1. Introduction
Despite their limited neuronal capacity [
1], herbivorous insects must face a continuous and challenging process of searching for, evaluating and selecting a host that may present mechanical and/or chemical defenses [
2]. These defenses can be drastically affected by a domestication process that makes them generally more vulnerable than their wild relative [
3,
4,
5]. Such is the case with maize, which is the result of a long process of domestication of teocintle (
Zea mays ssp. parviglumis) begun approximately 10,000 years ago [
6]. This process reduces their defenses [
7], making them more susceptible to herbivores [
8].
Native to the Americas and established in Europe [
9], Africa [
10,
11] and Asia [
12],
Spodoptera frugiperda (Smith, J.E. 1797) (Lepidoptera: Noctuidae) is one of the generalist insects to have taken advantage of the low defenses of many domesticated crops (mainly grasses [
13,
14]) and become their key pest. Chemical control is the main way to combat these pests. However, the strict regulations imposed on chemical insecticides has increased the interest in environmentally friendly pest management strategies, among which are behavioral manipulation methods [
15]. The latter consists of using chemical compounds that stimulate or inhibit the insect-pest’s behavior and, consequently, its expression [
16]. Of these techniques, one of the few implemented that is effective is the push–pull strategy [
17], which consists of manipulating the insect-pest’s behavior by using a repellent stimulus to expel it from the main crop, and an attractive stimulus to attract it to an alternative source, where it can be eliminated or controlled [
18]. The successful design of an effective push–pull strategy requires knowledge of the insect-pest’s biology and its interaction with the candidate species to be integrated into the system. This knowledge should go beyond an understanding of repellent or attractive properties [
19].
Plant colonization differs between specialist and generalist insects [
20]. While specialists use more specific stimuli from a selected number of hosts, and are more tolerant of their host’s defenses [
3,
21,
22], generalists such as
S. frugiperda inhabit a wide range of hosts and are more sensitive to their defenses. Unlike specialists, generalists are less efficient in host selection and more vulnerable to their natural enemies [
1]. Therefore, although generalists have advantages, such as greater availability of resources and a better nutritional balance [
23], there is a general tendency in herbivorous insects to specialize [
24]. Consequently, generalists are considered more vulnerable, more manipulable, and relatively easier to repel than specialists [
3]. Fortunately, although most insects are specialists [
1], most of those that become important agricultural pests are generalists. These present a host acceptability hierarchy, which is exploited in pest management strategy design, and mainly in trap plant selection [
25].
In the establishment of push–pull systems, unlike the selection of repellent plants that is based mainly on antixenosis studies, attractive plant (trap) selection is based on a greater number of criteria (antixenosis, antibiosis and tolerance), generated by greater plant–insect interaction. Because of this larger number of criteria, trap plant selection can be highly subjective. Therefore, multivariate data reduction techniques can be used to construct selection indexes (SI) that simultaneously express these multiple dimensions in a simple way for interpretation [
26]. Principal Component Analysis (PCA) is the best index construction method [
27]. Based on their SI values, plants can be ordered according to characteristics that indicate their degrees of antixenosis (preference or non-preference), antibiosis and tolerance, in relation to a certain insect-pest.
The broad knowledge generated in recent decades regarding push–pull [
28,
29,
30,
31,
32,
33] and its successful use for the management of
S. frugiperda in Africa [
17,
32,
34], in addition to the empirical knowledge of Mesoamerican milpas (traditional local systems of maize polyculture), constitutes a foundation for the proposal of an efficient push–pull system for
S. frugiperda management in Mexico. Therefore, this study selected attractive and repellent plants for the design of push–pull strategies for
S. frugiperda management in maize crops (
Z. mays), in the municipality of Yautepec, Morelos, México.
4. Discussion
Plant selection for the design of push–pull strategies was carried out by studying the relationship of
S. frugiperda with potential attractive or repellent plants.
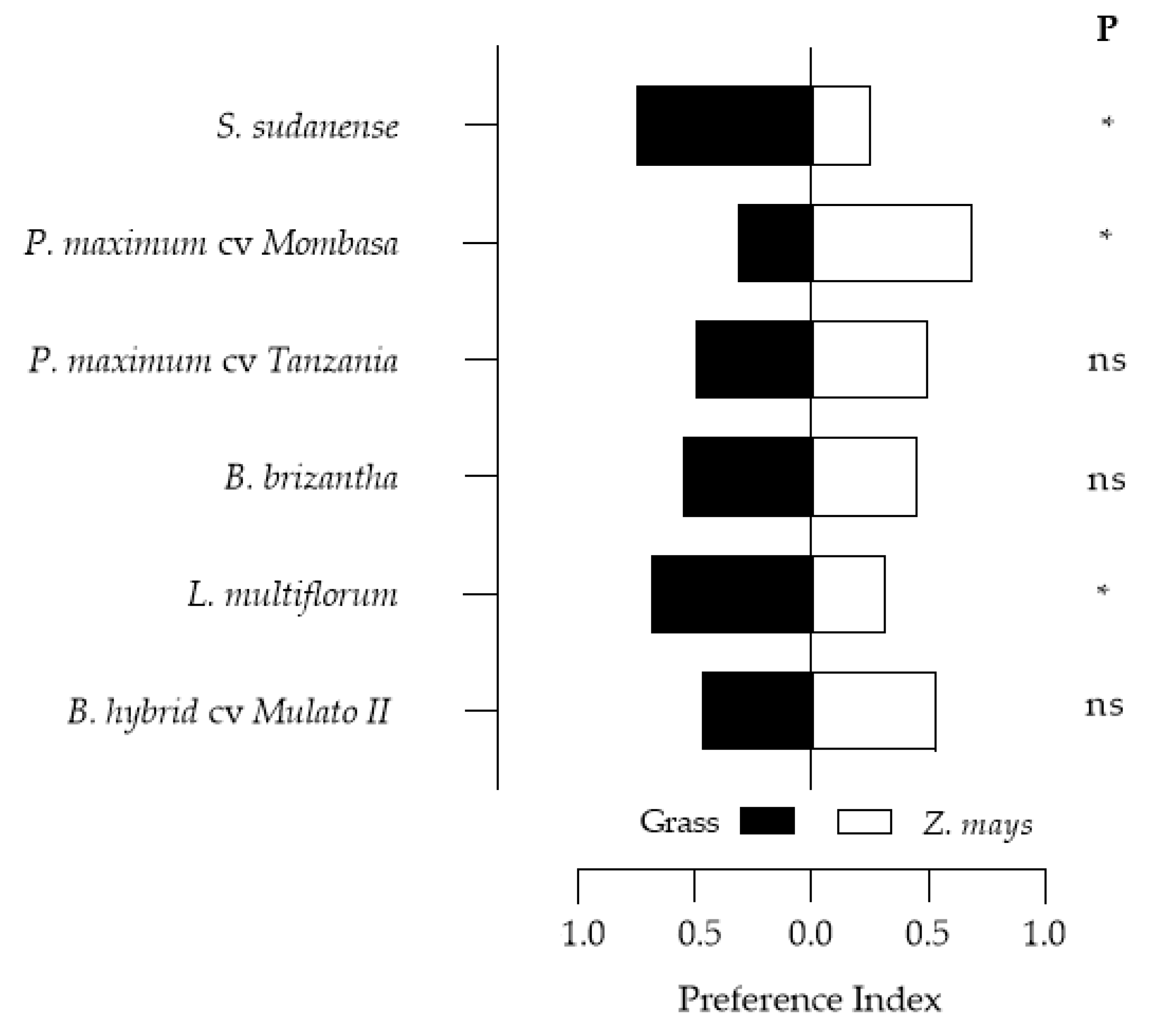
B. brizantha was the only grass with an oviposition deterrent effect (indicated by negative OPI index), the other grasses stimulated oviposition (
Figure 3 and
Figure 4). The oviposition preference for
P. maximum cv. Mombasa,
L. multiflorum and
S. sudanense agrees with Pitre et al. [
58], who report
L. multiflorum among the grasses preferred by
S. frugiperda for oviposition. This species preferred to oviposit on the leaves’ abaxial surfaces, as reported by other authors (Pitre et al. [
58], Ali et al. [
59] and Beserra et al. [
60]).
Trichome density is one of the main causes of antixenosis in generalist lepidoptera oviposition. In this study, the highest densities were observed in
B. hybrid cv. Mulato II and
B. brizantha (
Table S2). The values found for these species are lower than those reported by Cheruiyot et al. [
61]. Except for Mombasa, the trichome density decreased from the leaf base to the leaf apex, as reported by Rendón-Carmona et al. [
38].
The
B. brizantha–Z. mays assay revealed a strong negative correlation between trichome density and oviposition (
Table 2). Preference for the species with the lowest density of trichomes may indicate that these structures inhibit oviposition, as reported by Kumar [
62]. However, this result differs from those of Pitre et al. [
58], who associate
S. frugiperda oviposition preference with leaf color. Negative correlations were also observed in the
P. maximum cv. Mombasa and
S. sudanense assays. On the other hand, a significant positive correlation was observed between oviposition and trichomes density in the
L. multiflorum–Z. mays test, which could indicate that females preferred to oviposit in plants with higher trichomes density, or that trichomes were not determinants for oviposition, as the densities observed in both species were low. It was also perceived that the high density of trichomes of
B. hybrid cv. Mulato II could not inhibit oviposition, which was like the density observed in maize. Volatile compounds attractive to
S. frugiperda and released by Mulato II may have counteracted the supposed trichome inhibitory effect. This leads us to infer that oviposition site selection may depend on the balance of physical or tactile stimuli, chemical stimuli (volatile compounds) and visual stimuli (color and shape) [
20]. Kumar [
62] asserts that tactile stimuli are more decisive. For
S. frugiperda, Rojas et al.’s [
63] results confirm the greater importance of these tactile stimuli (rough surfaces) over chemicals (leaf volatiles). These authors report that
S. frugiperda prefers to oviposit on rough surfaces, not to be confused with trichomes, which are plants’ defense structures. From the inconclusive results of the assays with Mulato II and
L. multiflorum, it follows that, although tactile stimuli (in this case the trichomes) are decisive, they can be weighted by the females in the presence of chemical attractants. Among the grasses evaluated,
S. frugiperda preferred
P. maximum cv. Mombasa,
L. multiflorum and
S. sudanense over maize. Although the amounts of oviposited eggs in Tanzania and Mulato II were not significantly higher than those recorded in maize, they were higher, and by this criterion (oviposition), it is considered that they would be optimal
S. frugiperda trap plants, as well as
P. maximum cv. Mombasa,
L. multiflorum and
S. sudanense.
B. brizantha would be inadequate as a trap plant in the push–pull design for the pathosystem
Z. mays–S. frugiperda.
The oviposition site chosen by
S. frugiperda may be inappropriate for larval development [
64], as the neonates can leave that oviposition site by walking or ballooning [
65]. Neonates, despite their limited olfactory system, use chemical signals that are crucial for host selection, mainly in their early stages [
66]. In the olfactometry studies carried out in this study, the attraction exerted by Mombasa and Tanzania on
S. frugiperda larvae (
Figure 5) is probably due to green leaf volatiles (GLVs) that are generally the mediators of generalist Lepidoptera attraction to Poaceae [
67]. Among these GLVs stand out α-pinene and linalool, which have proven to be generators of significant antennal responses in
S. frugiperda [
68]. A similar trend was observed with
S. sudanense and Mulato II, although the attractions of these were not significantly superior to those of maize.
The repellent activity of
T. erecta (
Figure 5) agrees with Díaz and Serrato [
69], who state that species of the genus
Tagetes contain, in their aerial parts, secondary metabolites that can be repellent and/or toxic for numerous insect-pests. Several studies report repellent activities of
T. erecta against many lepidopteran pests. Among them are those of Calumpang and Ohsawa [
70], who found that this species is repellent to
Leucinodes orbonalis, and attributed that repellency to citral and 1-dodecene, two of the seven volatile organic compounds found in the GC–MS (Gas Chromatography–Mass Spectrometry) analysis of that species. In addition to the repellent activity found here,
T. erecta also has an antialimentary activity that can cause up to 72% mortality of
S. frugiperda larvae [
71]. This capacity, added to its great utility, makes this species a crop with great potential for pest management through behavioral manipulation strategies such as push–pull. The repellency of
C. juncea to
S. frugiperda can be attributed to the presence, in its leaves, of triterpenes, alkaloids, flavonoids and phenolic compounds, as reported by Al-Snafi [
72]. The repellent activity of
C. ambrosioides is probably due to the volatile terpenoid compounds reported by Sagrero-Nieves and Bartley [
73], which are generally repellents to herbivorous insect. For a better understanding of the relationship of
S. frugiperda with the plants analyzed here, more detailed studies (GC–MS analysis and electrophysiological bioassays) will be necessary. In the larval dispersion bioassay, the uneven distribution of neonates observed (
Figure 6) may be indicative of the presence of toxins in the grasses [
65].
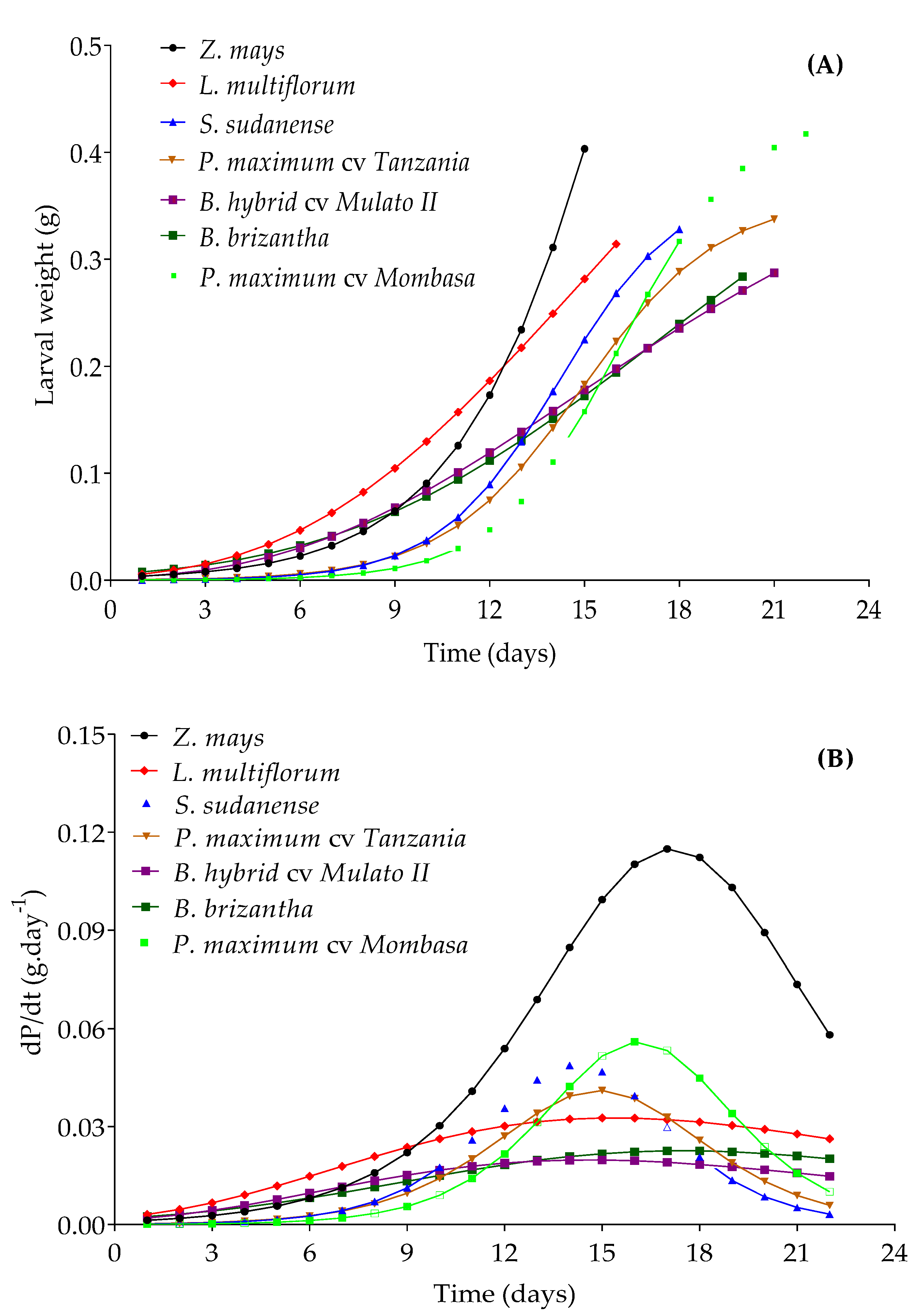
The lower growth rate in larval weight in the other grasses compared to maize resulted in an extension of the duration of larval stages (
Figure 8;
Table 3 and
Table 4). This can be attributed to the low nutritional quality of the plant tissues of these species and/or a higher density of trichomes, mainly in Mulato II and
B. brizantha, which may have inhibited the feeding of
S. frugiperda larvae [
74]. Secondary metabolites can also influence larval performance by being stimulants or deterrents of their feeding. This is probably the case of Mombasa, the feeding preference of which was lower than that of maize (
Figure 7) despite its low trichomes density. Low host quality is compensated by the lengthening of larval stage duration [
75,
76], as occurred with Mombasa, Tanzania, Mulato II and
B. brizantha. The increase in the length of the life cycle (
Table 4) would reduce the population growth of the insect-pest and increase its exposure to natural enemies [
77]. On the other hand, the larval stage of
S. frugiperda was shorter in
Z. mays,
L. multiflorum and
S. sudanense, due to a higher rate of larval development in these species. This latter will produce a greater number of generations per year, which would favor the population growth of the pest.
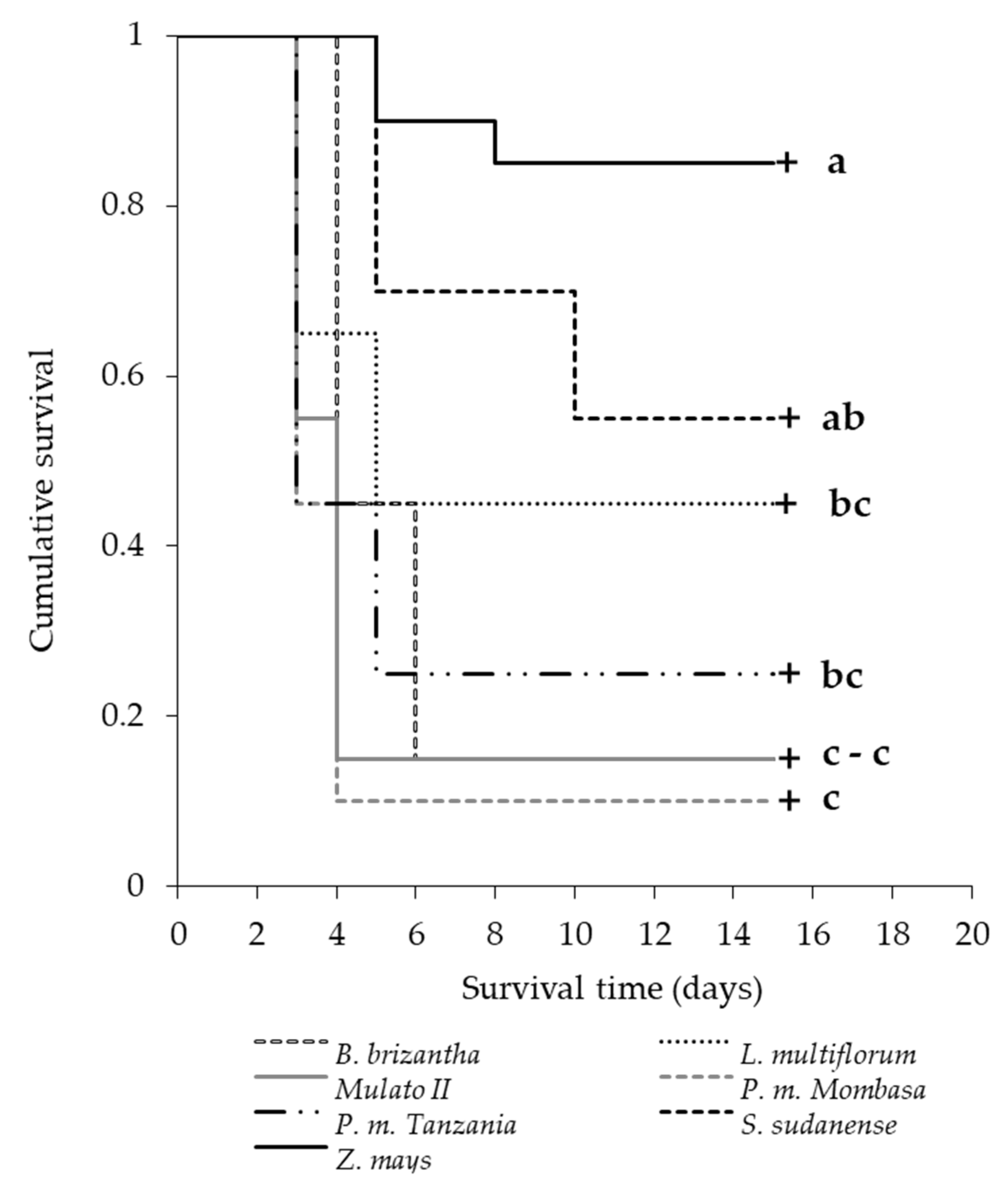
According to Slansky and Feeny [
78], the low nutritional quality mentioned above could lead to higher larval mortality, as in our study. On the fifth day of the survival assessment, the species Mombasa, Tanzania, Mulato II and
B. brizantha produced larvae mortality rates of 90%, 75%, 85% and 55%, respectively (
Figure 9). This high mortality of neonates and larvae of the first two instars is probably due to the antialimentary, insecticidal, antimicrobial and allelopathic [
79,
80,
81] effect of benzoxazinoids [
82,
83].
S. frugiperda larvae ingesting these phytochemicals (DIMBOA and MBOA) detoxify them via N-glycosylation [
84], preventing them from having negative effects on their growth [
85]. However, in some grasses, the combined effect of chemical (including benzoxazinoids at different concentrations) and physical (trichome) defenses can affect neonates’ growth, and even become lethal. Numerous factors can affect the concentrations of DIMBOA in grasses, age being one of them. Generally, higher concentrations of these compounds are found in younger plants [
82,
86], information that must be considered for the timing of the establishment of grasses as trap plants in push–pull systems in the field. The high larval mortality before reaching the most voracious larval stages (third and fourth instars for
S. frugiperda) (
Figure 9), and the lengthening of the larval period (greater exposure to natural enemies) of those who survived (
Table 4), could inhibit pest development in the grasses [
87].
L. multiflorum,
B. brizantha and
S. sudanense fulfilled the hypothesis of “the mother knows best”, according to which the females oviposit in plants suitable for the development of their offspring [
88,
89]. However, this hypothesis, also known as Jaenike’s preference–performance hypothesis, is not always met, and the larvae of some species can make their own choices if those of their parents are not suitable for their development [
66,
90]. Females can prioritize their needs over those of their offspring in oviposition site selection, and this is known as the “optimal bad motherhood” principle [
91,
92,
93]. It was observed that Mombasa, Tanzania and Mulato II were preferred by the females, but these grasses provided poor survival, and poor and slow development, to their offspring. In generalist insects such as
S. frugiperda, this can occur due to its wide range of potential hosts [
94], oviposition pressure [
95], the mobility of the larvae from one plant to another to correct the poor choice of their parent [
96], Hopkins’ host-selection principle [
97], and the enemy-free space hypothesis [
98], which leads some species to oviposit outside their host plant, possibly as a strategy to evade their natural enemies. It may be due to the latter hypothesis that
S. frugiperda tends to oviposit outside its host plants, both in experimental conditions (observations in this study and results of Rojas et al. [
63]) and in the field.
The nine push–pull strategies (
Figure 10) can be used for
S. frugiperda management in maize crops in Morelos, and probably in other regions where the plants that make up these systems develop properly. However, its field effectiveness needs to be studied in future studies.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}