
Myo/Nog Cells Increase in Response to Elevated Intraocular Pressure and Mitigate Ganglion Cell Death in a Mouse Model of Glaucoma
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Induction of Glaucomatous State
2.3. M/N Cell Extraction and Labeling
2.4. Treatment Groups
2.5. Embedding and Sectioning
2.6. Quantitation of RGCs and Measurement of GCL-NFL Thickness
2.7. Immunofluorescence Microscopy
2.8. Statistical Analyses
3. Results
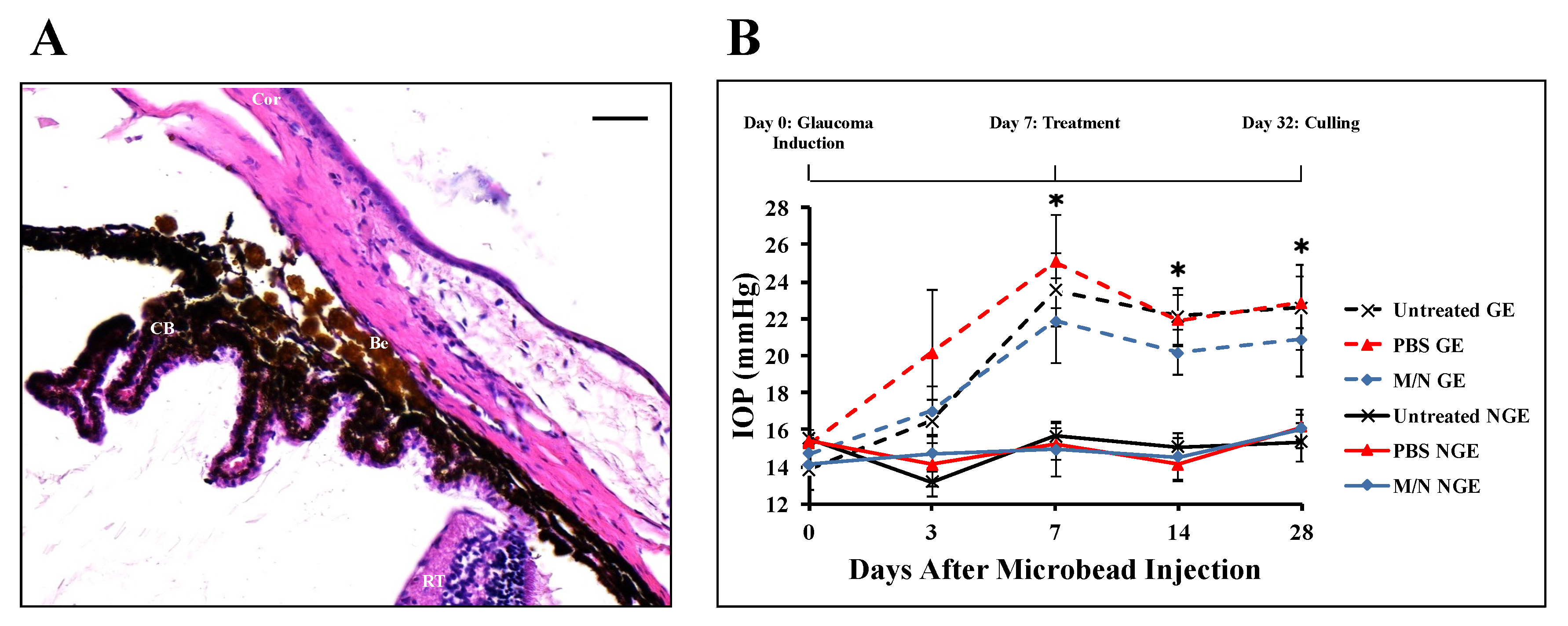
3.1. Effect of Injecting Beads into the AC on IOP
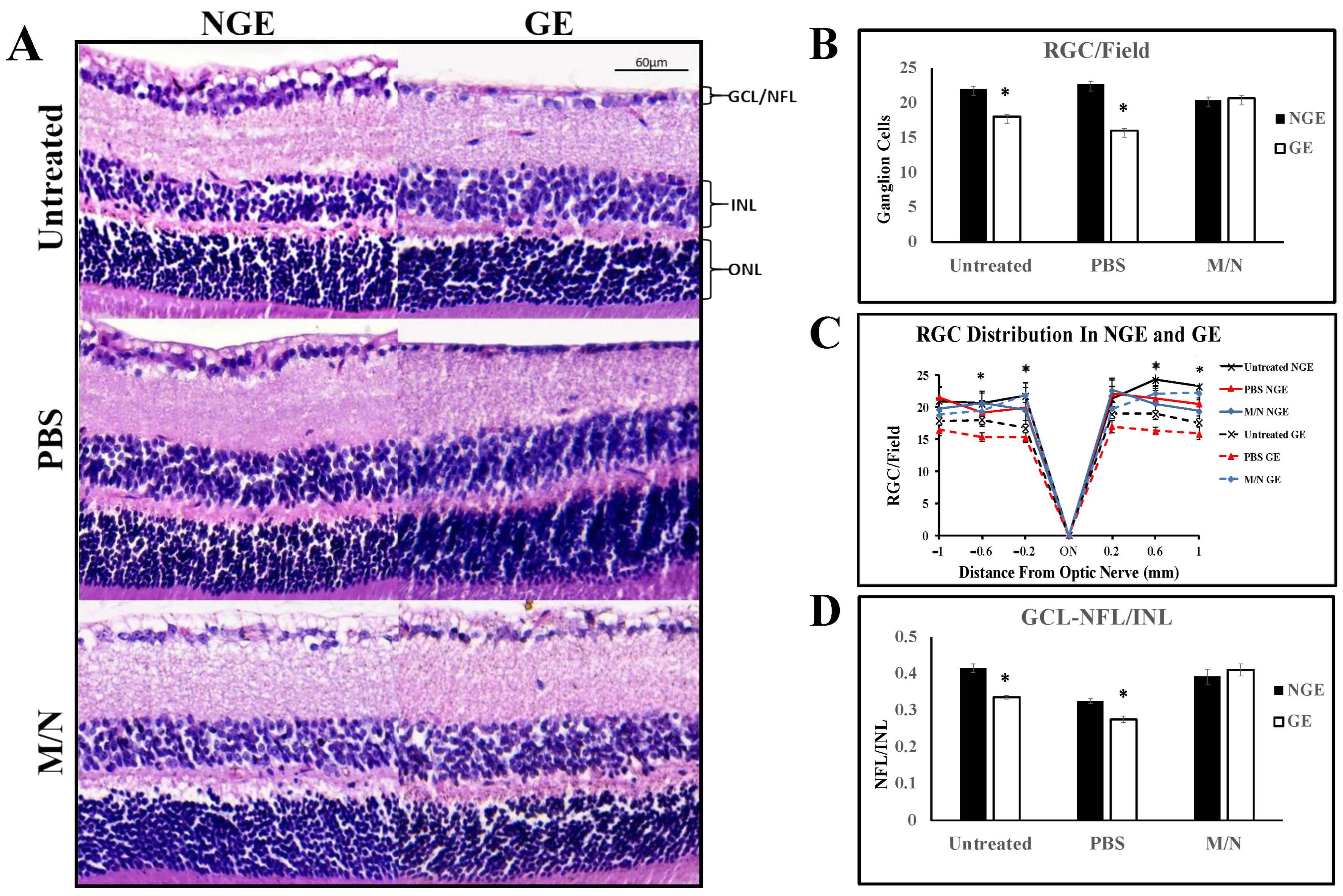
3.2. Effects of Elevated IOP on the Retina
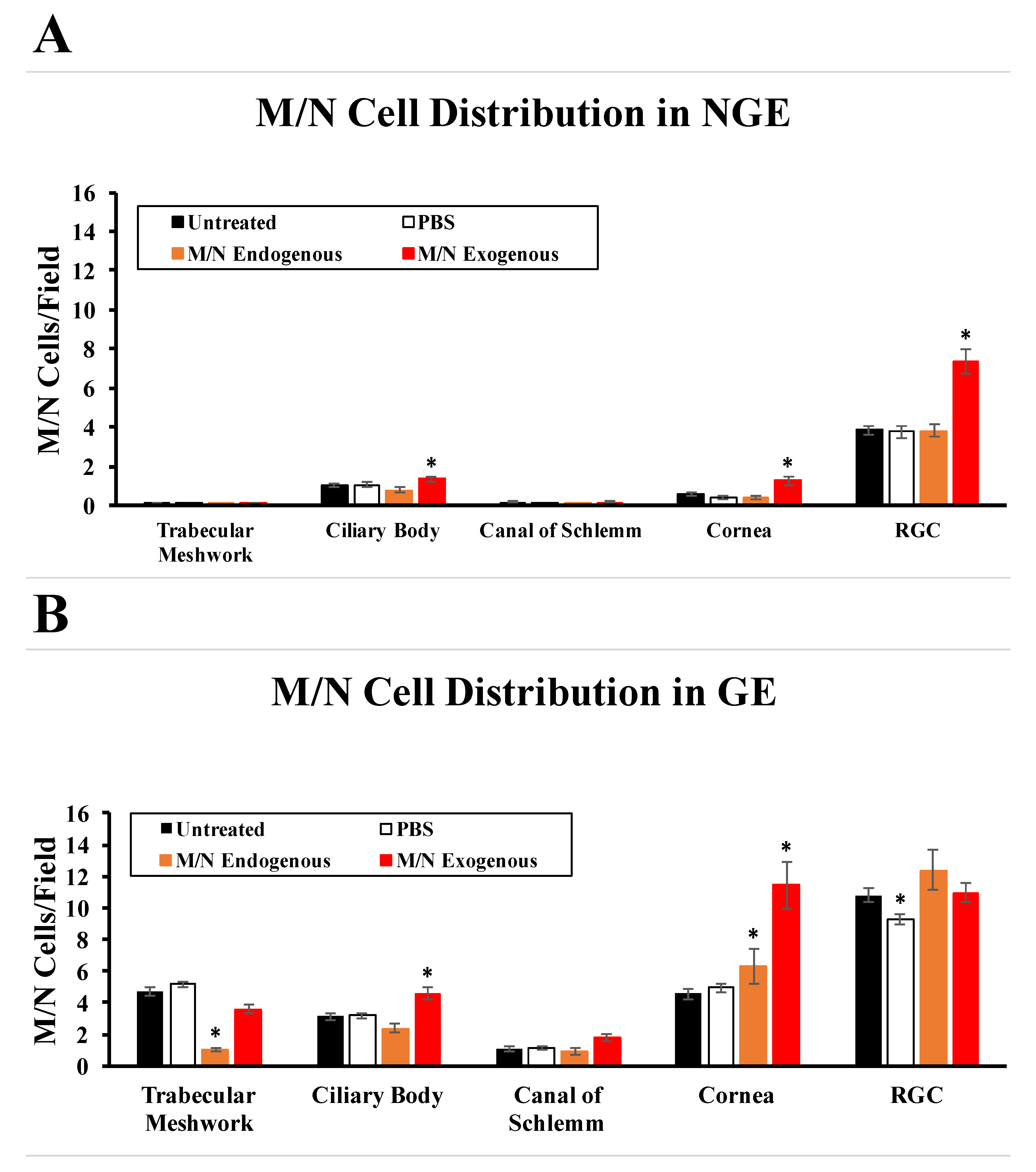
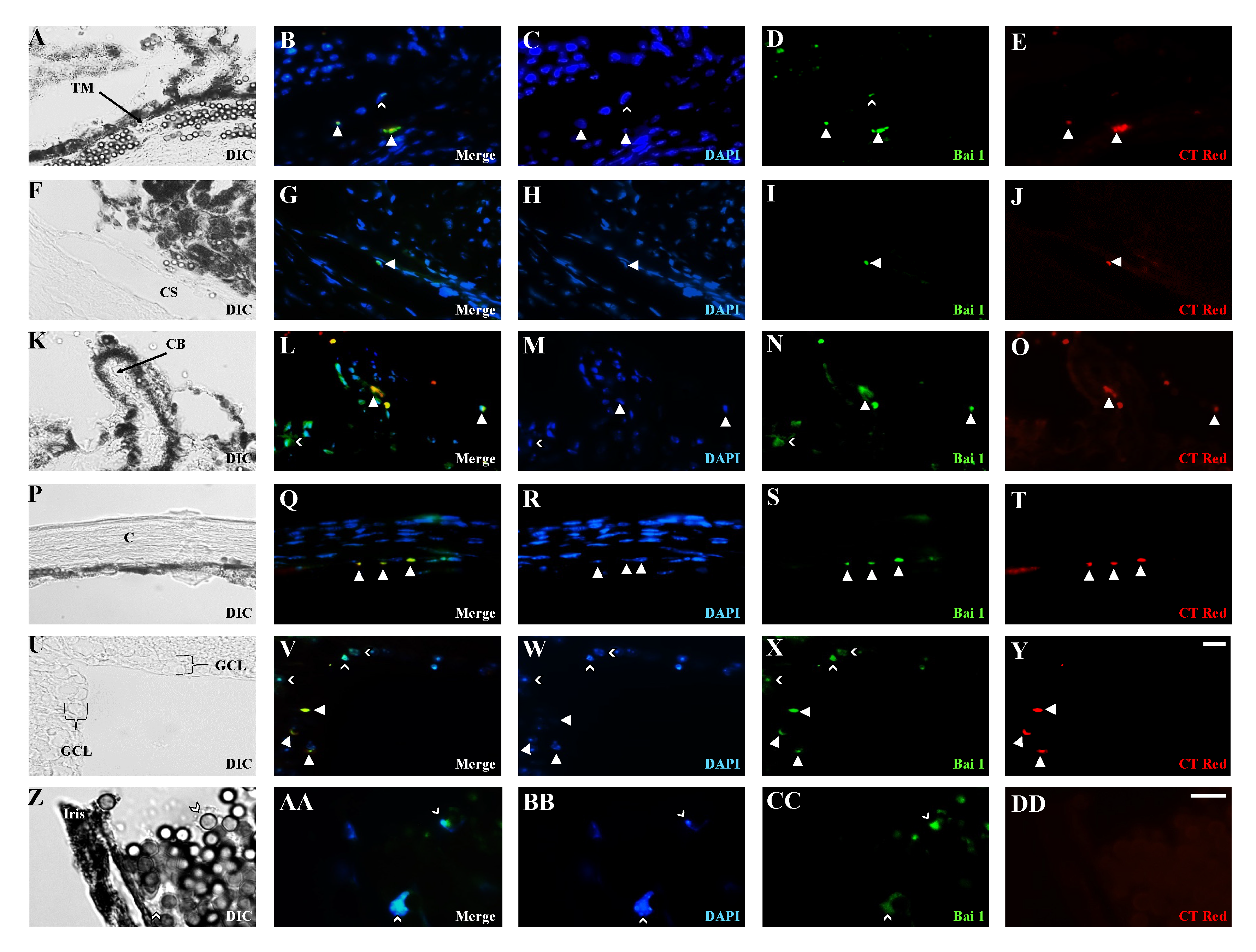
3.3. Effects of Elevated IOP on the Distribution of Endogenous M/N Cells
3.4. Distribution of Exogenous M/N Cells in Glaucomatous and Non-Glaucomatous Eyes
3.5. Effects of Exogenous M/N Cells on Endogenous M/N Cells, IOP, and RGCs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IOP | Intraocular Pressure |
| AC | Anterior Chamber |
| BAI1 | Brain-Specific Angiogenesis Inhibitor 1 |
| + | Positive |
| RGC | Retinal Ganglion Cell |
| NFL/INL | Nerve Fiber Layer/Inner Nuclear Layer |
References
- Parihar, J.K. Glaucoma: The ‘Black hole’ of irreversible blindness. Med. J. Armed Forces India 2016, 72, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Varma, R.; Peeples, P.; Walt, J.G.; Bramley, T.J. Disease progression and the need for neuroprotection in glaucoma management. Am. J. Manag. Care 2008, 14, S15–S19. [Google Scholar] [PubMed]
- Baudouin, C.; Kolko, M.; Melik-Parsadaniantz, S.; Messmer, E.M. Inflammation in Glaucoma: From the back to the front of the eye, and beyond. Prog. Retin. Eye Res. 2021, 83, 100916. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Tirendi, S.; Bassi, A.M.; Traverso, C.E.; Sacca, S.C. Neuroinflammation in Primary Open-Angle Glaucoma. J. Clin. Med. 2020, 9, 3172. [Google Scholar] [CrossRef] [PubMed]
- Baudouin, C.; Denoyer, A.; Rostene, W. Glaucoma today: Detection and therapeutic progress. Biol. Aujourdhui 2013, 207, 87–95. [Google Scholar] [CrossRef]
- Anholt, R.R.; Carbone, M.A. A molecular mechanism for glaucoma: Endoplasmic reticulum stress and the unfolded protein response. Trends Mol. Med. 2013, 19, 586–593. [Google Scholar] [CrossRef]
- Fingert, J.H. Primary open-angle glaucoma genes. Eye 2011, 25, 587–595. [Google Scholar] [CrossRef]
- Kwon, Y.H.; Fingert, J.H.; Kuehn, M.H.; Alward, W.L. Primary open-angle glaucoma. N. Engl. J. Med. 2009, 360, 1113–1124. [Google Scholar] [CrossRef]
- Kanagavalli, J.; Pandaranayaka, E.; Krishnadas, S.R.; Krishnaswamy, S.; Sundaresan, P. A review of genetic and structural understanding of the role of myocilin in primary open angle glaucoma. Indian. J. Ophthalmol. 2004, 52, 271–280. [Google Scholar]
- Ray, K.; Mukhopadhyay, A.; Acharya, M. Recent advances in molecular genetics of glaucoma. Mol. Cell Biochem. 2003, 253, 223–231. [Google Scholar] [CrossRef]
- WuDunn, D. Genetic basis of glaucoma. Curr. Opin. Ophthalmol. 2002, 13, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Khazaeni, B.; Khazaeni, L. Acute Closed Angle Glaucoma; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Dave, S.D.; Meyer, J.J. Chronic Closed Angle Glaucoma; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Jindra, L.F. Closed-angle glaucoma: Diagnosis and management. Hosp. Pract. 1984, 19, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Mapstone, R. The syndrome of closed-angle glaucoma. Br. J. Ophthalmol. 1976, 60, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.S.; Penteado, R.C.; Papoyan, V.; Voskanyan, L.; Weinreb, R.N. Aqueous Angiographic Outflow Improvement after Trabecular Microbypass in Glaucoma Patients. Ophthalmol. Glaucoma 2019, 2, 11–21. [Google Scholar] [CrossRef]
- Ianchulev, T.; Weinreb, R.N.; Kamthan, G.; Calvo, E.; Pamnani, R.; Ahmed, I.K. Biotissue stent for supraciliary outflow in open-angle glaucoma patients: Surgical procedure and first clinical results of an aqueous drainage biostent. Br. J. Ophthalmol. 2023, 1–6. [Google Scholar] [CrossRef]
- Evangelho, K.; Mogilevskaya, M.; Losada-Barragan, M.; Vargas-Sanchez, J.K. Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: A review of the literature. Int. Ophthalmol. 2019, 39, 259–271. [Google Scholar] [CrossRef]
- Jayaram, H.; Kolko, M.; Friedman, D.S.; Gazzard, G. Glaucoma: Now and beyond. Lancet 2023, 402, 1788–1801. [Google Scholar] [CrossRef]
- Nobl, M.; Priglinger, S.G. Glaucoma—An overview for general practitioners. MMW Fortschr. Med. 2023, 165, 36–38. [Google Scholar] [CrossRef]
- Brazuna, R.; Alonso, R.S.; Salomao, M.Q.; Fernandes, B.F.; Ambrosio, R., Jr. Ocular Biomechanics and Glaucoma. Vision 2023, 7, 36. [Google Scholar] [CrossRef]
- Hsu, E.; Desai, M. Glaucoma and Systemic Disease. Life 2023, 13, 1018. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.; Chakrabarti, A.; Nazm, N.; Mehta, R.; Edward, D.P. Newer advances in medical management of glaucoma. Indian J. Ophthalmol. 2022, 70, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Lusthaus, J.; Goldberg, I. Current management of glaucoma. Med. J. Aust. 2019, 210, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.A.; Belforte, N.; Cueva Vargas, J.L.; Di Polo, A. A Magnetic Microbead Occlusion Model to Induce Ocular Hypertension-Dependent Glaucoma in Mice. J. Vis. Exp. 2016, 109, e53731. [Google Scholar] [CrossRef]
- Samsel, P.A.; Kisiswa, L.; Erichsen, J.T.; Cross, S.D.; Morgan, J.E. A novel method for the induction of experimental glaucoma using magnetic microspheres. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1671–1675. [Google Scholar] [CrossRef]
- Gerhart, J.; Baytion, M.; DeLuca, S.; Getts, R.; Lopez, C.; Niewenhuis, R.; Nilsen, T.; Olex, S.; Weintraub, H.; George-Weinstein, M. DNA dendrimers localize MyoD mRNA in presomitic tissues of the chick embryo. J. Cell Biol. 2000, 149, 825–834. [Google Scholar] [CrossRef]
- Gerhart, J.; Bast, B.; Neely, C.; Iem, S.; Amegbe, P.; Niewenhuis, R.; Miklasz, S.; Cheng, P.F.; George-Weinstein, M. MyoD-positive myoblasts are present in mature fetal organs lacking skeletal muscle. J. Cell Biol. 2001, 155, 381–392. [Google Scholar] [CrossRef]
- Gerhart, J.; Elder, J.; Neely, C.; Schure, J.; Kvist, T.; Knudsen, K.; George-Weinstein, M. MyoD-positive epiblast cells regulate skeletal muscle differentiation in the embryo. J. Cell Biol. 2006, 175, 283–292. [Google Scholar] [CrossRef]
- Strony, R.; Gerhart, J.; Tornambe, D.; Perlman, J.; Neely, C.; Dare, J.; Stewart, B.; George-Weinstein, M. NeuroM and MyoD are expressed in separate subpopulations of cells in the pregastrulating epiblast. Gene Expr. Patterns 2005, 5, 387–395. [Google Scholar] [CrossRef]
- Krause, C.; Guzman, A.; Knaus, P. Noggin. Int. J. Biochem. Cell Biol. 2011, 43, 478–481. [Google Scholar] [CrossRef]
- Secondini, C.; Wetterwald, A.; Schwaninger, R.; Thalmann, G.N.; Cecchini, M.G. The role of the BMP signaling antagonist noggin in the development of prostate cancer osteolytic bone metastasis. PLoS ONE 2011, 6, e16078. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Gao, M.; He, Z.; Guo, P.; Liu, Z.; Zhang, P.; Zhang, F.; Chen, D.; Zhou, G.; Zhou, Z. Noggin, an inhibitor of bone morphogenetic protein signaling, antagonizes TGF-beta1 in a mouse model of osteoarthritis. Biochem. Biophys. Res. Commun. 2021, 570, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, J.; Bowers, J.; Gugerty, L.; Gerhart, C.; Martin, M.; Abdalla, F.; Bravo-Nuevo, A.; Sullivan, J.T.; Rimkunas, R.; Albertus, A.; et al. Brain-specific angiogenesis inhibitor 1 is expressed in the Myo/Nog cell lineage. PLoS ONE 2020, 15, e0234792. [Google Scholar] [CrossRef]
- Gerhart, J.; Pfautz, J.; Neely, C.; Elder, J.; DuPrey, K.; Menko, A.S.; Knudsen, K.; George-Weinstein, M. Noggin producing, MyoD-positive cells are crucial for eye development. Dev. Biol. 2009, 336, 30–41. [Google Scholar] [CrossRef]
- Gerhart, J.; Greenbaum, M.; Scheinfeld, V.; Fitzgerald, P.; Crawford, M.; Bravo-Nuevo, A.; Pitts, M.; George-Weinstein, M. Myo/Nog cells: Targets for preventing the accumulation of skeletal muscle-like cells in the human lens. PLoS ONE 2014, 9, e95262. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Nuevo, A.; Brandli, A.A.; Gerhart, J.; Nichols, J.; Pitts, M.; Sutera, C.K.; Assali, S.; Scheinfeld, V.; Prendergast, G.C.; Stone, J.; et al. Neuroprotective effect of Myo/Nog cells in the stressed retina. Exp. Eye Res. 2016, 146, 22–25. [Google Scholar] [CrossRef]
- Brandli, A.; Gerhart, J.; Sutera, C.K.; Purushothuman, S.; George-Weinstein, M.; Stone, J.; Bravo-Nuevo, A. Role of Myo/Nog Cells in Neuroprotection: Evidence from the Light Damaged Retina. PLoS ONE 2017, 12, e0169744. [Google Scholar] [CrossRef]
- Gerhart, J.; Withers, C.; Gerhart, C.; Werner, L.; Mamalis, N.; Bravo-Nuevo, A.; Scheinfeld, V.; FitzGerald, P.; Getts, R.; George-Weinstein, M. Myo/Nog cells are present in the ciliary processes, on the zonule of Zinn and posterior capsule of the lens following cataract surgery. Exp. Eye Res. 2018, 171, 101–105. [Google Scholar] [CrossRef]
- Gerhart, J.; Behling, K.; Paessler, M.; Milton, L.; Bramblett, G.; Garcia, D.; Pitts, M.; Hurtt, R.; Crawford, M.; Lackman, R.; et al. Rhabdomyosarcoma and Wilms tumors contain a subpopulation of noggin producing, myogenic cells immunoreactive for lens beaded filament proteins. PLoS ONE 2019, 14, e0214758. [Google Scholar] [CrossRef]
- Gerhart, J.; Greenbaum, M.; Casta, L.; Clemente, A.; Mathers, K.; Getts, R.; George-Weinstein, M. Antibody-Conjugated, DNA-Based Nanocarriers Intercalated with Doxorubicin Eliminate Myofibroblasts in Explants of Human Lens Tissue. J. Pharmacol. Exp. Ther. 2017, 361, 60–67. [Google Scholar] [CrossRef]
- Gerhart, J.; Werner, L.; Mamalis, N.; Infanti, J.; Withers, C.; Abdalla, F.; Gerhart, C.; Bravo-Nuevo, A.; Gerhart, O.; Getts, L.; et al. Depletion of Myo/Nog Cells in the Lens Mitigates Posterior Capsule Opacification in Rabbits. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, J.; Morrison, N.; Gugerty, L.; Telander, D.; Bravo-Nuevo, A.; George-Weinstein, M. Myo/Nog cells expressing muscle proteins are present in preretinal membranes from patients with proliferative vitreoretinopathy. Exp. Eye Res. 2020, 197, 108080. [Google Scholar] [CrossRef]
- Crispin, M.; Gerhart, J.; Heffer, A.; Martin, M.; Abdalla, F.; Bravo-Nuevo, A.; Philp, N.J.; Kuriyan, A.E.; George-Weinstein, M. Myo/Nog Cells Give Rise to Myofibroblasts During Epiretinal Membrane Formation in a Mouse Model of Proliferative Vitreoretinopathy. Investig. Ophthalmol. Vis. Sci. 2023, 64, 1. [Google Scholar] [CrossRef] [PubMed]
- Joseph-Pauline, S.; Morrison, N.; Braccia, M.; Payne, A.; Gugerty, L.; Mostoller, J.; Lecker, P.; Tsai, E.J.; Kim, J.; Martin, M.; et al. Acute Response and Neuroprotective Role of Myo/Nog Cells Assessed in a Rat Model of Focal Brain Injury. Front. Neurosci. 2021, 15, 780707. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.J.; Zhao, E.J.; Chiao, M.; Lim, C.J. Co-culture of induced pluripotent stem cells with cardiomyocytes is sufficient to promote their differentiation into cardiomyocytes. PLoS ONE 2020, 15, e0230966. [Google Scholar] [CrossRef]
- Ferrer, E. Trabecular meshwork as a new target for the treatment of glaucoma. Drug News Perspect. 2006, 19, 151–158. [Google Scholar] [CrossRef]
- Gong, H.; Freddo, T.F. The washout phenomenon in aqueous outflow--why does it matter? Exp. Eye Res. 2009, 88, 729–737. [Google Scholar] [CrossRef]
- Braunger, B.M.; Fuchshofer, R.; Tamm, E.R. The aqueous humor outflow pathways in glaucoma: A unifying concept of disease mechanisms and causative treatment. Eur. J. Pharm. Biopharm. 2015, 95, 173–181. [Google Scholar] [CrossRef]
- Gerhart, J.; Scheinfeld, V.L.; Milito, T.; Pfautz, J.; Neely, C.; Fisher-Vance, D.; Sutter, K.; Crawford, M.; Knudsen, K.; George-Weinstein, M. Myo/Nog cell regulation of bone morphogenetic protein signaling in the blastocyst is essential for normal morphogenesis and striated muscle lineage specification. Dev. Biol. 2011, 359, 12–25. [Google Scholar] [CrossRef]
- Gerhart, J.; Hayes, C.; Scheinfeld, V.; Chernick, M.; Gilmour, S.; George-Weinstein, M. Myo/Nog cells in normal, wounded and tumor-bearing skin. Exp. Dermatol. 2012, 21, 466–468. [Google Scholar] [CrossRef]
- Gerhart, J.; Gugerty, L.; Lecker, P.; Abdalla, F.; Martin, M.; Gerhart, O.; Gerhart, C.; Johal, K.; Bernstein, J.; Spikes, J.; et al. Myo/Nog cells are nonprofessional phagocytes. PLoS ONE 2020, 15, e0235898. [Google Scholar] [CrossRef] [PubMed]
- Samanta, J.; Alden, T.; Gobeske, K.; Kan, L.; Kessler, J.A. Noggin protects against ischemic brain injury in rodents. Stroke 2010, 41, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Du, Y.; Wu, W.; Yip, H.K. Bone morphogenetic proteins mediate cellular response and, together with Noggin, regulate astrocyte differentiation after spinal cord injury. Exp. Neurol. 2010, 221, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Dizon, M.L.; Maa, T.; Kessler, J.A. The bone morphogenetic protein antagonist noggin protects white matter after perinatal hypoxia-ischemia. Neurobiol. Dis. 2011, 42, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.H.; Brenner, G.J.; Omura, T.; Samad, O.A.; Costigan, M.; Inquimbert, P.; Niederkofler, V.; Salie, R.; Sun, C.C.; Lin, H.Y.; et al. The BMP coreceptor RGMb promotes while the endogenous BMP antagonist noggin reduces neurite outgrowth and peripheral nerve regeneration by modulating BMP signaling. J. Neurosci. 2011, 31, 18391–18400. [Google Scholar] [CrossRef] [PubMed]
- Gulino, R.; Gulisano, M. Noggin and Sonic hedgehog are involved in compensatory changes within the motoneuron-depleted mouse spinal cord. J. Neurol. Sci. 2013, 332, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Koda, M.; Furuya, T.; Murata, A.; Yamazaki, M.; Takahashi, K. Intrathecal Noggin administration in rats temporally ameliorates mechanical allodynia induced by a chronic constriction injury. eNeurologicalSci 2016, 4, 4–9. [Google Scholar] [CrossRef]
- Al-Sammarraie, N.; Mahmood, M.; Ray, S.K. Neuroprotective role of Noggin in spinal cord injury. Neural Regen. Res. 2023, 18, 492–496. [Google Scholar] [CrossRef]
- Su, X.; Ying, J.; Xiao, D.; Qiu, X.; Li, S.; Zhao, F.; Tang, J. Activin A rescues preterm brain injury through a novel Noggin/BMP4/Id2 signaling pathway. Int. J. Mol. Med. 2023, 51, 12. [Google Scholar] [CrossRef]
- Park, D.; Tosello-Trampont, A.C.; Elliott, M.R.; Lu, M.; Haney, L.B.; Ma, Z.; Klibanov, A.L.; Mandell, J.W.; Ravichandran, K.S. BAI1 is an engulfment receptor for apoptotic cells upstream of the ELMO/Dock180/Rac module. Nature 2007, 450, 430–434. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lecker, P.; Johal, K.; McGrath, A.; Spikes, J., II; Bernstein, J.; MacPherson, V.; Brahmbhatt, R.; Fadl, N.; Weyback-Liogier, E.; Adams, S.; et al. Myo/Nog Cells Increase in Response to Elevated Intraocular Pressure and Mitigate Ganglion Cell Death in a Mouse Model of Glaucoma. Appl. Sci. 2023, 13, 12423. https://doi.org/10.3390/app132212423
Lecker P, Johal K, McGrath A, Spikes J II, Bernstein J, MacPherson V, Brahmbhatt R, Fadl N, Weyback-Liogier E, Adams S, et al. Myo/Nog Cells Increase in Response to Elevated Intraocular Pressure and Mitigate Ganglion Cell Death in a Mouse Model of Glaucoma. Applied Sciences. 2023; 13(22):12423. https://doi.org/10.3390/app132212423
Chicago/Turabian StyleLecker, Paul, Karanveer Johal, Alexa McGrath, John Spikes, II, Jake Bernstein, Victoria MacPherson, Rushil Brahmbhatt, Nada Fadl, Edgar Weyback-Liogier, Sarah Adams, and et al. 2023. "Myo/Nog Cells Increase in Response to Elevated Intraocular Pressure and Mitigate Ganglion Cell Death in a Mouse Model of Glaucoma" Applied Sciences 13, no. 22: 12423. https://doi.org/10.3390/app132212423