
Genetic Diversity and Population Structure of the Asian Tiger Mosquito (Aedes albopictus) in Vietnam: Evidence for Genetic Differentiation by Climate Region


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Materials
2.2. DNA Extraction and Sequencing
2.3. Genetic Diversity and Demographic History
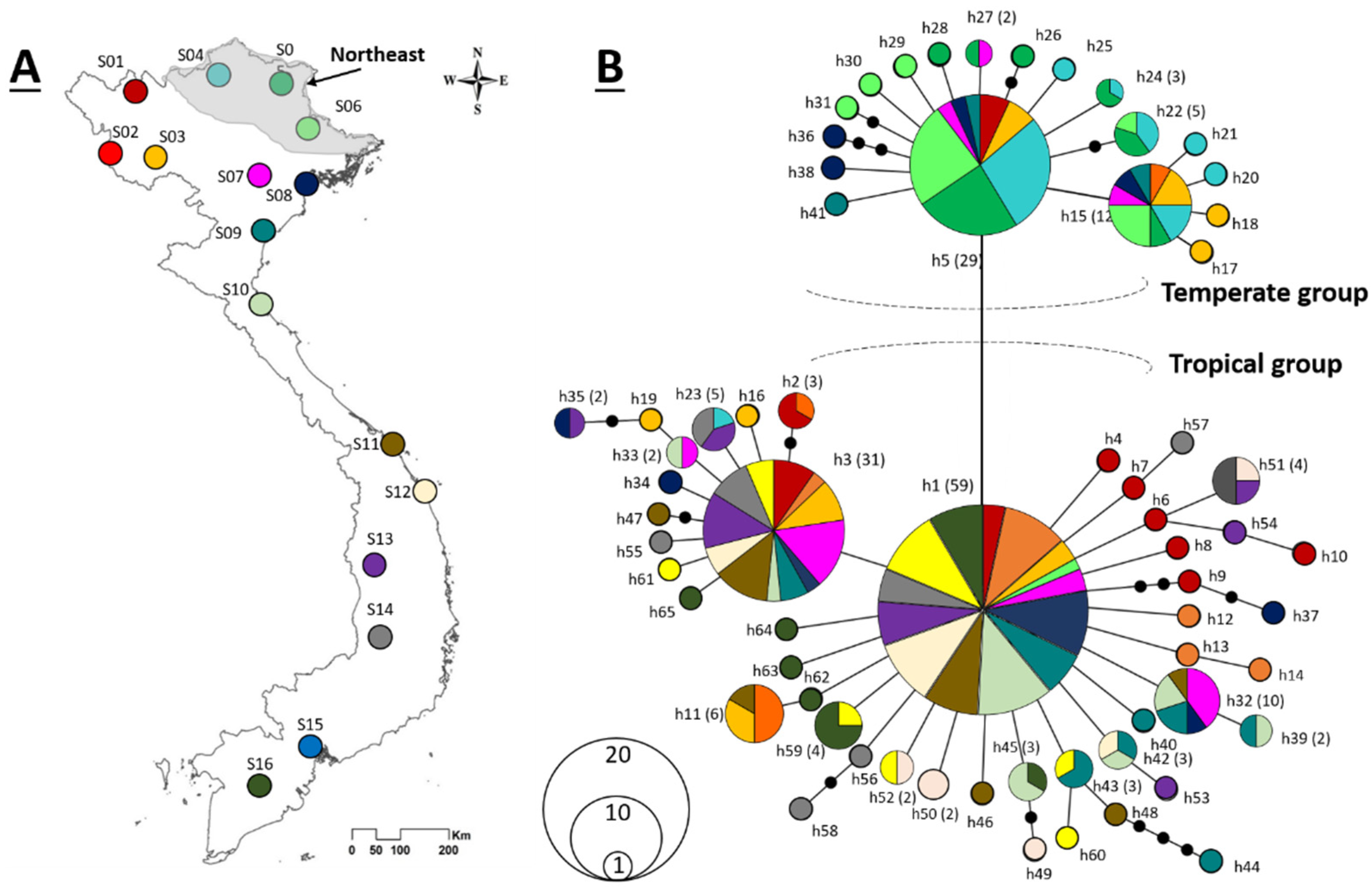
2.4. Haplotype Network Analysis
2.5. Population Differentiation and Structure
2.6. Effect of Geography and Climate on Genetic Variation
~ geography (geo1+geo2+geo3)
3. Results
3.1. Genetic Diversity
3.2. Haplotype Network
3.3. Genetic Differentiation and Population Structure
3.4. Effect of Geography and Climate on Genetic Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, An arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1785. [Google Scholar] [CrossRef]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface. 2012, 9, 2708–2717. [Google Scholar] [CrossRef] [Green Version]
- Hawley, W.A.; Reiter, P.; Copeland, R.S.; Pumpuni, C.B.; Craig, G.B. Aedes albopictus in North America: Probable introduction in used tires from northern Asia. Science 1987, 236, 1114–1116. [Google Scholar] [CrossRef]
- Scholte, E.J.; Schaffner, F. Waiting for the tiger: Establishment and spread of the Asian tiger mosquito in Europe. Emerg. Pests Vector-Borne Dis. Eur. 2007, 14, 241–261. [Google Scholar]
- Urbanski, J.M.; Benoit, J.B.; Michaud, M.R.; Denlinger, D.L.; Armbruster, P. The molecular physiology of increased egg desiccation resistance during diapause in the invasive mosquito, Aedes albopictus. Proc. R. Soc. B. Biol. Sci. 2010, 277, 2683–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Franco, J.G.; Craig, G.B. Biology, relationships, and control of Aedes albopictus. Pan. Am. Health Org. Tech. Pap. 2002, 42, 1–61. [Google Scholar]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control. Assoc. Suppl. 1988, 1, 1–39. [Google Scholar]
- Birungi, J.; Munstermann, L.E. Genetic structure of Aedes albopictus (Diptera: Culicidae) populations based on mitochondrial ND5 sequences: Evidence for an independent invasion into Brazil and United States. Ann. Entomol. Soc. Am. 2006, 95, 125–132. [Google Scholar] [CrossRef]
- Maynard, A.J.; Ambrose, L.; Cooper, R.D.; Chow, W.K.; Davis, J.B.; Muzari, M.O.; Hurk, A.F.; Hall-Mendelin, S.; Hasty, J.M.; Burkot, T.R.; et al. Tiger on the prowl: Invasion history and spatio-temporal genetic structure of the Asian tiger mosquito Aedes albopictus (Skuse 1894) in the Indo-Pacific. PLoS Negl. Trop. Dis. 2017, 11, e0005546. [Google Scholar] [CrossRef]
- Motoki, M.T.; Fonseca, D.M.; Miot, E.F.; Demari-Silva, B.; Thammavong, P.; Chonephetsarath, S.; Phommavanh, N.; Hertz, J.C.; Kittayapong, P.; Brey, P.T.; et al. Population genetics of Aedes albopictus (Diptera: Culicidae) in its native range in Lao People’s Democratic Republic. Parasites Vectors 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Zhong, D.; Lo, E.; Hu, R.; Metzger, M.E.; Cummings, R.; Bonizzoni, M.; Fujioka, K.K.; Sorvillo, T.E.; Kluh, S.; Healy, S.P.; et al. Genetic analysis of invasive Aedes albopictus populations in Los Angeles County, California and its potential public health impact. PLoS ONE 2013, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratz, N.G. Critical review of the vector status of Aedes Albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Rezza, G. Aedes albopictus and the reemergence of Dengue. BMC Public Health 2012, 12, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenzie, B.A.; Wilson, A.E.; Zohdy, S. Aedes albopictus is a competent vector of Zika virus: A meta-analysis. PLoS ONE 2019, 14, e0216794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagès, F.; Peyrefitte, C.N.; Mve, M.T.; Jarjaval, F.; Brisse, S.; Iteman, I.; Gravier, P.; Nkoghe, D.; Grandadam, M. Aedes albopictus mosquito: The main vector of the 2007 chikungunya outbreak in Gabon. PLoS ONE 2009, 4, e4691. [Google Scholar] [CrossRef]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef] [Green Version]
- Rollins, L.A.; Woolnough, A.P.; Sherwin, W.B. Population genetic tools for pest management: A review. Wildl. Res. 2006, 33, 251–261. [Google Scholar] [CrossRef]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- Alto, B.W.; Juliano, S.A. Precipitation and temperature effects on populations of Aedes albopictus (Diptera: Culicidae): Implications for range expansion. J. Med. Entomol. 2001, 38, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Kamgang, B.; Brengues, C.; Fontenille, D.; Njiokou, F.; Simard, F.; Paupy, C. Genetic structure of the tiger mosquito, Aedes albopictus, in Cameroon (Central Africa). PLoS ONE 2011, 6, e20257. [Google Scholar] [CrossRef] [Green Version]
- Mousson, L.; Dauga, C.; Garrigues, T.; Schaffner, F.; Vazeille, M.; Failloux, A.B. Phylogeography of Aedes (Stegomyia) aegypti (L.) and Aedes (Stegomyia) albopictus (Skuse) (Diptera: Culicidae) based on mitochondrial DNA variations. Genet. Res. 2005, 86, 1–11. [Google Scholar] [CrossRef]
- Raharimalala, F.N.; Ravaomanarivo, L.H.; Ravelonandro, P.; Rafarasoa, L.S.; Zouache, K.; Tran, V.V.; Mousson, L.; Failloux, A.; Hellard, E.; Moro, C.V.; et al. Biogeography of the two major arbovirus mosquito vectors, Aedes aegypti and Aedes albopictus (Diptera, Culicidae), in Madagascar. Parasit. Vectors. 2012, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.N.; Nguyen, T.H. Climate and Climate Resources in Vietnam; Agriculture Publisher: Hanoi, Vietnam, 2004. [Google Scholar]
- Higa, Y.; Nguyen, T.Y.; Kawada, H.; Tran, H.S.; Nguyen, T.H.; Takagi, M. Geographic distribution of Aedes aegypti and Aedes albopictus collected from used tires in Vietnam. J. Am. Mosq. Control. Assoc. 2010, 26, 1–9. [Google Scholar] [CrossRef]
- Schmid, M.; Guillaume, F. The role of phenotypic plasticity on population differentiation. Heredity 2017, 119, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, R.; Phan, T.N.; Nguyen, T.Y.; Hoshino, K.; Isawa, H.; Higa, Y.; Nguyen, V.H.; Bui, M.T.; Do, P.L.; Tran, V.P.; et al. Surveillance of Japanese encephalitis virus infection in mosquitoes in Vietnam from 2006 to 2008. Am. J. Trop. Med. Hyg. 2013, 88, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.K.L.; Briant, L.; Gavotte, L.; Labbe, P.; Perriat-Sanguinet, M.; Cornillot, E.; Vu, T.D.; Nguyen, T.Y.; Tran, V.P.; Nguyen, V.S.; et al. Incidence of dengue and chikungunya viruses in mosquitoes and human patients in border provinces of Vietnam. Parasit. Vectors 2017, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.Q.; Duong, T.H.K.; Rabaa, M.; Nguyen, M.T.; Tran, T.V.; Le, V.T.; Nguyen, T.H.; Ha, M.T.; Ta, V.T.; Nguyen, L.D.H.; et al. Chikungunya and Zika virus cases detected against a backdrop of endemic dengue transmission in Vietnam. Am. J. Trop. Med. Hyg. 2017, 97, 146–150. [Google Scholar]
- Nguyen, T.T.; Lundkvist, Å.; Lindahl, J. Urban transmission of mosquito-borne flaviviruses–a review of the risk for humans in Vietnam. Infect. Ecol. Epidemiol. 2019, 9, 1660129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-S.; Mogasale, V.; Lim, J.K.; Carabali, M.; Lee, K.-S.; Sirivichayakul, C.; Dang, D.A.; Palencia-Florez, D.C.; Nguyen, T.H.A.; Riewpaiboon, A.; et al. A multi-country study of the economic burden of dengue fever: Vietnam, Thailand, and Colombia. PLoS Negl. Trop. Dis. 2017, 11, e0006037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harving, M.; Ronsholt, F. The economic impact of dengue hemorrhagic fever on family level in Southern Vietnam. Dan. Med. Bull. 2007, 54, 170–172. [Google Scholar] [PubMed]
- Huang, Y.M. The subgenus stegomyia of aedes in the oriental region with keys to the species (Diptera: Culicidae). Med. Entomol. Stud. 1979, 15, 1–79. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Mol. Ecol. Notes. 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Cira, D.; Arish, D.; Kilroy, A.; Lozano, N.; Wang, H.G. Vietnam Urbanization Review. 2012. Available online: https://www.researchgate.net/publication/272088348_Vietnam_Urbanization_Review (accessed on 23 August 2020).
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Wagner, J. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/package=vegan (accessed on 19 July 2020).
- Rousset, F. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genetics 1997, 145, 1219–1228. [Google Scholar] [CrossRef]
- Moore, M.; Sylla, M.; Goss, L.; Burugu, M.W.; Sang, R.; Kamau, L.W.; Kenya, E.U.; Bosio, C.; Munoz, M.L.; Sharakova, M.; et al. Dual African origins of global Aedes aegypti s.l. populations revealed by mitochondrial DNA. PLoS Negl. Trop. Dis. 2013, 7, e2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Stephan, W. Inferring the demographic history and rate of adaptive substitution in drosophila. PLoS Genet. 2006, 2, e166. [Google Scholar] [CrossRef] [Green Version]
- Bergsten, J.; Bilton, D.T.; Fujisawa, T.; Elliott, M.; Monaghan, M.T.; Balke, M.; Hendrich, L.; Geijer, J.; Herrmann, J.; Foster, G.N.; et al. The effect of geographical scale of sampling on DNA barcoding. Syst. Biol. 2012, 61, 851–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, V.; Gabrieli, P.; Brandini, S.; Capodiferro, M.R.; Javier, P.A.; Chen, X.G.; Achillo, A.; Semino, O.; Gomulski, L.M.; Malacrida, A.R.; et al. The worldwide spread of the tiger mosquito as revealed by mitogenome haplogroup diversity. Front. Genet. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsakiozi, P.; Richardson, J.B.; Pichler, V.; Favia, G.; Martins, A.J.; Urbanelli, S.; Armbruster, P.A.; Caccone, A. Population genomics of the Asian tiger mosquito, Aedes albopictus: Insights into the recent worldwide invasion. Ecol. Evol. 2017, 7, 10143–10157. [Google Scholar] [CrossRef] [Green Version]
- Žitko, T.; Kovavic, A.; Desdevises, Y.; Puizina, J. Genetic variation in East-Adriatic populations of the Asian tiger mosquito, Aedes albopictus (Diptera: Culicidae), inferred from NADH5 and COI sequence variability. Eur. J. Entomol. 2011, 108, 501–508. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Zhang, J.; Wu, R.; Xue, B.; Qian, Q.; Gao, B. Genetic polymorphism study on Aedes albopictus of different geographical regions based on DNA barcoding. BioMed. Res. Int. 2018, 2018, 1501430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawani, M.K.N.; Abu, H.A.; Sazaly, A.B.; Zary, S.Y.; Darlina, M.N. Population genetic structure of Aedes albopictus in Penang, Malaysia. Genet. Mol. Res. 2014, 13, 8184–8196. [Google Scholar] [CrossRef] [PubMed]
- Porretta, D.; Mastrantonio, V.; Bellini, R.; Somboon, P.; Urbanelli, S. Glacial history of a modern invader: Phylogeography and species distribution modelling of the Asian tiger mosquito Aedes Albopictus. PLoS ONE 2012, 7, e44515. [Google Scholar] [CrossRef]
- Lee, E.; Yang, S.C.; Kim, T.K.; Noh, B.E.; Lee, H.S.; Kim, H.; Roh, J.Y.; Lee, W.G. Geographical genetic variation and sources of Korean Aedes albopictus (Diptera: Culicidae) populations. J. Med. Entomol. 2020, 57, 1057–1068. [Google Scholar] [CrossRef]
- Ismail, N.A.; Adilah-Amrannudin, N.; Hamsidi, M.; Ismail, R.; Dom, N.C.; Ahmad, A.H.; Mastuki, M.F.; Camalxaman, A.N. The genetic diversity, haplotype analysis, and phylogenetic relationship of Aedes albopictus (Diptera: Culicidae) based on the Cytochrome oxidase 1 marker: A Malaysian scenario. J. Med. Entomol. 2017, 54, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Gomulski, L.M.; Aketarawong, N.; Tait, G.; Scolari, F.; Somboon, P.; Guglielmino, C.R.; Malacrida, A.R.; Gasperi, G. Molecular markers for analyses of intraspecific genetic diversity in the Asian tiger mosquito, Aedes albopictus. Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.M.H.; Phan, V.T.; Le, N.Q.; Nguyen, Q.T. Extreme climatic events over Vietnam from observational data and RegCM3 projections. Clim. Res. 2011, 49, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, S.V.; Vu, M.T.; Liong, S.Y. Regional climate simulations over Vietnam using the WRF model. Theor. Appl. Climatol. 2016, 126, 161–182. [Google Scholar] [CrossRef]
- Phan, V.T.; Ngo, D.T. Seasonal and interannual variations of surface climate elements over Vietnam. Clim. Res. 2009, 40, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Mathias, D.; Reed, L.K.; Bradshaw, W.E.; Holzapfel, C.M. Evolutionary divergence of circadian and photoperiodic phenotypes in the pitcher-plant mosquito, Wyeomyia smithii. J. Biol. Rhythm. 2006, 21, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, W.E.; Zani, P.A.; Holzapfel, C.M. Adaptation to temperate climates. Evolution 2004, 58, 1748–1762. [Google Scholar] [CrossRef]
- Armbruster, P.A. Photoperiodic diapause and the establishment of Aedes albopictus (Diptera: Culicidae) in North America. J. Med. Entomol. 2016, 53, 1013–1023. [Google Scholar] [CrossRef] [Green Version]
- Sota, T.; Mogi, M. Survival time and resistance to desiccation of diapause and non-diapause eggs of temperate Aedes (Stegomyia) mosquitoes. Entomol. Exp. Appl. 1992, 63, 155–161. [Google Scholar] [CrossRef]
- Batz, Z.A.; Armbruster, P.A. Diapause-associated changes in the lipid and metabolite profiles of the Asian tiger mosquito, Aedes albopictus. J. Exp. Biol. 2018, 221, jeb189480. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, T.; Chaves, L.F.; Nguyen, T.T.G.; Nguyen, T.Y.; Takagi, M. Winter activity and diapause of Aedes albopictus (Diptera: Culicidae) in Hanoi, northern Vietnam. J. Med. Entomol. 2015, 52, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Morales Vargas, R.E.; Phumala-Morales, N.; Tsunoda, T.; Apiwathnasorn, C.; Dujardin, J.P. The phenetic structure of Aedes albopictus. Infect. Genet. Evol. 2013, 13, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Ouyang, Z.; Nguyen, D.D.; Nguyen, T.T.H.; Park, H.; Chen, J. Urbanization, economic development, environmental and social changes in transitional economies: Vietnam after Doimoi. Landsc. Urban. Plan. 2019, 187, 145–155. [Google Scholar] [CrossRef]
- Li, Y.; Kamara, F.; Zhou, G.; Puthiyakunnon, S.; Li, C.; Liu, Y.; Zhou, Y.; Yao, L.; Yan, G.; Chen, X.G. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 2014, 8, e3301. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | CR | Locality | N | No. Haplotypes (NH) | Haplotype Diversity (Hd) (SD) | Nucleotide Diversity (π) (SD) | Tajima’s D | Fu’s Fs |
|---|---|---|---|---|---|---|---|---|
| S01 | NW | Lai Chau province | 15 | 10 | 0.9429 (0.0403) | 0.0037 (0.0024) | −1.3221 | −5.1246 |
| S02 | Dien Bien province | 15 | 8 | 0.8286 (0.0849) | 0.0030 (0.0020) | −1.3545 | −3.1153 | |
| S03 | Son La province | 15 | 9 | 0.9333 (0.0397) | 0.0036 (0.0023) | −0.1473 | −3.7633 | |
| S04 | NE | Ha Giang province | 17 | 8 | 0.7794 (0.0985) | 0.0025 (0.0021) | −1.6227 | −3.3956 |
| S05 | Cao Bang province | 15 | 7 | 0.7810 (0.1016) | 0.0021 (0.0016) | −1.5728 | −3.0275 | |
| S06 | Lang Son province | 15 | 7 | 0.7714 (0.1001) | 0.0019 (0.0016) | −1.7724 | −3.3800 | |
| S07 | ND | Hanoi city | 15 | 7 | 0.8381 (0.0680) | 0.0024 (0.0016) | 0.5623 | −2.6767 |
| S08 | Hai Phong city | 15 | 10 | 0.8571 (0.0901) | 0.0042 (0.0026) | −1.5855 | −4.4915 | |
| S09 | NC | Thanh Hoa province | 17 | 11 | 0.9338 (0.0426) | 0.0032 (0.0021) | −1.5294 | −6.8099 |
| S10 | Nghe An province | 15 | 7 | 0.7810 (0.1016) | 0.0020 (0.0014) | −1.1519 | −3.4900 | |
| S11 | SC | Da Nang city | 14 | 7 | 0.8242 (0.0781) | 0.0025 (0.0017) | −1.3494 | −2.7610 |
| S12 | Quang Ngai province | 14 | 7 | 0.8132 (0.1158) | 0.0023 (0.0017) | −1.7663 | −2.9501 | |
| S13 | CH | Gia Lai province | 14 | 7 | 0.8571 (0.0652) | 0.0031 (0.0020) | −1.3491 | −1.9899 |
| S14 | Dak Lak province | 14 | 8 | 0.9121 (0.0440) | 0.0032 (0.0021) | −0.6081 | −3.1394 | |
| S15 | S | Ho Chi Minh city | 13 | 7 | 0.7949 (0.1091) | 0.0020 (0.0015) | −1.2441 | −3.8060 |
| S16 | Can Tho province | 13 | 7 | 0.8333 (0.0861) | 0.0020 (0.0015) | −1.5939 | −3.7332 | |
| Total | 236 | 65 | 0.9020 | 0.0033 | −2.3260 | −26.8672 |
| Hypothesis | Source of Variation | Percent of Variation | F-Statistic | P-Value |
|---|---|---|---|---|
| Two groups: PCoA plot and haplotype distribution map - Northeast (S04, S05, S06) - The other populations. | ΦCT | 29.14 | 0.2914 | <0.001 |
| ΦSC | 3.15 | 0.0445 | <0.001 | |
| ΦST | 67.71 | 0.3229 | <0.001 | |
| Three groups: spatial distribution of haplotypes (Figure 2A) - Region 1 (S04, S05, S06) - Region 2 (S01, S02, S03, S07, S08, S09) - Region 3 (S10, S11, S12, S13, S14, S15, S16) | ΦCT | 18.36 | 0.1836 | <0.001 |
| ΦSC | 2.71 | 0.0302 | <0.01 | |
| ΦST | 78.93 | 0.2107 | <0.001 | |
| Seven groups, geography and climate - Northwest (S01, S02, S03) - Northeast (S04, S05, S06) - North Delta (S07, S08) - North Central (S09, S10) - South Central (S11, S12) - Central Highlands (S13, S14) - The South (S15, S16) | ΦCT | 15.99 | 0.1599 | <0.001 |
| ΦSC | 1.45 | 0.0173 | 0.06158 | |
| ΦST | 80.25 | 0.1744 | <0.01 | |
| Three groups: level of urbanization - Class I, municipalities (S7, S8, S11, S15, S16) - Class II, provincial cities (S1, S2, S3, S4, S5, S6, S13, S14) - Class III, provincial towns (S9, S10, S12) | ΦCT | 3.23 | 0.0323 | 0.1329 |
| ΦSC | 13.88 | 0.1434 | <0.001 | |
| ΦST | 82.89 | 0.1711 | <0.001 |
| Variable | Inertia | Percent of Variation (%) | P-Value |
|---|---|---|---|
| Global dbRDA | |||
| Geographic distance + Climates | 0.2961 | 75.98 | 0.002 |
| Partial dbRDA | |||
| Climates | 0.2516 | 64.55 | 0.001 |
| Geographic distance | 0.1314 | 33.73 | 0.010 |
| Climates | Geography | 0.1646 | 42.25 | 0.002 |
| Geography | Climates | 0.0445 | 11.42 | 0.020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duong, C.-V.; Kang, J.-H.; Nguyen, V.-V.; Bae, Y.-J. Genetic Diversity and Population Structure of the Asian Tiger Mosquito (Aedes albopictus) in Vietnam: Evidence for Genetic Differentiation by Climate Region. Genes 2021, 12, 1579. https://doi.org/10.3390/genes12101579
Duong C-V, Kang J-H, Nguyen V-V, Bae Y-J. Genetic Diversity and Population Structure of the Asian Tiger Mosquito (Aedes albopictus) in Vietnam: Evidence for Genetic Differentiation by Climate Region. Genes. 2021; 12(10):1579. https://doi.org/10.3390/genes12101579
Chicago/Turabian StyleDuong, Cuong-Van, Ji-Hyoun Kang, Vinh-Van Nguyen, and Yeon-Jae Bae. 2021. "Genetic Diversity and Population Structure of the Asian Tiger Mosquito (Aedes albopictus) in Vietnam: Evidence for Genetic Differentiation by Climate Region" Genes 12, no. 10: 1579. https://doi.org/10.3390/genes12101579