
Modulation of Extracellular Matrix Composition and Chronic Inflammation with Pirfenidone Promotes Scar Reduction in Retinal Wound Repair

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Laser-Induced Retinal Damage
2.2. Pharmacological Treatment
2.3. Histology
2.4. Immunohistochemistry
2.5. Western Blot Analysis
2.6. RNA Extraction and RT-qPCR
2.7. RT2 Profiler PCR Array
2.8. Statistical Analysis
3. Results
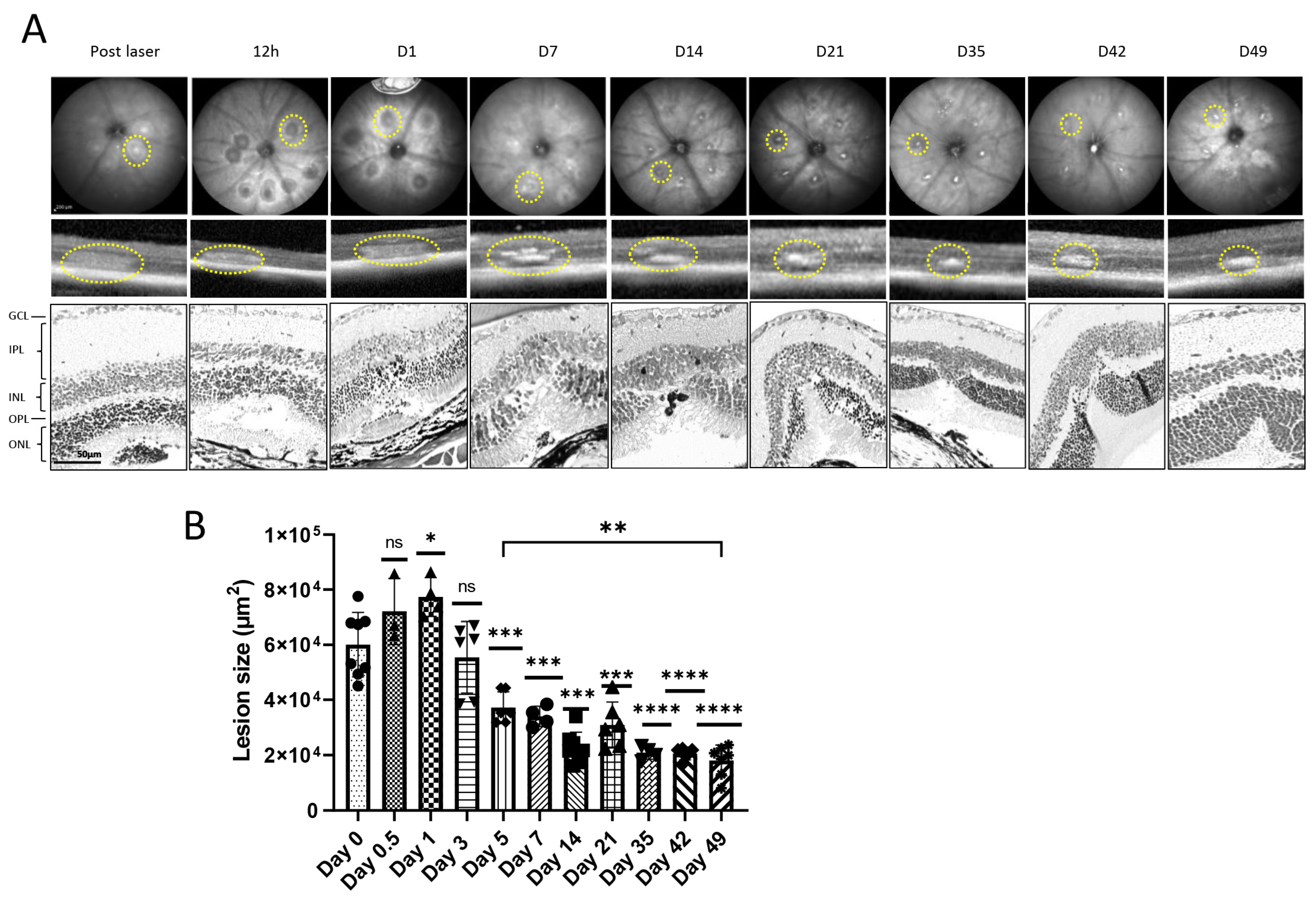
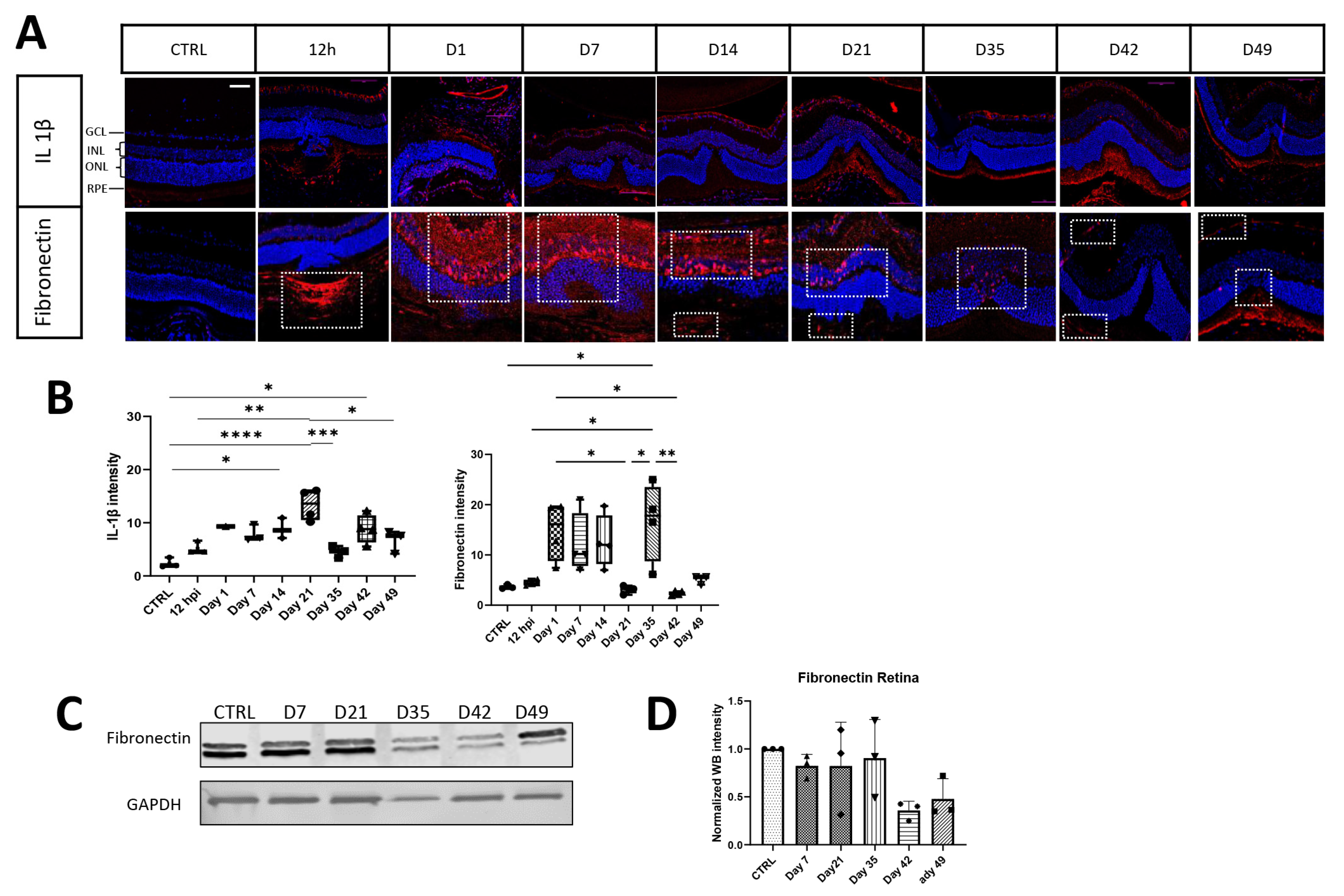
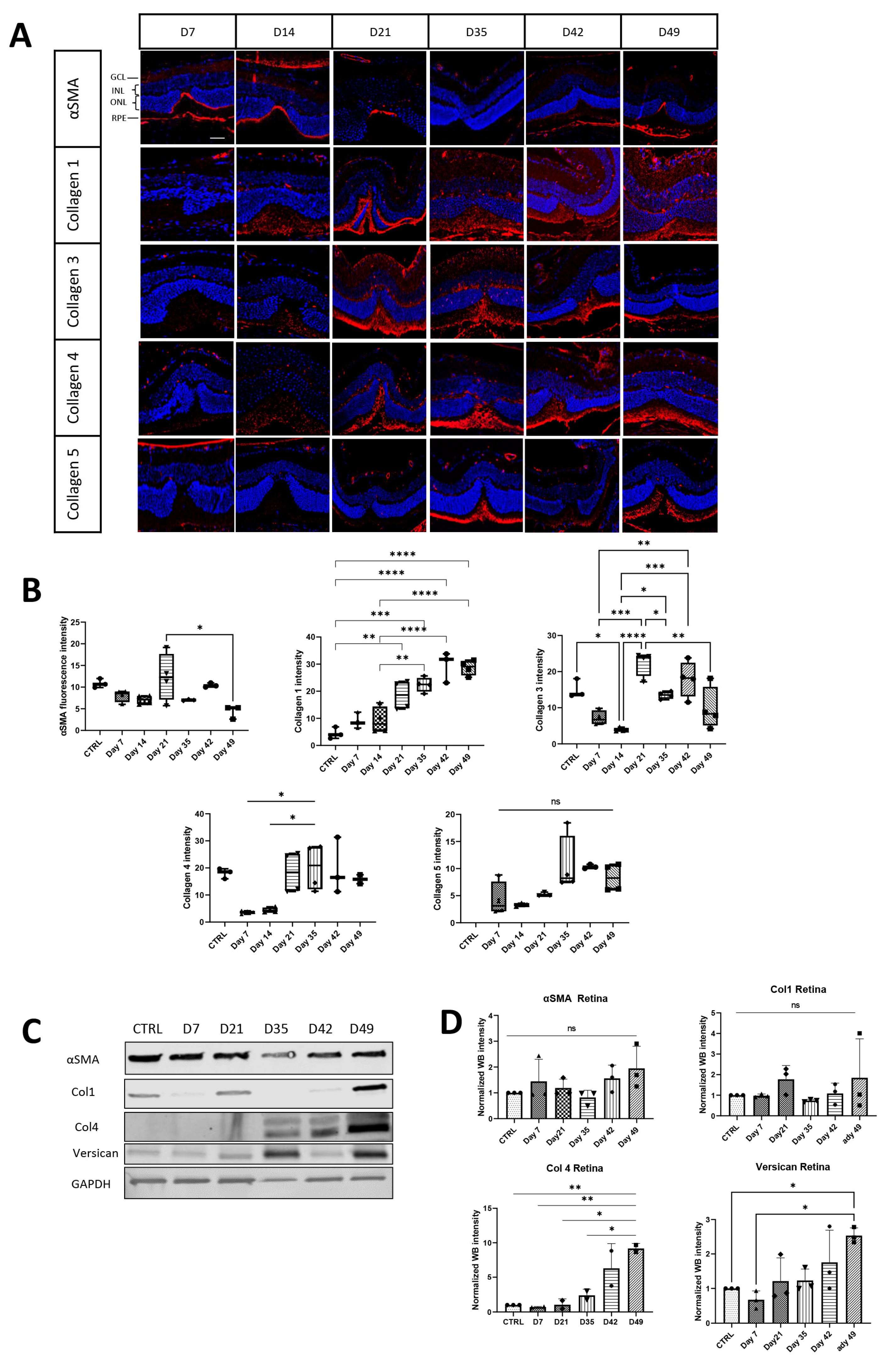
3.1. Extracellular Matrix and Fibrosis Development in Laser-Damaged Retina over Time
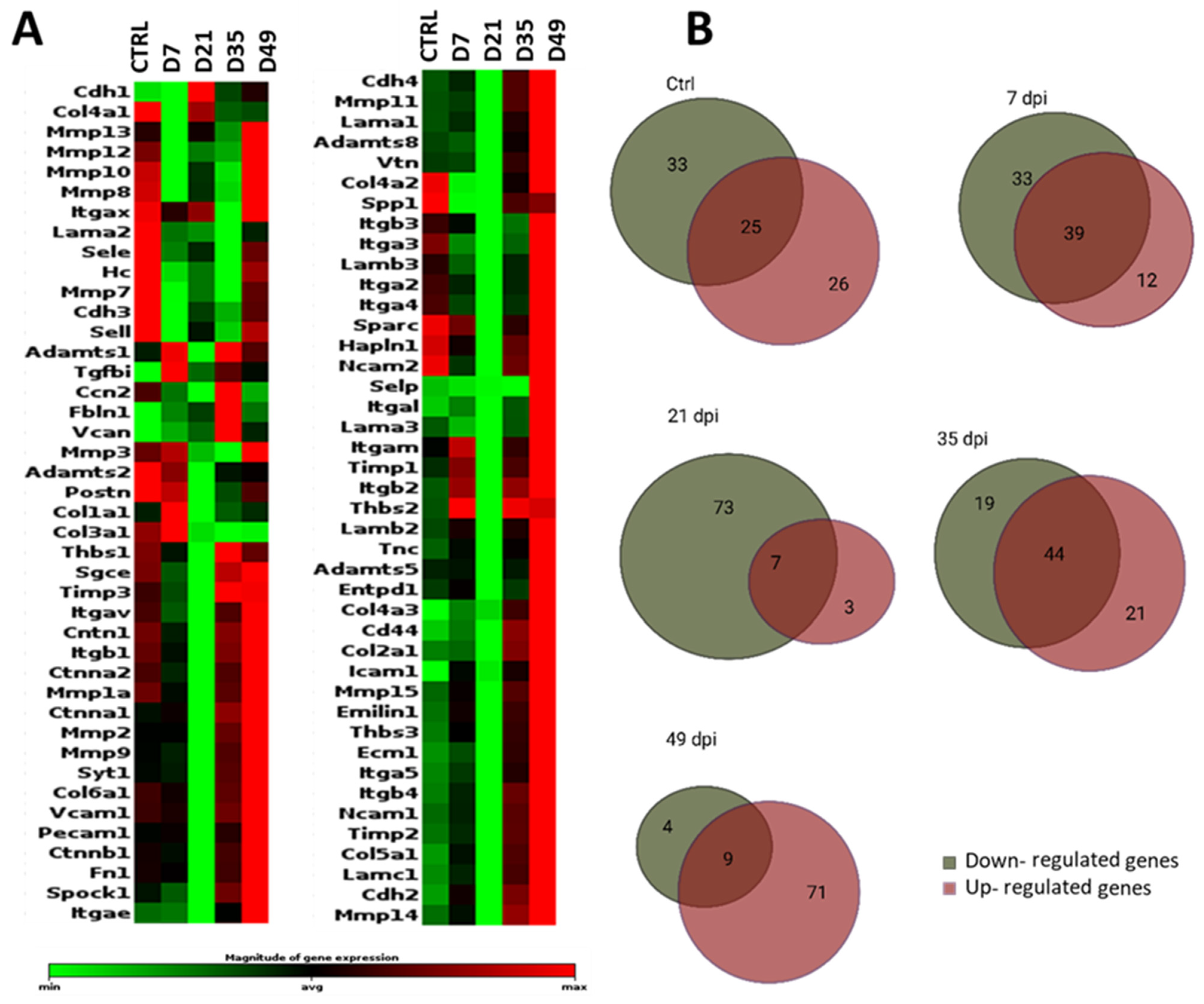
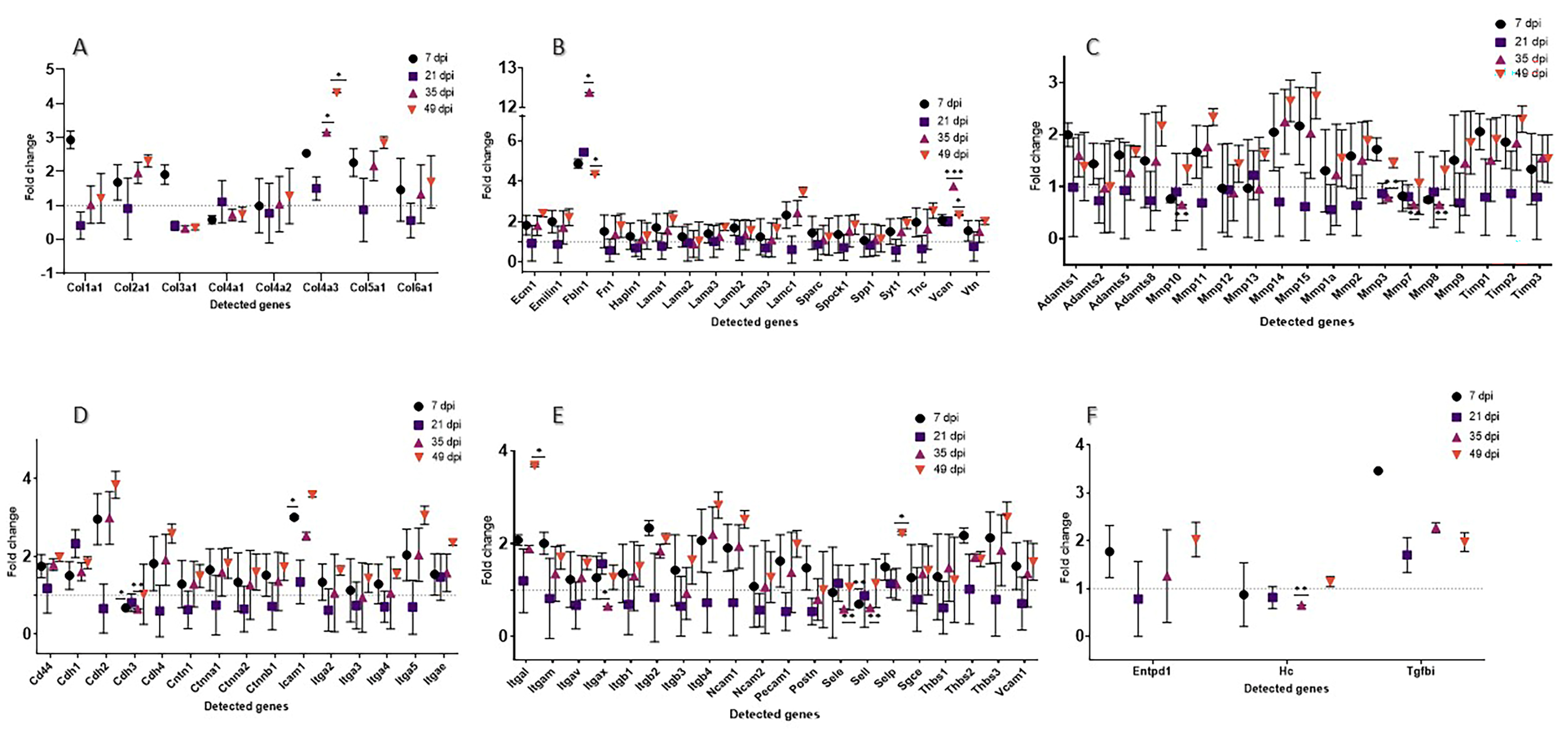
3.2. Extracellular Matrix-Related Genes Show Strong Dynamics during Tissue Replacement
3.2.1. General Comparison
3.2.2. Extracellular Matrix Components
3.2.3. Transmembrane Molecules and Adhesion Molecules
3.2.4. Proteases and Inhibitors
3.2.5. Other ECM Proteins
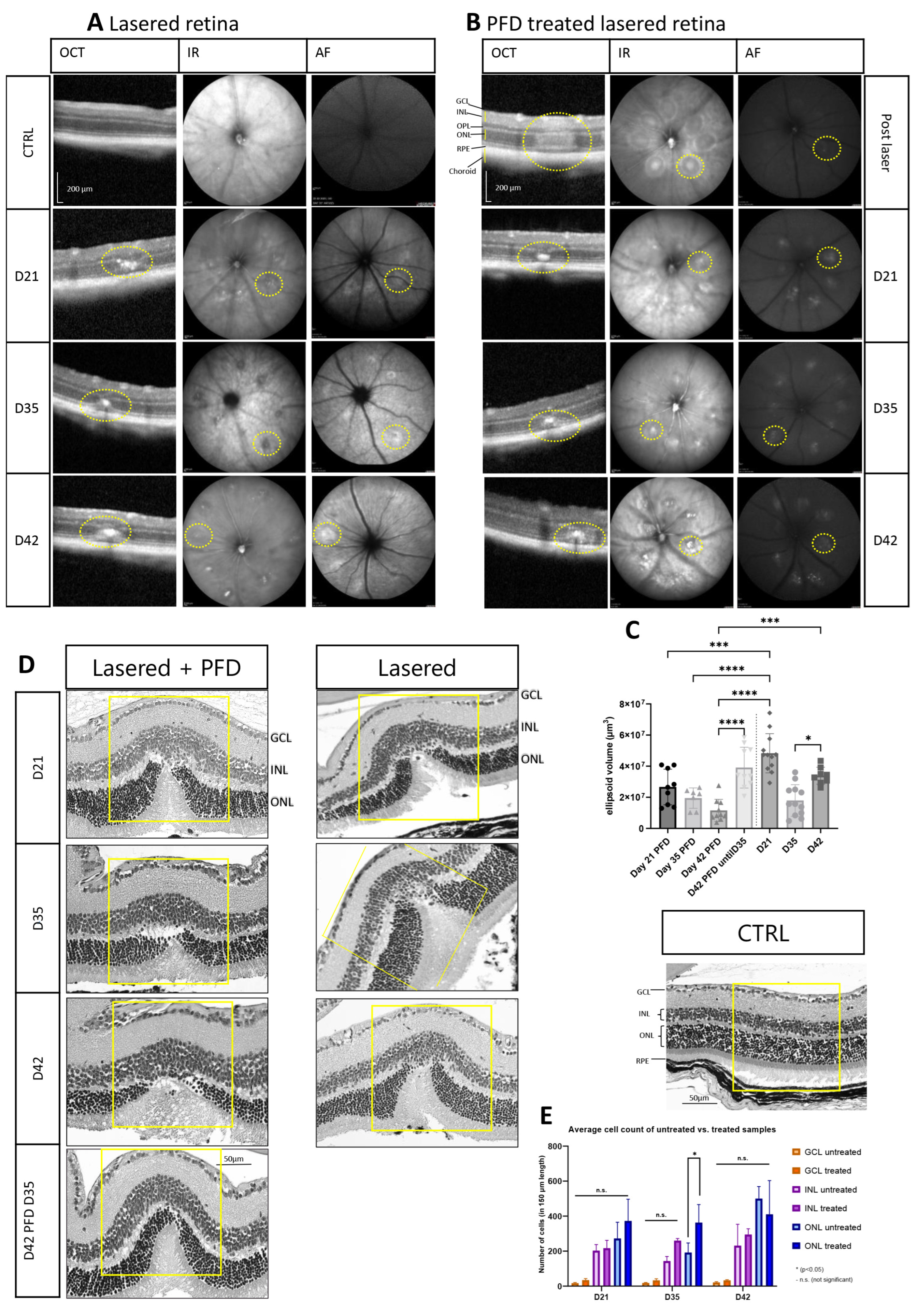
3.3. Extracellular Matrix and Fibrosis Development after Treatment with Pirfenidone
3.3.1. Treatment with Pirfenidone Alters the Cell Count in the Outer Nuclear Layer and Results in Reduced Scar Formation
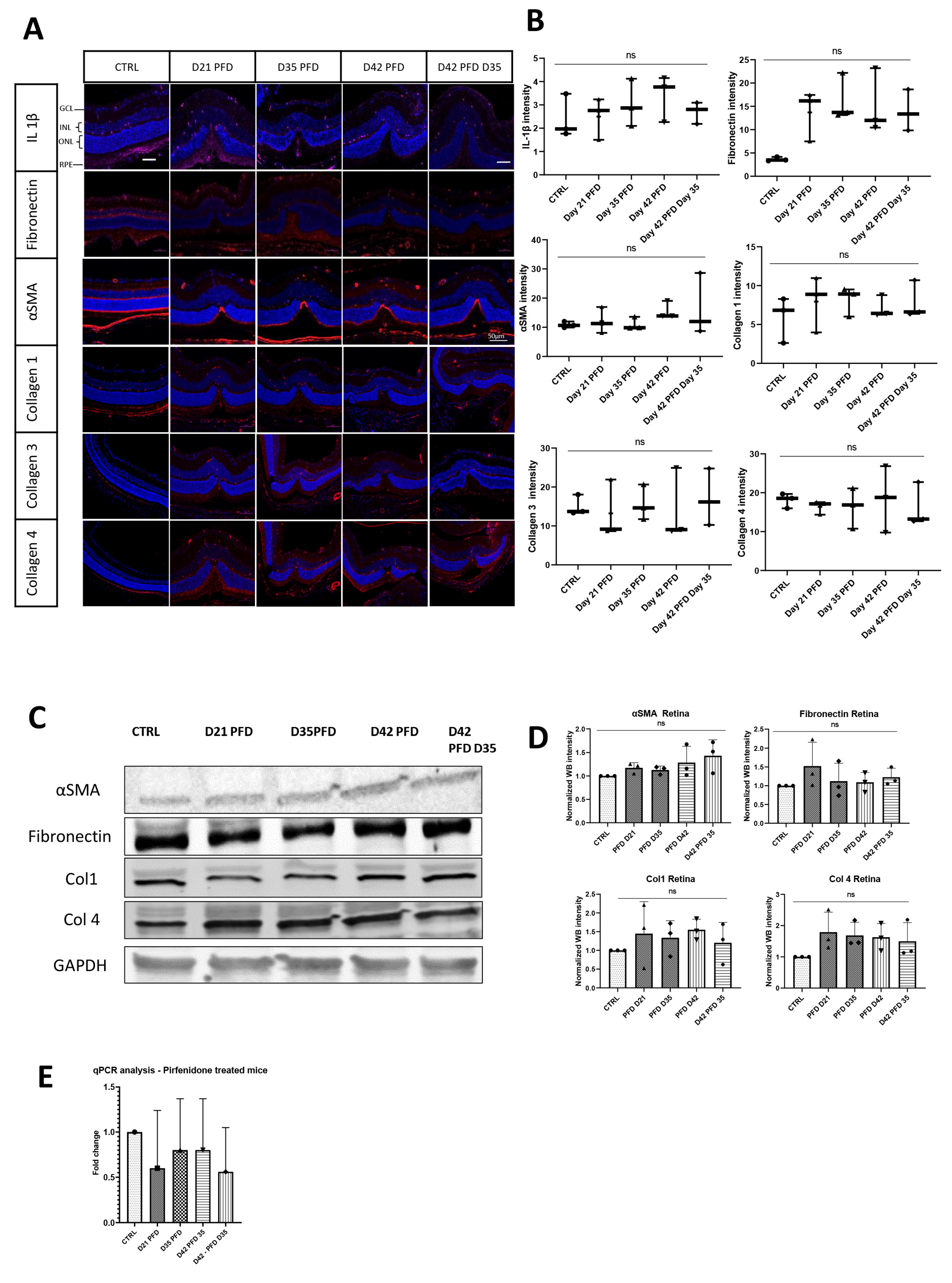
3.3.2. Pirfenidone Prevents Changes in Collagen Expression during Fibrotic Development
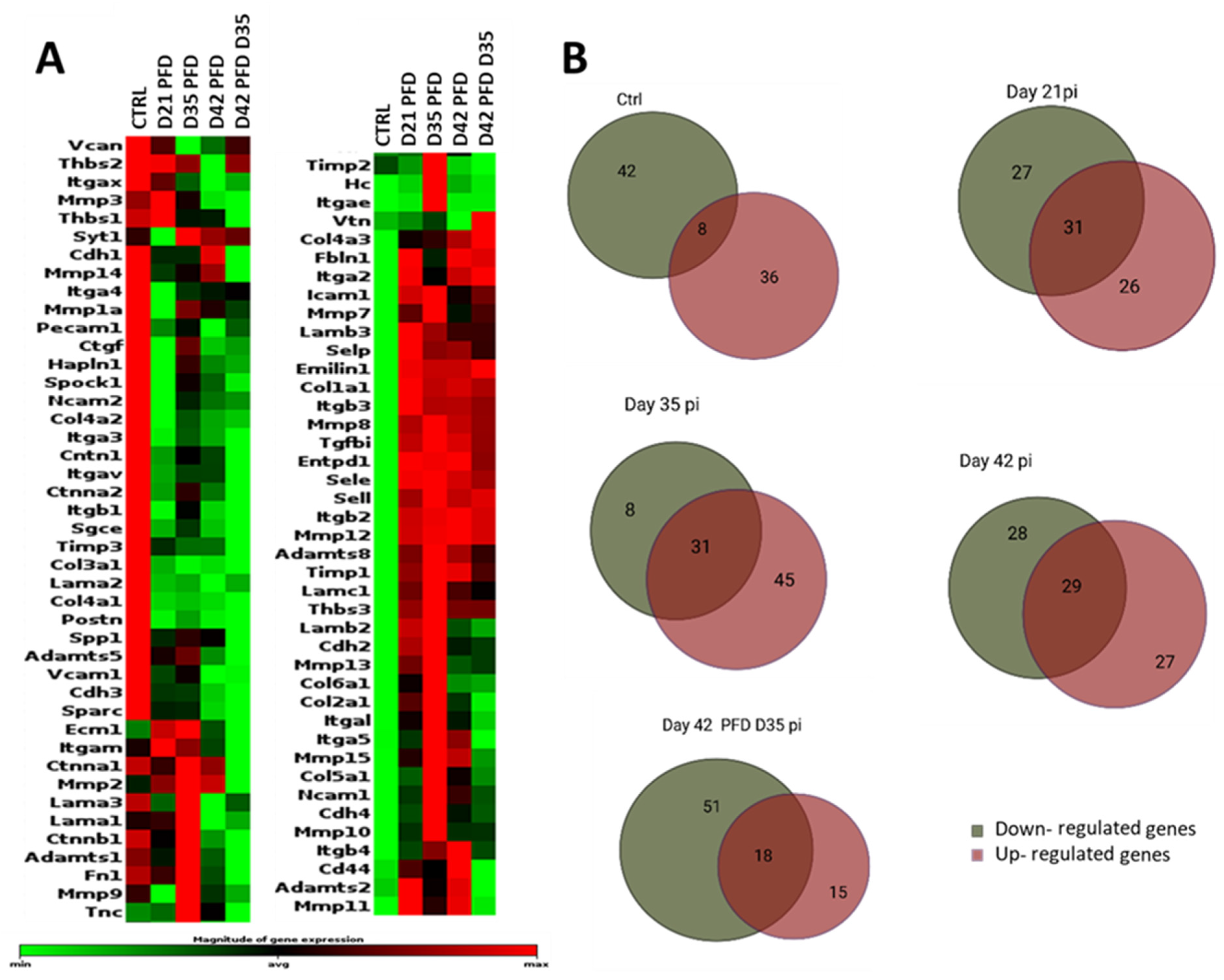
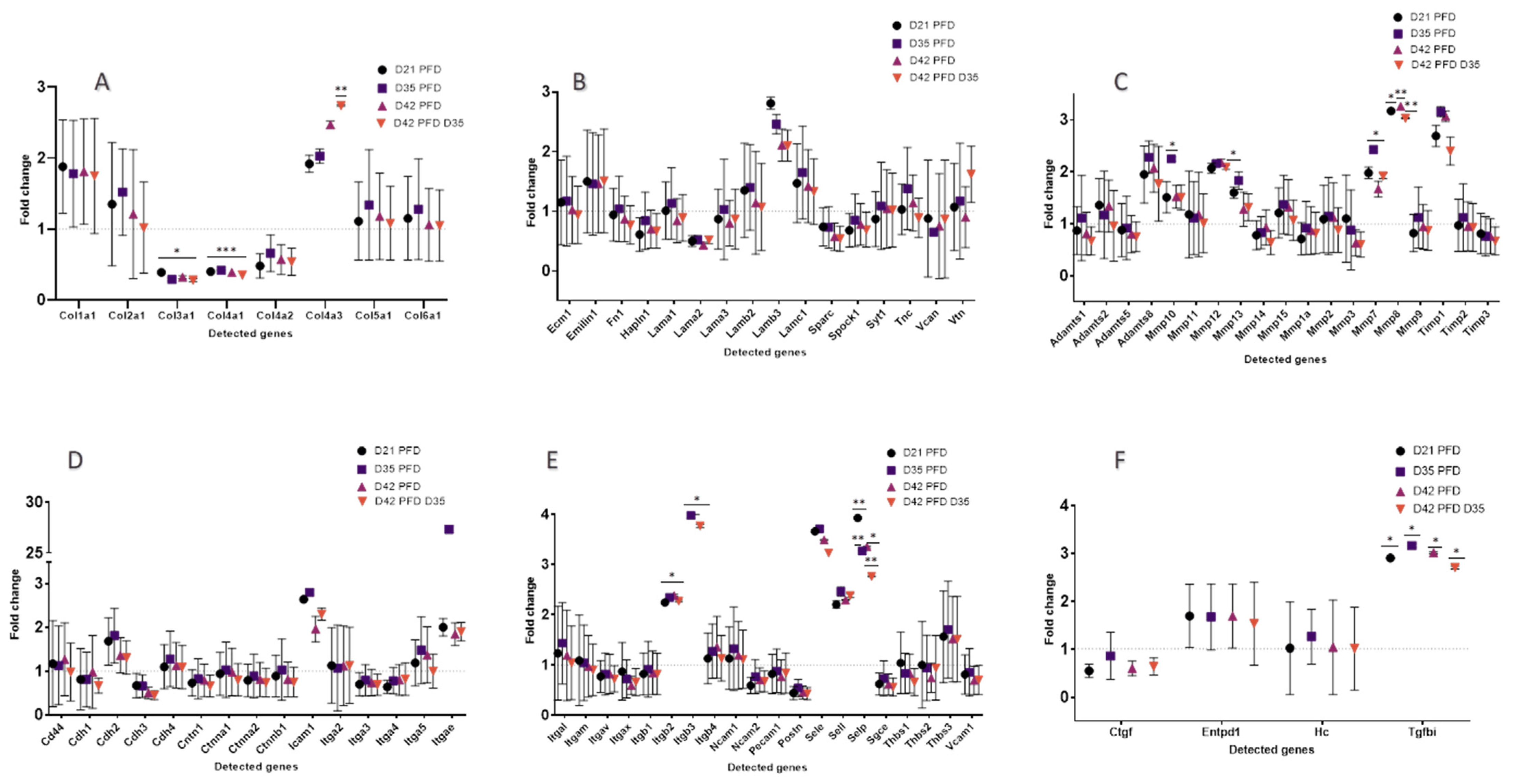
3.3.3. Screening of ECM and Adhesion Molecules Involved in the ECM Formation during Damage Response after PFD Treatment
3.3.4. General Comparisons
3.3.5. Extracellular Matrix Components
3.3.6. Transmembrane Molecules and Adhesion Molecules
3.3.7. Proteases and Inhibitors
3.3.8. Other ECM Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishikawa, K.; Kannan, R.; Hinton, D.R. Molecular mechanisms of subretinal fibrosis in age-related macular degeneration. Exp. Eye Res. 2016, 142, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Gabbiani, G. The myofibroblast in wound healing and fibrocontractive diseases. J. Pathol. 2003, 200, 500–503. [Google Scholar] [CrossRef]
- Van Leeuwen, E.M.; Emri, E.; Merle, B.M.J.; Colijn, J.M.; Kersten, E.; Cougnard-Gregoire, A.; Dammeier, S.; Meester-Smoor, M.; Pool, F.M.; de Jong, E.K.; et al. A new perspective on lipid research in age-related macular degeneration. Prog. Retin Eye Res. 2018, 67, 56–86. [Google Scholar] [CrossRef] [PubMed]
- Litwińska, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Machaliński, B.; Machalińska, A. The Interplay Between Systemic Inflammatory Factors and MicroRNAs in Age-Related Macular Degeneration. Front. Aging Neurosci. 2019, 11, 286. [Google Scholar] [CrossRef] [PubMed]
- Wooff, Y.; Man, S.M.; Aggio-Bruce, R.; Natoli, R.; Fernando, N. IL-1 Family Members Mediate Cell Death, Inflammation and Angiogenesis in Retinal Degenerative Diseases. Front. Immunol. 2019, 10, 1618. [Google Scholar] [CrossRef] [PubMed]
- Natoli, R.; Fernando, N.; Madigan, M.; Chu-Tan, J.A.; Valter, K.; Provis, J.; Rutar, M. Microglia-derived IL-1β promotes chemokine expression by Müller cells and RPE in focal retinal degeneration. Mol. Neurodegener. 2017, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Spindler, J.; Zandi, S.; Pfister, I.B.; Gerhardt, C.; Garweg, J.G. Cytokine profiles in the aqueous humor and serum of patients with dry and treated wet age-related macular degeneration. PLoS ONE 2018, 13, e0203337. [Google Scholar] [CrossRef]
- Miller, J.W. Beyond VEGF-The Weisenfeld Lecture. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6911–6918. [Google Scholar] [CrossRef]
- Viviani, B.; Corsini, E.; Binaglia, M.; Galli, C.L.; Marinovich, M. Reactive oxygen species generated by glia are responsible for neuron death induced by human immunodeficiency virus-glycoprotein 120 in vitro. Neuroscience 2001, 107, 51–58. [Google Scholar] [CrossRef]
- Viviani, B.; Bartesaghi, S.; Gardoni, F.; Vezzani, A.; Behrens, M.M.; Bartfai, T.; Binaglia, M.; Corsini, E.; Di Luca, M.; Galli, C.L.; et al. Interleukin-1β Enhances NMDA Receptor-Mediated Intracellular Calcium Increase through Activation of the Src Family of Kinases. J. Neurosci. 2003, 23, 8692–8700. [Google Scholar] [CrossRef]
- Wang, H.; Han, X.; Wittchen, E.S.; Hartnett, M.E. TNF-α mediates choroidal neovascularization by upregulating VEGF expression in RPE through ROS-dependent β-catenin activation. Mol. Vis. 2016, 22, 116–128. [Google Scholar]
- Alshoubaki, Y.K.; Nayer, B.; Das, S.; Martino, M.M. Modulation of the Activity of Stem and Progenitor Cells by Immune Cells. Stem Cells Transl. Med. 2022, 11, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Olivares-González, L.; Velasco, S.; Campillo, I.; Rodrigo, R. Retinal Inflammation, Cell Death and Inherited Retinal Dystrophies. Int. J. Mol. Sci. 2021, 22, 2096. [Google Scholar] [CrossRef]
- Gao, C.; Cao, X.; Huang, L.; Bao, Y.; Li, T.; Di, Y.; Wu, L.; Song, Y. Pirfenidone Alleviates Choroidal Neovascular Fibrosis through TGF-β/Smad Signaling Pathway. J. Ophthalmol. 2021, 2021, 8846708. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Silverman, S.M.; Zhao, L.; Villasmil, R.; Campos, M.M.; Amaral, J.; Wong, W.T. Absence of TGFβ signaling in retinal microglia induces retinal degeneration and exacerbates choroidal neovascularization. eLife 2019, 8, e42049. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, T.D.; Kelynack, K.J.; Tait, M.G.; Martic, M.; Jones, C.L.; Margolin, S.B.; Becker, G.J. Pirfenidone reduces in vitro rat renal fibroblast activation and mitogenesis. J. Nephrol. 2001, 14, 453–460. [Google Scholar]
- Ruwanpura, S.M.; Thomas, B.J.; Bardin, P.G. Pirfenidone: Molecular Mechanisms and Potential Clinical Applications in Lung Disease. Am. J. Respir. Cell Mol. Biol. 2020, 62, 413–422. [Google Scholar] [CrossRef]
- Lopez-de la Mora, D.A.; Sanchez-Roque, C.; Montoya-Buelna, M.; Sanchez-Enriquez, S.; Lucano-Landeros, S.; Macias-Barragan, J.; Armendariz-Borunda, J. Role and New Insights of Pirfenidone in Fibrotic Diseases. Int. J. Med. Sci. 2015, 12, 840–847. [Google Scholar] [CrossRef]
- Meng, X.-M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Diaz-Palomera, C.D.; Vidal-Paredes, I.A.; Navarro-Partida, J.; Cid-Hernandez, M.; Rosales-Rivera, L.C.; De la Rosa-Bibiano, R.; Monroy-Ramirez, H.C.; Santos, A.; Armendariz-Borunda, J. Topical Pirfenidone-Loaded Liposomes Ophthalmic Formulation Reduces Haze Development after Corneal Alkali Burn in Mice. Pharmaceutics 2022, 14, 316. [Google Scholar] [CrossRef]
- Khanum, B.N.M.K.; Guha, R.; Sur, V.P.; Nandi, S.; Basak, S.K.; Konar, A.; Hazra, S. Pirfenidone inhibits post-traumatic proliferative vitreoretinopathy. Eye 2017, 31, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Talpan, D.; Salla, S.; Seidelmann, N.; Walter, P.; Fuest, M. Antifibrotic Effects of Caffeine, Curcumin and Pirfenidone in Primary Human Keratocytes. Int. J. Mol. Sci. 2023, 24, 1461. [Google Scholar] [CrossRef]
- Bao, Y.; Huang, L.; Huang, X.; Gao, C.; Chen, Y.; Wu, L.; Zhu, S.; Song, Y. Pirfenidone ameliorates the formation of choroidal neovascularization in mice. Mol. Med. Rep. 2020, 21, 2162–2170. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.V.; Balani, P.; Lopez, A.R.; Nobleza, C.M.N.; Siddiqui, M.; Khan, S. A Review of Pirfenidone as an Anti-Fibrotic in Idiopathic Pulmonary Fibrosis and Its Probable Role in Other Diseases. Cureus 2021, 13, e12482. [Google Scholar] [CrossRef] [PubMed]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef]
- Zandi, S.; Li, Y.; Jahnke, l.; Schweri-Olac, A.; Ishikawa, K.; Wada, I.; Nakao, S.; Zinkernagel, M.S.; Enzmann, V. Animal model of subretinal fibrosis without active choroidal neovascularization. Exp. Eye Res. 2023, 229, 109428. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Zhang, C.; Zhang, J.; Luo, X.; Qiu, Q.; Luo, D.; Zhang, J. TGF-β promotes pericyte-myofibroblast transition in subretinal fibrosis through the Smad2/3 and Akt/mTOR pathways. Exp. Mol. Med. 2022, 54, 673–684. [Google Scholar] [CrossRef]
- Shu, D.Y.; Butcher, E.; Saint-Geniez, M. EMT and EndMT: Emerging Roles in Age-Related Macular Degeneration. Int. J. Mol. Sci. 2020, 21, 4271. [Google Scholar] [CrossRef]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef]
- An, D.; Chung-Wah-Cheong, J.; Yu, D.Y.; Balaratnasingam, C. Alpha-Smooth Muscle Actin Expression and Parafoveal Blood Flow Pathways Are Altered in Preclinical Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2022, 63, 8. [Google Scholar] [CrossRef]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in Wound Repair: Molecular and Cellular Mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Damico, F.M.; Gasparin, F.; Scolari, M.R.; Pedral, L.S.; Takahashi, B.S. New approaches and potential treatments for dry age-related macular degeneration. Arq. Bras. Oftalmol. 2012, 75, 71–76. [Google Scholar] [CrossRef]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s double life: Intracellular regulator and extracellular signal. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1008–1022. [Google Scholar] [CrossRef] [PubMed]
- Budatha, M.; Zhang, J.; Schwartz, M.A. Fibronectin-Mediated Inflammatory Signaling Through Integrin α5 in Vascular Remodeling. J. Am. Heart Assoc. 2021, 10, e021160. [Google Scholar] [CrossRef]
- Van Der Rest, M.; Garrone, R. Collagen family of proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Rai, V.; Agrawal, D.K. Regulation of Collagen I and Collagen III in Tissue Injury and Regeneration. Cardiol. Cardiovasc. Med. 2023, 7, 5–16. [Google Scholar] [CrossRef]
- Halfter, W.; Willem, M.; Mayer, U. Basement membrane-dependent survival of retinal ganglion cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1000–1009. [Google Scholar] [CrossRef]
- Tomino, Y.; Suzuki, S.; Azushima, C.; Shou, I.; Iijima, T.; Yagame, M.; Wang, L.N.; Chen, H.C.; Lai, K.N.; Tan, S.Y.; et al. Asian multicenter trials on urinary type IV collagen in patients with diabetic nephropathy. J. Clin. Lab. Anal. 2001, 15, 188–192. [Google Scholar] [CrossRef]
- Urushiyama, H.; Terasaki, Y.; Nagasaka, S.; Terasaki, M.; Kunugi, S.; Nagase, T.; Fukuda, Y.; Shimizu, A. Role of α1 and α2 chains of type IV collagen in early fibrotic lesions of idiopathic interstitial pneumonias and migration of lung fibroblasts. Lab. Investig. 2015, 95, 872–885. [Google Scholar] [CrossRef]
- Ge, Q.; Chen, L.; Jaffar, J.; Argraves, W.S.; Twal, W.O.; Phil Hansbro, P.; Black, J.L.; Burgess, J.K.; Oliver, B. Fibulin1C peptide induces cell attachment and extracellular matrix deposition in lung fibroblasts. Sci. Rep. 2015, 5, 9496. [Google Scholar] [CrossRef]
- Wight, T.N.; Kang, I.; Evanko, S.P.; Harten, I.A.; Chang, M.Y.; Pearce, O.M.T.; Allen, C.E.; Frevert, C.W. Versican-A Critical Extracellular Matrix Regulator of Immunity and Inflammation. Front. Immunol. 2020, 11, 512. [Google Scholar] [CrossRef]
- Song, I.; Dityatev, A. Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 2018, 136, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Hirose, M.; Hirose, J.; Nagakubo, D.; Plaas, A.H.; Miyasaka, M. Binding of a large chondroitin sulfate/dermatan sulfate proteoglycan, versican, to L-selectin, P-selectin, and CD44. J. Biol. Chem. 2000, 275, 35448–35456. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.-S.; Vais, D.; Lapierre, D.; Liang, Y.-Y.; Lee, V.; Yang, B.L.; Yang, B.B. PG-M/versican binds to P-selectin glycoprotein ligand-1 and mediates leukocyte aggregation. J. Cell Sci. 2004, 117, 5887–5895. [Google Scholar] [CrossRef]
- Wu, Y.J.; Pierre, D.P.L.A.; Wu, J.; Yee, A.J.; Yang, B.B. The interaction of versican with its binding partners. Cell Res. 2005, 15, 483–494. [Google Scholar] [CrossRef]
- Hirano, Y.; Sakurai, E.; Matsubara, A.; Ogura, Y. Suppression of ICAM-1 in Retinal and Choroidal Endothelial Cells by Plasmid Small-Interfering RNAs In Vivo. Investig. Ophthalmol. Vis. Sci. 2010, 51, 508–515. [Google Scholar] [CrossRef]
- Granot, E.; Shouval, D.; Ashur, Y. Cell adhesion molecules and hyaluronic acid as markers of inflammation, fibrosis and response to antiviral therapy in chronic hepatitis C patients. Mediat. Inflamm. 2001, 10, 274192. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chung, J.-Y.; Rai, U.; Esumi, N. Cadherins in the retinal pigment epithelium (RPE) revisited: P-cadherin is the highly dominant cadherin expressed in human and mouse RPE in vivo. PLoS ONE 2018, 13, e0191279. [Google Scholar] [CrossRef]
- Polisetti, N.; Sharaf, L.; Martin, G.; Schlunck, G.; Reinhard, T. P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus. Cells 2022, 11, 1975. [Google Scholar] [CrossRef]
- Aimo, A.; Spitaleri, G.; Nieri, D.; Tavanti, L.M.; Meschi, C.; Panichella, G.; Lupón, J.; Pistelli, F.; Carrozzi, L.; Bayes-Genis, A.; et al. Pirfenidone for Idiopathic Pulmonary Fibrosis and Beyond. Card. Fail. Rev. 2022, 8, e12. [Google Scholar] [CrossRef]
- Adler, M.; Mayo, A.; Zhou, X.; Franklin, R.A.; Meizlish, M.L.; Medzhitov, R.; Kallenberger, S.M.; Alon, A. Principles of Cell Circuits for Tissue Repair and Fibrosis. iScience 2020, 23, 100841. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.I.; Lau, L.F. Resolution of organ fibrosis. J. Clin. Investig. 2018, 128, 97–107. [Google Scholar] [CrossRef]
- Rodrigues, R.M.; He, Y.; Hwang, S.; Bertola, A.; Mackowiak, B.; Ahmed, Y.A.; Seo, W.; Ma, J.; Wang, X.; Park, S.H.; et al. E-Selectin-Dependent Inflammation and Lipolysis in Adipose Tissue Exacerbate Steatosis-to-NASH Progression via S100A8/9. Cell Mol. Gastroenterol. Hepatol. 2022, 13, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Liu, Z.; Yao, L.; Mehta-D’souza, P.; McEver, R.P. P-Selectin Expressed by a Human SELP Transgene Is Atherogenic in Apolipoprotein E-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Corona, A.; Blobe, G.C. The role of the extracellular matrix protein TGFBI in cancer. Cell Signal. 2021, 84, 110028. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Jahnke, L.; Zandi, S.; Elhelbawi, A.; Conedera, F.M.; Enzmann, V. Characterization of Macroglia Response during Tissue Repair in a Laser-Induced Model of Retinal Degeneration. Int. J. Mol. Sci. 2023, 24, 9172. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Species | Type | Dilution | Company | Catalog. No. |
|---|---|---|---|---|---|
| αSMA | Rabbit | monoclonal | 1:200 (IHC) 1:1000 (WB) | Novus Biologicals, Centennial, CO, USA | NBP2-67436 |
| GS | Rabbit | monoclonal | 1:200 (IHC) | Abcam, Cambridge, UK | ab16802 |
| Isolectin GS-IB4 | Griffonia simplicifolia | - | 1:500 (IHC) | ThermoFisher Scientific, Reinach, Switzerland | I21412 |
| GFAP | Mouse | monoclonal | 1:1000 (IHC) 1:1000 (WB) | Novus Biologicals | NBP1-05197 |
| Collagen 1 | Rabbit | polyclonal | 1:200 (IHC) 1:1000 (WB) | Novus Biologicals | NB600-408 |
| Collagen 3 | Rabbit | polyclonal | 1:200 (IHC) | Novus Biologicals | NB600-594 |
| Collagen 4 | Rabbit | polyclonal | 1:200 (IHC) 1:1000 (WB) | Novus Biologicals | PA1-28534 |
| Collagen 5 | Rabbit | polyclonal | 1:200 (IHC) | Novus Biologicals | NBP1-68938 |
| Fibronectin | Rabbit | polyclonal | 1:200 (IHC) 1:1000 (WB) | Novus Biologicals | NBP1-91258 |
| GAPDH | Mouse | monoclonal | 1:1000 (WB) | Novus Biologicals | NB300-221 |
| IL-1β | Rabbit | polyclonal | 1:200 (IHC) | Abcam | ab9722 |
| Versican | Rabbit | monoclonal | 1:200 (WB) | Novus Biologicals | NBP2-75706 |
| Ingredients | Concentration |
|---|---|
| Tris-HCl (pH 7.5) | 20 mM |
| NaCl | 150 mM |
| EDTA | 5 mM |
| Na-Pyrophosphate | 5 mM |
| NaH2PO4 (pH 7.6) | 20 mM |
| Na-β-glycerophosphate | 3 mM |
| NaF | 10 mM |
| Gene | Forward 5′-3′ | Reverse 5′-3′ | Annealing Temp. |
|---|---|---|---|
| IL-1β | GCC CAT CCT CTG TGA CTC AT | AGG CCA CAG GTA TTT TGT CG | 58 °C |
| GAPDH | TGC ACC ACC AAC TGC TTA GC | GGC ATG GAC TGT GGT CAT GAG | 58 °C |
| Cell Adhesion Molecules | Transmembrane Receptors | Cd44, Cdh1 (E-cadherin), Cdh2 (N-caderin), Cdh3, Cdh4, Entpd1, Icam1, Itga2, Itga3, Itga4 (Cd49d), Itga5, Itgae, Itgal, Itgam, Itgav, Itgb1, Itgb2, Itgb3, Itgb4, Mmp14, Mmp15, Ncam1, Ncam2, Pecam1, Sele, Sell (Lecam-1), Selp, Sgce, Syt1, Vcam1 |
| Cell–Cell Adhesion | Cdh1 (E-cadherin), Icam1, Vcam1 | |
| Cell–Extracellular Matrix (ECM) Adhesion | Ccn2, Itga2, Itga3, Itga4 (Cd49d), Itga5, Itgae, Itgal, Itgam, Itgav, Itgax, Itgb1, Itgb2, Itgb3, Itgb4, Spp1 | |
| Other Cell Adhesion Molecules | Cntn1, Col5a1, Col6a1, Ctnna1, Ctnna2, Ctnnb1, Emilin1, Fn1, Hapln1, Lama1, Lama2, Lama3, Lamb2, Lamb3, Lamc1, Postn, Tgfbi, Thbs1 (TSP-1), Thbs2, Thbs3, Vcan, Vtn | |
| Extracellular Matrix (ECM) Molecules | Basement Membrane Constituents | Col4a1, Col4a2, Col4a3, Entpd1, Itgb4, Lama1, Lama2, Lama3, Lamb2, Lamb3, Lamc1, Sparc, Timp1, Timp2, Timp3 |
| Collagens and Extracellular Matrix (ECM) Structural Constituents | Col1a1, Col2a1, Col3a1, Col4a1, Col4a2, Col4a3, Col5a1, Col6a1, Hapln1, Lama1 | |
| ECM Proteases | Adamts1, Adamts2, Adamts5, Adamts8, Mmp10, Mmp11, Mmp12, Mmp13, Mmp14, Mmp15, Mmp1a, Mmp2, Mmp3, Mmp7, Mmp8, Mmp9 | |
| ECM Protease Inhibitors | Col4a3, Timp1, Timp2, Timp3 | |
| Other ECM Molecules | Ccn2, Ecm1, Emilin1, Fbln1, Fn1, Hc, Postn, Spock1, Spp1, Tgfbi, Thbs1 (TSP-1), Tnc, Vcan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahnke, L.; Perrenoud, V.; Zandi, S.; Li, Y.; Conedera, F.M.; Enzmann, V. Modulation of Extracellular Matrix Composition and Chronic Inflammation with Pirfenidone Promotes Scar Reduction in Retinal Wound Repair. Cells 2024, 13, 164. https://doi.org/10.3390/cells13020164
Jahnke L, Perrenoud V, Zandi S, Li Y, Conedera FM, Enzmann V. Modulation of Extracellular Matrix Composition and Chronic Inflammation with Pirfenidone Promotes Scar Reduction in Retinal Wound Repair. Cells. 2024; 13(2):164. https://doi.org/10.3390/cells13020164
Chicago/Turabian StyleJahnke, Laura, Virginie Perrenoud, Souska Zandi, Yuebing Li, Federica Maria Conedera, and Volker Enzmann. 2024. "Modulation of Extracellular Matrix Composition and Chronic Inflammation with Pirfenidone Promotes Scar Reduction in Retinal Wound Repair" Cells 13, no. 2: 164. https://doi.org/10.3390/cells13020164