
Probiotic Bacillus Strains Enhance T Cell Responses in Chicken
by
 , , and
, , and
Filip Larsberg
1,
Maximilian Sprechert
1,
Deike Hesse
1,
Gunnar Loh
2,
Gudrun A. Brockmann
1 and
Susanne Kreuzer-Redmer
3,* 1
Albrecht Daniel Thaer-Institute, Breeding Biology and Molecular Genetics, Humboldt Universität zu Berlin, Unter den Linden 6, 10099 Berlin, Germany
2
Evonik Operations GmbH-Research, Development & Innovation Nutrition & Care, Kantstraße 2, 33790 Halle, Germany
3
Institute of Animal Nutrition, Nutrigenomics, University of Veterinary Medicine Vienna, Veterinärplatz 1, 1210 Vienna, Austria
*
Author to whom correspondence should be addressed.
Microorganisms 2023, 11(2), 269; https://doi.org/10.3390/microorganisms11020269
Submission received: 20 December 2022
/
Revised: 10 January 2023
/
Accepted: 11 January 2023
/
Published: 19 January 2023
(This article belongs to the Section Veterinary Microbiology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Banning antibiotic growth promotors and other antimicrobials in poultry production due to the increasing antimicrobial resistance leads to increased feeding of potential alternatives such as probiotics. However, the modes of action of those feed additives are not entirely understood. They could act even with a direct effect on the immune system. A previously established animal-related in vitro system using primary cultured peripheral blood mononuclear cells (PBMCs) was applied to investigate the effects of immune-modulating feed additives. Here, the immunomodulation of different preparations of two probiotic Bacillus strains, B. subtilis DSM 32315 (BS), and B. amyloliquefaciens CECT 5940 (BA) was evaluated. The count of T-helper cells and activated T-helper cells increased after treatment in a ratio of 1:3 (PBMCs: Bacillus) with vital BS (CD4+: p < 0.05; CD4+CD25+: p < 0.01). Furthermore, vital BS enhanced the proliferation and activation of cytotoxic T cells (CD8+: p < 0.05; CD8+CD25+: p < 0.05). Cell-free probiotic culture supernatants of BS increased the count of activated T-helper cells (CD4+CD25+: p < 0.1). UV-inactivated BS increased the proportion of cytotoxic T cells significantly (CD8+: p < 0.01). Our results point towards a possible involvement of secreted factors of BS in T-helper cell activation and proliferation, whereas it stimulates cytotoxic T cells presumably through surface contact. We could not observe any effect on B cells after treatment with different preparations of BS. After treatment with vital BA in a ratio of 1:3 (PBMCs:Bacillus), the count of T-helper cells and activated T-helper cells increased (CD4+: p < 0.01; CD4+CD25+: p < 0.05). Cell-free probiotic culture supernatants of BA as well as UV-inactivated BA had no effect on T cell proliferation and activation. Furthermore, we found no effect of BA preparations on B cells. Overall, we demonstrate that the two different Bacillus strains enhanced T cell activation and proliferation, which points towards an immune-modulating effect of both strains on chicken immune cells in vitro. Therefore, we suggest that administering these probiotics can improve the cellular adaptive immune defense in chickens, thereby enabling the prevention and reduction of antimicrobials in chicken farming.
1. Introduction
Poultry is the cheapest source of animal protein worldwide and the most common source of meat. In 2030, poultry meat is expected to represent about 41% of all the protein from meat sources [1]. Furthermore, increased poultry production in developing countries [2] is accompanied by an elevated overall usage of antimicrobials. However, there is growing concern about the resistance of pathogenic bacteria against antibiotics and other antimicrobials, the residual effects of antibiotics in meat products [3], and the public health risk from zoonotic pathogens such as Salmonella and Campylobacter. Consequently, the subtherapeutic usage of antibiotic growth promotors and other antimicrobials has been forbidden in farming in the EU since 2006 (Regulation (EC) No 1831/2003). In the last decade, alternatives such as prebiotics [4,5], probiotics [4,5], synbiotics [6], and organic acids [7] have arisen to improve the health of animals and to reduce the usage of antibiotics and other antimicrobials. The most promising feed additives comprise probiotics, representing mainly live bacteria, fungi, and yeast, which supplement the gastrointestinal flora and promote the maintenance of a healthy digestive system. Probiotic usage has increased over the last years due to antibiotic-free poultry production and its well-researched benefits [8]. Each probiotic strain possesses varying levels of beneficial effects for the host [8]. These include the secretion of antimicrobial substances in the gastrointestinal tract (GIT), competitive adherence to the gut mucosa. the epithelium, as well as the improvement of the gut epithelial barrier and its integrity [9]. Moreover, probiotics improve growth performance [10], nutrient digestibility [11,12], and modulate the immune system of the host [13,14,15] as the interaction between host cells and the bacteria or their structural components may lead to modulation of either local or systemic T lymphocyte- (T cell) and B lymphocyte (B cell)-mediated immune responses [16]. Among the most widely and commonly used in-feed probiotics are Bacillus spp., which are spore-forming bacteria with resistance to high temperatures and harsh storing conditions [17]. These are generally considered as safe strains for use in poultry production [18,19] and comprise different positive effects on chicken health. Thus, the commercially available probiotic strains Bacillus amyloliquefaciens CECT 5940 (BA) and Bacillus subtilis DSM 32315 (BS) were reported to be effective in reducing or eliminating the negative effects caused by pathogenic bacteria such as Escherichia coli and Clostridium perfringens [20,21,22,23,24,25]. Moreover, they were shown to improve growth performance, increase feed efficiency, and manipulate the intestinal structure and microbial composition of the gut [20,24,26]. Furthermore, a positive effect on the immune system was reported in vivo [26,27]. An intact immune system in the GIT, the main portal of entry for pathogenic microorganisms, is indispensable. The two major cell types of the adaptive immune system are T and B cells. B cells contribute to the adaptive immune response through the secretion of antibodies, known as the humoral immune response, and can be recognized with an antibody against the chB6 (Bu-1) marker, a highly glycosylated protein of unique structure in chickens [28]. T cells mainly act as the cellular component of the adaptive immune response. The chicken T cell receptor (TCR) can be assigned to three subgroups which are identified by specific monoclonal antibodies [29]. γδ T cells, encompassing up to 50% of the peripheral blood T cells [30], can be recognized by the TCR1 antibody, whereas the two discrete subsets of αβ T cells can be labeled and recognized by TCR2 and TCR3 antibodies [29]. Like in humans and mice, αβ T cells can be further distinguished and subdivided by the expression of their co-receptors, cluster of differentiation 4 (CD4) and CD8 representing CD4+ T-helper cells and CD8+ cytotoxic T cells [31,32]. T and B cells exhibit distinct functions in mediating the immune response and are known to confer immunological protection by developing pathogen-specific memory [33].
This study aimed to assess, if the two probiotic Bacillus strains BA and BS are able to directly modulate two major components of the adaptive immune response, T and B cells, in chicken. A recently reported in vitro chicken immune cell assay utilizing peripheral blood mononuclear cells (PBMCs) [34] was used to investigate possible effects on the immune system.
By increasing the understanding of the functional mechanisms of probiotics on immune cells, this study can contribute to strategizing prevention measures and thereby help the poultry industry in raising healthy chickens. In particular, it will help to prevent and treat diseases by targeted administration of probiotics and thus prevent and reduce the usage of the subtherapeutic administration of antimicrobials.
2. Materials and Methods
2.1. Animals, Housing, Feeding, and Tissue Collection
The study was approved by the local state office of occupational health and technical safety “Landesamt für Gesundheit und Soziales Berlin” (LaGeSo Reg. T 0151/19). In total, 55 four-to-six-week-old broiler chickens of the commercial layer Cobb500 (Cobb Germany Avimex GmbH, Wiedemar, Germany) were used. The birds were kept and fed as described previously [34]. In brief, all chickens were fed a starter diet from day 1 to day 14 post hatch and a grower diet afterwards (H. Wilhelm Schaumann GmbH, Pinneberg, Germany). The ration was fed on an ad libitum basis and water was always available. The chickens were kept in groups of approximately 20 broiler chickens in 4 m² floor pens. For experiments, 3 to 6 birds per week were stunned and blood was sampled in sodium citrate (Na-citrate) pre-filled polystyrene tubes (VACUETTE®, Greiner Bio-One, Kremsmünster, Austria) by neck cutting.
2.2. PBMC Isolation and Culture
PBMCs were isolated using combined dextran–ficoll isolation as described previously [34,35]. In brief, blood was diluted with Dulbecco’s phosphate-buffered saline (DPBS, Gibco™, ThermoFisher Scientific, Waltham, MA, USA) containing 2 mM ethylenediaminetetraacetic acid (EDTA, Carl Roth, Karlsruhe, Germany) 1:1, mixed with 3% dextran (Carl Roth) in a ratio of 1:0.4, and centrifuged at 50× g for 20 min. Thereafter, the upper phase was collected, layered onto an equal volume of Histopaque®-1077 (Sigma-Aldrich, St. Louis, MO, USA), and centrifuged at 900× g for 30 min. The PBMCs at the interphase were collected, washed twice with DPBS/EDTA in a new 50 mL tube, and centrifuged at 400× g for 10 min. Isolated PBMCs were adjusted to 5×106 cells/mL living cells using Tali™ image-based cytometer (ThermoFisher Scientific) with propidium iodide (PI, ThermoFisher Scientific) as a viability marker. PBMCs were cultured in RPMI 1640 medium (Gibco™, ThermoFisher Scientific) with 10% chicken serum (Gibco™, ThermoFisher Scientific) and 1% penicillin (10,000 U/mL)-streptomycin (10,000 µg/mL) (Pen/Strep, Gibco™, ThermoFisher Scientific) at 41 °C with 5% CO2. The next day, further experiments were performed.
2.3. Bacterial Strains and Culture
RPMI 1640 medium was inoculated with either B. amyloliquefaciens CECT 5940 (BA) or B. subtilis DSM 32315 (BS) and bacteria were cultured overnight at 37 °C and 120 rpm. To define the actual colony forming units per milliliter (cfu/mL), the bacterial growth was measured on a Tecan Infinite® M200 Pro plate reader with Magellan™ v. 7.1 software (Tecan Group AG, Männedorf, Switzerland) and bacterial cultures were plated on tryptic-soy agar (TSA, Carl Roth) on Petri dishes (Greiner Bio-One, Frickenhausen, Germany) and counted the next day.
2.4. Probiotic Treatment/Co-culture
Co-culture experiments with probiotic bacterial strains were performed with 1×106 PBMCs with different preparations of either BS or BA in a total volume of 1 mL in 24-well plates (Eppendorf, Hamburg, Germany) for 24 h. All experiments were performed in RPMI 1640 medium containing 10% chicken serum without Pen/Strep at 41 °C with 5% CO2. For all co-culture experiments, concanavalin A (conA, Vector Laboratories, Newark, California, USA) was used as a positive control. PBMCs were treated at a concentration of 10 µg/mL conA (Figure S1).
2.4.1. Treatment with Vital Probiotic Bacteria
To investigate the effect of vital probiotic bacteria, 1×106 PBMCs were co-cultured with either BS or BA in a ratio of 1:3 (PBMCs:Bacillus).
2.4.2. Treatment with Supernatants of Probiotic Culture
For treatment with cell-free supernatants of the probiotic bacterial cultures, 40 mL RPMI 1640 medium was inoculated with either BS or BA, and bacteria were cultured overnight at 37 °C and 100 rpm. The next day, the bacterial cultures were sterile-filtered using a vacuum filter system with a polyethersulfone membrane possessing a pore size of 0.22 μm (Corning, Corning, NY, USA). The supernatants of the cultures were then split into two new 50 mL tubes. For thermal inactivation of the secreted factors of the bacterial strains, one 50 mL tube was heat-treated (10 min at 80 °C), and the other 50 mL tube was left untreated. Heat-treated and untreated pure RPMI 1640 medium served as a negative control. 1×106 PBMCs were exposed to either 500 µL heat-treated supernatants, untreated supernatants, heat-treated RPMI 1640 medium, or untreated RPMI 1640 medium. Treatment occurred in a total volume of 1 mL for 24 h. All experiments with cell-free bacterial culture supernatants were performed in RPMI 1640 medium containing 10% chicken serum and without Pen/Strep at 41 °C with 5% CO2. To validate that cell-free supernatants were used, bacterial culture supernatants were plated on TSA agar plates before and after sterile filtering.
2.4.3. Treatment with UV-inactivated Probiotic Bacteria
To investigate whether secreted factors or cell surface proteins could influence immune cell phenotypes, the bacteria were inactivated by UV light. Therefore, 20 mL RPMI 1640 medium was inoculated with either BS or BA and cultured overnight at 37 °C. On the next day, bacterial cultures were transferred into Petri dishes and irradiated with UV light (200 nm) for 5 h. Thereafter, the bacteria were harvested and stored at 4 °C overnight for co-culture experiments. The next day, 1×106 PBMCs were treated with either UV-inactivated BS or BA in a ratio of 2:1 (PBMCs:Bacillus) in a total volume of 1 mL for 24 h. The number of bacteria used for treatment was determined before the UV inactivation. To validate the UV inactivation, bacterial cultures were plated on TSA agar plates before and after UV light exposure.
2.5. Immunophenotyping
For immunophenotyping using flow cytometry, cells were harvested, centrifuged for 10 min at 400× g, washed once with DPBS/EDTA containing 0.05% bovine serum albumin (BSA, Sigma-Aldrich), and stained with different panels of monoclonal antibodies. In this study, CD4+ T cells were stained with mouse anti-chicken CD4-phycoerythrin (PE) or CD4-SpectralRed (SPRD) (CT-4, SouthernBiotech, Birmingham, Alabama, USA). CD8+ T cells were labeled with CD8-allophycocyanin (APC) (CT-8, ThermoFisher Scientific). In addition, human anti-chicken CD25-FITC (AbD13504, Bio-Rad Laboratories, Hercules, California, USA) and CD28-PE (AV7, SouthernBiotech) were used to investigate the activation of CD4+ T-helper cells (CD4+CD25+, CD4+CD28+) and CD8+ cytotoxic T cells (CD8+CD25+, CD8+CD28+). All CD3+ T cells were labeled with the pan-T-cell marker CD3-APC (CT-3, ThermoFisher Scientific). Bu-1+ B cells were stained with Bu-1-FITC (AV20, ThermoFisher Scientific). As a viability marker, 1.5 µL of 1 mg/mL 4′, 6-diamino-2-phenylindole (DAPI, ThermoFisher Scientific) was used. 20,000 vital PBMCs were recorded on a BD FACSCanto™ II (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) flow cytometer.
2.6. Statistical Analysis
The data obtained by flow cytometry was analyzed using a gating strategy (Figure S3). The cell count of antibody-positive cells was calculated relative to the vital PBMCs measured on a BD FACSCanto™ II. Due to high variation in the immune response capacity, the results are displayed as the ∆ relative cell count, representing the difference of the relative cell count between the treatment and the respective untreated negative control for every biological replicate. Statistical analysis was performed by a one sample Student’s t-test. All tests were performed using GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, USA). Differences between groups were considered statistically significant at p < 0.05 (* = p < 0.05, ** = p < 0.01, *** = p < 0.001, **** = p < 0.0001). A statistical tendency is shown as +, p < 0.1.
3. Results
3.1. Effects of Vital BS and BA on T and B Cells in a PBMC Composite
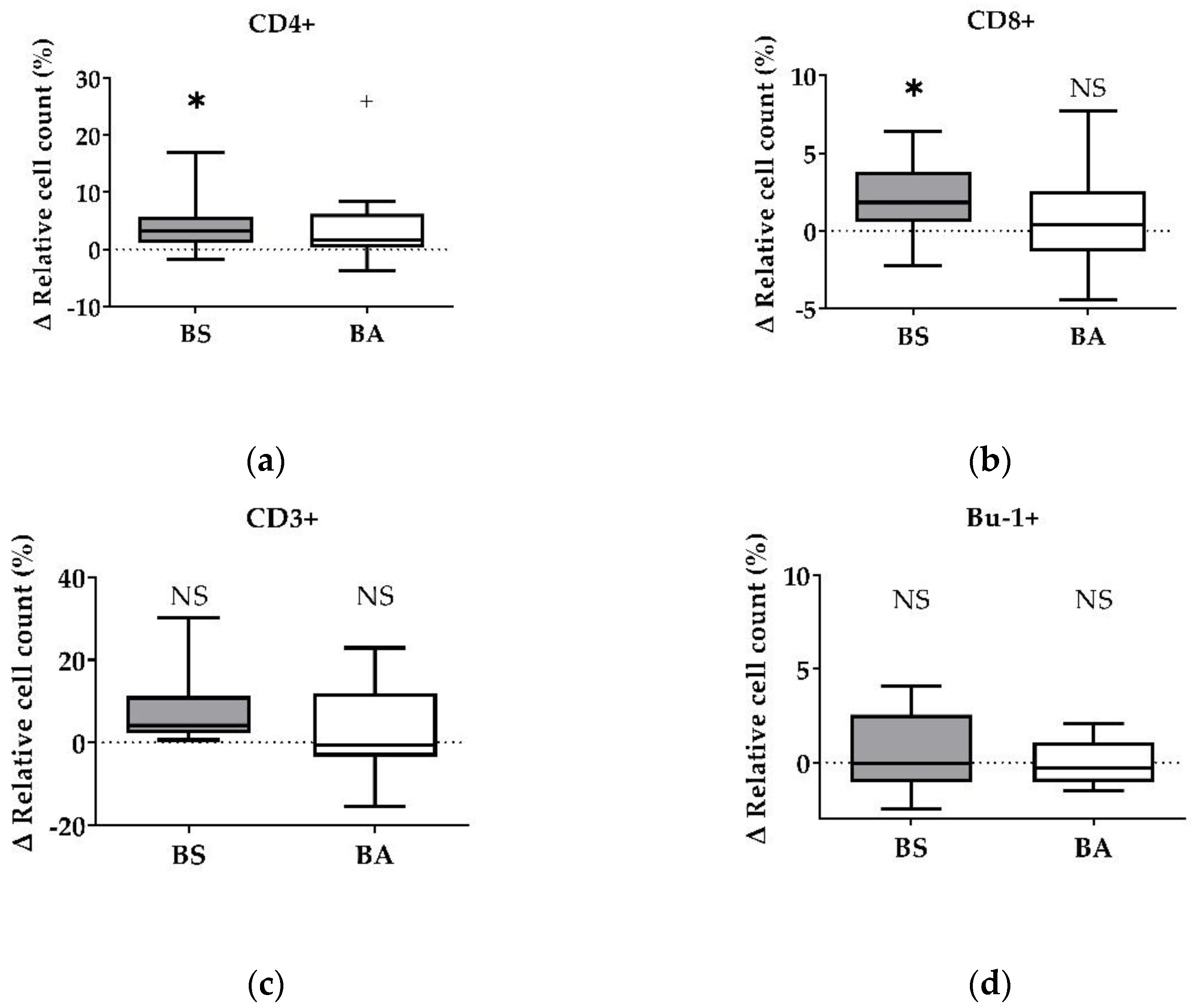
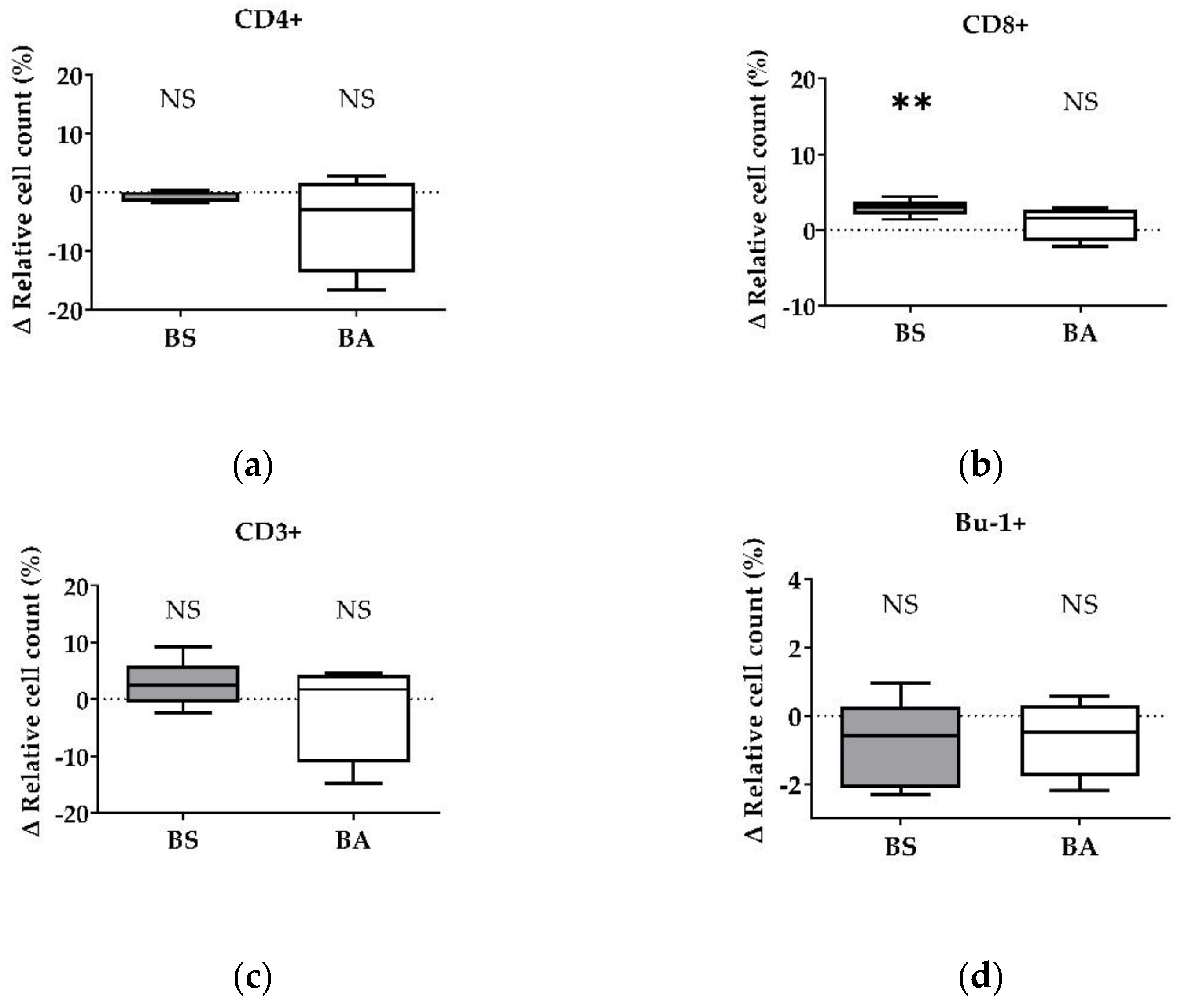
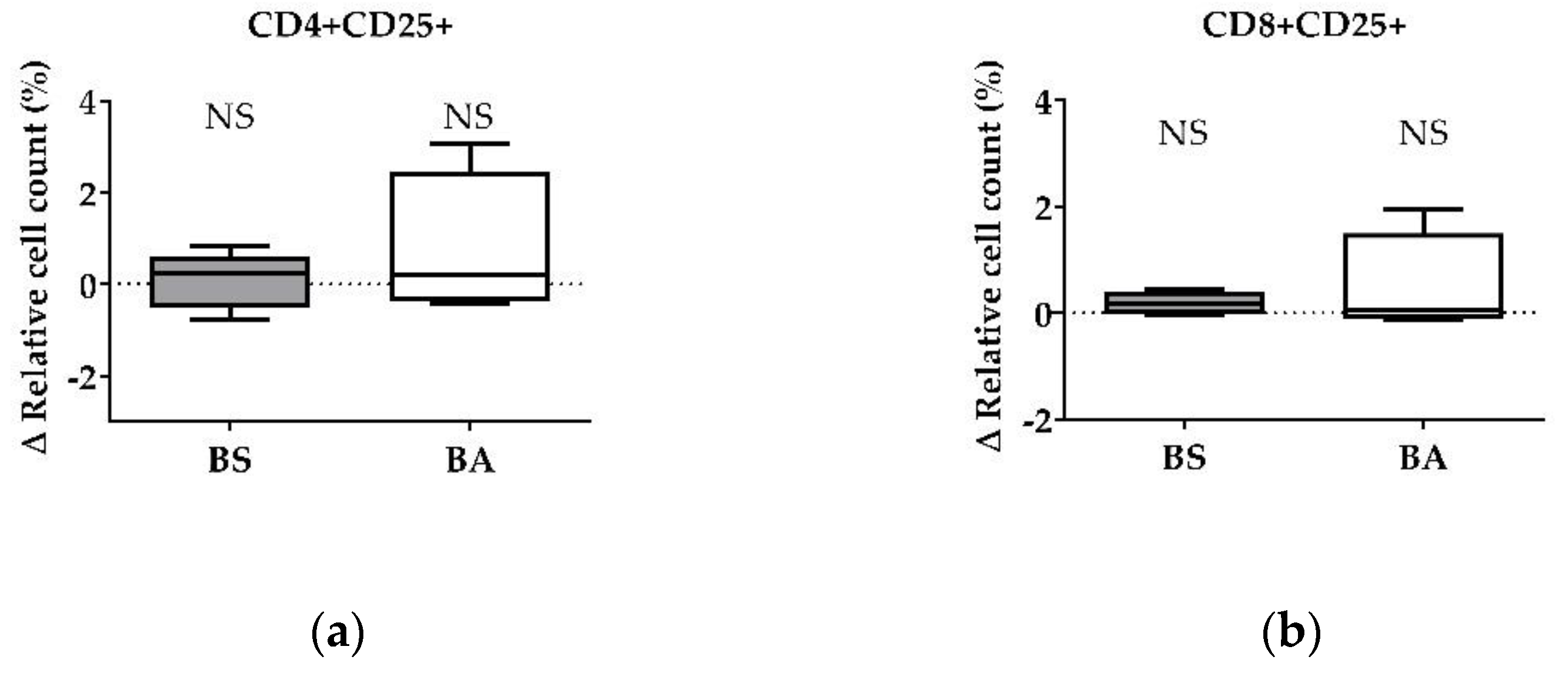
The effect of vital BS and BA on chicken adaptive immune cells was evaluated by measures of the proportions of the adaptive immune cell populations. In particular, the proliferation of CD4+ T-helper cells (Figure 1a), CD8+ cytotoxic T cells (Figure 1b), and all CD3+ T cells (Figure 1c) were measured. Furthermore, the effect on the proliferation of Bu1+ B cells was investigated (Figure 1d). Additional T cell activation markers (CD25, CD28) allowed to display the effect on CD4+CD25+ activated T-helper cells (Figure 2a), CD8+CD25+ activated cytotoxic T cells (Figure 2b), and all CD28+ αβ T cells (Figure 2c).
The treatment of PBMCs with vital BS revealed an increase of the ∆ relative cell count of CD4+ T-helper cells by 4.10% (p < 0.05, Figure 1a) and CD8+ cytotoxic T cells by 1.79% (p < 0.05, Figure 1b). Furthermore, the ∆ relative cell count of all CD3+ T cells increased by 7.83% (Figure 1c). However, this result was not significant. Additionally, treatment with vital BS had no effect on Bu-1+ B cells (Figure 1d).
After treatment with vital BA, the ∆ relative cell count of CD4+ T-helper cells increased by 2.38% (p < 0.1, Figure 1a), while the count of CD8+ cytotoxic T cells (Figure 1b) remained unaffected. The ∆ relative cell count of all CD3+ T cells increased by 2.64% (Figure 1c), however, the result was not significant. The ∆ relative cell count of Bu-1+ B cells did not change after treatment with vital BA (Figure 1d).
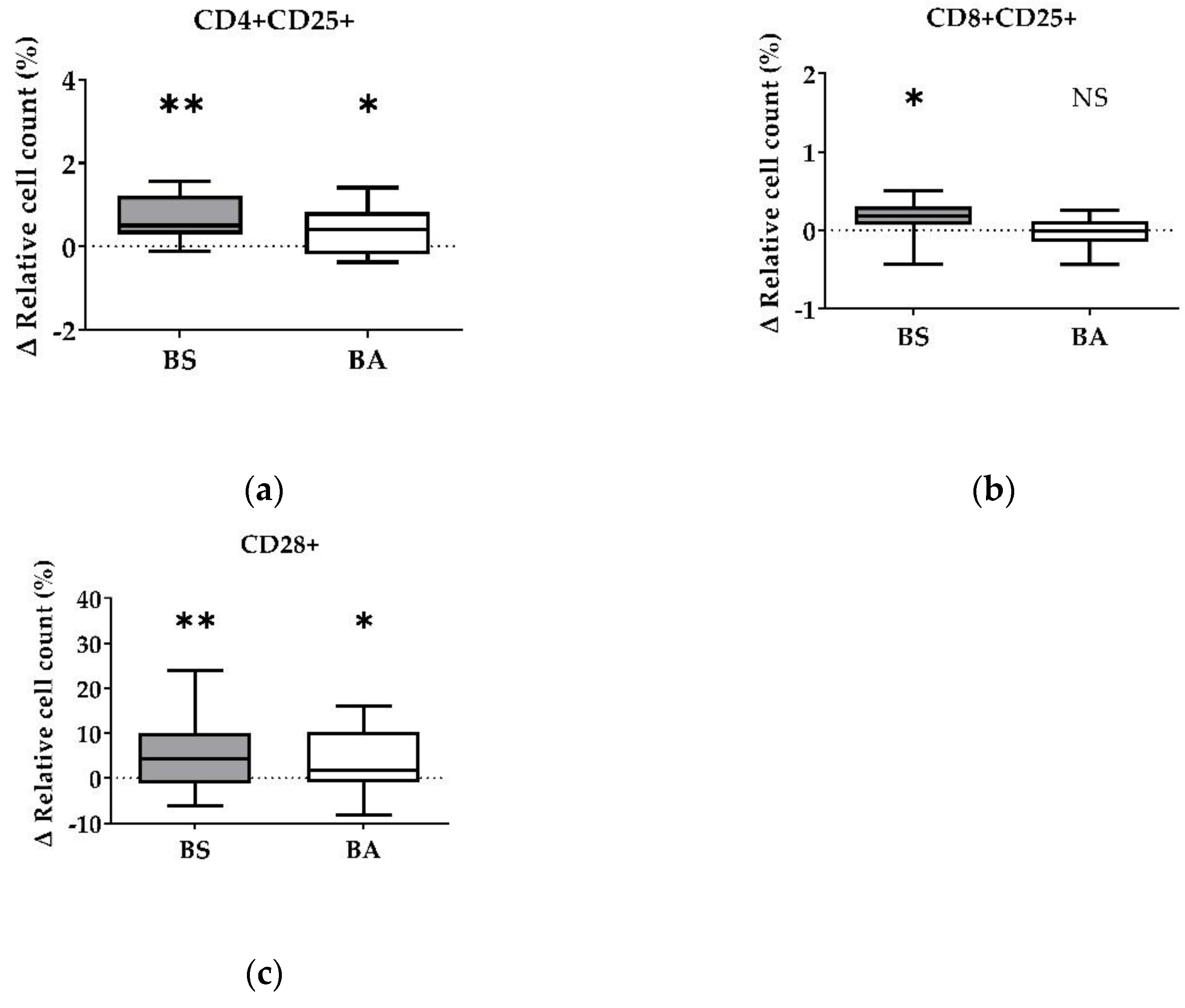
Corresponding to the increase of the ∆ relative cell count of CD4+ T-helper cells and CD8+ cytotoxic T cells, vital BS enhanced the count of CD4+CD25+ activated T-helper cells by 0.67% (p < 0.01, Figure 2a) and CD8+CD25+ activated cytotoxic T cells (p < 0.05, Figure 2b) by 0.16%. Additionally, activated CD28+ αβ T cells increased by 5.20% (p < 0.01, Figure 2c) after treatment with BS.
The addition of vital BA resulted in an elevated number of the CD4+CD25+ activated T-helper cell proportion by 0.37% (p < 0.05, Figure 2a). The ∆ relative cell count of CD8+CD25+ activated cytotoxic T cells (Figure 2b) remained unaffected after treatment with vital BA, while all CD28+ αβ T cells increased by 3.81% (p < 0.05, Figure 2c).
Summarized, vital BS stimulated the proliferation and activation of T-helper cells (CD4+, CD4+CD25+), cytotoxic T cells (CD8+, CD8+CD25+) and all αβ T cells (CD28+), while vital BA stimulated the activation and proliferation of T-helper cells (CD4+, CD4+CD25+) and all αβ T cells (CD28+).
3.2. Effects of Cell-Free Culture Supernatants of BS and BA on T and B cells in a PBMC Composite
After treatment with vital probiotic bacteria, we evaluated which component of the probiotics BS and BA is involved in the stimulation of the immune system, especially T cells. Therefore, in a first step, we analyzed the effects of cell-free bacterial culture supernatants of BS and BA on PBMCs by measuring the proliferation (Figure 3a–d) and the activation status (Figure 4a,b) of the adaptive immune cell populations, T and B cells.
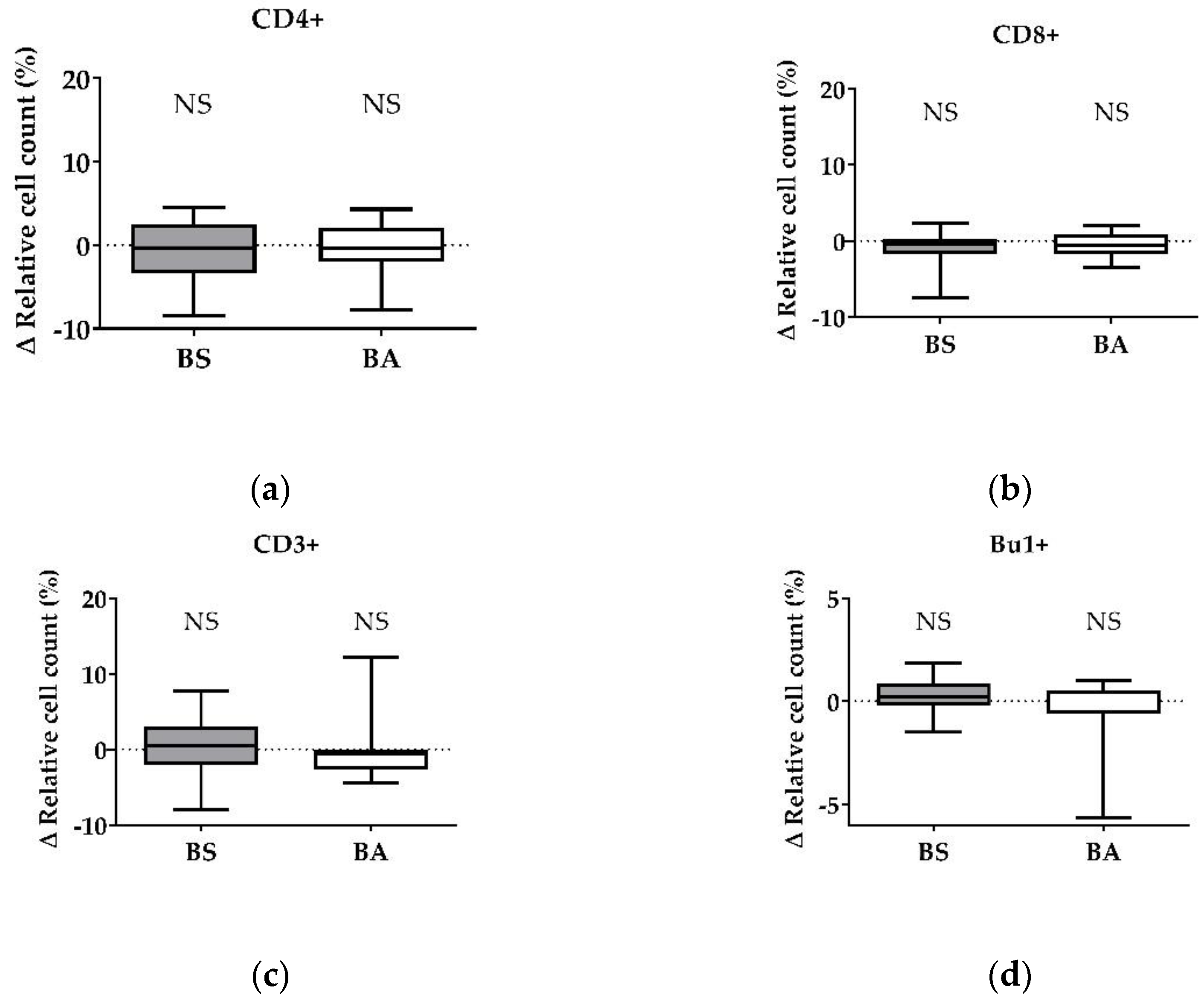
PBMC treatment with cell-free culture supernatants of the probiotic BS did not affect the CD4+ T-helper cells (Figure 3a) and CD8+ cytotoxic T cells (Figure 3b). Furthermore, the ∆ relative cell count of all CD3+ T cells (Figure 3c) and Bu-1+ B cells (Figure 3d) remained unaffected after treatment. Interestingly, we found a decreased number of CD4+ T-helper cells after treatment with the negative control, heat-treated cell-free culture supernatants of BS (Figure S2a).
After treatment of PBMCs with cell-free culture supernatants of BA, we found no effect on the Δ relative cell count of CD4+ T-helper cells (Figure 3a), CD8+ cytotoxic T cells (Figure 3b), all CD3+ T cells (Figure 3c), and Bu-1+ B cells (Figure 3d). In addition, we found a decreased count of Bu-1+ B cells after exposure to heat-treated cell-free culture supernatants of BA (Figure S2f).
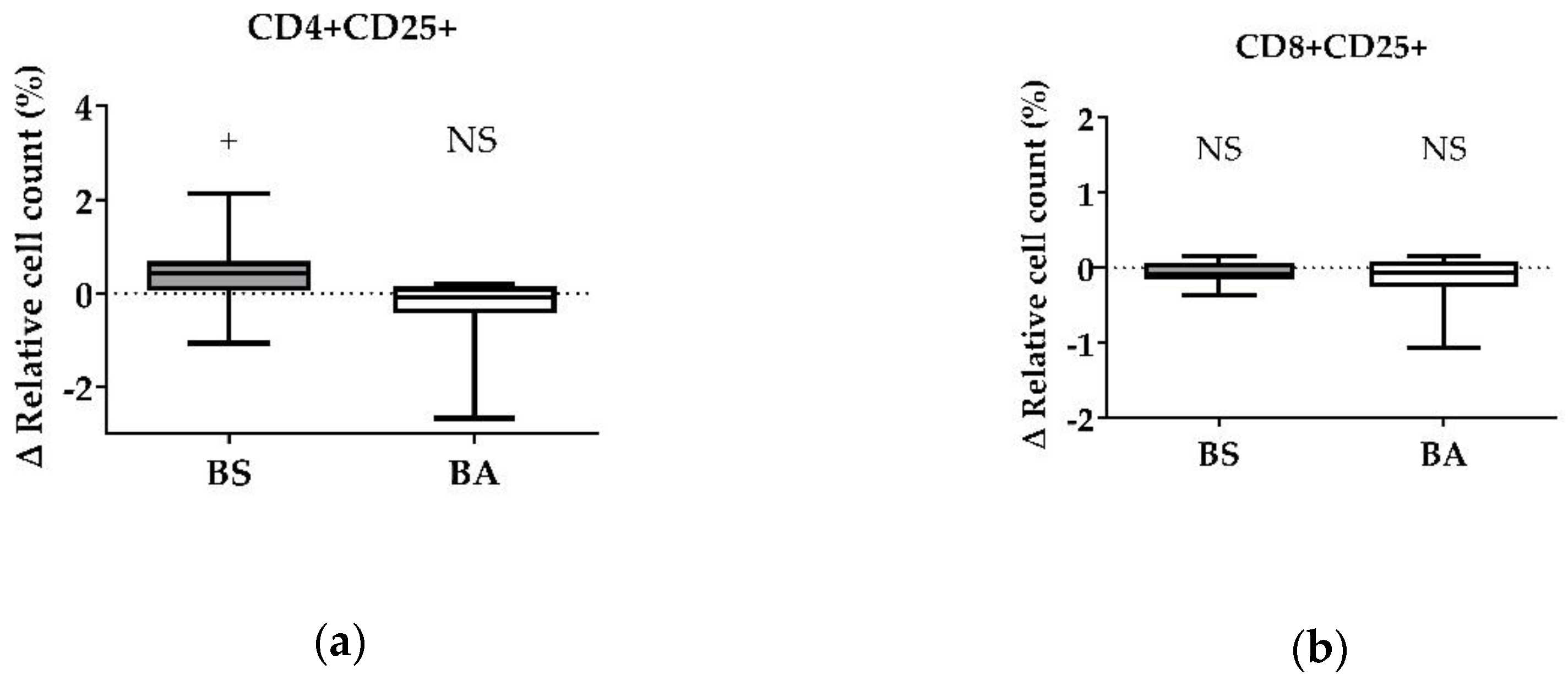
For CD4+CD25+ activated T-helper cells, the ∆ relative cell count was tendentially increased by 0.39% (p < 0.1, Figure 4a) after treatment with cell-free culture supernatants of BS, whereas CD8+CD25+ activated cytotoxic T cells (Figure 4b) were not affected.
After treatment of PBMCs with cell-free culture supernatants of BA, we found no effect on the Δ relative cell count of CD4+CD25+ activated T-helper cells (Figure 3a) and CD8+CD25+ activated cytotoxic T cells (Figure 3c,d).
Summarized, secreted factors, expected in the cell-free culture supernatants of BS, stimulated T-helper cell activation (CD4+CD25+). Furthermore, heat-treated cell-free culture supernatants, which are supposed to contain denaturated proteins, decreased the CD4+ T-helper cell count. Cell-free culture supernatants of BA had no effect on T and B cells. However, heat-treated cell-free culture supernatants decreased the count of Bu-1+ B cells.
3.3. Effects of UV-Inactivated BS and BA on T and B Cells
As we could partially confirm the involvement of secreted factors in the stimulation of T-helper cells by BS and could further exclude the involvement of secreted factors in the stimulation of cytotoxic T cells by BS and of T-helper cells by BA, we investigated the effect of the bacterial cell surface. We hypothesized that direct contact of the bacterial cell surface and the adaptive immune cells constitutes the major component of adaptive immune cell activation. To this end, the bacteria were inactivated by UV light. After UV light exposure, we assumed that the bacteria possessed an intact cell surface, as reported earlier [36], which could be involved in a possible immunomodulatory effect. The proliferation (Figure 5a–d) and the activation status (Figure 6a,b) of the adaptive immune cell populations, T and B cells, were investigated.
After the treatment of PBMCs with UV-inactivated BS, we did not observe a difference in the Δ relative cell count of CD4+ T-helper cells (Figure 5a). In contrast, treatment with UV-inactivated BS significantly increased the CD8+ cytotoxic T cell count by 2.96% (p < 0.01, Figure 5b). Furthermore, the CD3+ T cell population was enhanced after treatment with UV-inactivated BS by 2.61% (Figure 5c), although, not significantly. The Bu-1+ B cell count remained unaffected after treatment with UV-inactivated bacteria (Figure 5d).
Treatment with UV-inactivated BA did not result in a change of the Δ relative cell count of CD4+ T-helper cells (Figure 5a), CD8+ cytotoxic T cells (Figure 5b), the CD3+ T cell population (Figure 5c), and Bu-1+ B cells (Figure 5d).
We did not observe an effect on CD4+CD25+ activated T-helper cells (Figure 6a) after treatment with UV-inactivated BS. In contrast, the ∆ relative cell count of the CD8+CD25+ activated cytotoxic T cell population seemed to be increased by 0.19% (Figure 6b) after treatment with UV-inactivated BS, however, the result was not significant.
Treatment with UV-inactivated BA did not result in a change of the Δ relative cell count of CD4+CD25+ activated T-helper cells (Figure 6a) and CD8+CD25+ activated cytotoxic T cells (Figure 6b).
In summary, we observed that UV-inactivated BS increased the count of CD8+ cytotoxic T cells and seemed to stimulate cytotoxic T cell activation (CD8+CD25+). In contrast, UV-inactivated BA had no effect on CD4+ T-helper cell, CD8+ cytotoxic T cell, and Bu-1+ B cell responses in PBMCs of broiler chicken.
4. Discussion
The use of antimicrobials, especially antibiotics, in broiler production triggers the development of antimicrobial-resistant pathogenic bacteria with implications for human and animal health. Innovative feeding strategies, including the use of probiotics, have the potential to support animal health and, thus, may help to reduce the use of antimicrobials in poultry production. Proposed modes of probiotic action include immunomodulatory effects of specific strains. Primary cell culture systems represent a good alternative to animal experiments, more closely mimicking the in vivo model compared to conventional cell lines, to investigate the mechanisms of potentially immune-modulating feed additives. In this study, we investigated the immunomodulatory potential of two probiotic Bacillus strains, BS and BA, which are both commercially available probiotics for chicken farming, using an in vitro cell culture system with chicken immune cells [34].
We detected stimulating effects of both probiotic Bacillus strains on the proliferation (percentage of CD4+ T-helper cells) (Figure 1a), as well as on the activation (percentage of CD4+CD25+ activated T-helper cells) (Figure 2a) of T-helper cells. Furthermore, the count of activated CD28+ αβ T cells (Figure 2c) increased after treatment with both bacterial strains, which is in line with the increase of the CD4+ and the CD4+CD25+ activated T-helper cell proportion (Figure 1a and Figure 2a). CD28, a costimulatory molecule important for T cell activation [37], is reported to be a late activation marker (48 h) and only for CD8+ cytotoxic T cells [38]. In our study, the CD28 signal increased significantly 24 h after treatment with BS and BA. These results hint towards an in vitro stimulation of CD4+ and CD4+CD25+ activated T-helper cells through vital BS and BA bacteria. CD4+CD25+ T cells are further reported to possess properties similar to that of mammalian regulatory T cells (Tregs) [39,40]. Therefore, chicken Tregs are required to maintain immune homeostasis and self-antigen tolerance. Interestingly, the CD4+CD25+ T cell population showed no downregulation of the cytotoxic T cell activity in our study and therefore did not seem to possess suppressing properties, as it was reported earlier [41]. The proliferation (percentage of CD8+ T cells) and activation (percentage of CD8+CD25+ T cells) of cytotoxic T cells increased after treatment with vital BS (Figure 1b and Figure 2b). In another study, the applicability of chicken homologs to well-known mammalian T cell activation markers was investigated [38]. Next to CD28, CD5, MHC-II, CD44, and CD45, the marker CD25 was used to measure T cell activation. The frequencies of CD25+ T cells were increased after conA stimulation for 24 h, which demonstrated that CD25 could also be used for chicken PBMCs as an activation marker [38]. Therefore, we used CD25 as a T cell activation marker. In addition to the effect of vital bacteria, the activation of T-helper cells tended to increase after treatment with cell-free culture supernatants of BS (CD4+CD25+, Figure 4b). This result points towards an involvement of secreted factors of the bacteria, which remained in the supernatants, in the proliferation (CD4+) and activation (CD4+CD25+) of T-helper cells. Interestingly, we found a decrease in the CD4+ T-helper cell count after treatment with heat-treated cell-free culture supernatants of BS (Figure S2a). This result possibly indicates important compounds for CD4+ T-helper cell survival, which were inactivated by heat in the cell-free bacterial culture supernatants. In the experiments with vital BS, we found stimulating effects on the proliferation (CD4+) and activation (CD4+CD25+) of T-helper cells (Figure 1a and Figure 2a) as well as on the proliferation (CD8+) and activation (CD8+CD25+) of cytotoxic T cells (Figure 1b and Figure 2b). This finding is in line with observations in a previous in vivo study in chicken with probiotic Lactobacillus acidophilus LA5 [42]. Moreover, B. subtilis spores, as adjuvants, were shown to enhance CD4+ and CD8+ T cell responses in vivo against avian influenza H9N2 [43]. In a necrotic enteritis challenge, B. subtilis DSM 32315, the same bacterial strain used in our study, was demonstrated to reduce pathology and improve the performance of broilers [23]. We show that UV-inactivated BS bacteria led to an increase of CD8+ cytotoxic T cells (Figure 5b), whereas the median after treatment with cell-free supernatants of both bacterial strains rather tended to decrease (Figure 3b). This result indicates an involvement of direct contact of the cell surface of BS with the immune cells for cytotoxic T cell activation. UV irradiation can denature the DNA of microorganisms, causing death or inactivation [44], however, the cell surface of UV-inactivated bacteria was reported to remain intact [36]. Additionally, the result obtained for all CD3+ T cells after treatment with UV-inactivated BS (Figure 5c) points to the same direction regarding an effect on the T cell population. However, probably partially due to a smaller sample size in this experiment, those results were not significant. In mice, B. subtilis was shown to induce anti-inflammatory M2 macrophages inhibiting T cell-mediated immune responses [45]. From our results, we suggest that BS may improve the performance of the animals against infection. Thus, BS helps to prevent and treat disease through the modulation of T cell responses in chicken.
B cells, another important cell type of the adaptive immune response, play a major role in the antibody-mediated humoral immune response. Probiotic effects on B cells have mainly focused on antibody production, like immunoglobulin A (IgA) production in the GIT [33,46,47]. Moreover, probiotics were reported to affect B cells by altering or increasing the number of antigen-specific B cells, as it was shown after in vitro stimulation of human PBMCs with different lactic acid bacterial (LAB) strains [48] and in chicken after vaccination with inactivated avian influenza H9N2 together with a B. subtilis spore adjuvant [43]. In pigs, B cell stimulation in PBMCs was also reported after in vivo administration of probiotic Enterococcus faecium NCIMB 10415 in piglets [49]. For Bu-1+ B cells, we found no effect of both vital probiotic Bacillus strains, BS and BA (Figure 1d). Furthermore, we found no differences after treatment with cell-free supernatants (Figure 3d) and UV-inactivated bacteria (Figure 5d). However, after treatment with heat-treated cell-free culture supernatants, we found a significant decrease in the Bu-1+ B cell population (Figure S2f), possibly indicating an apoptosis-inducing effect of the denatured proteins or the loss of factors necessary for B cell survival.
This study provides evidence of a direct immunomodulatory effect of BS and BA on chicken T cells. Treatment with BS increased the proliferation (CD8+) and activation (CD8+CD25+) of cytotoxic T cells, which may enhance the cellular immune response, in particular by destroying pathogens through perforins and granzymes or inducing apoptosis of infected host cells [50]. Furthermore, treatment with both probiotic bacteria, BS and BA, increased the proliferation (CD4+) and activation (CD4+CD25+) of T-helper cells. However, the CD4+CD25+ T cell population could further represent Tregs, regulating and suppressing excessive T-helper cell 1 (Th1) and T-helper cell 2 (Th2) immune responses and maintaining immunological unresponsiveness and tolerance to self-antigens [40,50,51].
5. Conclusions
Our results point towards a possible involvement of secreted factors of BS in T-helper cell activation and proliferation, whereas BS stimulated cytotoxic T cells presumably through surface contact. For BA, more experiments are necessary to unravel the mechanism behind the action on T cells. All in all, we observed a T cell stimulation by both tested probiotic Bacillus strains in vitro. Therefore, we suggest that these probiotic Bacillus strains could improve the health of animals and their defense against infection and, thus, help to prevent and reduce the usage of antimicrobials in chicken farming.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11020269/s1, Figure S1: Influence of conA on activation and proliferation of T cells and B cells; Figure S2: Influence of heat-treated cell-free culture supernatants of B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) activation and proliferation of T and B cells; Figure S3: Example of the gating strategy for chicken PBMCs.
Author Contributions
Conceptualization, G.A.B. and S.K.-R.; data curation, F.L.; formal analysis, F.L.; investigation, F.L. and M.S.; supervision, D.H., G.A.B. and S.K.-R.; writing—original draft, F.L.; writing—review and editing, D.H., G.A.B., S.K.-R., G.L. and M.S. All authors will be informed about each step of manuscript processing including submission, revision, revision reminder, etc. via emails from our system or assigned assistant editor. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Bundesministerium für Bildung und Forschung (BMBF, Federal Ministry of Education and Research, Funding number: 031B0743).
Data Availability Statement
Data supporting reported results will be provided upon request.
Acknowledgments
The article processing charge was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)–491192747 and the Open Access Publication Fund of Humboldt-Universität zu Berlin. We thank Evonik Operations GmbH for providing the bacterial strains. Michael Modrow, Annett Kannegießer, and Jens Fuchs for taking care of the animals and collecting samples.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- OECD; Food, Agriculture Organization of the United Nations. OECD-FAO Agricultural Outlook 2022–2031; OECD: Paris, France, 2022. [Google Scholar]
- Kumar, A.; Patyal, A. Impacts of intensive poultry farming on ‘one health’ in developing countries: Challenges and remedies. Explor. Anim. Med. Res. 2020, 10, 100–111. [Google Scholar]
- Sato, Y.; Kuroki, Y.; Oka, K.; Takahashi, M.; Rao, S.; Sukegawa, S.; Fujimura, T. Effects of Dietary Supplementation With Enterococcus faecium and Clostridium butyricum, Either Alone or in Combination, on Growth and Fecal Microbiota Composition of Post-weaning Pigs at a Commercial Farm. Front. Vet. Sci. 2019, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Elgeddawy, S.A.; Shaheen, H.M.; El-Sayed, Y.S.; Abd Elaziz, M.; Darwish, A.; Samak, D.; Batiha, G.E.; Mady, R.A.; Bin-Jumah, M.; Allam, A.A.; et al. Effects of the dietary inclusion of a probiotic or prebiotic on florfenicol pharmacokinetic profile in broiler chicken. J. Anim. Physiol. Anim. Nutr. 2020, 104, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Alloui, M.N.; Szczurek, W.; Świątkiewicz, S. The Usefulness of Prebiotics and Probiotics in Modern Poultry Nutrition: A Review / Przydatność prebiotyków i probiotyków w nowoczesnym żywieniu drobiu—przegląd. Ann. Anim. Sci. 2013, 13, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Sławinska, A.; Siwek, M.Z.; Bednarczyk, M.F. Effects of synbiotics injected in ovo on regulation of immune-related gene expression in adult chickens. Am. J. Vet. Res. 2014, 75, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Qiu, K.; Zhang, H.-J.; Wu, S.-G.; Han, Y.-M.; Wu, Y.-Y.; Qi, G.-H.; Wang, J. Organic Acids as Alternatives for Antibiotic Growth Promoters Alter the Intestinal Structure and Microbiota and Improve the Growth Performance in Broilers. Front. Microbiol 2021, 11, 618144. [Google Scholar] [CrossRef]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (Direct-Fed Microbials) in Poultry Nutrition and Their Effects on Nutrient Utilization, Growth and Laying Performance, and Gut Health: A Systematic Review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2020, 11, 1054. [Google Scholar] [CrossRef] [Green Version]
- Rathnapraba, S.; Kanagaraju, P.; Vijayarani, K. Effect of dietary supplementation of probiotic and BMD on the growth performance of broiler chickens challenged with Clostridium perfringens induced necrotic enteritis. Int. J. Chem. Stud. 2018, 6, 13–15. [Google Scholar]
- Sen, S.; Ingale, S.L.; Kim, Y.W.; Kim, J.S.; Kim, K.H.; Lohakare, J.D.; Kim, E.K.; Kim, H.S.; Ryu, M.H.; Kwon, I.K.; et al. Effect of supplementation of Bacillus subtilis LS 1-2 to broiler diets on growth performance, nutrient retention, caecal microbiology and small intestinal morphology. Res. Vet. Sci. 2012, 93, 264–268. [Google Scholar] [CrossRef]
- Hossain, M.M.; Begum, M.; Kim, I.H. Effect of Bacillus subtilis, Clostridium butyricum and Lactobacillus acidophilus endospores on growth performance, nutrient digestibility, meat quality, relative organ weight, microbial shedding and excreta noxious gas emission in broilers. Vet. Med. Czech. 2015, 60, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.P.; Wu, A.M.; Ding, X.M.; Lei, Y.; Bai, J.; Zhang, K.Y.; Chio, J.S. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Hart, A.L.; Kamm, M.A.; Stagg, A.J.; Knight, S.C. Mechanisms of action of probiotics: Recent advances. Inflamm. Bowel Dis. 2009, 15, 300–310. [Google Scholar] [CrossRef]
- Erickson, K.L.; Hubbard, N.E. Probiotic immunomodulation in health and disease. J. Nutr. 2000, 130, 403S–409S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, F.; Akram, K.; Saleh Alghamdi, E.; Arshad, Q.; Siddique, A. The immunomodulatory role of probiotics. In Prebiotics and Probiotics—From Food to Health; Franco-Robles, E., Ed.; IntechOpen: London, UK, 2022. [Google Scholar]
- Ogbuewu, I.P.; Mabelebele, M.; Sebola, N.A.; Mbajiorgu, C. Bacillus Probiotics as Alternatives to In-feed Antibiotics and Its Influence on Growth, Serum Chemistry, Antioxidant Status, Intestinal Histomorphology, and Lesion Scores in Disease-Challenged Broiler Chickens. Front. Vet. Sci. 2022, 9, 876725. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Penaloza-Vazquez, A.; Ma, L.M.; Rayas-Duarte, P. Isolation and characterization of Bacillus spp. strains as potential probiotics for poultry. Can. J. Microbiol. 2019, 65, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Gharib-Naseri, K.; de Paula Dorigam, J.C.; Doranalli, K.; Kheravii, S.; Swick, R.A.; Choct, M.; Wu, S.-B. Modulations of genes related to gut integrity, apoptosis, and immunity underlie the beneficial effects of Bacillus amyloliquefaciens CECT 5940 in broilers fed diets with different protein levels in a necrotic enteritis challenge model. J. Anim. Sci. Biotechnol. 2020, 11, 104. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Serpa Vieira, B.; de Paula Dorigam, J.C.; Menconi, A.; Sokale, A.; Doranalli, K.; Applegate, T.J. Bacillus subtilis DSM 32315 Supplementation Attenuates the Effects of Clostridium perfringens Challenge on the Growth Performance and Intestinal Microbiota of Broiler Chickens. Microorganisms 2019, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Piao, X.; Ru, Y.; Zhang, H.; Péron, A.; Zhang, H. Effect of Bacillus amyloliquefaciens-based Direct-fed Microbial on Performance, Nutrient Utilization, Intestinal Morphology and Cecal Microflora in Broiler Chickens. Asian-Australas. J. Anim. Sci. 2015, 28, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Whelan, R.A.; Doranalli, K.; Rinttilä, T.; Vienola, K.; Jurgens, G.; Apajalahti, J. The impact of Bacillus subtilis DSM 32315 on the pathology, performance, and intestinal microbiome of broiler chickens in a necrotic enteritis challenge. Poult. Sci. 2019, 98, 3450–3463. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.J.K.; de Sakomura, N.K.; de Paula Dorigam, J.C.; Doranalli, K.; Soares, L.; Da Viana, G.S. Bacillus amyloliquefaciens CECT 5940 alone or in combination with antibiotic growth promoters improves performance in broilers under enteric pathogen challenge. Poult. Sci. 2019, 98, 4391–4400. [Google Scholar] [CrossRef] [PubMed]
- Bodinga, B.M.; Hayat, K.; Liu, X.; Zhou, J.; Yang, X.; Ismaila, A.; Soomro, R.N.; Ren, Z.; Zhang, W.; Yang, X. Effects of Bacillus subtilis DSM 32315 on Immunity, Nutrient Transporters and Functional Diversity of Cecal Microbiome of Broiler Chickens in Necrotic Enteritis Challenge. J. World’s Poult. Res. 2020, 10, 527–544. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.; Liu, M.; Li, J.; Lai, W.; Geng, S.; Yuan, T.; Liu, Y.; Di, Y.; Zhang, W.; et al. Effects of dietary Bacillus amyloliquefaciens CECT 5940 supplementation on growth performance, antioxidant status, immunity, and digestive enzyme activity of broilers fed corn-wheat-soybean meal diets. Poult. Sci. 2022, 101, 101585. [Google Scholar] [CrossRef]
- Tregaskes, C.A.; Bumstead, N.; Davison, T.F.; Young, J.R. Chicken B-cell marker chB6 (Bu-1) is a highly glycosylated protein of unique structure. Immunogenetics 1996, 44, 212–217. [Google Scholar] [CrossRef]
- Chen, C.H.; Göbel, T.W.; Kubota, T.; Cooper, M.D. T cell development in the chicken. Poult. Sci. 1994, 73, 1012–1018. [Google Scholar] [CrossRef]
- Pieper, J.; Methner, U.; Berndt, A. Heterogeneity of avian gammadelta T cells. Vet. Immunol. Immunopathol. 2008, 124, 241–252. [Google Scholar] [CrossRef]
- Arstila, T.P.; Vainio, O.; Lassila, O. Central role of CD4+ T cells in avian immune response. Poult. Sci. 1994, 73, 1019–1026. [Google Scholar] [CrossRef]
- Seliger, C.; Schaerer, B.; Kohn, M.; Pendl, H.; Weigend, S.; Kaspers, B.; Härtle, S. A rapid high-precision flow cytometry based technique for total white blood cell counting in chickens. Vet. Immunol. Immunopathol. 2012, 145, 86–99. [Google Scholar] [CrossRef]
- Azad, M.A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. Biomed. Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsberg, F.; Sprechert, M.; Hesse, D.; Brockmann, G.A.; Kreuzer-Redmer, S. Chicken Immune Cell Assay to Model Adaptive Immune Responses In Vitro. Animals 2021, 11, 3600. [Google Scholar] [CrossRef] [PubMed]
- Jergović, M.; Nedeljković, G.; Cvetić, Ž.; Gottstein, Ž.; Bendelja, K. Combined dextran and ficoll separation yields pure populations of chicken peripheral blood mononuclear cells—short communication. Vet. Arhiv. 2017, 87, 623–631. [Google Scholar] [CrossRef]
- Weigel, K.M.; Nguyen, F.K.; Kearney, M.R.; Meschke, J.S.; Cangelosi, G.A. Molecular Viability Testing of UV-Inactivated Bacteria. Appl. Environ. Microbiol. 2017, 83, e00331-17. [Google Scholar] [CrossRef] [Green Version]
- Arstila, T.P.; Vainio, O.; Lassila, O. Evolutionarily conserved function of CD28 in αβ T cell activation. Scand. J. Immunol. 1994, 40, 368–371. [Google Scholar] [CrossRef]
- Naghizadeh, M.; Hatamzade, N.; Larsen, F.T.; Kjaerup, R.B.; Wattrang, E.; Dalgaard, T.S. Kinetics of activation marker expression after in vitro polyclonal stimulation of chicken peripheral T cells. Cytometry A 2022, 101, 45–56. [Google Scholar] [CrossRef]
- Lee, I.K.; Gu, M.J.; Ko, K.H.; Bae, S.; Kim, G.; Jin, G.-D.; Kim, E.B.; Kong, Y.-Y.; Park, T.S.; Park, B.-C.; et al. Regulation of CD4+CD8-CD25+ and CD4+CD8+CD25+ T cells by gut microbiota in chicken. Sci. Rep. 2018, 8, 8627. [Google Scholar] [CrossRef] [Green Version]
- Shanmugasundaram, R.; Selvaraj, R.K. Regulatory T cell properties of chicken CD4+CD25+ cells. J. Immunol. 2011, 186, 1997–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Zheng, G.; Zhou, D.; Huang, L.; Dong, J.; Cheng, Z. High-frequency and activation of CD4+CD25+ T cells maintain persistent immunotolerance induced by congenital ALV-J infection. Vet. Res. 2021, 52, 119. [Google Scholar] [CrossRef]
- Asgari, F.; Madjd, Z.; Falak, R.; Bahar, M.A.; Nasrabadi, M.H.; Raiani, M.; Shekarabi, M. Probiotic feeding affects T cell populations in blood and lymphoid organs in chickens. Benef. Microbes 2016, 7, 669–675. [Google Scholar] [CrossRef]
- Lee, J.E.; Kye, Y.-C.; Park, S.-M.; Shim, B.-S.; Yoo, S.; Hwang, E.; Kim, H.; Kim, S.-J.; Han, S.H.; Park, T.S.; et al. Bacillus subtilis spores as adjuvants against avian influenza H9N2 induce antigen-specific antibody and T cell responses in White Leghorn chickens. Vet. Res. 2020, 51, 68. [Google Scholar] [CrossRef] [PubMed]
- Summerfelt, S.T. Ozonation and UV irradiation—an introduction and examples of current applications. Aquac. Eng. 2003, 28, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Paynich, M.L.; Jones-Burrage, S.E.; Knight, K.L. Exopolysaccharide from Bacillus subtilis Induces Anti-Inflammatory M2 Macrophages That Prevent T Cell-Mediated Disease. J. Immunol. 2017, 198, 2689–2698. [Google Scholar] [CrossRef]
- Kemgang, T.S.; Kapila, S.; Shanmugam, V.P.; Kapila, R. Cross-talk between probiotic lactobacilli and host immune system. J. Appl. Microbiol. 2014, 117, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, M.; Hitaka, A.; Fujino, H.; Matsumoto, T.; Hasegawa, T.; Morimatsu, F.; Fujiki, T.; Katakura, Y. Identification of Immunopotentiating Lactic Acid Bacteria that Induce Antibody Production by in vitro Stimulated Human Peripheral Blood Mononuclear Cells. Biosci. Microbiota Food Health 2012, 31, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuzer, S.; Machnowska, P.; Aßmus, J.; Sieber, M.; Pieper, R.; Schmidt, M.F.; Brockmann, G.A.; Scharek-Tedin, L.; Johne, R. Feeding of the probiotic bacterium Enterococcus faecium NCIMB 10415 differentially affects shedding of enteric viruses in pigs. Vet. Res. 2012, 43, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef]
Figure 1.
Influence of vital B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with BS or BA in a ratio of 1:3 (PBMCs:Bacillus) or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent the following numbers of biological replicates: 11 (BS, BA) for CD4+, 13 (BS), and 11 (BA) for CD8+, 6 (BS), and 9 (BA) for CD3+ and Bu-1+. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as +, p < 0.1; *, p < 0.05; NS: not significant.
Figure 1.
Influence of vital B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with BS or BA in a ratio of 1:3 (PBMCs:Bacillus) or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent the following numbers of biological replicates: 11 (BS, BA) for CD4+, 13 (BS), and 11 (BA) for CD8+, 6 (BS), and 9 (BA) for CD3+ and Bu-1+. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as +, p < 0.1; *, p < 0.05; NS: not significant.

Figure 2.
Influence of vital B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on activation of T cells. Isolated PBMCs were treated with BS or BA in a ratio of 1:3 (PBMCs: Bacillus), or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells, (b) CD8+CD25+ activated cytotoxic T cells, (c) activated CD28+ αβ T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent the following numbers of biological replicates: 11 (BS, BA) for CD4+CD25+, 13 (BS) and 11 (BA) for CD8+CD25+, and 24 (BS) and 22 (BA) for CD28+. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as *, p < 0.05; **, p < 0.01; NS: not significant.
Figure 2.
Influence of vital B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on activation of T cells. Isolated PBMCs were treated with BS or BA in a ratio of 1:3 (PBMCs: Bacillus), or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells, (b) CD8+CD25+ activated cytotoxic T cells, (c) activated CD28+ αβ T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent the following numbers of biological replicates: 11 (BS, BA) for CD4+CD25+, 13 (BS) and 11 (BA) for CD8+CD25+, and 24 (BS) and 22 (BA) for CD28+. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as *, p < 0.05; **, p < 0.01; NS: not significant.

Figure 3.
Influence of cell-free culture supernatants of B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with cell-free supernatants of BS or BA, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to heat-treated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 13 (BS) or 12 (BA) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as NS: not significant.
Figure 3.
Influence of cell-free culture supernatants of B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with cell-free supernatants of BS or BA, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to heat-treated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 13 (BS) or 12 (BA) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as NS: not significant.

Figure 4.
Influence of cell-free culture supernatants of B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the activation of T cells. Isolated PBMCs were treated with either cell-free supernatants of BS or BA, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to heat-treated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells and (b) CD8+CD25+ activated cytotoxic T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 13 (BS) or 12 (BA) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as +, p < 0.1; NS: not significant.
Figure 4.
Influence of cell-free culture supernatants of B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the activation of T cells. Isolated PBMCs were treated with either cell-free supernatants of BS or BA, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to heat-treated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells and (b) CD8+CD25+ activated cytotoxic T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 13 (BS) or 12 (BA) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and minimum to maximum as the whiskers. Significance is shown as +, p < 0.1; NS: not significant.

Figure 5.
Influence of UV-inactivated B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with either UV-inactivated BA or BS or left untreated (negative control). After treatment for 24 h, immune cells were labeled with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 4 (BA) or 5 (BS) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and min to max as the whiskers. Significance is shown as **, p < 0.01; NS: not significant.
Figure 5.
Influence of UV-inactivated B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the proliferation of T and B cells. Isolated PBMCs were treated with either UV-inactivated BA or BS or left untreated (negative control). After treatment for 24 h, immune cells were labeled with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+ T-helper cells, (b) CD8+ cytotoxic T cells, (c) all CD3+ T cells, and (d) Bu-1+ B cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 4 (BA) or 5 (BS) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and min to max as the whiskers. Significance is shown as **, p < 0.01; NS: not significant.

Figure 6.
Influence of UV-inactivated B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the activation of T cells. Isolated PBMCs were treated with either UV-inactivated BA or BS, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells and (b) CD8+CD25+ activated cytotoxic T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 4 (BA) or 5 (BS) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and min to max as the whiskers. Significance is shown as NS: not significant.
Figure 6.
Influence of UV-inactivated B. subtilis DSM 32315 (grey) and B. amyloliquefaciens CECT 5940 (white) on the activation of T cells. Isolated PBMCs were treated with either UV-inactivated BA or BS, or left untreated (negative control). After treatment for 24 h, immune cells were stained with monoclonal antibodies. The relative cell count was related to untreated controls. DAPI was used as a viability marker. (a) CD4+CD25+ activated T-helper cells and (b) CD8+CD25+ activated cytotoxic T cells relative to the vital PBMC cell count. 20,000 vital PBMCs were recorded on a BD FACSCanto II flow cytometer. Data represent 4 (BA) or 5 (BS) biological replicates. Results are presented as box and whisker plots showing the median, with a 25–75 percentile range as the box and min to max as the whiskers. Significance is shown as NS: not significant.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Larsberg, F.; Sprechert, M.; Hesse, D.; Loh, G.; Brockmann, G.A.; Kreuzer-Redmer, S. Probiotic Bacillus Strains Enhance T Cell Responses in Chicken. Microorganisms 2023, 11, 269. https://doi.org/10.3390/microorganisms11020269
AMA Style
Larsberg F, Sprechert M, Hesse D, Loh G, Brockmann GA, Kreuzer-Redmer S. Probiotic Bacillus Strains Enhance T Cell Responses in Chicken. Microorganisms. 2023; 11(2):269. https://doi.org/10.3390/microorganisms11020269
Chicago/Turabian StyleLarsberg, Filip, Maximilian Sprechert, Deike Hesse, Gunnar Loh, Gudrun A. Brockmann, and Susanne Kreuzer-Redmer. 2023. "Probiotic Bacillus Strains Enhance T Cell Responses in Chicken" Microorganisms 11, no. 2: 269. https://doi.org/10.3390/microorganisms11020269
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.