
Factors Influencing the Reproductive Ability of Male Bees: Current Knowledge and Further Directions


Key Laboratory for Insect-Pollinator Biology of the Ministry of Agriculture, Institute of Apicultural Research, Chinese Academy of Agricultural Science, Beijing 100093, China
*
Authors to whom correspondence should be addressed.
Insects 2021, 12(6), 529; https://doi.org/10.3390/insects12060529
Submission received: 9 April 2021
/
Revised: 20 May 2021
/
Accepted: 31 May 2021
/
Published: 7 June 2021
Abstract
:Simple Summary
Bumblebees and honeybees are well known as the dominant and most important pollinators in natural and agricultural ecosystems. The quality characteristics of their colonies depend greatly on the reproductive ability/quality of the parents (queens and drones). Male bees, despite their exclusive reproductive role and ability to determine colony quality, have been less considered than female bees, especially bumblebees. We reviewed the current studies on environmental factors and inherent characteristics that affect the mating success and fecundity of male honeybees and bumblebees. Temperature, nutrients, pesticides, body size, weight and age affect reproduction in male bees and consequently the progeny colony quality. However, more studies, especially in male bumblebees, are still needed to address the impacts of these factors in detail to confront the requirements of agricultural pollination and declining wild bee pollinators worldwide.
Abstract
Bumblebees and honeybees are very important pollinators and play a vital role in agricultural and natural ecosystems. The quality of their colonies is determined by the queens and the reproductive drones of mother colonies, and mated drones transmit semen, including half of the genetic materials, to queens and enhance their fertility. Therefore, factors affecting drone fecundity will also directly affect progeny at the colony level. Here, we review environmental and bee-related factors that are closely related to drone reproductive ability. The environmental factors that mainly affect the sperm count and the viability of males include temperature, nutrients and pesticides. In addition, the inherent characteristics of male bees, such as body size, weight, age, seminal fluid proteins and proteins of the spermathecal fluid, contribute to mating success, sperm quality during long-term storage in the spermathecae and the reproductive behaviors of queens. Based on the results of previous studies, we also suggest that the effects of somatotype dimorphism in bumblebee males on sperm quality and queen fecundity and the indispensable and exploitable function of gland proteins in the fecundity of males and queens should be given more attention in further studies.
1. Life and Role of Male Bees in Colonies
Honeybees and bumblebees are the most important pollinators in agricultural systems. Similar to other eusocial species in the order Hymenoptera, honeybees and bumblebees live in large colonies containing a queen, workers, and males [1,2]. The nests and life cycles of the two bee taxa differ in some ways. Honeybee colonies are perennial, consisting of workers and drones numbering in the thousands and a healthy fertilized queen that can live for up to five years [1]. Bumblebees are seasonal with an annual life cycle, with a few hundred workers and males composing a colony, and in some species, a population peak of over 1000 bees has been documented [2,3].
Males mainly hatch from haploid unfertilized eggs, while females (workers and queens) develop from diploid fertilized eggs [1,2]. Diploid males are also produced in both honeybees and bumblebees, and their production may pose an increased risk to the survival of the bee colonies [4,5]. The laying of male-destined eggs and the production of males are normally initiated during the reproductive season, when a sufficient population of attending workers and food resources are available [1]. Development from egg to adult differs among species, taking a relatively long time in bumblebees. On average, development in bumblebee males takes 24–28 days [6], while development takes approximately 24 days in male honeybees [1,7]. Several factors, such as the overall colony condition, temperature (brood nest), number of workers, and quality and availability of food resources (nutrients), have a crucial influence on the development time and quality of the male bumblebees produced [2].
The primary role of males in bee colonies is mating, which is a fundamental reason why they are produced only when needed. Their reproductive ability begins and completes development at early stages before emergence (larval and pupal stages). After emergence, males gradually mature to copulate and transmit sperm to the spermathecae of queens. Estimates of sexual maturation range between 6–20 days after emergence in bumblebees [6] and 6–16 days in honeybees [7]. Efficient copulation and sperm viability are closely related to male quality characteristics such as birth weight, age, body size, and environmental conditions. Apart from their exclusive reproductive role, male bees contribute to heat regulation in the colony, although their contribution is no more than that of worker bees [8].
Male honeybees are unable to engage in foraging behavior as they lack food collection body parts [9]. Usually, during the dearth of food or at the end of the reproductive season, the laying of male-destined eggs halts, and adult males are no longer needed and are thus forcefully evicted from the colony [1,9,10]. Other factors have also been suggested to contribute to the eviction of drones, e.g., reduced environmental temperature, the presence of workers with developed ovaries, the presence of a queen, the age of the queen, the amount of sealed and unsealed brood, the activity of the colony, the amount of forage being collected, the amount and condition of honey stores, and the strain of the bees [10]. Evicted males starve since they lack food collection body parts such as a honey stomach and pollen basket for foraging [1,9]. In contrast, multiple studies have documented the foraging and pollination abilities of male bumblebees [11,12,13,14,15]. Mature male bumblebees leave their nest in search of virgin queens with which to mate and engage in foraging and flower pollination [13,15]. Fliszkiewicz and colleagues noted the pollination effect of male Bombus terrestris to be similar to that of worker bees [14], and when trained to identify fodder color, freely flying male B. terrestris behave as workers in terms of learning speed and accuracy [12]. However, they need to make a trade-off between foraging and mating with queens. Male bumblebees can also act as an excellent alternative model system for further studies of pollination capacity due to their ability to successfully pollinate some crops, such as the blackcurrant Ribes nigrum and Gentiana parryi (Gentianaceae) [13,14,15].
2. Reproductive Ability of Male Bees
Mating behavior is an aspect of the reproductive ability of male bees that influences the offspring of the colony. Although successful mating is fatal for male honeybees, honeybee queens can mate with multiple males to store sufficient sperm. This possibility of nonvirgin queens mating again is unfavorable for Apis mellifera drones in terms of passing on their genes [16,17]. To prevent the mated queens from flying out of the colonies to mate again, male semen changes the expression of genes associated with vision in the queens’ brains, weakening the reaction of their compound eyes and ocelli to light. This makes it difficult for a mated queen that has flown to mate again to return to the colony, causing her to face the risk of death [18]. The selective pressure of sperm competition may cause the production of higher-quality sperm in male honeybees than those monandrous bumblebee species [19]. In contrast to the fatal fate of male honeybees after successful mating, male bumblebees are able to mate multiple times. Moreover, the mating frequency of queens varies widely between different species of bumblebees, for instance, the queens of B. terrestris, B. lantschouensis, B. patagiatus and B. ignitus mate only once in their lifetime, while queens of B. hypnorum can mate up to six times [20,21]. Queens that mate with nonvirgin males can produce more workers and males and are thus more successful in building colonies [22]. Multiple mating may be closely related to the length of sperm and the selection for longer sperm, which can be related to body size [23]. Male bumblebees prevent the sperm of other males from entering the queen’s genital tract with the placement of a mating plug that reduces sperm competition. For instance, in B. terrestris, the ubiquitous and rather unspecific fatty acid linoleic acid composition of the male mating plug prevents females (queens) from further mating [24]. Male characteristics such as age and weight influence the duration of copulation and mating success. A short copulation duration could delay the establishment of colonies, although it has been found to have no impact on the production of new queens and males [25]. Environmental factors that may affect the mating process have been described in previous reviews, such as the temperature of the hive and drone nutrients [11,26]. Furthermore, the mating rate differs in different species and has been found to be affected by internal factors in both bumblebees and honeybees. In this regard, there might be some unique factors that control the mating behavior of males of different bumblebee species, which is very important to the knowledge of social insects.
In addition to mating behavior, sperm quality is a key factor influencing successful reproduction in bee colonies since semen will be stored for more than three years in the spermathecae of honeybee queens [1] and for several months in the spermathecae of bumblebee queens after insemination. Several parameters have been used to evaluate the reproductive ability of male bees, such as sperm motility, sperm viability, and acrosomal integrity. Sperm motility is important for the transfer of sperm into the queen’s spermathecae [27] and has been used as an essential indicator for estimating sperm quality [28,29]. Sperm viability is closely linked to the fertility of queens and is thus widely assayed to evaluate semen quality [30,31]. However, sperm quality is irrelevant to queen longevity, which is associated with the mating behavior or substances received from mating in eusocial insects such as ants [32]. However, sperm influence the hibernation success and fitness of inseminated females, and sperm of different male genotypes show distinct female longevity in generally monandrous B. terrestris [33,34]. Most studies related to the cryopreservation of frozen semen regard acrosomal integrity as an indicator of digestion of the zona pellucida and the fusion ability of sperm [35,36]. Yaniz and colleagues summarized most of the parameters and several kinds of methods for the analysis of semen quality in honeybees [37]. Poor male reproductive ability may affect health and productivity by influencing queen fecundity, which determines subsequent worker behavioral or physiological traits and is influenced by semen fluid and viable sperm [38]. Moreover, multiple mating increases the genetic diversity of sperm in queens, swarms headed by which show great advantages in the speed of founding new colonies, foraging rates, food storage, drone population, winter survival and pathogen resistance [39,40,41]. In the current paper, we review the factors affecting sperm quality in ejaculate and spermathecae in both honeybees and bumblebees (Figure 1).
3. Influence of External Environmental Factors on the Sperm Quality of Drones
3.1. Nutrients
Nutrients have been proven to be important for improving the reproductive quality of drones. Nectar and pollen are the prime nutrient resources used for adult and larval development in bees, and the nutritional requirements of different bees, such as honeybees and bumblebees, exhibit preferences [42,43]. Limited access to pollen during larval development in honeybee drones results in less semen and an increased likelihood of ejaculation failure [44]. The pollen species composition is a very important factor that greatly influences the population of bee colonies [45]. A colony of B. terriestris fed pollen with a high protein content produces the highest number of males compared to those fed pollen with a low protein content [46]. When colonies were fed sucrose syrup and protein supplements, the weight and abdominal size of the drones increased significantly. In contrast, the sperm volume and motility of each mature drone declined significantly in the colonies that received no additional nutrients [47]. A lack of protein intake after the emergence of drones does not affect sperm motility, indicating that sperm development is completed before emergence and that sexually mature drones do not need additional protein [48]. Pelletier and McNeil found that in bumblebees, food supplementation can increase the number of males and the probability of producing gynes (young queens) [49]. In addition, the bumblebee adult body size depends on the amount of food received [50,51]. Relatively large males have an advantage in male–male competition in some species [52,53,54].
The season also shows an obvious effect on the semen volume and sperm number. The maximum number of honeybee drone broods are produced in June and July [55]. Drones release significantly higher semen volumes in spring than in summer and autumn. However, autumn-reared drones produce more sperm than summer- and spring-reared drones, which produce the least sperm [56]. Seasonal factors influence drone production, including the day length and temperature, size of the colonies and availability of food [57].
However, fewer studies have addressed the effects of nutrients and seasons on the quality of sperm in bumblebees than in honeybees. Based on the species-specific characteristics of reproduction in bumblebees, more detailed and explicit studies on the influence of the nutrient dose, nutrient composition, nutrient ratio and feeding time point on drone quality attributes, such as the sperm count and viability, should be performed among different species in future research.
3.2. Temperature
A. mellifera drones incubated at the capped brood stage at a relatively low temperature (32–35 °C) deliver less semen but have higher sperm viability [58]. In the process of the in vitro storage of drone sperm, extreme temperatures can significantly reduce the viability of stored sperm. Preserving the fresh ejaculate of A. mellifera at high or low temperatures resulted in a significant increase in dead sperm by up to 40% [59,60]. Moreover, temperatures are variable during A. mellifera young queen shipment, and both low and high temperatures significantly decrease sperm viability [61]. Exposure to high-temperature extremes (45 °C) can cause more than 50% of sperm to die in the spermathecae of queens [59]. The optimal temperature (30–35 °C) significantly increases the number of living sperm after insemination [59]. Thus, the optimal temperature is a critical environmental factor affecting the maintenance of the sperm quality of drones and the storage of sperm in vitro at certain times.
A study of drones of twenty-one bumblebee species showed that male bumblebees tend to evolve larger body sizes in locations where rain occurs, mostly during summer, when the overall temperature is warmer [3]. The male size can directly impact the B. terrestris mating success [62]. In comparison to honeybees, bumblebees exhibit greater species diversity and occur over more complex environments and thus serve as a good model for studying the influence of climate change on bees and other insects [63]. Studies on the impact of high temperatures on the reproductive ability of bumblebees are crucial for the evaluation of the threat of global warming to insect colonies. Moreover, for bumblebee queens that live alone to pass through diapause at cold temperatures, the effects of cold temperatures on the storage of sperm in queens must be evaluated because they have important influences on the reproduction and protection of bumblebees.
3.3. Pesticides
Widely used pesticides have caused controversy due to their adverse effects on bee health, including those related to sperm viability in male bees and the storage of sperm in queen bees. Neonicotinoids, which are registered for use on over 140 crops, are the most widely used pesticides in more than 120 countries [64]. The neonicotinoids imidacloprid and thiamethoxam have significantly negative effects on the sperm quality of male bees. After exposure to thiamethoxam at a concentration of 4.5 ppb, the life span of male bees was significantly shortened, and the number of living sperm decreased by 39% [65]. After being exposed to an extremely high concentration of imidacloprid (0.02 ppm), the sperm motility of drones significantly decreased, although this value significantly varied among different colonies [29,66]. When bee colonies were exposed to neonicotinoid pesticides, the asymmetry of forewing veins and wing morphology fluctuation significantly increased in honeybee drones. The honeybee queen spermatheca is also highly susceptible to pesticides. Antioxidant genes in the spermatheca are inhibited after treatment with a low dose of coumaphos (5 ppm) and a sublethal dose of imidacloprid (0.02 ppm) [67]. Exposure to a sublethal dose of imidacloprid (0.02 ppm) or a high dose of coumaphos (100 ppm) can result in a decline in sperm viability by 50% or 33% in the spermatheca, respectively [67].
Another common type of chemical exposure is exposure to pesticides used for bee disease prevention by beekeepers, such as amitraz, which is used to control Varroa destructor, a major pest of honeybees. Sublethal doses of amitraz had no effect on the viability of sperm stored in honeybee queens under laboratory conditions or sperm viability. However, it affected development and downregulated genes related to detoxification, cyclic adenosine monophosphate (cAMP)-dependent protein kinase, immunity, and antioxidant capacity [68].
In addition, fipronil syrup is a phenylpyrazole insecticide. After eating it at a concentration of 0.1 mg/L, the sperm concentration and sperm motility of honeybee drones were found to decrease significantly, and the sperm metabolic rate increased, leading to reduced fertility in the drones [69]. Many moderately toxic and low-toxicity pesticides, such as fungicides and herbicides, also have lethal and sublethal impacts on bees [6].
Overall, research studies on the effects of pesticides on bees are mainly conducted in Europe and North America, assessed in honeybees under laboratory conditions, and measured at the individual level [70]. Low doses or nonlethal doses of diverse pesticides might not cause the death of bees but may decrease some of their abilities, such as reproduction. Therefore, we suggest that studies should combine the effects on individual bees and on bee colonies of more bee species to discuss the toxicity of neonicotinoids at the population level and how the effects pose a threat to the reproductive ability of male bees. The effects of low or nonlethal doses of diverse pesticides on the sperm quality and foraging behavior of male bees should be evaluated in detail. Moreover, some insecticides used by beekeepers to prevent bee diseases should be assessed further in future studies, evaluating their influence on the sperm quality of drones and the storage of sperm in queens.
4. Influence of the Internal Environment on the Sperm Quality of Drones
4.1. Age and Birth Weight
Several studies have reported that honeybee sperm production, maturation and viability are closely related to age. Hölldobler and Bartz reported that sperm production in social insect males ceases upon eclosion [71]. The honeybee drone also ecloses with all the sperm [72], and then the sperm move from the testes to the seminal vesicles, during which the sperm or seminal fluid gradually matures with age (6–14 days after eclosion). The semen volume in the seminal vesicles gradually decreases, the sperm density increases, and the composition of the ejaculate changes [73]. The age of drones displays a continuing effect on the semen volume and sperm concentration. For instance, 14-day-old and 21-day-old drones release more semen volume in the ejaculate than 35-day-old drones, and 21-day-old drones release higher sperm counts in the ejaculate than 14-day-old and 35-day-old drones that share similar sperm numbers [56]. Ruttner and Tarpy noted that the sperm counts in the seminal vesicles reached 7,390,000 at 20 days after emergence and decreased significantly 30 days later [74]. In addition to the semen volume and sperm counts, sperm viability is closely correlated with drone age. Locke and Peng reported that sperm viability decreased significantly with increasing drone age, while motility patterns did not change [75]. Sturup found that the sperm viability of A. mellifera decreases with drone age, with differences among colonies, some of which could delay ejaculation senescence [48]. However, recent studies show that drone age and time of breeding have no effect on sperm viability [74,76]. Moreover, sperm DNA fragmentation and the coagulation and proteolysis of semen are closely related to drone age and have notable effects on sperm. The coagulation of semen occurs in old drones, and these sperm display the lowest sperm surface protease activities, with both characteristics negatively affecting sperm viability [77,78]. The correlation between sperm DNA fragmentation and male age is controversial in animals based on current studies and exhibits a significantly negative effect on male fertility [79]. Unfortunately, studies paid scant attention to the sperm DNA fragmentation alongside drone age in bees. Currently, we only know that 14-day-old honeybee drones possess significantly higher DNA fragmentation rates in sperm than those collected directly from queens of different ages [80], hence, it is meaningful to explore the DNA fragmentation rate in male bees from different age and the connection with sperm quality. Studies in the bumblebee B. terrestris have noted that age and weight influence the duration of copulation. Younger males copulated more quickly and copulated for a shorter duration than old males, and relatively heavy males of B. terrestris copulated more rapidly than lighter males [62]. However, studies on the relationships between the age, birth weight, and sperm viability of males are currently lacking for bumblebees. The ages of males and queens are critical to mating success and the development of offspring colonies in bumblebees.
4.2. Body Size
Czekonska reported that the weight of honeybee drones at birth is an important factor in predicting the survival of drones in colonies and that males that were heavier at birth reached sexual maturity 15 days after emergence and lived longer than males that were lighter at birth [81]. In queenright colonies (QRCs) of honeybees where the queens are in charge, reproductive workers produce only 7% of the unfertilized eggs in the colony. When the queen is lost, the expression level of vitellogenin in nonreproductive workers increases, and workers from different subfamilies of honeybees all tend to lay eggs with different success rates [82,83]. Drones produced in the drone cells (DCs) of QRCs are 17% heavier than those produced in the DCs of laying worker colonies (LWCs), while drones produced in worker cells (WCs) in LWCs have the smallest body size and lightest weight [84]. The length of the wing is thought to be an effective indicator for distinguishing large drones (LDs) from small drones (SDs) since the length and width of the front wing of LDs are significantly larger than those of SDs [85]. Body size is closely related to the sperm count of male bees. The average sperm count and accessory mucous gland and seminal vesicle weights of LDs were found to be significantly higher than those of SDs [84,86]. The SDs are only 87% of those of normal drones on average, and the sperm count is also significantly lower than that of normal-size drones [87].
Moreover, body size also affects the mating success of male bees. Couvillon and colleagues found that within a day, the flight activity of LDs peaked at noon, which is consistent with the peak in queen activity, while the flight activity of SDs was more uniform throughout the day [88]. Gençer and Kahya quantified the sperm competitiveness of SDs reared in laying worker colonies (LWCs) against LDs reared in QRCs and found that SDs were not as successful as LDs in terms of sperm competition. Gencer and Kahya mixed the sperm of two types of male bees of A. mellifera in different proportions for artificial insemination of queens. The paternal frequencies in the offspring of SDs were all lower than expected, while large males produced more offspring than expected [89]. Although SDs remained slightly behind LDs in regard to sperm competition, this does not imply that SDs reared in LWCs are useless. In addition, the concentration of sperm in the ejaculate affects the reproductive success of drones, and evolution forces drones to produce not only plentiful but also particularly concentrated semen [89].
The somatotype dimorphism of males is more obvious in bumblebees than in honeybees. Bumblebee workers are also capable of producing male offspring from unfertilized, haploid eggs, but parentage analyses suggest that only a low proportion (approximately 5%) of adult males are worker produced [90,91]. Lacking queen-specific chemical cues, worker-laid eggs can be distinguished and quickly cleared by other workers in the bumblebee B. terrestris [92,93]. An observation in our lab showed that males from the QRCs of B. terrestris have a distinctly larger body size than that produced by the workers (unpublished data). Large males may have relatively large genitalia that act as particularly strong forceps for attachment to the queen’s sting apparatus [94,95]. Large/young drones initiate mating more quickly and copulate for less time than lighter, older drones. The lengths of the front and hind feet affect mating success as age becomes less important in competitive situations, such that drones with longer feet have higher mating success than drones with shorter feet [62]. Moreover, males from a queenright bumblebee colony also exhibited diverse body sizes. Sperm length was found to be positively correlated with male body size in B. terrestris and possibly in B. hypnorum [23]. Therefore, somatotype dimorphism in male bees, especially bumblebees, can best explain the influence of drone size on the colony. However, to the best of our knowledge, no research explains its influence on sperm quality and queen fecundity. Body size is an important factor for breeding queens in both bumblebees and honeybees and should be given more attention in further research.
4.3. Gland Proteins
Male insect semen contains sperm cells and seminal fluid components [96,97,98,99,100]. Seminal fluid is composed of proteins and other small molecules, including peptides, sugars, and lipids and is primarily derived from the male accessory glands [96,101]. Furthermore, how seminal fluid and seminal fluid proteins (SFPs) influence male and female fertility and behavior, sperm viability, and susceptibility has been intensively studied for decades in Drosophila. Proteins from the accessory glands of D. melanogaster males have been proven to be an essential factor influencing the transfer of sperm and play an important role in the storage of sperm in females. For example, accessory gland proteins are needed for morphological changes in the female reproductive tract that may be important for sperm storage [102]. Seminal fluids have been studied in other insects, including mosquitos, crickets, ants, moths, and beetles [103,104,105,106,107,108,109,110,111,112,113].
In honeybees, the activity of sperm in ejaculated semen is usually higher than that of sperm stored in the spermathecae, which benefit from the proteins in the semen [114]. Greeff found that the proportion of dead sperm in the testes of bumblebee drones was significantly lower than that in the spermathecae of newly mated queens and old queens [115]. The higher sperm viability might result from the components of the semen. The semen of A. mellifera is so effective at keeping sperm alive that the positive effects can last up to 24 h. The presence of proteins in semen and their structural integrity are critical to this effect, which cannot be replicated by ordinary protein substitutes [116]. Moreover, semen fluids are largely responsible for stimulating postmating changes in queen behavior and physiology, e.g., reduced sexual receptivity, reduced attraction to light, reduced mating flight, ovary activation, ovulation, modulation of pheromone production and transcriptional changes in queens [117,118]. Honeybee drone seminal fluid could induce a decline in queen vision by causing substantial gene expression changes in the brain and perturbing the phototransduction pathway to reduce queen promiscuity across mating flights [18]. This regulatory mechanism is comparable to earlier findings in Drosophila, with changes triggered by the sex peptide [119]. Similar changes in gene expression of the phototransduction pathway, neuroactive ligand-receptor interactions, the Hippo signaling pathway and the phagosome pathway were found in the postmating queens of B. terrestris [120]. Mass spectrometry has been used to identify the proteins in honeybee drone semen [99], accessory gland-associated proteins [99], seminal fluid [96,121,122,123,124], and sperm cell-associated proteins [114]. Approximately 260 proteins have been identified in honeybee seminal fluid [122]. Proteins in the accessory mucous gland of drones are the main component of semen, and the proteins in accessory glandular fluid significantly improve sperm motility [125]. Further research on Drosophila has identified some specific proteins that reduce female sexual receptivity and benefit rival sperm (i.e., sex peptide) [126], maintain sperm viability (i.e., Acp29AB), promote uterine contractions (i.e., Acp36DE), and promote ovulation (i.e., ovulin [127]). Interestingly, homologs of these specific proteins have not been identified within the honeybee genome, underscoring the uniqueness of the honeybee mating system and the need for specific investigations into this system [38]. Furthermore, the homologs of these specific proteins have not been identified in known bumblebee genomes that exhibit some distinctions from the honeybee genome.
In addition, both bumblebee and honeybee queens have a special organ, the spermatheca, for the long-term storage of sperm throughout their lifetime. This is especially true for most bumblebee species, which must experience a diapause period at cold temperatures. Insect spermathecae have associated secretory cells (spermathecal secretory cells or SSCs) that produce proteins and other molecules that function in sperm storage [128,129,130]. Female reproductive fluid (FRF), which significantly affects sperm traits, including chemoattraction and alterations in sperm velocity, has been shown to exert positive phenotypic effects on sperm competition in males [131]. After mating in Drosophila [132], Apis [133], Crematogaster [134], and Anopheles [135], molecules involved in the immune response, carbohydrate and lipid metabolism, cellular transport and oxidative stress have been identified in the transcriptional and proteomic profiles of sperm storage organs and might play protective roles in sperm and/or mediate female postmating processes. Interestingly, the spermathecae of A. mellifera and Crematogaster osakensis ant queens also possess some important proteins that can significantly improve sperm motility and are involved in the long-term maintenance of stored sperm [125,134]. The vesicular gland is the main contributor to the proteome of the spermathecae, but there is a more complete protein network in this organ that is conducive to long-term sperm storage [96,133]. Hundreds of proteins representing the main components of spermathecal fluid have been identified. They belong to a series of different functional groups, the most obvious of which are energy metabolism enzymes and antioxidant defense enzymes [133]. The transcripts of two antioxidative enzymes, catalase and glutathione-transferase (GST), are ten to twenty times higher in the spermathecae of mating queens than in those of unmated queens, and the expression levels of the antioxidant genes TXN2 and TXNRD1 in the spermathecae of mating queens are also higher than those in the spermathecae of nonmating queens to protect the sperm from damage [105,136,137]. Furthermore, some metabolites were identified and found to be enriched in metabolic pathways, including glycerophospholipid (GPL) metabolism, the biosynthesis of amino acids, and the mTOR signaling pathway, likely contributing to the long-term maintenance and protection of sperm [138]. In summary, SFPs and gland proteins in spermathecae exhibit diverse functions and play a significant role in the reproduction of insects (Table 1). Therefore, the function of SFPs and gland proteins in spermathecae in terms of sperm quality should be studied in bumblebees and honeybees, as this information is very important for the long-term storage of sperm in vitro and the artificial regulation of the reproduction of bumblebees and honeybees in the future.
5. Further Research Directions
Throughout the review, knowledge of the effects of age, birth weight, and body size on sperm viability is currently lacking for bumblebees, which is important for copulation by useful drones for bumblebee breeding. Fewer studies have reported the effects of nutrients on the quality of sperm in bumblebees compared to honeybees, and this should be addressed in further research [44,47]. Furthermore, the impacts of pollen diversity and seasons on male development and sperm quality need to be clarified. Moreover, the SFPs and gland proteins in the spermathecae of bumblebees should be identified, and the functions of SFPs and gland proteins in spermathecae in terms of sperm quality in long-term storage of sperm and postmating changes of queens in bumblebees and honeybees should be studied in further research. Moreover, the mechanism by which SFPs or sperm contribute to the hibernation success of bumblebees should be given more attention. Current studies on the exposure of drones to pesticides mainly focus on the negative effect on sperm viability [66,67,68,69]; however, pesticides may change the mating behavior of drones, which is crucial to mating success. Therefore, we suggest that more research on the impacts of various pesticides on drones should cover the mating behaviors of drones exposed to pesticides.
In summary, the current studies on male bumblebees are still much less thorough than those on male honeybees. Hence, more studies should be systematically carried out on male bumblebees that differ from male honeybees in terms of biology and lifecycle. More in-depth research regarding the factors that influence male reproduction is needed in honeybees and bumblebees, which will benefit the comprehensive evaluation of male bees’ reproductive ability in bee breeding. We highlight the knowledge gaps of current studies on the reproductive ability of male bees, it is meaningful for researchers to pay close attention and conduct in-depth research to fill these knowledge gaps. More factors, such as DNA fragmentation, genetic abnormalities, mitochondrial DNA mutations, infections, and signaling mechanisms involved in sperm motility that are thought to affect the sperm quality of humans, might also need to be considered in further research.
Author Contributions
Conceptualization, Y.L. and J.A.; writing—original draft preparation, H.Z.; writing—review and editing, S.K.M., Y.L. and J.A.; supervision, J.A.; funding acquisition, Y.L. and J.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by and the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2015-IAR) and the Special Program for Basic Scientific Research of Chinese Academy of Agricultural Sciences (CAAS-2021-LiuYanJie).
Acknowledgments
We are very thankful to Jianghong Li and three anonymous reviewers for their comments to this paper.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Flottum, K. The Backyard Beekeeper: An Absolute Beginner’s Guide to Keeping Bees in Your Yard and Garden, 4th ed.; Quarry Books: Beverly, MA, USA, 2018. [Google Scholar]
- Goulson, D. Bumblebees Behaviour and Ecology; Oxford University Press: New York, NY, USA, 2003. [Google Scholar]
- Cueva del Castillo, R.; Sanabria-Urbán, S.; Serrano-Meneses, M.A. Trade-offs in the evolution of bumblebee colony and body size: A comparative analysis. Ecol. Evol. 2015, 5, 3914–3926. [Google Scholar] [CrossRef] [PubMed]
- Betti, M.I.; Lee, I. The effects of diploid male production on honey bee colony evolution and survival. Theor. Popul. Biol. 2020, 135, 49–55. [Google Scholar] [CrossRef]
- Duchateau, M.J. Sexual biology of haploid and diploid males in the bumble bee Bombus terrestris. Insect Soc. 1995, 42, 255–266. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Effects of fungicide and herbicide chemical exposure on Apis and non-Apis bees in agricultural landscape. Front. Environ. Sci. 2020, 8, 81. [Google Scholar] [CrossRef]
- Rangel, J.; Fisher, A. Factors affecting the reproductive health of honey bee (Apis mellifera) drones—A review. Apidologie 2019, 50, 759–778. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.M. Roles of individual honeybee workers and drones in colonial thermogenesis. J. Exp. Biol. 1987, 129, 53–61. [Google Scholar] [CrossRef]
- Rhodes, J.W. Drone Honey Bees—Rearing and Maintenance. Agnote DAI. 2002. Available online: https://powell.ca.uky.edu/files/drone-bee-rearing-and-maintenance_002_0.pdf (accessed on 1 June 2021).
- Free, J.; Williams, I.H. Factors determining the rearing and rejection of drones by the honeybee colony. Anim. Behav. 1975, 23, 650–675. [Google Scholar] [CrossRef]
- Belsky, J.E.; Camp, A.A.; Lehmann, D.M. The importance of males to bumble bee (Bombus species) nest development and colony viability. Insects 2020, 11, 506. [Google Scholar] [CrossRef]
- Wolf, S.; Chittka, L. Male bumblebees, Bombus terrestris, perform equally well as workers in a serial colour-learning task. Anim. Behav. 2016, 111, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Ogilvie, J.E.; Thomson, J.D. Male bumble bees are important pollinators of a late-blooming plant. Arthropod-Plant Inte. 2015, 9, 205–213. [Google Scholar] [CrossRef]
- Fliszkiewicz, M.; Giejdasz, K.; Wilkaniec, Z. The importance of male red mason bee (Osmia rufa L.) and male bufftailed bumblebee (Bombus terrestris L.) pollination in blackcurrant (Ribes nigrum L.). J. Hortic. Sci. Biotechnol. 2011, 86, 457–460. [Google Scholar] [CrossRef]
- Oatevik, K.; Manson, J.; Thompson, J.; Ostevik, K.; Thomson, J. Pollination potential of male bumble bees (Bombus impatiens): Movement patterns and pollen-transfer efficiency. J. Pollinat. Ecol. 2010, 2, 21–26. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Page, J.R.E. No behavioral control over mating frequency in queen honey bees (Apis mellifera L.): Implications for the evolution of extreme polyandry. Am. Nat. 2000, 155, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdelkader, F.; Kairo, G.; Tchamitchian, S.; Cousin, M.; Senechal, J.; Crauser, D.; Vermandere, J.P.; Alaux, C.; Le Conte, Y.; Belzunces, L.P.; et al. Semen quality of honey bee drones maintained from emergence to sexual maturity under laboratory, semi-field and field conditions. Apidologie 2014, 45, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Liberti, J.; Görner, J.; Welch, M.; Dosselli, R.; Schiøtt, M.; Ogawa, Y.; Castleden, I.; Hemmi, J.M.; Baer-Imhoof, B.; Boomsma, J.J.; et al. Seminal fluid compromises visual perception in honeybee queens reducing their survival during additional mating flights. eLife 2019, 8, e45009. [Google Scholar] [CrossRef] [PubMed]
- Hunter, F.; Birkhead, T. Sperm viability and sperm competition in insects. Curr. Biol. 2002, 12, 121–123. [Google Scholar] [CrossRef] [Green Version]
- Paxton, R.J.; Thorén, P.A.; Estoup, A.; Tengö, J. Queen–worker conflict over male production and the sex ratio in a facultatively polyandrous bumblebee, Bombus hypnorum: The consequences of nest usurpation. Mol. Ecol. 2001, 10, 2489–2498. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z.; Huang, J.; Yuan, X.; Ding, G.; An, J. Queen traits and colony size of four bumblebee species of China. Insect Soc. 2018, 65, 537–547. [Google Scholar] [CrossRef]
- Gosterit, A.; Gurel, F. Male remating and its influences on queen colony foundation success in the bumblebee, Bombus terrestris. Apidologie 2016, 47, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Baer, B.; Schmid-Hempel, P.; Boomsma, J.J. Sperm length, sperm storage and mating system characteristics in bumblebees. Insect Soc. 2003, 50, 101–108. [Google Scholar] [CrossRef]
- Baer, B.; Morgan, E.D.; Schmid-Hempel, P. A nonspecific fatty acid within the bumblebee mating plug prevents females from remating. Proc. Natl. Acad. Sci. USA 2001, 98, 3926–3928. [Google Scholar] [CrossRef] [Green Version]
- Amin, R.; Than, K.K.; Kwon, Y.J. Copulation duration of bumblebee Bombus terrestris (Hymenoptera: Apidae): Impacts on polyandry and colony parameters. J. Asia-Pac. Èntomol. 2009, 12, 141–144. [Google Scholar] [CrossRef]
- Rezende, E.L.; Bozinovic, F.; Szilágyi, A.; Santos, M. Predicting temperature mortality and selection in natural Drosophila populations. Science 2020, 369, 1242–1245. [Google Scholar] [CrossRef]
- Ruttner, F.; Koeniger, G. Die Füllung der Spermatheka der Bienenkünigin. J. Comp. Physiol. A 1971, 72, 411–422. [Google Scholar] [CrossRef]
- Wegener, J.; May, T.; Knollmann, U.; Kamp, G.; Müller, K.; Bienefeld, K. In vivo validation of in vitro quality tests for cryopreserved honey bee semen. Cryobiology 2012, 65, 126–131. [Google Scholar] [CrossRef]
- Ciereszko, A.; Wilde, J.; Dietrich, G.J.; Siuda, M.; Bąk, B.; Judycka, S.; Karol, H. Sperm parameters of honeybee drones exposed to imidacloprid. Apidologie 2017, 48, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.; Donoghue, A. Viability assessment of honey bee, Apis mellifera, sperm using dual fluorescent staining. Theriogenology 1999, 51, 1513–1523. [Google Scholar] [CrossRef]
- Collins, A.M. Relationship between semen quality and performance of instrumentally inseminated honey bee queens. Apidologie 2000, 31, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Schrempf, A.; Heinze, J.; Cremer, S. Sexual cooperation: Mating increases longevity in ant queens. Curr. Biol. 2005, 15, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.; Schmid-Hempel, P. Sperm influences female hibernation success, survival and fitness in the bumble–bee Bombus terrestris. Proc. R. Soc. B Biol. Sci. 2005, 272, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Korner, P.; Schmid-Hempel, P. Effects of sperm on female longevity in the bumble-bee Bombus terrestris L. Proc. R. Soc. B Biol. Sci. 2003, 270, S227–S229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hezavehei, M.; Sharafi, M.; Kouchesfahani, H.M.; Henkel, R.; Agarwal, A.; Esmaeili, V.; Shahverdi, A. Sperm cryopreservation: A review on current molecular cryobiology and advanced approaches. Reprod. BioMed. Online 2018, 37, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Alcay, S.; Cakmak, S.; Cakmak, I.; Mulkpinar, E.; Gokce, E.; Ustuner, B.; Sen, H.; Nur, Z. Successful cryopreservation of honey bee drone spermatozoa with royal jelly supplemented extenders. Cryobiology 2019, 87, 28–31. [Google Scholar] [CrossRef]
- Yániz, J.L.; Silvestre, M.A.; Santolaria, P. Sperm quality assessment in honey bee drones. Biology 2020, 9, 174. [Google Scholar] [CrossRef]
- Brutscher, L.M.; Baer, B.; Niño, E.L. Putative drone copulation factors regulating honey bee (Apis mellifera) queen reproduction and health: A review. Insects 2019, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.D.; Tarpy, D. Queen promiscuity lowers disease within honeybee colonies. Proc. R. Soc. B Biol. Sci. 2006, 274, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Mattila, H.R.; Seeley, T.D. Genetic diversity in honey bee colonies enhances productivity and fitness. Science 2007, 317, 362–364. [Google Scholar] [CrossRef] [Green Version]
- Tarpy, D.R. Genetic diversity within honeybee colonies prevents severe infections and promotes colony growth. Proc. R. Soc. B Biol. Sci. 2003, 270, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee nutrition and floral resource restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Èntomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Czekońska, K.; Chuda-Mickiewicz, B.; Samborski, J. Quality of honeybee drones reared in colonies with limited and unlimited access to pollen. Apidologie 2014, 46, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vanderplanck, M.; Moerman, R.; Rasmont, P.P.; Lognay, G.C.; Wathelet, B.; Wattiez, R.; Michez, D. How does pollen chemistry impact development and feeding behaviour of polylectic bees? PLoS ONE 2014, 9, e86209. [Google Scholar] [CrossRef] [PubMed]
- Billiet, A.; Meeus, I.; Van Nieuwerburgh, F.; Deforce, D.; Wäckers, F.; Smagghe, G. Impact of sugar syrup and pollen diet on the bacterial diversity in the gut of indoor-reared bumblebees (Bombus terrestris). Apidologie 2015, 47, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, A.; Giovenazzo, P. Optimizing drone fertility with spring nutritional supplements to honey bee (Hymenoptera: Apidae) colonies. J. Econ. Èntomol. 2016, 109, 1009–1014. [Google Scholar] [CrossRef]
- Sturup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When every sperm counts: Factors affecting male fertility in the honeybee Apis mellifera. Behav. Ecol. 2013, 24, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, L.; McNeil, J.N. The effect of food supplementation on reproductive success in bumblebee field colonies. Oikos 2003, 103, 688–694. [Google Scholar] [CrossRef]
- Sutcliffe, G.H.; Plowright, R.C. The effects of pollen availability on development time in the bumble bee Bombus terricola K. (Hymenoptera: Apidae). Can. J. Zoöl. 1990, 68, 1120–1123. [Google Scholar] [CrossRef]
- Sutcliffe, G.; Plowright, R. The effects of food supply on adult size in the bumble bee Bombus terricola kirby (Hymenoptera: Apidae). Can. Èntomol. 1988, 120, 1051–1058. [Google Scholar] [CrossRef]
- Paxton, R.J. Male mating behaviour and mating systems of bees: An overview. Apidologie 2005, 36, 145–156. [Google Scholar] [CrossRef]
- Williams, P.H. The bumble bees of the Kashmir Himalaya (Hymenoptera: Apidae, Bombini). Bull. Br. Mus. 1991, 60, 1–204. [Google Scholar]
- Alcock, J.P. Male behaviour in two bumblebees, Bombus nevudensis auricomus and B. griseicollis (Hymenoptera: Apidae). J. Zool. 1983, 200, 561–570. [Google Scholar] [CrossRef]
- Rowland, C.M. Seasonal changes of drone numbers in a colony of the honeybee, Apis mellifera. Ecol. Model. 1987, 37, 155–166. [Google Scholar] [CrossRef]
- Rhodes, J.W.; Harden, S.; Spooner-Hart, R.; Anderson, D.L.; Wheen, G. Effects of age, season and genetics on semen and sperm production in Apis mellifera drones. Apidologie 2011, 42, 29–38. [Google Scholar] [CrossRef]
- Boes, K.E. Honeybee colony drone production and maintenance in accordance with environmental factors: An interplay of queen and worker decisions. Insect Soc. 2010, 57, 1–9. [Google Scholar] [CrossRef]
- Czekońska, K.; Chuda-Mickiewicz, B.; Chorbiński, P. The effect of brood incubation temperature on the reproductive value of honey bee (Apis mellifera) drones. J. Apic. Res. 2013, 52, 96–105. [Google Scholar] [CrossRef]
- Bienkowska, M.; Panasiuk, B.; Wegrzynowicz, P.; Gerula, D. The effect of different thermal conditions on drone semen quality and number of spermatozoa entering the spermatheca of queen bee. J. Apic. Sci. 2011, 55, 161–168. [Google Scholar]
- McAfee, A.; Chapman, A.; Higo, H.; Underwood, R.; Milone, J.; Foster, L.J.; Guarna, M.M.; Tarpy, D.R.; Pettis, J.S. Vulnerability of honey bee queens to heat-induced loss of fertility. Nat. Sustain. 2020, 3, 367–376. [Google Scholar] [CrossRef]
- Rousseau, A.; Houle, É.; Giovenazzo, P. Effect of shipping boxes, attendant bees, and temperature on honey bee queen sperm quality (Apis mellifera). Apidologie 2020, 1–12. [Google Scholar] [CrossRef]
- Amin, M.R.; Bussière, L.F.; Goulson, D. Effects of male age and size on mating success in the bumblebee Bombus terrestris. J. Insect Behav. 2011, 25, 362–374. [Google Scholar] [CrossRef]
- Peter, S.; Tim, N.; Jeremy, K. Climate change contributes to widespread declines among bumble bees across continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Straub, L.; Villamar-Bouza, L.; Bruckner, S.; Chantawannakul, P.; Gauthier, L.; Khongphinitbunjong, K.; Retschnig, G.; Troxler, A.M.; Vidondo, B.; Neumann, P.; et al. Neonicotinoid insecticides can serve as inadvertent insect contraceptives. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160506. [Google Scholar] [CrossRef] [Green Version]
- Inouri-Iskounen, A.; Sadeddine-Zennouche, O.; Nait Mouloud, M.; Kebieche, M.; Iguer-Ouada, M. In vitro effects of imidacloprid on honey bee sperm: Evaluation using computer-aided sperm analysis (CASA). J. Apic. Res. 2020, 59, 343–349. [Google Scholar] [CrossRef]
- Chaimanee, V.; Evans, J.D.; Chen, Y.; Jackson, C.; Pettis, J.S. Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 2016, 89, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chaimanee, V.; Pettis, J.S. Gene expression, sperm viability, and queen (Apis mellifera) loss following pesticide exposure under laboratory and field conditions. Apidologie 2019, 50, 304–316. [Google Scholar] [CrossRef]
- Kairo, G.; Provost, B.; Tchamitchian, S.; Ben Abdelkader, F.; Bonnet, M.; Cousin, M.; Sénéchal, J.; Benet, P.; Kretzschmar, A.; Belzunces, L.P.; et al. Drone exposure to the systemic insecticide Fipronil indirectly impairs queen reproductive potential. Sci. Rep. 2016, 6, 31904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundin, O.; Rundlöf, M.; Smith, H.G.; Fries, I.; Bommarco, R. Neonicotinoid insecticides and their impacts on bees: A systematic review of research approaches and identification of knowledge gaps. PLoS ONE 2015, 10, e0136928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölldobler, B.A.B. Sociobiology of reproduction in ants. In Experimental Behavioral Ecology and Sociobiology; Hölldobler, B., Lindauer, M., Eds.; Gustav Fisher: Stuttgart, Germany, 1985; Volume 31, pp. 237–257. [Google Scholar]
- Baer, B. Sexual selection in Apis bees. Apidologie 2005, 36, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Satoh, T. Sperm maturation process occurs in the seminal vesicle following sperm transition from testis in honey bee males. Apidologie 2019, 50, 369–378. [Google Scholar] [CrossRef]
- Metz, B.N.; Tarpy, D. Reproductive senescence in drones of the honey bee (Apis mellifera). Insects 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Locke, S.J.; Peng, Y.-S. The effects of drone age, semen storage and contamination on semen quality in the honey bee (Apis mellifera). Physiol. Èntomol. 1993, 18, 144–148. [Google Scholar] [CrossRef]
- Rousseau, A.; Fournier, V.; Giovenazzo, P. Apis mellifera (Hymenoptera: Apidae) drone sperm quality in relation to age, genetic line, and time of breeding. Can. Èntomol. 2015, 147, 702–711. [Google Scholar] [CrossRef]
- Borsuk, G.; Olszewski, K.; Strachecka, A.; Paleolog, J.; Gagos, M. Microscopic image of honeybee drone spermatozoa in three diluents. J. Apic. Sci. 2011, 55, 5–13. [Google Scholar]
- Borsuk, G.; Strachecka, A.; Olszewski, K.; Paleolog, J.; Chobotow, J.; Anusiewicz, M. Proteolytic system of the sperm of Apis mellifera drones. Biologia 2013, 68, 533–538. [Google Scholar] [CrossRef]
- Kumaresan, A.; Das Gupta, M.; Datta, T.K.; Morrell, J.M. Sperm DNA integrity and male fertility in farm animals: A review. Veter- Sci. 2020, 7, 321. [Google Scholar] [CrossRef] [PubMed]
- Borsuk, G.; Kozłowska, M.; Anusiewicz, M.; Olszewski, K. A scientific note on DNA fragmentation rates in sperm collected from drones and spermathecae of queens of different age, with possible implications on the scattered brood phenomenon. Apidologie 2018, 49, 803–806. [Google Scholar] [CrossRef] [Green Version]
- Czekońska, K.; Szentgyörgyi, H.; Tofilski, A. Body mass but not wing size or symmetry correlates with life span of honey bee drones. Bull. Èntomol. Res. 2018, 109, 383–389. [Google Scholar] [CrossRef]
- Page, R.E.; Erickson, E.H. Reproduction by worker honey bees (Apis mellifera L.). Behav. Ecol. Sociobiol. 1988, 23, 117–126. [Google Scholar] [CrossRef]
- Peso, M.; Even, N.; Søvik, E.; Naeger, N.L.; Robinson, G.E.; Barron, A.B. Physiology of reproductive worker honey bees (Apis mellifera): Insights for the development of the worker caste. J. Comp. Physiol. A 2015, 202, 147–158. [Google Scholar] [CrossRef]
- Gençer, H.V.; Firatli, Ç. Reproductive and morphological comparisons of drones reared in queenright and laying worker colonies. J. Apic. Res. 2005, 44, 163–167. [Google Scholar] [CrossRef]
- Utaipanon, P.; Holmes, M.J.; Oldroyd, B.P. Queenless colonies contribute to the male breeding population at honey bee drone congregation areas. Insect Soc. 2019, 66, 593–599. [Google Scholar] [CrossRef]
- Gençer, H.V.; Kahya, Y. Are sperm traits of drones (Apis mellifera L.) from laying worker colonies noteworthy? J. Apic. Res. 2011, 50, 130–137. [Google Scholar] [CrossRef]
- Schlüns, H.; Schlüns, E.A.; Van Praagh, J.; Moritz, R.F. Sperm numbers in drone honeybees (Apis mellifera) depend on body size. Apidologie 2003, 34, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Couvillon, M.J.; Hughes, W.O.; Pérez-Sato, J.A.; Martin, S.J.; Roy, G.G.; Ratnieks, F.L. Sexual selection in honey bees: Colony variation and the importance of size in male mating success. Behav. Ecol. 2010, 21, 520–525. [Google Scholar] [CrossRef] [Green Version]
- Gençer, H.V.; Kahya, Y. Sperm competition in honey bees (Apis mellifera L.): The role of body size dimorphism in drones. Apidologie 2019, 51, 1–17. [Google Scholar] [CrossRef]
- Lopez-Vaamonde, C.; Koning, J.W.; Brown, R.M.; Jordan, W.C.; Bourke, A.F.G. Social parasitism by male-producing reproductive workers in a eusocial insect. Nature 2004, 430, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Savarit, F.; Jaisson, P.; Hefetz, A. Does the queen win it all? Queen-worker conflict over male production in the bumblebee, Bombus terrestris. Naturwissenschaften 2004, 91, 400–403. [Google Scholar] [CrossRef]
- Zanette, L.R.S.; Miller, S.D.L.; Faria, C.M.A.; Almond, E.; Huggins, T.J.; Jordan, W.C.; Bourke, A.F.G. Reproductive conflict in bumblebees and the evolution of worker policing. Evolution 2012, 66, 3765–3777. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.K. Reproductive conflict in honey bees: A stalemate of worker egg-laying and policing. Behav. Ecol. Sociobiol. 1996, 39, 237–244. [Google Scholar] [CrossRef]
- Duvoisin, N.; Baer, B.; Schmid-Hempel, P. Sperm transfer and male competition in a bumblebee. Anim. Behav. 1999, 58, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.H. A preliminary cladistic investigation of relationships among bumblebees (Hymenoptera, Apidae). Syst. Entomol. 1985, 10, 239–255. [Google Scholar] [CrossRef]
- Baer, B.; Heazlewood, J.L.; Taylor, N.L.; Eubel, H.; Millar, A.H. The seminal fluid proteome of the honeybee Apis mellifera. Proteom. 2009, 9, 2085–2097. [Google Scholar] [CrossRef]
- Findlay, G.D.; Yi, X.; MacCoss, M.J.; Swanson, W.J. Proteomics reveals novel Drosophila seminal fluid proteins transferred at mating. PLoS Biol. 2008, 6, e178. [Google Scholar] [CrossRef] [PubMed]
- Pilch, B.; Mann, M. Large-scale and high-confidence proteomic analysis of human seminal plasma. Genome Biol. 2006, 7, R40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.M.; Caperna, T.J.; Williams, V.; Garrett, W.M.; Evans, J.D. Proteomic analyses of male contributions to honey bee sperm storage and mating. Insect Mol. Biol. 2006, 15, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Colonello, N.A.; Hartfelder, K. She’s my girl-male accessory gland products and their function in the reproductive biology of social bees. Apidologie 2005, 36, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Poiani, A. Complexity of seminal fluid: A review. Behav. Ecol. Sociobiol. 2006, 60, 289–310. [Google Scholar] [CrossRef]
- Tram, U.; Wolfner, M.F. Male seminal fluid proteins are essential for sperm storage in Drosophila melanogaster. Genetics 1999, 153, 837–844. [Google Scholar] [CrossRef]
- Wagner, W.E.W., Jr.; Kelley, R.J.; Tucker, K.R.; Harper, C.J. Females receive a life-span benefit from male ejaculates in a field cricket. Evolution 2001, 55, 994. [Google Scholar] [CrossRef]
- Fan, Y.; Rafaeli, A.; Gileadi, C.; Kubli, E.; Applebaum, S.W. Drosophila melanogaster sex peptide stimulates juvenile hormone synthesis and depresses sex pheromone production in Helicoverpa armigera. J. Insect Physiol. 1999, 45, 127–133. [Google Scholar] [CrossRef]
- Goenaga, J.; Yamane, T.; Rönn, J.; Arnqvist, G. Within-species divergence in the seminal fluid proteome and its effect on male and female reproduction in a beetle. BMC Evol. Biol. 2015, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kingan, T.G.; Bodnar, W.M.; Raina, A.K.; Shabanowitz, J.; Hunt, D.F. The loss of female sex pheromone after mating in the corn earworm moth Helicoverpa zea: Identification of a male pheromonostatic peptide. Proc. Natl. Acad. Sci. USA 1995, 92, 5082–5086. [Google Scholar] [CrossRef] [Green Version]
- Rogers, D.W.; Baldini, F.; Battaglia, F.; Panico, M.; Dell, A.; Morris, H.R.; Catteruccia, F. Transglutaminase-mediated semen coagulation controls sperm storage in the Malaria Mosquito. PLoS Biol. 2009, 7, e1000272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wang, Q. Seminal fluid reduces female longevity and stimulates egg production and sperm trigger oviposition in a moth. J. Insect Physiol. 2011, 57, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Goenaga, J.; Rönn, J.L.; Arnqvist, G. Male seminal fluid substances affect sperm competition success and female reproductive behavior in a seed beetle. PLoS ONE 2015, 10, e0123770. [Google Scholar] [CrossRef] [Green Version]
- Yamane, T.; Miyatake, T. Reduced female mating receptivity and activation of oviposition in two Callosobruchus species due to injection of biogenic amines. J. Insect Physiol. 2010, 56, 271–276. [Google Scholar] [CrossRef]
- Boer, S.D.; Baer, B.; Boomsma, J.J. Seminal fluid mediates ejaculate competition in social insects. Science 2010, 327, 1506–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, B. Bumblebees as model organisms to study male sexual selection in social insects. Behav. Ecol. Sociobiol. 2003, 54, 521–533. [Google Scholar] [CrossRef]
- Yamane, T.; Kimura, Y.; Katsuhara, M.; Miyatake, T. Female mating receptivity inhibited by injection of male-derived extracts in Callosobruchus chinensis. J. Insect Physiol. 2008, 54, 501–507. [Google Scholar] [CrossRef]
- Poland, V.; Eubel, H.; King, M.; Solheim, C.; Millar, A.H.; Baer, B. Stored sperm differs from ejaculated sperm by proteome alterations associated with energy metabolism in the honeybee Apis mellifera. Mol. Ecol. 2011, 20, 2643–2654. [Google Scholar] [CrossRef] [Green Version]
- Greeff, M.; Schmid-Hempel, P. Sperm viability in the male accessory testes and female spermathecae of the bumblebee Bombus terrestris (Hymenoptera: Apidae). Eur. J. Èntomol. 2008, 105, 849–854. [Google Scholar] [CrossRef] [Green Version]
- King, M.; Eubel, H.; Millar, A.H.; Baer, B. Proteins within the seminal fluid are crucial to keep sperm viable in the honeybee Apis mellifera. J. Insect Physiol. 2011, 57, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Jasper, W.C.; Brutscher, L.M.; Grozinger, C.M.; Niño, E.L. Injection of seminal fluid into the hemocoel of honey bee queens (Apis mellifera) can stimulate post-mating changes. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Johnson, J.N.; Hardgrave, E.; Gill, C.; Moore, D. Absence of consistent diel rhythmicity in mated honey bee queen behavior. J. Insect Physiol. 2010, 56, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Gioti, A.; Wigby, S.; Wertheim, B.; Schuster, E.; Martinez, P.; Pennington, C.J.; Partridge, L.; Chapman, T. Sex peptide of Drosophila melanogaster males is a global regulator of reproductive processes in females. Proc. R. Soc. B Biol. Sci. 2012, 279, 4423–4432. [Google Scholar] [CrossRef] [Green Version]
- Manfredini, F.; Romero, A.E.; Pedroso, I.; Paccanaro, A.; Sumner, S.; Brown, M.J.F. Neurogenomic signatures of successes and failures in life-history transitions in a key insect pollinator. Genome Biol. Evol. 2017, 9, 3059–3072. [Google Scholar] [CrossRef] [Green Version]
- Grassl, J.; Peng, Y.; Baer-Imhoof, B.; Welch, M.; Millar, A.H.; Baer, B. Infections with the sexually transmitted pathogen Nosema apis trigger an immune response in the seminal fluid of honey bees (Apis mellifera). J. Proteome Res. 2016, 16, 319–334. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Grassl, J.; Millar, A.H.; Baer, B. Seminal fluid of honeybees contains multiple mechanisms to combat infections of the sexually transmitted pathogen Nosema apis. Proc. R. Soc. B Biol. Sci. 2016, 283, 20151785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurze, C.; Dosselli, R.; Grassl, J.; Le Conte, Y.; Kryger, P.; Baer, B.; Moritz, R.F. Differential proteomics reveals novel insights into Nosema–honey bee interactions. Insect Biochem. Mol. Biol. 2016, 79, 42–49. [Google Scholar] [CrossRef]
- Baer, B.; Zareie, R.; Paynter, E.; Poland, V.; Millar, A.H. Seminal fluid proteins differ in abundance between genetic lineages of honeybees. J. Proteom. 2012, 75, 5646–5653. [Google Scholar] [CrossRef] [PubMed]
- Boer, S.P.D.; Boomsma, J.J.; Baer, B. Honey bee males and queens use glandular secretions to enhance sperm viability before and after storage. J. Insect Physiol. 2009, 55, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Misra, S.; Wolfner, M.F. Drosophila seminal sex peptide associates with rival as well as own sperm, providing SP function in polyandrous females. eLife 2020, 9, 58322. [Google Scholar] [CrossRef]
- Heifetz, Y.; Lung, O.; Frongillo, E.A.; Wolfner, M.F. The Drosophila seminal fluid protein Acp26Aa stimulates release of oocytes by the ovary. Curr. Biol. 2000, 10, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Pascini, T.V.; Ramalho-Ortigão, M.; Ribeiro, J.M.; Jacobs-Lorena, M.; Martins, G.F. Transcriptional profiling and physiological roles of Aedes aegypti spermathecal-related genes. BMC Genom. 2020, 21, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Pascini, T.V.; Martins, G.F. The insect spermatheca: An overview. Zoology 2017, 121, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Wolfner, M.F. Precious essences: Female secretions promote sperm storage in Drosophila. PLoS Biol. 2011, 9, e1001191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparini, C.; Pilastro, A.; Evans, J.P. The role of female reproductive fluid in sperm competition. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20200077. [Google Scholar] [CrossRef]
- Prokupek, A.M.; Kachman, S.D.; Ladunga, I.; Harshman, L.G. Transcriptional profiling of the sperm storage organs of Drosophila melanogaster. Insect Mol. Biol. 2009, 18, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.; Eubel, H.; Taylor, N.L.; O’Toole, N.; Millar, A.H. Insights into female sperm storage from the spermathecal fluid proteome of the honeybee Apis mellifera. Genome Biol. 2009, 10, R67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, A.; Shigenobu, S.; Yamaguchi, K.; Kobayashi, S.; Ito, F.; Tsuji, K. Transcriptome profiling of the spermatheca identifies genes potentially involved in the long-term sperm storage of ant queens. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, W.R.; Teodori, E.; Mitchell, S.N.; Baldini, F.; Gabrieli, P.; Rogers, D.W.; Catteruccia, F. Mating activates the heme peroxidase HPX15 in the sperm storage organ to ensure fertility in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2014, 111, 5854–5859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.N.; Ing, N.; Rangel, J. Upregulation of antioxidant genes in the spermathecae of honey bee (Apis mellifera) queens after mating. Apidologie 2018, 49, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.M.; Williams, V.; Evans, J.D. Sperm storage and antioxidative enzyme expression in the honey bee, Apis mellifera. Insect Mol. Biol. 2004, 13, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, F.; Li, G.; Chi, X.; Wang, Y.; Wang, H.; Ma, L.; Han, K.; Zhao, G.; Guo, X. Metabolite support of long-term storage of sperm in the spermatheca of honeybee (Apis mellifera) queens. Front. Physiol. 2020, 11, 1303. [Google Scholar] [CrossRef] [PubMed]
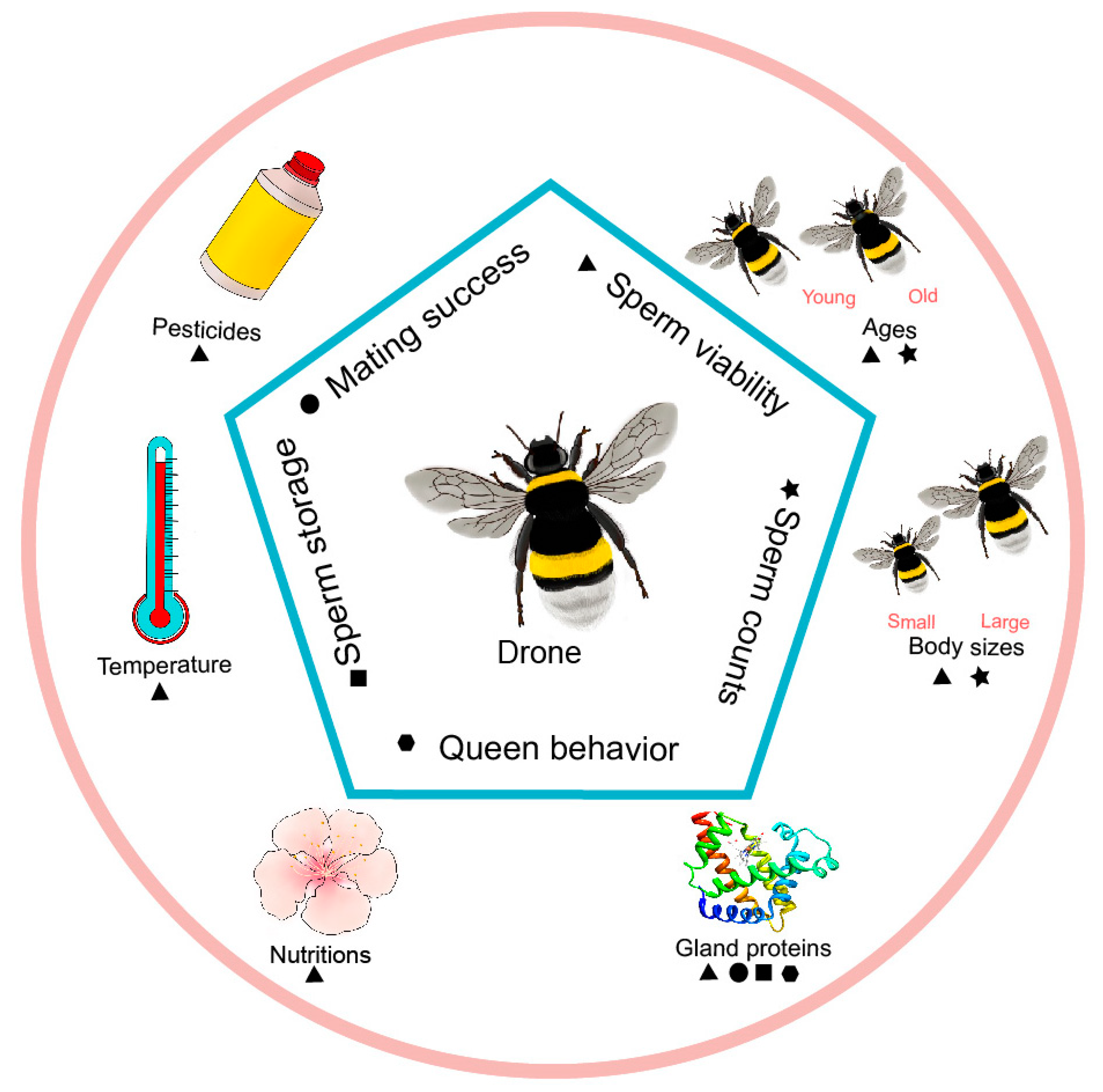
Figure 1.
The main aspects of the reproductive ability of male bees are affected by various environmental factors and inherent characteristics, including ▲ sperm viability, ★ sperm counts, ⬣ queen behavior, ■ sperm storage, ● mating success.
Figure 1.
The main aspects of the reproductive ability of male bees are affected by various environmental factors and inherent characteristics, including ▲ sperm viability, ★ sperm counts, ⬣ queen behavior, ■ sperm storage, ● mating success.


{kind=link}
Table 1.
Functions of proteins in accessory gland of male insects and in spermathecae of queen insects presented throughout the review.
Table 1.
Functions of proteins in accessory gland of male insects and in spermathecae of queen insects presented throughout the review.
| Organ | Key Points | Descriptions | Species | References |
|---|---|---|---|---|
| Accessory gland in male | Maintenance of sperm viability | The semen is so effective at keeping sperm alive that the positive effects can last up to 24 h cannot be replicated by ordinary protein substitutes | Apis mellifera Bombus terrestris | [115,116,125] |
| Contribution to sperm transfer and storage | Alter expression of several genes and change morphology of queen’s reproductive tract. | Drosophila melanogaster | [102] | |
| Sperm competition | Change the expression of genes in the queens’ brains associated with vision to restrain queens to mate again, reduce sexual receptivity. Directly benefit the previous sperm stored in females. | Apis mellifera Drosophila | [18,117,126] | |
| Stimulate changes of post-mating queens | Minimize the risk to sexually transmit the parasite to the queen and colony. Stimulate the release of oocytes by the ovary. | Apis mellifera Drosophila melanogaster | [121,127] | |
| Spermathecae in queens | Maintenance of sperm viability | Higher expression of glutathione-stransferase, catalase, thioredoxin 2, and thioredoxin reductase 1 may assist in maintaining sperm viability inside the spermathecae. | Apis mellifera | [136] |
| Facilitate long-term storage of sperm | Proteins in spermathecae fall into a range of different functional groups, most notably enzymes of energy metabolism and antioxidant defense. Seminal receptacle was enriched for genes involved in localization, signaling and ion transport. GPL metabolism, biosynthesis of amino acids, and the mTOR signaling pathway were also enriched. Mating-induced heme peroxidase (HPX) 15 as an important factor in long-term fertility. | Apis mellifera Drosophila melanogaster Crematogaster osakensis Anopheles gambiae | [132,133,134,135,137,138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, H.; Mashilingi, S.K.; Liu, Y.; An, J. Factors Influencing the Reproductive Ability of Male Bees: Current Knowledge and Further Directions. Insects 2021, 12, 529. https://doi.org/10.3390/insects12060529
AMA Style
Zhao H, Mashilingi SK, Liu Y, An J. Factors Influencing the Reproductive Ability of Male Bees: Current Knowledge and Further Directions. Insects. 2021; 12(6):529. https://doi.org/10.3390/insects12060529
Chicago/Turabian StyleZhao, Huiyue, Shibonage K. Mashilingi, Yanjie Liu, and Jiandong An. 2021. "Factors Influencing the Reproductive Ability of Male Bees: Current Knowledge and Further Directions" Insects 12, no. 6: 529. https://doi.org/10.3390/insects12060529
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.