

Ecological Responses of Meiofauna to a Saltier World—A Case Study in the Van Uc River Continuum (Vietnam) in the Dry Season

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
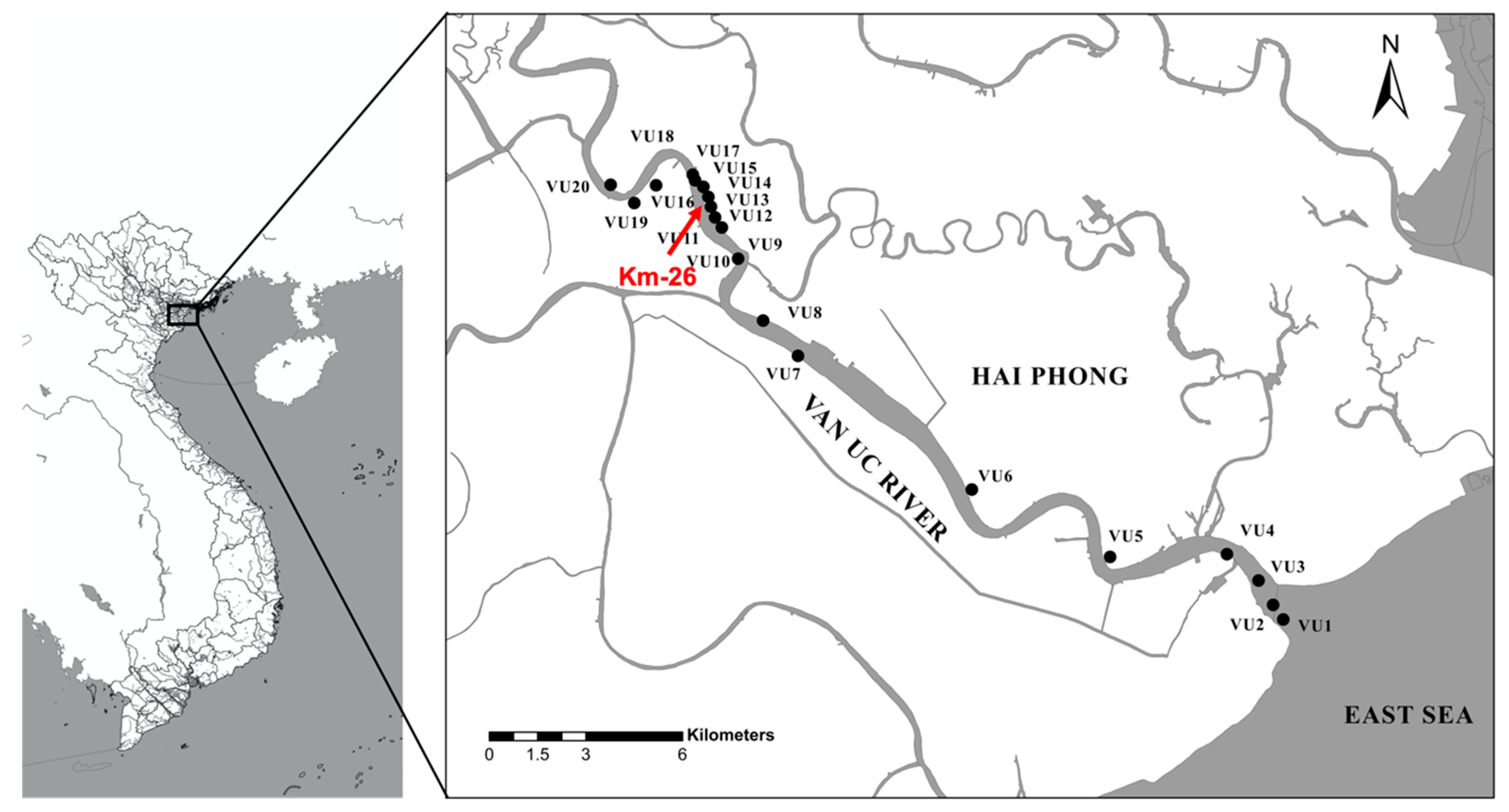
2.1. Study Area
2.2. Sampling and Sample Processing
2.3. Data Analysis
3. Results
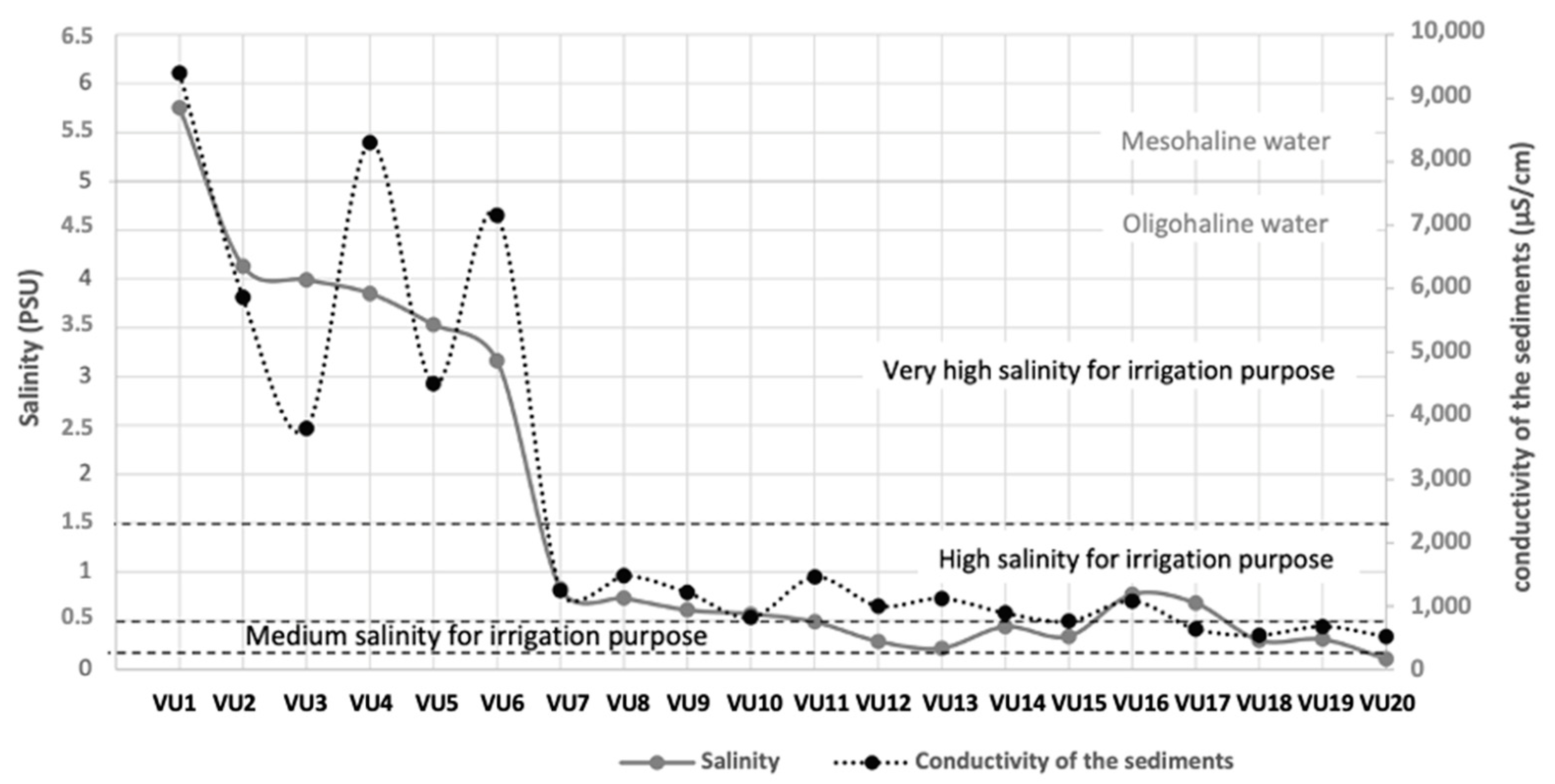
3.1. Environmental Variables
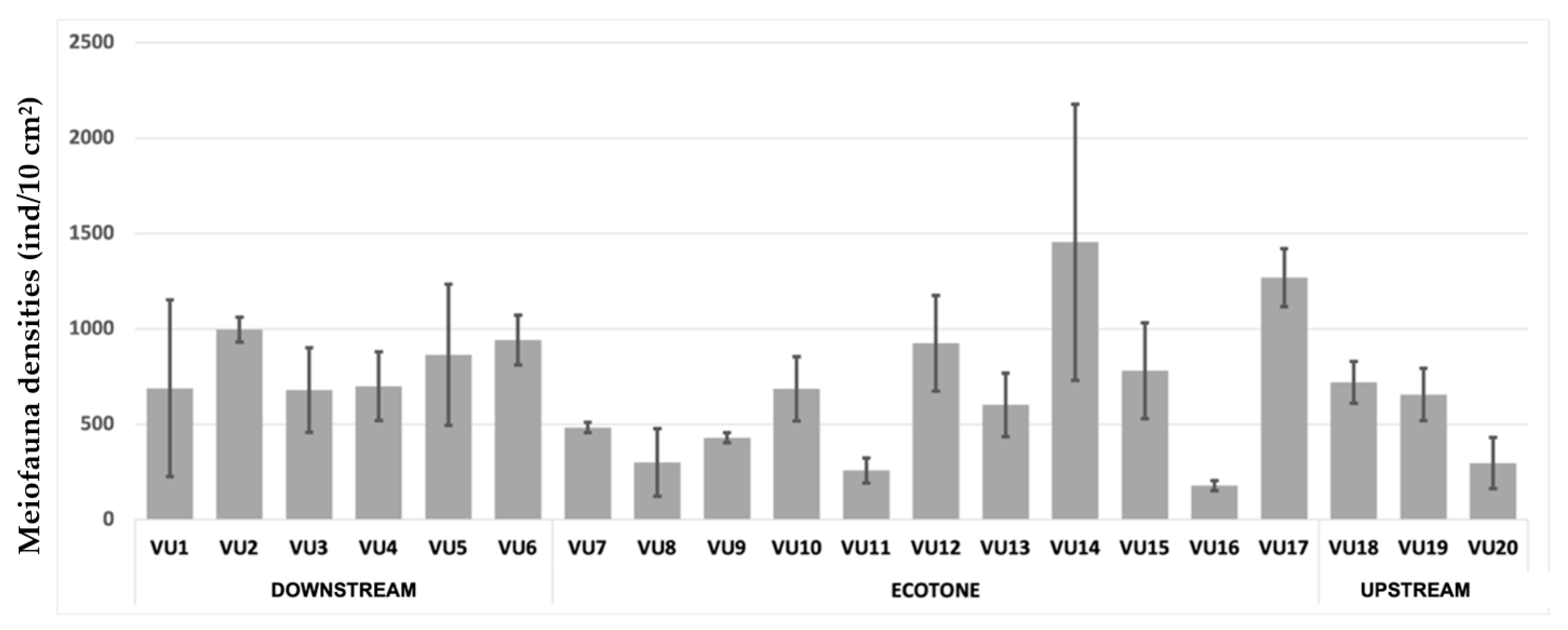
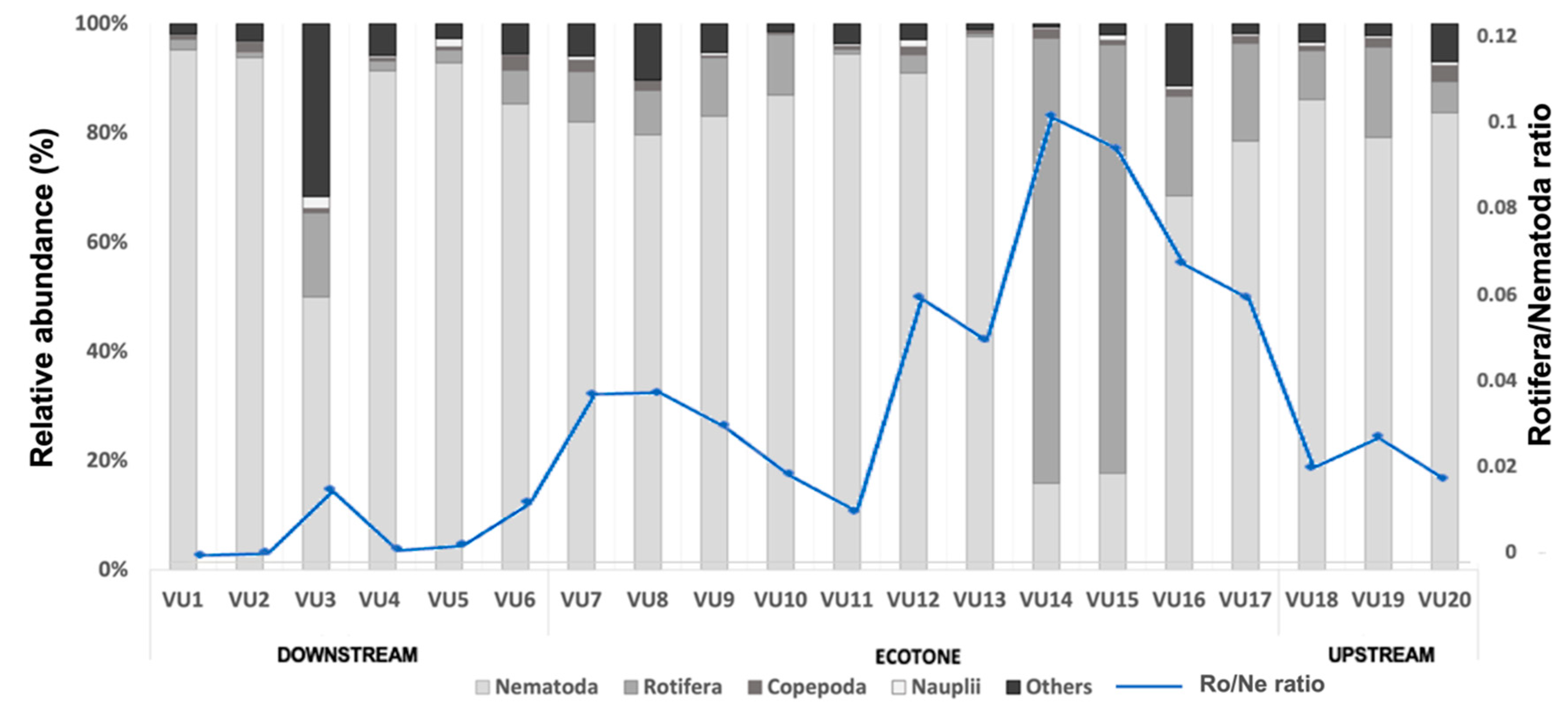
3.2. Meiofauna Assemblage along the Van Uc River
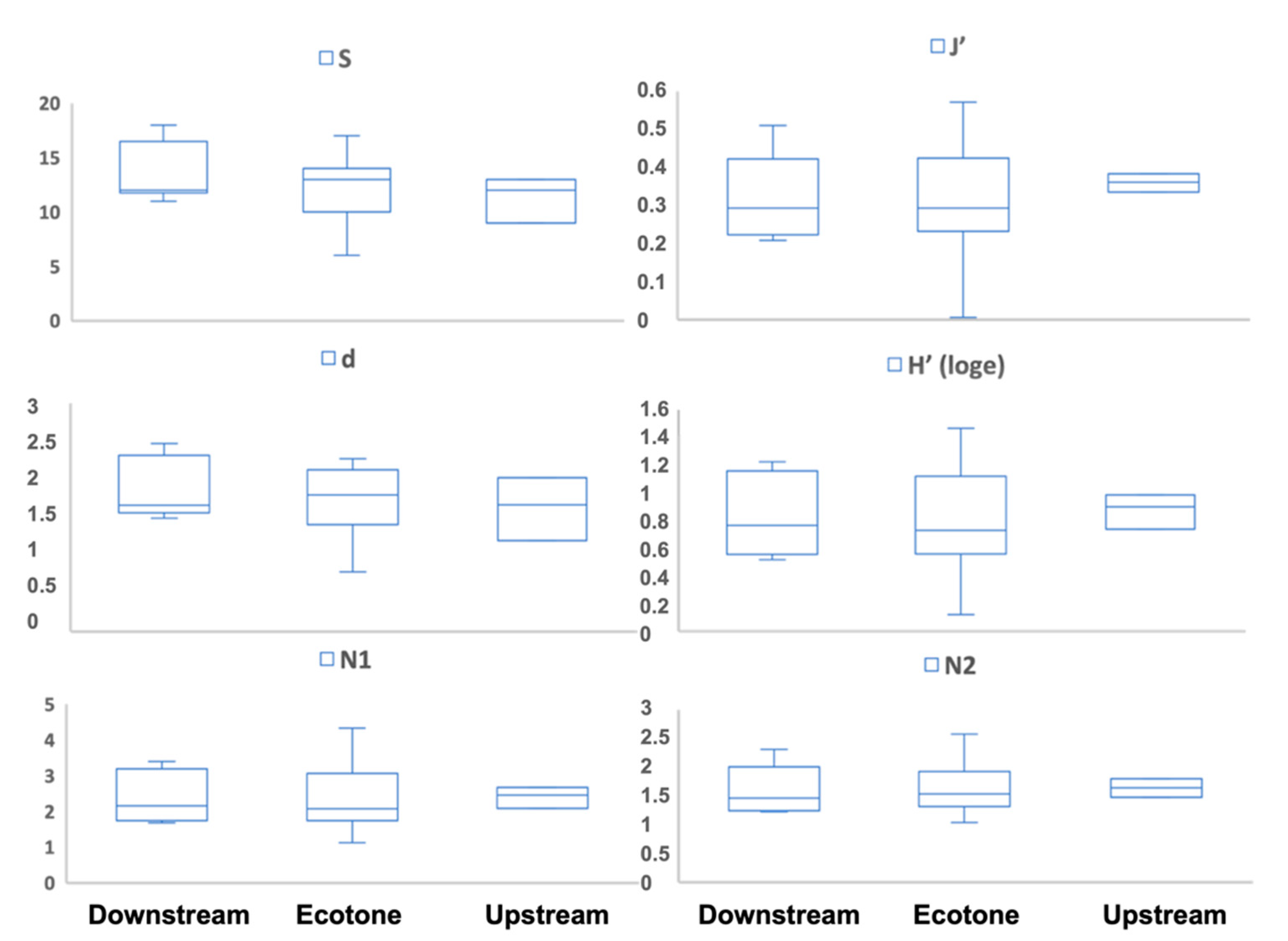
3.3. Meiofaunal Ecological Indices
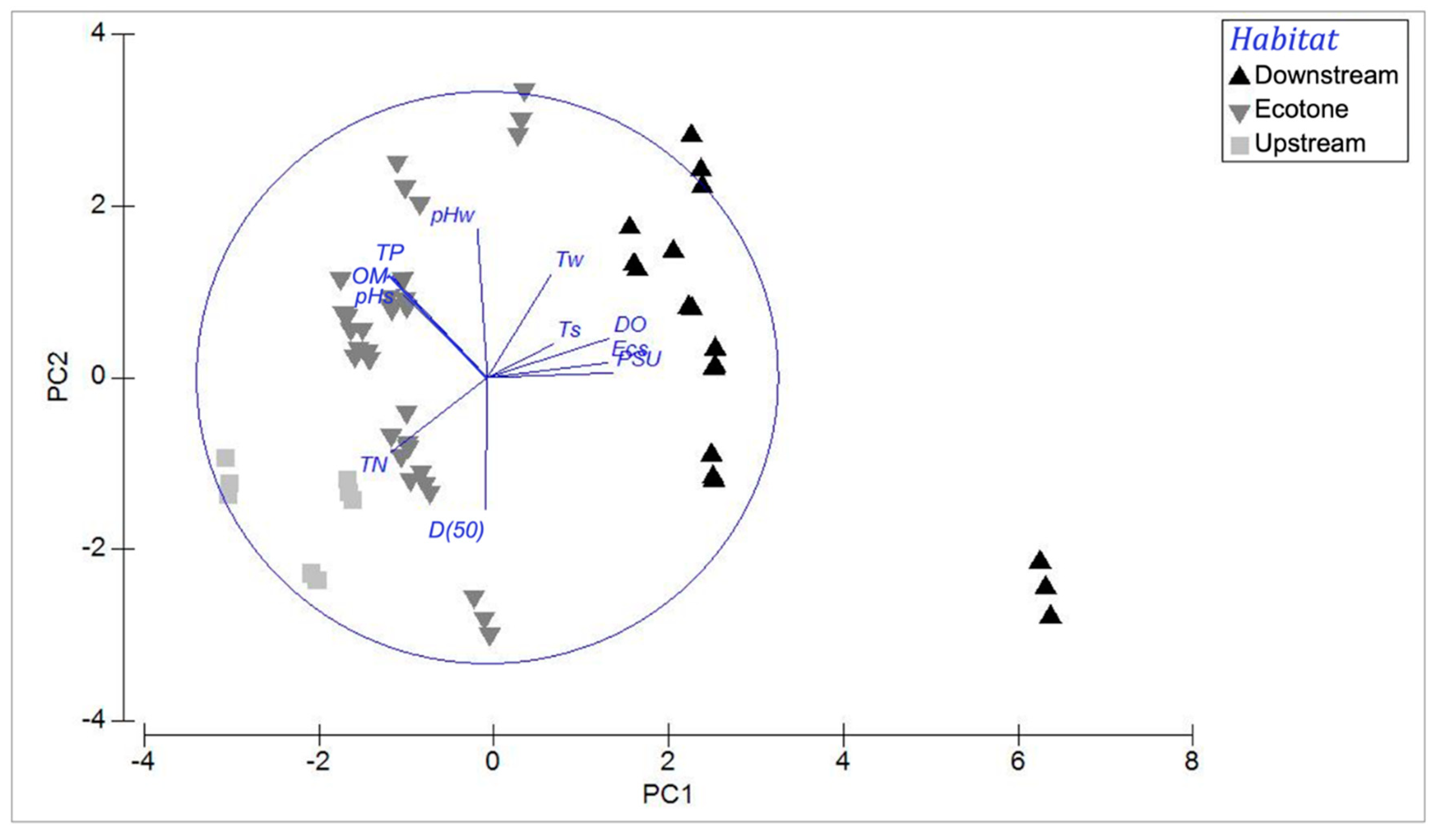
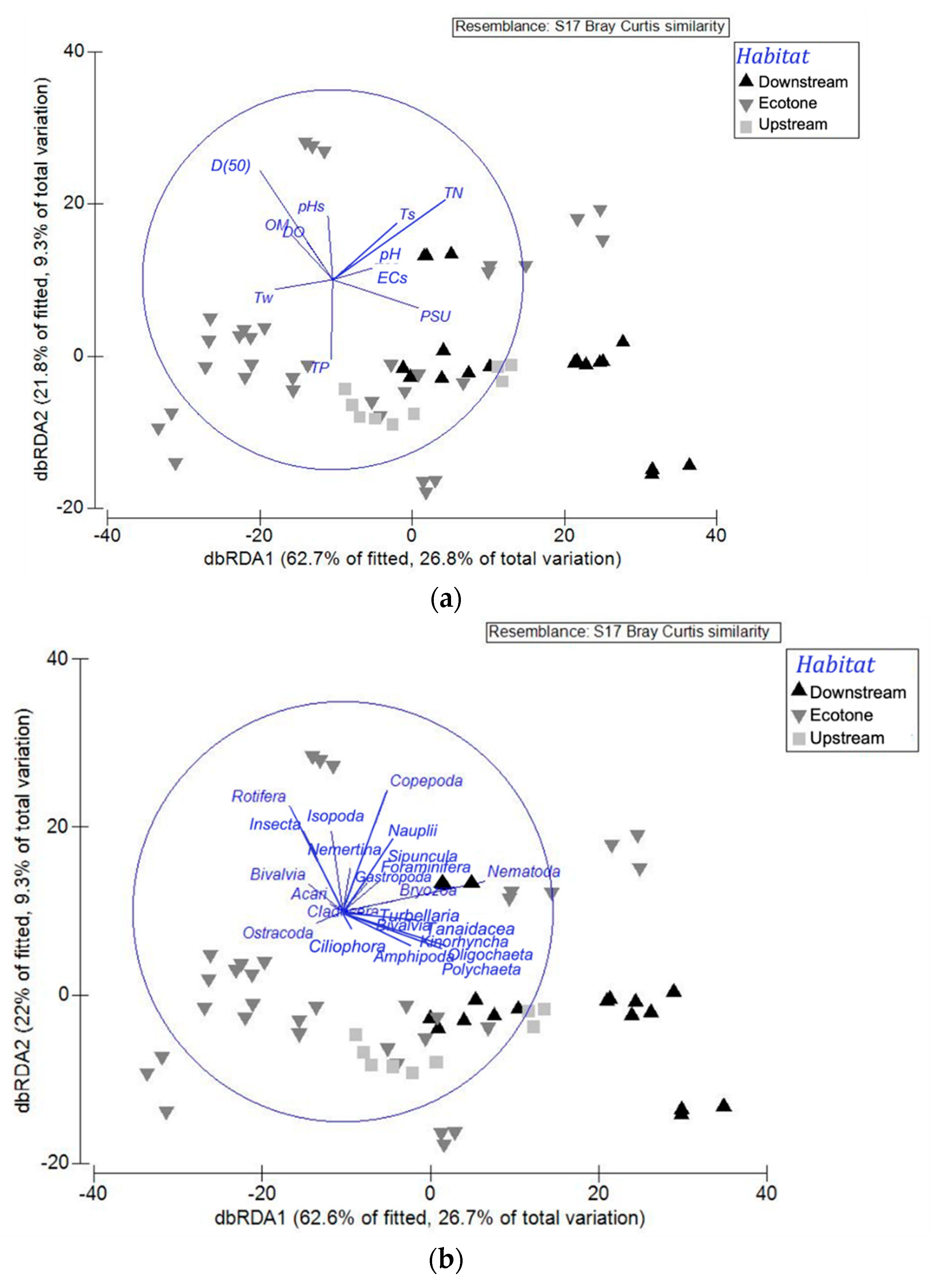
3.4. Meiofauna in Relation to Environmental Variations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sabater, S.; Elosegui, A. Balancing conservation needs with uses of river ecosystems. Acta Biol. Colomb. 2013, 19, 3. [Google Scholar] [CrossRef]
- Tickner, D.; Parker, H.; Moncrieff, C.R.; Oates, N.E.M.; Ludi, E.; Acreman, M. Managing Rivers for Multiple Benefits—A Coherent Approach to Research, Policy and Planning. Front. Environ. Sci. 2017, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Strokal, M.; Bai, Z.; Franssen, W.; Hofstra, N.; Koelmans, A.A.; Ludwig, F.; Ma, L.; Van Puijenbroek, P.; Spanier, J.E.; Vermeulen, L.C.; et al. Urbanization: An Increasing Source of Multiple Pollutants to Rivers in the 21st Century. NPJ Urban Sustain. 2021, 1, 24. [Google Scholar] [CrossRef]
- Siddha, S.; Sahu, P. Impact of Climate Change on the River Ecosystem. In Ecological Significance of River Ecosystems; Elsevier: Amsterdam, The Netherlands, 2022; pp. 79–104. ISBN 978-0-323-85045-2. [Google Scholar]
- Piton, V.; Ouillon, S.; Vinh, V.D.; Many, G.; Herrmann, M.; Marsaleix, P. Seasonal and Tidal Variability of the Hydrology and Suspended Particulate Matter in the Van Uc Estuary, Red River, Vietnam. J. Mar. Syst. 2020, 211, 103403. [Google Scholar] [CrossRef]
- Vinh, V.D.; Ouillon, S.; Thanh, T.D.; Chu, L.V. Impact of the Hoa Binh Dam (Vietnam) on Water and Sediment Budgets in the Red River Basin and Delta. Hydrol. Earth Syst. Sci. 2014, 18, 3987–4005. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.H.; Swennen, R.; Cappuyns, V.; Vassilieva, E.; Neyens, G.; Rajabali, M.; Van Tran, T. Assessment on Pollution by Heavy Metals and Arsenic Based on Surficial and Core Sediments in the Cam River Mouth, Haiphong Province, Vietnam. Soil Sediment Contam. Int. J. 2013, 22, 415–432. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Ouillon, S.; Vu, D.V. Sea-Level Rise in Hai Phong Coastal Area (Vietnam) and Its Response to Enso—Evidence from Tide Gauge Measurement of 1960–2020. Vietnam. J. Earth Sci. 2022, 44, 109–126. [Google Scholar] [CrossRef]
- Nguyen, Y.T.B.; Kamoshita, A.; Dinh, V.T.H.; Matsuda, H.; Kurokura, H. Salinity Intrusion and Rice Production in Red River Delta under Changing Climate Conditions. Paddy Water Environ. 2017, 15, 37–48. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Roberto, R.; Vu, M.C. Potential Use of Reservoirs for Mitigating Saline Intrusion in the Coastal Areas of Red River Delta. In Proceedings of the 2018 IEEE International Conference on Environment and Electrical Engineering and 2018 IEEE Industrial and Commercial Power Systems Europe (EEEIC/I&CPS Europe), Palermo, Italy, 12–15 June 2018; pp. 1–6. [Google Scholar]
- Nguyen, T.H.; Vu, M.C.; Roberto, R. Upstream Effects on Salinity Dynamics in the Red River Delta. In APAC 2019; Trung Viet, N., Xiping, D., Thanh Tung, T., Eds.; Springer: Singapore, 2020; pp. 1439–1443. ISBN 978-9-81150-290-3. [Google Scholar]
- Pereira, C.S.; Lopes, I.; Abrantes, I.; Sousa, J.P.; Chelinho, S. Salinization Effects on Coastal Ecosystems: A Terrestrial Model Ecosystem Approach. Phil. Trans. R. Soc. B 2019, 374, 20180251. [Google Scholar] [CrossRef] [Green Version]
- Du Laing, G.; De Vos, R.; Vandecasteele, B.; Lesage, E.; Tack, F.M.G.; Verloo, M.G. Effect of Salinity on Heavy Metal Mobility and Availability in Intertidal Sediments of the Scheldt Estuary. Estuar. Coast. Shelf Sci. 2008, 77, 589–602. [Google Scholar] [CrossRef]
- Acosta, J.A.; Jansen, B.; Kalbitz, K.; Faz, A.; Martínez-Martínez, S. Salinity Increases Mobility of Heavy Metals in Soils. Chemosphere 2011, 85, 1318–1324. [Google Scholar] [CrossRef]
- Szöcs, E.; Coring, E.; Bäthe, J.; Schäfer, R.B. Effects of Anthropogenic Salinization on Biological Traits and Community Composition of Stream Macroinvertebrates. Sci. Total Environ. 2014, 468–469, 943–949. [Google Scholar] [CrossRef]
- Sowa, A.; Krodkiewska, M. Impact of Secondary Salinisation on the Structure and Diversity of Oligochaete Communities. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 6. [Google Scholar] [CrossRef]
- Tully, K.; Gedan, K.; Epanchin-Niell, R.; Strong, A.; Bernhardt, E.S.; Bendor, T.; Mitchell, M.; Kominoski, J.; Jordan, T.E.; Neubauer, S.C.; et al. Corrigendum: The Invisible Flood: The Chemistry, Ecology, and Social Implications of Coastal Saltwater Intrusion. BioScience 2019, 69, 760. [Google Scholar] [CrossRef] [Green Version]
- Goss, K.F. Environmental Flows, River Salinity and Biodiversity Conservation: Managing Trade-Offs in the Murray—Darling Basin. Aust. J. Bot. 2003, 51, 619. [Google Scholar] [CrossRef]
- Mooney, H.; Larigauderie, A.; Cesario, M.; Elmquist, T.; Hoegh-Guldberg, O.; Lavorel, S.; Mace, G.M.; Palmer, M.; Scholes, R.; Yahara, T. Biodiversity, Climate Change, and Ecosystem Services. Curr. Opin. Environ. Sustain. 2009, 1, 46–54. [Google Scholar] [CrossRef]
- Venâncio, C.; Ribeiro, R.; Lopes, I. Seawater Intrusion: An Appraisal of Taxa at Most Risk and Safe Salinity Levels. Biol. Rev. 2022, 97, 361–382. [Google Scholar] [CrossRef]
- Nguyen, P.T.B.; Koedsin, W.; McNeil, D.; Van, T.P.D. Remote Sensing Techniques to Predict Salinity Intrusion: Application for a Data-Poor Area of the Coastal Mekong Delta, Vietnam. Int. J. Remote Sens. 2018, 39, 6676–6691. [Google Scholar] [CrossRef]
- Li, L.; Zheng, B.; Liu, L. Biomonitoring and Bioindicators Used for River Ecosystems: Definitions, Approaches and Trends. Procedia Environ. Sci. 2010, 2, 1510–1524. [Google Scholar] [CrossRef] [Green Version]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The Natural Indicator of Environmental Pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Hoppit, G.; Schmidt, D.N. A Regional View of the Response to Climate Change: A Meta-Analysis of European Benthic Organisms’ Responses. Front. Mar. Sci. 2022, 9, 896157. [Google Scholar] [CrossRef]
- Oertel, N.; Salánki, J. Biomonitoring and Bioindicators in Aquatic Ecosystems. In Modern Trends in Applied Aquatic Ecology; Ambasht, R.S., Ambasht, N.K., Eds.; Springer: Boston, MA, USA, 2003; pp. 219–246. ISBN 978-1-4613-4972-3. [Google Scholar]
- Pletterbauer, F.; Melcher, A.; Graf, W. Climate Change Impacts in Riverine Ecosystems. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 203–223. ISBN 978-3-319-73249-7. [Google Scholar]
- Ferchichi, H.; Ben Hamouda, M.F.; Farhat, B.; Ben Mammou, A. Assessment of Groundwater Salinity Using GIS and Multivariate Statistics in a Coastal Mediterranean Aquifer. Int. J. Environ. Sci. Technol. 2018, 15, 2473–2492. [Google Scholar] [CrossRef]
- Li, Y.; Huang, D.; Sun, W.; Sun, X.; Yan, G.; Gao, W.; Lin, H. Characterizing Sediment Bacterial Community and Identifying the Biological Indicators in a Seawater-Freshwater Transition Zone during the Wet and Dry Seasons. Environ. Sci. Pollut. Res. 2022, 29, 41219–41230. [Google Scholar] [CrossRef] [PubMed]
- Balsamo, M.; Albertelli, G.; Ceccherelli, V.U.; Coccioni, R.; Colangelo, M.A.; Curini-Galletti, M.; Danovaro, R.; D’Addabbo, R.; De Leonardis, C.; Fabiano, M.; et al. Meiofauna of the Adriatic Sea: Present Knowledge and Future Perspectives. Chem. Ecol. 2010, 26, 45–63. [Google Scholar] [CrossRef]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Arbizu, P.M.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.N.; Sørensen, M.V.; et al. Is the Meiofauna a Good Indicator for Climate Change and Anthropogenic Impacts? Mar. Biodivers. 2015, 45, 505–535. [Google Scholar] [CrossRef] [Green Version]
- Ridall, A.; Ingels, J. Suitability of Free-Living Marine Nematodes as Bioindicators: Status and Future Considerations. Front. Mar. Sci. 2021, 8, 685327. [Google Scholar] [CrossRef]
- Pusceddu, A.; Gambi, C.; Corinaldesi, C.; Scopa, M.; Danovaro, R. Relationships between Meiofaunal Biodiversity and Prokaryotic Heterotrophic Production in Different Tropical Habitats and Oceanic Regions. PLoS ONE 2014, 9, e91056. [Google Scholar] [CrossRef]
- Semprucci, F.; Sbrocca, C.; Rocchi, M.; Balsamo, M. Temporal Changes of the Meiofaunal Assemblage as a Tool for the Assessment of the Ecological Quality Status. J. Mar. Biol. Assoc. 2015, 95, 247–254. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Fiasca, B.; Di Cicco, M.; Cifoni, M.; Galassi, D.M.P. Taxonomic and Functional Trait Variation along a Gradient of Ammonium Contamination in the Hyporheic Zone of a Mediterranean Stream. Ecol. Indic. 2021, 132, 108268. [Google Scholar] [CrossRef]
- Cifoni, M.; Boggero, A.; Rogora, M.; Ciampittiello, M.; Martínez, A.; Galassi, D.M.P.; Fiasca, B.; Di Lorenzo, T. Effects of Human-Induced Water Level Fluctuations on Copepod Assemblages of the Littoral Zone of Lake Maggiore. Hydrobiologia 2022, 849, 3545–3564. [Google Scholar] [CrossRef]
- Broman, E.; Raymond, C.; Sommer, C.; Gunnarsson, J.S.; Creer, S.; Nascimento, F.J.A. Salinity Drives Meiofaunal Community Structure Dynamics across the Baltic Ecosystem. Mol. Ecol. 2019, 28, 3813–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semprucci, F.; Gravina, M.F.; Magni, P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System. Water 2019, 11, 1488. [Google Scholar] [CrossRef] [Green Version]
- Leasi, F.; Sevigny, J.L.; Hassett, B.T. Meiofauna as a Valuable Bioindicator of Climate Change in the Polar Regions. Ecol. Indic. 2021, 121, 107133. [Google Scholar] [CrossRef]
- Udalov, A.; Chikina, M.; Azovsky, A.; Basin, A.; Galkin, S.; Garlitska, L.; Khusid, T.; Kondar, D.; Korsun, S.; Kremenetskiy, V.; et al. Integrity of Benthic Assemblages along the Arctic Estuarine-Coastal System. Ecol. Indic. 2021, 121, 107115. [Google Scholar] [CrossRef]
- Baia, E.; Rollnic, M.; Venekey, V. Seasonality of Pluviosity and Saline Intrusion Drive Meiofauna and Nematodes on an Amazon Freshwater-Oligohaline Beach. J. Sea Res. 2021, 170, 102022. [Google Scholar] [CrossRef]
- Officer, C.B. Physical Dynamics of Estuarine Suspended Sediments. Marine Geology 1981, 40, 1–14. [Google Scholar] [CrossRef]
- Karydis, M.; Tsirtsis, G. Ecological Indices: A Biometric Approach for Assessing Eutrophication Levels in the Marine Environment. Sci. Total Environ. 1996, 186, 209–219. [Google Scholar] [CrossRef]
- Richards, L.A. Diagnosis and Improvement of Saline Alkali Soils, Agriculture, 160, Handbook 60; US Department of Agriculture: Washington, DC, USA, 1954.
- Anderson, M.J.; Gorley, R.N.; Clarke, K.N. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Cunillera-Montcusí, D.; Beklioğlu, M.; Cañedo-Argüelles, M.; Jeppesen, E.; Ptacnik, R.; Amorim, C.A.; Arnott, S.E.; Berger, S.A.; Brucet, S.; Dugan, H.A.; et al. Freshwater Salinisation: A Research Agenda for a Saltier World. Trends Ecol. Evol. 2022, 37, 440–453. [Google Scholar] [CrossRef]
- Root, T.L.; Hall, K.R.; Herzog, M.P.; Howell, C.A. (Eds.) Biodiversity in a Changing Climate: Linking Science and Management in Conservation; University of California Press: Oakland, CA, USA, 2015; ISBN 978-0-520-27885-1. [Google Scholar]
- Bulger, A.J.; Hayden, B.P.; Monaco, M.E.; Nelson, D.M.; McCormick-Ray, M.G. Biologically-Based Estuarine Salinity Zones Derived from a Multivariate Analysis. Estuaries 1993, 16, 311. [Google Scholar] [CrossRef]
- Attrill, M.J. A Testable Linear Model for Diversity Trends in Estuaries. J. Anim. Ecol. 2002, 71, 262–269. [Google Scholar] [CrossRef] [Green Version]
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem; Oxford University Press: Oxford, UK, 2004; ISBN 978-0-19-852508-0. [Google Scholar]
- Soetaert, K.; Vincx, M.; Wittoeck, J.; Tulkens, M. Meiobenthic Distribution and Nematode Community Structure in Five European Estuaries. Hydrobiologia 1995, 311, 185–206. [Google Scholar] [CrossRef]
- Coull, B.C. Long-Term Variability of Estuarine Meiobenthos: An 11 Year Study. Mar. Ecol. Prog. Ser. 1985, 24, 205–218. [Google Scholar] [CrossRef]
- Vincx, M. Diversity of the Nematode Communities in the Southern Bight of the North Sea. Neth. J. Sea Res. 1990, 25, 181–188. [Google Scholar] [CrossRef]
- Coull, B.C. Role of Meiofauna in Estuarine Soft-Bottom Habitats. Austral. Ecol. 1999, 24, 327–343. [Google Scholar] [CrossRef]
- Bouwman, L.A. A Survey of Nematoda from the Ems Estuary: Species Assemblages and Associations. Zool. Jahrb. Syst. Okologie Geogr. Tiere 1983, 110, 345–376. [Google Scholar]
- Heip, C.; Vincx, M.; Vranken, G. The Ecology of Marine Nematodes. In Oceanography and Marine Biology: An Annual Review, 23; Aberdeen University Press: Aberdeen, UK, 1985; pp. 399–489. ISBN 0-08-030397-8. [Google Scholar]
- Pavlyuk, O.; Trebukhova, Y.; Thanh, N.V.; Tu, N.D. Meiobenthos in Estuary Part of Ha Long Bay (Gulf of Tonkin, South China Sea, Vietnam). Ocean Sci. J. 2008, 43, 153–160. [Google Scholar] [CrossRef]
- Adão, H. Dynamic of Meiofauna Communities in Association with Zostera Noltii Seegrass Beds in the Mira SW Portugal. Ph.D. Thesis, University of Evora, Evora, Portugal, 2003. [Google Scholar]
- Alves, A.S.; Adão, H.; Patrício, J.; Neto, J.M.; Costa, M.J.; Marques, J.C. Spatial Distribution of Subtidal Meiobenthos along Estuarine Gradients in Two Southern European Estuaries (Portugal). J. Mar. Biol. Assoc. 2009, 89, 1529–1540. [Google Scholar] [CrossRef]
- Ngo, X.Q.; Smol, N.; Vanreusel, A. The Meiofauna Distribution in Correlation with Environmental Characteristics in 5 Mekong Estuaries, Vietnam. Cah. Biol. Mar. 2013, 54, 71–83. [Google Scholar] [CrossRef]
- Beusen, A.H.W.; Doelman, J.C.; Van Beek, L.P.H.; Van Puijenbroek, P.J.T.M.; Mogollón, J.M.; Van Grinsven, H.J.M.; Stehfest, E.; Van Vuuren, D.P.; Bouwman, A.F. Exploring River Nitrogen and Phosphorus Loading and Export to Global Coastal Waters in the Shared Socio-Economic Pathways. Glob. Environ. Change 2022, 72, 102426. [Google Scholar] [CrossRef]
- Moens, T.; Vincx, M. Temperature, Salinity and Food Thresholds in Two Brackish-Water Bacterivorous Nematode Species: Assessing Niches from Food Absorption and Respiration Experiments. J. Exp. Mar. Biol. Ecol. 2000, 243, 137–154. [Google Scholar] [CrossRef]
- Adão, H.; Alves, A.S.; Patrício, J.; Neto, J.M.; Costa, M.J.; Marques, J.C. Spatial Distribution of Subtidal Nematoda Communities along the Salinity Gradient in Southern European Estuaries. Acta Oecologica. 2009, 35, 287–300. [Google Scholar] [CrossRef]
- Haegerbaeumer, A.; Höss, S.; Ristau, K.; Claus, E.; Heininger, P.; Traunspurger, W. The Use of Meiofauna in Freshwater Sediment Assessments: Structural and Functional Responses of Meiobenthic Communities to Metal and Organics Contamination. Ecol. Indic. 2017, 78, 512–525. [Google Scholar] [CrossRef]
- Warwick, R.; Gee, J. Community Structure of Estuarine Meiobenthos. Mar. Ecol. Prog. Ser. 1984, 18, 97–111. [Google Scholar] [CrossRef]
- Smol, N.; Willems, K.A.; Govaere, J.C.R.; Sandee, A.J.J. Composition, Distribution and Biomass of Meiobenthos in the Oosterschelde Estuary (SW Netherlands). Hydrobiologia 1994, 282–283, 197–217. [Google Scholar] [CrossRef] [Green Version]
- Sinh, N.V.; Kim Phuong, N.T.; Quang, N.X. The Distribution of Meiofauna Community Related to Salinity Gradient in the Ham Luong Estuary, Mekong River. Acad. J. Biol. 2014, 35, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. The Role of Soft-Bottom Benthic Communities in Tropical Mangrove and Coral Reef Ecosystems. Rev. Aquat. Sci. 1989, 1, 243–280. [Google Scholar]
- Fonseca, G.; Netto, S.A. Shallow Sublittoral Benthic Communities of the Laguna Estuarine System, South Brazil. Braz. J. Oceanogr. 2006, 54, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. Intertidal Zonation and Seasonality of Meiobenthos in Tropical Mangrove Estuaries. Mar. Biol. 1987, 95, 447–458. [Google Scholar] [CrossRef]
- Majdi, N.; Threis, I.; Traunspurger, W. It’s the Little Things That Count: Meiofaunal Density and Production in the Sediment of Two Headwater Streams: Meiofauna in Streams. Limnol. Oceanogr. 2017, 62, 151–163. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Nandini, S.; Morales-Ventura, J.; Delgado-Martínez, I.; González-Valverde, L. Effects of NaCl Salinity on the Population Dynamics of Freshwater Zooplankton (Rotifers and Cladocerans). Aquat. Ecol. 2006, 40, 349–360. [Google Scholar] [CrossRef]
- Medeiros, A.; Barbosa, J.; Medeiros, P.; Rocha, R.; Silva, L. Salinity and Freshwater Discharge Determine Rotifer Distribution at the Mossoró River Estuary (Semiarid Region of Brazil). Braz. J. Biol. 2010, 70, 551–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutkowska, A.; Paturej, E.; Kowalska, E. Rotifer trophic state indices as ecosystem indicators in brackish coastal waters. Oceanologia 2013, 55, 887–899. [Google Scholar] [CrossRef]
- Arcifa, M.S.; De Souza, B.B.; De Morais-Junior, C.S.; Bruno, C.G.C. Functional Groups of Rotifers and an Exotic Species in a Tropical Shallow Lake. Sci. Rep. 2020, 10, 14698. [Google Scholar] [CrossRef] [PubMed]
- Widbom, B.; Elmgren, R. Response of Benthic Meiofauna to Nutrient Enrichment of Experimental Marine Ecosystems. Mar. Ecol. Prog. Ser. 1988, 42, 257–268. [Google Scholar] [CrossRef]
- Ristau, K.; Faupel, M.; Traunspurger, W. The Effects of Nutrient Enrichment on a Freshwater Meiofaunal Assemblage: Meiofaunal Response to Nutrient Enrichment. Freshw. Biol. 2012, 57, 824–834. [Google Scholar] [CrossRef]
- De Troch, M.; Roelofs, M.; Riedel, B.; Grego, M. Structural and Functional Responses of Harpacticoid Copepods to Anoxia in the Northern Adriatic: An Experimental Approach. Biogeosciences 2013, 10, 4259–4272. [Google Scholar] [CrossRef] [Green Version]
- Veit-Köhler, G.; Gerdes, D.; Quiroga, E.; Hebbeln, D.; Sellanes, J. Metazoan Meiofauna within the Oxygen-Minimum Zone off Chile: Results of the 2001-PUCK Expedition. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 1105–1111. [Google Scholar] [CrossRef]
- Neira, C.; Ingels, J.; Mendoza, G.; Hernandez-Lopez, E.; Levin, L.A. Distribution of Meiofauna in Bathyal Sediments Influenced by the Oxygen Minimum Zone Off Costa Rica. Front. Mar. Sci. 2018, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Cook, A.A.; John, D.; Lambshead, P.; Hawkins, L.E.; Mitchell, N.; Levin, L.A. Nematode Abundance at the Oxygen Minimum Zone in the Arabian Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 75–85. [Google Scholar] [CrossRef]
- Singh, R.; Ingole, B.S. Structure and Function of Nematode Communities across the Indian Western Continental Margin and Its Oxygen Minimum Zone. Biogeosciences 2016, 13, 191–209. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, T.E.; Jacobsen, D.; Demars, B.O.L.; Brittain, J.E.; Søli, G.; Friberg, N. Effects of Pollution-Induced Changes in Oxygen Conditions Scaling up from Individuals to Ecosystems in a Tropical River Network. Sci. Total Environ. 2022, 814, 151958. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-G.; Kwon, O.; Cho, Y. A Study of Bioindicator Selection for Long-Term Ecological Monitoring. J. Ecol. Environ. 2015, 38, 119–122. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Station | Geographical Location | Description | |
|---|---|---|---|---|
| Latitude | Longitude | |||
| Downstream | VU1 | 20.678319 | 106.700849 | Estuary, dense mangrove forest |
| Downstream | VU2 | 20.682121 | 106.698022 | |
| Downstream | VU3 | 20.688487 | 106.693951 | Mangrove forest, next to a freshwater outlet from agriculture irrigation channel |
| Downstream | VU4 | 20.695359 | 106.685127 | Scattered distribution of mangroves |
| Downstream | VU5 | 20.69464 | 106.652553 | |
| Downstream | VU6 | 20.712204 | 106.613992 | Close to livestock farm (pig) |
| Ecotone | VU7 | 20.747069 | 106.565513 | Close to rice field |
| Ecotone | VU8 | 20.756267 | 106.555787 | Close to rice field |
| Ecotone | VU9 | 20.77239 | 106.548795 | Close to rice field |
| Ecotone | VU10 | 20.772398 | 106.548794 | Close to polychaeta farm |
| Ecotone | VU11 | 20.780523 | 106.544285 | Close to rice field |
| Ecotone | VU12 | 20.783151 | 106.542376 | Close to rice field (26 km from the estuary) |
| Ecotone | VU13 | 20.785923 | 106.541231 | Close to polychaeta farm |
| Ecotone | VU14 | 20.788484 | 106.540525 | Close to rice field |
| Ecotone | VU15 | 20.791128 | 106.53912 | Riverbank under construction |
| Ecotone | VU16 | 20.792836 | 106.536795 | Close to rice field |
| Ecotone | VU17 | 20.794296 | 106.536181 | Close to polychaeta farm, on fertilized soil/mud in the dry season |
| Upstream | VU18 | 20.791533 | 106.525937 | Industrial zone |
| Upstream | VU19 | 20.786903 | 106.519853 | |
| Upstream | VU20 | 20.791663 | 106.513265 | Close to rice field |
| Abiotic Factor | Downstream | Ecotone | Upstream | One-Way PERMANOVA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Min.–Max. | Mean ± SD | Min.–Max. | Mean ± SD | Min.–Max. | df | MS | Pseudo-F | p | |
| PSU | 4.07 ± 0.84 | 3.16–5.75 | 0.54 ± 0.19 | 0.22–0.83 | 0.24 ± 0.1 | 0.11–0.31 | 2 | 27.272 | 348.8 | 0.001 |
| ECs (μScm−1) | 6503 ± 2048 | 3800–9400 | 1064 ± 270 | 640–1480 | 583 ± 75 | 520–680 | 2 | 24.8 | 150.39 | 0.001 |
| Ts (°C) | 26.48 ± 0.49 | 25.8–27.1 | 26.23 ± 0.42 | 25.8–27.2 | 25.87 ± 0.43 | 25.4–26.4 | 2 | 5.0129 | 5.8345 | 0.007 |
| Tw (°C) | 25.94 ± 0.19 | 25.7–26.2 | 25.90 ± 0.16 | 25.6–26.2 | 25.44 ± 0.02 | 25.4–25.5 | 2 | 4.9071 | 5.6221 | 0.04 |
| TN (%) | 0.72 ± 0.24 | 0.39–1.04 | 2.13 ± 0.79 | 0.48–3.52 | 2.49 ± 0.09 | 2.36–2.57 | 2 | 16.92 | 38.335 | 0.001 |
| TP (%) | 0.14 ± 0.02 | 0.10–0.16 | 0.16 ± 0.01 | 0.14–0.18 | 0.16 ± 0.0005 | 0.16–0.161 | 2 | 5.3827 | 6.3609 | 0.006 |
| OM (%) | 3.39 ± 0.45 | 2.81–3.99 | 4.04 ± 0.48 | 2.77–4.66 | 3.95 ± 0.63 | 3.32–4.74 | 2 | 7.7696 | 10.19 | 0.001 |
| D(50) (μm) | 49.21 ± 27.27 | 22–98.9 | 56.21 ± 22.44 | 27.7–109 | 60.07 ± 15.61 | 38.6–76.7 | 2 | 0.8199 | 0.8148 | 0.45 |
| DO (mgL−1) | 5.59 ± 0.46 | 4.89–6.42 | 4.18 ± 0.23 | 3.88–4.73 | 3.22 ± 0.24 | 2.9–3.41 | 2 | 25.685 | 191.91 | 0.001 |
| pHw | 7.78 ± 0.1 | 7.54–7.92 | 7.81 ± 0.08 | 7.64–8.05 | 7.67 ± 0.06 | 7.61–7.77 | 2 | 7.2054 | 9.2108 | 0.001 |
| pHs | 7.2 ± 0.33 | 6.49–7.59 | 7.39 ± 0.12 | 7.2–7.72 | 7.38 ± 0.08 | 7.29–7.49 | 2 | 4.5527 | 5.201 | 0.016 |
| One-Way PERMANOVA | ||||||
| Source | df | SS | MS | Pseudo-F | p (perm) | Permutation |
| Habitat | 2 | 4133.4 | 2066.7 | 3.0274 | 0.003 | 998 |
| Res | 57 | 38,911 | 682.65 | |||
| Total | 59 | 43,045 | ||||
| Pairwise Test | ||||||
| Groups | t | p (perm) | perms | p (MC) | ||
| Downstream, Ecotone | 2.1901 | 0.001 | 999 | 0.001 | ||
| Downstream, Upstream | 1.4951 | 0.032 | 999 | 0.056 | ||
| Ecotone, Upstream | 1.1445 | 0.236 | 999 | 0.26 | ||
| Station | Average Similarity (%) | Taxa Contributions (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Nematodes | Rotifers | Polychaeta | Turbellaria | Nauplii | Copepods | Amphipoda | Bivalves | ||
| VU1 | 60.85 | 95.24 | |||||||
| VU2 | 92.88 | 95.86 | |||||||
| VU3 | 73.88 | 93.23 | 1.56 | 1.48 | |||||
| VU4 | 68.43 | 73.42 | 17.07 | 1.98 | 2.74 | ||||
| VU5 | 66.21 | 83.88 | 2.14 | 7.25 | 3.84 | ||||
| VU6 | 75.45 | 80.79 | 10.33 | 2.58 | 2.88 | ||||
| VU7 | 81.70 | 81.54 | 8.19 | 3.24 | 3.74 | ||||
| VU8 | 63.80 | 91.18 | 5.44 | ||||||
| VU9 | 91.28 | 73.22 | 14.93 | 5.54 | 5.02 | ||||
| VU10 | 74.75 | 89.51 | 7.94 | ||||||
| VU11 | 82.46 | 97.21 | |||||||
| VU12 | 77.85 | 93.22 | 3.05 | ||||||
| VU13 | 80.13 | 99.14 | |||||||
| VU14 | 65.11 | 23.95 | 72.93 | ||||||
| VU15 | 70.26 | 16.82 | 81.50 | ||||||
| VU16 | 79.76 | 75.75 | 12.09 | 9.16 | |||||
| VU17 | 88.69 | 90.98 | 2.99 | 2.18 | |||||
| VU18 | 82.48 | 93.34 | 2.53 | ||||||
| VU19 | 80.44 | 83.35 | 8.71 | 4.12 | |||||
| VU20 | 65.94 | 82.13 | 3.72 | 2.77 | 3.06 | 5.30 | |||
| Marginal Tests | Sequential Tests | ||||||
|---|---|---|---|---|---|---|---|
| Variables | Pseudo-F | p | Variables | R2 | Pseudo-F | p | Res. df |
| pHw | 0.80628 | 0.483 | (+) pHw | 0.013 | 0.80628 | 0.459 | 58 |
| pHs | 0.8369 | 0.498 | (+) pHs | 0.025 | 1.5032 | 0.207 | 57 |
| DO | 2.2615 | 0.072 | (+) DO | 0.056 | 1.0279 | 0.409 | 56 |
| Ecs | 3.0077 | 0.03 | (+) Ecs | 0.084 | 1.7092 | 0.142 | 55 |
| PSU | 4.3192 | 0.01 | (+) PSU | 0.152 | 4.2724 | 0.01 | 54 |
| OM | 1.6501 | 0.171 | (+) OM | 0.173 | 1.3829 | 0.231 | 53 |
| TP | 1.466 | 0.235 | (+) TP | 0.228 | 3.6862 | 0.009 | 52 |
| TN | 1.7807 | 0.131 | (+) TN | 0.287 | 4.2498 | 0.008 | 51 |
| Tw | 0.35302 | 0.842 | (+) Tw | 0.299 | 0.82954 | 0.451 | 50 |
| Ts | 2.4743 | 0.064 | (+) Ts | 0.371 | 5.5701 | 0.001 | 49 |
| D(50) | 4.5717 | 0.006 | (+) D(50) | 0.428 | 4.7905 | 0.002 | 48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, H.T.; Gourdon, L.; Bui, H.V.; Dao, D.T.; Mai, H.; Do, H.M.; Nguyen, T.V.; Ouillon, S. Ecological Responses of Meiofauna to a Saltier World—A Case Study in the Van Uc River Continuum (Vietnam) in the Dry Season. Water 2023, 15, 1278. https://doi.org/10.3390/w15071278
Nguyen HT, Gourdon L, Bui HV, Dao DT, Mai H, Do HM, Nguyen TV, Ouillon S. Ecological Responses of Meiofauna to a Saltier World—A Case Study in the Van Uc River Continuum (Vietnam) in the Dry Season. Water. 2023; 15(7):1278. https://doi.org/10.3390/w15071278
Chicago/Turabian StyleNguyen, Hien Thanh, Lucie Gourdon, Hoi Van Bui, Duong Thanh Dao, Huong Mai, Hao Manh Do, Thanh Vu Nguyen, and Sylvain Ouillon. 2023. "Ecological Responses of Meiofauna to a Saltier World—A Case Study in the Van Uc River Continuum (Vietnam) in the Dry Season" Water 15, no. 7: 1278. https://doi.org/10.3390/w15071278