
Association of the Infant Gut Microbiome with Temperament at Nine Months of Age: A Michigan Cohort Study
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Data Collection
2.3. Fecal Microbiota Analysis
2.4. Statistical Analysis
3. Results
3.1. Study Population
3.2. Alpha Diversity and Temperament Scores
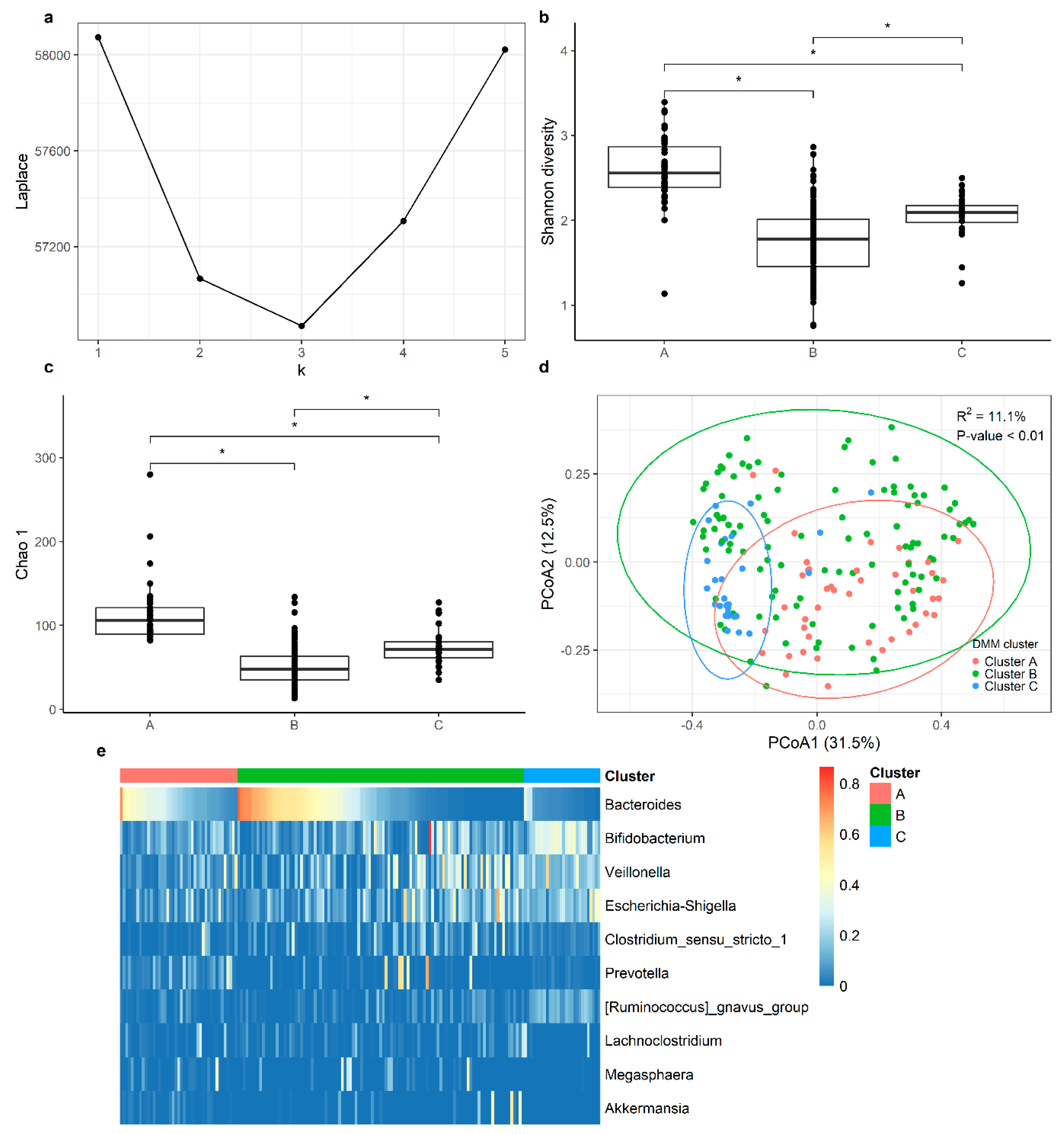
3.3. Cluster Analysis
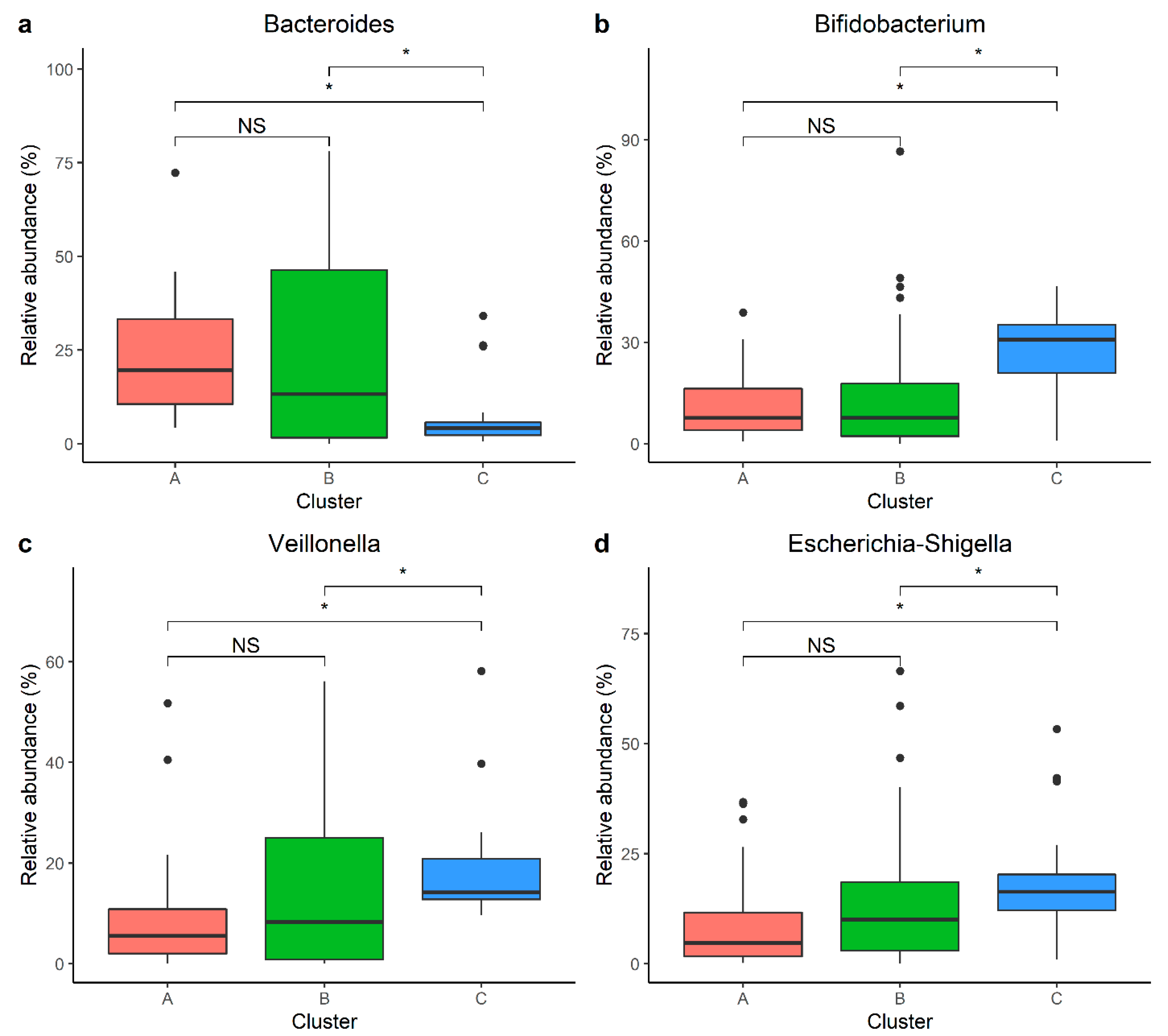
3.4. Individual Taxa Analysis
4. Discussion
4.1. Alpha Diversity
4.2. Cluster Analysis
4.3. Individual Taxa
4.4. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Scale | Cluster | No Vitamin D (59) | Vitamin D (100) | ||||
|---|---|---|---|---|---|---|---|
| Beta | p-Value | q-Value 2 | Beta | p-Value | q-Value 2 | ||
| Positive affect/surgency | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.06 (−0.38, 0.50) | 0.78 | 0.84 | 0.14 (−0.27, 0.56) | 0.50 | 0.50 | |
| Cluster C | 0.31 (−0.22, 0.84) | 0.24 | 0.36 | 0.28 (−0.21, 0.76) | 0.26 | 0.26 | |
| Negative Emotionality | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.58 (−0.06, 1.23) | 0.08 | 0.24 | 0.60 (−0.04, 1.23) | 0.06 | 0.18 | |
| Cluster C | 0.94 (0.17, 1.71) | 0.02 * | 0.06 | 0.59 (−0.15, 1.33) | 0.11 | 0.26 | |
| Orienting/regulatory capacity | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.05 (−0.47, 0.57) | 0.84 | 0.84 | 0.19 (−0.27, 0.65) | 0.42 | 0.50 | |
| Cluster C | −0.03 (0.64, 0.59) | 0.93 | 0.93 | 0.37 (−0.16, 0.91) | 0.17 | 0.26 | |
| Scale | Cluster | Male (84) | Female (75) | ||||
|---|---|---|---|---|---|---|---|
| Beta | p-Value | q-Value 2 | Beta | p-Value | q-Value 2 | ||
| Positive affect/surgency | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.20 (−0.21, 0.62) | 0.34 | 0.34 | 0.06 (−0.30, 0.43) | 0.73 | 0.73 | |
| Cluster C | 0.26 (−0.23, 0.74) | 0.30 | 0.45 | 0.29 (−0.21, 0.79) | 0.25 | 0.38 | |
| Negative Emotionality | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.86 (0.27, 1.46) | 0.005 * | 0.02 * | 0.18 (−0.40, 0.76) | 0.53 | 0.73 | |
| Cluster C | 0.92 (0.25, 1.60) | 0.008 * | 0.02 * | 0.52 (−0.27, 1.30) | 0.19 | 0.38 | |
| Orienting/regulatory capacity | Cluster A | ref | - | - | ref | - | - |
| Cluster B | 0.39 (−0.12, 0.90) | 0.13 | 0.20 | −0.12 (−0.50, 0.26) | 0.54 | 0.73 | |
| Cluster C | 0.20 (−0.38, 0.78) | 0.49 | 0.50 | 0.08 (−0.44, 0.59) | 0.76 | 0.76 | |
| Phylum | Family | Genus | DESeq2 | ANCOMBC | MaAslin |
|---|---|---|---|---|---|
| Firmicutes | Christensenellaceae | R-7 group | 1.87 | 0.39 | 0.52 |
| Firmicutes | Oscillospiraceae | UCG-002 | 1.69 | 0.37 | 0.52 |
| Firmicutes | Peptostreptococcaceae | Clostridioides | −1.26 | −0.44 | −0.49 |
| Firmicutes | Ruminococcaceae | Subdoligranulum | 1.13 | 0.38 | 0.51 |
References
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B.; Galla, S.; Chakraborty, S.; Cheng, X.; Yeo, J.; Mell, B.; Zhang, H.; et al. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Hakansson, A.; Molin, G. Gut microbiota and inflammation. Nutrients 2011, 3, 637–682. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Gut bacteria in health and disease. Gastroenterol. Hepatol. 2013, 9, 560–569. [Google Scholar]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.-F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; McVey Neufeld, K.-A. Gut–brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Burokas, A.; Moloney, R.D.; Dinan, T.G.; Cryan, J.F. Microbiota regulation of the Mammalian gut-brain axis. Adv. Appl. Microbiol. 2015, 91, 1–62. [Google Scholar]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Kubo, C.; Koga, Y.; Yu, X.-N. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, K.-A.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Effects of intestinal microbiota on anxiety-like behavior. Commun. Integr. Biol. 2011, 4, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Cruz, N.J.; Kang, D.-W.; Gandal, M.J.; Wang, B.; Kim, Y.-M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell 2019, 177, 1600–1618.e17. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse Microbiota Models: Comparing Germ-Free Mice and Antibiotics Treatment as Tools for Modifying Gut Bacteria. Front. Physiol. 2018, 9, 1534. [Google Scholar] [CrossRef] [PubMed]
- Fiebiger, U.; Bereswill, S.; Heimesaat, M.M. Dissecting the Interplay Between Intestinal Microbiota and Host Immunity in Health and Disease: Lessons Learned from Germfree and Gnotobiotic Animal Models. Eur. J. Microbiol. Immunol. 2016, 6, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e3. [Google Scholar] [CrossRef]
- Desbonnet, L.; Clarke, G.; Traplin, A.; O’sullivan, O.; Crispie, F.; Moloney, R.D.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015, 48, 165–173. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.-L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Sampson, T.R.; Mazmanian, S.K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe 2015, 17, 565–576. [Google Scholar] [CrossRef]
- Cenit, M.C.; Sanz, Y.; Codoñer-Franch, P. Influence of gut microbiota on neuropsychiatric disorders. World J. Gastroenterol. 2017, 23, 5486–5498. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-N.; Yun, Y.; Ryu, S.; Chang, Y.; Kwon, M.-J.; Cho, J.; Shin, H.; Kim, H.-L. Correlation between gut microbiota and personality in adults: A cross-sectional study. Brain Behav. Immun. 2018, 69, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Białecka-Dębek, A.; Granda, D.; Szmidt, M.K.; Zielińska, D. Gut Microbiota, Probiotic Interventions, and Cognitive Function in the Elderly: A Review of Current Knowledge. Nutrients 2021, 13, 2514. [Google Scholar] [CrossRef] [PubMed]
- Borre, Y.E.; O’keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.-W.; Park, J.G.; Ilhan, Z.E.; Wallstrom, G.; LaBaer, J.; Adams, J.B.; Krajmalnik-Brown, R. Reduced Incidence of Prevotella and Other Fermenters in Intestinal Microflora of Autistic Children. PLoS ONE 2013, 8, e68322. [Google Scholar] [CrossRef] [PubMed]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 24. [Google Scholar] [CrossRef]
- Gondalia, S.V.; Palombo, E.A.; Knowles, S.R.; Cox, S.B.; Meyer, D.; Austin, D.W. Molecular characterisation of gastrointestinal microbiota of children with autism (with and without gastrointestinal dysfunction) and their neurotypical siblings. Autism Res. 2012, 5, 419–427. [Google Scholar] [CrossRef]
- Son, J.S.; Zheng, L.J.; Rowehl, L.M.; Tian, X.; Zhang, Y.; Zhu, W.; Litcher-Kelly, L.; Gadow, K.D.; Gathungu, G.; Robertson, C.E.; et al. Comparison of Fecal Microbiota in Children with Autism Spectrum Disorders and Neurotypical Siblings in the Simons Simplex Collection. PLoS ONE 2015, 10, e0137725. [Google Scholar] [CrossRef]
- Sordillo, J.E.; Korrick, S.; Laranjo, N.; Carey, V.; Weinstock, G.M.; Gold, D.R.; O’Connor, G.; Sandel, M.; Bacharier, L.B.; Beigelman, A. Association of the Infant Gut Microbiome With Early Childhood Neurodevelopmental Outcomes: An Ancillary Study to the VDAART Randomized Clinical Trial. JAMA Netw. Open 2019, 2, e190905. [Google Scholar] [CrossRef]
- Loughman, A.; Ponsonby, A.-L.; O’Hely, M.; Symeonides, C.; Collier, F.; Tang, M.L.; Carlin, J.; Ranganathan, S.; Allen, K.; Pezic, A.; et al. Gut microbiota composition during infancy and subsequent behavioural outcomes. EBioMedicine 2020, 52, 102640. [Google Scholar] [CrossRef]
- Carlson, A.L.; Xia, K.; Azcarate-Peril, M.A.; Rosin, S.P.; Fine, J.P.; Mu, W.; Zopp, J.B.; Kimmel, M.C.; Styner, M.A.; Thompson, A.L.; et al. Infant gut microbiome composition is associated with non-social fear behavior in a pilot study. Nat. Commun. 2021, 12, 3294. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Stilling, R.M.; Stanton, C.; Cryan, J.F. Collective unconscious: How gut microbes shape human behavior. J. Psychiatr. Res. 2015, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, M.K.; Sheese, B.E.; Rueda, M.R.; Posner, M.I. Developing Mechanisms of Self-Regulation in Early Life. Emot. Rev. 2011, 3, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, M.K.; Ahadi, S.A.; Evans, D.E. Temperament and personality: Origins and outcomes. J. Personal. Soc. Psychol. 2000, 78, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Rettew, D.C.; McKee, L. Temperament and its role in developmental psychopathology. Harv. Rev. Psychiatry 2005, 13, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Crawford, H.; Morales, S.; Degnan, K.A.; Pine, D.S.; Fox, N.A. Infant behavioral inhibition predicts personality and social outcomes three decades later. Proc. Natl. Acad. Sci. USA 2020, 117, 9800–9807. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, J.V.; Gibbons, R.D.; Christoffel, K.K.; Arend, R.; Rosenbaum, D.; Binns, H.; Dawson, N.; Sobel, H.; Isaacs, C. Prevalence rates and correlates of psychiatric disorders among preschool children. J. Am. Acad. Child Adolesc. Psychiatry 1996, 35, 204–214. [Google Scholar] [CrossRef]
- Tamana, S.K.; Tun, H.M.; Konya, T.; Chari, R.S.; Field, C.J.; Guttman, D.S.; Becker, A.B.; Moraes, T.J.; Turvey, S.E.; Subbarao, P.; et al. Bacteroides-dominant gut microbiome of late infancy is associated with enhanced neurodevelopment. Gut Microbes 2021, 13, 1930875. [Google Scholar] [CrossRef]
- About|CHARM Study. Available online: https://charmstudy.epibio.msu.edu/ (accessed on 1 April 2022).
- Tylavsky, F.A.; Ferrara, A.; Catellier, D.J.; Oken, E.; Li, X.; Law, A.; Dabelea, D.; Rundle, A.; Gilbert-Diamond, D.; Hivert, M.-F.; et al. Understanding childhood obesity in the US: The NIH environmental influences on child health outcomes (ECHO) program. Int. J. Obes. 2020, 44, 617–627. [Google Scholar] [CrossRef]
- Haddad, E.N.; Comstock, S.S. Archive for Research in Child Health (ARCH) and Baby Gut: Study Protocol for a Remote, Prospective, Longitudinal Pregnancy and Birth Cohort to Address Microbiota Development and Child Health. Methods Protoc. 2021, 4, 52. [Google Scholar] [CrossRef]
- Ma, T.; Bu, S.; Paneth, N.; Kerver, J.M.; Comstock, S.S. Vitamin D Supplementation in Exclusively Breastfed Infants Is Associated with Alterations in the Fecal Microbiome. Nutrients 2022, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Putnam, S.P.; Helbig, A.L.; Gartstein, M.A.; Rothbart, M.K.; Leerkes, E. Development and assessment of short and very short forms of the infant behavior questionnaire-revised. J. Pers. Assess. 2014, 96, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, M.K. Measurement of Temperament in Infancy. Child Dev. 1981, 52, 569–578. [Google Scholar] [CrossRef]
- Rothbart, M.K.; Ahadi, S.A.; Hershey, K.L.; Fisher, P. Investigations of temperament at three to seven years: The Children’s Behavior Questionnaire. Child Dev. 2001, 72, 1394–1408. [Google Scholar] [CrossRef] [PubMed]
- Sugino, K.Y.; Paneth, N.; Comstock, S.S. Michigan cohorts to determine associations of maternal pre-pregnancy body mass index with pregnancy and infant gastrointestinal microbial communities: Late pregnancy and early infancy. PLoS ONE 2019, 14, e0213733. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Nigam, K.; Mccallum, A.K.; Thrun, S.; Mitchell, T. Text Classification from Labeled and Unlabeled Documents using EM. Mach. Learn. 2000, 39, 103–134. [Google Scholar] [CrossRef]
- Holmes, I.; Harris, K.; Quince, C. Dirichlet Multinomial Mixtures: Generative Models for Microbial Metagenomics. PLoS ONE 2012, 7, e30126. [Google Scholar] [CrossRef] [PubMed]
- DirichletMultinomial: Dirichlet-Multinomial Mixture Model Machine Learning for Microbiome Data Version 1.32.0 from Bioconductor. Available online: https://rdrr.io/bioc/DirichletMultinomial/ (accessed on 21 October 2022).
- Acuña, I.; Cerdó, T.; Ruiz, A.; Torres-Espínola, F.J.; López-Moreno, A.; Aguilera, M.; Suárez, A.; Campoy, C. Infant Gut Microbiota Associated with Fine Motor Skills. Nutrients 2021, 13, 1673. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Fox, M.; Lee, S.M.; Wiley, K.S.; Lagishetty, V.; Sandman, C.A.; Jacobs, J.P.; Glynn, L.M. Development of the infant gut microbiome predicts temperament across the first year of life. Dev. Psychopathol. 2021, 34, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Peddada, S.D. Analysis of compositions of microbiomes with bias correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable association discovery in population-scale meta-omics studies. PLoS Comput. Biol. 2021, 17, e1009442. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLOS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Kelsey, C.M.; Prescott, S.; McCulloch, J.A.; Trinchieri, G.; Valladares, T.L.; Dreisbach, C.; Alhusen, J.; Grossmann, T. Gut microbiota composition is associated with newborn functional brain connectivity and behavioral temperament. Brain Behav. Immun. 2021, 91, 472–486. [Google Scholar] [CrossRef]
- Rothbart, M.K. Becoming Who We Are: Temperament and Personality in Development; Guilford Press: New York, NY, USA, 2011. [Google Scholar]
- Lemery, K.S.; Essex, M.J.; Smider, N.A. Revealing the relation between temperament and behavior problem symptoms by eliminating measurement confounding: Expert ratings and factor analyses. Child Dev. 2002, 73, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Lengua, L.J.; Wolchik, S.A.; Sandler, I.N.; West, S.G. The additive and interactive effects of parenting and temperament in predicting adjustment problems of children of divorce. J. Clin. Child Psychol. 2000, 29, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Aatsinki, A.-K.; Lahti, L.; Uusitupa, H.-M.; Munukka, E.; Keskitalo, A.; Nolvi, S.; O’Mahony, S.; Pietilä, S.; Elo, L.L.; Eerola, E.; et al. Gut microbiota composition is associated with temperament traits in infants. Brain Behav. Immun. 2019, 80, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Ogbu, D.; Xia, E.; Sun, J. Gut instincts: Vitamin D/vitamin D receptor and microbiome in neurodevelopment disorders. Open Biol. 2020, 10, 200063. [Google Scholar] [CrossRef] [PubMed]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Jašarević, E.; Morrison, K.E.; Bale, T.L. Sex differences in the gut microbiome–brain axis across the lifespan. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150122. [Google Scholar] [CrossRef]
- Alur, P. Sex Differences in Nutrition, Growth, and Metabolism in Preterm Infants. Front. Pediatr. 2019, 7, 22. Available online: https://www.frontiersin.org/article/10.3389/fped.2019.00022 (accessed on 1 April 2022). [CrossRef]
- Loughman, A.; Quinn, T.; Nation, M.L.; Reichelt, A.; Moore, R.J.; Van, T.T.H.; Sung, V.; Tang, M.L.K. Infant microbiota in colic: Predictive associations with problem crying and subsequent child behavior. J. Dev. Orig. Health Dis. 2021, 12, 260–270. [Google Scholar] [CrossRef]
- Mounsey, A.; Smith, K.L.; Reddy, V.C.; Nickolich, S. Clostridioides difficile Infection: Update on Management. Am. Fam. Physician 2020, 101, 168–175. [Google Scholar]
- Schutze, G.E.; Willoughby, R.E.; Committee on Infectious Diseases. Clostridium difficile infection in infants and children. Pediatrics 2013, 131, 196–200. [Google Scholar]
- Al-Owain, M.; Kaya, N.; Al-Shamrani, H.; Al-Bakheet, A.; Qari, A.; Al-Muaigl, S.; Ghaziuddin, M. Autism Spectrum Disorder in a Child with Propionic Acidemia. JIMD Rep. 2012, 7, 63–66. [Google Scholar] [PubMed]
- Cong, X.; Henderson, W.A.; Graf, J.; McGrath, J.M. Early Life Experience and Gut Microbiome: The Brain-Gut-Microbiota Signaling System. Adv. Neonatal Care 2015, 15, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]


| Scale | Positive Affect/Surgency | Negative Emotionality | Orienting/Regulatory Capacity | ||||
|---|---|---|---|---|---|---|---|
| Variable 1 | n (%) or Mean (SD) | Mean (SD) or β (95% CI) | p-Value | Mean (SD) or β (95% CI) | p-Value | Mean (SD) or β (95% CI) | p-Value |
| Categorical variable | |||||||
| Maternal education level, n (%) | |||||||
| Did not finish high school | 4 (2.5%) | 5.92 (0.98) | 0.18 | 4.92 (1.05) | 0.22 | 4.90 (1.10) | 0.004 * |
| High school graduate or GED | 23 (14.5%) | 5.74 (0.69) | 4.30 (1.0) | 5.76 (0.53) | |||
| Some college | 40 (25.2%) | 5.59 (0.67) | 4.13 (0.95) | 5.49 (0.70) | |||
| College graduate or above | 92 (57.9%) | 5.46 (0.64) | 4.02 (0.96) | 5.33 (0.67) | |||
| Race of mother, n (%) | |||||||
| White | 126 (80.3%) | 5.46 (0.63) | <0.001 * | 4.02 (0.94) | 0.03 * | 5.29 (0.69) | 0.002 * |
| Black | 21 (13.2%) | 5.82 (0.73) | 4.53 (0.88) | 5.85 (0.58) | |||
| Other | 12 (7.5%) | 6.04 (0.70) | 4.42 (1.25) | 5.66 (0.56) | |||
| Infant sex, n (%) | |||||||
| Male | 84 (52.8%) | 5.72 (0.64) | <0.001 * | 4.14 (0.96) | 0.69 | 5.40 (0.73) | 0.77 |
| Female | 75 (47.2%) | 5.36 (0.65) | 4.08 (0.98) | 5.37 (0.65) | |||
| Breastfeeding status, n (%) | |||||||
| Exclusive breastfeeding | 90 (56.6%) | 5.51 (0.67) | 0.37 | 4.17 (0.95) | 0.68 | 5.35 (0.73) | 0.10 |
| Partial breastfeeding | 26 (16.4%) | 5.72 (0.63) | 4.03 (0.96) | 5.57 (0.76) | |||
| Not breastfeeding | 43 (27.0%) | 5.53 (0.68) | 4.02 (1.02) | 5.49 (0.67) | |||
| Continuous variable | |||||||
| Maternal age (year), mean (SD) | 31.4 (5.1) | −0.03 (−0.05, −0.008) | 0.006 * | −0.04 (−0.07, −0.01) | 0.009 * | −0.006 (−0.03, 0.02) | 0.61 |
| Pre-pregnancy BMI, mean (SD) | 27.8 (7.2) | 0.01 (−0.005, 0.02) | 0.18 | 0.001 (−0.02, 0.02) | 0.96 | 0.01 (−0.002, 0.03) | 0.09 |
| Gestational age at delivery (week), mean (SD) | 38.8 (1.5) | 0.03 (−0.04, 0.10) | 0.38 | 0.02 (−0.08, 0.13) | 0.67 | −0.05 (−0.13, 0.02) | 0.14 |
| Infant age at IBQ measurement (month), mean (SD) | 9.4 (0.7) | 0.004 (−0.001, 0.008) | 0.14 | 0.04 (−0.003, 0.01) | 0.32 | 0.002 (−0.004, 0.007) | 0.55 |
| Shannon | Chao 1 | |||
|---|---|---|---|---|
| Scale | Beta | p-Value | Beta | p-Value |
| Positive affect/surgency | 0.11 | 0.27 | 0.001 | 0.63 |
| Negative Emotionality | −0.18 | 0.24 | −0.003 | 0.12 |
| Orienting/regulatory capacity | 0.01 | 0.92 | −0.001 | 0.62 |
| Scale | Cluster | Univariate Model | Multivariate Model | ||||
|---|---|---|---|---|---|---|---|
| Beta | p-Value | q-Value | Beta | p-Value | q-Value 2 | ||
| Positive affect/surgency | Cluster C | ref | - | - | ref | - | - |
| Cluster B | 0.17 (−0.11, 0.46) | 0.22 | 0.22 | 0.14 (−0.13, 0.41) | 0.30 | 0.31 | |
| Cluster A | 0.41 (0.08, 0.74) | 0.01 * | 0.03 * | 0.29 (−0.03, 0.61) | 0.07 | 0.11 | |
| Negative emotionality | Cluster C | ref | - | ref | - | - | |
| Cluster B | 0.44 (0.03, 0.85) | 0.04 * | 0.12 | 0.44 (0.02, 0.86) | 0.04 * | 0.12 | |
| Cluster A | 0.57 (0.09, 1.05) | 0.02 * | 0.03 * | 0.55 (0.05, 1.04) | 0.03 * | 0.09 | |
| Orienting/regulatory capacity | Cluster C | ref | - | ref | - | - | |
| Cluster B | 0.20 (−0.11, 0.51) | 0.21 | 0.22 | 0.16 (−0.15, 0.47) | 0.31 | 0.31 | |
| Cluster A | 0.24 (−0.12, 0.61) | 0.20 | 0.20 | 0.20 (−0.17, 0.57) | 0.30 | 0.30 | |
| Phylum | Family | Genus | Q Value for DESeq2 2 | Q Value for ANCOMBC | Q Value for MaAslin |
|---|---|---|---|---|---|
| Firmicutes | Christensenellaceae | Christensenellaceae R-7 group | 6.3 × 10−7 | 0.03 | 0.11 |
| Firmicutes | Oscillospiraceae | UCG-002 | 6.3 × 10−7 | 0.03 | 0.11 |
| Firmicutes | Peptostreptococcaceae | Clostridioides | 1.0 × 10−3 | 0.11 | 0.16 |
| Firmicutes | Ruminococcaceae | Subdoligranulum | 8.7 × 10−4 | 0.23 | 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Bu, S.; Nzerem, A.C.; Paneth, N.; Kerver, J.M.; Cavalieri, C.N.; Comstock, S.S. Association of the Infant Gut Microbiome with Temperament at Nine Months of Age: A Michigan Cohort Study. Microorganisms 2024, 12, 214. https://doi.org/10.3390/microorganisms12010214
Ma T, Bu S, Nzerem AC, Paneth N, Kerver JM, Cavalieri CN, Comstock SS. Association of the Infant Gut Microbiome with Temperament at Nine Months of Age: A Michigan Cohort Study. Microorganisms. 2024; 12(1):214. https://doi.org/10.3390/microorganisms12010214
Chicago/Turabian StyleMa, Tengfei, Sihan Bu, Adannaya C. Nzerem, Nigel Paneth, Jean M. Kerver, Cybil Nicole Cavalieri, and Sarah S. Comstock. 2024. "Association of the Infant Gut Microbiome with Temperament at Nine Months of Age: A Michigan Cohort Study" Microorganisms 12, no. 1: 214. https://doi.org/10.3390/microorganisms12010214