
Conjugative RP4 Plasmid-Mediated Transfer of Antibiotic Resistance Genes to Commensal and Multidrug-Resistant Enteric Bacteria In Vitro
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Chemicals, and Reagents
2.2. Isolation and Characterization of a Human-Derived Donor Bacterial Strain
2.3. Creation of a Fluorescently Labeled Commensal Donor Strain
2.4. Donor and Recipient Strains
2.5. In Vitro Conjugation Experiments to Confirm Plasmid Transfer
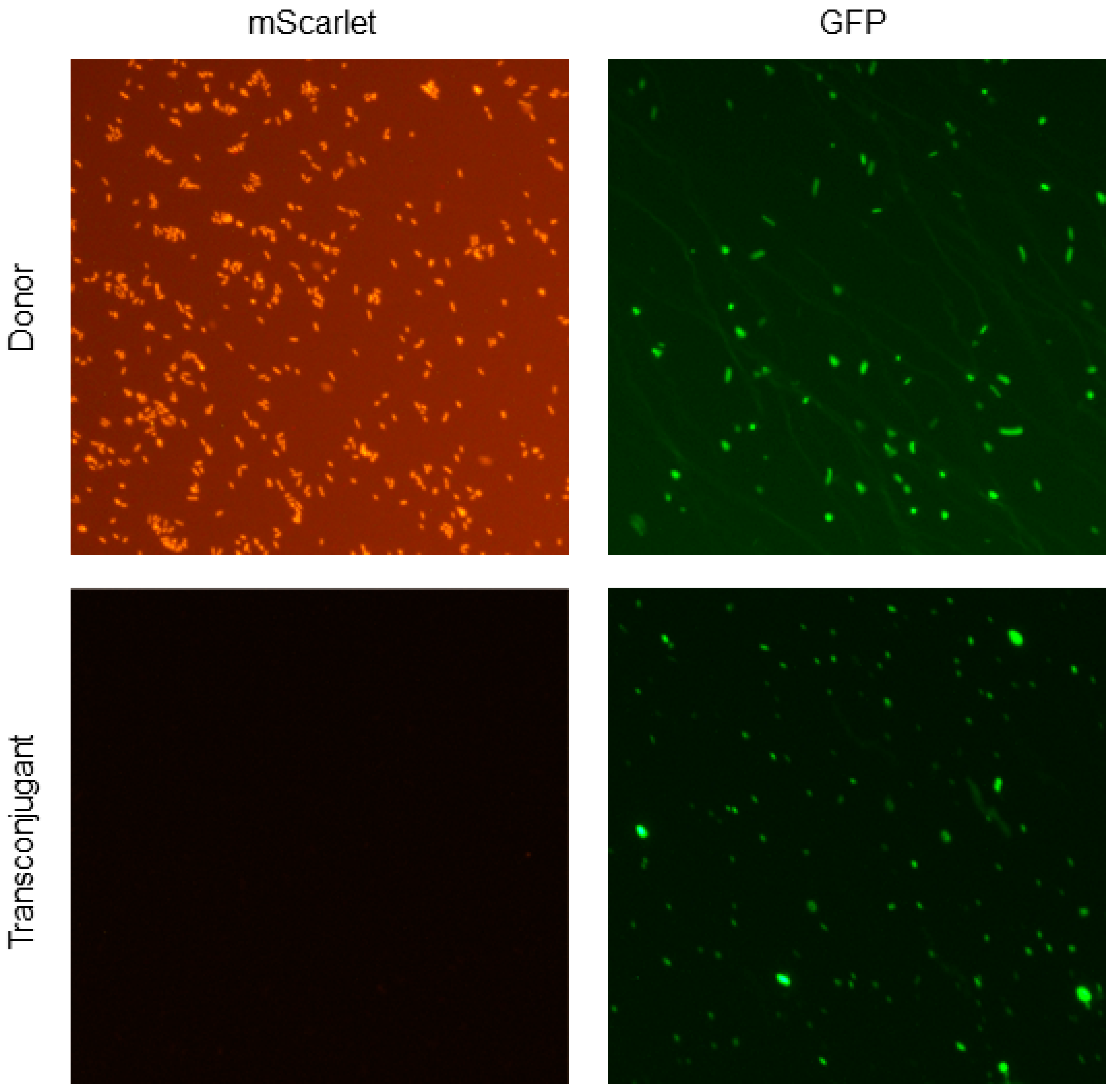
2.6. Detection of Fluorescence in Donor and Transconjugant Bacteria
2.7. Confirmation of the Plasmid in Transconjugant Bacteria Using Colony PCR
2.8. Bacteriocin Assay
2.9. Plasmid Persistence Assay
2.10. Statistical Analysis
3. Results
3.1. Characterization of Donor and Recipient Strains
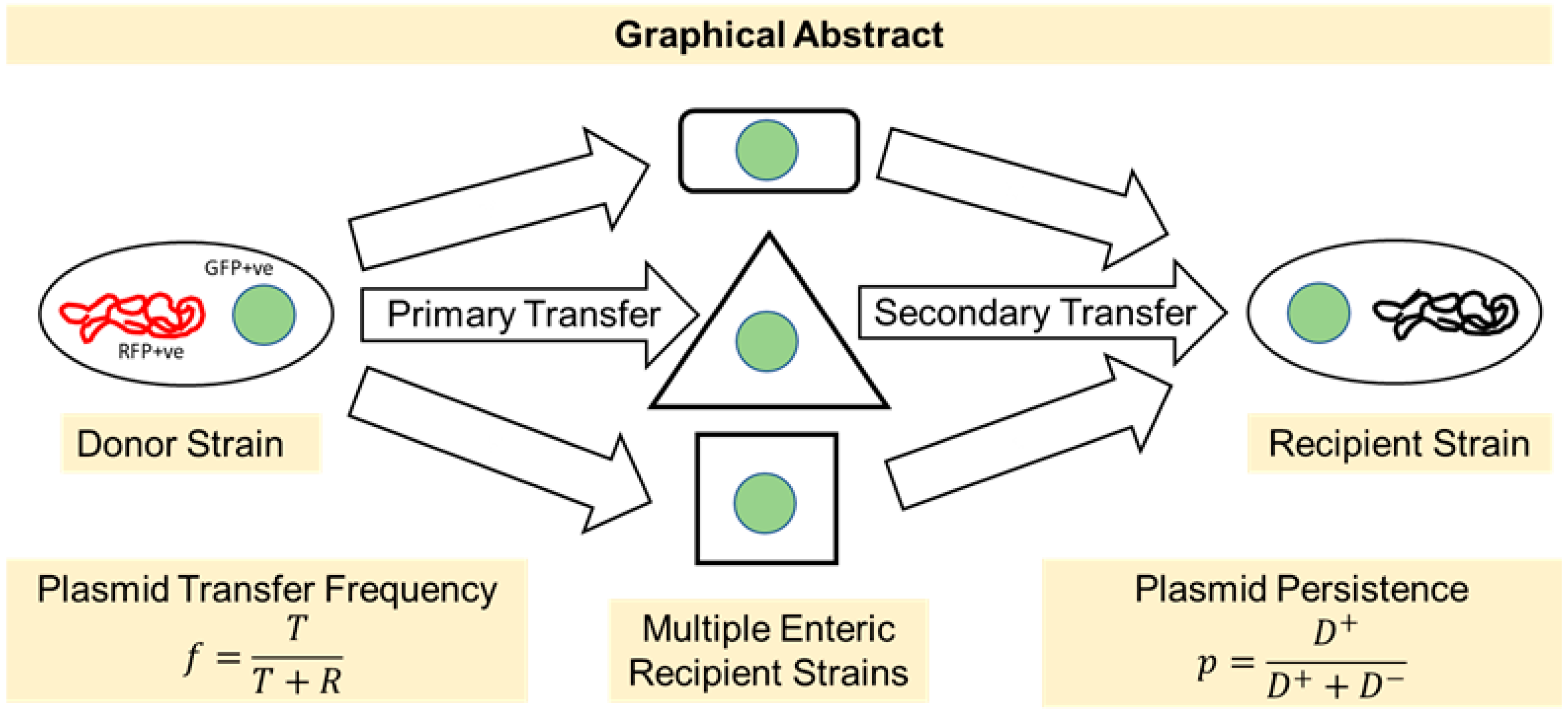
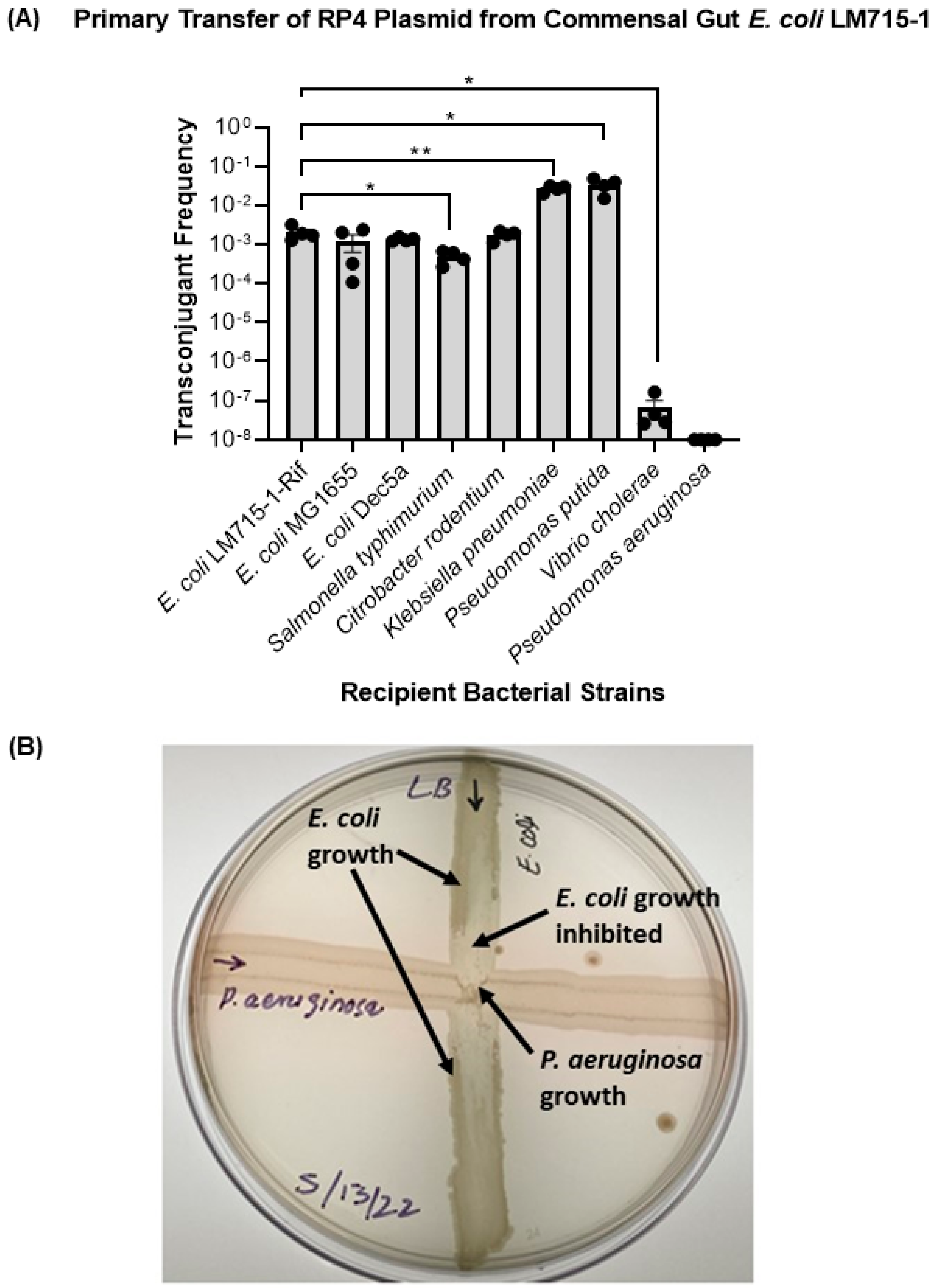
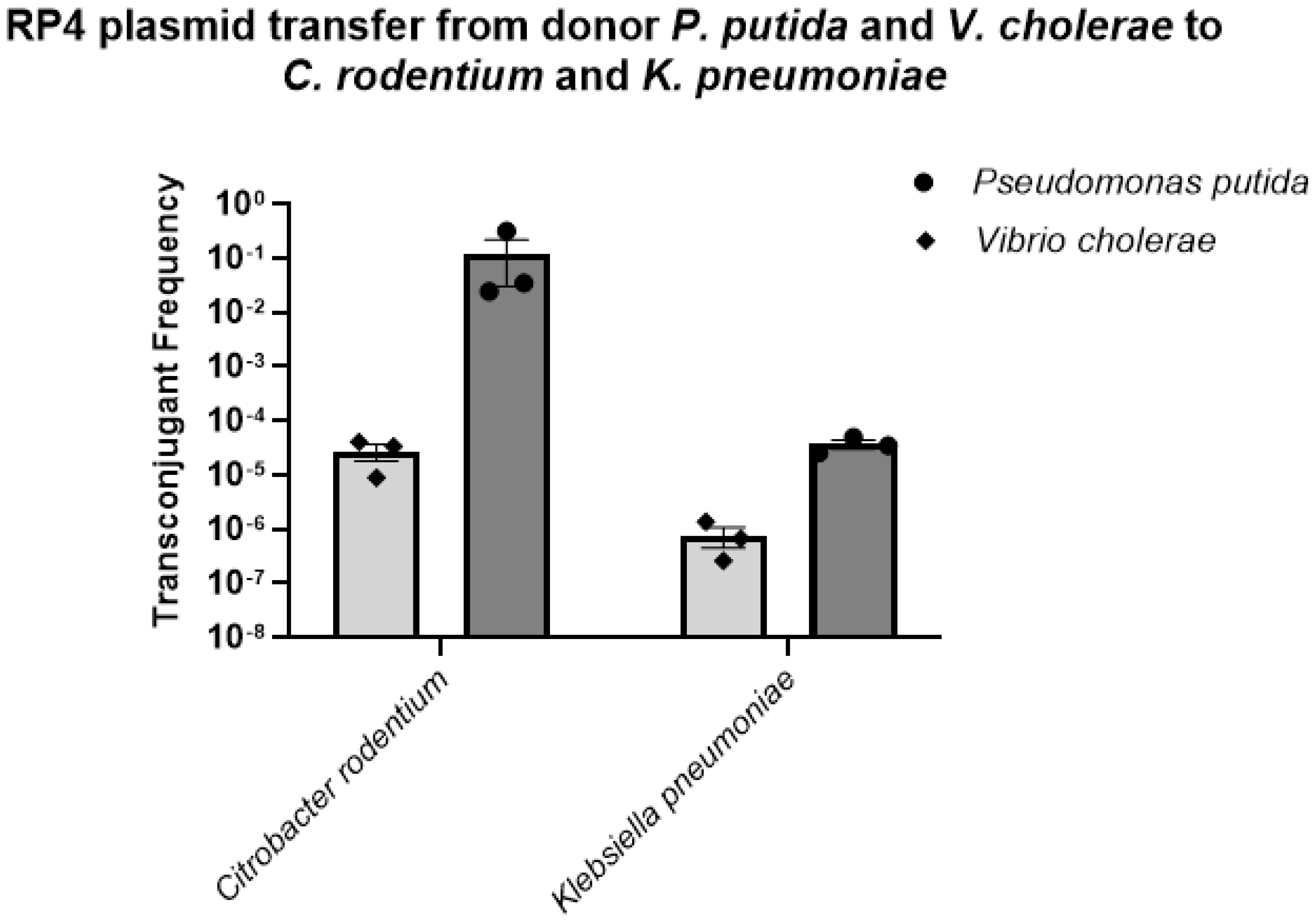
3.2. The Labeled RP4 Plasmid Transferred to Multiple Drug-Resistant Bacterial Strains
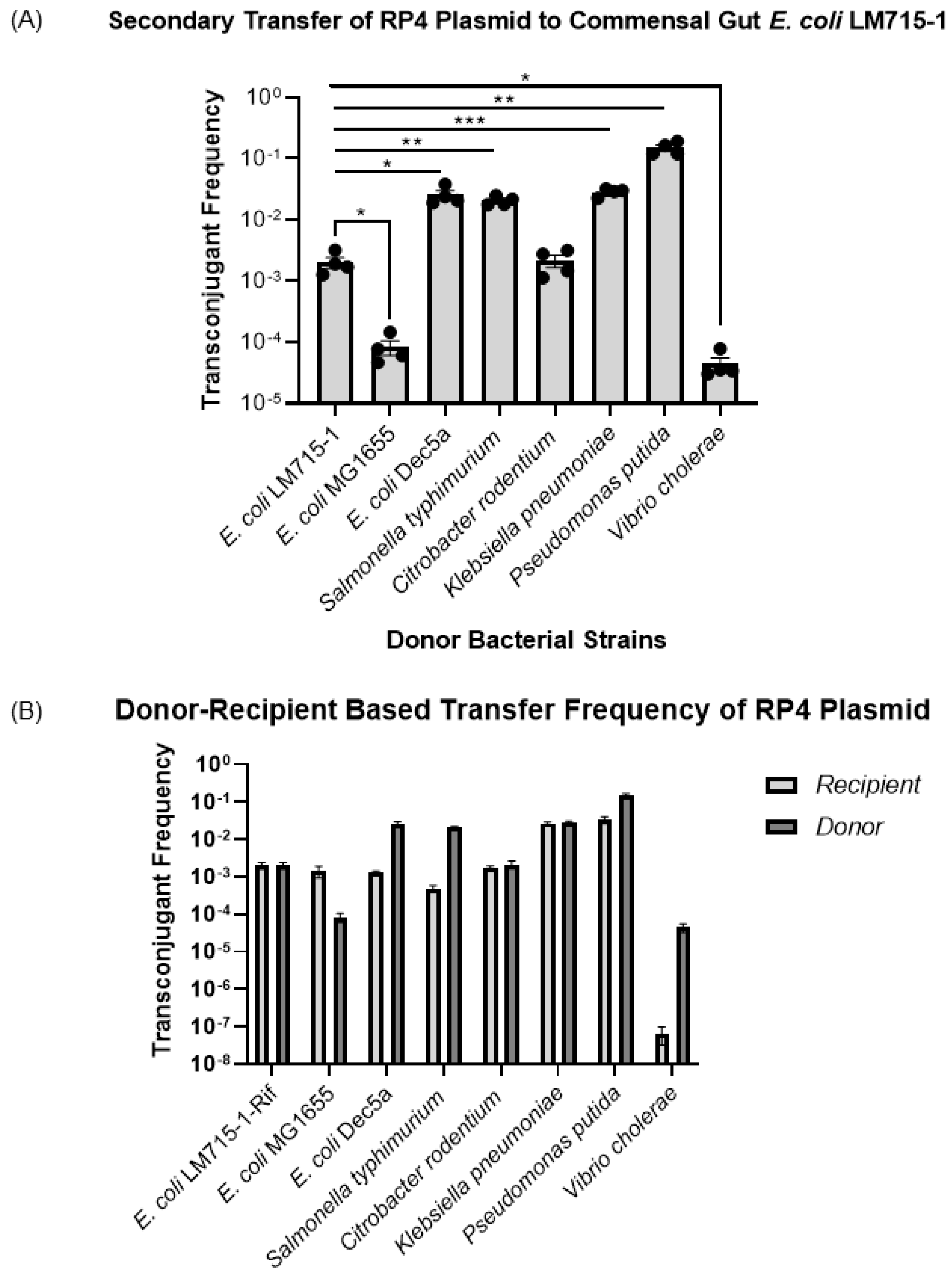
3.3. All Recipient Bacterial Strains Can Mediate Secondary Transfer of the BHR Plasmid to the Human Commensal E. coli LM715-1 Recipient Strain
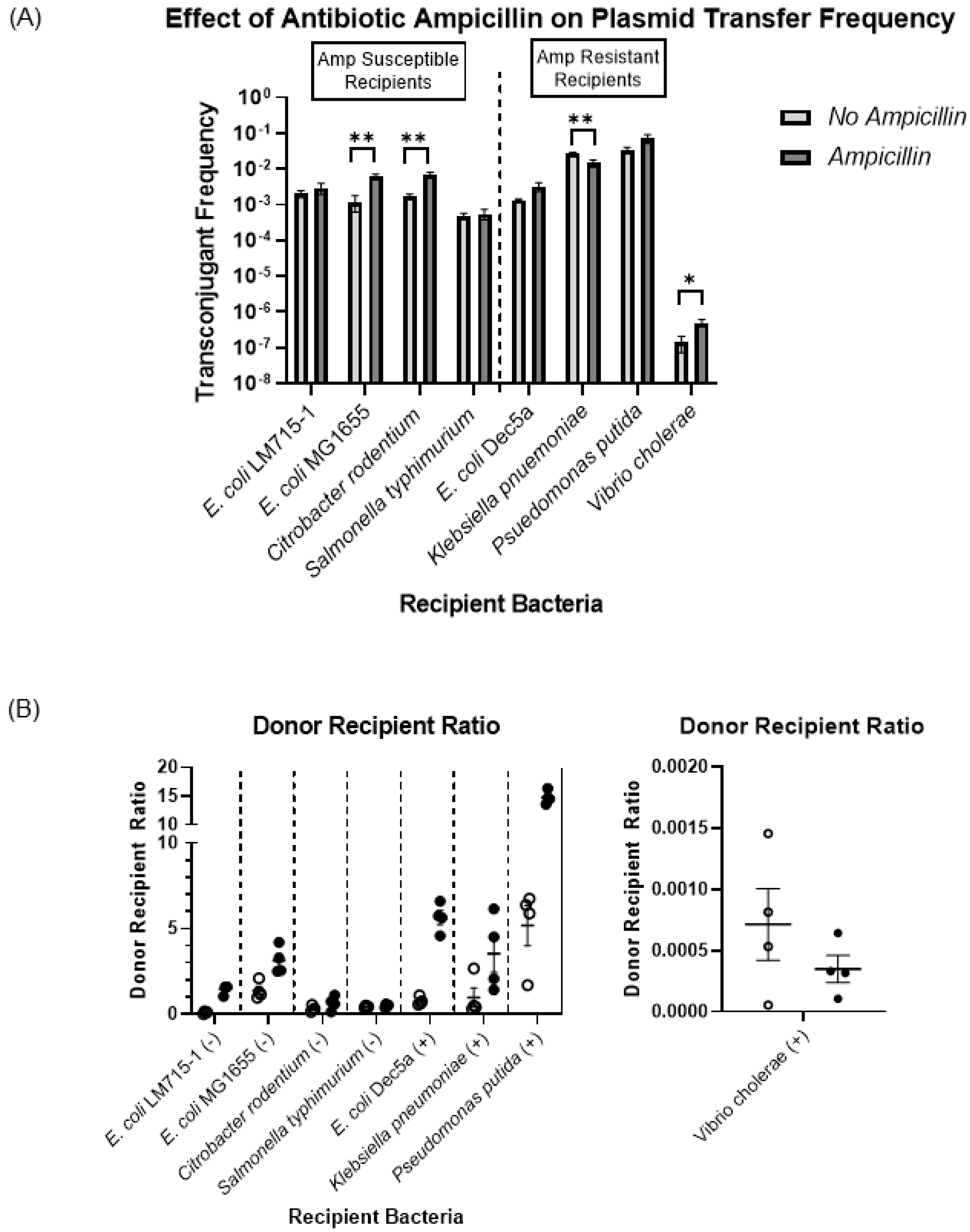
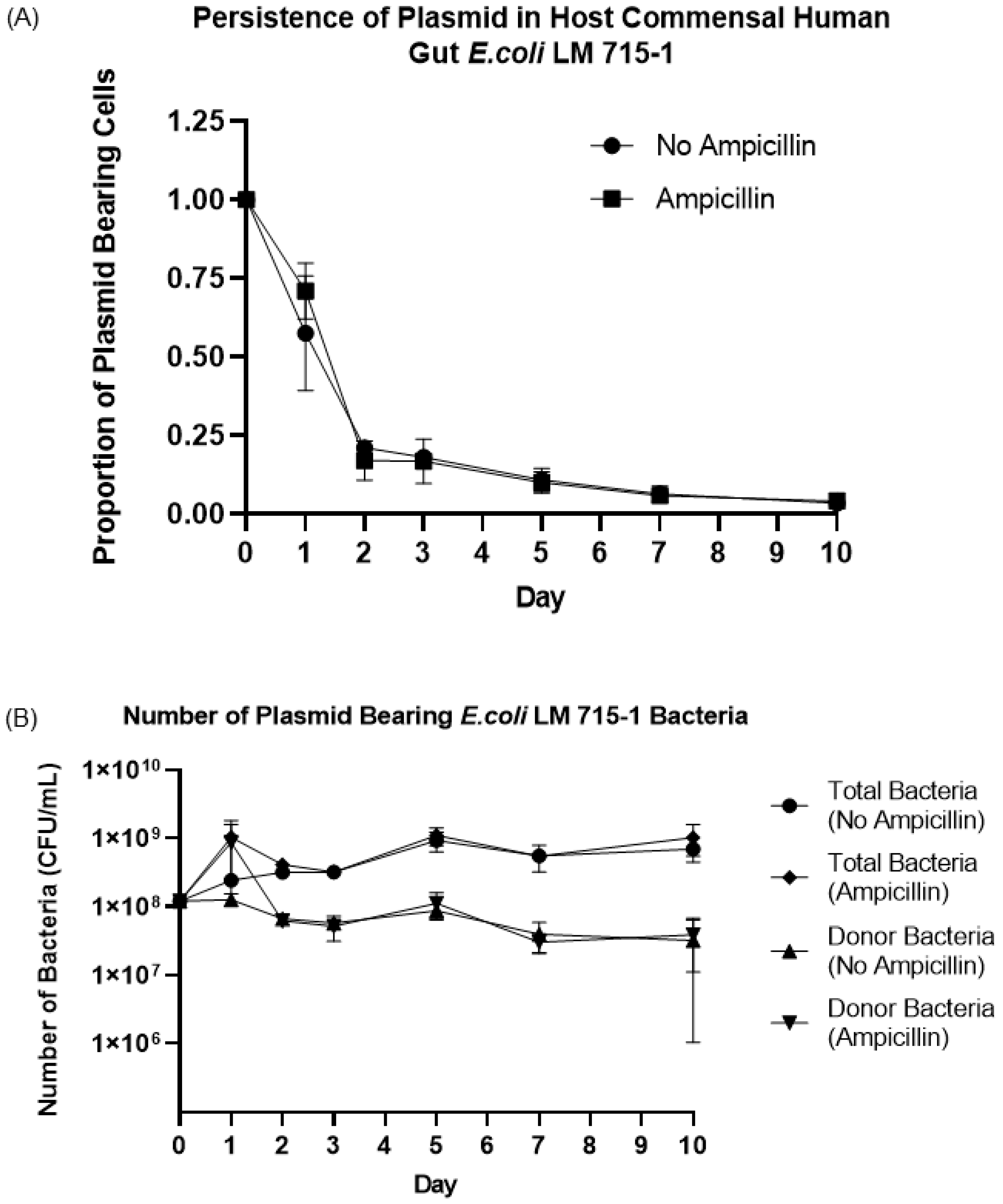
3.4. Fitness Cost and Persistence of Broad-Host-Range Plasmid during Adaptation and Evolution in a Naïve Host Bacterium
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Shunji Ikuta, K.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019; p. 148. [CrossRef] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. In The Review on Antimicrobial Resistance; HM Government and the Wellcome Trust: London, UK, 2016. [Google Scholar]
- Jain, A.; Srivastava, P. Broad host range plasmids. FEMS Microbiol. Lett. 2013, 348, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlechter, R.O.; Jun, H.; Bernach, M.; Oso, S.; Boyd, E.; Muñoz-Lintz, D.A.; Dobson, R.C.J.; Remus, D.M.; Remus-Emsermann, M.N.P. Chromatic Bacteria—A Broad Host-Range Plasmid and Chromosomal Insertion Toolbox for Fluorescent Protein Expression in Bacteria. Front. Microbiol. 2018, 9, 3052. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I. Horizontal gene exchange in environmental microbiota. Front. Microbiol. 2011, 2, 158. [Google Scholar] [CrossRef] [Green Version]
- Datta, N.; Hedges, R.W. R factors of compatibility group A. J. Gen. Microbiol. 1973, 74, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [Green Version]
- Popowska, M.; Krawczyk-Balska, A. Broad-host-range IncP-1 plasmids and their resistance potential. Front. Microbiol. 2013, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Klümper, U.; Riber, L.; Dechesne, A.; Sannazzarro, A.; Hansen, L.H.; Sørensen, S.J.; Smets, B.F. Broad host range plasmids can invade an unexpectedly diverse fraction of a soil bacterial community. ISME J. 2015, 9, 934–945. [Google Scholar] [CrossRef]
- Lu, X.; Hu, Y.; Luo, M.; Zhou, H.; Wang, X.; Du, Y.; Li, Z.; Xu, J.; Zhu, B.; Xu, X.; et al. MCR-1.6, a new MCR variant carried by an IncP plasmid in a colistin-resistant Salmonella enterica serovar Typhimurium isolate from a healthy individual. Antimicrob. Agents Chemother. 2017, 61, e02632-16. [Google Scholar] [CrossRef] [Green Version]
- Pansegrau, W.; Lanka, E.; Barth, P.T.; Figurski, D.H.; Guiney, D.G.; Haas, D.; Helinski, D.R.; Schwab, H.; Stanisich, V.A.; Thomas, C.M. Complete nucleotide sequence of birmingham IncPα plasmids. Compilation and comparative analysis. J. Mol. Biol. 1994, 239, 623–663. [Google Scholar] [CrossRef]
- Datta, N.; Hedges, R.W. Compatibility Groups among fi− R Factors. Nature 1971, 234, 222–223. [Google Scholar] [CrossRef]
- Adamczyk, M.; Jagura-Burdzy, G. Spread and survival of promiscuous IncP-1 plasmids. Acta Biochim. Pol. 2003, 50, 425–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, P.T.; Grinter, N.J. Map of plasmid RP4 derived by insertion of transposon C. J. Mol. Biol. 1977, 113, 455–474. [Google Scholar] [CrossRef]
- Musovic, S.; Klümper, U.; Dechesne, A.; Magid, J.; Smets, B.F. Long-term manure exposure increases soil bacterial community potential for plasmid uptake. Environ. Microbiol. Rep. 2014, 6, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johannes Sørensen, S.; Schyberg, T.; Rønn, R. Predation by protozoa on Escherichia coli K12 in soil and transfer of resistance plasmid RP4 to indigenous bacteria in soil. Appl. Soil Ecol. 1999, 11, 79–90. [Google Scholar] [CrossRef]
- Liu, Z.; Klümper, U.; Liu, Y.; Yang, Y.; Wei, Q.; Lin, J.-G.; Gu, J.-D.; Li, M. Metagenomic and metatranscriptomic analyses reveal activity and hosts of antibiotic resistance genes in activated sludge. Environ. Int. 2019, 129, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Geisenberger, O.; Ammendola, A.; Christensen, B.B.; Molin, S.; Schleifer, K.-H.; Eberl, L. Monitoring the conjugal transfer of plasmid RP4 in activated sludge and in situ identification of the transconjugants. FEMS Microbiol. Lett. 1999, 174, 9–17. [Google Scholar] [CrossRef]
- Soda, S.; Otsuki, H.; Inoue, D.; Tsutsui, H.; Sei, K.; Ike, M. Transfer of antibiotic multiresistant plasmid RP4 from escherichia coli to activated sludge bacteria. J. Biosci. Bioeng. 2008, 106, 292–296. [Google Scholar] [CrossRef]
- Heß, S.; Kneis, D.; Virta, M.; Hiltunen, T. The spread of the plasmid RP4 in a synthetic bacterial community is dependent on the particular donor strain. FEMS Microbiol. Ecol. 2022, 97, fiab147. [Google Scholar] [CrossRef]
- Shintani, M.; Sanchez, Z.K.; Kimbara, K. Genomics of microbial plasmids: Classification and identification based on replication and transfer systems and host taxonomy. Front. Microbiol. 2015, 6, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A. Resistance Plasmid Families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Millan, A.; MacLean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Dechesne, A.; Madsen, J.S.; Nesme, J.; Sørensen, S.J.; Smets, B.F. Plasmids persist in a microbial community by providing fitness benefit to multiple phylotypes. ISME J. 2020, 14, 1170–1181. [Google Scholar] [CrossRef]
- Brooks, P.T.; Brakel, K.A.; Bell, J.A.; Bejcek, C.E.; Gilpin, T.; Brudvig, J.M.; Mansfield, L.S. Transplanted human fecal microbiota enhanced Guillain Barré syndrome autoantibody responses after Campylobacter jejuni infection in C57BL/6 mice. Microbiome 2017, 5, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; Fan, Y.; Achtman, M. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Schlechter, R.O.; Remus-Emsermann, M.N. Delivering “Chromatic Bacteria” Fluorescent Protein Tags to Proteobacteria Using Conjugation. Bio Protoc. 2019, 9, e3199. [Google Scholar] [CrossRef]
- Reid, S.D.; Betting, D.J.; Whittam, T.S. Molecular Detection and Identification of Intimin Alleles in Pathogenic Escherichia Coli by Multiplex PCR. J. Clin. Microbiol. 1999, 37, 2719. [Google Scholar] [CrossRef]
- Jagnow, J.; Clegg, S. Klebsiella Pneumoniae MrkD-Mediated Biofilm Formation on Extracellular Matrix- and Collagen-Coated Surfaces. Microbiology 2003, 149, 2397–2405. [Google Scholar] [CrossRef] [Green Version]
- Sambanthamoorthy, K.; Gokhale, A.A.; Lao, W.; Parashar, V.; Neiditch, M.B.; Semmelhack, M.F.; Lee, I.; Waters, C.M. Identification of a Novel Benzimidazole That Inhibits Bacterial Biofilm Formation in a Broad-Spectrum Manner. Antimicrob. Agents Chemother. 2011, 55, 4369–4378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.; Janagama, H.; Dwivedi, H.P.; Senthil Kumar, T.M.A.; Jaykus, L.A.; Schefers, J.; Sreevatsan, S. Selection, Characterization, and Application of DNA Aptamers for the Capture and Detection of Salmonella Enterica Serovars. Mol. Cell Probes 2009, 23, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Helene Thelin, K.; Taylor, R.K. Toxin-Coregulated Pilus, but Not Mannose-Sensitive Hemagglutinin, Is Required for Colonization by Vibrio Cholerae O1 El Tor Biotype and O139 Strains. Infect. Immun. 1996, 64, 2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trieu-Cuot, P.; Courvalin, P. Transposition Behavior of IS75 and Its Progenitor IS75A: Are Cointegrates Exclusive End Products? Plasmid 1985, 14, 80–89. [Google Scholar] [CrossRef]
- Mani, R.J.; Thachil, A.J.; Ramachandran, A. Discrimination of Streptococcus equi subsp. equi and Streptococcus equi subsp. zooepidemicus using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Vet. Diagn. Investig. 2017, 29, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Skinner, S.O.; Sepúlveda, L.A.; Xu, H.; Golding, I. Measuring mRNA copy-number in individual Escherichia coli cells using single-molecule fluorescent in situ hybridization (smFISH). Nat. Protoc. 2013, 8, 1100. [Google Scholar] [CrossRef] [PubMed]
- Williston, E.H.; Zia-Walrath, P.; Youmans, G.P. Plate Methods for Testing Antibiotic Activity of Actinomycetes against Virulent Human Type Tubercle Bacilli. J. Bacteriol. 1947, 54, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Welch, B.L. The significance of the difference between two means when the population variances are unequal. Biometrika 1938, 29, 350–362. [Google Scholar] [CrossRef]
- Toval, F.; Köhler, C.D.; Vogel, U.; Wagenlehner, F.; Mellmann, A.; Fruth, A.; Schmidt, M.A.; Karch, H.; Bielaszewska, M.; Dobrindt, U. Characterization of Escherichia coli Isolates from Hospital Inpatients or Outpatients with Urinary Tract Infection. J. Clin. Microbiol. 2014, 52, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H.; et al. Heteropathogenic virulence and phylogeny reveal phased pathogenic metamorphosis in Escherichia coli O2:H6. EMBO Mol. Med. 2014, 6, 347–357. [Google Scholar] [CrossRef]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and Evolution of Hybrid Shiga Toxin-Producing and Uropathogenic Escherichia coli Strains of Sequence Type 141. J. Clin. Microbiol. 2020, 58, e01309-19. [Google Scholar] [CrossRef] [PubMed]
- Zhi, S.; Stothard, P.; Banting, G.; Scott, C.; Huntley, K.; Ryu, K.; Otto, S.; Ashbolt, N.; Checkley, S.; Dong, T.; et al. Characterization of water treatment-resistant and multidrug-resistant urinary pathogenic Escherichia coli in treated wastewater. Water Res. 2020, 182, 115827. [Google Scholar] [CrossRef]
- Yun, K.W.; Kim, H.Y.; Park, H.K.; Kim, W.; Lim, I.S. Virulence factors of uropathogenic Escherichia coli of urinary tract infections and asymptomatic bacteriuria in children. J. Microbiol. Immunol. Infect. 2014, 47, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli isolates that carry vat, fyua, chua, and yfcv efficiently colonize the urinary tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipour, T.; Poursina, F. The frequency of hybrid Enteroaggregative/Uropathogenic Escherichia coli isolated from clinical samples of Isfahan hospitals, Iran. Gene Rep. 2021, 23, 101042. [Google Scholar] [CrossRef]
- Zhang, Y.; Hong, P.Y.; LeChevallier, M.W.; Liu, W.T. Phenotypic and phylogenetic identification of coliform bacteria obtained using 12 coliform methods approved by the U.S. Environmental Protection Agency. Appl. Environ. Microbiol. 2015, 81, 6012–6023. [Google Scholar] [CrossRef] [Green Version]
- Brenner, D.J.; Farmer, J.J. Enterobacteriaceae. In Bergey’s Manual of Systematics Archaea and Bacteria; John Wiley & Sons, Inc., in association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015; Volume 15, pp. 1–24. [Google Scholar] [CrossRef]
- Farmer, J.J.; Janda, J.M. Vibrionaceae. In Bergey’s Manual of Systematics Archaea and Bacteria; John Wiley & Sons, Inc., in association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015; pp. 1–7. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Pseudomonadaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc., in association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Figurski, D.H.; Helinski, D.R. Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Proc. Natl. Acad. Sci. USA 1979, 76, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Huddleston, J.R. Horizontal gene transfer in the human gastrointestinal tract: Potential spread of antibiotic resistance genes. Infect. Drug Resist. 2014, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Benz, F.; Huisman, J.S.; Bakkeren, E.; Herter, J.A.; Stadler, T.; Ackermann, M.; Diard, M.; Egli, A.; Hall, A.R.; Hardt, W.-D.; et al. Plasmid- and strain-specific factors drive variation in ESBL-plasmid spread in vitro and in vivo. ISME J. 2021, 15, 862–878. [Google Scholar] [CrossRef]
- Headd, B.; Bradford, S.A. Physicochemical factors that favor conjugation of an antibiotic resistant plasmid in non-growing bacterial cultures in the absence and presence of antibiotics. Front. Microbiol. 2018, 9, 2122. [Google Scholar] [CrossRef] [PubMed]
- Lopatkin, A.J.; Huang, S.; Smith, R.P.; Srimani, J.K.; Sysoeva, T.A.; Bewick, S.; Karig, D.K.; You, L. Antibiotics as a selective driver for conjugation dynamics. Nat. Microbiol. 2016, 1, 16044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- Cairns, J.; Ruokolainen, L.; Hultman, J.; Tamminen, M.; Virta, M.; Hiltunen, T. Ecology determines how low antibiotic concentration impacts community composition and horizontal transfer of resistance genes. Commun. Biol. 2018, 1, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes 2020, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and Barriers to, Horizontal Gene Transfer between Bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Purdy, D.; O’Keeffe, T.A.T.; Elmore, M.; Herbert, M.; McLeod, A.; Bokori-Brown, M.; Ostrowski, A.; Minton, N.P. Conjugative transfer of clostridial shuttle vectors from Escherichia coli to Clostridium difficile through circumvention of the restriction barrier. Mol. Microbiol. 2002, 46, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Melamed, S.; Ofir, G.; Leavitt, A.; Lopatina, A.; Keren, M.; Amitai, G.; Sorek, R. Systematic discovery of antiphage defense systems in the microbial pangenome. Science 2018, 359, eaar4120. [Google Scholar] [CrossRef] [Green Version]
- Price, V.J.; McBride, S.W.; Hullahalli, K.; Chatterjee, A.; Duerkop, B.A.; Palmer, K.L. Enterococcus faecalis CRISPR-Cas Is a Robust Barrier to Conjugative Antibiotic Resistance Dissemination in the Murine Intestine. mSphere 2019, 4, e00464-19. [Google Scholar] [CrossRef] [Green Version]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Ghequire, M.G.K.; Öztürk, B. A Colicin M-Type Bacteriocin from Pseudomonas aeruginosa Targeting the HxuC Heme Receptor Requires a Novel Immunity Partner. Appl. Environ. Microbiol. 2018, 84, e00716-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Händel, N.; Otte, S.; Jonker, M.; Brul, S.; Kuile, B.H.T. Factors That Affect Transfer of the IncI1 β-Lactam Resistance Plasmid pESBL-283 between E. coli Strains. PLoS ONE 2015, 10, e0123039. [Google Scholar] [CrossRef] [PubMed]
- David, X.; Bradley, E. Determination of pili by conjugative bacterial drug resistance plasmids of incompatibility groups B, C, H, J, K, M, V, and X. J. Bacteriol. 1980, 141, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Lenski, R.E.; Bouma, J.E. Effects of segregation and selection on instability of plasmid pACYC184 in Escherichia coli B. J. Bacteriol. 1987, 169, 5314–5316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouma, J.E.; Lenski, R.E. Evolution of a bacteria/plasmid association. Nature 1988, 335, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Porse, A.; Schønning, K.; Munck, C.; Sommer, M.O.A. Survival and Evolution of a Large Multidrug Resistance Plasmid in New Clinical Bacterial Hosts. Mol. Biol. Evol. 2016, 33, 2860–2873. [Google Scholar] [CrossRef] [PubMed]
- Ronda, C.; Chen, S.P.; Cabral, V.; Yaung, S.J.; Wang, H.H. Metagenomic engineering of the mammalian gut microbiome in situ. Nat. Methods 2019, 16, 167–170. [Google Scholar] [CrossRef]
- Lopatkin, A.J.; Meredith, H.R.; Srimani, J.K.; Pfeiffer, C.; Durrett, R.; You, L. Persistence and reversal of plasmid-mediated antibiotic resistance. Nat. Commun. 2017, 8, 1689. [Google Scholar] [CrossRef] [Green Version]
- San Millan, A. Evolution of Plasmid-Mediated Antibiotic Resistance in the Clinical Context. Trends Microbiol. 2018, 26, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Hughes, D. Persistence of antibiotic resistance in bacterial populations. FEMS Microbiol. Rev. 2011, 35, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Bahl, M.I.; Hansen, L.H.; Sørensen, S.J. Persistence Mechanisms of Conjugative Plasmids. Methods Mol. Biol. 2009, 532, 73–102. [Google Scholar] [CrossRef] [PubMed]
- Bottery, M.J.; Wood, A.J.; Brockhurst, M.A. Temporal dynamics of bacteria-plasmid coevolution under antibiotic selection. ISME J. 2018, 13, 559–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Beltran, J.; Hernandez-Beltran, J.C.R.; Delafuente, J.; Escudero, J.A.; Fuentes-Hernandez, A.; MacLean, R.C.; Peña-Miller, R.; Millan, A.S. Multicopy plasmids allow bacteria to escape from fitness trade-offs during evolutionary innovation. Nat. Ecol. Evol. 2018, 2, 873–881. [Google Scholar] [CrossRef]
- Stewart, F.M.; Levin, B.R. The Population Biology of Bacterial Plasmids: A PRIORI Conditions for the Existence of Conjugationally Transmitted Factors. Genetics 1977, 87, 209–228. [Google Scholar] [CrossRef]
- Bahl, M.I.; Hansen, L.H.; Licht, T.R.; Sørensen, S.J. Conjugative transfer facilitates stable maintenance of IncP-1 plasmid pKJK5 in Escherichia coli cells colonizing the gastrointestinal tract of the germfree rat. Appl. Environ. Microbiol. 2007, 73, 341–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salyers, A.A.; Am’abile, C.F.; Am’abile-Cuevas, A. MINIREVIEW Why Are Antibiotic Resistance Genes So Resistant to Elimination? Antimicrob. Agents Chemother. 1997, 41, 2321–2325. [Google Scholar] [CrossRef]
- Nicoloff, H.; Hjort, K.; Levin, B.R.; Andersson, D.I. The high prevalence of antibiotic heteroresistance in pathogenic bacteria is mainly caused by gene amplification. Nat. Microbiol. 2019, 4, 504–514. [Google Scholar] [CrossRef]
- Zeng, X.; Lin, J. Beta-lactamase induction and cell wall metabolism in Gram-negative bacteria. Front. Microbiol. 2013, 4, 128. [Google Scholar] [CrossRef] [Green Version]
- Moya, B.; Dötsch, A.; Juan, C.; Blázquez, J.; Zamorano, L.; Haussler, S.; Oliver, A. Beta-lactam resistance response triggered by inactivation of a nonessential penicillin-binding protein. PLoS Pathog. 2009, 5, e1000353. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description of Donor and Recipient Strains Included in This Study | |||
|---|---|---|---|
| Bacterial Strain | Antibiotic Resistance Phenotype | Isolate Type | Source (Attached References) |
| Escherichia coli LM715-1 | Human E. coli Strain—ST 259 | Linda S. Mansfield lab (This study) | |
| Escherichia coli LM715-1 | CamR, KanR | Human E. coli Strain—ST 259 | Linda S. Mansfield lab (This study) |
| Escherichia coli LM715-1 | RifR | Human E. coli Strain—ST 259 | Linda S. Mansfield lab (This study) |
| Escherichia coli MG1655 | RifR | K-12 E. coli Laboratory Strain | ATCC/Lixin Zhang lab |
| Escherichia coli DEC 5a TW00587 | AmpR, RifR | Human Diarrheagenic E. coli Strain (DEC)—ST 73 | STEC/Shannon Manning lab [31] |
| Citrobacter rodentium ATCC 51459 | RifR | Pathogen Strain | ATCC/Linda S. Mansfield lab |
| Pseudomonas putida KT2440 | AmpR, CtxR, RifR | Environmental Strain | ATCC/Lixin Zhang lab |
| Pseudomonas aeruginosa | AmpR, CtxR, CamR | Human Pathogen Strain Isolated from CF Patient | Robert Quin Lab (personal communication) |
| Klebsiella pneumoniae IA565 | AmpR, RifR | Human Pathogen Strain | Christopher Waters lab [32,33] |
| Salmonella enterica serovar Typhimurium | RifR | Clinical Strain Isolated from Chicken | Srinand Sreevatsan lab [34] |
| Vibrio cholerae O1 biotype El Tor C6706str2 | AmpR, StrepR | Human Pathogen Strain | Christopher Waters lab [35] |
| Primers Used for Donor and Transconjugant Confirmation | ||||
|---|---|---|---|---|
| Primer | Product Size (in bp) | Primer Sequence (5′-3′) | Target Gene | Gene Bank Accession No. References |
| gfpF | 182 | GGTGAAGGTGAAGGTGATGC | gfp | U73901.1 |
| gfpR | CTTCTGGCATGGCAGACTTG | |||
| mScarletF | 371 | CGCGTGATGAACTTTGAAGA | mScarlet-I | KY021424.1 |
| mScarletR | TCGCTGCGTTCATACTGTTC | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sher, A.A.; VanAllen, M.E.; Ahmed, H.; Whitehead-Tillery, C.; Rafique, S.; Bell, J.A.; Zhang, L.; Mansfield, L.S. Conjugative RP4 Plasmid-Mediated Transfer of Antibiotic Resistance Genes to Commensal and Multidrug-Resistant Enteric Bacteria In Vitro. Microorganisms 2023, 11, 193. https://doi.org/10.3390/microorganisms11010193
Sher AA, VanAllen ME, Ahmed H, Whitehead-Tillery C, Rafique S, Bell JA, Zhang L, Mansfield LS. Conjugative RP4 Plasmid-Mediated Transfer of Antibiotic Resistance Genes to Commensal and Multidrug-Resistant Enteric Bacteria In Vitro. Microorganisms. 2023; 11(1):193. https://doi.org/10.3390/microorganisms11010193
Chicago/Turabian StyleSher, Azam A., Mia E. VanAllen, Husnain Ahmed, Charles Whitehead-Tillery, Sonia Rafique, Julia A. Bell, Lixin Zhang, and Linda S. Mansfield. 2023. "Conjugative RP4 Plasmid-Mediated Transfer of Antibiotic Resistance Genes to Commensal and Multidrug-Resistant Enteric Bacteria In Vitro" Microorganisms 11, no. 1: 193. https://doi.org/10.3390/microorganisms11010193