Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea)

Abstract
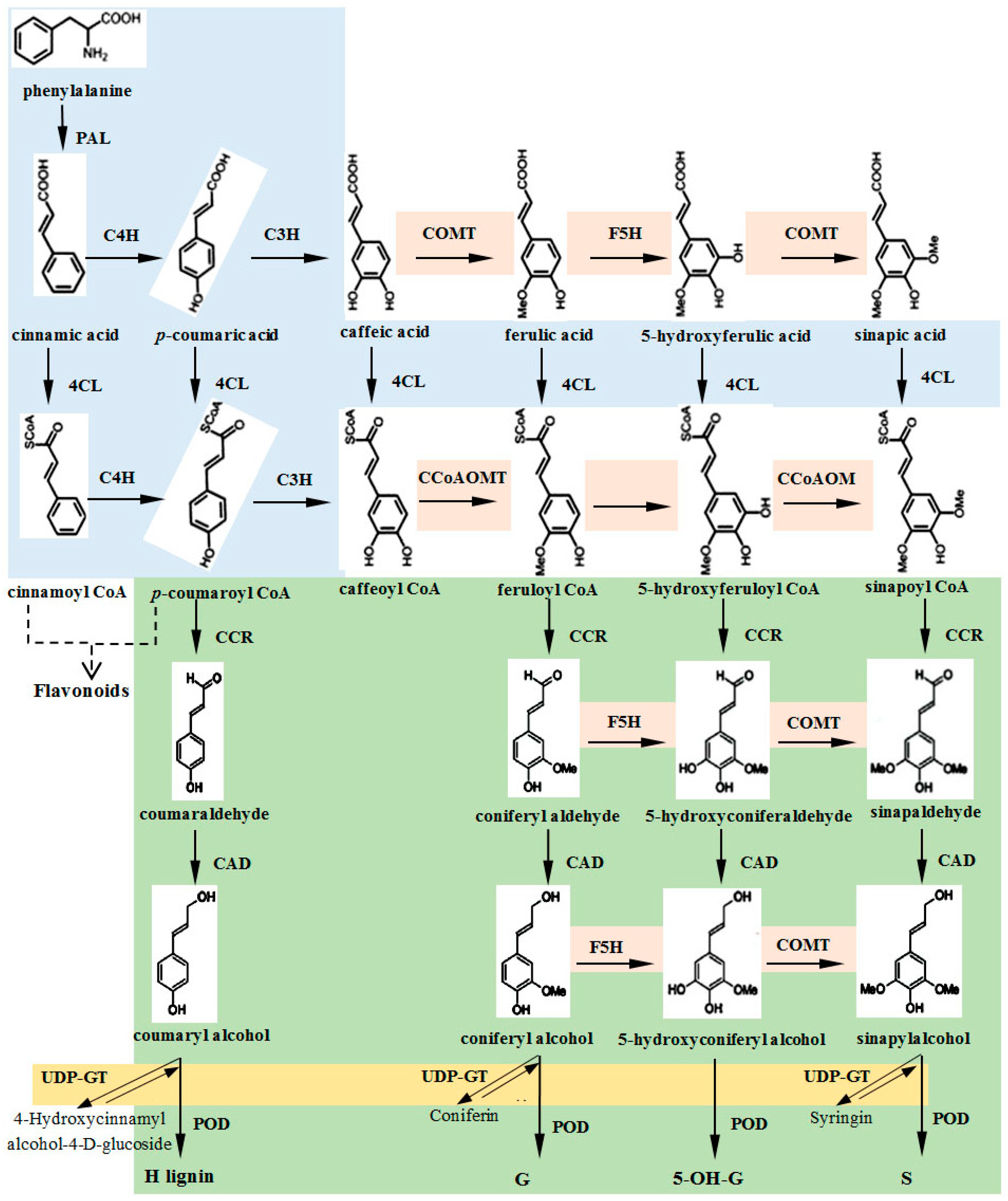
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Total RNA Extraction and Integrity Testing
2.3. Determination of Lignin Content
2.4. qRT-PCR Analysis
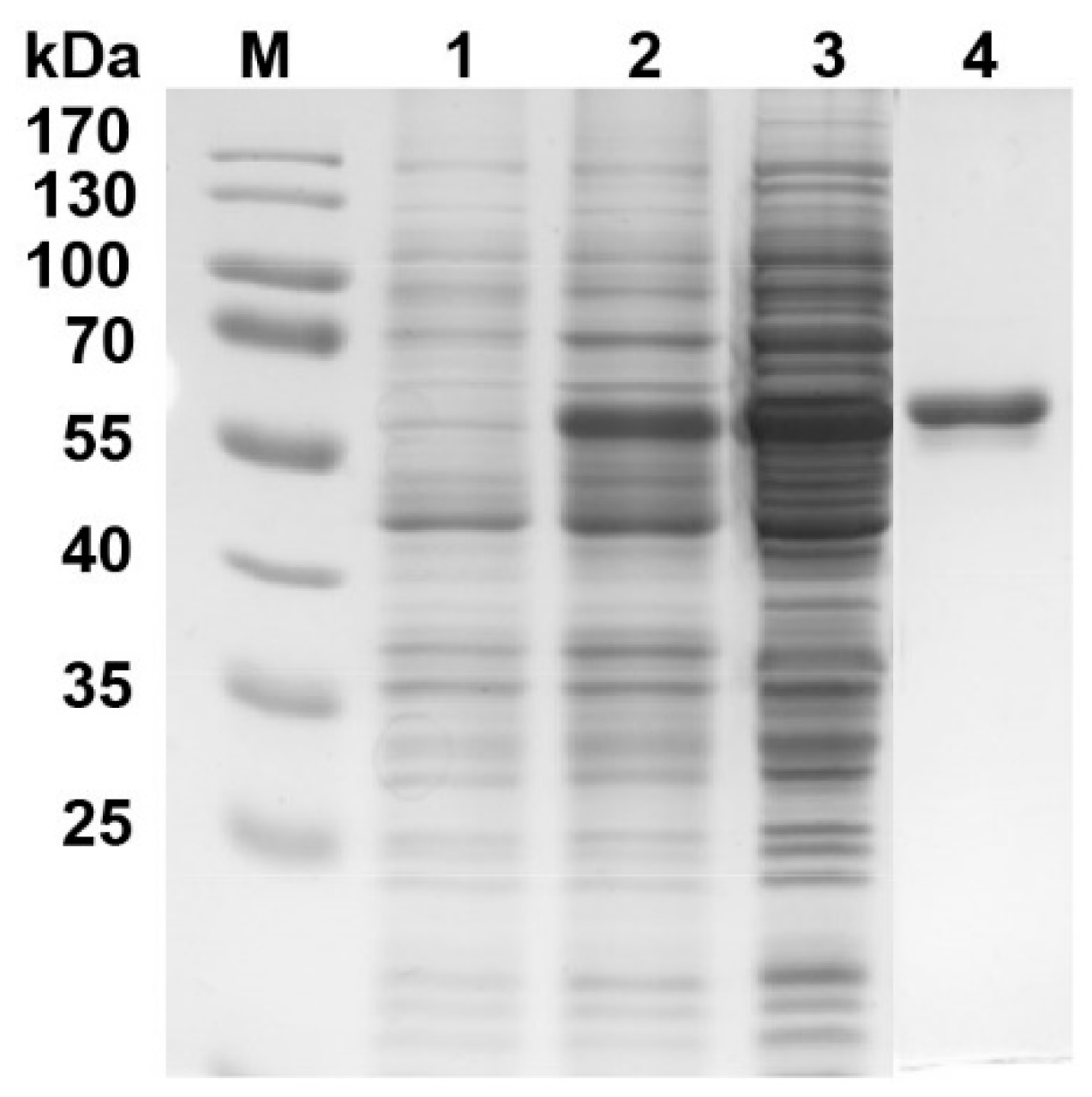
2.5. Heterologous Expression and Purification of Recombinant 4CL3
2.6. Determination of Enzyme Activity
2.7. Data Processing and Analysis
3. Results
3.1. Lignin Content in the Xylem and Phloem of Ramie Stems during the Three Periods
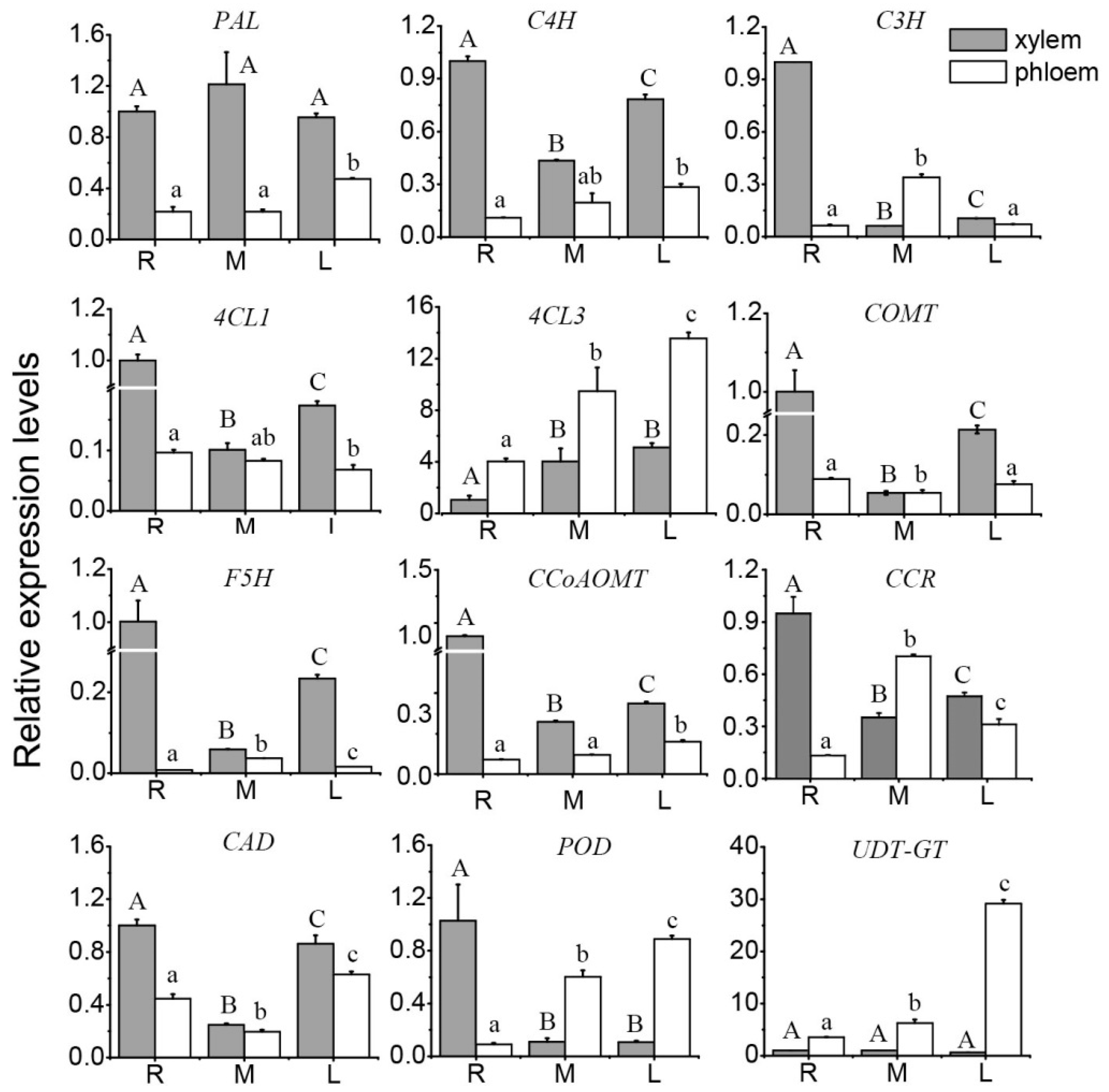
3.2. Expression Profile of Key Genes for Lignin Synthesis in the Stem
3.3. Correlation between the Lignin Content in Stem Segments and the Expression Level of Key Genes
3.4. Enzyme Assay of the Recombinant 4CL3 Protein of Ramie
4. Discussion
4.1. Lignin Formation in the Xylem and Phloem of the Ramie Stem
4.2. Correlation between Lignin Content in the Ramie Stem and the Key Genes for Lignin Synthesis
4.3. Enzyme Assay of the Recombinant 4CL3 Protein of Ramie
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kandimalla, R.; Kalita, S.; Choudhury, B.; Devi, D.; Kalita, D.; Kalita, K.; Dash, S.; Kotoky, J. Fiber from ramie plant (Boehmeria nivea): A novel suture biomaterial. Mater. Sci. Eng. C 2016, 62, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.F.; Chen, J.H.; Mu, B.; Zeng, M.; Zhang, H.; Yu, J.; Zhao, J.; Luan, M.B. Advances in Medicinal Health Protection Studies of Boehmeria Jacq. Spp. Plant Fiber Sci. Chin. 2016, 38, 237–241. [Google Scholar]
- He, H.B.; Bai, C.H.; Zou, K.; Wang, W.Z.; Hu, J.Z.; Tai, Y.L. Study and exploitation of ramie leaf. Feed Res. 2013, 4, 83–87. [Google Scholar]
- Xu, N.; Yang, S.; Wang, D. Development of ramie planting industry to promote the control of soil erosion in the southern slope. Water Res. Dev. Manag. 2012, 10, 47–48. [Google Scholar]
- Xu, H.Y. A review: Chemical modification on ramie fiber in China. J. Henan Inst. Eng. 2012, 24, 20–24. [Google Scholar]
- Whettena, R.; Sederoff, R. Lignin biosynthesis. Plant Cell. 2003, 7, 1001–1013. [Google Scholar] [CrossRef]
- Delwiche, C.F.; Graham, L.E.; Thomson, N. Lignin-like compounds and sporopollenin coleochaete, an algal model for land plant ancestry. Science 1989, 245, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Ligrone, R.; Carafa, A.; Duckett, J.G.; Renzaglia, K.S.; Ruel, K. Immunocytochemical detection of lignin-related epitopes in cell walls in bryophytes and the charalean alga Nitella. Plant Syst. Evol. 2007, 270, 257–272. [Google Scholar] [CrossRef]
- Sorensen, I.; Pettolino, F.A.; Bacic, A.; Ralph, J.; Lu, F.; O’Neill, M.A.; Fei, Z.; Rose, J.K.; Domozych, D.S.; Willats, W.G. The charophycean green algae provide insights into the early origins of plant cell walls. Plant J. 2011, 68, 201–211. [Google Scholar] [CrossRef]
- de Vries, J.; de Vries, S.; Slamovits, C.H.; Rose, L.E.; Archibald, J.M. How Embryophytic is the Biosynthesis of Phenylpropanoids and their Derivatives in Streptophyte Algae? Plant Cell Physiol. 2017, 58, 934–945. [Google Scholar] [CrossRef]
- Kroken, S.B.; Graham, L.E.; Cook, M.E. Occurrence and evolutionary significance of resistant cell walls in charophytes and bryophytes. Am. J. Bot. 1996, 83, 1241–1254. [Google Scholar] [CrossRef]
- Oliva, J.; Rommel, S.; Fossdal, C.G.; Hietala, A.M.; Nemesio-Gorriz, M.; Solheim, H.; Elfstrand, M. Transcriptional responses of Norway spruce (Picea abies) inner sapwood against Heterobasidion parviporum. Tree Physiol. 2015, 35, 1007–1015. [Google Scholar] [CrossRef]
- Danielsson, M.; Lundén, K.; Elfstrand, M.; Hu, J.; Zhao, T.; Arnerup, J.; Ihrmark, K.; Swedjemark, G.; Borg-Karlson, A.-K.; Stenlid, J. Chemical and transcriptional responses of Norway spruce genotypes with different susceptibility to Heterobasidion spp. infection. BMC Plant Biol. 2011, 11, 154–169. [Google Scholar] [CrossRef]
- Fan, L. Bioinformatic and Functional Analysis of Mulberry Genes Involved in Lignin Biosynthesis; Southwest University: Chongqing, China, 2013. (In Chinese) [Google Scholar]
- Wang, J.; Chen, X.B.; Gao, Y.; Zhang, Y.H.; Long, S.H.; Deng, X.; He, D.F.; Wang, Y.F. Expression of Critical Lignin Metabolism Genes in Flax (Linum usitatissimum). Acta Agron. Sin. 2009, 35, 1468–1473. [Google Scholar] [CrossRef]
- Hu, D.; Liu, X.B.; Wang, C.; Yang, H.; Li, H.X.; Ruan, R.W.; Yuan, X.H.; Yi, Z.L. Expression Analysis of Key Enzyme Genes in Lignin Synthesis of Culm Among Different Lodging Resistances of Common Buckwheat (Fagopyrum esculentum Moench). Sci. Agric. Sin. 2015, 48, 1864–1872. [Google Scholar]
- Huang, J.H. Lodging Resistant Traits and Lignin Related Gene Analysis in B.napus under Drought Stress; Southwest University: Chongqing, China, 2013. (In Chinese) [Google Scholar]
- Deng, J.; Liu, F.; Guo, Q.Q.; Chen, J.R.; Zhang, X.W. Construction of the RNAi vector of ramie CCoAOMT gene and its transformation in tobacco. J. Hunan Agric. Uni. (Nat. Sci.) 2008, 2, 132–135. [Google Scholar]
- Huang, C.Q.; Guo, A.P.; Zhang, X.Y.; Liu, G.D. cDNA Cloning and Sequence Analysis of COMT Gene from Ramie. Chin. Agric. Sci. 2008, 5, 386–391. [Google Scholar]
- Tang, Y.H.; Liu, F.; Mao, K.Q.; Xing, H.C.; Chen, J.R.; Guo, Q.Q. Cloning and characterization of the key 4-coumarate CoA ligase genes in Boehmeria nivea. S. Afr. J. Bot. 2018, 116, 123–130. [Google Scholar] [CrossRef]
- Tang, Y.H.; Chen, J.R.; Liu, F.; Yuan, Y.M.; Guo, Q.Q.; Chang, H.T. cDNA Cloning and Analysis of Cinnamoyl-CoA Reductase Gene from Boehmeria nivea. Acta Agron. Sin. 2015, 41, 1324. [Google Scholar] [CrossRef]
- Zhu, W.Y.; Rong, C.J.; Yan, P.; Weng, Z.X.; Quan, G.Q.; Yan, Z. cDNA cloning and expression analysis of a lignin synthetase CAD gene in ramie (Boehmeria nivea (L.) Gaud.). J. Hunan Agric. Univ. (Nat. Sci.) 2014, 40, 483–488. [Google Scholar]
- Liu, F.; Chen, J.R.; Tang, Y.H.; Chang, H.T.; Yuan, Y.M.; Guo, Q.Q. Isolation and characterization of cinnamate 4-hydroxylase gene from cultivated ramie (Boehmeria nivea). Biotechnol. Biotechnol. Equip. 2018, 32, 324–331. [Google Scholar] [CrossRef]
- Li, X.Q.; Peng, Z.H.; Gao, Z.M.; Hu, T. The Effects of Depressing Expression of COMT on Lignin Synthesis of Transgenic Tobacco. Mol. Plant Breed. 2013, 10, 689–692. [Google Scholar]
- Li, Y.; Kim, J.I.; Pysh, L.; Chapple, C. Four Isoforms of Arabidopsis 4-Coumarate:CoA Ligase Have Overlapping yet Distinct Roles in Phenylpropanoid Metabolism. Plant Physiol. 2015, 169, 2409–2421. [Google Scholar]
- Xiu, Z.M.; Peng, X.P.; Chen, J.H. Effect of down regulation of C3H and HCT on the spatial morphology of cell wall in poplar lignin synthesis. J. Shandong For. Sci. Technol. 2017, 47, 37–40. [Google Scholar]
- Chen, J.R.; Liu, F.; Tang, Y.H.; Yuan, Y.M.; Guo, Q.Q. Transcriptome Sequencing and Profiling of Expressed Genes in Phloem and Xylem of Ramie (Boehmeria nivea L. Gaud). PLoS ONE 2014, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.G.; Shi, C.Y.; Yin, Y.P.; Wang, Z.L.; Shi, Y.H.; Peng, D.L.; Ni, Y.L.; Cai, T. Relationship between Lignin Metabolism and Lodging Resistance in Wheat. Acta Agron. Sin. 2011, 37, 1616–1622. [Google Scholar] [CrossRef]
- Ma, X.; Tan, X.L.; Xing, H.C.; Liao, S.B. Establishment of real- time Quantitative PCR system for ramie. Plant Fiber Sci. China 2017, 39, 111–119. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Angelini, L.G.; Tavarini, S. Ramie [Boehmeria nivea (L.) Gaud.] as a potential new fibre crop for the Mediterranean region: Growth, crop yield and fibre quality in a long-term field experiment in Central Italy. Ind. Crops Prod. 2013, 51, 138–144. [Google Scholar] [CrossRef]
- Deng, J.L.; Zhang, G.Y.; Huang, S.Q.; Li, D.F. Isolation, overexpression vector construction and Transformation of Arabidopsis thaliana of peroxidase gene of Hibiscus cannabinus. Gen. Appl. Biol. 2017, 36, 1570–1574. [Google Scholar]
- Silber, M.V.; Meimberg, H.; Ebel, J. Identification of a 4-coumarate: CoA ligase gene family in the moss, Physcomitrella patens. Phytochemistry 2008, 69, 2449–2456. [Google Scholar] [CrossRef]
- Gao, S.; Yu, H.N.; Xu, R.X.; Cheng, A.X.; Lou, H.X. Cloning and functional characterization of a 4-coumarate CoA ligase from liverwort Plagiochasma appendiculatum. Phytochemistry 2015, 111, 48–58. [Google Scholar] [CrossRef]
- Schneider, K.; Hovel, K.; Witzel, K.; Hamberger, B.; Schomburg, D.; Kombrink, E.; Stuible, H.P. The substrate specificity-determining amino acid code of 4-coumarate: CoA ligase. Proc. Natl. Acad. Sci. USA 2003, 8, 8601–8606. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | PCR Amplicon Size (bp) | Tm (°C) |
|---|---|---|---|
| PAL | P1:GAGCAGCACAACCAAGACG P2:CCGTGGTCAGCACCTTCTT | 202 | 49.2 |
| C4H | P1:CATTCCTGCGAGGCTACTTG P2:CTGCTGAGCGTCAAGAATGTG | 164 | 49.2 |
| C3H | P1:GAGGTAAACGAGCAAGGACAAG P2:GAACAACCGCAACCAAGGA | 105 | 58.9 |
| 4CL1 | P1:AGAGGTCCCTGTTGCCTTTGTTG P2:TTTTGCCTGAAGGGGATTTAGTGA | 152 | 60.0 |
| 4CL3 | P1:ACCGCCGTGCTGCTGTATT P2:CAGCCCGAAGACGTGAAACA | 171 | 61.8 |
| COMT | P1:GACCAAGAACAACGACGGC P2:AATCTCGGATCTGTCCCATGG | 171 | 60.0 |
| F5H | P1:GTGCGAAGGTGAATGAGAGC P2:TGCACCTTCTTTAGATCCTCTG | 175 | 60.0 |
| CCoAOMT | P1:GGATGCTGACAAGGACAACT P2:GAGCCATTCCATAGGGTGT | 102 | 49.2 |
| CCR | P1:CCCGATGTTGTGGTTGATGAGTC P2:ACCAAATCCACGCCTTTCTCC | 137 | 60.0 |
| CAD | P1:GGCATGAGATTGTTGGAATTG P2:CATGGTTGAAGGTGTAAACGG | 171 | 60.0 |
| POD | P1:CCCAGGATAGCCATCAACAT P2:TTCCGACTCGTTTTCACCC | 230 | 58.0 |
| UDP-GT | P1:GGAGCAACAAGTCAACGCCT P2:CTCTTTTACCCTTTTCCTTATCGC | 199 | 60.0 |
| Actin I | P1:CGTTGAACCCTAAGGC P2:ATCCAGCACGATACCAG | 137 | reference gene |
| Tissue | Fast-Growing Period | Maturation Period | Late Maturation Period |
|---|---|---|---|
| Xylem | 46.09 ± 3.091 cC | 62.19 ± 1.013 bB | 71.46 ± 3.669 aA |
| Phloem | 8.04 ± 0.771 eD | 11.74 ± 0.485 deD | 15.95 ± 0.636 dD |
| Variation Source | SS | DF | MS | F-Value |
|---|---|---|---|---|
| Tissue | 10,369.440 | 1.000 | 10,369.440 | 368.635 ** |
| Periods | 840.654 | 2.000 | 420.327 | 14.943 ** |
| Error | 393.810 | 14.000 | 28.129 |
| Periods | PAL | C4H | C3H | 4CL1 | 4CL3 | COMT | F5H | CCoAOMT | CCR | CAD | POD | UDP-GT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fast-growing period | 0.983 ** | 0.981 ** | 0.986 ** | 0.981 ** | −0.961 ** | 0.981 ** | 0.978 ** | 0.987 ** | 0.999 ** | 0.994 ** | 0.961 ** | −0.987 ** |
| Maturation period | 0.953 * | 0.972 ** | −0.998 ** | 0.945 ** | −0.916 * | 0.025 | 0.985 ** | 0.997 ** | −0.996 ** | 0.915 * | −0.990 ** | −0.989 ** |
| Late maturation period | 0.998 ** | 0.997 ** | 0.934 ** | 0.992 ** | −0.991 ** | 0.985 ** | 0.983 ** | 0.988 ** | 0.969 ** | 0.944 ** | −0.988 ** | −0.992 ** |
| Substrate | Km (μM) | Vmax (nkat mg−1 Protein) | Kcat (s−1) | Kcat/Km (s−1·mM−1) |
|---|---|---|---|---|
| p-Coumaric acid | 498.491 | 1.607 | 0.097 | 0.194 |
| Cinnamic acid | 449.926 | 4.842 | 0.291 | 0.248 |
| Caffeic acid | 194.426 | 0.746 | 0.045 | 0.231 |
| Ferulic acid | ND | ND | ND | ND |
| Sinapic acid | ND | ND | ND | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Liu, F.; Xing, H.; Mao, K.; Chen, G.; Guo, Q.; Chen, J. Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea). Genes 2019, 10, 389. https://doi.org/10.3390/genes10050389
Tang Y, Liu F, Xing H, Mao K, Chen G, Guo Q, Chen J. Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea). Genes. 2019; 10(5):389. https://doi.org/10.3390/genes10050389
Chicago/Turabian StyleTang, Yinghong, Fang Liu, Hucheng Xing, Kaiquan Mao, Guo Chen, Qingquan Guo, and Jianrong Chen. 2019. "Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea)" Genes 10, no. 5: 389. https://doi.org/10.3390/genes10050389