domains of high and low polarized mitochondria in mouse and human oocytes and early embryos. Hum. Reprod. 2002, 17, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. Beobachtungen uber die oxydationsprozesse im seeigelei. Hoppe Seylers Z. Physiol. Chem. 1908, 57, 1–16. [Google Scholar] [CrossRef]
- Schatten, H.; Sun, Q.Y.; Prather, R. The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reprod. Biol. Endocrinol. 2014, 12, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Emidio, G.; Santini, S.J.; D’Alessandro, A.M.; Vetuschi, A.; Sferra, R.; Artini, P.G.; Carta, G.; Falone, S.; Amicarelli, F.; Tatone, C. SIRT1 participates in the response to methylglyoxal-dependent glycative stress in mouse oocytes and ovary. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1389–1401. [Google Scholar] [CrossRef]
- Lord, T.; Aitken, R.J. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction 2013, 146, R217–R227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsi, M.B.; Govindaraj, P.; Chawla, L.; Malhotra, N.; Singh, N.; Mittal, S.; Talwar, P.; Thangaraj, K.; Dada, R. Mitochondrial DNA variations in ova and blastocyst: Implications in assisted reproduction. Mitochondrion 2013, 13, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Fragouli, E.; Spath, K.; Alfarawati, S.; Kaper, F.; Craig, A.; Michel, C.E.; Kokocinski, F.; Cohen, J.; Munne, S.; Wells, D. Altered levels of mitochondrial DNA are associated with female age, aneuploidy, and provide an independent measure of embryonic implantation potential. PLoS Genet. 2015, 11, e1005241. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar]
- Eichenlaub-Ritter, U.; Vogt, E.; Yin, H.; Gosden, R. Spindles, mitochondria and redox potential in ageing oocytes. Reprod. Biomed. Online 2004, 8, 45–58. [Google Scholar] [CrossRef]
- Mikwar, M.; MacFarlane, A.J.; Marchetti, F. Mechanisms of oocyte aneuploidy associated with advanced maternal age. Mutat. Res. 2020, 785, 108320. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Simsek-Duran, F.; Li, F.; Ford, W.; Swanson, R.J.; Jones, H.W.; Castora, F.J. Age associated metabolic and morphologic changes in mitochondria of individual mouse and hamster oocytes. PLoS ONE 2013, 8, e64955. [Google Scholar] [CrossRef] [Green Version]
- Takeo, S.; Goto, H.; Kuwayama, T.; Monji, Y.; Iwata, H. Effect of maternal age on the ratio of cleavage and mitochondrial DNA copy number in early developmental stage bovine embryos. J. Reprod. Dev. 2013, 59, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Cree, L.M.; Hammond, E.R.; Shelling, A.N.; Berg, M.C.; Peek, J.C.; Green, M.P. Maternal age and ovarian stimulation independently affect oocyte mtDNA copy number and cumulus cell gene expression in bovine clones. Hum. Reprod. 2015, 30, 1410–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragouli, E.; McCaffrey, C.; Ravichandran, K.; Spath, K.; Grifo, J.A.; Munné, S.; Wells, D. Clinical implications of mitochondrial DNA quantification on pregnancy outcomes: A blinded prospective non-selection study. Hum. Reprod. 2017, 32, 2340–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thouas, G.A.; Trounson, A.O.; Wolvetang, E.J.; Jones, G.M. Mitochondrial dysfunction in mouse oocytes results in preimplantation embryo arrest in vitro. Biol. Reprod. 2004, 71, 1936–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarta, E.; de Los Santos, M.J.; Escribá, M.J.; Pellicer, A.; Herraiz, S. Mitochondria as a tool for oocyte rejuvenation. Fertil. Steril. 2019, 111, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeets, H.J.M. Preventing the transmission of mitochondrial DNA disorders: Selecting the good guys or kicking out the bad guys. Reprod. BioMed Online 2013, 27, 599–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, P.; Tachibana, M.; Sparman, M.; Mitalipov, S. Three-parent in vitro fertilization: Gene replacement for the prevention of inherited mitochondrial diseases. Fertil. Steril. 2014, 101, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Mitalipov, S.; Wolf, D.P. Clinical and ethical implications of mitochondrial gene transfer. Trends Endocrinol. Metab. 2014, 25, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.; Scott, R.; Alikani, M.; Schimmel, T.; Munne, S.; Levron, J.; Wu, L.; Brenner, C.; Warner, C.; Willadsen, S. Ooplasmic transfer in mature human oocytes. Mol. Hum. Reprod. 1998, 4, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Lanzendorf, S.E.; Mayer, J.F.; Toner, J.; Oehninger, S.; Saffan, D.S.; Muasher, S. Pregnancy following transfer of ooplasm from cryopreserved-thawed donor oocytes into recipient oocytes. Fertil. Steril. 1999, 71, 575–577. [Google Scholar] [CrossRef]
- Barritt, J.A.; Brenner, C.A.; Malter, H.E.; Cohen, J. Mitochondria in human offspring derived from ooplasmic transplantation. Hum. Reprod. 2001, 16, 513–516. [Google Scholar] [CrossRef]
- Mobarak, H.; Heidarpour, M.; Tsai, P.J.; Rezabakhsh, A.; Rahbarghazi, R.; Nouri, M.; Mahdipour, M. Autologous mitochondrial microinjection; a strategy to improve the oocyte quality and subsequent reproductive outcome during aging. Cell Biosci. 2019, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Potdar, N.; Gelbaya, T.A.; Nardo, L.G. Oocyte vitrification in the 21st century and post-warming fertility outcomes: A systematic review and meta-analysis. Reprod. Biomed. Online 2014, 29, 159–176. [Google Scholar] [CrossRef] [Green Version]
- Glujovsky, D.; Riestra, B.; Sueldo, C.; Fiszbajn, G.; Repping, S.; Nodar, F.; Papier, S.; Ciapponi, A. Vitrification versus slow freezing for women undergoing oocyte cryopreservation. Cochrane Database Syst. Rev. 2014, 5, CD010047. [Google Scholar] [CrossRef]
- Kool, E.M.; Bos, A.M.E.; van der Graaf, R.; Fauser, B.C.J.M.; Bredenoord, A.L. Ethics of oocyte banking for third-party assisted reproduction: A systematic review. Hum. Reprod. Update 2018, 24, 615–635. [Google Scholar] [CrossRef]
- Chen, C. Pregnancy after human oocyte cryopreservation. Lancet 1986, 1, 884–886. [Google Scholar] [CrossRef]
- Van Uem, J.F.; Siebzehnrubl, E.R.; Schuh, B.; Koch, R.; Trotnow, S.; Lang, N. Birth after cryopreservation of unfertilized oocytes. Lancet 1987, 1, 752–753. [Google Scholar] [CrossRef]
- Kuwayama, M.; Vajta, G.; Kato, O.; Leibo, S.P. Highly efficient vitrification method for cryopreservation of human oocytes. Reprod. Biomed. Online 2005, 11, 300–308. [Google Scholar] [CrossRef]
- Practice Committees of American Society for Reproductive Medicine and Society for Assisted Reproductive Technology. Mature oocyte cryopreservation: A guideline. Fertil. Steril. 2013, 99, 37–43. [Google Scholar] [CrossRef]
- Cao, Y.X.; Xing, Q.; Li, L.; Cong, L.; Zhang, Z.G.; Wei, Z.L.; Zhou, P. Comparison of survival and embryonic development in human oocytes cryopreserved by slow-freezing and vitrification. Fertil. Steril. 2009, 92, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Serafini, P.C.; Fioravanti, J.; Yadid, I.; Coslovsky, M.; Hassun, P.; Alegretti, J.R.; Motta, E.L. Prospective randomized comparison of human oocyte cryopreservation with slow-rate freezing or vitrification. Fertil. Steril. 2010, 94, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Cobo, A.; Diaz, C. Clinical application of oocyte vitrification: A systematic review and meta-analysis of randomized controlled trials. Fertil. Steril. 2011, 96, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Edgar, D.H.; Gook, D.A. A critical appraisal of cryopreservation (slow cooling versus vitrification) of human oocytes and embryos. Hum. Reprod. Update 2012, 18, 536–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cil, A.P.; Bang, H.; Oktay, K. Age-specific probability of live birth with oocyte cryopreservation: An individual patient data meta-analysis. Fertil. Steril. 2013, 100, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, M.; McClure, N.; Lewis, S.E.M. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef]
- Dalcin, L.; Silva, R.C.; Paulini, F.; Silva, B.D.M.; Neves, J.P.; Lucci, C.M. Cytoskeleton structure, pattern of mitochondrial activity and ultrastructure of frozen or vitrified sheep embryos. Cryobiology 2013, 67, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, E.; Valdebenito, I.; Zepeda, A.B.; Figueroa, C.A.; Dumorne´, K.; Castillo, R.L.; Farias, J.G. Effects of cryopreservation on mitochondria of fish spermatozoa. Rev. Aquacult. 2017, 9, 76–87. [Google Scholar] [CrossRef]
- Jones, A.; Van Blerkom, J.; Davis, P.; Toledo, A.A. Cryopreservation of metaphase II human oocytes effects mitochondrial membrane potential: Implications for developmental competence. Hum. Reprod. 2004, 19, 1861–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualtieri, R.; Iaccarino, M.; Mollo, V.; Prisco, M.; Iaccarino, S.; Talevi, R. Slow cooling of human oocytes: Ultrastructural injuries and apoptotic status. Fertil. Steril. 2009, 91, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Guo, N.; Tan, M.H.; Li, Y.F. Effect of mouse oocyte vitrification on mitochondrial membrane potential and distribution. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2014, 34, 99–102. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Barbaro, R.; Vento, M.; Ciriminna, R.; Artini, P.G. Effects of reproductive aging and postovulatory aging on the maintenance of biological competence after oocyte vitrification: Insights from the mouse model. Theriogenology 2011, 76, 864–873. [Google Scholar] [CrossRef]
- Nohales-Córcoles, M.; Sevillano-Almerich, G.; Di Emidio, G.; Tatone, C.; Cobo, A.C.; Dumollard, R.; De Los Santos Molina, M.J. Impact of vitrification on the mitochondrial activity and redox homeostasis of human oocyte. Hum. Reprod. 2016, 31, 1850–1858. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Du, W.H.; Wang, D.; Hao, H.S.; Liu, Y.; Qin, T.; Zhu, H.B. Effect of cyclosporine pretreatment on mitochondrial function in vitrified bovine mature oocytes. Fertil. Steril. 2011, 95, 2786–2788. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Hao, H.S.; Li, C.Y.; Zhao, Y.H.; Wang, H.Y.; Yan, C.L.; Du, W.H.; Wang, D.; Liu, Y.; Pang, Y.W.; et al. Calcium ion regulation by BAPTA-AM and ruthenium red improved the fertilisation capacity and developmental ability of vitrified bovine oocytes. Sci. Rep. 2017, 7, 10652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Hammett, M.F.; Cho, M.S. Multifaceted freezing injury in human polymorphonuclear cells at high subfreezing temperatures. Cryobiology 1985, 22, 215–236. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z.Y.; Cai, M.D.; Li, X.X.; Li, Y.H.; Lei, Y.; Yu, X.L. Effect of vitrification temperature and cryoprotectant concentrations on the mRNA transcriptome of bovine mature oocytes after vitrifying at immature stage. Theriogenology 2020, 148, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Daddangadi, A.; Uppangala, S.; Kalthur, G.; Talevi, R.; Adiga, S.K. Germinal stage vitrification is superior to MII stage vitrification in prepubertal mouse oocytes. Cryobiology 2020, 93, 49–55. [Google Scholar] [CrossRef]
- Liu, M.H.; Zhou, W.H.; Chu, D.P.; Fu, L.; Sha, W.; Li, Y. Ultrastructural changes and methylation of human oocytes vitrified at the germinal vesicle stage and matured in vitro after thawing. Gynecol. Obstet. Investig. 2017, 82, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Han, S.; Liu, W.; Wang, Y.; Huang, G. Effect of vitrification on mitochondrial membrane potential in human metaphase II oocytes. J. Assist. Reprod. Genet. 2012, 29, 1045–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larman, M.G.; Sheehan, C.B.; Gardner, D.K. Calcium-free vitrification reduces cryoprotectant-induced zona pellucida hardening and increases fertilization rates in mouse oocytes. Reproduction 2006, 131, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Larman, M.G.; Katz-Jaffe, M.G.; Sheehan, C.B.; Gardner, D.K. 1,2-propanediol and the type of cryopreservation procedure adversely affect mouse oocyte physiology. Hum. Reprod. 2007, 22, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Igarashi, H.; Doshida, M.; Takahashi, K.; Nakahara, K.; Tezuka, N.; Kurachi, H. Lowering intracellular and extracellular calcium contents prevents cytotoxic effects of ethylene glycol-based vitrification solution in unfertilized mouse oocytes. Mol. Reprod. Dev. 2004, 68, 250–258. [Google Scholar] [CrossRef]
- Marques, C.C.; Santos-Silva, C.; Rodrigues, C.; Matos, J.E.; Moura, T.; Baptista, M.C.; Horta, A.E.M.; Bessa, R.J.B.; Alves, S.P.; Soveral, G.; et al. Bovine oocyte membrane permeability and cryosurvival: Effects of different cryoprotectants and calcium in the vitrification media. Cryobiology 2018, 81, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Budd, S.L.; Nicholls, D.G. Mitochondria, calcium regulation, and acute glutamate excitotoxicity in cultured cerebellar granule cells. J. Neurochem. 1996, 67, 2282–2291. [Google Scholar] [CrossRef]
- Galluzzi, L.; Morselli, E.; Kepp, O.; Kroemer, G. Targeting post-mitochondrial effectors of apoptosis for neuroprotection. Biochim. Biophys. Acta 2009, 1787, 402–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrod, J.W.; Wong, R.; Mishra, S.; Vagnozzi, R.J.; Sakthievel, B.; Goonasekera, S.A.; Karch, J.; Gabel, S.; Farber, J.; Force, T.; et al. Cyclophilin D controls mitochondrial pore-dependent Ca(2+) exchange, metabolic flexibility, and propensity for heart failure in mice. J. Clin. Investig. 2010, 120, 3680–3687. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Ouyang, X.; Wan, R.; Cheng, H.; Mattson, M.P.; Cheng, A. Mitochondrial superoxide production negatively regulates neural progenitor proliferation and cerebral cortical development. Stem Cells 2012, 30, 2535–2547. [Google Scholar] [CrossRef] [Green Version]
- Ozil, J.P.; Huneau, D. Activation of rabbit oocytes: The impact of the Ca2+ signal regime on development. Development 2001, 128, 917–928. [Google Scholar] [PubMed]
- Ducibella, T.; Huneau, D.; Angelichio, E.; Xu, Z.; Schultz, R.M.; Kopf, G.S.; Fissore, R.; Madoux, S.; Ozil, J.P. Egg-to-embryo transition is driven by differential responses to Ca(2+) oscillation number. Dev. Biol. 2002, 250, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Ozil, J.P.; Banrezes, B.; Toth, S.; Pan, H.; Schultz, R.M. Ca2+oscillatory pattern in fertilized mouse eggs affects gene expression and development to term. Dev. Biol. 2006, 300, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Borini, A.; Sciajno, R.; Bianchi, V.; Sereni, E.; Flamigni, C.; Coticchio, G. Clinical outcome of oocyte cryopreservation after slow cooling with a protocol utilizing a high sucrose concentration. Hum. Reprod. 2006, 21, 512–517. [Google Scholar] [CrossRef] [Green Version]
- La Sala, G.B.; Nicoli, A.; Villani, M.T.; Pescarini, M.; Gallinelli, A.; Blickstein, I. Outcome of 518 salvage oocyte-cryopreservation cycles performed as a routine procedure in an in vitro fertilization program. Fertil. Steril. 2006, 86, 1423–1427. [Google Scholar] [CrossRef]
- Levi Setti, P.E.; Albani, E.; Novara, P.V.; Cesana, A.; Morreale, G. Cryopreservation of supernumerary oocytes in IVF/ICSI cycles. Hum. Reprod. 2006, 21, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Gualtieri, R.; Mollo, V.; Barbato, V.; Fiorentino, I.; Iaccarino, M.; Talevi, R. Ultrastructure and intracellular calcium response during activation in vitrified and slow-frozen human oocytes. Hum. Reprod. 2011, 26, 2452–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.Y.; Yoon, S.Y.; Cha, S.K.; Kwak, K.H.; Fissore, R.A.; Parys, J.B.; Yoon, T.K.; Lee, D.R. Alterations in calcium oscillatory activity in vitrified mouse eggs impact on egg quality and subsequent embryonic development. Pflug. Arch. 2011, 461, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Bonte, D.; Thys, V.; De Sutter, P.; Boel, A.; Leybaert, L.; Heindryckx, B. Vitrification negatively affects the Ca2+-releasing and activation potential of mouse oocytes, but vitrified oocytes are potentially useful for diagnostic purposes. Reprod. Biomed. Online 2020, 40, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Wu, C.; Muneri, C.W.; Niu, Y.; Zhang, S.; Rui, R.; Zhang, D. Changes in mitochondrial function in porcine vitrified MII-stage oocytes and their impacts on apoptosis and developmental ability. Cryobiology 2015, 71, 291–298. [Google Scholar] [CrossRef]
- Dai, J.J.; Yang, J.H.; Zhang, S.S.; Niu, Y.F.; Chen, Y.N.; Wu, C.F.; Zhang, D.F. Partial recovery of mitochondrial function of vitrified porcine MII stage oocytes during post-thaw incubation. Cryo Lett. 2018, 39, 39–44. [Google Scholar]
- Karlsson, J.O.; Eroglu, A.; Toth, T.L.; Cravalho, E.G.; Toner, M. Fertilization and development of mouse oocytes cryopreserved using a theoretically optimized protocol. Hum. Reprod. 1996, 11, 1296–1305. [Google Scholar] [CrossRef] [Green Version]
- Oktay, K.; Cil, A.P.; Bang, H. Efficiency of oocyte cryopreservation: A meta-analysis. Fertil Steril. 2006, 86, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, B.; Szurek, E.A.; Schall, P.; Latham, K.E.; Eroglu, A. Probing lasting cryoinjuries to oocyte-embryo transcriptome. PLoS ONE 2020, 15, e0231108. [Google Scholar] [CrossRef] [Green Version]
- Ruihuan, G.; Zhichao, L.; Song, G.; Jing, F.; Yijuan, S.; Xiaoxi, S. Oocyte vitrification temporarily turns on oxidation-reduction process genes in mouse preimplantation embryos. Reprod. Sci. 2020. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Arnanz, A.; De Munck, N.; Bayram, A.; El-Damen, A.; Abdalla, A.; ElKhatib, I.; Melado, L.; Lawrenz, B.; Fatemi, H.M. Blastocyst mitochondrial DNA (mtDNA) is not affected by oocyte vitrification: A sibling oocyte study. J. Assist. Reprod. Genet. 2020, 37, 1387–1397. [Google Scholar] [CrossRef]
- Dai, J.J.; Niu, Y.F.; Wu, C.F.; Zhang, S.H.; Zhang, D.F. Both death receptor and mitochondria mediated apoptotic pathways participated the occurrence of apoptosis in porcine vitrified mii stage oocytes. Cryo Lett. 2016, 37, 129–136. [Google Scholar]
- Pan, B.; Yang, H.; Wu, Z.; Qazi, I.H.; Liu, G.; Han, H.; Meng, Q.; Zhou, G. Melatonin Improves Parthenogenetic Development of Vitrified–Warmed Mouse Oocytes Potentially by Promoting G1/S Cell Cycle Progression. Int. J. Mol. Sci. 2018, 19, 4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukunoki, A.; Takeo, T.; Nakagata, N. N-acetyl cysteine restores the fertility of vitrified-warmed mouse oocytes derived through ultrasuperovulation. PLoS ONE 2019, 14, e0224087. [Google Scholar] [CrossRef] [PubMed]
- Borjizadeh, A.; Ahmadi, H.; Daneshi, E.; Roshani, D.; Fathi, F.; Abdi, M.; Nasseri, S.; Abouzaripour, M. The effect of adding rosmarinic and ascorbic acids to vitrification media on fertilization rate of the mice oocyte: An experimental study. Int. J. Reprod. Biomed. 2019, 17, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, I.; Tagiri, M.; Ogawa, H.; Tashima, K.; Takashima, S.; Hara, H.; Hirabayashi, M.; Hochi, S. High revivability of vitrified-warmed bovine mature oocytes after recovery culture with α-tocopherol. Reproduction 2015, 149, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Moawad, A.R.; Tan, S.L.; Xu, B.; Chen, H.Y.; Taketo, T. L-carnitine supplementation during vitrification of mouse oocytes at the germinal vesicle stage improves preimplantation development following maturation and fertilization in vitro. Biol. Reprod. 2013, 88, 104. [Google Scholar] [CrossRef] [PubMed]
- Trapphoff, T.; Heiligentag, M.; Simon, J.; Staubach, N.; Seidel, T.; Otte, K.; Fröhlich, T.; Arnold, G.J.; Eichenlaub-Ritter, U. Improved cryotolerance and developmental potential of in vitro and in vivo matured mouse oocytes by supplementing with a glutathione donor prior to vitrification. Mol. Hum. Reprod. 2016, 22, 867–881. [Google Scholar]
- García-Martínez, T.; Vendrell-Flotats, M.; Martínez-Rodero, I.; Ordóñez-León, E.A.; Álvarez-Rodríguez, M.; López-Béjar, M.; Yeste, M.; Mogas, T. Glutathione ethyl ester protects in vitro-maturing bovine oocytes against oxidative stress induced by subsequent vitrification/warming. Int. J. Mol. Sci. 2020, 21, 7547. [Google Scholar] [CrossRef]
- Li, Z.; Gu, R.; Lu, X.; Zhao, S.; Feng, Y.; Sun, Y. Preincubation with glutathione ethyl ester improves the developmental competence of vitrified mouse oocytes. J. Assist. Reprod. Genet. 2018, 35, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Rashedi, A.S.; de Roo, S.F.; Ataman, L.M.; Edmonds, M.E.; Silva, A.A.; Scarella, A.; Horbaczewska, A.; Anazodo, A.; Arvas, A.; Ramalho de Carvalho, B.; et al. Survey of fertility preservation options available to patients with cancer around the globe. JCO Glob. Oncol. 2020, 6, JGO.2016.008144. [Google Scholar] [CrossRef]
- Eyden, B.; Radford, J.; Shalet, S.M.; Thomas, N.; Brison, D.R.; Lieberman, B.A. Ultrastructural preservation of ovarian cortical tissue cryopreserved in dimethylsulfoxide for subsequent transplantation into young female cancer patients. Ultrastruct. Pathol. 2004, 28, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gook, D.A.; Edgar, D.H.; Stern, C. The effects of cryopreservation regimens on the morphology of human ovarian tissue. Mol. Cell. Endocrinol. 2000, 169, 99–103. [Google Scholar] [CrossRef]
- Salehnia, M.; Abbasian, M.E.; Rezazadeh, V.M. Ultrastructure of follicles after vitrification of mouse ovarian tissue. Fertil. Steril. 2002, 78, 644–645. [Google Scholar] [CrossRef]
- Talevi, R.; Barbato, V.; Mollo, V.; Fiorentino, I.; De Stefano, C.; Guarino, F.M.; Gualtieri, R. Replacement of sodium with choline in slow-cooling media improves human ovarian tissue cryopreservation. Reprod. Biomed. Online 2013, 27, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, R.; Vicenti, R.; Macciocca, M.; Martino, N.A.; Dell’Aquila, M.E.; Pasquinelli, G.; Morselli-Labate, A.M.; Seracchioli, R.; Paradisi, R. Morphological, ultrastructural and functional imaging of frozen/thawed and vitrified/warmed human ovarian tissue retrieved from oncological patients. Hum. Reprod. 2016, 31, 1838–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talevi, R.; Barbato, V.; Fiorentino, I.; Braun, S.; De Stefano, C.; Ferraro, R.; Sudhakaran, S.; Gualtieri, R. Successful slush nitrogen vitrification of human ovarian tissue. Fertil. Steril. 2016, 105, 1523–1531.e1. [Google Scholar] [CrossRef] [Green Version]
- Barbato, V.; Gualtieri, R.; Capriglione, T.; Pallotta, M.M.; Braun, S.; Di Nardo, M.; Costanzo, V.; Ferraro, R.; Catapano, G.; Talevi, R. Slush nitrogen vitrification of human ovarian tissue does not alter gene expression and improves follicle health and progression in long-term in vitro culture. Fertil. Steril. 2018, 110, 1356–1366. [Google Scholar] [CrossRef]
- Mathias, F.J.; D’Souza, F.; Uppangala, S.; Salian, S.R.; Kalthur, G.; Adiga, S.K. Ovarian tissue vitrification is more efficient than slow freezing in protecting oocyte and granulosa cell DNA integrity. Syst. Biol. Reprod. Med. 2014, 60, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Salehnia, M.; Töhönen, V.; Zavareh, S.; Inzunza, J. Does cryopreservation of ovarian tissue affect the distribution and function of germinal vesicle oocytes mitochondria? Biomed. Res. Int. 2013, 2013, 489032. [Google Scholar] [CrossRef]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin inhibits oxidative stress and apoptosis in cryopreserved ovarian tissues via Nrf2/HO-1 signaling pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef]
- Arkoun, B.; Galas, L.; Dumont, L.; Rives, A.; Saulnier, J.; Delessard, M.; Rondanino, C.; Rives, N. Vitamin E but not GSH decreases reactive oxygen species accumulation and enhances sperm production during in vitro maturation of frozen-thawed prepubertal mouse testicular tissue. Int. J. Mol. Sci. 2019, 20, 5380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.G.; Wang, Y.H.; Han, C.; Hu, S.; Wang, L.Q.; Hu, J.H. Effects of trehalose supplementation on cell viability and oxidative stress variables in frozen-thawed bovine calf testicular tissue. Cryobiology 2015, 70, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, A.E.; Taha, E.A.; Younis, A.; Fakhry, M.E.; Morsy, H. Testicular tissue oxidative stress in azoospermic patients: Effect of cryopreservation. Andrologia 2020, 52, e13817. [Google Scholar] [CrossRef] [PubMed]
- Keros, V.; Rosenlund, B.; Hultenby, K.; Aghajanova, L.; Levkov, L.; Hovatta, O. Optimizing cryopreservation of human testicular tissue: Comparison of protocols with glycerol, propanediol and dimethylsulphoxide as cryoprotectants. Hum. Reprod. 2005, 20, 1676–1687. [Google Scholar] [CrossRef] [Green Version]
- Ha, S.J.; Kim, B.G.; Lee, Y.A.; Kim, Y.H.; Kim, B.J.; Jung, S.E.; Pang, M.G.; Ryu, B.Y. Effect of antioxidants and apoptosis inhibitors on cryopreservation of murine germ cells enriched for spermatogonial stem cells. PLoS ONE 2016, 11, e0161372. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, R.; Pasquinelli, G.; Bracone, G.; Orrico, C.; Paradisi, R.; Seraccioli, R.; Venturoli, S. Fetal calf serum versus human serum: Ultrastructural evaluation of protein support influence on human ovarian tissue cryopreservation. Ultrastruct. Pathol. 2006, 30, 253–260. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gualtieri, R.; Kalthur, G.; Barbato, V.; Di Nardo, M.; Adiga, S.K.; Talevi, R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants 2021, 10, 337. https://doi.org/10.3390/antiox10030337
Gualtieri R, Kalthur G, Barbato V, Di Nardo M, Adiga SK, Talevi R. Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells. Antioxidants. 2021; 10(3):337. https://doi.org/10.3390/antiox10030337
Chicago/Turabian StyleGualtieri, Roberto, Guruprasad Kalthur, Vincenza Barbato, Maddalena Di Nardo, Satish Kumar Adiga, and Riccardo Talevi. 2021. "Mitochondrial Dysfunction and Oxidative Stress Caused by Cryopreservation in Reproductive Cells" Antioxidants 10, no. 3: 337. https://doi.org/10.3390/antiox10030337