
Characterization of Escherichia coli Cefotaxime-Resistance in Al-Ahsa, KSA: Predominance of CTX-15 and First Report of blaCMY-42 Gene
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
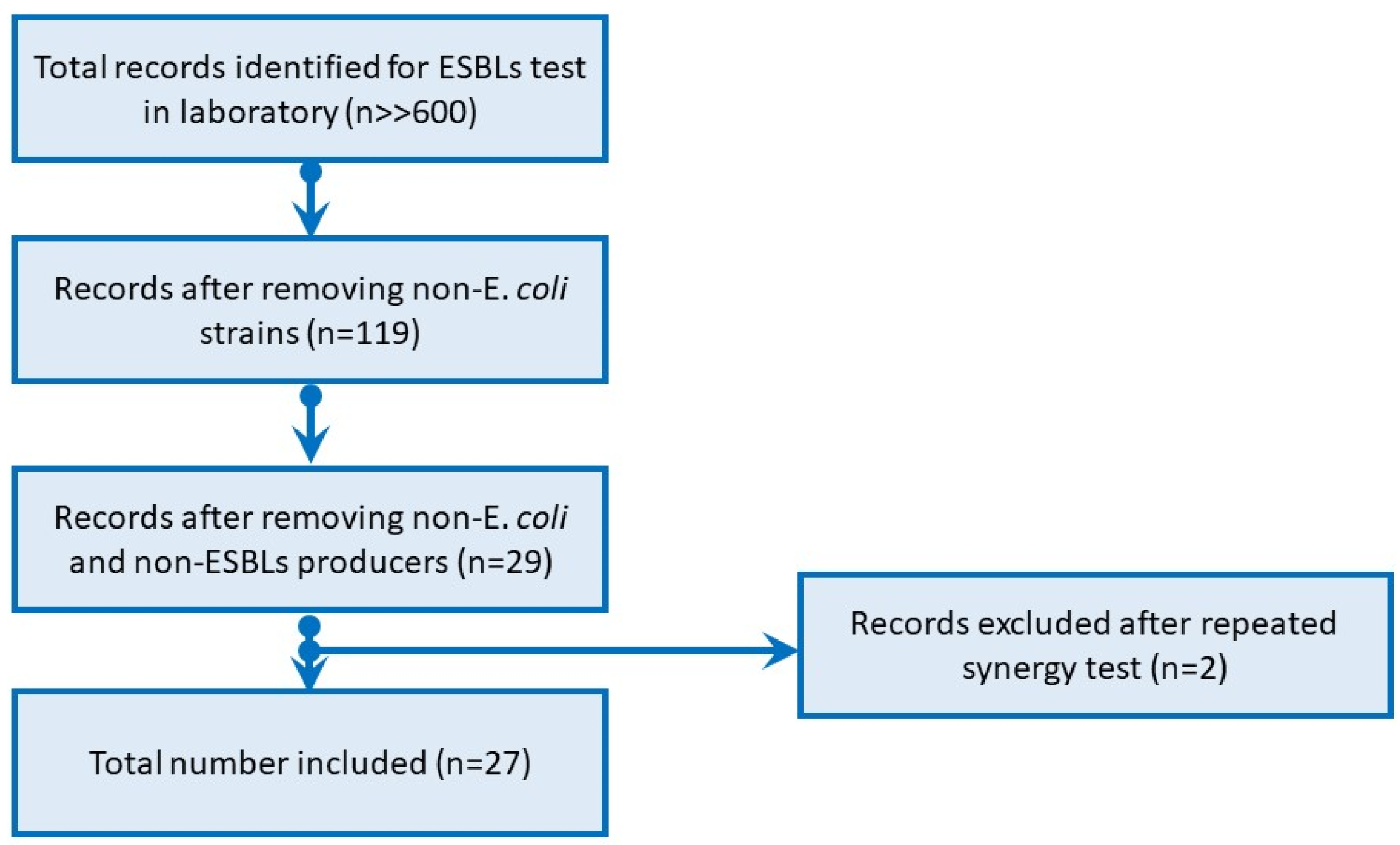
2.1. Study Design
2.2. Antimicrobial Susceptibility Tests and Cefotaxime Resistance Transfer Assays
2.3. Characterization of Antibiotic Resistance Genes
2.4. Quinolone Resistance Genes, Integrons, and Plasmid Incompatibility Groups
2.5. Phylogenetic Analysis, Genetic Relatedness, and ST131 Identification
2.6. Ethical Approval
3. Results
3.1. Antibiotic Susceptibilities and Cefotaxime Resistance Transfer Assays
3.2. Characterization of Antibiotic Resistance
3.3. Quinolone Resistance Genes, Integrons, and Plasmid Incompatibility
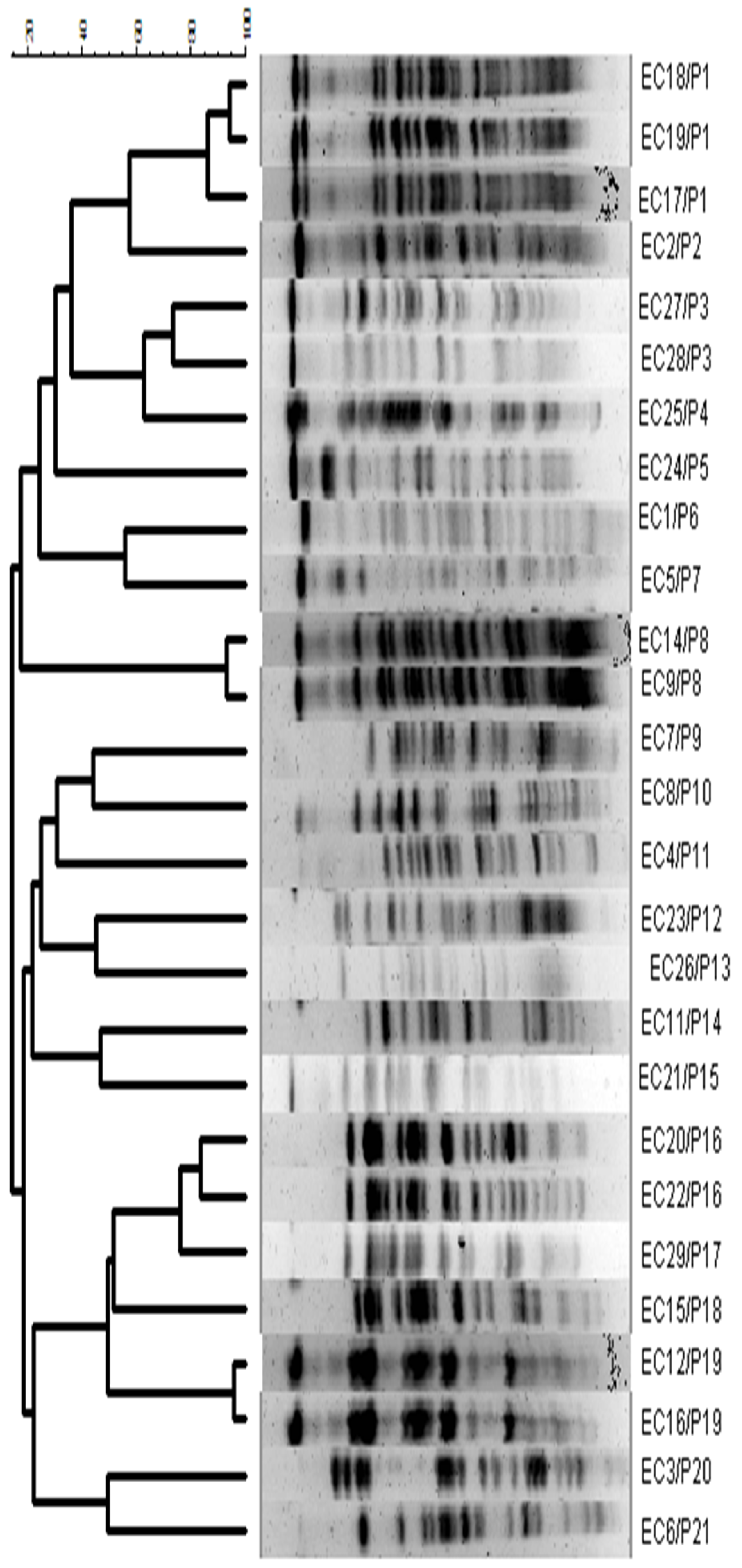
3.4. Genetic Relationship, Phylogenetic Groups, and Identification of the ST131 Clone
3.5. Occurrence of Virulence Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anas, A.; Sobhanan, J.; Sulfiya, K.; Jasmin, C.; Sreelakshmi, P.; Biju, V. Advances in photodynamic antimicrobial chemotherapy. J. Photochem. Photobiol. C Photochem. Rev. 2021, 49, 100452. [Google Scholar] [CrossRef]
- Bokhary, H.; Pangesti, K.N.A.; Rashid, H.; Abd El Ghany, M.; Hill-Cawthorne, G.A. Travel-Related Antimicrobial Resistance: A Systematic Review. Trop. Med. Infect. Dis. 2021, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Guerra, B.; Gonzalez-Lara, M.; De-Leon-Cividanes, N.; Tamez-Torres, K.; Roman-Montes, C.; Rajme-Lopez, S.; Villalobos-Zapata, G.; Lopez-Garcia, N.; Martínez-Gamboa, A.; Sifuentes-Osornio, J.; et al. Antimicrobial Resistance Patterns and Antibiotic Use during Hospital Conversion in the COVID-19 Pandemic. Antibiotics 2021, 10, 182. [Google Scholar] [CrossRef]
- Saini, V.; Jain, C.; Singh, N.; Alsulimani, A.; Gupta, C.; Dar, S.; Haque, S.; Das, S. Paradigm Shift in Antimicrobial Resistance Pattern of Bacterial Isolates during the COVID-19 Pandemic. Antibiotics 2021, 10, 954. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Stell, A.L. Extended Virulence Genotypes of Escherichia coli Strains from Patients with Urosepsis in Relation to Phylogeny and Host Compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals|EFSA. Available online: https://www.efsa.europa.eu/it/efsajournal/pub/2322 (accessed on 1 October 2022).
- Al-Agamy, M.H.; Shibl, A.M.; Hafez, M.M.; Al-Ahdal, M.N.; Memish, Z.A.; Khubnani, H. Molecular characteristics of extended-spectrum β-lactamase-producing Escherichia coli in Riyadh: Emergence of CTX-M-15-producing E. coli ST131. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alharbi, N.S.; Khaled, J.M.; Kadaikunnan, S.; Alobaidi, A.S.; Sharafaddin, A.H.; Alyahya, S.A.; Almanaa, T.N.; Alsughayier, M.A.; Shehu, M.R. Prevalence of Escherichia coli strains resistance to antibiotics in wound infections and raw milk. Saudi J. Biol. Sci. 2018, 26, 1557–1562. [Google Scholar] [CrossRef] [PubMed]
- Majumder, A.A.; Rahman, S.; Cohall, D.; Bharatha, A.; Singh, K.; Haque, M.; Hilaire, M.G.-S. Antimicrobial Stewardship: Fighting Antimicrobial Resistance and Protecting Global Public Health. Infect. Drug Resist. 2020, 13, 4713–4738. [Google Scholar] [CrossRef]
- Ben Aissa, M.; Fernea, A.H.; Bouraoui, H.; Ben Aissa, R.; Mercuri, P.S.; Manai, M. Molecular characterization of plasmid-encoded ACC-1a4 β-lactamase produced by Salmonella strains isolated from water in a Tunisian hospital. Ann. Microbiol. 2011, 62, 31–36. [Google Scholar] [CrossRef]
- Samarasinghe, S.; Reid, R.; Varnakulasingam, A. The Distribution of ESBL-Producing Enterobacteriaceae: Leicestershire UK Compared to Worldwide. Am. J. Biomed. Sci. Res. 2019, 3, 70–72. [Google Scholar] [CrossRef]
- Nosheen, S.; Bukhari, N.I.; Junaid, K.; Anwar, N.; Ahmad, F.; Younas, S.; Ejaz, H. Phylogenetic diversity and mutational analysis of New Delhi Metallo-β-lactamase (NDM) producing E. coli strains from pediatric patients in Pakistan. Saudi J. Biol. Sci. 2021, 28, 5875–5883. [Google Scholar] [CrossRef] [PubMed]
- Chedid, M.; Waked, R.; Haddad, E.; Chetata, N.; Saliba, G.; Choucair, J. Antibiotics in treatment of COVID-19 complications: A review of frequency, indications, and efficacy. J. Infect. Public Health 2021, 14, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Ojer-Usoz, E.; González, D.; Vitas, A.I. Clonal Diversity of ESBL-Producing Escherichia coli Isolated from Environmental, Human and Food Samples. Int. J. Environ. Res. Public Health 2017, 14, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasir, M.; Farman, M.; Shah, M.W.; Jiman-Fatani, A.A.; Othman, N.A.; Almasaudi, S.B.; Alawi, M.; Shakil, S.; Al-Abdullah, N.; Ismaeel, N.A.; et al. Genomic and antimicrobial resistance genes diversity in multidrug-resistant CTX-M-positive isolates of Escherichia coli at a health care facility in Jeddah. J. Infect. Public Health 2019, 13, 94–100. [Google Scholar] [CrossRef]
- Zgurskaya, H.I. Molecular analysis of efflux pump-based antibiotic resistance. Int. J. Med Microbiol. 2002, 292, 95–105. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Ciesielczuk, H.; Hornsey, M.; Choi, V.; Woodford, N.; Wareham, D. Development and evaluation of a multiplex PCR for eight plasmid-mediated quinolone-resistance determinants. J. Med Microbiol. 2013, 62, 1823–1827. [Google Scholar] [CrossRef]
- Sáenz, Y.; Zarazaga, M.; Briñas, L.; Lantero, M.; Larrea, F.R.; Torres, C. Antibiotic resistance in Escherichia coli isolates obtained from animals, foods and humans in Spain. Int. J. Antimicrob. Agents 2001, 18, 353–358. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Cantón, R.; Horcajada, J.P.; Oliver, A.; Garbajosa, P.R.; Vila, J. Inappropriate use of antibiotics in hospitals: The complex relationship between antibiotic use and antimicrobial resistance. Enferm. Infecc. Microbiol. Clin. 2013, 31, 3–11. [Google Scholar] [CrossRef]
- Takahashi, T.; Masuda, T.; Koyama, Y. Enhanced gene targeting frequency in ku70 and ku80 disruption mutants of Aspergillus sojae and Aspergillus oryzae. Mol. Genet. Genom. 2006, 275, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Dhanji, H.; Upton, M.; Gibreel, T.; Fox, A.; Boyd, D.; Mulvey, M.R.; Nordmann, P.; Ruppe, E.; Sarthou, J.L.; et al. Rapid detection of the O25b-ST131 clone of Escherichia coli encompassing the CTX-M-15-producing strains. J. Antimicrob. Chemother. 2009, 64, 274–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, H.; Török, M.E. Corrigendum: Extended-spectrum β-lactamase-producing and carbapenemase-producing Enterobacteriaceae. Microb. Genom. 2018, 4, e000218. [Google Scholar] [CrossRef] [PubMed]
- Alqasim, A.; Abu Jaffal, A.; Alyousef, A.A. Prevalence of Multidrug Resistance and Extended-Spectrum β-Lactamase Carriage of Clinical Uropathogenic Escherichia coli Isolates in Riyadh, Saudi Arabia. Int. J. Microbiol. 2018, 2018, 302685. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, M.A.; Sadoma, H.H.M.; Mathew, S.; Alghamdi, S.; Malik, J.A.; Anwar, S. Retrospective Analysis of Antimicrobial Susceptibility of Uropathogens Isolated from Pediatric Patients in Tertiary Hospital at Al-Baha Region, Saudi Arabia. Healthcare 2021, 9, 1564. [Google Scholar] [CrossRef]
- Senok, A.; Khanfar, H.S.; Bindayna, K.M.; Botta, G.A. Extended spectrum beta-lactamases (ESBL) in Escherichia coli and Klebsiella pneumoniae: Trends in the hospital and community settings. J. Infect. Dev. Ctries. 2009, 3, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Bi, W.; Li, B.; Song, J.; Hong, Y.; Zhang, X.; Liu, H.; Lu, H.; Zhou, T.; Cao, J. Antimicrobial susceptibility and mechanisms of fosfomycin resistance in extended-spectrum β-lactamase-producing Escherichia coli strains from urinary tract infections in Wenzhou, China. Int. J. Antimicrob. Agents 2017, 50, 29–34. [Google Scholar] [CrossRef]
- Sghaier, S.; Abbassi, M.S.; Pascual, A.; Serrano, L.; De-Alba, P.D.; Ben Said, M.; Hassen, B.; Ibrahim, C.; Hassen, A.; López-Cerero, L. Extended-spectrum β-lactamase-producing Enterobacteriaceae from animal origin and wastewater in Tunisia: First detection of O25b-B23-CTX-M-27-ST131 Escherichia coli and CTX-M-15/OXA-204-producing Citrobacter freundii from wastewater. J. Glob. Antimicrob. Resist. 2019, 17, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Al-Agamy, M.H.; Aljallal, A.; Radwan, H.H.; Shibl, A.M. Characterization of carbapenemases, ESBLs, and plasmid-mediated quinolone determinants in carbapenem-insensitive Escherichia coli and Klebsiella pneumoniae in Riyadh hospitals. J. Infect. Public Health 2017, 11, 64–68. [Google Scholar] [CrossRef]
- Yasir, M.; Ajlan, A.M.; Shakil, S.; Jiman-Fatani, A.A.; Almasaudi, S.B.; Farman, M.; Baazeem, Z.M.; Baabdullah, R.; Alawi, M.; Al-Abdullah, N.; et al. Molecular characterization, antimicrobial resistance and clinico-bioinformatics approaches to address the problem of extended-spectrum β-lactamase-producing Escherichia coli in western Saudi Arabia. Sci. Rep. 2018, 8, 14847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentschke, M.; Kotsakis, S.D.; Wolters, M.; Heisig, P.; Miriagou, V.; Aepfelbacher, M. CMY-42, a Novel Plasmid-Mediated CMY-2 Variant AmpC Beta-Lactamase. Microb. Drug Resist. 2011, 17, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, P.; Xie, Y.; Wang, X.; McNally, A.; Zong, Z. Escherichia coli of sequence type 3835 carrying blaNDM-1, blaCTX-M-15, blaCMY-42 and blaSHV-12. Sci. Rep. 2015, 5, 12275. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance Plasmid Families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madec, J.-Y.; Haenni, M. Antimicrobial resistance plasmid reservoir in food and food-producing animals. Plasmid 2018, 99, 72–81. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D.D. The Role of Epidemic Resistance Plasmids and International High-Risk Clones in the Spread of Multidrug-Resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | Numbers | Percentage (%) | |
|---|---|---|---|
| Gender | Male | 13 | 48.1 |
| Female | 14 | 51.9 | |
| Age | ≤37 | 5 | 18.5 |
| 38–55 | 12 | 44.4 | |
| 56–73 | 6 | 22.2 | |
| 74+ | 4 | 14.8 | |
| Specimen | Sputum | 2 | 7.4 |
| Surgical tissue | 2 | 7.4 | |
| Urine | 14 | 51.9 | |
| Wound | 9 | 33.3 |
| Strain | β-Lactam Resistance Phenotypes Detected * | Non-β-Lactam Resistance Phenotypes Detected * |
|---|---|---|
| EC 1 | CAZ, CTX, FEP | NAL, CIP, TET, FOS, SXT |
| EC 2 | AMC, CAZ, CTX, FEP | TOB, NAL, CIP, FOS, CHL, SXT |
| EC 3 | AMC, CTX, FEP | NAL, TET, FOS, CHL, SXT |
| EC 4 | AMC, CTX | NAL, CIP, TET, FOS, CHL, SXT |
| EC 5 | AMC, CTX, FEP | NAL, TET, MNO, FOS, CHL, SXT |
| EC 6 | AMC, CAZ, CTX, FEP | NAL, CIP, TET, MNO, FOS, CHL, SXT |
| EC 7 | AMC, CTX | NAL, CIP, FOS |
| EC 8 | AMC, CAZ, CTX, FOX | TOB, NAL, CIP, TET, MNO, FOS, CHL, SXT |
| EC 9 | AMC, CAZ, CTX, FEP | NAL, CIP, FOS, CHL, SXT |
| EC 11 | AMC, CAZ, CTX, FEP | GMN, TOB, NAL, CIP, TET, FOS, SXT |
| EC 12 | AMC, CAZ, CTX, FEP | NAL, FOS, SXT |
| EC 14 | AMC, CAZ, CTX, FEP | GMN, TOB, NET, NAL, CIP, TET, FOS, CHL, SXT |
| EC 15 | AMC, CTX, FEP | NAL, CIP, FOS, CHL, SXT |
| EC 16 | AMC, CAZ, CTX, PEP | GMN, TOB, NAL, CIP, TET, FOS, CHL, SXT |
| EC 17 | AMC, CTX, FEP | NAL, CIP, TET, FOS, CHL, SXT, |
| EC 18 | CAZ, CTX, FEP | NET, NAL, CIP, FOS, CHL, |
| EC 19 | AMC, CTX, FEP | NAL, CIP, FOS |
| EC 20 | CTX, FEP | NAL, CIP, FOS, SXT |
| EC 21 | AMC, CAZ, CTX, FEP | NAL, CIP, TET, MNO, FOS, SXT |
| EC 22 | AMC, CTX | NAL, CIP, TET, FOS, CHL, SXT |
| EC 23 | AMC, CTX | NAL, CIP, FOS, CHL, SXT |
| EC 24 | AMC, CTX | TOB, NAL, CIP, FOS, CHL, SXT |
| EC 25 | AMC, CTX, FEP | TOB, NAL, CIP, FOS, SXT |
| EC 26 | AMC, CTX, FEP | NAL, CIP, TET, FOS, CHL |
| EC 27 | AMC, CAZ, CTX, FEP | GMN, TOB, NAL, CIP, TET, FOS, CHL, SXT |
| EC 28 | AMC, CAZ, CTX, FEP | GMN, TOB, NAL, CIP, TET, FOS, CHL, SXT |
| EC 29 | AMC, CTX, FOX | NAL, CIP, FOS, CHL, SXT |
| Strains | bla and PMQR* Genes Identified | Virulence Profile | Virulence Score |
|---|---|---|---|
| EC 1 | blaCTX-M-15, qnrS1 | fimH-papGII-iha-iutA-traT-malX-usp-ompT | 8 |
| EC 2 | blaCMY-42, blaCTX-M-15, blaSHV-1 | fimH-papGII-iutA-malX | 4 |
| EC 3 | blaCTX-M-15, blaCTX-M-27, blaTEM | papGII-ompT | 2 |
| EC 4 | blaCTX-M-27 | fimH-papGII-iha-malX-usp-ompT | 6 |
| EC 5 | blaCTX-M-15, blaTEM-1 | malX-ompT | 2 |
| EC 6 | blaCTX-M-15, blaTEM-1 | papGII-fyuA-malX-ompT | 4 |
| EC 7 | blaCTX-M-15, qnrS1 | fimH-papGII-iha-iutA-kpsMTII-malX-usp-ompT | 8 |
| EC 8 | blaCMY-42, blaTEM-1, blaSHV-1, qnrS1, aac(6′)Ib-cr | fimH-papGII | 2 |
| EC 9 | blaCTX-M-15, blaTEM-1 | papGII-malX | 2 |
| EC 11 | blaCTX-M-15, blaSHV, qnrS1 | papGII-iha-traT-malX-usp-ompT | 6 |
| EC 12 | blaCTX-M-15, qnrS1, aac(6′)Ib-cr | papGI-iha-traT-malX-usp | 5 |
| EC 14 | blaCTX-M-15, blaTEM-1 | papGIII-malX-usp-ompT | 4 |
| EC 15 | blaCTX-M-15 | papGII-iha-usp-ompT | 4 |
| EC 16 | blaCTX-M-15, blaTEM-1, blaSHV-1 | papGII-iha-iutA-traT-usp-ompT | 6 |
| EC 17 | blaCTX-M-15, blaTEM-1 | fimH-papGII-iha--iutA-ompT | 5 |
| EC 18 | blaCTX-M-15, blaTEM-1 | fimH-iha-iutA-kpsMTII-malX-usp-ompT | 7 |
| EC 19 | blaCTX-M-15, blaTEM-1, qnrS1 | fimH-iha-iutA-kpsMTII-malX-usp-ompT | 7 |
| EC 20 | blaCTX-M-27, blaTEM-1, qnrS1 | fimH-papGII-iha-traT-usp-ompT | 6 |
| EC 21 | blaCTX-M-15, blaTEM-1, qnrS1 | fimH-iha-kpsMTII-usp-ompT | 5 |
| EC 22 | blaCTX-M-27, blaTEM-1 | fimH-papGII-iha-usp-ompT | 5 |
| EC 23 | blaCTX-M-27, blaTEM-1 | papGII-ompT | 2 |
| EC 24 | blaCTX-M-15, blaTEM-1, qnrS1, aac(6′)Ib-cr | - | - |
| EC 25 | blaCTX-M-15, blaSHV-1 | fimH-papGII-iha-iutA-malX-usp-ompT | 7 |
| EC 26 | blaCTX-M-15, blaTEM-1 | fimH-papGII-iutA-ompT | 4 |
| EC 27 | blaCTX-M-15, blaTEM-1, qnrS1, aac(6′)Ib-cr | papGI-papGII-iha-kpsMTII-hlyA traT-usp-ompT | 8 |
| EC 28 | blaCTX-M-15, blaTEM-1-1, qnrS1, aac(6′)Ib-cr | fimH-papGI-iha-iutA-kpsMTII-traT-malX-usp-ompT | 9 |
| EC 29 | blaCTX-M-9, blaCTX-M-27, blaTEM-1, qnrS1, aac(6′)Ib-cr | fimH-papGII-malX-usp | 4 |
| Strains | int Genes | PRT | PG | PFGE |
|---|---|---|---|---|
| EC 1 | - | F | B2 | P6 |
| EC 2 | int1 | F | D | P2 |
| EC 3 | int2 | FIA | D | P20 |
| EC 4 | - | - | B2 | P11 |
| EC 5 | - | - | A | P7 |
| EC 6 | - | - | B2 | P21 |
| EC 7 | - | - | D | P9 |
| EC 8 | int2 | FIA-F | B1 | P10 |
| EC 9 | int1 | - | B2 | P8 |
| EC 11 | - | F | B2 | P14 |
| EC 12 | int1 | FIA-F | B2 | P19 |
| EC 14 | - | - | B2 | P8 |
| EC 15 | int1 | FIA | B2 | P18 |
| EC 16 | int1 | - | B2 | P19 |
| EC 17 | - | FIA | B2 * | P1 |
| EC 18 | - | FIA | B2 * | P1 |
| EC 19 | - | FIA | B2 * | P1 |
| EC 20 | - | - | B2 | P16 |
| EC 21 | - | - | B2 | P15 |
| EC 22 | - | - | B2 | P16 |
| EC 23 | - | - | D | P12 |
| EC 24 | - | FIA-F | D | P5 |
| EC 25 | - | - | B2 | P4 |
| EC 26 | - | - | A | P13 |
| EC 27 | int1 | FIA-F | B2 | P3 |
| EC 28 | int1 | FIA-F | B2 | P3 |
| EC 29 | - | - | B2 | P1 |
| Strains | Non-β-Lactams Resistance Phenotypes Detected | bla and PMQR Genes | int Genes | PRT |
|---|---|---|---|---|
| EC 1 | NAL, CIP, TET, FOS, SXT | blaCTX-M-15, blaTEM, qnrS1 | - | F |
| * Tc EC 1 | TET | blaCTX-M-15 | - | F |
| EC 7 | NAL, CIP, FOS | blaCTX-M-15, qnrS1 | - | - |
| Tc EC 7 | - | blaCTX-M-15 | - | - |
| EC 8 | TOB, NAL, CIP, TET, MNO, FOS, CHL, SXT | blaCMY-42, blaTEM, blaSHV, qnrS1, aac(6′)Ib-cr | int2 | FIA-F |
| Tc EC 8 | TET, MNO | blaCMY-42, blaTEM, qnrS1 | - | FIA |
| EC 11 | GMN, TOB, NAL, CIP, TET, FOS, SXT | blaCTX-M-15, blaSHV, qnrS1 | - | F |
| Tc EC 11 | TET | blaCTX-M-15 | - | F |
| EC 12 | NAL, FOS, SXT | blaCTX-M-15, qnrS1, aac(6′)Ib-cr | int1 | FIA-F |
| Tc EC 12 | - | blaCTX-M-15 | int1 | FIA |
| EC 19 | NAL, CIP, FOS | blaCTX-M-15, blaTEM, qnrS1 | - | FIA |
| Tc EC 19 | - | blaCTX-M-15 | - | FIA |
| EC 20 | NAL, CIP, FOS, SXT | blaCTX-M-27, blaTEM, qnrS1 | - | - |
| Tc EC 20 | SXT | blaCTX-M-27, blaTEM, qnrS1 | - | - |
| EC 21 | NAL, CIP, TET, MNO, FOS, SXT | blaCTX-M-15, blaTEM, qnrS1 | - | - |
| Tc EC 21 | TET, MNO, SXT | blaCTX-M-15, blaTEM, qnrS1 | - | - |
| EC 24 | TOB, NAL, CIP, FOS, CHL, SXT | blaCTX-M-15, blaTEM, qnrS1, aac(6′)Ib-cr | - | FIA-F |
| Tc EC 24 | TOB, CHL, SXT | blaCTX-M-15, blaTEM, qnrS1, aac(6′)Ib-cr | - | FIA-F |
| EC 27 | GMN, TOB, NAL, CIP, TET, FOS, CHL, SXT | blaCTX-M-15, blaTEM, qnrS1, aac(6′)Ib-cr | int1 | FIA-F |
| Tc EC 27 | TET | blaCTX-M-15 | int1 | F |
| EC 28 | GMN, TOB, NAL, CIP, TET, FOS, CHL, SXT | blaCTX-M-15, blaTEM, qnrS1, aac(6′)Ib-cr | int1 | FIA-F |
| Tc EC 28 | TET, SXT | blaCTX-M-15, blaTEM, qnrS1 | int1 | FIA-F |
| EC 29 | NAL, CIP, FOS, CHL, SXT | blaCTX-M-9, blaCTX-M-27, blaTEM, qnrS1, aac(6′)Ib-cr | - | - |
| Tc EC 29 | CHL, SXT | blaCTX-M-9, blaTEM, qnrS1 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aissa, M.B.; Ferjani, S.; Abassi, M.S.; Al-Suwailem, N.; Boutiba, I. Characterization of Escherichia coli Cefotaxime-Resistance in Al-Ahsa, KSA: Predominance of CTX-15 and First Report of blaCMY-42 Gene. Appl. Sci. 2022, 12, 9964. https://doi.org/10.3390/app12199964
Aissa MB, Ferjani S, Abassi MS, Al-Suwailem N, Boutiba I. Characterization of Escherichia coli Cefotaxime-Resistance in Al-Ahsa, KSA: Predominance of CTX-15 and First Report of blaCMY-42 Gene. Applied Sciences. 2022; 12(19):9964. https://doi.org/10.3390/app12199964
Chicago/Turabian StyleAissa, Melek Ben, Sana Ferjani, Mohamed Salah Abassi, Nada Al-Suwailem, and Ilhem Boutiba. 2022. "Characterization of Escherichia coli Cefotaxime-Resistance in Al-Ahsa, KSA: Predominance of CTX-15 and First Report of blaCMY-42 Gene" Applied Sciences 12, no. 19: 9964. https://doi.org/10.3390/app12199964