
Transcriptome Analysis Reveals Differentially Expressed circRNAs Associated with Fecundity in Small-Tail Han Sheep Thyroid with Different FecB Genotypes

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. RNA Extraction, Library Construction, and Sequencing
2.3. Sequencing Data Filtering, Comparative Analysis, and Splicing
2.4. circRNA Identification
2.5. Differential Expression Analysis
2.6. Enrichment Analysis of the Source Genes of circRNAs
2.7. Validation of Sequencing Data
- (1)
- Extraction and identification of total RNA
- (2)
- Reverse transcription
- (3)
- Primer design and synthesis
2.8. Validation of circRNA–miRNA–mRNA ceRNA Mechanism
- (1)
- Carrier construction
- (2)
- Cell culture and transfection
- (3)
- Analysis of dual luciferase activity
- (4)
- Statistical analysis
3. Results
3.1. circRNA Characterization
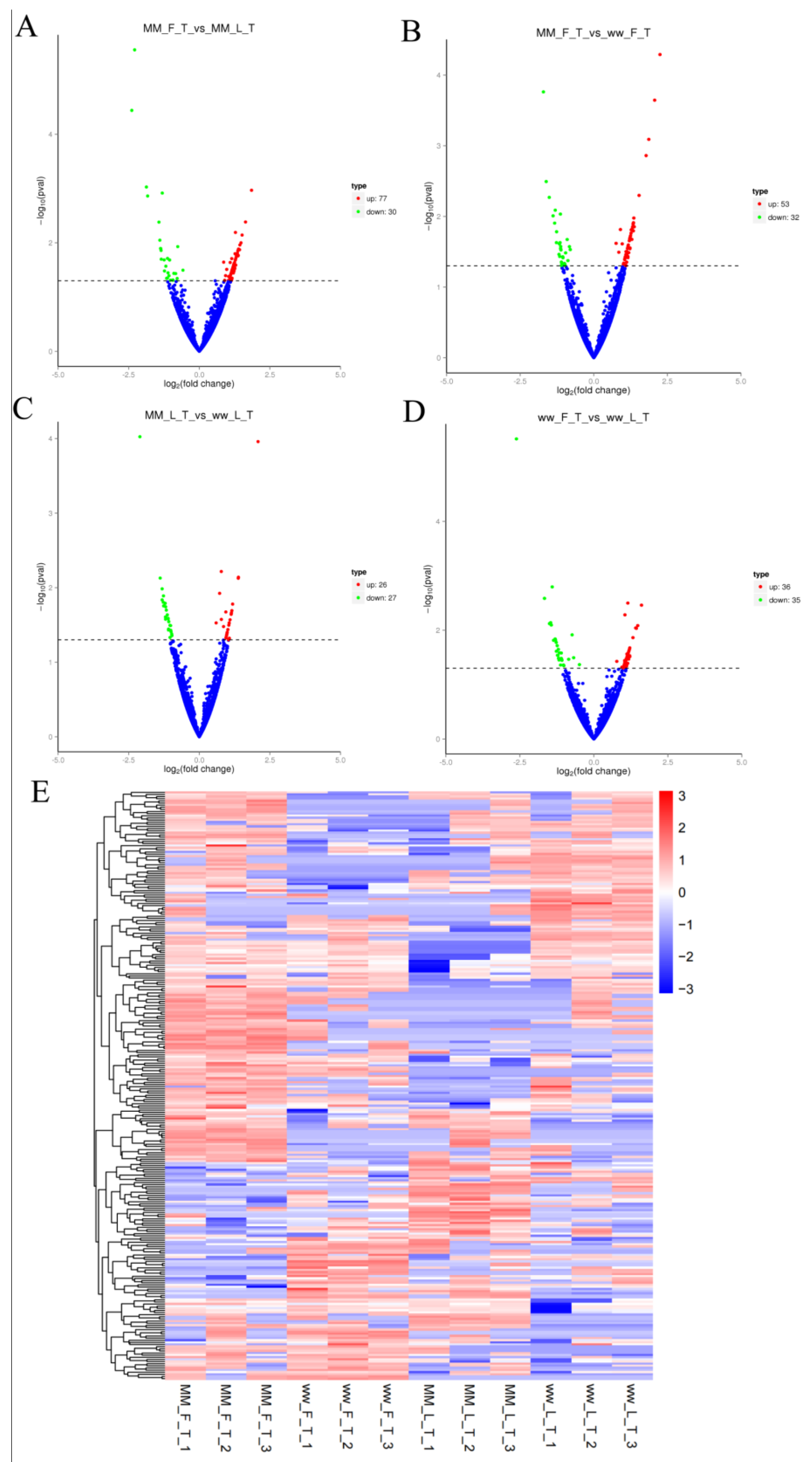
3.2. Differential Expression and Analysis of circRNA
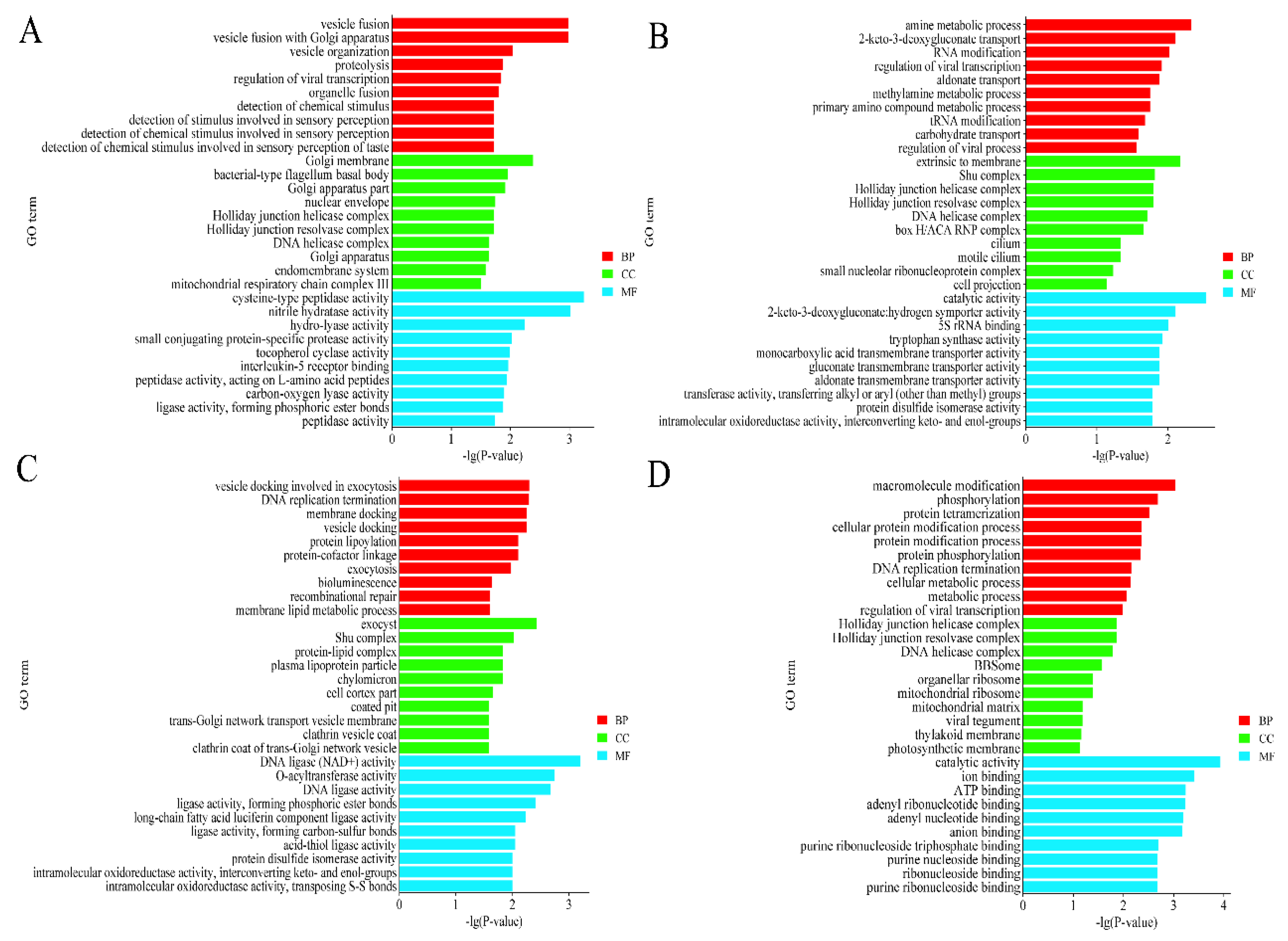
3.3. Functional Enrichment Analysis of DECs-Derived Genes
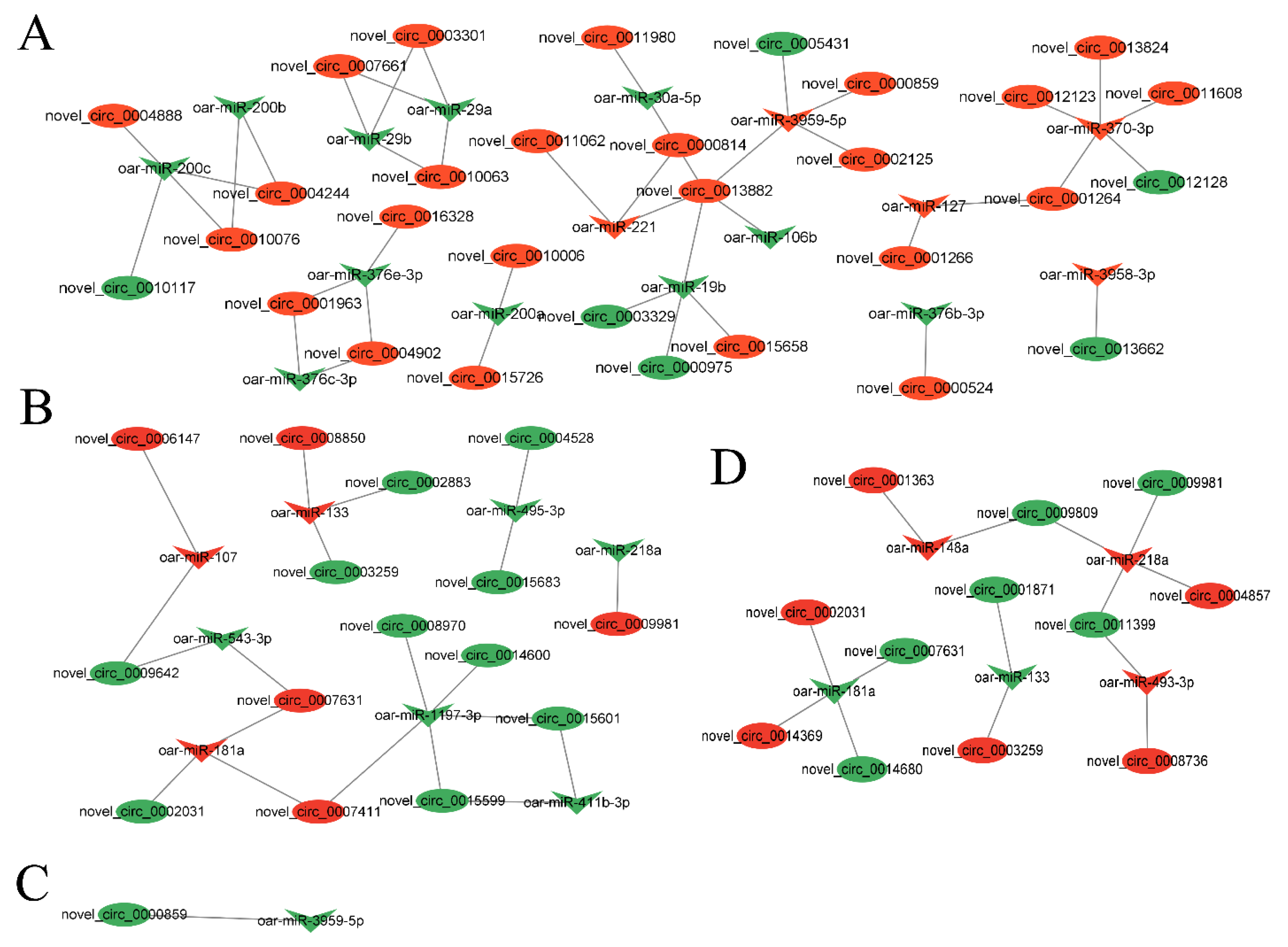
3.4. Analysis of circRNA–miRNA Co-Expression Network
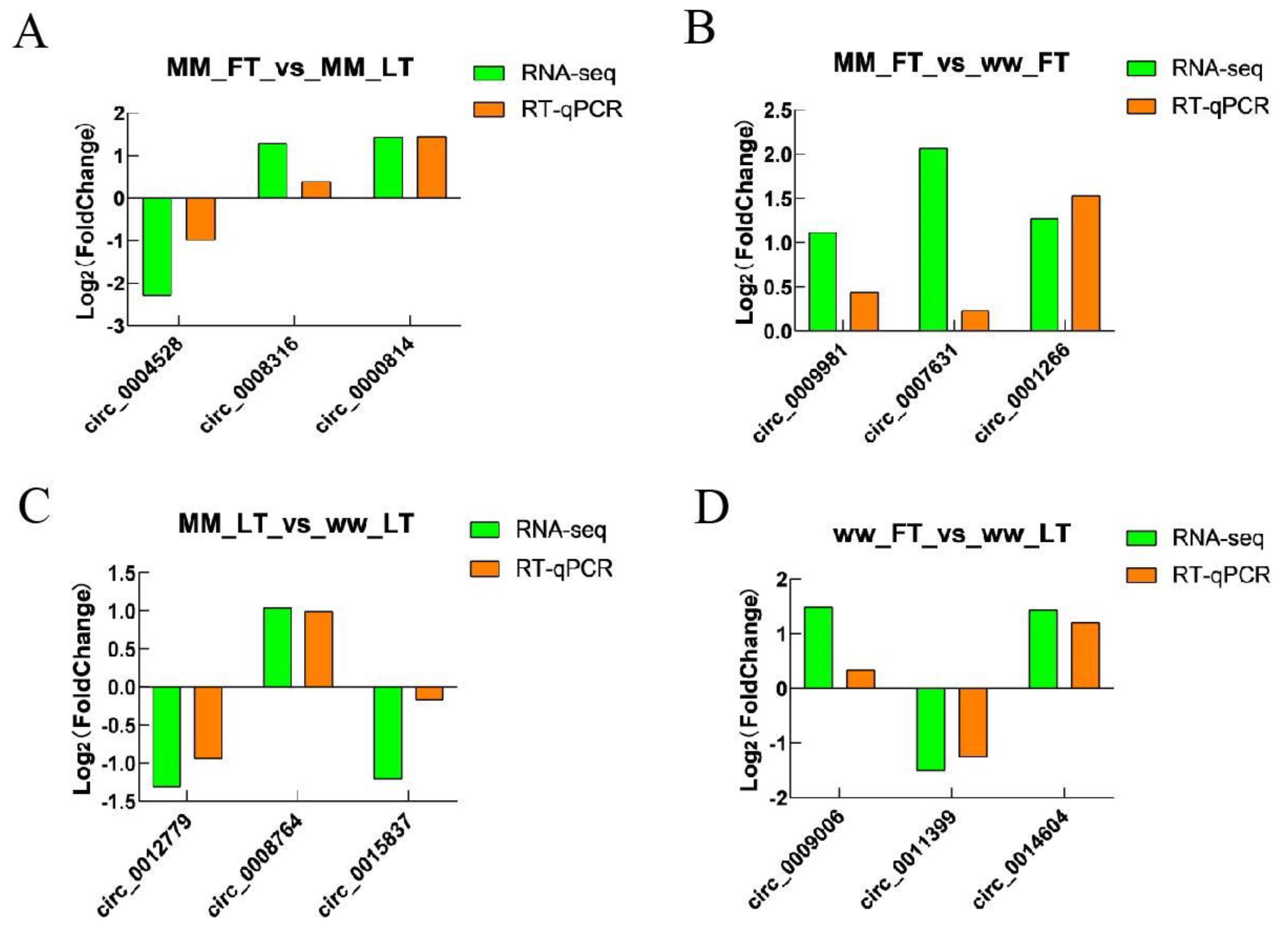
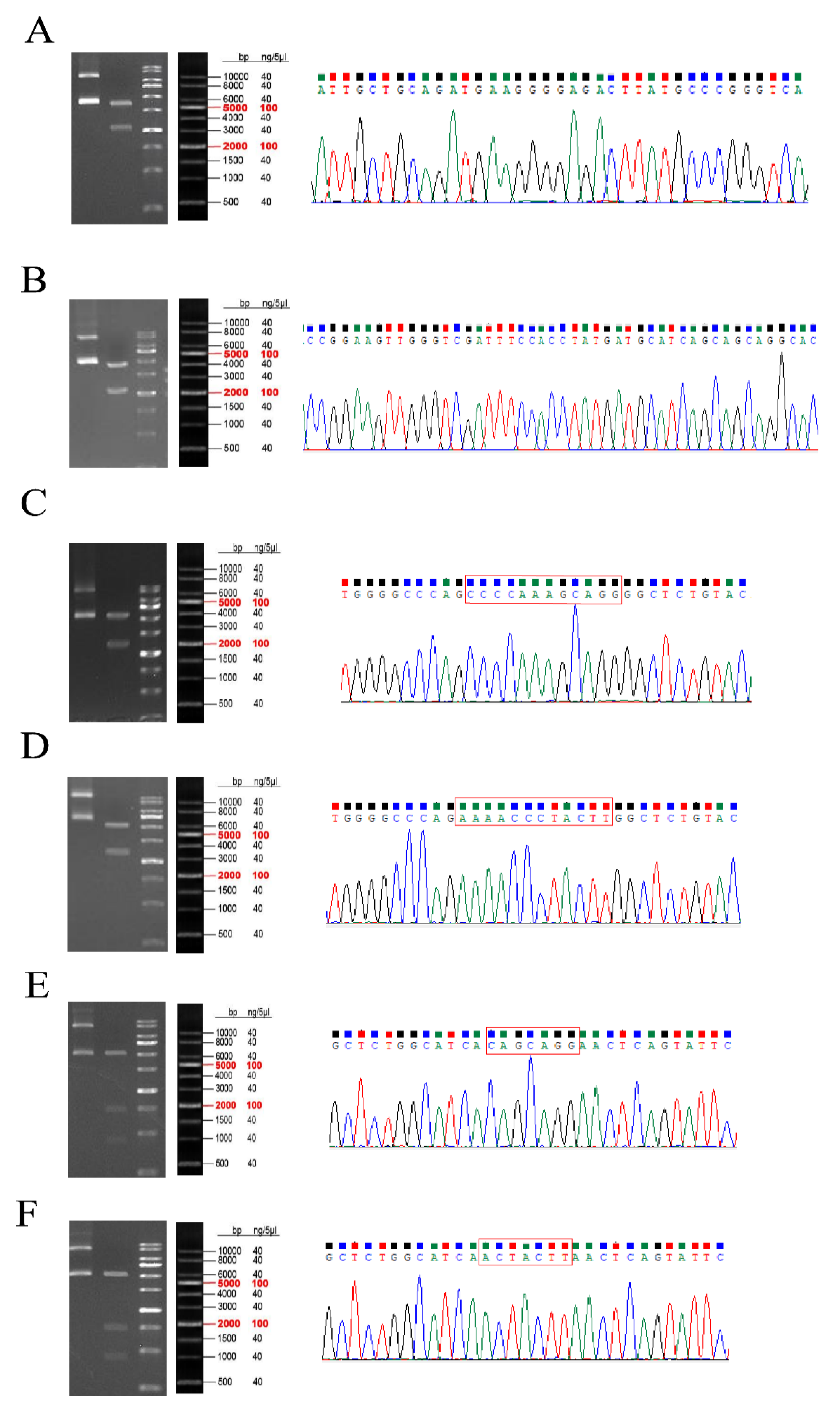
3.5. Validation of Sequencing Data
3.6. Plasmid Construction
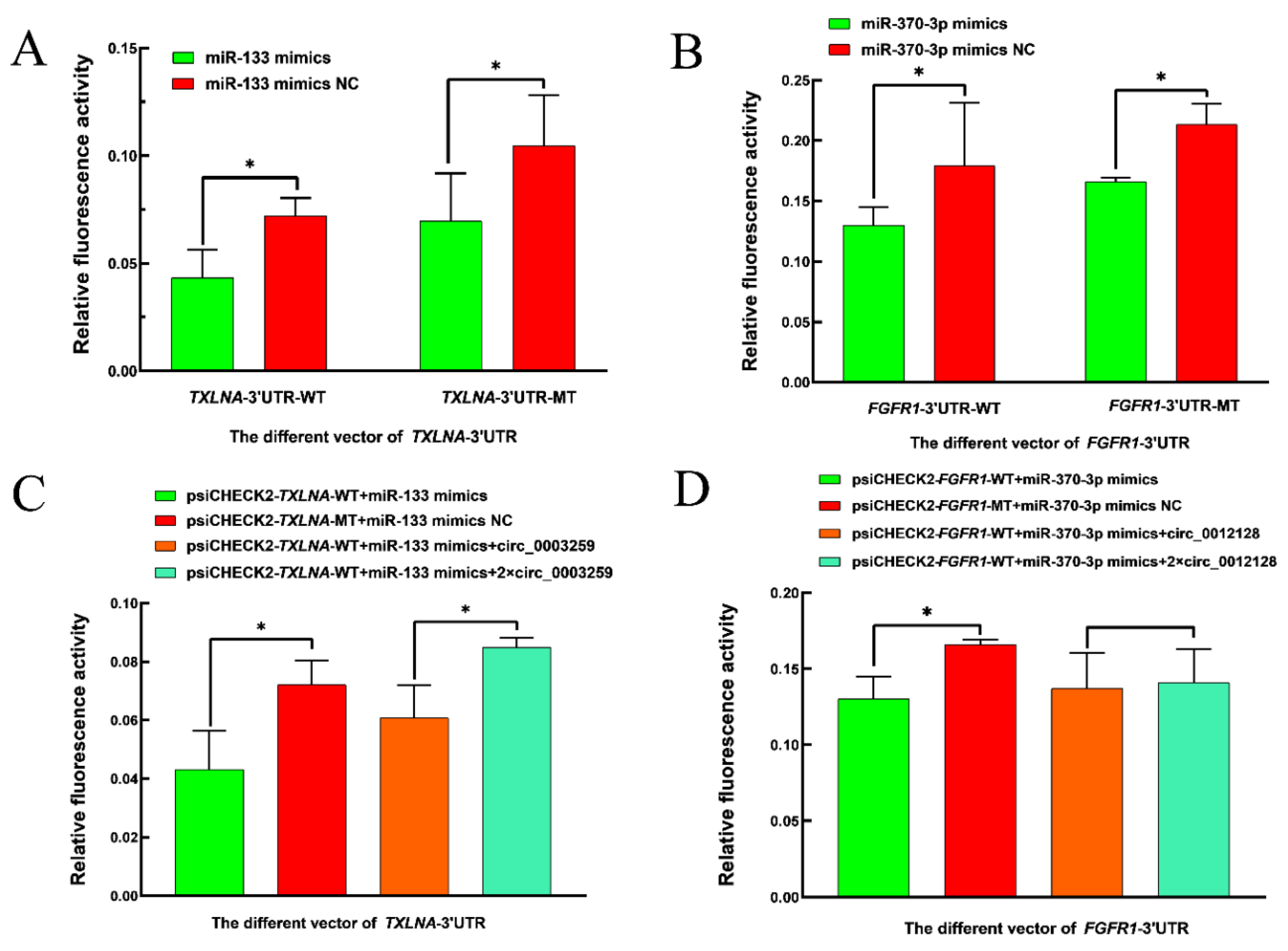
3.7. Experimental Validation of Dual Luciferase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, X.; Li, B.; Fu, S.; Wang, B.; Qi, Y.; Da, L.; Te, R.; Sun, S.; Liu, Y.; Zhang, W. Identification of piRNAs in the testes of Sunite and Small-tailed Han sheep. Anim. Biotechnol. 2021, 32, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chu, M.; Pan, L.; Wang, X.; He, X.; Zhang, R.; Tao, L.; La, Y.; Ma, L.; Di, R. Polymorphism detection of GDF9 gene and its association with litter size in Luzhong mutton sheep (Ovis aries). Animals 2021, 11, 571. [Google Scholar] [CrossRef] [PubMed]
- El-Seedy, A.S.; Hashem, N.M.; El-Azrak, K.M.; Nour El-Din, A.; Ramadan, T.A.; Taha, T.A.; Salem, M.H. Genetic screening of FecB, FecX(G) and FecX(I) mutations and their linkage with litter size in Barki and Rahmani sheep breeds. Reprod. Domest. Anim. 2017, 52, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; La, Y.; Zhou, X.; Zhang, X.; Li, F.; Liu, B. The genetic polymorphisms of TGFβ superfamily genes are associated with litter size in a Chinese indigenous sheep breed (Hu sheep). Anim. Reprod. Sci. 2018, 189, 19–29. [Google Scholar] [CrossRef]
- Dardente, H.; Simonneaux, V. GnRH and the photoperiodic control of seasonal reproduction: Delegating the task to kisspeptin and RFRP-3. J. Neuroendocrinol. 2022, 34, e13124. [Google Scholar] [CrossRef]
- Dufourny, L.; Skinner, D.C. Colocalization of progesterone receptors and thyroid hormone receptors alpha in the ovine diencephalon: No effect of estradiol. Neuroendocrinology 2003, 77, 51–58. [Google Scholar] [CrossRef]
- Abdoli, R.; Zamani, P.; Mirhoseini, S.Z.; Ghavi Hossein-Zadeh, N.; Nadri, S. A review on prolificacy genes in sheep. Reprod. Domest. Anim. 2016, 51, 631–637. [Google Scholar] [CrossRef]
- Liu, J.; Guo, M.; Hu, X.; Weng, X.; Tian, Y.; Xu, K.; Heng, D.; Liu, W.; Ding, Y.; Yang, Y.; et al. Effects of thyroid dysfunction on reproductive hormones in female rats. Chin. J. Physiol. 2018, 61, 152–162. [Google Scholar] [CrossRef]
- Kowalczyk-Zieba, I.; Staszkiewicz-Chodor, J.; Boruszewska, D.; Lukaszuk, K.; Jaworska, J.; Woclawek-Potocka, I. Hypothyroidism affects uterine function via the modulation of prostaglandin signaling. Animals 2021, 11, 2636. [Google Scholar] [CrossRef]
- He, T.; Chen, Q.; Tian, K.; Xia, Y.; Dong, G.; Yang, Z. Functional role of circRNAs in the regulation of fetal development, muscle development, and lactation in livestock. Biomed. Res. Int. 2021, 2021, 5383210. [Google Scholar] [CrossRef]
- Liu, A.; Chen, X.; Liu, M.; Zhang, L.; Ma, X.; Tian, S. Differential expression and functional analysis of circRNA in the ovaries of low and high fecundity hanper sheep. Animals 2021, 11, 1863. [Google Scholar] [CrossRef]
- Cao, Z.; Gao, D.; Xu, T.; Zhang, L.; Tong, X.; Zhang, D.; Wang, Y.; Ning, W.; Qi, X.; Ma, Y.; et al. Circular RNA profiling in the oocyte and cumulus cells reveals that circARMC4 is essential for porcine oocyte maturation. Aging 2019, 11, 8015–8034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tang, J.; He, X.; Zhu, M.; Gan, S.; Guo, X.; Zhang, X.; Zhang, J.; Hu, W.; Chu, M. Comparative transcriptomics identify key hypothalamic circular RNAs that participate in sheep (Ovis aries) reproduction. Animals 2019, 9, 557. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hong, Q.; Liu, Y.; He, X.; Di, R.; Wang, X.; Ren, C.; Zhang, Z.; Chu, M. Characterization of circular RNA profiles of oviduct reveal the potential mechanism in prolificacy trait of goat in the estrus cycle. Front. Physiol. 2022, 13, 990691. [Google Scholar] [CrossRef]
- Chang, C.; He, X.; Di, R.; Wang, X.; Han, M.; Liang, C.; Chu, M. Thyroid Transcriptomic Profiling Reveals the Follicular Phase Differential Regulation of lncRNA and mRNA Related to Prolificacy in Small Tail Han Sheep with Two FecB Genotypes. Genes 2022, 13, 849. [Google Scholar] [CrossRef] [PubMed]
- Guo, X. Study on Molecular Mechanism of FecB Gene for Fecundity in Small Tail Han Sheep. Ph.D. Thesis, China Agricultural University, Beijing, China, 2018. [Google Scholar]
- Chang, C.; He, X.; Di, R.; Wang, X.; Han, M.; Liang, C.; Chu, M. Thyroid Transcriptomics Revealed the Reproductive Regulation of miRNA in the Follicular and Luteal Phases in Small-Tail Han Sheep with Different FecB Genotypes. Genes 2023, 14, 2024. [Google Scholar] [CrossRef]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef]
- Fengzhi, L. Transcriptome Expression Analysis of Follicular and Luteal Phase Oviducts in FecB~(BB) and FecB~(++) Type Small-Tailed Cold Sheep. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2021. [Google Scholar]
- Wallace, J.M.; Martin, G.B.; Mcneilly, A.S. Changes in the secretion of LH pulses, FSH and prolactin during the preovulatory phase of the oestrous cycle of the ewe and the influence of treatment with bovine follicular fluid during the luteal phase. J. Endocrinol. 1988, 116, 123–135. [Google Scholar] [CrossRef]
- Chen, H.; Yao, W.; Jin, D.; Xia, T.; Chen, X.; Lei, T.; Zhou, L.; Yang, Z. Cloning, expression pattern, chromosomal localization, and evolution analysis of Porcine gnaq, gna11, and gna14. Biochem. Genet. 2008, 46, 398–405. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, H.; Yang, H.; Zhao, Z.; Blair, H.; Zhai, M.; Yu, Q.; Wu, P.; Fang, C.; Xie, M. Polymorphisms and association of GRM1, GNAQ and HCRTR1 genes with seasonal reproduction and litter size in three sheep breeds. Reprod. Domest. Anim. 2022, 57, 532–540. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, H.; Yang, H.; Zhao, Z.; Blair, H.T.; Liang, H.; Wu, P.; Yu, Q. Targeting GNAQ in hypothalamic nerve cells to regulate seasonal estrus in sheep. Theriogenology 2022, 181, 79–88. [Google Scholar] [CrossRef]
- Yurchenko, A.A.; Deniskova, T.E.; Yudin, N.S.; Dotsev, A.V.; Khamiruev, T.N.; Selionova, M.I.; Egorov, S.V.; Reyer, H.; Wimmers, K.; Brem, G. High-density genotyping reveals signatures of selection related to acclimation and economically important traits in 15 local sheep breeds from Russia. BMC Genom. 2019, 20 (Suppl. S3), 294. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, A.; Hughes, C.H.K.; Murphy, B.D. Orphan nuclear receptors in angiogenesis and follicular development. Reproduction 2021, 162, R35–R54. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.G.; Hosoe, M.; Fujii, S.; Kanahara, H.; Sakumoto, R. Temporal expression and localization of vascular endothelial growth factor family members in the bovine uterus during peri-implantation period. Theriogenology 2019, 133, 56–64. [Google Scholar] [CrossRef]
- El-Sheikh, M.; Mesalam, A.; Khalil, A.A.K.; Idrees, M.; Ahn, M.J.; Mesalam, A.A.; Kong, I.K. Downregulation of PI3K/AKT/mTOR pathway in juglone-treated bovine oocytes. Antioxidants 2023, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Cajas, Y.N.; Cañón-Beltrán, K.; Ladrón De Guevara, M.; Millán de la Blanca, M.G.; Ramos-Ibeas, P.; Gutiérrez-Adán, A.; Rizos, D.; González, E.M. Antioxidant nobiletin enhances oocyte maturation and subsequent embryo development and quality. Int. J. Mol. Sci. 2020, 21, 5340. [Google Scholar] [CrossRef]
- Guo, B.; Qu, X.; Chen, Z.; Yu, J.; Yan, L.; Zhu, H. Transcriptome analysis reveals transforming growth factor-β1 prevents extracellular matrix degradation and cell adhesion during the follicular-luteal transition in cows. J. Reprod. Dev. 2022, 68, 12–20. [Google Scholar] [CrossRef]
- Yaacobi-Artzi, S.; Kalo, D.; Roth, Z. Seasonal variation in the morphokinetics of in-vitro-derived bovine embryos is associated with the blastocyst developmental competence and gene expression. Front. Reprod. Health 2022, 4, 1030949. [Google Scholar] [CrossRef] [PubMed]
- Roach, C.M.; Bidne, K.L.; Romoser, M.R.; Ross, J.W.; Baumgard, L.H.; Keating, A.F. Impact of heat stress on prolactin-mediated ovarian JAK-STAT signaling in postpubertal gilts. J. Anim. Sci. 2022, 100, skac118. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, X.; Xie, J.; Wang, W.; Xin, J.; Kong, F.; Wu, Q.; Ling, Y.; Cao, X.; Li, Q. Identification of transcriptome differences in goat ovaries at the follicular phase and the luteal phase using an RNA-Seq method. Theriogenology 2020, 158, 239–249. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, M.; He, N.; Ahmed, M.F.; Wang, Y.; Zhao, R.; Yu, X.; Jin, J.; Song, J.; Zuo, Q. Hsd3b2 associated in modulating steroid hormone synthesis pathway regulates the differentiation of chicken embryonic stem cells into spermatogonial stem cells. J. Cell. Biochem. 2018, 119, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Z.; Zhao, W.; Fu, Y.; Li, B.; Cheng, J.; Deng, Y.; Li, S.; Li, H. TGF-β1 induces type I collagen deposition in granulosa cells via the AKT/GSK-3β signaling pathway-mediated MMP1 down-regulation. Reprod. Biol. 2022, 22, 100705. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, J.; Ma, S.; Zhao, F.; Zhao, W.; Chen, F.; Fu, Y.; Li, B.; Cheng, J.; Deng, Y. TGF-β1 suppresses de novo cholesterol biosynthesis in granulosa-lutein cells by down-regulating DHCR24 expression via the GSK-3β/EZH2/H3K27me3 signaling pathway. Int. J. Biol. Macromol. 2023, 224, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Wang, S.; Du, X.; Lin, C.; Zhang, J.; Pan, Z.; Li, Q. SMAD4 inhibits granulosa cell apoptosis via the miR-183-96-182 cluster and FoxO1 axis. Reprod. Sci. 2022, 29, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, D.; Zhang, J.; Li, L.; Qiu, J.; Gou, K.; Cui, S. Casein kinase 1α regulates murine spermatogenesis via p53-Sox3 signaling. Development 2022, 149, dev200205. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, Z.; Wu, G.; Zang, Z.; Zhang, J.Q.; Li, X.; Tao, J.; Shen, M.; Liu, H. FOXO1 mediates hypoxia-induced G0/G1 arrest in ovarian somatic granulosa cells by activating the TP53INP1-p53-CDKN1A pathway. Development 2021, 148, dev199453. [Google Scholar] [CrossRef]
- Wang, J.; Jia, R.; Gong, H.; Celi, P.; Zhuo, Y.; Ding, X.; Bai, S.; Zeng, Q.; Yin, H.; Xu, S.; et al. The effect of oxidative stress on the chicken ovary: Involvement of microbiota and melatonin interventions. Antioxidants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Niu, X.; Huang, Y.; Lu, H.; Li, S.; Huang, S.; Ran, X.; Wang, J. CircRNAs in Xiang pig ovaries among diestrus and estrus stages. Porc. Health Manag. 2022, 8, 29. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Z.; He, X.; Jiang, Y.; Ouyang, Y.; Hong, Q.; Chu, M. Differentially expressed circular RNA profile signatures identified in prolificacy trait of Yunshang black goat ovary at estrus cycle. Front. Physiol. 2022, 13, 820459. [Google Scholar] [CrossRef]
- Aglan, H.S.; Gebremedhn, S.; Salilew-Wondim, D.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Regulation of Nrf2 and NF-κB during lead toxicity in bovine granulosa cells. Cell Tissue Res. 2020, 380, 643–655. [Google Scholar] [CrossRef]
- Tan, W.; Dai, F.; Yang, D.; Deng, Z.; Gu, R.; Zhao, X.; Cheng, Y. MiR-93-5p promotes granulosa cell apoptosis and ferroptosis by the NF-kB signaling pathway in polycystic ovary syndrome. Front. Immunol. 2022, 13, 967151. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Shaukat, I.; Rajput, S.A.; Shukat, R.; Hanif, S.; Shaukat, I.; Zhang, X.; Chen, C.; Sun, X.; Ye, T.; et al. Ginsenoside rb1 mitigates escherichia coli lipopolysaccharide-induced endometritis through TLR4-mediated NF-κB pathway. Molecules 2021, 26, 7089. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gong, T.; Shi, F.; Xu, H.; Chen, X. Taste receptors affect male reproduction by influencing steroid synthesis. Front. Cell Dev. Biol. 2022, 10, 956981. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Thyroid hormones and female reproduction. Biol. Reprod. 2018, 99, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Fukuta, N.; Yamazaki, J.; Tominaga, M. Identification and classification of a new TRPM3 variant (γ subtype). J. Physiol. Sci. 2019, 69, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Ito, R.; Yamamoto, H.; Otsubo, S.; Matsumoto, R.; Ojima, H.; Komori, Y.; Matsuda, K.; Ihara, M. Effects of cofactors RIC-3, TMX3 and UNC-50, together with distinct subunit ratios on the agonist actions of imidacloprid on Drosophila melanogaster Dα1/Dβ1 nicotinic acetylcholine receptors expressed in Xenopus laevis oocytes. Pestic. Biochem. Physiol. 2022, 187, 105177. [Google Scholar] [CrossRef]
- Brooks, A.N.; Kilgour, E.; Smith, P.D. Molecular pathways: Fibroblast growth factor signaling: A new therapeutic opportunity in cancer. Clin. Cancer Res. 2012, 18, 1855–1862. [Google Scholar] [CrossRef]
- Wierman, M.E.; Kiseljak-Vassiliades, K.; Tobet, S. Gonadotropin-releasing hormone (GnRH) neuron migration: Initiation, maintenance and cessation as critical steps to ensure normal reproductive function. Front. Neuroendocrinol. 2011, 32, 43–52. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Z.; He, X.; Tao, L.; Jiang, Y.; Lan, R.; Hong, Q.; Chu, M. Integrated analyses of miRNA-mRNA expression profiles of ovaries reveal the crucial interaction networks that regulate the prolificacy of goats in the follicular phase. BMC Genom. 2021, 22, 812. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Volume |
|---|---|
| PrimeScript RT Enzyme Mix I | 1 µL |
| Oligo dT Primer | 1 µL |
| Random 6 Mers | 1 µL |
| 5 × PrimeScript Buffer | 4 µL |
| Total RNA (500 ng/µL) | 2 µL |
| RNase-Free ddH2O | 11 µL |
| Gene Name | Primer Sequences (5’-3’) | Accession No. | Tm (°C) |
|---|---|---|---|
| circ_0004528 | F: TGATGAGCAGCTTTGCAGAAGA | 101123020 | 60 |
| R: CTCGTGCATGCGTCTCTTGAC | |||
| circ_0008316 | F: AGCCAACAAGATGAGATCGACAG | 101103576 | 60 |
| R: CTCAAGTTCCTCGTGCTGGGA | |||
| circ_0000814 | F: AGGGGAAAAGTCCTTGATGCAA | 101106049 | 60 |
| R: GCTACATCCATCTTTGAACGTGC | |||
| circ_0009981 | F: GGGATCAAGGTCATTCACAAGC | 101102742 | 60 |
| R: TTTCTCCCCAGCCAGTACAAAG | |||
| circ_0007631 | F: GGCTGTCTCTATTTTGGGAGATG | 101112983 | 60 |
| R: GCATAGGCCTTTCCTTCTGTG | |||
| circ_0001266 | F: TCAGCTACACAGTTGCCCCAA | 101115254 | 60 |
| R: TCTTTCCCCTGAGTTGCCTCG | |||
| circ_0012779 | F: GCAACTGCTACTGGAAGAGGAG | 101101794 | 60 |
| R: ACTGGCTCATGCTTCACTGGA | |||
| circ_0008764 | F: CTGGTCTCTGGAGGTGTTCTG | 101105760 | 60 |
| R: CGTTGCTGCAAGTTCACTCC | |||
| circ_0015837 | F: TGTGTCCGTCTACAGATGTGG | 101105629 | 60 |
| R: GTCATGAGTTCATCCACACTGG | |||
| circ_0009006 | F: GTCCTGGACTTGGCCGTGTAT | 101102570 | 60 |
| R: CCGGCACCACTTGTCAATGTG | |||
| circ_0011399 | F: CAGTTCCCTGGGTTTGCACAC | 101121697 | 60 |
| R: GGGACACACACTGAACCAGCT | |||
| circ_0014604 | F: ACGTGAACCTTGCTTCTGGC | 101119353 | 60 |
| R: TGCGATGGAGCAGAGCAGTT | |||
| GRPDH | F: ATCGCCAATGCCAACTC | NM_001190390.1 | 60 |
| R: CCTTTCGCTTACCTATACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.; He, X.; Di, R.; Wang, X.; Han, M.; Liang, C.; Chu, M. Transcriptome Analysis Reveals Differentially Expressed circRNAs Associated with Fecundity in Small-Tail Han Sheep Thyroid with Different FecB Genotypes. Animals 2024, 14, 105. https://doi.org/10.3390/ani14010105
Chang C, He X, Di R, Wang X, Han M, Liang C, Chu M. Transcriptome Analysis Reveals Differentially Expressed circRNAs Associated with Fecundity in Small-Tail Han Sheep Thyroid with Different FecB Genotypes. Animals. 2024; 14(1):105. https://doi.org/10.3390/ani14010105
Chicago/Turabian StyleChang, Cheng, Xiaoyun He, Ran Di, Xiangyu Wang, Miaoceng Han, Chen Liang, and Mingxing Chu. 2024. "Transcriptome Analysis Reveals Differentially Expressed circRNAs Associated with Fecundity in Small-Tail Han Sheep Thyroid with Different FecB Genotypes" Animals 14, no. 1: 105. https://doi.org/10.3390/ani14010105