
Biofilm Formation of Clostridioides difficile, Toxin Production and Alternatives to Conventional Antibiotics in the Treatment of CDI

Department of Microbiology, Stellenbosch University, Stellenbosch 7600, South Africa
Microorganisms 2023, 11(9), 2161; https://doi.org/10.3390/microorganisms11092161
Submission received: 29 June 2023
/
Revised: 16 August 2023
/
Accepted: 24 August 2023
/
Published: 26 August 2023
(This article belongs to the Section Gut Microbiota)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Clostridioides difficile is considered a nosocomial pathogen that flares up in patients exposed to antibiotic treatment. However, four out of ten patients diagnosed with C. difficile infection (CDI) acquired the infection from non-hospitalized individuals, many of whom have not been treated with antibiotics. Treatment of recurrent CDI (rCDI) with antibiotics, especially vancomycin (VAN) and metronidazole (MNZ), increases the risk of experiencing a relapse by as much as 70%. Fidaxomicin, on the other hand, proved more effective than VAN and MNZ by preventing the initial transcription of RNA toxin genes. Alternative forms of treatment include quorum quenching (QQ) that blocks toxin synthesis, binding of small anion molecules such as tolevamer to toxins, monoclonal antibodies, such as bezlotoxumab and actoxumab, bacteriophage therapy, probiotics, and fecal microbial transplants (FMTs). This review summarizes factors that affect the colonization of C. difficile and the pathogenicity of toxins TcdA and TcdB. The different approaches experimented with in the destruction of C. difficile and treatment of CDI are evaluated.
1. Introduction
Clostridioides difficile is a Gram-positive, sporulating, rod-shaped cell, and obligatory anaerobic [1]. Although C. difficile is considered a nosocomial pathogen that flairs up in patients exposed to antibiotic treatment [2,3], four out of ten patients diagnosed with Clostridioidis difficile infection (CDI) acquired the infection from non-hospitalized individuals [4], many of whom have not been treated with antibiotics [5]. Contracting C. difficile may also be through contact with infected animals, including reptiles and birds [6,7]. One to three percent of adults are asymptomatic carriers of C. difficile [8].
Approximately half a million people in the USA are hospitalized with CDI annually, and 5 to 6% die within the first month of diagnosis [9]. According to Mada and Alam [9], antibiotic use remains the leading risk factor for C. difficile infection. Of these, penicillins, cephalosporins, fluoroquinolones, and clindamycin have been implicated as the most possible cause [9]. Other risk factors associated with CDI include advanced age, chemotherapy, use of proton pump inhibitors, chronic renal disease, chronic liver disease, and malnutrition [9]. Based on the latest report from the Center for Disease Control (CDC), recurrent C. difficile infection (RCDI) was reported in 12.0% of the 4301 cases studied, with a sharp increase in 2020 during the onset of the COVID-19 pandemic [10]. An earlier report by Miranda-Katz et al. [11] stated that only 17 to 24 in 100,000 children develop CDI, which is eightfold lower than that reported in adults over the age of 65. The resistance of infants and young children to CDI may be ascribed to the immunoglobulin in breast milk that inhibits the binding of TcdA to intestinal receptors, along with the lack of intestinal receptors in newborns that recognize the toxin [12,13]. With aging, changes in diet and the secretion of bile acids render intestinal cells more susceptible to C. difficile. Drastic changes in the gut microbiota, as observed with prolonged antibiotic treatment, prevent the conversion of primary bile acids to secondary bile acids. This favors C. difficile colonization [14,15]. Biofilm formation protects cells from oxygen and antibiotics, including metronidazole (MNZ) and vancomycin (VAN), which are commonly used to treat CDI [16,17].
Treatment of recurrent CDI (rCDI) with antibiotics, especially vancomycin (VAN) and metronidazole (MNZ), increases the risk of experiencing a relapse by as much as 70% [16,17]. Strains becoming resistant to both antibiotics are on the increase. Fidaxomicin, which prevents the initial transcription of RNA toxin genes, proved more effective than VAN and MNZ. The efficacy of antibiotics is, however, hampered by their poor ability to penetrate biofilms. More research is required on alternatives to antibiotics, such as non-antimicrobial agents sequestering or inactivating toxin production, quenching of genes (quorum quenching, QQ), immunization, and bacteriotherapy, including fecal microbial transplants (FMTs).
The first part of this paper summarizes the factors affecting the colonization of C. difficile in the gastrointestinal tract (GIT), toxin production, and the pathogenicity of toxins TcdA and TcdB. The advantages and disadvantages of antibiotics are discussed, and results obtained with fidaxomicin are compared to treatment with MNZ and VAN. Alternatives to antibiotics, such as non-antimicrobial agents sequestering or inactivating toxin production, quenching of genes (quorum quenching, QQ), immunization, bacteriotherapy, and microbiome replacement therapies are reviewed.
2. Colonization of C. difficile to Human Intestinal Cells
CDI is contracted through the ingestion of endospores [18]. Germination of spores is controlled by the concentration and type of primary bile salts in the upper part of the GIT [19,20]. Chenodeoxycholate (CDCA) represses spore germination, whereas cholate (CA) induces germination [20]. Most of the primary bile acids (95%), conjugated with taurine and glycine or unconjugated, are absorbed in the terminal ileum and through the hepatic system [19,21]. Primary bile acids that reach the large intestine are converted by gut microbiota into secondary bile acids, for example, ω-muricholate (ωMCA), hyodeoxycholate (HDCA), ursodeoxycholate (UDCA), lithocholate (LCA), and deoxycholate (DCA) [19,22].
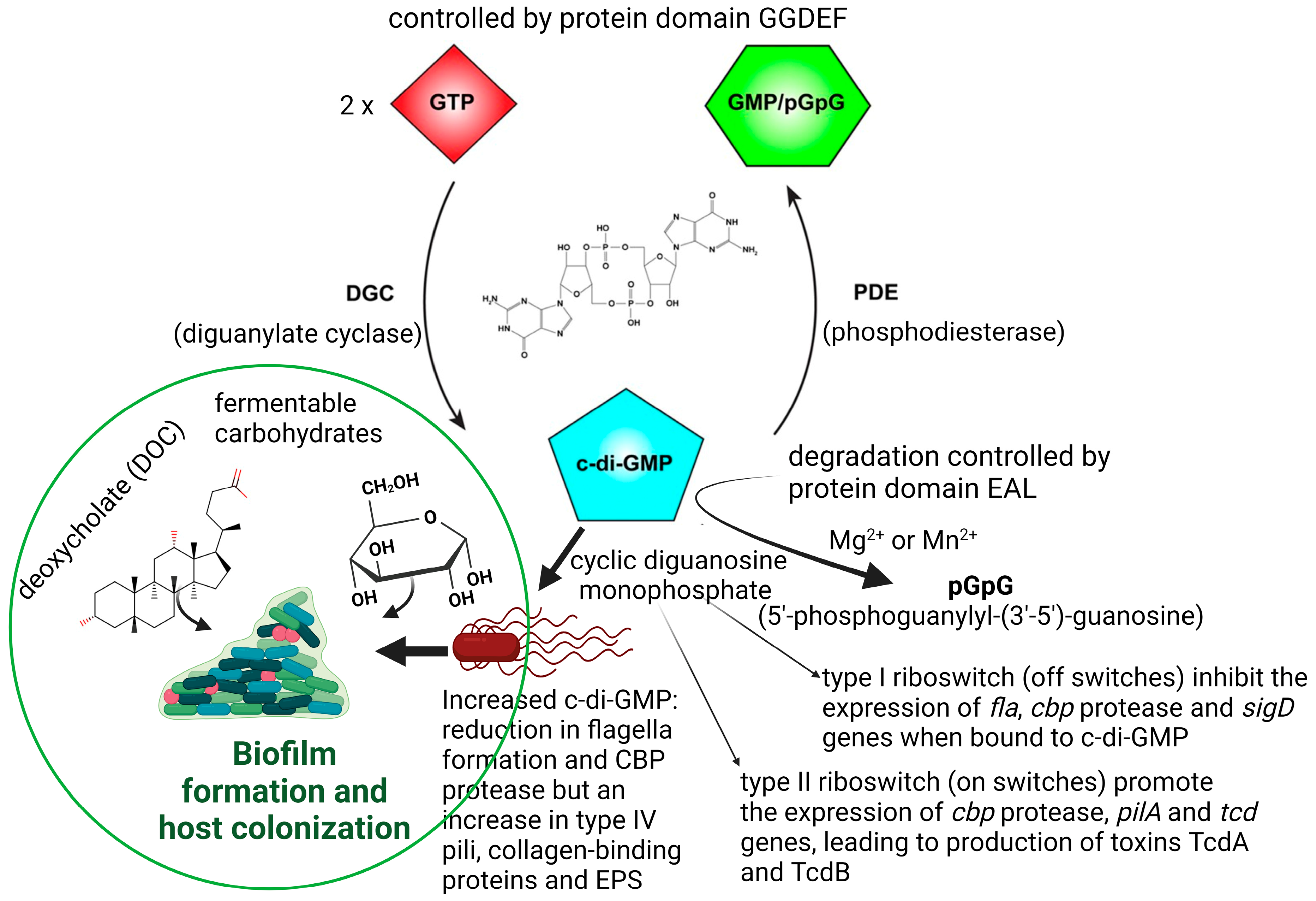
Physiological conditions, such as an excess of fermentable carbohydrates or an increase in deoxycholate (DOC, Figure 1), stimulate C. difficile to form biofilms in the human GIT, which may lead to recurrent episodes of CDI [23,24]. Biofilm formation is also regulated by quorum sensing (QS) signals such as cyclic diguanosine monophosphate (c-di-GMP; Figure 1).
Increased production of c-di-GMP represses motility and stimulates biofilm formation [25]. The synthesis of c-di-GMP is controlled by the protein domain GGDEF, which is widely present in free-living bacteria [26,27,28,29]. Increased c-di-GMP levels reduce the expression of tcdA, tcdB, and tcdR [25,26,27,28,29,30]. The tcdR gene encodes an alternative sigma factor SigD (FliA; σ28) that activates the expression of tcdA and tcdB in response to c-di-GMP [25,30]. Degradation of c-di-GMP is controlled by the protein domain EAL (Figure 1) [26,27,28,29].
Changes in c-di-GMP levels influence the response of riboswitches, which in turn control the expression of flagellar genes. Biological functions have been assigned to 11 of the 16 riboswitches described for C. difficile [31,32]. Seven of the riboswitches, classified as class I, behave in an “off” position in the presence of high levels of c-di-GMP, that is, they terminate gene transcription. The remaining four functional riboswitches, defined as class II, react as “on” switches and trigger gene expressions. Elevated levels of c-di-GMP prevent the transcription of flagellar genes [33,34] and result in biofilm formation (Figure 1, left panel). Strains with mutations in flagellar genes fliC and fliD produced higher levels of TcdA and TcdB [35]. A decrease in the transcription of sigD, located on operon flgB, represses the expression of genes encoding the synthesis of chemotaxis proteins, cell wall proteins (e.g., collagen-binding protein CbpA), and putative membrane transport proteins [36]. Mutations in sigD and flagellar genes fliF, fliG, and fliM resulted in loss of motility and a significant decrease in the expression of toxin genes [30].
Binding of c-di-GMP to Cdi-2-4, one of the four class II c-di-GMP riboswitches located directly upstream of the type IV pili (T4P) primary locus, upregulates the transcription of TFP (type IV pilus) genes and stimulates the aggregation of C. difficile cells [37]. In the absence of c-di-GMP, Cdi-2-4 induces transcription termination and prevents the expression of TFP genes [38]. A 2.54-fold reduction in the expression of fliC in the hypervirulent C. difficile strain R20291 stimulated flagellin production and biofilm formation on glass beads after 7 days [39,40]. Downregulation of other flagellar biosynthesis genes such as flhA, flbD, flgE, and flgD also resulted in biofilm formation [41].
Our understanding of biofilm formation by C. difficile is far from complete, as stimuli for the aggregation of hypervirulent strains (e.g., strains 630 and R20291) differ [42]. Valiente et al. [43] reported an increase in cell hydrophobicity for strain R20291 that lacked flagellar post-transcriptional modification. Only two of the five mutants studied had reduced motility; however, all five mutants showed an increase in biofilm formation. This led the authors to conclude that biofilm formation by C. difficile is not influenced by motility but by hydrophobicity due to the presence of glycan on the flagella. Dapa et al. [44] suggested that flagella play an important role in the late stages of biofilm formation, as shown with the mutant R20291 fliC ClosTron. Since the pilin pilA1 gene (CD3513) in C. difficile is regulated by c-di-GMP acting on the upstream riboswitch Cdi-2-4 [32,45], pili may be required for initial adhesion to epithelial cells and initiate biofilm formation [34]. Pili does, however, not promote late-stage biofilm formation [39].
The importance of cell structure in biofilm formation cannot be ignored. Poquet et al. [41] showed that cells in biofilms have upregulated phospholipid metabolism, active acyl carrier proteins, and increased fatty acid synthesis. Increased production of fatty acids was also reported for cells of Bacillus subtilis in biofilms [46]. Mutants of C. difficile with a deficient lcpB gene and inability to deposit PSII teichoic acids at the cell surface [41] were elongated, larger in diameter, formed abnormal septa, and grew slower [47,48,49]. Cell wall-binding protein Cwp11 (CD2795), cell surface protein Cwp10 (CD2796), and calcium-binding adhesion protein (CD2797) are regulated by c-di-GMP [32]. Cwp11 is released in the “secretome” during biofilm formation [41,50].
A mutation in the prkC gene of C. difficile 630Δerm resulted in increased biofilm formation after 24 h, but only in the presence of bile salt DOC [51]. The function of prkC in C. difficile remains unknown [51]. Mutation of the dnaK gene of strain 630Δerm resulted in the disruption of DnaK synthesis and thus protein folding but also led to a significant increase in biofilm formation and cell elongation [52]. Similar results were recorded when lexA, which encodes the transcriptional repressor LexA in C. difficile R20291, was disrupted [47,53]. Other genes attributed to C. difficile biofilm formation are spo0A [23,24,54], quorum sensing regulator luxS [44,45], and germination receptor sleC [54]. Inactivation of the chaperones dnaK and hfq changes the cells to become temperature-sensitive and increases biofilm formation [52,55].
Iron plays a key role in the growth of pathogenic bacteria, including C. difficile [56]. Ferrous iron is required for C. difficile to colonize the large bowel [57]. C. difficile regulates iron transport with three membrane-bound ferrous iron transporters (FeoBs), of which FeoB1 is produced in the highest quantity under iron-limiting conditions [58]. Although iron stimulates the growth of C. difficile and renders the species more resistant to MNZ [59], it is not known whether an increase in FeoB1 leads to elevated levels of TcdA and TcdB.
Extracellular DNA (eDNA) is a major component of C. difficile biofilms [44,60]. Hypervirulent C. difficile 027 strains are rich in prophages and mobile genetic elements [61]. DNA released from lysed cells may support biofilm formation, as observed for Staphylococcus aureus and Pseudomonas aeruginosa. In both these species, cell lysis in biofilms is controlled by signals regulating quorum sensing [62,63,64]. In C. difficile biofilms, S-ribosylhomocysteinase (LuxS) induces prophages, which likely contribute to biofilm formation [55]. In LuxS mutants, on the other hand, downregulated prophage loci are conserved among C. difficile strains, specifically region 2 encoding a phiC2-like phi-027 phage [40,61,65].
In a complex system, such as the human GIT, metabolites and enzymes produced by bacteria have a profound influence on the microbial population. Bile salt hydrolase (BSH), for instance, produced by Bacteroides ovatus, inhibited the growth of C. difficile [66]. Bacteroides fragilis inhibited the growth of wild-type strains of C. difficile in biofilms when cultured together [55]. Elevated levels of succinate produced by B. fragilis increased the regulation of succinate metabolism by C. difficile [41,67]. With the upregulation of sucD (succinate-CoA ligase [ADP-forming] subunit alpha), increased expressions of accB (biotin carboxyl carrier protein of acetyl-CoA carboxylase), abfH (4-hydroxybutyrate dehydrogenase), abfT (4-hydroxybutyrate CoA-transferase), abfD (4-hydroxybutyryl-CoA dehydratase/vinylacetyl-CoA-delta-isomerase), and cat1 (catalase-1) were noted [55]. Other genes of C. difficile were downregulated, e.g., bcd2 and idhA, encoding butyryl-CoA dehydrogenase and (r)-2-hydroxyisocaproate dehydrogenase, respectively [55].
When cells of C. difficile deficient in luxS were co-cultured with B. fragilis, biofilm formation by the mutant was much weaker than when the same experiment was performed with the wild-type strain of C. difficile [55]. This indicated that AI2/LuxS is involved in facilitating B. fragilis-induced inhibition of C. difficile. Poquet et al. [41] also reported the downregulation of genes involved in carbohydrate metabolism. Both studies have shown that changes in carbohydrate metabolism favor the growth of B. fragilis at the expense of C. difficile. Thus, repression of carbohydrate metabolism plays a major role in biofilm formation. However, the signaling molecules orchestrating the downregulation of genes involved in key metabolic pathways are unknown. Planktonic cells of B. fragilis and C. difficile have no effect on each other’s growth [55], suggesting that the cells must be in close contact with each other. Biofilm formation by C. difficile is a multifaceted and complex process. For more information on biofilms and hypervirulence, refer to Taggart et al. [42].
3. Intra- and Inter-Cellular Communication
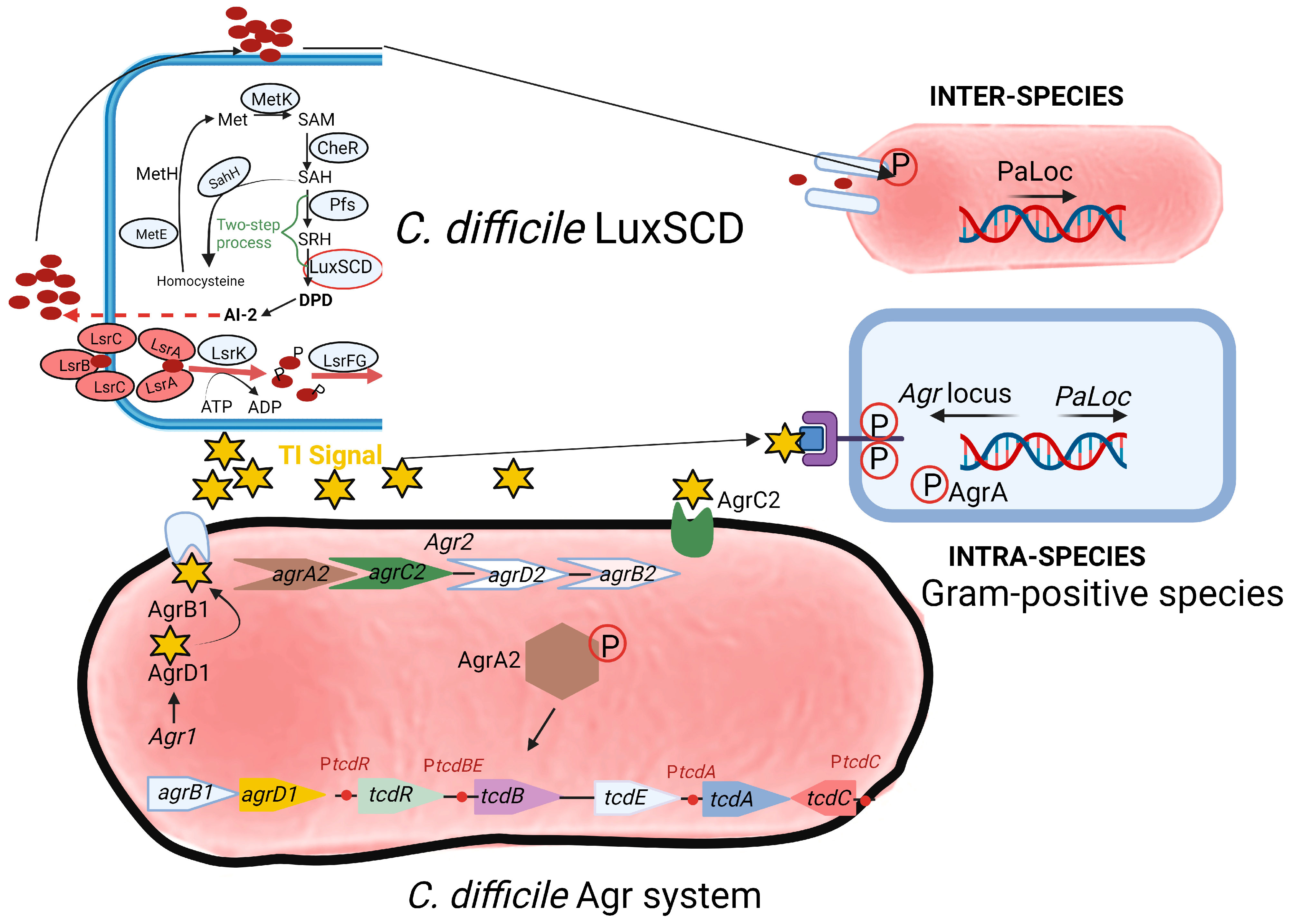
Two communication or quorum sensing (QS) systems have been identified in C. difficile, i.e., an inter-species LuxSCD system (top section of Figure 2) and an intra-species accessory gene regulator (Agr) system (bottom section of Figure 2). Genes encoding homologues of luxS have also been detected in C. difficile [68,69,70,71,72,73]. The luxS gene encodes AI-2 synthase (LuxS). Downstream of luxS are orfX and metH [74]. The function of orfX is unknown. The metH gene encodes 5-methyltetrahydrofolate-homocysteine methyltransferase. AI-2 molecules induce the transcription of the toxin genes tcdA, tcdB, and tcdE during the late exponential growth phase and modulate biofilm formation. Products of rolA and rolB upstream of the luxS gene act as negative regulators of AI-2 [74].
Studies conducted on a mutant of C. difficile with defective luxS have shown that biofilm formation could be restored by supplementing the growth medium with 4,5-dihydroxy-2, 3-pentanedione (DPD), the precursor of AI-2 [55]. As little as 100 nM of DPD was sufficient to restore biofilm formation. This indicates that AI-2 may be involved in signaling among C. difficile biofilm cells. Interestingly, no significant differences in biofilm formation or luxS expression were observed in C. difficile strains isolated from patients with recurrent and non-recurrent CDIs [75]. This suggests that other unknown regulatory systems or QS signals are involved in the colonization of C. difficile. Strains from recurrent CDI sporulated more [75]. This observation needs to be studied in more depth, as the genes involved in sporulation may influence colonization and biofilm formation.
The cell surface receptors for C. difficile have not yet been identified, and the LuxS/AI-2 mechanism, especially within biofilm communities, is unknown. A mutant defective in LuxS (strain R20291 luxS ClosTron mutant) did not produce AI-2 and could not form a biofilm in vitro [55]. RNA sequencing of genes expressed by R20291 luxS mutant cells in biofilms showed an increase in the expression of CDR20291_2554 (crr), a phosphotransferase (PTS) glucose-specific transporter subunit IIA; CDR20291_2927, a cellobiose phosphate-degrading protein; and CDR20291_2930 (treA), a trehalose-6-phosphate hydrolase [55]. An increase in the degradation of trehalose and the functioning of PTS (genes encoding these are on the same operon) provides C. difficile a competitive advantage over gut microbiota. Trehalose acts as an osmoprotectant [76] and prevents protein (and thus toxin) re-conformation during dehydration. Thus, it is possible that trehalose plays an important role in the formation of luxS-mediated C. difficile biofilms, similar to what has been reported for Candida albicans [77].
4. Toxin Production
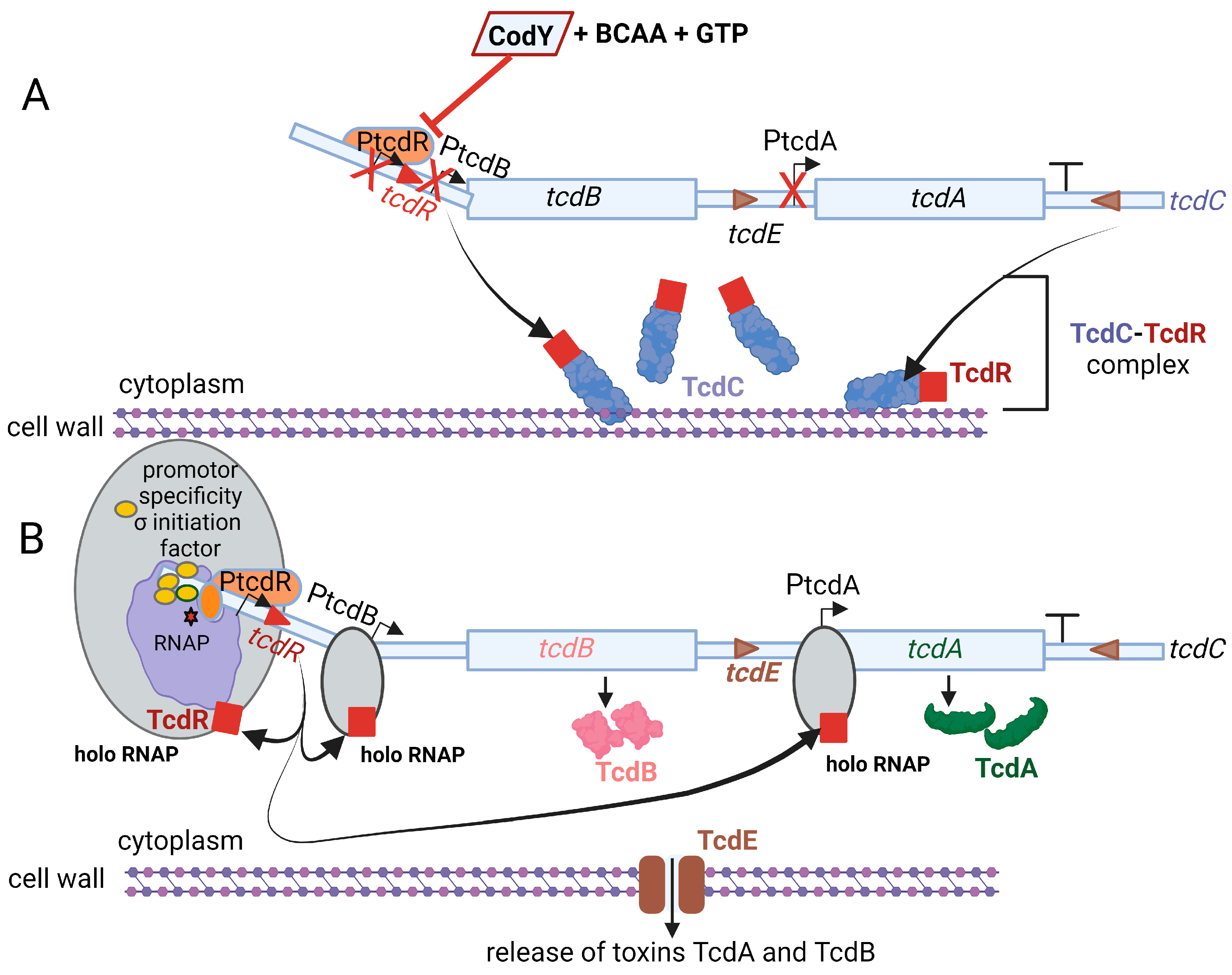
Toxin TcdA, classified as an enterotoxin (2710 amino acids; 308 kDa), causes accumulation of fluid in the ileum [78]. Toxin TcdB, a cytotoxin consisting of 2 366 amino acids (270 kDa), is 100- to 1000-fold more potent than TcdA [79]. Genes encoding the two toxins, tcdA and tcdB, are located on a 19.6 kb pathogenicity locus, PaLoc, together with regulatory genes tcdR, tcdC, and the toxin secretion gene tcdE, as shown in Figure 3 [80,81]. The tcdR gene encodes a sigma factor that controls the transcription of the toxin promoters and its own promoter [82,83]. The tcdC gene encodes an anti-sigma factor, TcdC, that deregulates toxin synthesis [84,85]. Neither the deletion of tcdC nor the altering of tcdC frameshift mutations influenced toxin synthesis [86,87]. Regulation of toxin synthesis is thus more complex and includes the involvement of other key regulatory elements. Darkoh et al. [88] have shown that toxin synthesis is regulated by an Agr QS system. sigD upregulates tcdR [25,30]. The expression of sigD is repressed by elevated levels of c-di-GMP, which in turn downregulate the expression of fliC and toxin production [25].
Pathogenic strains of C. difficile, such as R20291, contain two accessory gene regulator (Agr) loci, Agr1 and Agr2 (Figure 2). The Agr1 locus contains agrB1 (encoding a transmembrane protein AgrB1) and agrD1, encoding a prepeptide AgrD1, which produces an auto-inducer peptide (AIP) called thiolactone or “TI signal.” The latter acts as a signaling molecule to induce C. difficile toxin production once it reaches a sufficient concentration. Treatment of the prepeptide with hydroxylamine disrupted the thioester bonds [89,90,91] and resulted in the loss of activity, indicating that TI contains cysteine residues. According to Darkoh et al. [92], TI is less than 1000 Da in size and is constitutively produced in the GIT by hypervirulent strains of C. difficile. Deletion of the Agr1 locus resulted in the loss of toxin production in both the hypervirulent strain R20291 and non-hypervirulent strain 630 [93]. No significant mRNA transcripts from genes tcdA and tcdB were detected in any of the two strains. However, gene transcripts were detected in the R20291 Agr2 mutant and the wild-type strains R20291 and 630. No toxin activity was detected in cell-free supernatants collected from Agr1 mutants when tested in fibroblast cells. Based on these findings, the Agr1 locus appears to play a central role in toxin production. This finding was confirmed in vivo. The mutant strain of C. difficile (R20291 Agr1 mutant) colonized mice but did not develop CDI. The Agr2 locus contains quorum signal generation (agrB2D2) and response (agrC2A2) genes. Non-hypervirulent strains such as 630 contain only the Agr1 locus. At a certain threshold, TI peptides interact with the two-component AgrC2 histidine kinase, catalyzing the phosphorylation and dimerization of the response regulator AgrA2. The latter induces toxin production, either directly or indirectly. By altering the sensitivity of AgrA2 to TI, toxin production may be prevented. Whether this approach to CDI treatment is feasible remains uncertain. Strains of S. aureus lacking the agr gene (Δagr) are more prone to cause chronic infections and bacteremia. Thus, the treatment of S. aureus infections using the QQ approach is not an option. Part of the agr QS locus in S. aureus, agrDB (agr1) is also present in C. difficile [40,93].
In hypervirulent C. difficile strains belonging to RT (ribotype)-017 and RT-027, QS is dependent on the expression of agrACDB in the Agr2 operon [40,93,94]. This is similar to the agr QS locus agrACDB in S. aureus, which regulates virulence gene expression [94]. The Agr2 QS system in C. difficile uses the cyclic AIP encoded by agrD and exported by the transmembrane protein encoded by agrB [94]. Strains with mutations in AgrB1 and AgrD1 lost the ability to transcribe tcdA and tcdB. The virulence of the AgrB1D1 mutant was also reduced, as measured in a murine model of C. difficile infection [88,93]. Thus, the two-gene Agr1 system is important for C. difficile pathogenesis, but its influence on virulence factor gene expression in the absence of a two-component system is unclear.
A significant increase in agrD1 expression was noted in strains from recurrent CDI compared to non-recurrent CDI [75]. Strains with mutations in agrB1 and agrD1 of strains 630 and R20291 were deficient in both toxin A and B production [93]. A reduction in the expression of tcdA was observed in R20291 ClosTron mutants [94]. No differences in toxin expression (on RNA level) were detected between the R20291 luxS disruption mutant and the wild-type strain, suggesting that the LuxS QS system has little effect on toxin production in C. difficile [55]. An insertion mutation in agrA of strain R20291 also resulted in decreased expression of three genes encoding diguanylate cyclase (DGC) and phosphodiesterase (PDE) responsible for c-di-GMP production [94]. This implies that the Agr QS system is also involved in regulating the production of c-di-GMP [94]. Toxin production may be prevented by blocking AgrB1, eliminating the TI signal with analogs or antibodies, preventing the binding of AgrC2 to the TI signal, or preventing the phosphorylation or dimerization of AgrA2. Growth is not affected by the Agr system, which means that the strains may not become resistant to targeting the quorum signaling mechanism.
The Agr system in C. difficile 630 has multiple functions, as shown by Ahmed et al. [95]. Deletion of agrB1 and agrD1 by a Cas9 nickase system (CRISPR-Cas9n), or deletion of the entire locus, resulted in changes in gene expressions associated with sporulation. At the same time, the motility of C. difficile was reduced when these two genes were disrupted. Loss of AgrB1 resulted in the accumulation of AgrD1, which led to a 15-fold increase in the expression of tcdR, and a 20-fold and 5-fold increase in the expression of tcdA and tcdB, respectively. Deletion of agrB1 and agrD1 or deletion of only agrD1 did not significantly alter the expression of tcdR and tcdB but did result in a minor decrease in tcdA expression. In conclusion, the Agr1 system in C. difficile 630 performs multiple functions and not only AIP signaling. Agr1 influences sporulation efficiency, by requiring a combination of AgrB1 and AgrD1 [95]. Toxin expression is, however, only affected by the absence of AgrB1 and the intracellular accumulation of the AgrD1 peptide. Thus, Agr1 influences C. difficile activity via both AgrB1-dependent and AgrB1-independent mechanisms.
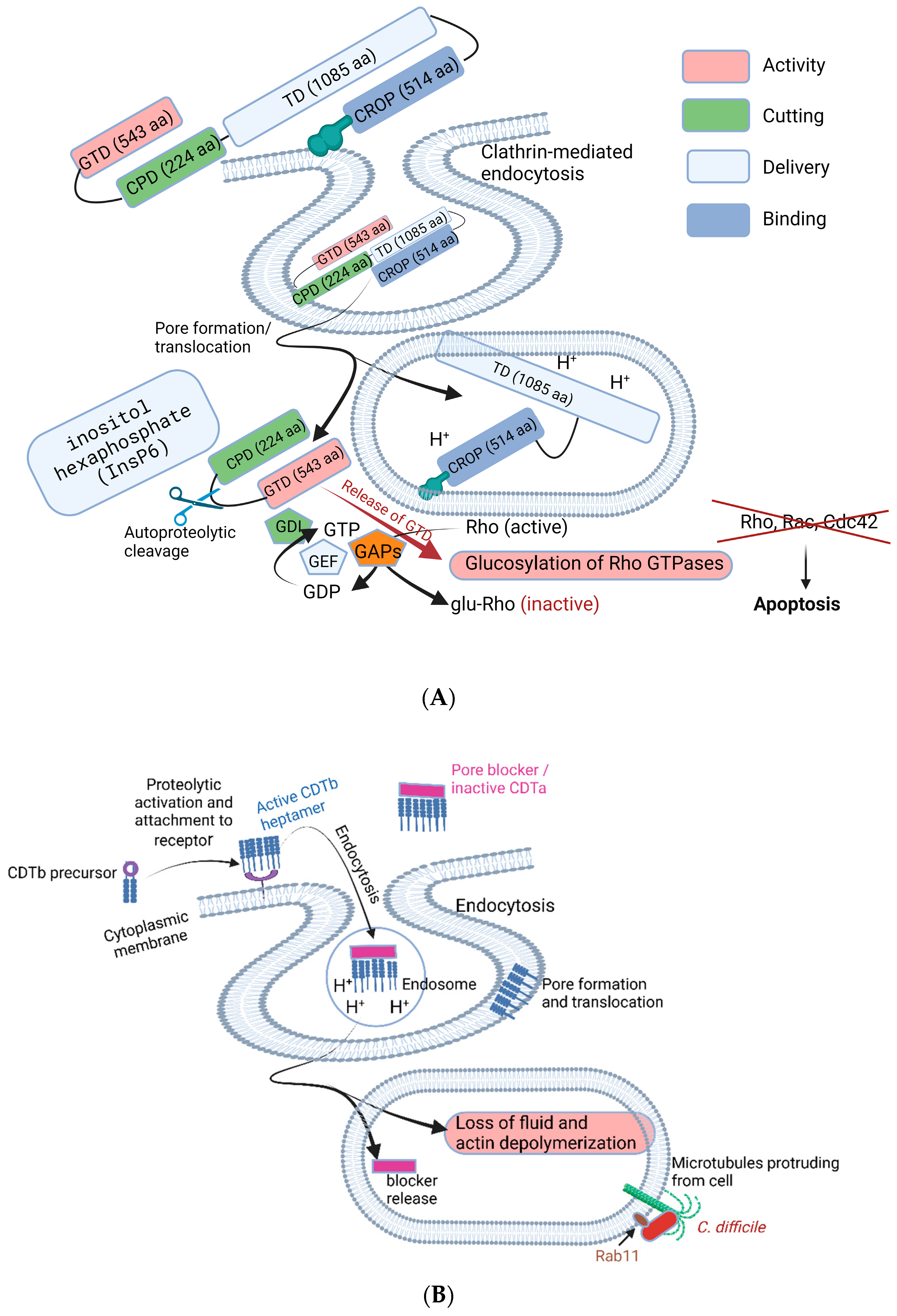
The production of TcdA and TcdB increases when cells in biofilms reach a certain threshold [25]. However, genes encoding the toxins are not triggered at the same rate and seem to be strain-related. The expression of tcdB increased significantly (2.83-fold) in biofilms of C. difficile R20291, in contrast to the expression of tcdA [39]. Toxins released into the colon are taken up by epithelial cells via receptor-mediated endocytosis. This causes mono-glucosylation of low-molecular-weight GTPases in the cytosol [79], resulting in the interruption of Rho GTPases, which leads to apoptosis, cell rounding, dysregulation of the actin cytoskeleton, and changes in cellular signaling [88]. These changes stimulate the release of several immunomodulatory mediators from epithelial cells, phagocytes, and mast cells, resulting in inflammation and the accumulation of neutrophils [88].
Biofilm cells of C. difficile 630Δerm grown in a continuous-flow microfermentor have shown a 1.03-fold decrease in the expression of tcdA but no significant changes in the expression of tcdB [41]. Although results generated by Maldarelli et al. [39] and Poquet et al. [41] made use of different strains and the results are not directly comparable, toxin production is clearly much higher amongst cells in the biofilm. It is interesting to note that the expression of tcdA by C. difficile 630 in human fecal water decreased fourfold [89]. This was ascribed to a decrease in butyrate production [89]. Genes involved in the metabolism of pyruvate, such as bcd2 and idhA, encoding butyryl-CoA dehydrogenase and (r)-2-hydroxyisocaproate dehydrogenase, respectively, were downregulated [55]. Such a shift in metabolism may stimulate the growth of Bacteroides fragilis and outcompete C. difficile. A decrease in toxin production coincided with a 300-fold increase in the expression of sporulation genes [89]. Concluded from these studies, there is no direct correlation between toxin production and endospore formation. This also suggests that pathogenicity, as far as toxin production is concerned, is highly dependent on biofilm formation. Tijerina-Rodrı’guez et al. [90] found no significant difference in the distribution of C. difficile ribotypes between recurrent and non-recurrent CDI cases. However, the authors reported significantly higher sporulation in recurrent CDI samples, especially increased expression of the sporulation genes spo0A and sigH. This implies that recurrent CDIs are more likely to be caused by an increase in the number of endospore-forming biofilm cells. It is, however, uncertain whether sporulation influences biofilm formation, as no significant differences in spore levels were recorded in 7-day-old biofilms of recurrent and non-recurrent CDIs [90]. This raises the question of whether sporulation plays a bigger role than biofilm formation in recurrent CDIs and requires more debate on the conditions required for toxin production.
5. Mode of Action of TcdA and TcdB
Both toxins, TcdA and TcdB, are composed of multimodular structures (a glucosyltransferase domain, GTD; cysteine protease domain, CPD; translocation domain, TD; and combined repetitive oligopeptide, CROP domain), as shown in Figure 4. The CROP domain at the C terminal is also referred to as the receptor-binding domain (RBD). Both toxins inactivate Rho- and Ras-GTPases in the host cytosol via mono-O-glucosylation (Figure 4). This leads to the disruption of tight junctions between intestinal cells, immune modulation [91], and inactivation of enzymes such as phospholipase D [92], protein and lipid kinases, and nicotinamide adenine dinucleotide-oxidase [93,94,95]. Inactivation of Rho proteins also leads to the induction of apoptosis [96], prevention of gene transcription, and the inhibition of phagocytosis [97], G1 cell cycle progression, microtubule dynamics, and vesicular transport pathways [93,94,95]. Notable symptoms caused by TcdA and TcdB are diarrhea, pseudomembranous colitis, and toxic megacolon [98,99].
6. Treatment of CDI with Antibiotics
Treatment of CDI with antibiotics is difficult because of biofilms that protect C. difficile [16,17]. MNZ is administered at 500 mg three times per day, whereas VAN is prescribed at 125 mg four times daily. In more complicated CDI cases (presence of hypotension, ileus, shock, or megacolon), oral VAN with or without intravenous MNZ is recommended [100]. VAN disrupts normal gut flora, whereas fidaxomicin, a narrow-spectrum macrocyclic antibiotic that inhibits the synthesis of RNA polymerase, causes less alteration of gut microbiota [101]. VAN and MNZ have been associated with the colonization of VAN-resistant enterococci (VRE) [102,103]. Recurrent infections occur in approximately 20% to 30% of patients [104,105], with higher CDI recurrence rates observed in patients who have experienced multiple episodes [106] and in subgroups of high-risk patients (oncology, renal impairment, concomitant antibiotics, increased age, and previous CDI episode) [104,107,108,109].
Clinical trials published in 2014 found MNZ inferior to VAN [110]. It is important to note that subinhibitory concentrations of MNZ stimulate C. difficile and increase biofilm formation [111,112]. It is also noteworthy that treatment of recurrent CDI (rCDI) with antibiotics increases the risk of experiencing a relapse by as much as 70% [18,113]. Failure of treatment with MNZ and VAN led to the treatment of CDI with fidaxomicin [114,115]. In 2011, the FDA approved fidaxomicin (previously known as OPT-80, PAR-101, Tiacumicin B and Difimicin) for the treatment of CDI in adults [116], and in 2020, for the treatment of CDI in children 6 months and older [117]. Since the approval of fidaxomicin, it has mainly been prescribed to treat multiple CDI recurrences [118,119]. In 2021, the Infectious Diseases Society of America (IDSA) and the Society for Healthcare Epidemiology of America (SHEA) updated CDI guidelines to encourage the use of fidaxomicin as first-line therapy in adults [120]. Moderate to severe cases of CDI are treated with fidaxomicin for 10 to 14 days [121]. The safety of fidaxomicin and its superiority to VAN was demonstrated in several clinical trials [105,122,123,124,125]. Patients that have been treated with VAN or fidaxomicin and experience their first relapse of CDI may be treated with the same antibiotics. Second and continuing relapses are treated with pulsed or tapered doses of vancomycin [126]. For more information on the resistance of C. difficile to antibiotics, the reader is referred to Dureja et al. [127].
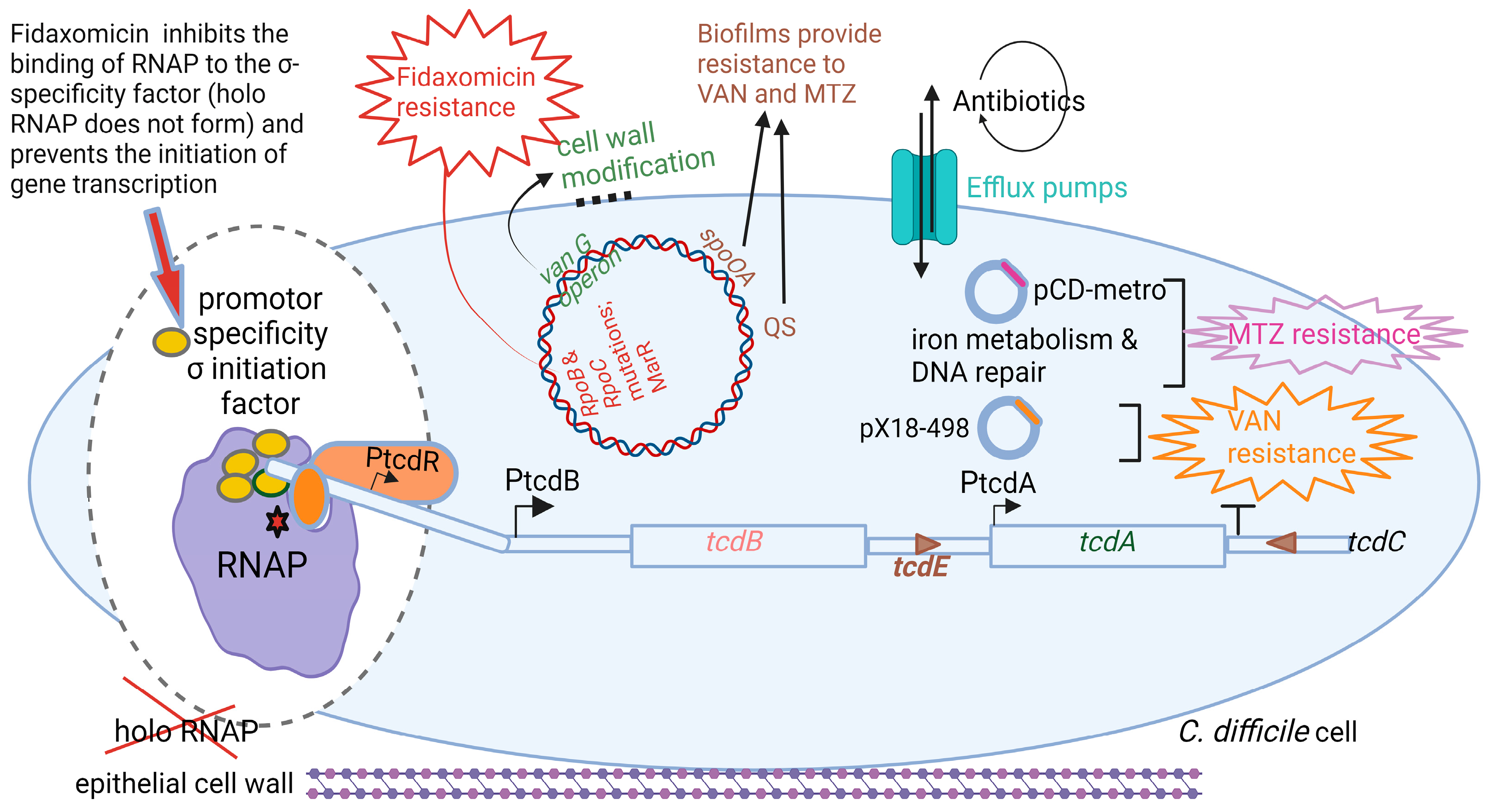
Fidaxomicin is produced by the actinomycete Dactylosporangium aurantiacum subsp. Hamdenesis. It is an unsaturated macrocyclic lactone ring with a hepta-carbohydrate at position 12 and a 6-deoxy sugar at position 21 [100]. Isobutyryl ester at the fourth position is converted by an unknown esterase to produce metabolite OP-1118, which is also active against C. difficile. Fidaxomicin has a narrow spectrum of antibacterial activity and kills strains of C. difficile resistant to cephalosporins, fluoroquinolones, clindamycin, and rifamycin [128]. Isolates of C. difficile resistant to rifamycins or to other antimicrobial classes (cephalosporins, fluoroquinolones, and clindamycin) are not cross-resistant to fidaxomicin [129]. Artsimovitch et al. [115] were the first to show that fidaxomicin inhibits the binding of RNAP to the σ-specificity factor (Figure 5), thus preventing the opening and closing of the DNA:RNA clamp during the initiation of transcription. This means that holo RNAP is not formed. The addition of fidaxomicin after the formation of the transcription-competent open promoter complex did not inhibit transcription during the binding of the promoter to DNA. This clearly indicates that fidaxomicin blocks transcriptional initiation when template DNA strands separate and at the point before RNA synthesis starts. These transcription reaction experiments were performed in vitro in E. coli by adding fidaxomicin at specific time points during DNA translation. This mode of action distinguishes fidaxomicin from elongation inhibitors, such as streptolydigin, and transcription initiation inhibitors, such as myxopyronin and rifamycin [130,131]. Although rifamycins are also RNAP inhibitors, their efficiency and versatility are limited by the rapid increase in drug-resistant strains. This is because they act on the β-subunit, which is relatively dispensable [132,133].
Strains of C. difficile resistant to fidaxomicin had mutations in either the rpoB (Gln1074Lys or Val1143Phe) or rpoC (Asp237Tyr) genes close to the binding site of RNAP [130]. A Val1143Asp mutation resulted in delayed growth [134]. More than 92% of fidaxomicin, orally administered to adults at single doses of 200 and 300 mg, was recovered in feces, and approximately 0.59% in urine, indicating that it is minimally absorbed in the bloodstream. Fidaxomicin also inhibits the growth of methicillin-resistant S. aureus (MRSA), vancomycin-resistant S. aureus (VRSA), and Mycobacterium tuberculosis. However, the low solubility and low systemic bioavailability of fidaxomicin have precluded its use for the treatment of MRSA, VRSA, and tuberculosis.
7. Toxin Binding or Suppression
Targeting the Agr1 quorum signaling system, for example, blocking the synthesis of TI (the toxin-auto-inducing peptide) through AgrB1, sequestering the activity of TI with analogs or antibodies, preventing the binding of a histidine kinase (AgrC2) to TI, and preventing the phosphorylation or dimerization of the response regulator (AgrA2), may sequester toxin-induced damage and inflammation without drastic alterations of the gut microbiome. With a healthy gut microbiome and, hence, a healthy immune system, C. difficile should be naturally cleared from the GIT tract to decrease the risk of CDI relapse. This also lowers the chance of developing resistance, as in the case of antibiotic treatment.
Several steps of QS (Figure 2) may be blocked to prevent toxin production. QQ refers to (i) blocking AIP synthesis, (ii) interfering with QS by using analogs or antibodies, (iii) preventing the binding of AIP to AgrC2, or (iv) blocking the phosphorylation of AgrA2. Thus, quorum quenching can be used to control pathogenesis. To date, no QQ enzymes or QSIs active against C. difficile have been identified. Communication pathways of S. aureus share many similarities with those of C. difficile and may serve as a model in the search against anti-Agr compounds.
The first QQ molecule discovered in bacteria was AiiA, an enzyme from Bacillus species that inactivates acyl homoserine lactone (AH2), the QS signal of Erwinia carotovora (Pectobacterium carotovorum) [135]. Other examples of QQ enzymes are lactonase, acylase, oxidoreductase, and paraoxonase [136]. Ambuic acid, a fungal secondary metabolite, blocks the synthesis of AIPs in several Gram-positive bacteria, including S. aureus and Listeria innocua by inhibiting N-terminal cleavage by AgrB [137]. Apolipoprotein B sequestered QS signals produced by S. aureus [138]. Savirin, Naringenin, and Ω-hydroxy emodin (OHM) repress AgrA activity in S. aureus and inhibit the transcription of virulence factors [139]. Cochinmicin, avellanin, and solonamide B cyclodepsipeptide can also act as competitive inhibitors of the AgrC protein [140].
Anion-binding resins such as X-aptamers (small nucleic acid molecules) with high affinity to the N-terminal glucosyltransferase domain or the C-terminal receptor-binding domain of tcdA and tcdB may sequester or inactivate toxins [92]. Bile salts such as taurocholate, tolevamer, cholestyramines, and colestipol also have toxin-binding properties [92]. Tolevamer is a high-molecular-weight, soluble anionic polymer [141]. Darkoh et al. [92] reported that tolevamer, administered at 6 g per day, was as effective as vancomycin administered at 500 mg per day and was deemed effective in the treatment of mild to moderate C. difficile diarrhea. However, tolevamers are associated with an increased risk of hypokalemia. Nevertheless, patients treated with tolevamer reduced the severity of CDI [141] but were less efficient than vancomycin and metronidazole [110]. Cholestyramine and colestipol also attached to vancomycin and decreased its activity [141].
8. Immunotherapy
Immunization against C. difficile toxins, cell wall structures or outer-membrane proteins offers the prospect of a relatively low-cost approach to CDI prevention. Bezlotoxumab, approved by the FDA, was the first prophylactic antibody used to treat recurrent CDI [92]. The monoclonal antibody targeted TcdB. Actoxumab, a different antibody, targets the C-terminal receptor-binding domain of toxin A [142,143]. Actoxumab used in combination with bezlotoxumab prevented CDI when tested in mice and hamsters [142,144]. A prophylactic concentration of bezlotoxumab, alone or combined with actoxumab, provided 100% protection to piglets [145]. In humans, a combination of actoxumab and bezlotoxumab significantly reduced the rate of CDI relapse [146]. Several combinations of antibodies targeting toxins A and B are in the early stages of development and have been shown to be more effective than actoxumab and bezlotoxumab in the treatment and recurrence of CDI and diarrhea in hamsters [143].
Phase III clinical trials are underway to test multi-dose-inactivated C. difficile toxin-based (toxoid) vaccines. Sanofi Pasteur’s ACAM-CDIFF vaccine contains formalin-inactivated toxoid, and preliminary results from the Phase I trial demonstrated that it was safe and successful in eliciting an adequate neutralizing antibody response [121,147]. Pfizer is currently initiating a large-scale multicenter Phase III trial. Valneva et al. developed a vaccine using a recombinant fusion protein containing cell-binding domains from truncated forms of toxins A and B. It is not clear whether vaccination will be effective for primary or secondary prevention and whether it will prevent or reduce disease severity. Clinical utilization also depends on its efficacy, cost, and safety. Vaccination is unlikely to eliminate colonization; therefore, patient isolation is important for preventing CDI transmission.
Even though toxins A and B are the primary targets of most immunotherapies in development, several other virulence and colonization factors such as flagella, surface-layer proteins, Cpw84 proteins, and pilin are promising avenues for therapeutic intervention. Success in the development of drugs that target virulence and colonization factors may guide the next generation of CDI therapies. Currently, there are no therapies for CDI that are based on the direct inhibition of toxin production, toxin activity, or colonization factors. Bender et al. [148] are actively investigating novel approaches to CDI treatment. As antitoxin antibodies do not prevent C. difficile colonization [146], antibodies targeting cell wall proteins or adherent factors may be an answer. Previous studies have shown a significant decline in C. difficile colonization when mice were immunized with anti-flagellin (FliC) and flagellin filament cap protein (FliD) antibodies [149]. Hamsters orally administered purified FliD-specific antibodies were protected from CDI when challenged with C. difficile strain 630 [150]. Mice rectally vaccinated with FliD and cell wall extracts showed a significant decrease in C. difficile colonization [151]. Concluded from these studies, prevention of colonization may be the answer to the treatment of CDI.
9. Probiotics
Probiotics have been used effectively in the treatment of a variety of diseases, including CDIs, but their efficacy is strain- and disease-specific [152]. Probiotics may produce proteases that degrade TcdA and TcdB and compete with toxins for attachment to the gut wall [153]. Lactobacillus acidophilus GPIB, isolated from swine, reduced C. difficile virulence by decreasing AI-2 activity [154]. Downregulation of virulence genes was also observed. Heat-inactivated cell extracts of L. fermentum Lim2 suppressed LuxS, tcdA, and tcdB of C. difficile 027 [155]. Upregulation of the negative regulator gene (tcdC) was also recorded. Conclusive proof that probiotic strains interfere with the QS system of C. difficile by producing QQ enzymes or QSIs is lacking. The molecular structures and biochemical pathways of these inhibitors may provide insights into the control of CDI and the development of next-generation probiotics. Further research is needed on strains from the phyla Bacteroidetes and Firmicutes, as they have shown promising results. Probiotics have immunoprotective properties, hamper the adherence of C. difficile to the intestinal lumen, and modulate the host’s immune response. Ofosu [156] showed that probiotics increase the intestinal secretion of IgA antitoxin and inhibit the production of IL-8, a proinflammatory cytokine. Lactobacillus and Bifidobacterium strains are the most frequently utilized probiotics, in addition to the yeast Saccharomyces boulardii as adjuvant treatment in CDI or as primary prevention therapy for patients receiving vancomycin [121,157]. S. boulardii may, however, induce fungicemia in immunocompromised patients [121,158,159].
Fecal microbial transplants (FMTs) may be an option [160], as seen in clinical trials having shown more than 85% effectiveness [160,161]. Some evidence suggests that host secretions or microbial metabolites may also be sufficient for treatment of CDI [162,163]. FMT proved effective in preventing recurrences of CDI and treating refractory cases [164]. In one study, 91% of CDI patients who were refractory to antibiotics showed a positive response to FMT [160]. In another study, a cure rate of 75% with a single FMT infusion and 100% with multiple FMT infusions (administered to patients with severe CDI refractory to antibiotics) was reported [165].
A Phase II clinical trial of a non-toxigenic C. difficile strain M3 (NTCD M3) developed by Viropharma, Inc., demonstrated efficacy in reducing CDI recurrence, with possible restoration of the intestinal flora to its normal state [159]. It was reported that 22 weeks after administration, NTCD M3 strains could not be detected in stools. This observation suggests that colonization of NTCD M3 strains may be transient and presumably occurs because of the restoration of the normal microbiota, which may then provide protection against subsequent CDI [159].
10. Bacteriophage Therapy
Bacteriophages isolated from patients with CDI are non-lytic and belong to the Myoviridae (phiC2, phiC5, phiC8) and Siphoviridae (phiC6) subfamilies of Caudovirales [166]. Several C. difficile phages are, however, not behaving in a lysogenic manner and are thus not integrated into the host genome but remain episomal [167]. This is specifically the case with larger phages of approximately 130 kbp (i.e., phiCD5763, phiCD5774, phiCD211), although phiCD38-2 (41 kbp) also exists as a plasmid prophage [168]. Phage phiHN10 binds to S-layer proteins [169]. Interestingly, the cell wall protein CwpV from C. difficile confers resistance to infection by different phages, including members of the Siphoviridae and Myoviridae families [170]. The mode of activity is to prevent phage DNA from entering the cell [170]. Some phages, e.g., phiCDHM1 contain homologs of agr genes [171] that may promote the survival and replication of phiCDHM1 and its host. Phage phiSemix9P1 has a functional binary toxin locus (CdtLoc) [172], suggesting that lysogenic phages may play an important role in the spreading of toxin genes.
Prophylactic treatment with phage phiCD27 reduced C. difficile cell numbers significantly and prevented the production of TcdA and TcdB [173]. Treatment of C. difficile strains CD105LC2 and CD105HE1 with phages showed limited clearing of the cultures in vitro [174]. The most effective phage (phiCDHM2) killed almost all cells of C. difficile within 5 h, but growth recovered 24 h later [174]. A four-phage cocktail (phiCDHM1-phiCDHM2-phiCDHM5-phiCDHM6) proved effective in reducing CD105LC2 biofilms in vitro [175]. Despite reduced colonization, C. difficile was still detectable in the cecum and colon of most animals [174]. This indicated that phage cocktail treatment could delay, but not prevent, CDI.
The use of phage-derived endolysins and phage tail-like particles (PTLPs) against C. difficile has been explored in a few studies [176]. Several strains of C. difficile produce PTLPs when the SOS response is induced, resulting in the killing of competing strains [177]. For further background on PTLPs, the reader is referred to the reviews of Dams et al. [178] and Heuler et al. [179].
An endolysin targeting C. difficile, e.g., phiCD27-derived CD27L, was successfully overexpressed in E. coli and showed activity against all 30 strains, including two strains of the hypervirulent ribotype 027 [180]. Truncation of the endolysin to its N-terminal catalytic domain (CD27L1−179) enhanced lytic activity and broadened the host range [181]. The PlyCD catalytic domain (PlyCD1−174) also showed superior lytic activity. Combined with a vancomycin pre-treatment, PlyCD1−174 significantly reduced the titer of Cd in vitro [182].
Bacteriophage therapy may be a good alternative to the treatment of CDI, as they are highly species- or strain-specific and are therefore far less detrimental to the intestinal microbiota. Further research on endolysins needs to be conducted, as they display high species specificity and may be used in combination with antibiotic treatment. One of the challenges to overcome would be to safeguard phages and endolysins against the destruction by gastric enzymes.
11. Conclusions
The emergence of hypervirulent strains owing to increased antibiotic use is one of the reasons why CDI is considered a high-risk pathogen. Those that are most at risk are individuals suffering from IBD, immunodeficiency, and hypoalbuminemia, who underwent an organ transplant, had malignant tumors, and received chemotherapy. The prevention of biofilm formation and toxin production is probably the most promising alternative treatment for CDI. However, no therapeutic agents are available to inhibit colonization and toxin production or to suppress toxin activity. Antibodies may be the answer, but recipients of such treatments may generate secondary antibodies and run the risk of developing autoimmune diseases. Competitive exclusion of C. difficile and prevention of adhesion to receptors in the GIT is another option worth exploring but requires in-depth knowledge of changes in the gut microbiome of patients with CDI. Although the use of probiotics to relieve CDI has been reported, results have not been conclusive. The use of non-toxigenic C. difficile strains to outcompete toxigenic strains has also been proposed and is currently being evaluated in clinical trials.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author has no conflict of interest.
Abbreviations
CDI, Clostridioides difficile infection; TI, thiolactone peptide; AIP, autoinducer peptide; TcdA and TcdB, toxins A and B, respectively; AIP, auto-inducer peptide; c-di-GMP, cyclic diguanosine monophosphate; CBP, collagen-binding protein; Agr, accessory gene regulator; QS, quorum sensing; QQ, quorum quenching; QSI, quorum sensing inhibitor; LuxS, S-ribosylhomocysteinase; VAN, vancomycin; MNZ, metronidazole; FMT, fecal microbial transplant; CDC, Center for Disease Control; RCDI, recurrent C. difficile infection; gastro-intestinal tract (GIT); CDCA, chenodeoxycholate; CA, cholate; ωMCA, ω-muricholate; HDCA, hyodeoxycholate; UDCA, ursodeoxycholate; LCA, lithocholate; DCA, deoxycholate; sigD, sigma factor; CbpA, collagen-binding protein A; CBPs, collagen-binding proteins; T4P, type IV pili; FeoBs, ferrous iron transporters; eDNA, extracellular DNA; BSH, bile salt hydrolase; DPD, 4,5-dihydroxy-2, 3-pentanedione; PTS, phosphotransferase; PaLoc, pathogenicity locus; GTP, guanosine triphosphate; BCAAs, branched-chain amino acids; RNAP, RNA polymerase; holo RNAP, holozyme RNAP complex; CROP, combined repetitive oligopeptide; RBD, receptor-binding domain; CPD, cysteine protease domain; TD, translocation domain; GTD, glucosyltransferase domain; GDIs, guanine nucleotide dissociation inhibitors; GEFs, guanine nucleotide exchange factors; GAPs, GTPase-activating proteins, VRE, VAN-resistant enterococci, FDA, Food and Drug Administration (USA); IDSA, Infectious Diseases Society of America; SHEA, Society for Healthcare Epidemiology of America; MRSA, methicillin-resistant Staphylococcus aureus; VRSA, vancomycin-resistant S. aureus; OHM, Ω-hydroxyemodin; PTLPs, phage tail-like particles.
References
- Sebaihia, M.; Wren, B.W.; Mullany, P.; Fairweather, N.F.; Minton, N.; Stabler, R.; Thomson, N.R.; Roberts, A.P.; Cerdeño-Tárraga, A.M.; Wang, H.; et al. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat. Genet. 2006, 38, 779–786. [Google Scholar] [CrossRef]
- Merrigan, M.; Venugopal, A.; Mallozzi, M.; Roxas, B.; Viswanathan, V.K.; Johnson, S.; Gerding, D.N.; Vedantam, G. Human hypervirulent Clostridium difficile strains exhibit increased sporulation as well as robust toxin production. J. Bacteriol. 2010, 192, 4904–4911. [Google Scholar] [CrossRef] [PubMed]
- Aliente, E.; Dawson, L.F.; Cairns, M.D.; Stabler, R.A.; Wren, B.W. Emergence of new PCR ribotypes from the hypervirulent Clostridium difficile 027 lineage. J. Med. Microbiol. 2012, 61, 49–56. [Google Scholar] [CrossRef]
- Lessa, F.C.; Winston, L.G.; McDonald, L.C. Emerging Infections Program C. difficile Surveillance Team. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 2369–2370. [Google Scholar] [CrossRef] [PubMed]
- Chitnis, A.S.; Holzbauer, S.M.; Belflower, R.M.; Winston, L.G.; Bamberg, W.M.; Lyons, C.; Farley, M.M.; Dumyati, G.K.; Wilson, L.E.; Beldavs, Z.G.; et al. Epidemiology of community-associated Clostridium difficile infection, 2009 through 2011. JAMA Intern. Med. 2013, 173, 1359–1367. [Google Scholar] [CrossRef]
- Gould, L.H.; Limbago, B. Clostridium difficile in food and domestic animals: A new foodborne pathogen. Clin. Infect. Dis. 2010, 51, 577–582. [Google Scholar] [CrossRef]
- Hensgens, M.P.; Keessen, E.C.; Squire, M.M.; Riley, T.V.; Koene, M.G.; de Boer, E.; Lipman, L.J.; Kuijper, E.J. Clostridium difficile infection in the community: A zoonotic disease? Clin. Microbiol. Infect. 2012, 18, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Vedantam, G.; Clark, A.; Chu, M.; McQuade, R.; Mallozzi, M.; Viswanathan, V.K. Clostridium difficile infection: Toxins and non-toxin virulence factors, and their contributions to disease establishment and host response. Gut Microbes. 2012, 3, 121–134. [Google Scholar] [CrossRef]
- Mada, P.K.; Alam, M.U. Clostridioides difficile Infection. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK431054/ (accessed on 23 January 2023).
- Okafor, C.M.; Clogher, P.; Olson, D.; Niccolai, L.; Hadler, J. Trends in and risk factors for recurrent Clostridioides difficile infection, New Haven County, Connecticut, USA, 2015–2020. Emerg. Infect. Dis. 2023, 29, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Katz, M.; Parmar, D.; Dang, R.; Alabaster, A.; Greenhow, T.L. Epidemiology and risk factors for community associated Clostridioides difficile in children. J. Pediatr. 2020, 221, 99–106. [Google Scholar] [CrossRef]
- Al-Jumaili, I.J.; Shibley, M.; Lishman, A.H.; Record, C.O. Incidence and origin of Clostridium difficile in neonates. J. Clin. Microbiol. 1984, 19, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Eglow, R.; Pothoulakis, C.; Itzkowitz, S.; Israel, E.J.; O’Keane, C.J.; Gong, D.; Gao, N.; Xu, Y.L.; Walker, W.A.; LaMont, J.T. Diminished Clostridium difficile toxin A sensitivity in newborn rabbit ileum is associated with decreased toxin A receptor. J. Clin. Investig. 1992, 90, 822–829. [Google Scholar] [CrossRef]
- Bassis, C.M.; Theriot, C.M.; Young, V.B. Alteration of the murine gastrointestinal microbiota by tigecycline leads to increased susceptibility to Clostridium difficile infection. Antimicrob. Agents Chemother. 2014, 58, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Theriot, C.M.; Bowman, A.A.; Young, V.B. Antibiotic-induced alterations of the gut microbiota alter secondary bile acid production and allow for Clostridium difficile spore germination and outgrowth in the large intestine. Host-Microbe Biol. 2016, 1, e00045-15. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. The efficacy of thuricin CD, tigecycline, vancomycin, teicoplanin, rifampicin and nitazoxanide, independently and in paired combinations against Clostridium difficile biofilms and planktonic cells. Gut Pathog. 2016, 8, 20. [Google Scholar] [CrossRef]
- James, G.A.; Chesnel, L.; Boegli, L.; Pulcini, E.D.; Fisher, S.; Stewart, P.S. Analysis of Clostridium difficile biofilms: Imaging and antimicrobial treatment. J. Antimicrob. Chemother. 2017, 73, 102–108. [Google Scholar] [CrossRef]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium difficile infection. Nat. Rev. Dis. Primers 2016, 2, 16020. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Bile salts and glycine as cogerminants for Clostridium difficile spores. J. Bacteriol. 2008, 190, 2505–2512. [Google Scholar] [CrossRef]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef]
- Kuipers, F.; Bloks, V.W.; Groen, A.K. Beyond intestinal soap–Bile acids in metabolic control. Nat. Rev. Endocrinol. 2014, 10, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.F.; Valiente, E.; Faulds-Pain, A.; Donahue, E.H.; Wren, B.W. Characterisation of Clostridium difficile biofilm formation, a role for Spo0A. PLoS ONE 2012, 7, e50527. [Google Scholar] [CrossRef] [PubMed]
- Dubois, T.; Tremblay, Y.D.N.; Hamiot, A.; Martin-Verstraete, I.; Deschamps, J.; Monot, M.; Briandet, R.; Dupuy, B. A microbiota generated bile salt induces biofilm formation in Clostridium difficile. NPJ Biofilms Microbiomes 2019, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.W.; Mangalea, M.R.; Purcell, E.B.; Borchardt, E.K.; Tamayo, R. The second messenger cyclic Di-GMP regulates Clostridium difficile toxin production by controlling expression of sigD. J. Bacteriol. 2013, 195, 5174–5185. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.; Weiser, S.; Amiot, N.C.; Chan, C.; Schirmer, T.; Giese, B.; Jenal, U. Cell cycle-dependent dynamic localization of a bacterial response regulator with a novel di-guanylate cyclase output domain. Genes Dev. 2004, 18, 715–727. [Google Scholar] [CrossRef]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef]
- Chan, C.; Paul, R.; Samoray, D.; Amiot, N.C.; Giese, B.; Jenal, U.; Schirmer, T. Structural basis of activity and allosteric control of diguanylate cyclase. Proc. Natl. Acad. Sci. USA 2004, 101, 17084–17089. [Google Scholar] [CrossRef]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J. Biol. Chem. 2005, 280, 30829–30837. [Google Scholar] [CrossRef]
- Aubry, A.; Hussack, G.; Chen, W.; KuoLee, R.; Twine, S.M.; Fulton, K.M.; Foote, S.; Carrillo, C.D.; Tanha, J.; Logan, S.M. Modulation of toxin production by the flagellar regulon in Clostridium difficile. Infect. Immun. 2012, 80, 3521–3532. [Google Scholar] [CrossRef]
- Lee, E.R.; Baker, J.L.; Weinberg, Z.; Sudarsan, N.; Breaker, R.R. An allosteric self-splicing ribozyme triggered by a bacterial second messenger. Science 2010, 329, 845–848. [Google Scholar] [CrossRef]
- McKee, R.W.; Aleksanyan, N.; Garrett, E.M.; Tamayo, R. Type IV pili promote Clostridium difficile adherence and persistence in a mouse model infection. Infect. Immun. 2018, 86, e00943-17. [Google Scholar] [CrossRef]
- Purcell, E.B.; McKee, R.W.; McBride, S.M.; Waters, C.M.; Tamayo, R. Cyclic diguanylate inversely regulates motility and aggregation in Clostridium difficile. J. Bacteriol. 2012, 194, 3307–3316. [Google Scholar] [CrossRef]
- McKee, R.W.; Harvest, C.K.; Tamayo, R. Cyclic diguanylate regulates virulence factor genes via multiple riboswitches in Clostridium difficile. mSphere 2018, 3, e00423-18. [Google Scholar] [CrossRef]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic analysis of the Clostridium difficile flagellar proteins, FliC and FliD, and their contribution to virulence in hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef]
- Tulli, L.; Marchi, S.; Petracca, R.; Shaw, H.A.; Fairweather, N.F.; Scarselli, M.; Soriani, M.; Leuzzi, R. CbpA: A novel surface exposed adhesin of Clostridium difficile targeting human collagen. Cell Microbiol. 2013, 15, 1674–1687. [Google Scholar] [PubMed]
- Bordeleau, E.; Purcell, E.B.; Lafontaine, D.A.; Fortier, L.C.; Tamayo, R.; Burrus, V. Cyclic di-GMP riboswitch-regulated type IV pili contribute to aggregation of Clostridium difficile. J. Bacteriol. 2015, 197, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.F.; Peltier, J.; Hall, C.L.; Harrison, M.A.; Derakhshan, M.; Shaw, H.A.; Fairweather, N.F.; Wren, B.W. Extracellular DNA, cell surface proteins and c-di-GMP promote biofilm formation in Clostridioides difficile. Sci. Rep. 2021, 5, 3244. [Google Scholar] [CrossRef] [PubMed]
- Maldarelli, G.A.; Piepenbrink, K.H.; Scott, A.J.; Freiberg, J.A.; Song, Y.; Achermann, Y.; Ernst, R.K.; Shirtliff, M.E.; Sundberg, E.J.; Donnenberg, M.S.; et al. Type IV pili promote early biofilm formation by Clostridium difficile. Pathog. Dis. 2016, 74, ftw061. [Google Scholar] [CrossRef] [PubMed]
- Stabler, R.A.; He, M.; Dawson, L.; Martin, M.; Valiente, E.; Corton, C.; Lawley, T.D.; Sebaihia, M.; Quail, M.A.; Rose, G.; et al. Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium. Genome Biol. 2009, 10, R102. [Google Scholar] [CrossRef]
- Poquet, I.; Saujet, L.; Canette, A.; Monot, M.; Mihajlovic, J.; Ghigo, J.-M.; Soutourina, O.; Briandet, R.; Martin-Verstraete, I.; Dupuy, B. Clostridium difficile biofilm: Remodeling metabolism and cell surface to build a sparse and heterogeneously aggregated architecture. Front. Microbiol. 2018, 9, 2084. [Google Scholar] [CrossRef] [PubMed]
- Taggart, M.G.; Snelling, W.J.; Naughton, P.J.; La Ragione, R.M.; Dooley, J.S.G.; Ternan, N.G. Biofilm regulation in Clostridioides difficile: Novel systems linked to hypervirulence. PLoS Pathog. 2021, 17, e1009817. [Google Scholar] [CrossRef] [PubMed]
- Valiente, E.; Bouché, L.; Hitchen, P.; Faulds-Pain, A.; Songane, M.; Dawson, L.F.; Donahue, E.; Stabler, R.A.; Panico, M.; Morris, H.R.; et al. Role of glycosyltransferases modifying type B flagellin of emerging hypervirulent Clostridium difficile lineages and their impact on motility and biofilm formation. J. Biol. Chem. 2016, 291, 25450–25461. [Google Scholar] [CrossRef] [PubMed]
- Ðapa, T.; Leuzzi, R.; Ng, Y.K.; Baban, S.T.; Adamo, R.; Kuehne, S.A.; Scarselli, M.; Minton, N.P.; Serruto, D.; Unnikrishnan, M. Multiple factors modulate biofilm formation by the anaerobic pathogen Clostridium difficile. J. Bacteriol. 2013, 195, 545–555. [Google Scholar] [PubMed]
- Purcell, E.B.; McKee, R.W.; Bordeleau, E.; Burrus, V.; Tamayo, R. Regulation of type IV pili contributes to surface behaviors of historical and epidemic strains of Clostridium difficile. J. Bacteriol. 2016, 198, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Pedrido, M.E.; de Oña, P.; Ramirez, W.; Leñini, C.; Goñi, A.; Grau, R. Spo0A links de novo fatty acid synthesis to sporulation and biofilm development in Bacillus subtilis. Mol. Microbiol. 2013, 87, 348–367. [Google Scholar] [CrossRef]
- Walter, B.M.; Cartman, S.T.; Minton, N.P.; Butala, M.; Rupnik, M. The SOS response master regulator LexA is associated with sporulation, motility and biofilm formation in Clostridium difficile. PLoS ONE 2015, 10, e0144763. [Google Scholar] [CrossRef]
- Chu, M.; Mallozzi, M.J.; Roxas, B.P.; Bertolo, L.; Monteiro, M.A.; Agellon, A.; Viswanathan, V.K.; Vedantam, G. A Clostridium difficile cell wall glycopolymer locus influences bacterial shape, polysaccharide production and virulence. PLoS Pathog. 2016, 12, e1005946. [Google Scholar] [CrossRef]
- Jain, S.; Smyth, D.; O’Hagan, B.M.G.; Heap, J.T.; McMullan, G.; Minton, N.P.; Ternan, N.G. Inactivation of the dnaK gene in Clostridium difficile 630 Δerm yields a temperature-sensitive phenotype and increases biofilm forming ability. Sci. Rep. 2017, 7, 17522. [Google Scholar] [CrossRef]
- Pantaléon, V.; Soavelomandroso, A.P.; Bouttier, S.; Briandet, R.; Roxas, B.; Chu, M.; Collignon, A.; Janoir, C.; Vedantam, G.; Candela, T. The Clostridium difficile protease Cwp84 modulates both biofilm formation and cell-surface properties. PLoS ONE 2015, 10, e0124971. [Google Scholar] [CrossRef] [PubMed]
- Cuenot, E.; Garcia-Garcia, T.; Douche, T.; Gorgette, O.; Courtin, P.; Denis-Quanquin, S.; Hoys, S.; Tremblay, Y.D.N.; Matondo, M.; Chapot-Chartier, M.-P.; et al. The Ser/Thr kinase PrkC participates in cell wall homeostasis and antimicrobial resistance in Clostridium difficile. Infect. Immun. 2019, 87, e00005-19. [Google Scholar] [CrossRef]
- Jain, S.; Graham, C.; Graham, R.L.J.; McMullan, G.; Ternan, N.G. Quantitative proteomic analysis of the heat stress response in Clostridium difficile strain 630. J. Proteome Res. 2011, 10, 3880–3890. [Google Scholar] [CrossRef]
- Johnston, J.L.; Sloan, J.; Fyfe, J.A.M.; Davies, J.K.; Rood, J.I. The recA gene from Clostridium perfringens is induced by methyl methanesulphonate and contains an upstream Cheo box. Microbiology 1997, 143, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Dapa, T.; Unnikrishnan, M. Biofilm formation by Clostridium difficile. Gut Microbes. 2013, 4, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Slater, R.T.; Frost, L.R.; Jossi, S.E.; Millard, A.D.; Unnikrishnan, M. Clostridioides difficile LuxS mediates inter-bacterial interactions within biofilms. Sci. Rep. 2019, 9, 9903. [Google Scholar] [CrossRef] [PubMed]
- Berges, M.; Michel, A.-M.; Lassek, C.; Nuss, A.M.; Beckstette, M.; Dersch, P.; Riedel, K.; Sievers, S.; Becher, D.; Otto, A.; et al. Iron Regulation in Clostridioides difficile. Front. Microbiol. 2018, 9, 3183. [Google Scholar] [CrossRef]
- Deshpande, A.; Olaitan, A.O.; Mckelvey, A.M.; Rutherford, T.J.T.; Hurdle, J.G. The ferrous iron transporter FeoB1 is essential for Clostridioides difficile toxin production and Pathogenesis in Mice. bioRxiv. 2022. [Google Scholar] [CrossRef]
- Ho, T.D.; Ellermeier, C.D. Ferric uptake regulator for control of putative iron acquisition systems in Clostridium difficile. J. Bacteriol. 2015, 197, 2930–2940. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, J.; Chawla, S.; Tong, S.; Lozada, K.A.; Yang, S. Iron effects on Clostridioides difficile toxin production and antimicrobial susceptibilities. Antibiotics 2022, 11, 537. [Google Scholar] [CrossRef]
- Semenyuk, E.G.; Laning, M.L.; Foley, J.; Johnston, P.F.; Knight, K.L.; Gerding, D.N.; Driks, A. Spore formation and toxin production in Clostridium difficile biofilms. PLoS ONE 2014, 9, e87757. [Google Scholar] [CrossRef]
- Nale, J.Y.; Shan, J.; Hickenbotham, P.T.; Fawley, W.N.; Wilcox, M.H.; Clokie, M.R. Diverse temperate bacteriophage carriage in Clostridium difficile 027 strains. PLoS ONE 2012, 7, e37263. [Google Scholar] [CrossRef]
- Ibanez de Aldecoa, A.L.; Zafra, O.; Gonzalez-Pastor, J.E. Mechanisms and regulation of extracellular DNA release and its biological roles in microbial communities. Front. Microbiol. 2017, 8, 1390. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Breyne, K.; De Rycke, R.; Vermote, A.; Van Nieuwerburgh, F.; Meyer, E.; Van Calenbergh, S.; Coenye, T. The quorum sensing inhibitor hamamelitannin increases antibiotic susceptibility of Staphylococcus aureus biofilms by affecting peptidoglycan biosynthesis and eDNA release. Sci. Rep. 2016, 6, 20321. [Google Scholar] [CrossRef]
- Svensson, S.L.; Pryjma, M.; Gaynor, E.C. Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni. PLoS ONE 2014, 9, e106063. [Google Scholar] [CrossRef]
- Sekulovic, O.; Fortier, L.C. Global transcriptional response of Clostridium difficile carrying the CD38 prophage. Appl. Environ. Microbiol. 2015, 81, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Yu, J.; McDowell, A.; Kim, S.H.; You, H.J.; Ko, G. Bile salt hydrolase-mediated inhibitory effect of Bacteroides ovatus on growth of Clostridium difficile. J. Microbiol. 2017, 55, 892–899. [Google Scholar] [CrossRef]
- Macy, J.M.; Ljungdahl, L.G.; Gottschalk, G. Pathway of succinate and propionate formation in Bacteroides fragilis. J. Bacteriol. 1978, 134, 84–91. [Google Scholar] [CrossRef]
- Dicks, L.M.T. How does quorum sensing of intestinal bacteria affect our health and mental status? Microorganisms 2022, 10, 1969. [Google Scholar] [CrossRef]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signaling in bacteria. FEMS Microbiol. Rev. 2013, 37, e156–e181. [Google Scholar] [CrossRef]
- Khan, C.M.A. The dynamic interactions between Salmonella and the microbiota, within the challenging niche of the gastrointestinal tract. Int. Sch. Res. Not. 2014, 2014, 846049. Available online: https://www.hindawi.com/journals/isrn/2014/846049/ (accessed on 14 June 2023). [CrossRef]
- Vendeville, A.; Winzer, K.; Heurlier, K.; Tang, K.; Hardie, K. Making “sense” of metabolism: Autoinducer-2, LuxS and pathogenic bacteria. Nature 2005, 3, 383–396. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Grenier, D.; Yi, L. Regulatory mechanisms of the LuxS/AI-2 system and bacterial resistance. Antimicrob. Agents Chemother. 2019, 63, e01186-19. [Google Scholar] [CrossRef]
- Carter, G.; Purdy, D.; Williams, P.; Minton, N. Quorum sensing in Clostridium difficile: Analysis of a luxS-type signaling system. J. Med. Microbiol. 2005, 54, 119–127. [Google Scholar] [CrossRef]
- Tijerina-Rodríguez, L.; Villarreal-Treviño, L.; Baines, S.D.; Morfín-Otero, R.; Camacho-Ortíz, A.; Flores-Treviño, S.; Maldonado-Garza, H.; Rodríguez-Noriega, E.; Garza-González, E. High sporulation and overexpression of virulence factors in biofilms and reduced susceptibility to vancomycin and linezolid in recurrent Clostridium [Clostridioides] difficile infection isolates. PLoS ONE 2019, 14, e0220671. [Google Scholar] [CrossRef] [PubMed]
- Tanghe, A.; Van Dijck, P.; Thevelein, J.M. Determinants of freeze tolerance in microorganisms, physiological importance, and biotechnological applications. Adv. Appl. Microbiol. 2003, 53, 129–176. [Google Scholar]
- Zhu, Z.; Wang, H.; Shang, Q.; Jiang, Y.; Cao, Y.; Chai, Y. Time course analysis of Candida albicans metabolites during biofilm development. J. Proteome Res. 2013, 12, 2375–2385. [Google Scholar] [CrossRef]
- Chen, S.; Sun, C.; Wang, H.; Wang, J. The role of Rho GTPases in toxicity of Clostridium difficile toxins. Toxins 2015, 7, 5254–5267. [Google Scholar] [CrossRef] [PubMed]
- Just, I.R.; Gerhard, R. Large clostridial cytotoxins. Rev. Physiol Biochem. Pharmacol. 2004, 152, 23–47. [Google Scholar] [PubMed]
- Govind, R.; Dupuy, B. Secretion of Clostridium difficile toxins A and B requires the holin-like protein TcdE. PLoS Pathog. 2012, 8, e1002727. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; England, P.; Dupuy, B. Clostridium difficile toxin expression is inhibited by the novel regulator TcdC. Mol. Microbiol. 2007, 64, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Mani, N.; Dupuy, B. Regulation of toxin synthesis in Clostridium difficile by an alternative RNA polymerase sigma factor. Proc. Natl. Acad. Sci. USA 2001, 98, 5844–5849. [Google Scholar] [CrossRef]
- Mani, N.; Lyras, D.; Barroso, L.; Howarth, P.; Wilkins, T.; Rood, J.I.; Sonenshein, A.L.; Dupuy, B. Environmental response and autoregulation of Clostridium difficile TxeR, a sigma factor for toxin gene expression. J. Bacteriol. 2002, 184, 5971–5978. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Douce, G.R.; Govind, R.; Howarth, P.M.; Mackin, K.E.; Spencer, J.; Buckley, A.M.; Antunes, A.; Kotsanas, D.; Jenkin, G.A.; et al. The anti-sigma factor TcdC modulates hypervirulence in an epidemic BI/NAP1/027 clinical isolate of Clostridium difficile. PLoS Pathog. 2011, 7, e1002317. [Google Scholar] [CrossRef] [PubMed]
- Hundsberger, T.; Braun, V.; Weidmann, M.; Leukel, P.; Sauerborn, M.; Von Eichel-Streiber, C. Transcription analysis of the genes tcdA-E of the pathogenicity locus of Clostridium difficile. Eur. J. Biochem. 1997, 244, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Cartman, S.T.; Kelly, M.L.; Heeg, D.; Heap, J.T.; Minton, N.P. Precise manipulation of the Clostridium difficile chromosome reveals a lack of association between the tcdC genotype and toxin production. Appl. Environ. Microbiol. 2012, 78, 4683–4690. [Google Scholar] [CrossRef]
- Bakker, D.; Smits, W.K.; Kuijper, E.J.; Corver, J. TcdC does not significantly repress toxin expression in Clostridium difficile 630ΔErm. PLoS ONE 2012, 7, e43247. [Google Scholar] [CrossRef]
- Darkoh, C.; DuPont, H.L.; Norris, S.J.; Kaplan, H.B. Toxin synthesis by Clostridium difficile is regulated through quorum signaling. MBio 2015, 6, e02569-14. [Google Scholar] [CrossRef]
- Buddelmeijer, N.; Young, R. The essential Escherichia coli apolipoprotein N-acyltransferase (Lnt) exists as an extracytoplasmic thioester acyl-enzyme intermediate. Biochemistry 2010, 49, 341–346. [Google Scholar] [CrossRef]
- List, F.; Bocola, M.; Haeger, M.C.; Sterner, R. Constitutively active glutaminase variants provide insights into the activation mechanism of anthranilate synthase. Biochemistry 2012, 51, 2812–2818. [Google Scholar] [CrossRef]
- Skaff, D.A.; Ramyar, K.X.; McWhorter, W.J.; Barta, M.L.; Geisbrecht, B.V.; Miziorko, H.M. Biochemical and structural basis for inhibition of Enterococcus faecalis hydroxymethylglutaryl-CoA synthase, mvaS, by hymeglusin. Biochemistry 2012, 51, 4713–4722. [Google Scholar] [CrossRef]
- Darkoh, C.; Deaton, M.; DuPont, H.L. Nonantimicrobial drug targets for Clostridium difficile infections. Future Microbiol. 2017, 12, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Darkoh, C.; Odo, C.; DuPont, H.L. Accessory gene regulator-1 locus is essential for virulence and pathogenesis of Clostridium difficile. MBio 2016, 7, e01237-16. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.J.; Clare, S.; Goulding, D.; Faulds-Pain, A.; Barquist, L.; Browne, H.P.; Pettit, L.; Dougan, G.; Lawley, T.D.; Wren, B.W. The agr locus regulates virulence and colonization genes in Clostridium difficile 027. J. Bacteriol. 2013, 195, 3672–3681. [Google Scholar] [CrossRef]
- Ahmed, U.K.B.; Shadid, T.M.; Larabee, J.L.; Ballard, J.D. Combined and distinct roles of Agr proteins in Clostridioides difficile 630 sporulation, motility, and toxin production. MBio 2020, 11, e03190-20. [Google Scholar] [CrossRef] [PubMed]
- Subauste, M.; Von Herrath, M.; Benard, V.; Chamberlain, C.E.; Chuang, T.-H.; Chu, K.; Bokoch, G.M.; Hahn, K.M. Rho family proteins modulate rapid apoptosis induced by cytotoxic T lymphocytes and Fas. J. Biol. Chem. 2000, 275, 9725–9733. [Google Scholar] [CrossRef]
- Caron, E.; Hall, A. Identification of two distinct mechanisms of phagocytosis controlled by different Rho GTPases. Science 1998, 282, 1717–1721. [Google Scholar] [CrossRef]
- Gerding, D.N.; Muto, C.A.; Owens, J.R.C. Treatment of Clostridium difficile infection. Clin. Infect. Dis. 2008, 46, S32–S42. [Google Scholar] [CrossRef] [PubMed]
- Voth, D.E.; Ballard, J.D. Clostridium difficile toxins: Mechanism of action and role in disease. Clin. Microbiol. Rev. 2005, 18, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G.; Walkty, A.J.; Karlowsky, J.A. Fidaxomicin: A novel agent for the treatment of Clostridium difficile infection. Can. J. Infect. Dis. Med. Microbiol. 2015, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Tannock, G.W.; Munro, K.; Taylor, C.; Lawley, B.; Young, W.; Byrne, B.; Emery, J.; Louie, T. A new macrocyclic antibiotic, fidaxomicin (OPT-80). Microbiology 2010, 156, 3354–3359. [Google Scholar] [CrossRef]
- Recommendations for Preventing the Spread of Vancomycin Resistance. Recommendations of the Hospital Infection Control Practices Advisory Committee (HICPAC). MMWR Recomm. Rep. 1995, 44, 105–113. [Google Scholar]
- ASHP therapeutic position statement on the preferential use of metronidazole for the treatment of Clostridium difficile-associated disease. Am. J. Health Syst. Pharm. 1998, 55, 1407–1411. [CrossRef]
- Cornely, O.A.; Miller, M.A.; Louie, T.J.; Crook, D.W.; Gorbach, S.L. Treatment of first recurrence of Clostridium difficile infection: Fidaxomicin versus vancomycin. Clin. Infect. Dis. 2012, 55, S154–S161. [Google Scholar] [CrossRef]
- Louie, T.J.; Miller, M.A.; Mullane, K.M.; Weiss, K.; Lentnek, A.; Golan, Y.; Gorbach, S.; Sears, P.; Shue, Y.-K. Fidaxomicin versus vancomycin for Clostridium difficile infection. N. Engl. J. Med. 2011, 364, 422–431. [Google Scholar] [CrossRef]
- Aslam, S.; Hamill, R.J.; Musher, D.M. Treatment of Clostridium difficile associated disease: Old therapies and new strategies. Lancet Infect. Dis. 2005, 5, 549–557. [Google Scholar] [CrossRef]
- Cornely, O.A.; Miller, M.A.; Fantin, B.; Mullane, K.; Kean, Y.; Gorbach, S. Resolution of Clostridium difficile-associated diarrhea in patients with cancer treated with fidaxomicin or vancomycin. J. Clin. Oncol. 2013, 31, 2493–2499. [Google Scholar] [CrossRef] [PubMed]
- Mullane, K.M.; Cornely, O.A.; Crook, D.W.; Golan, Y.; Louie, T.J.; Miller, M.A.; Josephson, M.A.; Gorbach, S.L. Renal impairment and clinical outcomes of Clostridium difficile infection in two randomized trials. Am. J. Nephrol. 2013, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mullane, K.M.; Miller, M.A.; Weiss, K.; Lentnek, A.; Golan, Y.; Sears, P.S.; Shue, Y.-K.; Louie, T.J.; Gorbach, S.L. Efficacy of fidaxomicin versus vancomycin as therapy for Clostridium difficile infection in individuals taking concomitant antibiotics for other concurrent infections. Clin. Infect. Dis. 2011, 53, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Louie, T.J.; Gerding, D.N.; Cornely, O.; Chasan-Taber, S.; Fitts, D.; Gelone, S.P.; Broom, C.; Davidson, D.M.; for the Polymer Alternative for CDI Treatment (PACT) investigators. Vancomycin, metronidazole, or tolevamer for Clostridium difficile infection: Results from two multinational, randomized, controlled trials. Clin. Infect. Dis. 2014, 59, 345–354. [Google Scholar] [CrossRef]
- Vuotto, C.; Moura, I.; Barbanti, F.; Donelli, G.; Spigaglia, P. Subinhibitory concentrations of metronidazole increase biofilm formation in Clostridium difficile strains. Pathog. Dis. 2016, 74, ftv114. [Google Scholar] [CrossRef]
- Alotaibi, B. The Role of Clostridium difficile Surface Structures in Virulence and Gut Colonization; Ulster University: Belfast, UK, 2017. [Google Scholar]
- Kelly, C.P. Can we identify patients at high risk of recurrent Clostridium difficile infection? Clin. Microbiol. Infect. 2012, 6, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, A.A.; Johnson, S. Fidaxomicin: A novel macrocyclic antibiotic approved for treatment of Clostridium difficile infection. Clin. Infect. Dis. 2012, 54, 568–574. [Google Scholar] [CrossRef]
- Artsimovitch, I.; Seddon, J.; Sears, P. Fidaxomicin is an inhibitor of the initiation of bacterial RNA synthesis. Clin. Infect. Dis. 2012, 55 (Suppl. S2), S127–S131. [Google Scholar] [CrossRef] [PubMed]
- Administration USFD. Search Orphan Drug Designation and Approval. 2022. Available online: https://www.accessdata.fda.gov/scripts/opdlisting/oopd/detailedIndex.cfm?cfgridkey=325210 (accessed on 21 June 2023).
- Merck. FDA Approves Merck’s DIFICID (Fidaxomicin) to Treat Clostridioides difficile in Children Aged Six Months and Older. 2020. Available online: https://www.merck.com/news/fda-approves-mercks-dificid-fidaxomicin-to-treat-clostridioides-difficile-in-children-aged-six-months-and-older/ (accessed on 21 June 2023).
- Giancola, S.E.; Williams RJ 2nd Gentry, C.A. Evaluation of fidaxomicin usage patterns and outcomes for Clostridium difficile infection across the United States veterans health administration. J. Clin. Pharm. Ther. 2018, 43, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Polivkovaa, S.; Krutovab, M.; Capekc, V.; Sykorovad, B.; Benesa, J. Fidaxomicin versus metronidazole, vancomycin and their combination for initial episode, first recurrence and severe Clostridioides difficile infection––An observational cohort study. Int. J. Infect. Dis. 2021, 103, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Lavergne, V.; Skinner, A.M.; Gonzales-Luna, A.J.; Garey, K.W.; Kelly, C.P.; Wilcox, M.H. Clinical practice guideline by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA): 2021 focused update guidelines on management of Clostridioides difficile infection in adults. Clin. Infect. Dis. 2021, 73, e1029–e1044. [Google Scholar] [CrossRef]
- Al-Jashaami, L.S.; Dupont, H.L. Management of Clostridium difficile infection. Gastroenterol Hepatol. 2016, 12, 609–616. [Google Scholar]
- Cornely, O.A.; Crook, D.W.; Esposito, R.; Poirier, A.; Somero, M.S.; Weiss, K.; Sears, P.; Gorbach, S. Fidaxomicin versus vancomycin for infection with Clostridium difficile in Europe, Canada, and the USA: A double-blind, non-inferiority, randomised controlled trial. Lancet Infect. Dis. 2012, 12, 281–289. [Google Scholar] [CrossRef]
- Weiss, K.; Allgren, R.L.; Sellers, S. Safety analysis of fidaxomicin in comparison with oral vancomycin for Clostridium difficile infections. Clin. Infect. Dis. 2012, 55, S110–S115. [Google Scholar] [CrossRef]
- Mikamo, H.; Tateda, K.; Yanagihara, K.; Kusachi, S.; Takesue, Y.; Miki, T.; Oizumi, Y.; Gamo, K.; Hashimoto, A.; Toyoshima, J.; et al. Efficacy and safety of fidaxomicin for the treatment of Clostridioides (Clostridium) difficile infection in a randomized, double-blind, comparative Phase III study in Japan. J. Infect. Chemother. 2018, 24, 744–752. [Google Scholar] [CrossRef]
- Nelson, R.L.; Suda, K.J.; Evans, C.T. Antibiotic treatment for Clostridium difficile-associated diarrhoea in adults. Cochrane Database Syst. Rev. 2017, 3, CD004610. [Google Scholar] [CrossRef]
- Tomas, M.E.; Mana, T.S.C.; Wilson, B.M.; Nerandzic, M.M.; Joussef-Piña, S.; Quiñones-Mateu, M.E.; Donskey, C.J. Tapering courses of oral vancomycin induce persistent disruption of the microbiota that provide colonization resistance to Clostridium difficile and vancomycin-resistant enterococci in mice. Antimicrob. Agents Chemother. 2018, 62, e02237-17. [Google Scholar] [CrossRef] [PubMed]
- Dureja, C.; Olaitan, A.O.; Hurdle, J.G. Mechanisms and impact of antimicrobial resistance in Clostridioides difficile. Curr. Opin. Microbiol. 2022, 66, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Wostem, M.M. Eubacterial sigma-factors. FEMS Microbiol. Rev. 1998, 22, 127–150. [Google Scholar] [CrossRef]
- Karlowsky, J.A.; Laing, N.M.; Zhanel, G.G. In vitro activity of OPT-80 tested against clinical isolates of toxin-producing Clostridium difficile. Antimicrob. Agents Chemother. 2008, 52, 4163–4165. [Google Scholar] [CrossRef]
- Babakhani, F.; Seddon, J.; Sears, P. Comparative microbiological studies of transcription inhibitors fidaxomicin and the rifamycins in Clostridium difficile. Antimicrob. Agents Chemother. 2014, 58, 2934–2937. [Google Scholar] [CrossRef]
- Gualtieri, M.; Villain-Guillot, P.; Latouche, J.; Leonetti, J.P.; Bastide, L. Mutation in the Bacillus subtilis RNA polymerase β’ subunit confers resistance to lipiarmycin. Antimicrob. Agents Chemother. 2006, 50, 401–402. [Google Scholar] [CrossRef]
- Artsimovitch, I.; Vassylyeva, M.N.; Svetlov, D.; Svetlov, V.; Perederina, A.; Igarashi, N.; Matsugaki, N.; Wakatsuki, S.; Tahirov, T.H.; Vassylyev, D.G. Allosteric modulation of the RNA polymerase catalytic reaction is an essential component of transcription control by rifamycins. Cell 2005, 122, 351–363. [Google Scholar] [CrossRef]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Dempster, A.W.; Heeg, D.; Longshaw, C.; Minton, N. Use of allelic exchange to characterize the impact of rpoB/C mutations on fitness of Clostridium difficile and sensitivity to fidaxomicin. In Proceedings of the 23rd European Congress of Clinical Microbiology and Infectious Diseases (ECCMID), Berlin, Germany, 27–30 April 2013. [Google Scholar]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef]
- Elmore, B.O.; Triplett, K.D.; Hall, P.R. Apolipoprotein B48, the structural component of chylomicrons, is sufficient to antagonize Staphylococcus aureus quorum-sensing. PLoS ONE 2015, 10, e0125027. [Google Scholar] [CrossRef]
- Horswill, A.R.; Gordon, C.P. Structure-activity relationship studies of small molecule modulators of the staphylococcal accessory gene regulator. J. Med. Chem. 2020, 63, 2705–2730. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, S.R.; Jiang, B.; Hu, X.M.; Li, S. Therapeutic targeting of the Staphylococcus aureus accessory gene regulator (agr) system. Front. Microbiol. 2018, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Louie, T.J.; Peppe, J.; Watt, C.K.; Johnson, D.; Mohammed, R.; Dow, G.; Weiss, K.; Simon, S.; John, J.F.; Garber, G.; et al. Tolevamer study investigator group. Tolevamer, a novel nonantibiotic polymer, compared with vancomycin in the treatment of mild to moderately severe Clostridium difficile-associated diarrhea. Clin. Infect. Dis. 2006, 43, 411–420. [Google Scholar] [CrossRef]
- Babcock, G.J.; Broering, T.J.; Hernandez, H.J.; Mandell, R.B.; Donahue, K.; Boatright, N.; Stack, A.M.; Lowy, I.; Graziano, R.; Molrine, D.; et al. Human monoclonal antibodies directed against toxins A and B prevent Clostridium difficile-induced mortality in hamsters. Infect. Immun. 2006, 74, 6339–6347. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.L.; Compson, J.E.; MacKenzie, B.; O’Dowd, V.L.; Oxbrow, A.K.F.; Heads, J.T.; Turner, A.; Sarkar, K.; Dugdale, S.L.; Jairaj, M.; et al. A mixture of functionally oligoclonal humanized monoclonal antibodies that neutralize Clostridium difficile TcdA and TcdB with high levels of in vitro potency shows in vivo protection in a hamster infection model. Clin. Vaccine Immunol. 2013, 20, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ramsey, J.; Hamza, T.; Zhang, Y.; Li, S.; Yfantis, H.G.; Lee, D.; Hernandez, L.D.; Seghezzi, W.; Furneisen, J.M.; et al. Mechanisms of protection against Clostridium difficile infection by the monoclonal antitoxin antibodies actoxumab and bezlotoxumab. Infect. Immun. 2015, 83, 822–831. [Google Scholar] [CrossRef]
- Steele, J.; Mukherjee, J.; Parry, N.; Tzipori, S. Antibody against TcdB, but not TcdA, prevents development of gastrointestinal and systemic Clostridium difficile disease. J. Infect. Dis. 2013, 207, 323–330. [Google Scholar] [CrossRef]
- Lowy, I.; Molrine, D.C.; Leav, B.A.; Blair, B.M.; Baxter, R.; Gerding, D.N.; Nichol, G.; Thomas, W.D.; Leney, M.; Sloan, S.; et al. Treatment with monoclonal antibodies against Clostridium difficile toxins. N. Engl. J. Med. 2010, 362, 197–205. [Google Scholar] [CrossRef]
- Kociolek, L.K.; Gerding, D.N. Breakthroughs in the treatment and prevention of Clostridium difficile infection. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 150–160. [Google Scholar] [CrossRef]
- Bender, K.O.; Garland, M.; Ferreyra, J.A.; Hryckowian, A.J.; Child, M.A.; Puri, A.W.; Solow-Cordero, D.E.; Higginbottom, S.K.; Segal, E.; Banaei, N.; et al. A small-molecule antivirulence agent for treating Clostridium difficile infection. Sci. Transl. Med. 2015, 7, 306ra148. [Google Scholar] [CrossRef]
- Bruxelle, J.F.; Mizrahi, A.; Hoÿs, S.; Collignon, A.; Janoir, C.; Péchiné, S. Clostridium difficile flagellin FliC: Evaluation as adjuvant and use in a mucosal vaccine against Clostridium difficile. PLoS ONE 2017, 12, e0187212. [Google Scholar] [CrossRef]
- Humphreys, D.P.; Wilcox, M.H. Antibodies for treatment of Clostridium difficile infection. Clin. Vaccine Immunol. 2014, 21, 913–923. [Google Scholar] [CrossRef]
- Péchiné, S.; Janoir, C.; Boureau, H.; Gleizes, A.; Tsapis, N.; Hoys, S.; Fattal, E.; Collignon, A. Diminished intestinal colonization by Clostridium difficile and immune response in mice after mucosal immunization with surface proteins of Clostridium difficile. Vaccine 2007, 25, 3946–3954. [Google Scholar] [CrossRef]
- Mills, J.P.; Rao, K.; Young, V.B. Probiotics for prevention of Clostridium difficile infection. Curr. Opin. Gastroenterol. 2018, 34, 3–10. [Google Scholar] [CrossRef]
- Romyasamit, C.; Thatrimontrichai, A.; Aroonkesorn, A.; Chanket, W.; Ingviya, N.; Saengsuwan, P.; Singkhamanan, K. Enterococcus faecalis isolated from infant feces inhibits toxigenic Clostridioides (Clostridium) difficile. Front. Pediatr. 2020, 8, 572633. [Google Scholar] [CrossRef]
- Yun, B.; Oh, S.; Griffiths, M. Lactobacillus acidophilus modulates the virulence of Clostridium difficile. J. Dairy Sci. 2014, 97, 4745–4758. [Google Scholar] [CrossRef]
- Yong, C.C.; Lim, J.; Kim, B.K.; Park, D.J.; Oh, S. Suppressive effect of Lactobacillus fermentum Lim2 on Clostridioides difficile 027 toxin production. Letts Appl. Microbiol. 2019, 68, 386–393. [Google Scholar] [CrossRef]
- Ofosu, A. Clostridium difficile infection: A review of current and emerging therapies. Ann. Gastroenterol. 2016, 29, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Hedge, D.D.; Strain, J.D.; Heins, J.R.; Farver, D.K. New advances in the treatment of Clostridium difficile infection (CDI). Ther. Clin. Risk Manag. 2008, 4, 949–964. [Google Scholar]
- Koon, H.W.; Su, B.; Xu, C.; Mussatto, C.C.; Tran, D.H.-N.; Lee, E.C.; Ortiz, C.; Wang, J.; Lee, J.E.; Ho, S.; et al. Probiotic Saccharomyces boulardii CNCM I-745 prevents outbreak-associated Clostridium difficile-associated cecal inflammation in hamsters. Am. J. Physiol. Gastrointest Liver Physiol. 2016, 311, G610–G623. [Google Scholar] [CrossRef]
- Gerding, D.N.; Meyer, T.; Lee, C.; Cohen, S.H.; Murthy, U.K.; Poirier, A. Administration of spores of nontoxigenic Clostridium difficile strain M3 for prevention of recurrent C. difficile infection: A randomized clinical trial. JAMA 2015, 313, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Sipe, B.; Cheng, Y.-W.; Phelps, E.; Rogers, N.; Sagi, S.; Bohm, M.; Xu, H.; Kassam, Z. Fecal microbiota transplant in severe and severe-complicated Clostridium difficile: A promising treatment approach. Gut Microbes. 2016, 8, 289–302. [Google Scholar] [CrossRef]
- Tariq, R.; Pardi, D.S.; Bartlett, M.G.; Khanna, S. Low cure rates in controlled trials of fecal microbiota transplantation for recurrent Clostridium difficile infection: A systematic review and meta-analysis. Clin. Infect. Dis. 2019, 68, 1351–1358. [Google Scholar] [CrossRef]
- Quraishi, M.N.; Widlak, M.; Bhala, N.; Moore, D.; Price, M.; Sharma, N.; Iqbal, T.H. Systematic review with meta-analysis: The efficacy of faecal microbiota transplantation for the treatment of recurrent and refractory Clostridium difficile infection. Aliment. Pharmacol. Ther. 2017, 46, 479–493. [Google Scholar] [CrossRef]
- Ott, S.J.; Waetzig, G.H.; Rehman, A.; Moltzau-Anderson, J.; Bharti, R.; Grasis, J.A.; Cassidy, L.; Tholey, A.; Fickenscher, H.; Seegert, D.; et al. Efficacy of sterile fecal filtrate transfer for treating patients with Clostridium difficile infection. Gastroenterology 2017, 152, 799–811.e7. [Google Scholar] [CrossRef]
- Juul, F.E.; Garborg, K.; Bretthauer, M.; Skudal, H.; Øines, M.N.; Wiig, H.; Rose, O.; Seip, B.; Lamont, J.T.; Midtvedt, T.; et al. Fecal microbiota transplantation for primary Clostridium difficile infection. N. Engl. J. Med. 2018, 378, 2535–2536. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Masucci, L.; Quaranta, G.; Simonelli, C.; Lopetuso, L.R.; Sanguinetti, M.; Gasbarrini, A.; Cammarota, G. Randomised clinical trial: Faecal microbiota transplantation by colonoscopy plus vancomycin for the treatment of severe refractory Clostridium difficile infection-single versus multiple infusions. Aliment. Pharmacol. Ther. 2018, 48, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Venhorst, J.; van der Vossen, J.M.B.M.; Agamennone, V. Battling enteropathogenic clostridia: Phage therapy for Clostridioides difficile and Clostridium perfringens. Front. Microbiol. 2022, 13, 891790. [Google Scholar] [CrossRef] [PubMed]
- Smits, W.K.; Roseboom, A.M.; Corver, J. Plasmids of Clostridioides difficile. Curr. Opin. Microbiol. 2022, 65, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Meessen-Pinard, M.; Fortier, L.C. Prophage-stimulated toxin production in Clostridium difficile NAP1/027 lysogens. J. Bacteriol. 2011, 193, 2726–2734. [Google Scholar] [CrossRef]
- Phothichaisri, W.; Ounjai, P.; Phetruen, T.; Janvilisri, T.; Khunrae, P.; Singhakaew, S.; Wangroongsarb, P.; Chankhamhaengdecha, S. Characterization of bacteriophages infecting clinical isolates of Clostridium difficile. Front. Microbiol. 2018, 9, 1701. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Ospina Bedoya, M.; Fivian-Hughes, A.S.; Fairweather, N.F.; Fortier, L.-C. The Clostridium difficile cell wall protein CwpV confers phase-variable phage resistance. Mol. Microbiol. 2015, 98, 329–342. [Google Scholar] [CrossRef]
- Hargreaves, K.R.; Kropinski, A.M.; Clokie, M.R.J. What does the talking? Quorum sensing signalling genes discovered in a bacteriophage genome. PLoS ONE 2014, 9, e85131. [Google Scholar] [CrossRef]
- Riedel, T.; Wittmann, J.; Bunk, B.; Schober, I.; Spröer, C.; Gronow, S.; Overmann, J. A Clostridioides difficile bacteriophage genome encodes functional binary toxin-associated genes. J. Biotechnol. 2017, 250, 23–28. [Google Scholar] [CrossRef]
- Meader, E.; Mayer, M.J.; Steverding, D.; Carding, S.R.; Narbad, A. Evaluation of bacteriophage therapy to control Clostridium difficile and toxin production in an in vitro human colon model system. Anaerobe 2013, 22, 25–30. [Google Scholar] [CrossRef]
- Nale, J.Y.; Spencer, J.; Hargreaves, K.R.; Buckley, A.M.; Trzepiński, P.; Douce, G.R.; Clokie, M.R.J. Bacteriophage combinations significantly reduce Clostridium difficile growth in vitro and proliferation in vivo. Antimicrob. Agents Chemother. 2016, 60, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Nale, J.Y.; Chutia, M.; Carr, P.; Hickenbotham, P.T.; Clokie, M.R.J. “Get in early”; biofilm and wax moth (Galleria mellonella) models reveal new insights into the therapeutic potential of Clostridium difficile bacteriophages. Front. Microbiol. 2016, 7, 1383. [Google Scholar] [CrossRef]
- Sangster, W.; Hegarty, J.P.; Stewart, D.B. Phage tail-like particles kill Clostridium difficile and represent an alternative to conventional antibiotics. Surgery 2015, 157, 96–103. [Google Scholar] [CrossRef]
- Gebhart, D.; Williams, S.R.; Bishop-Lilly, K.A.; Govoni, G.R.; Willner, K.M.; Butani, A.; Sozhamannan, S.; Martin, D.; Fortier, L.C.; Scholl, D. Novel high-molecular-weight, R-type bacteriocins of Clostridium difficile. J. Bacteriol. 2012, 194, 6240–6247. [Google Scholar] [CrossRef] [PubMed]
- Dams, D.; Brøndsted, L.; Drulis-Kawa, Z.; Briers, Y. Engineering of receptor-binding proteins in bacteriophages and phage tail-like bacteriocins. Biochem. Soc. Trans. 2019, 47, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Heuler, J.; Fortier, L.-C.; Sun, X. Clostridioides difficile phage biology and application. FEMS Microbiol. Rev. 2021, 45, fuab012. [Google Scholar] [CrossRef]
- Mayer, M.J.; Narbad, A.; Gasson, M.J. Molecular characterization of a Clostridium difficile bacteriophage and its cloned biologically active endolysin. J. Bacteriol. 2008, 190, 6734–6740. [Google Scholar] [CrossRef]
- Mayer, M.J.; Garefalaki, V.; Spoerl, R.; Narbad, A.; Meijers, R. Structure-based modification of a Clostridium difficile-targeting endolysin affects activity and host range. J. Bacteriol. 2011, 193, 5477–5486. [Google Scholar] [CrossRef]
- Wang, Q.; Euler, C.W.; Delaune, A.; Fischetti, V.A. Using a novel lysin to help control Clostridium difficile infections. Antimicrob. Agents Chemother. 2015, 59, 7447–7457. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cyclic di-GMP (c-di-GMP)-mediated riboswitches control the colonization of C. difficile to the host’s epithelial cells. Type I and II riboswitches control the expression of factors that are involved in motility, surface attachment, and virulence, including production of TcdA and TcdB. Type I riboswitches (off switches) inhibit translation following the binding of c-di-GMP, whereas type II riboswitches (on switches) promote the translation of target genes when bound to c-di-GMP. Increased levels of c-di-GMP stimulate the expression of adhesion factors, such as type IV pili and collagen-binding proteins (CBPs), and inhibit the expression of flagellar genes and CBP protease. Biofilm formation is altered by antimicrobials, QS signals, and the transcription factor spo0A that regulates sporulation. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.
Figure 1.
Cyclic di-GMP (c-di-GMP)-mediated riboswitches control the colonization of C. difficile to the host’s epithelial cells. Type I and II riboswitches control the expression of factors that are involved in motility, surface attachment, and virulence, including production of TcdA and TcdB. Type I riboswitches (off switches) inhibit translation following the binding of c-di-GMP, whereas type II riboswitches (on switches) promote the translation of target genes when bound to c-di-GMP. Increased levels of c-di-GMP stimulate the expression of adhesion factors, such as type IV pili and collagen-binding proteins (CBPs), and inhibit the expression of flagellar genes and CBP protease. Biofilm formation is altered by antimicrobials, QS signals, and the transcription factor spo0A that regulates sporulation. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.

Figure 2.
Quorum sensing (QS) in C. difficile. Two communication systems have been identified: an inter-species LuxSCD system (top section of diagram) and an intra-species accessory gene regulator (Agr) system (bottom section of diagram). The enzyme LuxSCD produces auto-inducer peptides (AI-2, red circles) that interact with receptors on another Gram-positive cell and is phosphorylated (top section of diagram). At a certain threshold, transcription of the PaLoc genes in the responding cell and the LuxSCD locus (not shown in responding cell) is activated. Intra-species communication (between two strains of C. difficile) occurs with TI (thiolactone peptide, depicted as yellow stars, bottom part of diagram). At a certain threshold, TI attaches to AgrC (a transmembrane protein, blue square in purple bracket), and AgrA is phosphorylated before it activates the transcription of the Agr and PaLoc loci. Two Agr loci (Agr1 and Agr2) have been identified. The Agr1 locus contains quorum signal generation genes agrB1 and agrD1 that encodes AgrB1 (a transmembrane protein) and AgrD1 (a prepeptide) responsible for production of TI. Sufficient levels of T1 may trigger TcdA and TcdB production. The Agr2 locus contains quorum signal generation genes agrB2 and agrD2, and response genes agrC2 and agrA2. Hypervirulent strains, such as R20291, have loci Agr1 and Agr2. Non-hypervirulent strains, such as 630, have only the Agr1 locus. TI interacts with a two-component AgrC2 histidine kinase, which phosphorylates and dimerizes the response regulator AgrA2. This induces toxin production directly or indirectly. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.
Figure 2.
Quorum sensing (QS) in C. difficile. Two communication systems have been identified: an inter-species LuxSCD system (top section of diagram) and an intra-species accessory gene regulator (Agr) system (bottom section of diagram). The enzyme LuxSCD produces auto-inducer peptides (AI-2, red circles) that interact with receptors on another Gram-positive cell and is phosphorylated (top section of diagram). At a certain threshold, transcription of the PaLoc genes in the responding cell and the LuxSCD locus (not shown in responding cell) is activated. Intra-species communication (between two strains of C. difficile) occurs with TI (thiolactone peptide, depicted as yellow stars, bottom part of diagram). At a certain threshold, TI attaches to AgrC (a transmembrane protein, blue square in purple bracket), and AgrA is phosphorylated before it activates the transcription of the Agr and PaLoc loci. Two Agr loci (Agr1 and Agr2) have been identified. The Agr1 locus contains quorum signal generation genes agrB1 and agrD1 that encodes AgrB1 (a transmembrane protein) and AgrD1 (a prepeptide) responsible for production of TI. Sufficient levels of T1 may trigger TcdA and TcdB production. The Agr2 locus contains quorum signal generation genes agrB2 and agrD2, and response genes agrC2 and agrA2. Hypervirulent strains, such as R20291, have loci Agr1 and Agr2. Non-hypervirulent strains, such as 630, have only the Agr1 locus. TI interacts with a two-component AgrC2 histidine kinase, which phosphorylates and dimerizes the response regulator AgrA2. This induces toxin production directly or indirectly. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.

Figure 3.
Pathogenicity locus (PaLoc) of C. difficile and regulation of toxin gene expression. For simplicity, genes in the PaLoc are depicted as light-blue rectangles. (A) During logarithmic growth, tcdA, tcdB, and tcdR (boxed) are expressed at low levels, regulated by a TcdR-independent promoter (PtcdR, orange). In the presence of GTP (guanosine triphosphate) and BCAAs (branched-chain amino acids, e.g., leucine, isoleucine, and valine), CodY (a transcriptional regulator of sporulation) binds to PtcdR and suppresses the expression of tcdR. This also leads to low expression of toxin genes tcdA and tcdB. The negative regulator gene, tcdC (on far right), is maximally expressed to produce the membrane-associated protein TcdC (in purple). The latter insulates minute amounts of TcdR (red squares) that may be expressed and further suppresses the expression of toxin genes. (B) During stationary (nutrient-limiting conditions or stress), the expression of toxin genes is induced. RNAP (depicted as a large purple circle), with an active site (red star), binds to a promoter-specificity σ initiation factor, also called the σ-specificity factor (yellow circle). The resulting holo RNAP (large gray circle on the left) reacts with promoters PtcdR, PtcdB, and PtcdA, respectively. The double-stranded DNA is loosely bound on the surface of RNAP. The two smaller gray circles also represent holo RNAP. In the absence of GTP and BCAA, CodY (not shown here) does not bind to PtcdR, and tcdR expression occurs unhindered. The tcdC gene (on far right) is not optimally expressed, which means TcdR is free to interact with RNAP and form the holozyme complex. The latter binds to the tcdR promoter (PtcdR) and promotors PtcdB and PtcdA, leading to maximal expression of tcdA, tcdB, and tcdR and thus high levels of toxins TcdA and TcdB. TcdE, a holin-like protein (depicted as brown rectangles in cell wall), facilitates the release of toxins TcdA and TcdB. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.
Figure 3.
Pathogenicity locus (PaLoc) of C. difficile and regulation of toxin gene expression. For simplicity, genes in the PaLoc are depicted as light-blue rectangles. (A) During logarithmic growth, tcdA, tcdB, and tcdR (boxed) are expressed at low levels, regulated by a TcdR-independent promoter (PtcdR, orange). In the presence of GTP (guanosine triphosphate) and BCAAs (branched-chain amino acids, e.g., leucine, isoleucine, and valine), CodY (a transcriptional regulator of sporulation) binds to PtcdR and suppresses the expression of tcdR. This also leads to low expression of toxin genes tcdA and tcdB. The negative regulator gene, tcdC (on far right), is maximally expressed to produce the membrane-associated protein TcdC (in purple). The latter insulates minute amounts of TcdR (red squares) that may be expressed and further suppresses the expression of toxin genes. (B) During stationary (nutrient-limiting conditions or stress), the expression of toxin genes is induced. RNAP (depicted as a large purple circle), with an active site (red star), binds to a promoter-specificity σ initiation factor, also called the σ-specificity factor (yellow circle). The resulting holo RNAP (large gray circle on the left) reacts with promoters PtcdR, PtcdB, and PtcdA, respectively. The double-stranded DNA is loosely bound on the surface of RNAP. The two smaller gray circles also represent holo RNAP. In the absence of GTP and BCAA, CodY (not shown here) does not bind to PtcdR, and tcdR expression occurs unhindered. The tcdC gene (on far right) is not optimally expressed, which means TcdR is free to interact with RNAP and form the holozyme complex. The latter binds to the tcdR promoter (PtcdR) and promotors PtcdB and PtcdA, leading to maximal expression of tcdA, tcdB, and tcdR and thus high levels of toxins TcdA and TcdB. TcdE, a holin-like protein (depicted as brown rectangles in cell wall), facilitates the release of toxins TcdA and TcdB. This illustration was made using BioRender (https://biorender.com/), accessed on 3 April 2023.

Figure 4.
Mechanisms of C. difficile toxin pathogenicity. (A) The presentation is that of TcdB. Both toxins, TcdB and TcdA, bind to the surface of intestinal epithelial cells. The GTD (glucosyltransferase domain, in pink), located at the N terminus, is linked to CPD (cysteine protease domain, in green) involved in cleaving of the toxin. The TD (translocation domain, light blue) is involved in pore formation, conformational changes, and the delivery of the GTD and CPD. The CROP (combined repetitive oligopeptide, dark blue) binds to receptors on the cell membrane. After binding, the toxin is internalized, followed by endocytosis (top half of diagram A). The intracellular acidic environment of the endosome causes the toxin to undergo conformational changes and initiate pore formation (bottom part of diagram A). CPD and GTD are translocated across the endosomal membrane and then cleaved by a protease activated by inositol hexaphosphate (InsP6). GTD is released into the host cytosol and glucosylate Rho proteins to inactive glu-Rho. Activation and signal transduction of the Rho GTPases are regulated by a classic GTPase cycle. The cycle is controlled by regulatory proteins acting as GDIs (guanine nucleotide dissociation inhibitors, green triangle) that extract inactive Rho GTPase from membranes, GEFs (guanine nucleotide exchange factors, white triangle) that catalyze nucleotide exchange and mediate activation, and GAPs (GTPase-activating proteins, orange triangle) that stimulate GTP hydrolysis to GDP. (B) The active CDTb heptamer binds to the cell membrane, is enclosed in an endosome, and is then translocated across the membrane once the blocker (in pink) is released. This results in fluid loss and irreversible actin depolymerization, which leads to the formation of microtubule-based protrusions (depicted in green). Rab11 = Ras-associated binding proteins (small GTPases) of the Ras superfamily that cycle between the cytosol and different membranes. Not shown here are Rab5 and Rab4. This illustration was made using BioRender (https://biorender.com/), accessed on 9 May 2023.
Figure 4.
Mechanisms of C. difficile toxin pathogenicity. (A) The presentation is that of TcdB. Both toxins, TcdB and TcdA, bind to the surface of intestinal epithelial cells. The GTD (glucosyltransferase domain, in pink), located at the N terminus, is linked to CPD (cysteine protease domain, in green) involved in cleaving of the toxin. The TD (translocation domain, light blue) is involved in pore formation, conformational changes, and the delivery of the GTD and CPD. The CROP (combined repetitive oligopeptide, dark blue) binds to receptors on the cell membrane. After binding, the toxin is internalized, followed by endocytosis (top half of diagram A). The intracellular acidic environment of the endosome causes the toxin to undergo conformational changes and initiate pore formation (bottom part of diagram A). CPD and GTD are translocated across the endosomal membrane and then cleaved by a protease activated by inositol hexaphosphate (InsP6). GTD is released into the host cytosol and glucosylate Rho proteins to inactive glu-Rho. Activation and signal transduction of the Rho GTPases are regulated by a classic GTPase cycle. The cycle is controlled by regulatory proteins acting as GDIs (guanine nucleotide dissociation inhibitors, green triangle) that extract inactive Rho GTPase from membranes, GEFs (guanine nucleotide exchange factors, white triangle) that catalyze nucleotide exchange and mediate activation, and GAPs (GTPase-activating proteins, orange triangle) that stimulate GTP hydrolysis to GDP. (B) The active CDTb heptamer binds to the cell membrane, is enclosed in an endosome, and is then translocated across the membrane once the blocker (in pink) is released. This results in fluid loss and irreversible actin depolymerization, which leads to the formation of microtubule-based protrusions (depicted in green). Rab11 = Ras-associated binding proteins (small GTPases) of the Ras superfamily that cycle between the cytosol and different membranes. Not shown here are Rab5 and Rab4. This illustration was made using BioRender (https://biorender.com/), accessed on 9 May 2023.

Figure 5.
Resistance mechanisms C. difficile developed against metronidazole (MTZ), vancomycin (VAN), and fidaxomicin. Resistance to MTZ is encoded by plasmid pCD-metro and genes involved in iron and redox metabolism. VAN resistance is either encoded by plasmid pX18-498 or the vanG operon that modifies the peptidoglycan by replacing the terminal D-Ala with D-Ser. Mutations in rpoB, rpoC, and MarR homolog CD2212 lead to resistance to fidaxomicin. Cells in biofilm are less affected by MTZ and VAN. Deletion of the ATP-binding cassette transporter CD2068 in efflux pumps reduces the activity of MTZ 1.4-fold. This illustration was made using BioRender (https://biorender.com/), accessed on 11 May 2023.
Figure 5.
Resistance mechanisms C. difficile developed against metronidazole (MTZ), vancomycin (VAN), and fidaxomicin. Resistance to MTZ is encoded by plasmid pCD-metro and genes involved in iron and redox metabolism. VAN resistance is either encoded by plasmid pX18-498 or the vanG operon that modifies the peptidoglycan by replacing the terminal D-Ala with D-Ser. Mutations in rpoB, rpoC, and MarR homolog CD2212 lead to resistance to fidaxomicin. Cells in biofilm are less affected by MTZ and VAN. Deletion of the ATP-binding cassette transporter CD2068 in efflux pumps reduces the activity of MTZ 1.4-fold. This illustration was made using BioRender (https://biorender.com/), accessed on 11 May 2023.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dicks, L.M.T. Biofilm Formation of Clostridioides difficile, Toxin Production and Alternatives to Conventional Antibiotics in the Treatment of CDI. Microorganisms 2023, 11, 2161. https://doi.org/10.3390/microorganisms11092161
AMA Style
Dicks LMT. Biofilm Formation of Clostridioides difficile, Toxin Production and Alternatives to Conventional Antibiotics in the Treatment of CDI. Microorganisms. 2023; 11(9):2161. https://doi.org/10.3390/microorganisms11092161
Chicago/Turabian StyleDicks, Leon M. T. 2023. "Biofilm Formation of Clostridioides difficile, Toxin Production and Alternatives to Conventional Antibiotics in the Treatment of CDI" Microorganisms 11, no. 9: 2161. https://doi.org/10.3390/microorganisms11092161
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.