
CREB1 Facilitates GABAergic Neural Differentiation of Human Mesenchymal Stem Cells through BRN2 for Pain Alleviation and Locomotion Recovery after Spinal Cord Injury

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
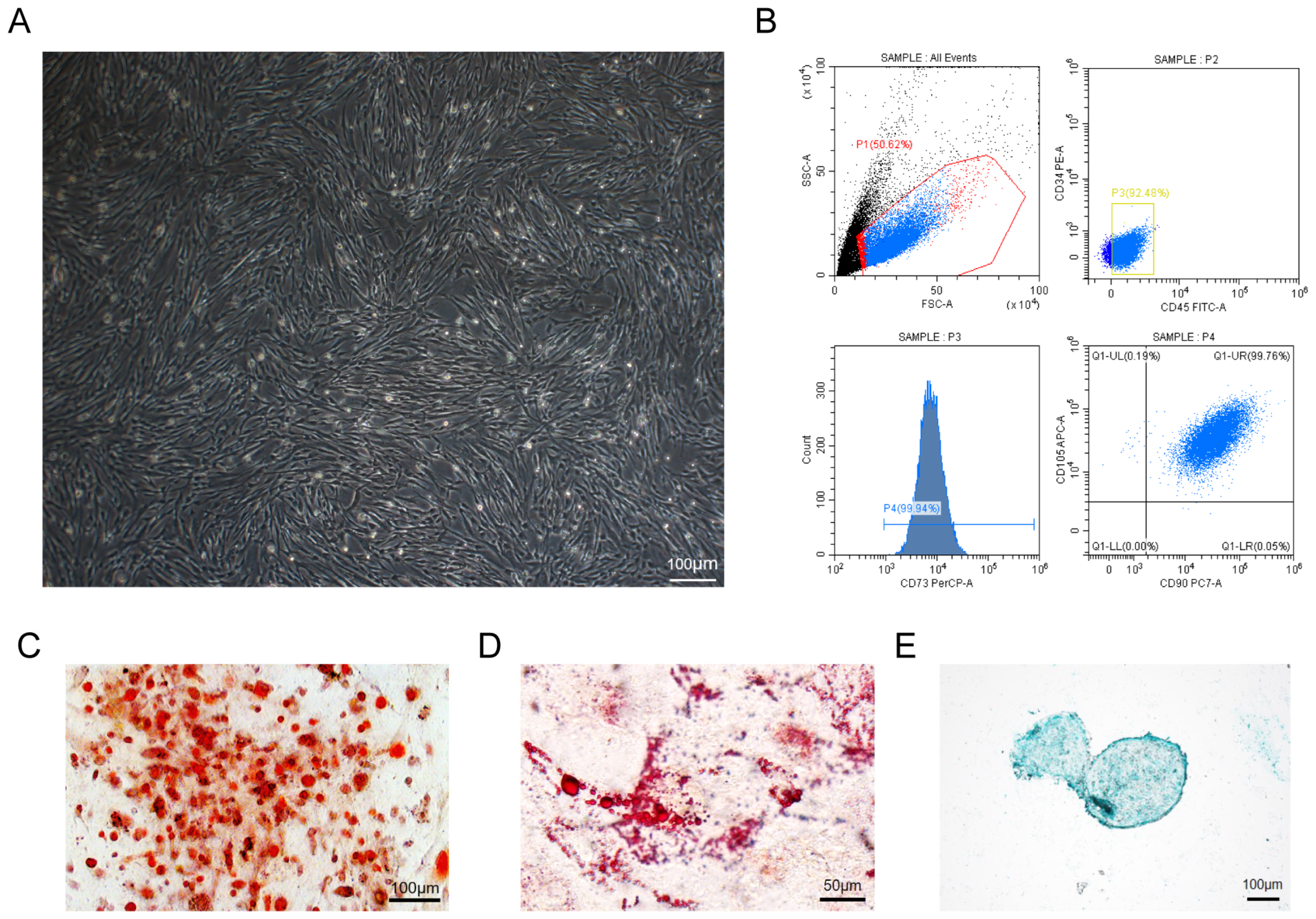
2.1. Acquisition and Characterization of Mesenchymal Stem Cells
2.2. Generation of iPSC-Derived GABA Neurons and iGNs
2.3. The Lentivirus Package and Cell Transfection
2.4. EdU Assay
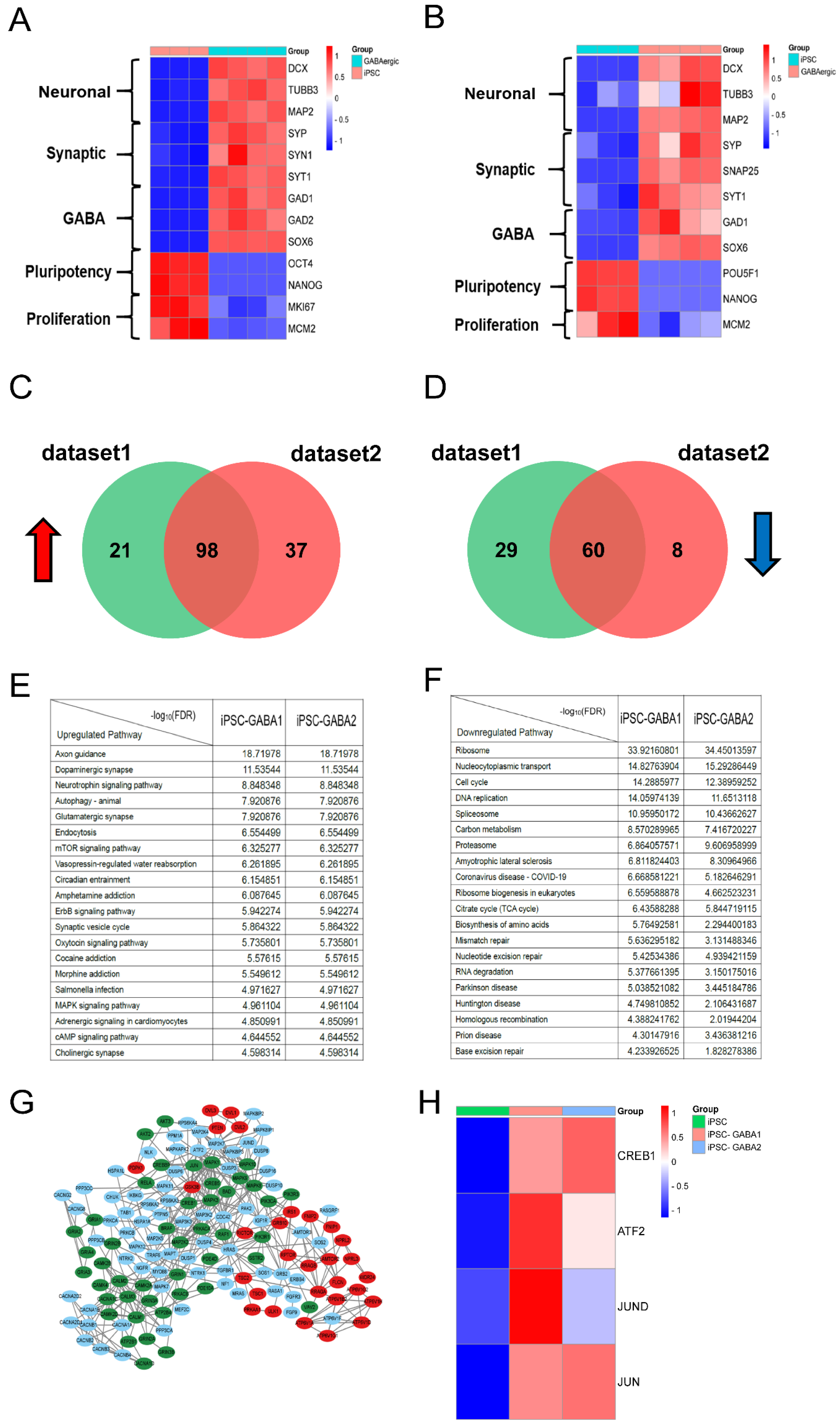
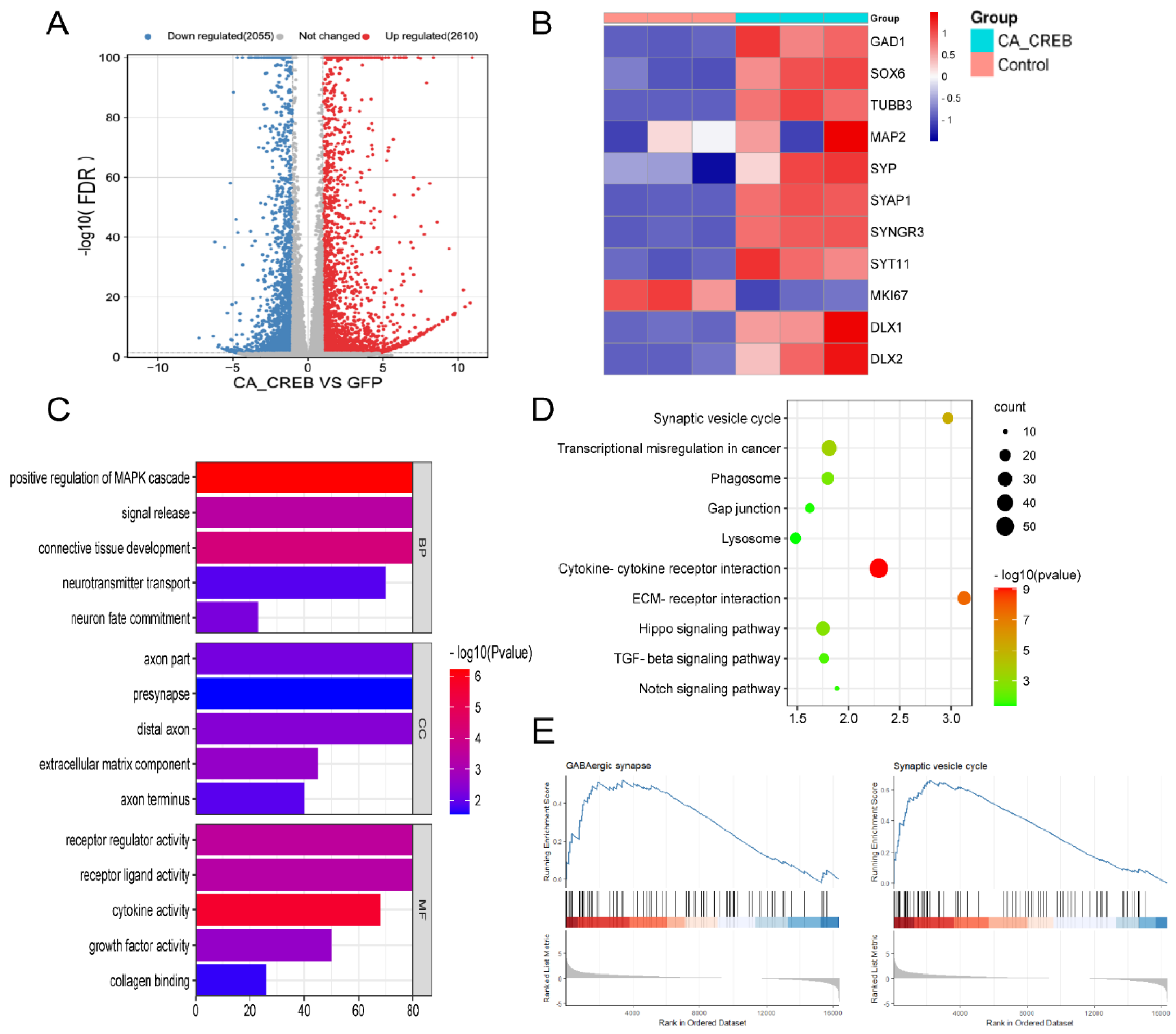
2.5. RNA-seq
2.6. Reverse Transcription and Quantitative PCR
2.7. Western Blotting
2.8. Chromatin Immunoprecipitation (ChIP)
2.9. Plasmid Construction and Dual-Luciferase Reporter Assay
2.10. Preparation of Transplanted Cell
2.11. Contusion SCI Model and Cell Transplantation
2.12. Perfusion and Sectioning
2.13. Immunofluorescence Staining
2.14. Behavioral Analysis
2.15. Statistical Analysis
3. Results
3.1. The Characterization of hUC-MSCs
3.2. Ectopic Expression of CA-CREB Induced MSC to Neuronal Phenotype In Vitro
3.3. RNA-seq Analysis Indicates That CREB1 Could Induce GABAergic Neuronal Expression at the Transcriptional Level
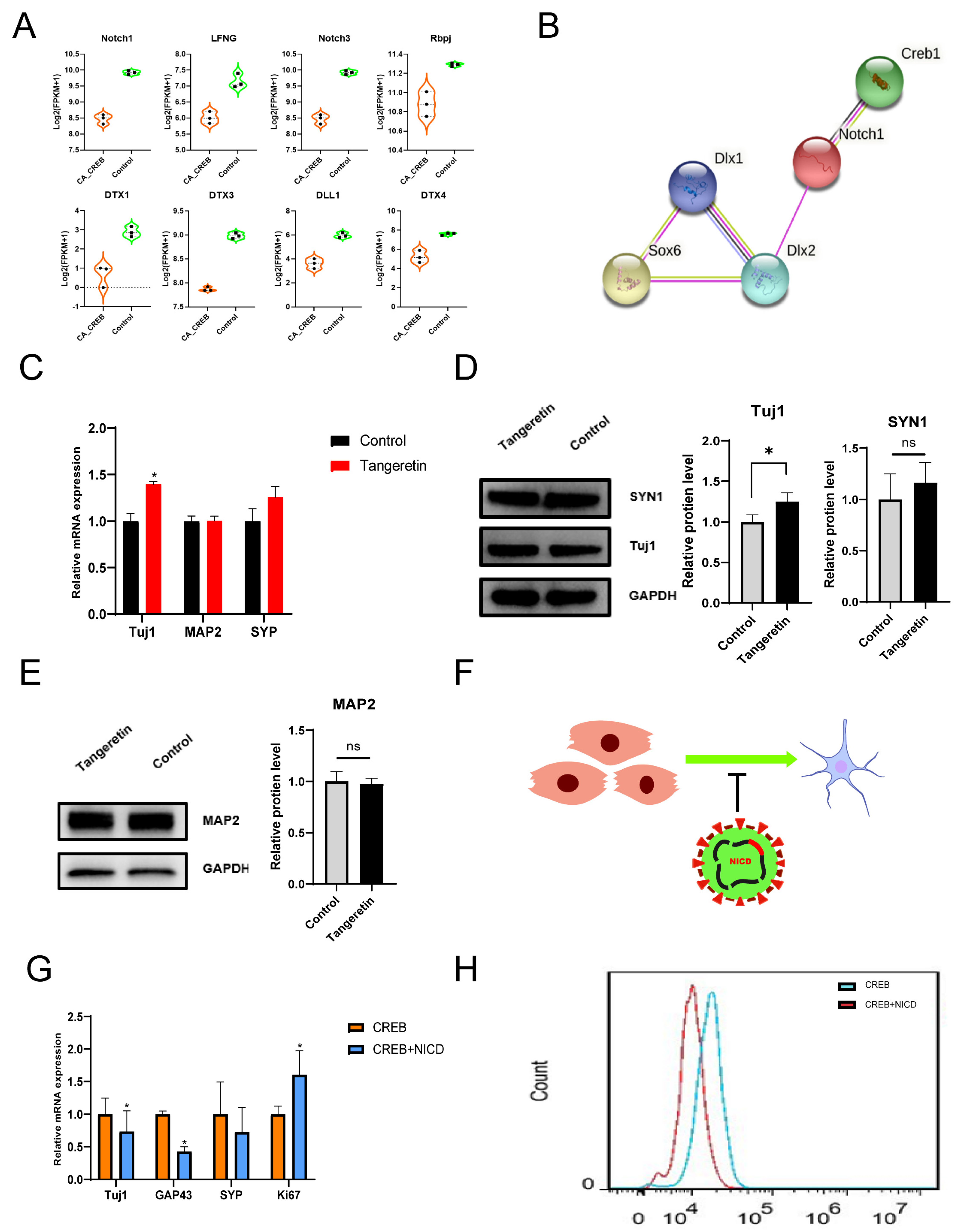
3.4. The Inhibition of the Notch Pathway Could Promote GABAergic Neural Differentiation In Vitro
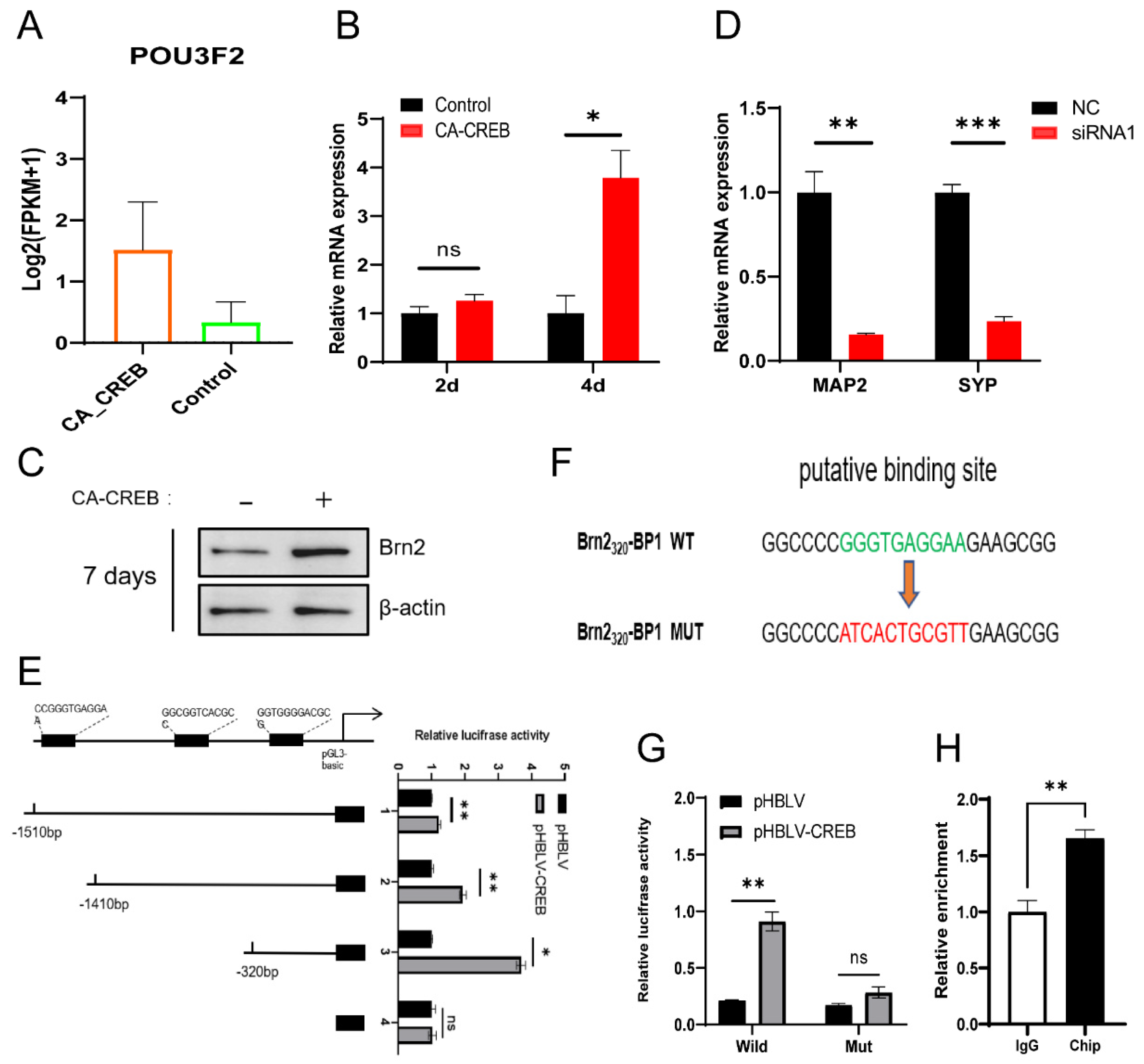
3.5. CREB1 Facilitates the Neuronal Maturation through Activating BRN2 Transcription In Vitro
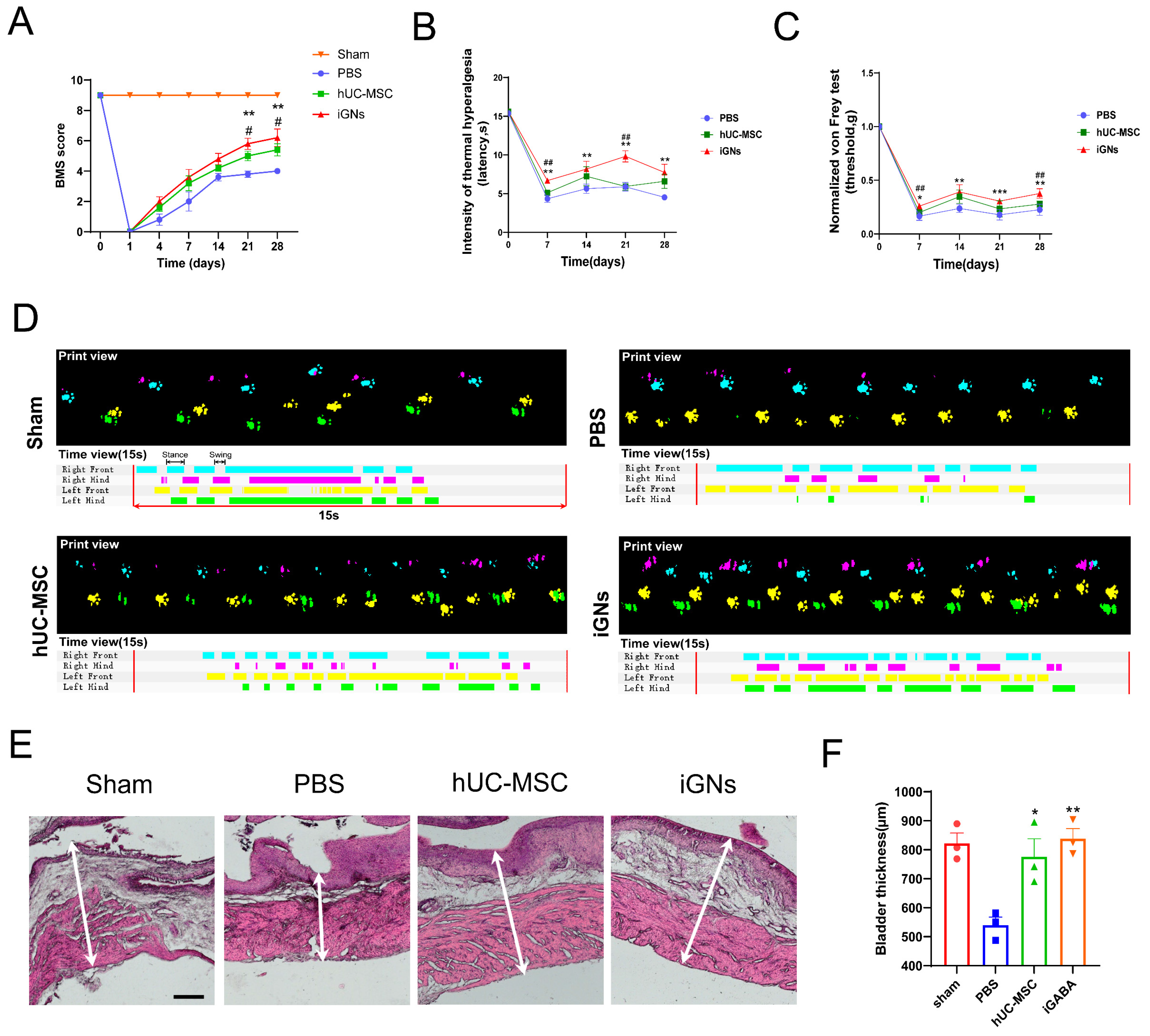
3.6. The Transfected hMSCs Could Differentiate to GABAergic Neuron Cells In Vivo and Promote Functional Recovery after SCI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SCI | spinal cord injury |
| hMSCs | human mesenchymal stem cells |
| hUC-MSCs | human umbilical cord mesenchymal stem cells |
| iGN | induced GABAergic neuron cell derived from hUC-MSCs |
| NSC | neural stem cell |
| NPC | neural progenitor cell |
| CsA | cyclosporine A |
| OPC | oligodendrocyte precursor cell |
| OEC | olfactory ensheathing cell |
| MSC | mesenchymal stem cell |
| MGE | medial ganglionic eminence |
| ESC-NPC | embryonic stem cell-derived neural precursor cell |
| iPSC | induced pluripotent stem cell |
| CA-CREB | constitutively active cAMP-responsive element-binding protein |
| DEG | differentially expressed gene |
| BPs | biological processes |
| CCs | cellular components |
| MFs | molecular functions |
| NICD | Notch1 intracellular domain |
| MEA | muse microelectrode array |
| BMS | Basso mouse scale |
References
- Fan, B.; Wei, Z.; Feng, S. Progression in translational research on spinal cord injury based on microenvironment imbalance. Bone Res. 2022, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Costăchescu, B.; Niculescu, A.-G.; Dabija, M.G.; Teleanu, R.I.; Grumezescu, A.M.; Eva, L. Novel Strategies for Spinal Cord Regeneration. Int. J. Mol. Sci. 2022, 23, 4552. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Nori, S.; Tetreault, L.; Wilson, J.; Kwon, B.; Harrop, J.; Choi, D.; Fehlings, M.G. Traumatic Spinal Cord Injury—Repair and Regeneration. Neurosurgery 2017, 80, S9–S22. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.A. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Wei, Z.; Yao, X.; Shi, G.; Cheng, X.; Zhou, X.; Zhou, H.; Ning, G.; Kong, X.; Feng, S. Microenvironment Imbalance of Spinal Cord Injury. Cell Transplant. 2018, 27, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-H.; Ning, G.-Z.; Feng, S.-Q.; Kong, X.-H.; Chen, J.-T.; Zheng, Y.-F.; Ban, D.-X.; Liu, T.; Li, H.; Wang, P. Transplantation of Autologous Activated Schwann Cells in the Treatment of Spinal Cord Injury: Six Cases, more than Five Years of Follow-up. Cell Transplant. 2012, 21 (Suppl. 1), S39–S47. [Google Scholar] [CrossRef]
- Curtis, E.; Martin, J.R.; Gabel, B.; Sidhu, N.; Rzesiewicz, T.K.; Mandeville, R.; Van Gorp, S.; Leerink, M.; Tadokoro, T.; Marsala, S. A First-in-Human, Phase I Study of Neural Stem Cell Transplantation for Chronic Spinal Cord Injury. Cell Stem Cell 2018, 22, 941–950.e6. [Google Scholar] [CrossRef]
- Kumamaru, H.; Kadoya, K.; Adler, A.F.; Takashima, Y.; Graham, L.; Coppola, G.; Tuszynski, M.H. Generation and post-injury integration of human spinal cord neural stem cells. Nat. Methods 2018, 15, 723–731. [Google Scholar] [CrossRef]
- van Gorp, S.; Leerink, M.; Kakinohana, O.; Platoshyn, O.; Santucci, C.; Galik, J.; Joosten, E.A.; Hruska-Plochan, M.; Goldberg, D.; Marsala, S.; et al. Amelioration of motor/sensory dysfunction and spasticity in a rat model of acute lumbar spinal cord injury by human neural stem cell transplantation. Stem Cell Res. Ther. 2013, 4, 57. [Google Scholar] [CrossRef]
- Bambakidis, N.C.; Miller, R.H. Transplantation of oligodendrocyte precursors and sonic hedgehog results in improved function and white matter sparing in the spinal cords of adult rats after contusion. Spine J. 2004, 4, 16–26. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, Y.; Zhu, W.; Bi, X.; Wang, Y.; Ye, M.; Zhou, G.-Q. The co-transplantation of human bone marrow stromal cells and embryo olfactory ensheathing cells as a new approach to treat spinal cord injury in a rat model. Cytotherapy 2008, 10, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Neuhuber, B.; Himes, B.T.; Shumsky, J.S.; Gallo, G.; Fischer, I. Axon growth and recovery of function supported by human bone marrow stromal cells in the injured spinal cord exhibit donor variations. Brain Res. 2005, 1035, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed]
- Mohammad-Gharibani, P.; Tiraihi, T.; Delshad, A.; Arabkheradmand, J.; Taheri, T. Improvement of contusive spinal cord injury in rats by co-transplantation of gamma-aminobutyric acidergic cells and bone marrow stromal cells. Cytotherapy 2013, 15, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Zheng, X.; Guo, F.; Wang, Y.; Zhang, S.; Chen, J.; Sun, X.; Shah, S.Z.A.; Zheng, Y.; Li, X.; et al. Human spinal GABA neurons alleviate spasticity and improve locomotion in rats with spinal cord injury. Cell Rep. 2021, 34, 108889. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Hahm, S.-C.; Choi, K.-A.; Park, S.-H.; Jeong, H.; Yea, J.-H.; Kim, J.; Hong, S. Intrathecal Transplantation of Embryonic Stem Cell-Derived Spinal GABAergic Neural Precursor Cells Attenuates Neuropathic Pain in a Spinal Cord Injury Rat Model. Cell Transplant. 2016, 25, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Askarian-Amiri, S.; Maleki, S.N.; Alavi, S.N.R.; Neishaboori, A.M.; Toloui, A.; Gubari, M.I.M.; Sarveazad, A.; Hosseini, M.; Yousefifard, M. The efficacy of GABAergic precursor cells transplantation in alleviating neuropathic pain in animal models: A systematic review and meta-analysis. Korean J. Pain 2022, 35, 43–58. [Google Scholar] [CrossRef]
- Manion, J.; Khuong, T.; Harney, D.; Littleboy, J.B.; Ruan, T.; Loo, L.; Costigan, M.; Larance, M.; Caron, L.; Neely, G.G. Human induced pluripotent stem cell-derived GABAergic interneuron transplants attenuate neuropathic pain. Pain 2020, 161, 379–387. [Google Scholar] [CrossRef]
- Zheng, Y.L. Some Ethical Concerns about Human Induced Pluripotent Stem Cells. Sci. Eng. Ethic- 2015, 22, 1277–1284. [Google Scholar] [CrossRef]
- Rohban, R.; Pieber, T.R. Mesenchymal Stem and Progenitor Cells in Regeneration: Tissue Specificity and Regenerative Potential. Stem Cells Int. 2017, 2017, 5173732. [Google Scholar] [CrossRef]
- Nagamura-Inoue, T.; Mukai, T. Umbilical Cord is a Rich Source of Mesenchymal Stromal Cells for Cell Therapy. Curr. Stem Cell Res. Ther. 2016, 11, 634–642. [Google Scholar] [CrossRef]
- Lv, F.; Lu, M.; MC Cheung, K.; Leung, V.Y.; Zhou, G. Intrinsic Properties of Mesemchymal Stem Cells from Human Bone Marrow, Umbilical Cord and Umbilical Cord Blood Comparing the Different Sources of MSC. Curr. Stem Cell Res. Ther. 2012, 7, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Elzat, E.Y.; Fan, X.; Yang, Z.; Yuan, Z.; Pang, Y.; Feng, S. Establishing a Mouse Contusion Spinal Cord Injury Model Based on a Minimally Invasive Technique. J. Vis. Exp. 2022, 187, e64538. [Google Scholar]
- Ran, N.; Li, W.; Zhang, R.; Lin, C.; Zhang, J.; Wei, Z.; Li, Z.; Yuan, Z.; Wang, M.; Fan, B.; et al. Autologous exosome facilitates load and target delivery of bioactive peptides to repair spinal cord injury. Bioact. Mater. 2023, 25, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, F.; Chen, H.; Liu, D.; Qu, T.; Li, X.; Xu, D.; Liu, F.; Yin, Z.; Chen, Y. Co-Transplantation of Human Umbilical Cord Mesenchymal Stem Cells and Human Neural Stem Cells Improves the Outcome in Rats with Spinal Cord Injury. Cell Transplant. 2019, 28, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.M.; Fisher, L.C.; Anderson, A.J.; Jakeman, L.B.; Mctigue, D.M.; Popovich, P.G. Basso Mouse Scale for Locomotion Detects Differences in Recovery after Spinal Cord Injury in Five Common Mouse Strains. J. Neurotrauma 2006, 23, 635–659. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, L.H.; Fehlings, M.G. Epidemiology, Demographics, and Pathophysiology of Acute Spinal Cord Injury. Spine 2001, 26, S2–S12. [Google Scholar] [CrossRef]
- Zipser, C.M.; Cragg, J.J.; Guest, J.D.; Fehlings, M.G.; Jutzeler, C.R.; Anderson, A.J.; Curt, A. Cell-based and stem-cell-based treatments for spinal cord injury: Evidence from clinical trials. Lancet Neurol. 2022, 21, 659–670. [Google Scholar] [CrossRef]
- Wang, H.; Xu, J.; Lazarovici, P.; Quirion, R.; Zheng, W. cAMP Response Element-Binding Protein (CREB): A Possible Signaling Molecule Link in the Pathophysiology of Schizophrenia. Front. Mol. Neurosci. 2018, 11, 255. [Google Scholar] [CrossRef]
- Zohny, S.M.; Habib, M.Z.; Mohamad, M.I.; Elayat, W.M.; Elhossiny, R.M.; El-Salam, M.F.A.; Hassan, G.A.M.; Aboul-Fotouh, S. Memantine/Aripiprazole Combination Alleviates Cognitive Dysfunction in Valproic Acid Rat Model of Autism: Hippocampal CREB/BDNF Signaling and Glutamate Homeostasis. Neurotherapeutics 2023, 20, 464–483. [Google Scholar] [CrossRef]
- Yao, D.; Zhao, X.; Zhao, S.; Shi, H.; Ma, Y.; Li, J. Characterization of the fatty acid binding protein 3 (FABP3) promoter and its transcriptional regulation by cAMP response element binding protein 1 (CREB1) in goat mammary epithelial cells. Anim. Biotechnol. 2023, 34, 1960–1967. [Google Scholar] [CrossRef] [PubMed]
- Whyte, W.A.; Orlando, D.A.; Hnisz, D.; Abraham, B.J.; Lin, C.Y.; Kagey, M.H.; Rahl, P.B.; Lee, T.I.; Young, R.A. Master Transcription Factors and Mediator Establish Super-Enhancers at Key Cell Identity Genes. Cell 2013, 153, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; Wu, W.; Wang, L.-L.; Ni, H.; Chen, C.; Yang, J.; Zang, T.; Zou, Y.; Xu, X.-M.; Zhang, C.-L. In vivo reprogramming of NG2 glia enables adult neurogenesis and functional recovery following spinal cord injury. Cell Stem Cell 2021, 28, 923–937.e4. [Google Scholar] [CrossRef] [PubMed]
- Tafoya, L.C.R.; Mameli, M.; Miyashita, T.; Guzowski, J.F.; Valenzuela, C.F.; Wilson, M.C. Expression and Function of SNAP-25 as a Universal SNARE Component in GABAergic Neurons. J. Neurosci. 2006, 26, 7826–7838. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Park, S.I.; Oh, J.H.; Kim, S.M.; Jeong, C.H.; Jun, J.A.; Lee, K.; Oh, W.; Lee, J.; Jeun, S. Brain-derived neurotrophic factor stimulates the neural differentiation of human umbilical cord blood-derived mesenchymal stem cells and survival of differentiated cells through MAPK/ERK and PI3K/Akt-dependent signaling pathways. J. Neurosci. Res. 2008, 86, 2168–2178. [Google Scholar] [CrossRef]
- Chen, J.; Dong, X.; Cheng, X.; Zhu, Q.; Zhang, J.; Li, Q.; Huang, X.; Wang, M.; Li, L.; Guo, W.; et al. Ogt controls neural stem/progenitor cell pool and adult neurogenesis through modulating Notch signaling. Cell Rep. 2021, 34, 108905. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, R.; Ohtsuka, T. The Notch-Hes pathway in mammalian neural development. Cell Res. 1999, 9, 179–188. [Google Scholar] [CrossRef]
- Cardano, M.; Zasso, J.; Ruggiero, L.; Di Giacomo, G.; Marcatili, M.; Cremona, O.; Conti, L. Epsins Regulate Mouse Embryonic Stem Cell Exit from Pluripotency and Neural Commitment by Controlling Notch Activation. Stem Cells Int. 2019, 2019, 4084351. [Google Scholar] [CrossRef]
- Antonios, J.P.; Farah, G.J.; Cleary, D.R.; Martin, J.R.; Ciacci, J.D.; Pham, M.H. Immunosuppressive mechanisms for stem cell transplant survival in spinal cord injury. Neurosurg. Focus 2019, 46, E9. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kao, Y.; Zhu, H.; Yang, Y.; Shen, W.; Song, W.; Zhang, R.; Liu, Y.; Liu, H.; Kong, X. CREB1 Facilitates GABAergic Neural Differentiation of Human Mesenchymal Stem Cells through BRN2 for Pain Alleviation and Locomotion Recovery after Spinal Cord Injury. Cells 2024, 13, 67. https://doi.org/10.3390/cells13010067
Kao Y, Zhu H, Yang Y, Shen W, Song W, Zhang R, Liu Y, Liu H, Kong X. CREB1 Facilitates GABAergic Neural Differentiation of Human Mesenchymal Stem Cells through BRN2 for Pain Alleviation and Locomotion Recovery after Spinal Cord Injury. Cells. 2024; 13(1):67. https://doi.org/10.3390/cells13010067
Chicago/Turabian StyleKao, Yanbing, Hanming Zhu, Yu Yang, Wenyuan Shen, Wei Song, Renjie Zhang, Yanchun Liu, Haoyun Liu, and Xiaohong Kong. 2024. "CREB1 Facilitates GABAergic Neural Differentiation of Human Mesenchymal Stem Cells through BRN2 for Pain Alleviation and Locomotion Recovery after Spinal Cord Injury" Cells 13, no. 1: 67. https://doi.org/10.3390/cells13010067