
Genomic Island-Encoded Diguanylate Cyclase from Vibrio alginolyticus Regulates Biofilm Formation and Motility in Pseudoalteromonas
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Plasmid Construction
2.3. Conjugation Assays
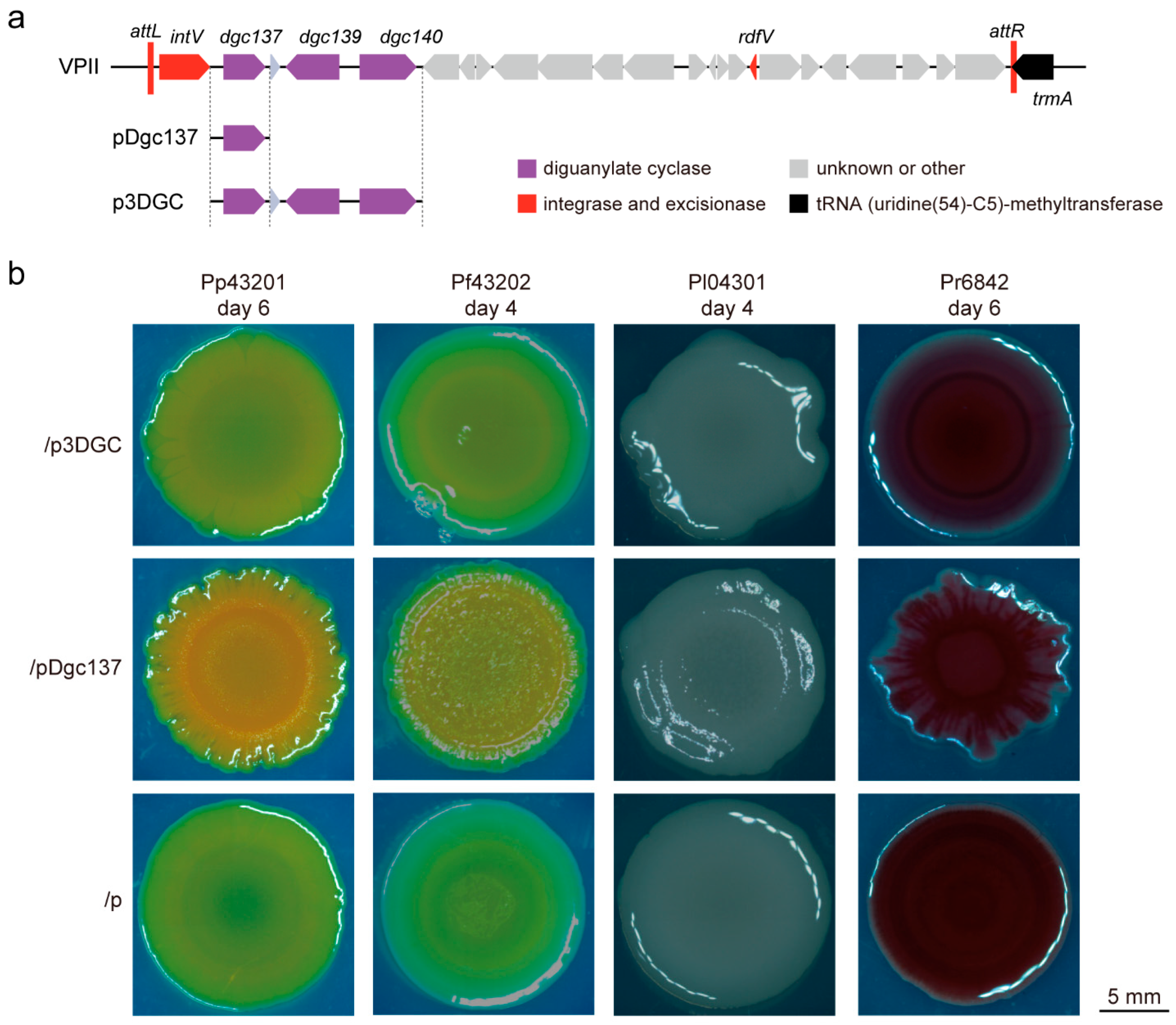
2.4. Observation of Colony Morphology
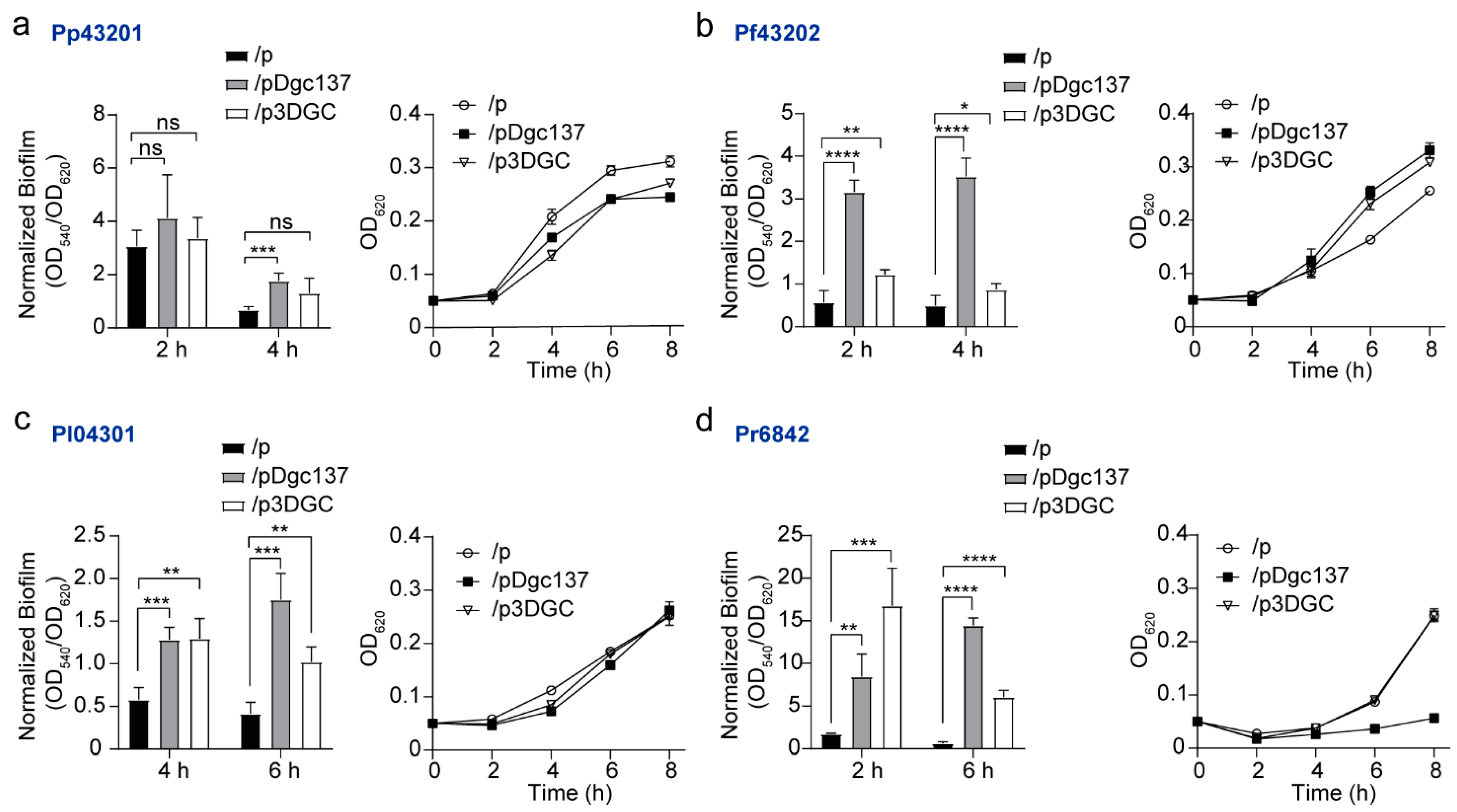
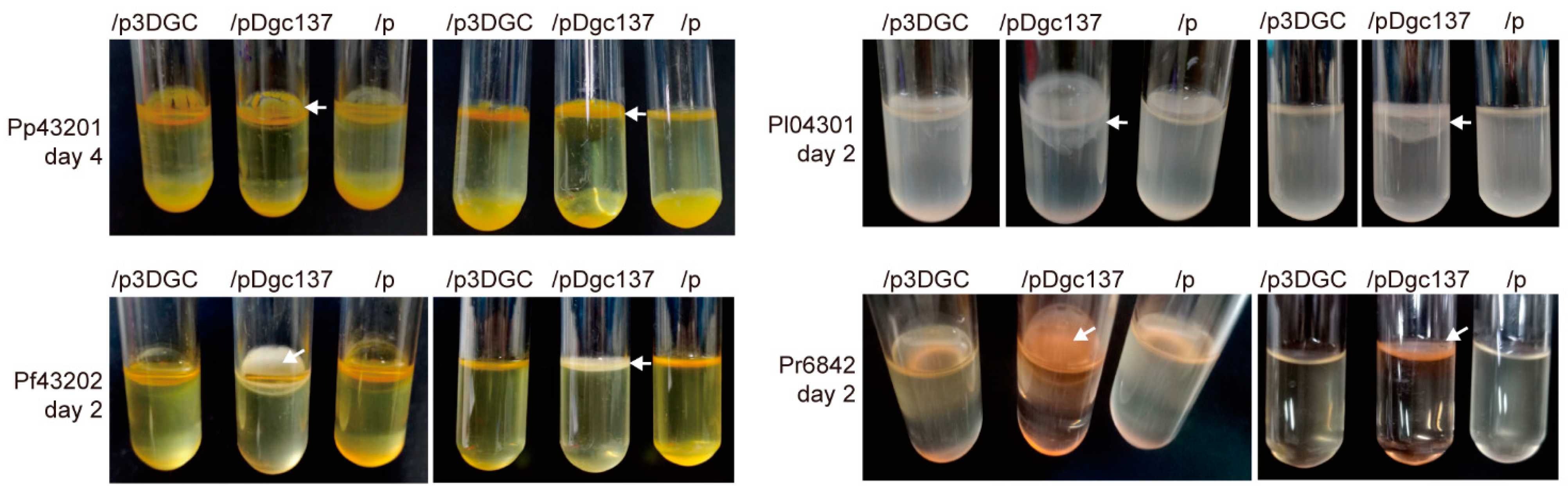
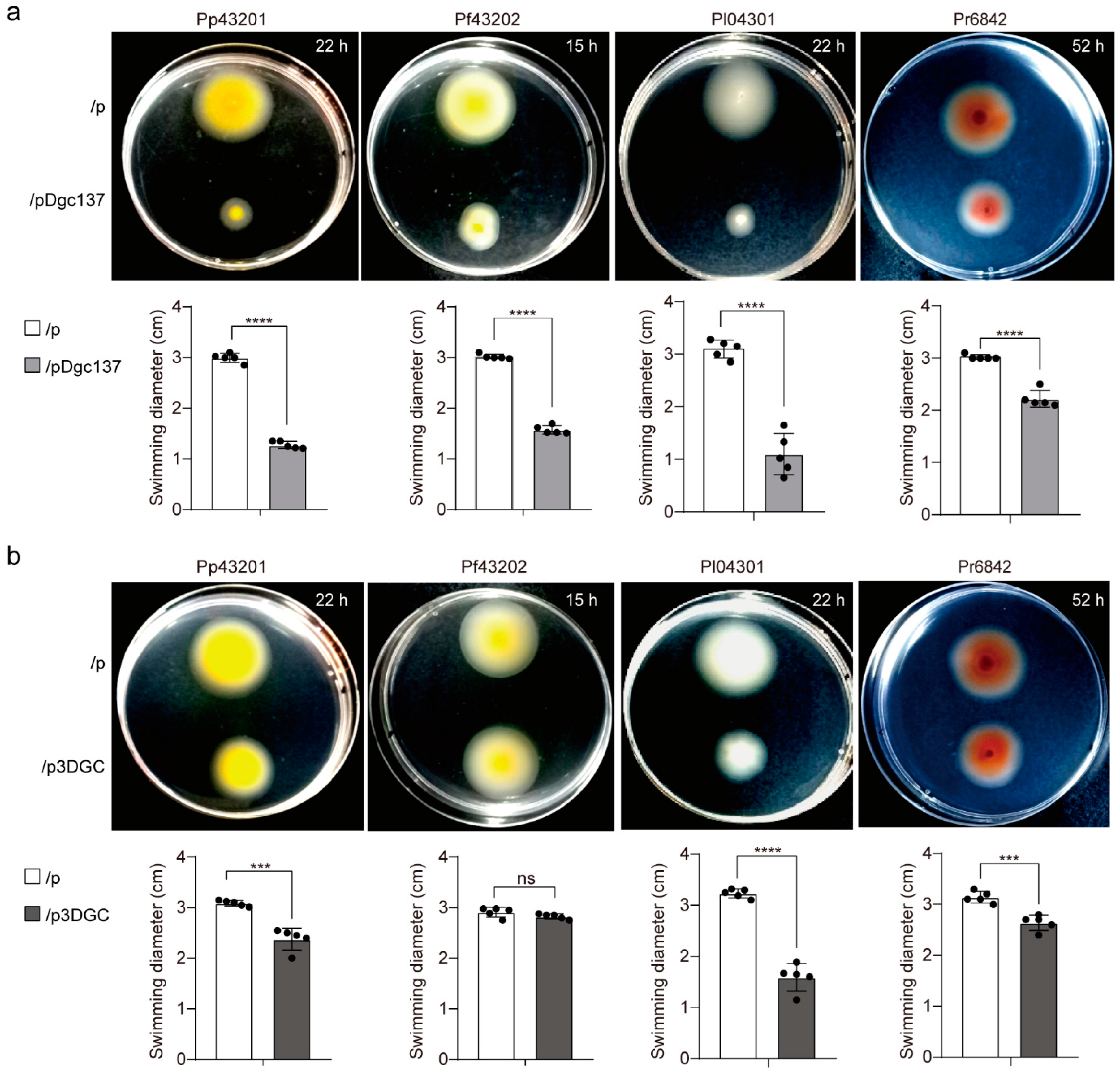
2.5. Motility and Biofilm Assays
2.6. Prediction of GGDEF Domain-Containing Proteins
2.7. Statistics and Reproducibility
3. Results
3.1. Dgc137 Regulates the Colony Biofilm of Pseudoalteromonas
3.2. Dgc137 Increases the Attached Biofilm of Pseudoalteromonas
3.3. Dgc137 Increases the Pellicle of Pseudoalteromonas
3.4. Dgc137 Decreases the Motility of Pseudoalteromonas
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brito, I.L. Examining horizontal gene transfer in microbial communities. Nat. Rev. Microbiol. 2021, 19, 442–453. [Google Scholar] [CrossRef]
- Horne, T.; Orr, V.T.; Hall, J.P.J. How do interactions between mobile genetic elements affect horizontal gene transfer? Curr. Opin. Microbiol. 2023, 73, 102282. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2022, 20, 206–218. [Google Scholar] [CrossRef]
- Webster, N.S.; Reusch, T.B.H. Microbial contributions to the persistence of coral reefs. ISME J. 2017, 11, 2167–2174. [Google Scholar] [CrossRef]
- Bioteau, A.; Durand, R.; Burrus, V. Redefinition and Unification of the SXT/R391 Family of Integrative and Conjugative Elements. Appl. Environ. Microbiol. 2018, 84, e00485-18. [Google Scholar] [CrossRef]
- Daccord, A.; Ceccarelli, D.; Rodrigue, S.; Burrus, V. Comparative analysis of mobilizable genomic islands. J. Bacteriol. 2013, 195, 606–614. [Google Scholar] [CrossRef]
- Wang, P.; Zeng, Z.; Wang, W.; Wen, Z.; Li, J.; Wang, X. Dissemination and loss of a biofilm-related genomic island in marine Pseudoalteromonas mediated by integrative and conjugative elements. Environ. Microbiol. 2017, 19, 4620–4637. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, Y.; Wang, W.; Lin, S.; Tang, K.; Liu, T.; Wood, T.K.; Wang, X. Mobile genetic elements used by competing coral microbial populations increase genomic plasticity. ISME J. 2022, 16, 2220–2229. [Google Scholar] [CrossRef]
- Tang, B.L.; Yang, J.; Chen, X.L.; Wang, P.; Zhao, H.L.; Su, H.N.; Li, C.Y.; Yu, Y.; Zhong, S.; Wang, L.; et al. A predator-prey interaction between a marine Pseudoalteromonas sp. and Gram-positive bacteria. Nat. Commun. 2020, 11, 285. [Google Scholar] [CrossRef]
- Luo, P.; He, X.; Wang, Y.; Liu, Q.; Hu, C. Comparative genomic analysis of six new-found integrative conjugative elements (ICEs) in Vibrio alginolyticus. BMC Microbiol. 2016, 16, 79. [Google Scholar] [CrossRef]
- Wozniak, R.A.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Daccord, A.; Ceccarelli, D.; Burrus, V. Integrating conjugative elements of the SXT/R391 family trigger the excision and drive the mobilization of a new class of Vibrio genomic islands. Mol. Microbiol. 2010, 78, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tang, K.; Zhang, D.; Li, Y.; Liu, Z.; Yao, J.; Wood, T.K.; Wang, X. Symbiosis of a P2-family phage and deep-sea Shewanella putrefaciens. Environ. Microbiol. 2019, 21, 4212–4232. [Google Scholar] [CrossRef] [PubMed]
- Chibani, C.M.; Roth, O.; Liesegang, H.; Wendling, C.C. Genomic variation among closely related Vibrio alginolyticus strains is located on mobile genetic elements. BMC Genom. 2020, 21, 354. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Ellabaan, M.M.H.; Munck, C.; Porse, A.; Imamovic, L.; Sommer, M.O.A. Forecasting the dissemination of antibiotic resistance genes across bacterial genomes. Nat. Commun. 2021, 12, 2435. [Google Scholar] [CrossRef]
- Camilli, A.; Bassler, B.L. Bacterial small-molecule signaling pathways. Science 2006, 311, 1113–1116. [Google Scholar] [CrossRef]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Romling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef]
- Bordeleau, E.; Brouillette, E.; Robichaud, N.; Burrus, V. Beyond antibiotic resistance: Integrating conjugative elements of the SXT/R391 family that encode novel diguanylate cyclases participate to c-di-GMP signalling in Vibrio cholerae. Environ. Microbiol. 2010, 12, 510–523. [Google Scholar] [CrossRef]
- Wang, W.; Tang, K.; Wang, P.; Zeng, Z.; Xu, T.; Zhan, W.; Liu, T.; Wang, Y.; Wang, X. The coral pathogen Vibrio coralliilyticus kills non-pathogenic holobiont competitors by triggering prophage induction. Nat. Ecol. Evol. 2022, 6, 1132–1144. [Google Scholar] [CrossRef]
- Zeng, Z.; Guo, X.P.; Li, B.; Wang, P.; Cai, X.; Tian, X.; Zhang, S.; Yang, J.L.; Wang, X. Characterization of self-generated variants in Pseudoalteromonas lipolytica biofilm with increased antifouling activities. Appl. Microbiol. Biotechnol. 2015, 99, 10127–10139. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, P.; Zeng, Z.; Cai, X.; Wang, G.; Wang, X. Complete genome sequence of Pseudoalteromonas rubra SCSIO 6842, harboring a putative conjugative plasmid pMBL6842. J. Biotechnol. 2016, 224, 66–67. [Google Scholar] [CrossRef]
- Wang, P.; Yu, Z.; Li, B.; Cai, X.; Zeng, Z.; Chen, X.; Wang, X. Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas. Microb. Cell Factories 2015, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, X.; Yao, J.; Guo, Y.; Li, B.; Li, Y.; Jiao, N.; Wang, X. Cold adaptation regulated by cryptic prophage excision in Shewanella oneidensis. ISME J. 2016, 10, 2787–2800. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Hernandez-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Tang, K.; Zhan, W.; Zhou, Y.; Xu, T.; Chen, X.; Wang, W.; Zeng, Z.; Wang, Y.; Wang, X. Antagonism between coral pathogen Vibrio coralliilyticus and other bacteria in the gastric cavity of scleractinian coral Galaxea fascicularis. Sci. China Earth Sci. 2020, 63, 157–166. [Google Scholar] [CrossRef]
- Zeng, Z.; Dai, S.; Xie, Y.; Tian, X.; Li, J.; Wang, X. Genome sequences of two Pseudoalteromonas strains isolated from the South china sea. Genome Announc. 2014, 2, e00305-14. [Google Scholar] [CrossRef]
- Ni, S.; Li, B.; Tang, K.; Yao, J.; Wood, T.K.; Wang, P.; Wang, X. Conjugative plasmid-encoded toxin-antitoxin system PrpT/PrpA directly controls plasmid copy number. Proc. Natl. Acad. Sci. USA 2021, 118, e2011577118. [Google Scholar] [CrossRef]
- Jiang, W.-X.; Li, P.-Y.; Chen, X.-L.; Zhang, Y.-S.; Wang, J.-P.; Wang, Y.-J.; Sheng, Q.; Sun, Z.-Z.; Qin, Q.-L.; Ren, X.-B.; et al. A pathway for chitin oxidation in marine bacteria. Nat. Commun. 2022, 13, 5899. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 2009, 7, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Beyhan, S.; Tischler, A.D.; Camilli, A.; Yildiz, F.H. Differences in gene expression between the classical and El tor biotypes of Vibrio cholerae O1. Infect. Immun. 2006, 74, 3633–3642. [Google Scholar] [CrossRef]
- Bridges, A.A.; Prentice, J.A.; Wingreen, N.S.; Bassler, B.L. Signal transduction network principles underlying Bacterial collective behaviors. Annu. Rev. Microbiol. 2022, 76, 235–257. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.G.; Zamorano-Sanchez, D.; Park, J.H.; Sondermann, H.; Yildiz, F.H. The ins and outs of cyclic di-GMP signaling in Vibrio cholerae. Curr. Opin. Microbiol. 2017, 36, 20–29. [Google Scholar] [CrossRef]
- Chou, K.T.; Lee, D.D.; Chiou, J.G.; Galera-Laporta, L.; Ly, S.; Garcia-Ojalvo, J.; Suel, G.M. A segmentation clock patterns cellular differentiation in a bacterial biofilm. Cell 2022, 185, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Manner, C.; Dias Teixeira, R.; Saha, D.; Kaczmarczyk, A.; Zemp, R.; Wyss, F.; Jaeger, T.; Laventie, B.J.; Boyer, S.; Malone, J.G.; et al. A genetic switch controls Pseudomonas aeruginosa surface colonization. Nat. Microbiol. 2023, 8, 1520–1533. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.B.; Waters, C.M. Switching on cyclic di-GMP heterogeneity in Pseudomonas aeruginosa biofilms. Nat. Microbiol. 2023, 8, 1380–1381. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Plasmids | Description | Reference |
|---|---|---|
| Va43097 | Vibrio alginolyticus SCSIO 43097, isolated from the gastric cavity of coral Galaxea fascicularis in China: Hainan Island | [8] |
| Pp43201 | Pseudoalteromonas peptidolylytica SCSIO 43201, isolated from the gastric cavity of coral Galaxea fascicularis in China: Hainan Island | [20] |
| Pf43202 | Pseudoalteromonas flavipulchra SCSIO 43202, isolated from the gastric cavity of coral Galaxea fascicularis in China: Hainan Island | [20] |
| Pl04301 | Pseudoalteromonas lipolytica SCSIO 04301, isolated from sediment at 63 m deep in the South China Sea (18°0′ = N, 109°42′ = E) | [21] |
| Pr6842 | Pseudoalteromona rubra SCSIO 6842, isolated from 150 m below surface at Bay of Bengal | [22] |
| VPII | Genomic island integrated in trmA of Va43097 | [8] |
| pBBR1Cm | pBBR1MCS-Cm, broad host range cloning vector containing chloramphenicol resistance gene | [21] |
| pDgc137 | DNA fragment containing gene dgc137 with its promoter cloned and inserted into pBBR1Cm | [8] |
| p3DGC | DNA fragment containing genes dgc137 to dgc140 of VPII cloned and inserted into pBBR1Cm | This study |
| Strains | Genome Content | Size (Mb) | Predicted Genes | GGDEF Domain-Containing Proteins a | Accession Numbers | Reference |
|---|---|---|---|---|---|---|
| Pp43201 | 2 chromosomes and 1 plasmid | 5.2 | 4374 | 51 | CP072683.1, CP072684.1, CP072685.1 | [20,26] |
| Pf43202 | 2 chromosomes | 5.4 | 4705 | 55 | CP024625.1, CP024626.1 | [20,26] |
| Pl04301 | 2 chromosomes | 4.7 | 4220 | 68 | GCA_000576675.1 | [27] |
| Pr6842 | 2 chromosomes and 1 plasmid | 5.9 | 4998 | 51 | CP013611.1, CP013612.1, CP013613.1, | [22,28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, T.; Tang, H.; Du, X.; Wang, W.; Tang, K.; Wang, X.; Liu, D.; Wang, P. Genomic Island-Encoded Diguanylate Cyclase from Vibrio alginolyticus Regulates Biofilm Formation and Motility in Pseudoalteromonas. Microorganisms 2023, 11, 2725. https://doi.org/10.3390/microorganisms11112725
Cai T, Tang H, Du X, Wang W, Tang K, Wang X, Liu D, Wang P. Genomic Island-Encoded Diguanylate Cyclase from Vibrio alginolyticus Regulates Biofilm Formation and Motility in Pseudoalteromonas. Microorganisms. 2023; 11(11):2725. https://doi.org/10.3390/microorganisms11112725
Chicago/Turabian StyleCai, Tongxuan, Huan Tang, Xiaofei Du, Weiquan Wang, Kaihao Tang, Xiaoxue Wang, Dong Liu, and Pengxia Wang. 2023. "Genomic Island-Encoded Diguanylate Cyclase from Vibrio alginolyticus Regulates Biofilm Formation and Motility in Pseudoalteromonas" Microorganisms 11, no. 11: 2725. https://doi.org/10.3390/microorganisms11112725