1. Introduction
Idiopathic pulmonary fibrosis is the most severe and fatal form of fibrosing lung diseases, with a median survival term of only 2 to 3 years after diagnosis [
1,
2]. Even with the poor prognosis of this disease, its etiology remains undetermined. Despite the poorly defined pathophysiology, it is thought to be caused by a chronic injury in the alveolar epithelial cells induced by aberrant wound healing [
3]. Unfortunately, current therapeutic strategies remain limited and can delay the disease progression rather than provide a cure [
4]. Therefore, there is an urgent need to develop new and efficient therapeutic strategies.
In addition to occurring in idiopathic pulmonary fibrosis, pulmonary fibrosis is also a common denominator of several lung disorders and is characterized by the accumulation of myofibroblasts and excessive deposition of extracellular matrix [
1,
5,
6]. These processes culminate in the formation of a fibrotic ‘scar’ which leads to the irreversible destruction of lung architecture and, ultimately, to death due to respiratory failure [
1,
5,
6]. There is increasing evidence that the immune system is involved in the progression of this pathologic condition. Macrophages are particularly prominent at sites of inflammation and fibrosis, rising as a crucial cell population in these processes [
7,
8].
During adulthood, macrophages are the central innate immune cells in the lung under homeostatic conditions and are known as the first line of defense in many organs [
9]. Diverse and distinct macrophage populations exist in the lung. Resident alveolar macrophages (AM) are mainly derived from precursors in the embryo and populate the airway space soon after birth [
10,
11,
12]. In a steady state, this population is capable of self-renewal and persists in the lung, the most represented macrophage population in normal adult mice lungs [
13]. Moreover, a population of recruited AM arises postnatally from circulating bone marrow-derived monocytes [
14]. AM are the best studied and understood lung macrophage population. However, interstitial macrophages (IM), infiltrated within the lung parenchyma, are rising as critical players in adult and murine lung fibrosis [
7,
13,
15]. The origin and function of IM are still not completely clarified, partly due to the difficulties in isolating this population [
13,
16]. It is believed that IM originate both from an embryonic yolk sac origin and postnatal bone marrow origin [
10,
17].
There is increasing evidence of the importance of interstitial and alveolar macrophages to pulmonary fibrosis [
18,
19,
20,
21]. However, mainly due to their heterogeneity and plasticity, depending on the microenvironment, which confers them the capacity to exert tissue-specific functions according to the transitory needs, but also due to, for example, the M1/M2 dichotomic metabolic state, among others (7, 8), their role may be controversial. Furthermore, macrophages can switch between two different activation states, which dictate their modulation of fibroproliferative responses. It has been suggested that classically activated macrophages (M1) maintain tissue inflammation, while alternatively activated macrophages (M2) play a role in resolving lung inflammation and aberrant wound healing cascade during fibrosis [
9,
22,
23].
Previous studies suggested that macrophages are present and adapt their phenotype and function when fibrogenesis is dominant. However, the detailed individual contribution of macrophages, both from the alveolar and interstitial compartments, has not been fully evinced in the early stages of the disease progression. Here, we use a bleomycin (BLM)-induced pulmonary fibrosis mouse model to characterize alveolar and interstitial macrophages in the initial stages of disease progression. In addition, we provide in vitro data to determine the paracrine effect played by IM in the advancement of lung fibrosis.
2. Materials and Methods
2.1. Mice
Eight-week-old gender- and age-matched C57BL/6 mice were bred under specific pathogen-free conditions and kept at the Life and Health Sciences Research Institute (ICVS) Animal Facility. Mice were fed ad libitum and kept under light/dark cycles of 12 h, temperature of 18–25 °C, and humidity of 40–60%.
All procedures in vivo followed the EU-adopted regulations (Directive 2010/63/EU). The ethical and regulatory approvals were consented to by the Direção Geral de Alimentação e Veterinária (DGAV, ref. 003671) and conducted according to the guidelines sanctioned by the Portuguese ethics committee for animal experimentation, DGAV.
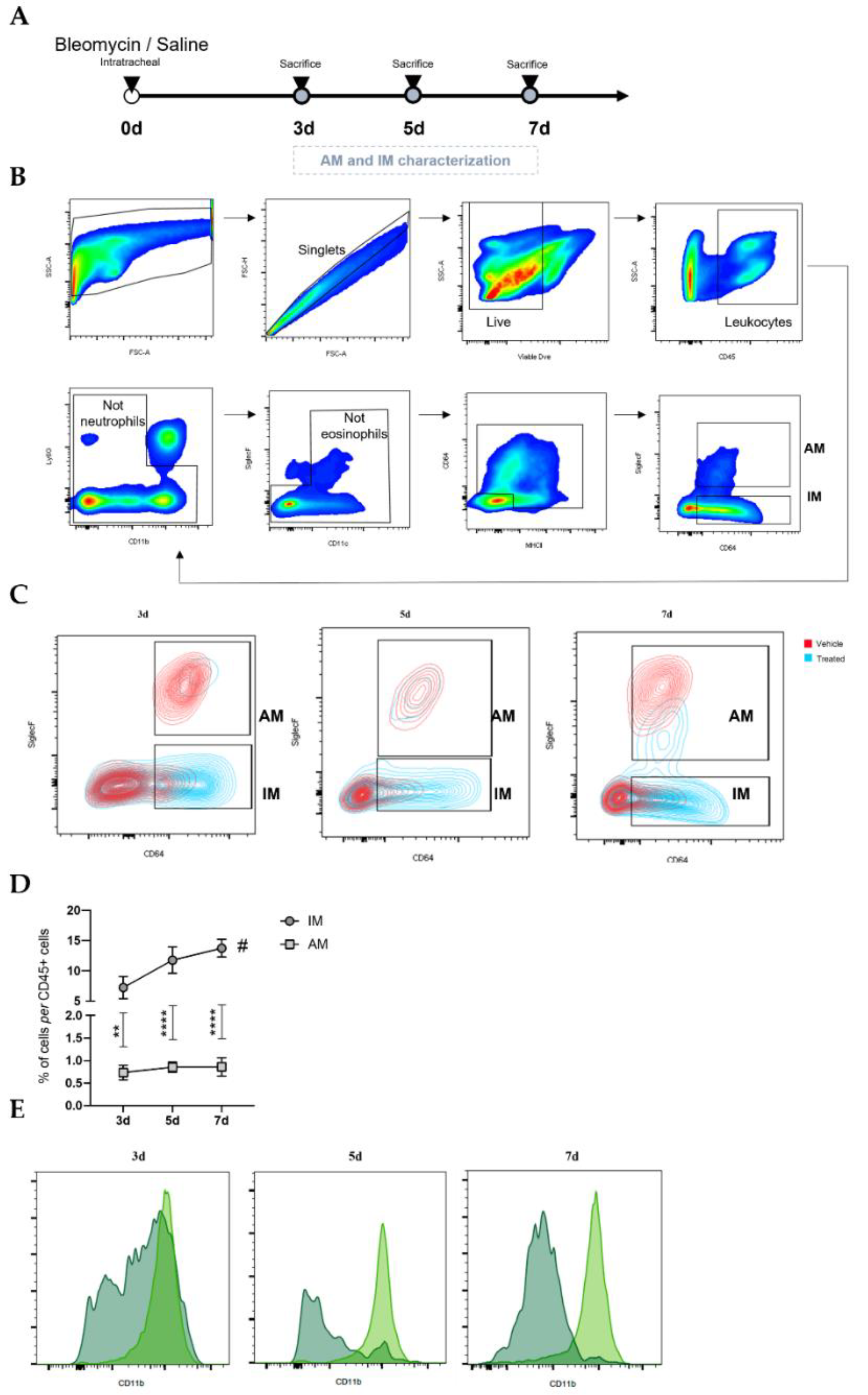
2.2. Bleomycin-Induced Pulmonary Fibrosis Mouse Model
Male C57BL/6J mice, 10 to 12 weeks old, were anesthetized with an intraperitoneal injection of ketamine and medetomidine, followed by intratracheal administration of BLM sulphate (BML-AP302-0010, Enzo Life Sciences, Miraflores, Portugal) as a single dose of 2 mg/kg body weight dissolved in 50 µL saline (treated) or, as control, with saline alone (untreated)—day 0. Weight was measured every single day throughout the experiment. At 3, 5, and 7 days after BLM administration, mice were euthanized. Mice were sacrificed by excessive doses of anesthesia to collect bronchoalveolar lavage fluid (BALF) and single-cell suspensions. Regarding the number of animals, 7–10 mice (per timepoint) were injected and sacrificed at day 3, 5, and 7 for flow cytometry analysis. For fluorescence-activated cell sorting, performed at day 7, 15 mice were injected with saline (untreated), and 15 mice were injected with bleomycin.
2.3. Bronchoalveolar Lavage (BAL)
In brief, the trachea was exposed by a midline incision and cannulated. Bilateral BALF was recovered by lavage three times with 1 mL PBS 1×. After collection, BALF cells were maintained on ice to be then added to single-cell suspensions obtained from tissue dissociation.
2.4. Preparation of Lung Single-Cell Suspensions
Briefly, mice were over-anesthetized intraperitoneally and perfused with 20 mL of sterile PBS 1×. After dissection, lungs were placed in dispase (11553550, Corning, Corning, NY, USA) while being minced into 2–3 mm3 pieces for 20 min at room temperature. Then, they were also digested in Dulbecco’s Modified Eagle Medium (DMEM) 1× (21969-035, Gibco, Waltham, MA, USA) containing 1 mg/mL DNase I (10104159001, Roche, Basel, Switzerland) and 5 mg/mL collagenase II (17101-015, Gibco). After an enzymatic digestion period of 10 min at 37 °C, tissue disruption was performed by mechanical titrations with P1000 tip, and then a second incubation period of 10 min was performed at 37 °C. After incubation, the disrupted tissue suspension was collected and sequentially filtered through 100 µm and 70 µm cell strainers. The pellet was resuspended in ACK lysis buffer (A1049201, Gibco), which was then incubated for 4 min. Cells were then resuspended in cold PBS 1× containing 4% fetal bovine serum (FBS-10270-106, Gibco). These lung single-cell suspensions were used to analyze the expression of various surface markers for immune cells by flow cytometry and magnetic (MACS)- and fluorescence (FACS)-activated cell-sorting-based separation.
2.5. Flow Cytometry
In brief, lung cells were harvested, washed, and resuspended in cold PBS containing 4% FBS. Then, 1 × 106 cells were preincubated with TruStain FcX anti-mouse CD16/32 antibody (Biolegend (San Diego, CA, USA), 101320) for 10 min at room temperature. This blocking of non-specific binding was followed by incubation for 30 min on ice, in the dark, with the following anti-mouse antibodies: BV510-conjugated CD45 (1:100—103138, Biolegend), PE-conjugated SiglecF (1:200—552126, BD Biosciences, San Jose, CA, USA), PE/Cy7-conjugated CD11b (1:100—101216, Biolegend), FITC-conjugated CD64 (1:200—139316, Biolegend), PerCP/Cy5.5-conjugated CD11c (1:100—117328, Biolegend), BV605-conjugated Ly6G (1:100—127639, Biolegend), Alexa Fluor 647-conjugated I-Ab (1:100—115310, Biolegend), APC/Cy7-conjugated CD24 (1:100—47-0242-80, Invitrogen, Waltham, MA, USA), BV650-conjugated CD80 (1:100—104731, Biolegend), and BV786-conjugated CD206 (1:100—141729, Biolegend). Then, cells were incubated with Viable Dye (VD) eFluor® 450 (65-0863-14, EBioscience, San Diego, CA, USA) for 30 min at 4 °C in the dark. Cells were resuspended and fixed in 4% paraformaldehyde for 20 min on ice in the dark.
Regarding intracellular staining with PE-Cy7-conjugated TNF-α (1:100—506323, Biolegend), before the membrane staining, cells were preincubated for 4 h at 37 °C with 10 ng of Phorbol 12-myristate 13-acetate (P8139, Sigma-Aldrich, St. Louis, MO, USA), 100 ng of ionomycin (I0634, Sigma-Aldrich), and 2 µg of brefeldin A (B7651, Sigma-Aldrich) in DMEM 1× supplemented with 10% FBS and 1% penicillin/streptomycin (P4333, Sigma-Aldrich). Then, the staining proceeded as mentioned in the previous paragraph, except for CD11b; in this case, we used APC-conjugated antibodies (1:200—553312, BD). After fixation, cells were permeabilized in 0.5% saponin in PBS 1×, and then intracellular staining was performed.
Data were acquired on a LSRII flow cytometer and analyzed using FlowJo Software.
For FACS-based separation of IM and AM contained in the CD45-positive fraction, the following antibodies were used: anti-mouse PE/Cy7-conjugated CD45 (1:200—552848, BD Biosciences), PE-conjugated SiglecF (1:200—552126, BD Biosciences), APC-conjugated CD11b (1:400—553312, BD Biosciences), FITC-conjugated CD64 (1:300—139316, Biolegend), PerCP/Cy5.5-conjugated CD11c (1:200—560584, BD Biosciences), and APC/Cy7-conjugated Ly6G (1:300—560600, BD Biosciences).
2.6. Reverse Transcription Quantitative PCR
At day 7 after BLM administration, FACS-sorted IM and AM were stored in RLT buffer and frozen at −80 °C. Total RNA was isolated from sorted interstitial and alveolar macrophages using RNeasy Plus Micro Kit following the instructions provided by the manufacturer (74034, Qiagen, Hilden, Germany).
First-strand cDNA was synthesized using the Super Script III First-Strand Synthesis Supermix kit, which was used to convert all the extracted RNA following the instructions provided by the manufacturer (18080400, Invitrogen).
Reverse transcription quantitative PCR (RT-qPCR) was conducted using the 7500 Fast Real-Time PCR System (Applied Biosystems, Waltham, MA, USA, Life Technologies, Carlsbad, CA, USA). RT-qPCR was performed using Taqman Universal PCR MasterMix (4304437, Thermo Fisher Scientific, Waltham, MA, USA) and one TaqMan® Gene Expression assay.
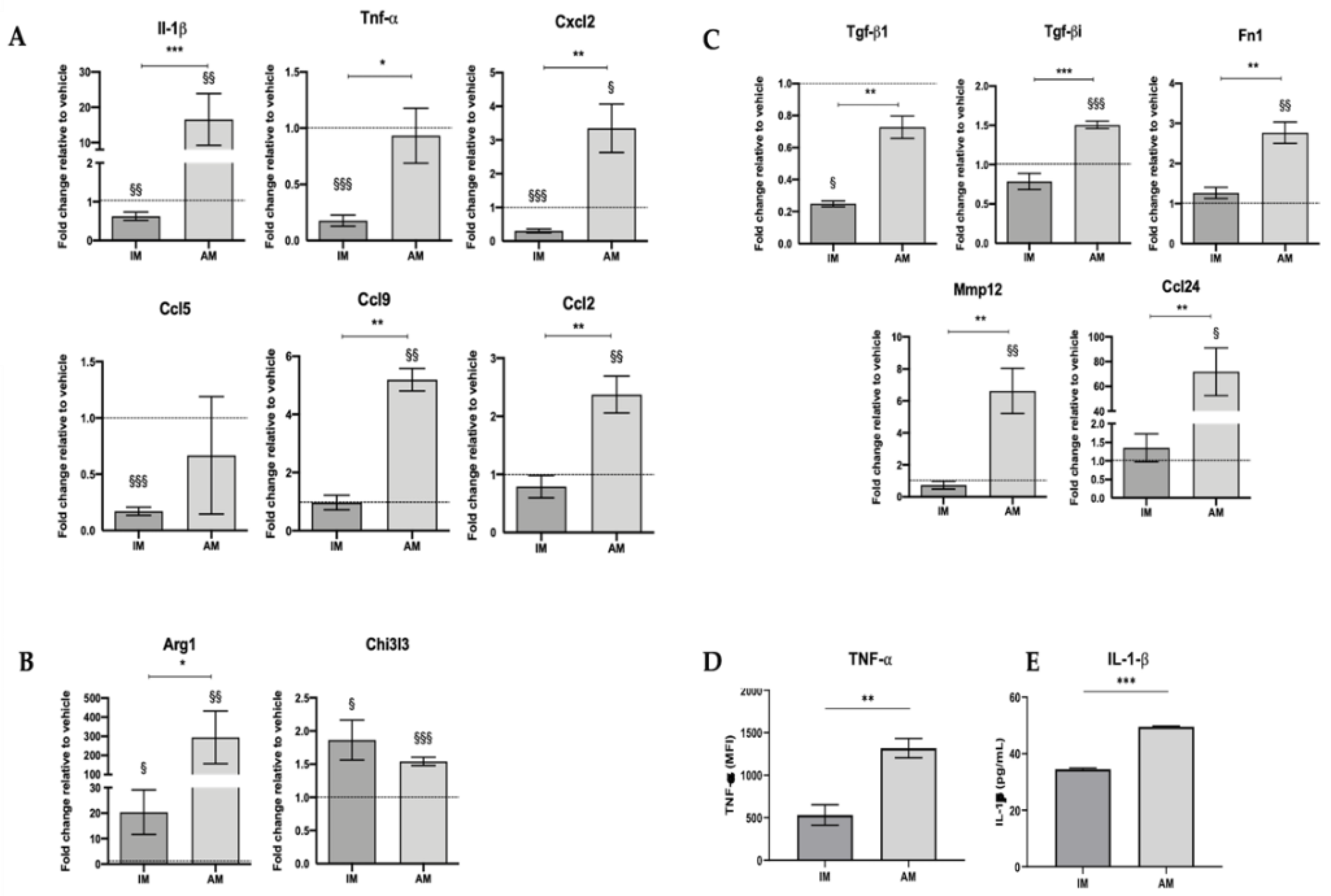
The following probes were used for each target gene: Gapdh (Mm99999915_g1), IL-1β (Mm00434228_m1), Tnf-α (Mm00443258_m1), Ccl2 (Mm00441242_m1), Cxcl2 (Mm00436450_m1), Ccl5 (Mm01302427_m1), Ccl6 (Mm01302419_m1), Ccl9 (Mm00441260_m1), Arg1 (Mm00475988_m1), Tgf-β1 (Mm01178820_m1), Tgf-bi (Mm01337605_m1), Chi3l3/ Ym1 (Mm00657889_m1), Ccl24 (Mm00444701_m1), Fibronectin 1 (Mm01256744_m1), Mmp12 (Mm00500554_m1).
Relative expression levels for each target gene were calculated using the 2−ΔΔCt method; Ct values of target genes were normalized to the reference gene Gapdh. Data were presented in fold change, which defined the gene’s relative expression in the respective vehicle group to 1.
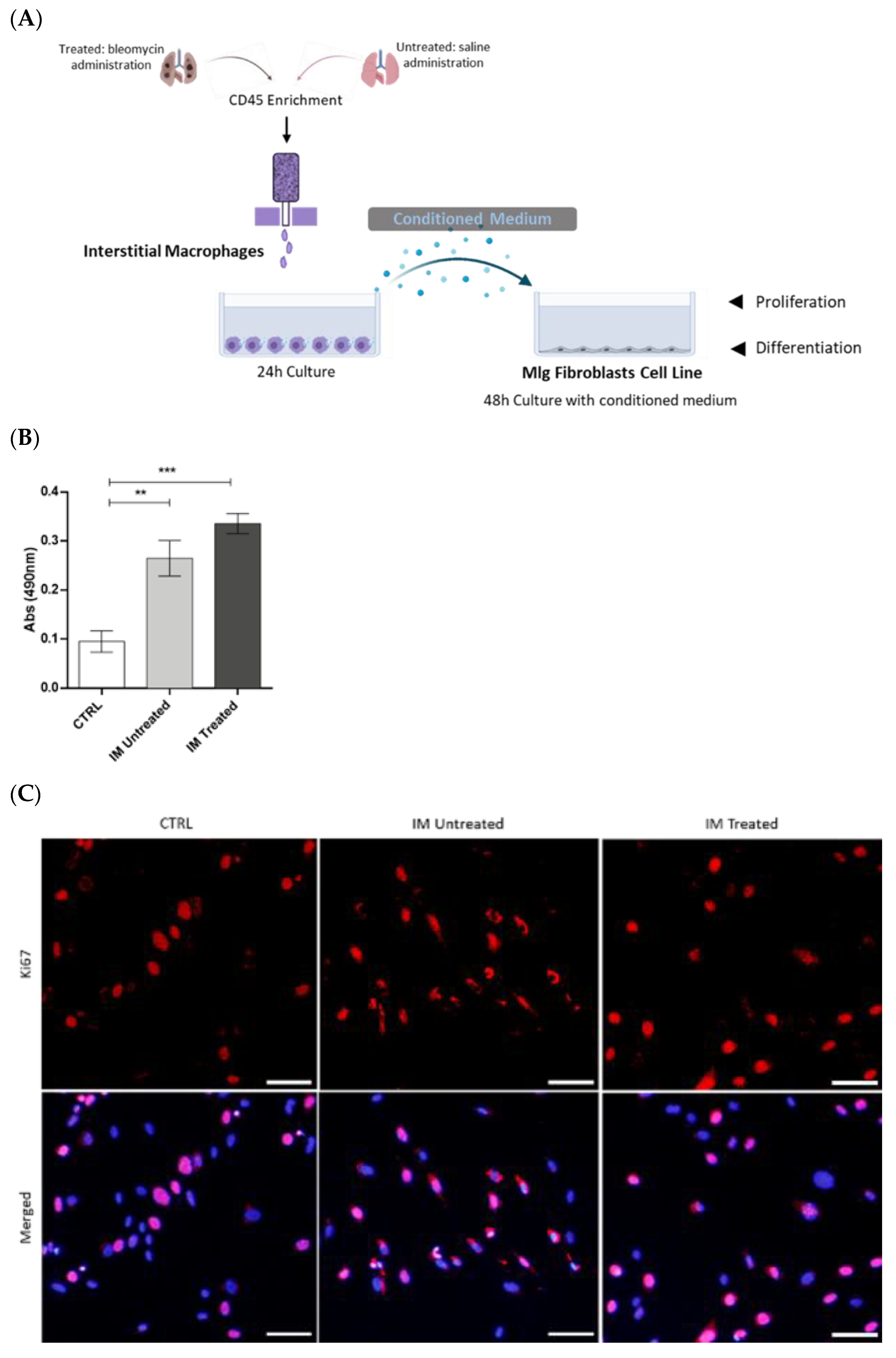
2.7. Interstitial Macrophages Fluorescence-Activated Cell Sorting and Culture
Untreated and treated animals were used to obtain interstitial macrophages for primary culture and to collect conditioned medium (CM).
The lung single-cell suspensions were incubated with anti-CD45 MicroBeads (130-052-301, Miltenyi Biotec, Bergisch Gladbach, Germany) to enrich the populations of interest, which are minor in the lung.
CD45+ cells were collected over MS column separation (130-042-201, Miltenyi Biotec) in the OctoMACS separator (130-042-109, Miltenyi Biotec) magnetic field. Viable cells were counted using trypan blue, and flow cytometry staining was performed to proceed further to the fluorescence-activated cell sorting procedure using BD FACSARIA II cell sorter (BD Bioscience).
Following cell sorting, 2.5 × 105 macrophages in DMEM 1× supplemented with 10% FBS, and 1% penicillin/streptomycin were plated in SterilinTM Square Petri Dishes (Thermo Fisher Scientific) for 24 h at 37 °C in a humidified atmosphere containing 5% CO2. After this time, the CM was collected, centrifuged at 1500 rpm for 5 min, and then stored at −80 °C.
2.8. Culture and Stimulation of MLg Cell Line
MLg (MLg 2908, ATCC® CCL206™), a mouse lung fibroblast cell line, was used in in vitro assays. MLg cells were maintained in Minimum Essential Medium (MEM; 31095-029, Gibco) supplemented with 10% FBS, 10 mM HEPES (15630-056, Gibco), and 1% penicillin–streptomycin solution. Cells were maintained at 37 °C in a humidified atmosphere containing 5% CO2. When cells reached 80–90% confluence, they were enzymatically dissociated using TrypLE Express Enzyme (12605-028, Gibco) for 5 min at 37 °C, and the cell suspension was passed to a new flask in a low dilution.
Based on Meziani et al. [
19], 5 × 10
3 or 1.5 × 10
5 MLg cells in DMEM 1× containing 10% FBS, 10 mM HEPES, and 1% penicillin–streptomycin were seeded in a regular 96-well plate (MTS assay), µ-Plates 96 Well Black (immunofluorescence—89626, Ibidi), or in a regular 6-well plate (Western blot), respectively, and were allowed to adhere overnight. Next, these cells were cultured for 48 h with untreated and treated, sorted, IM-derived CM with 10% FBS. In control conditions, MLg cells were cultured in the same medium without contact with IM-derived CM. The supplementation with 10% FBS was necessary as MLg cells do not survive when cultured without it, not even in lower percentages.
2.9. Metabolic Activity Assay
MLg proliferation after stimulation with control or CM for 48 h was assessed by MTS assay according to the manufacturer’s instructions (CellTiter 96 Aqueous One Solution, PROMEGA, Milan, Italy). Briefly, MLg were incubated in control medium with 1:10 MTS for 2.5 h at 37 °C. Next, 490 nm absorbances were read using a 96-well plate reader (Varioskan Flash—Thermo Fisher Scientific, Vantaa, Finland). All assays were performed in triplicate.
2.10. Immunofluorescence
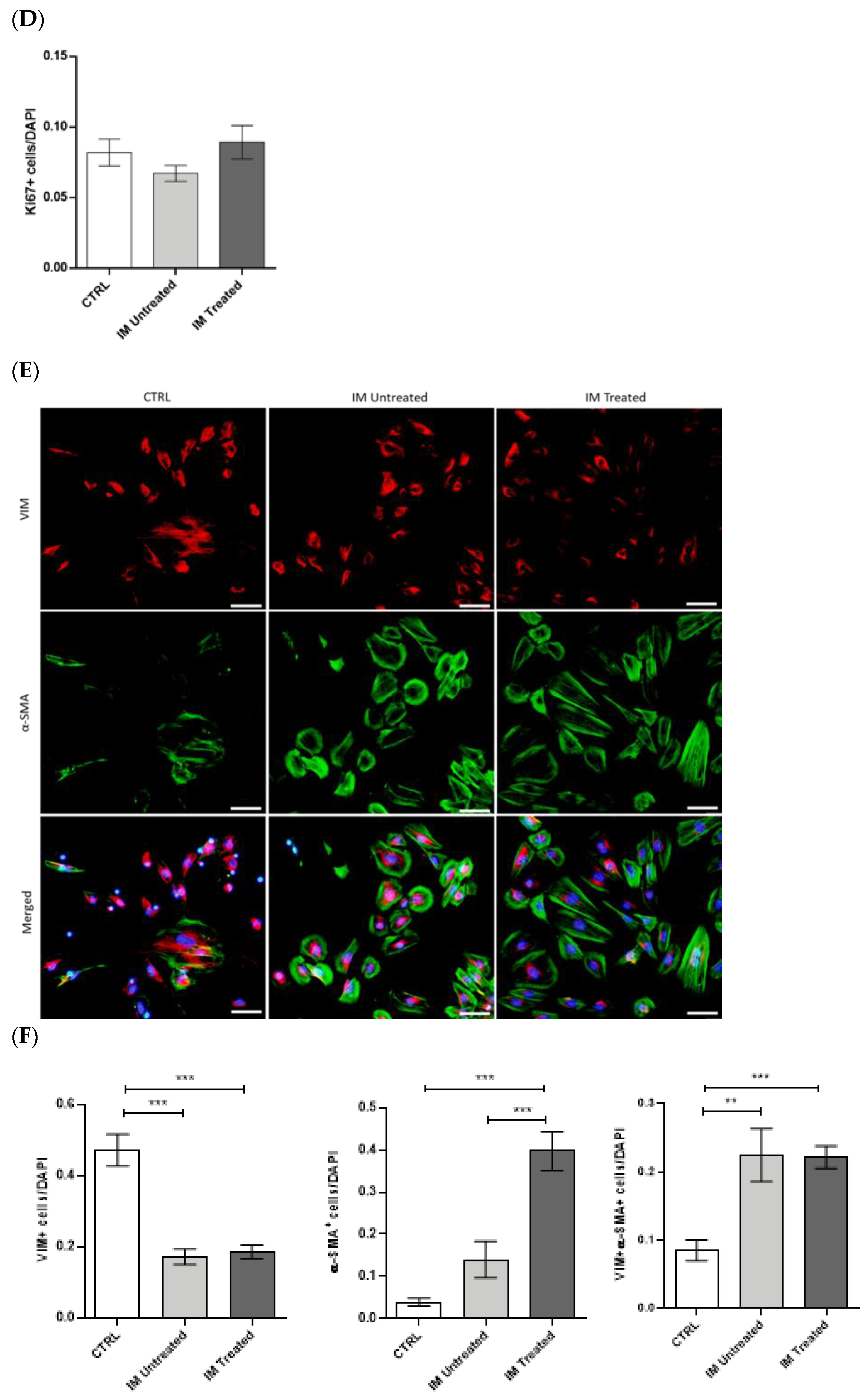
The proliferative and transdifferentiating state of MLg cells after CM stimulation was assessed by immunofluorescence. Following 48 h of stimulation with IM-derived conditioned medium, the medium was carefully removed, and cells were washed with PBS 1×. Then, adherent cells were fixed with PFA 4% (proliferation protocol) or methanol 100% (transdifferentiation protocol) for 30 min. Fixed cells were permeabilized in Triton-X-100 0.3% in PBS 1× for 20 min in the case of the proliferation protocol. Cells were washed twice with PBS Tween 0.5% for 10 min, and blockage of non-specific binding was performed with newborn calf serum (proliferation protocol; 1/10—186427, Biochrom) or normal goat serum (transdifferentiation protocol; 1/10—G9023-0010, Sigma-Aldrich) for 1 h. Next, cells were incubated with a rabbit anti-mouse Ki-67 (1:250 in blocking solution—AB9260, Millipore), rabbit polyclonal anti-mouse vimentin (1:150 in blocking solution—#5741, CellSignalling Technology, Danvers, MA, USA), and mouse anti-mouse α-smooth muscle actin (SMA, 1:100 in blocking solution—ab32575, Abcam, Cambridge, UK) primary antibodies overnight. Cells were washed and incubated for 2 h with goat anti-rabbit conjugated to Alexa Fluor 568 (1:250 in blocking solution—a11011, Thermo Fisher Scientific) and goat anti-mouse conjugated to Alexa Fluor 488 (1:250 in blocking solution—a11001, Thermo Fisher Scientific) secondary antibodies. Finally, cells were washed and incubated with DAPI for 5 min, mounted in Ibidi Mounting Medium (50001, Ibidi, Gräfelfing, Germany), and images were obtained with an inverted fluorescence microscope (Olympus widefield inverted microscope IX53, Olympus, Japan) using the 10× objective. Data were presented as positive cells per DAPI-positive cells.
2.11. Protein Extraction
The transdifferentiating state of MLg cells after CM stimulation was assessed by Western blot. Following 48 h of stimulation with IM-derived conditioned medium, the medium was carefully removed, and cells were washed with PBS 1×. Then, protein was extracted by scraping the cells after adding lysis buffer (50 mM Tris-Base, 150 mM NaCl, 5 mM EDTA, 1% Triton X-100, and 1:7 of cOmplete protease inhibitors cocktail (11697498001, Roche)). Lysed cells were collected, kept on ice for 15 min, and centrifuged at 10,000 rpm at 4 °C for 15 min. The supernatant was collected, and protein was quantified using Bradford reagent (B6916, Sigma-Aldrich). For quantification, 2 µL of protein extracts, as well as a BSA standard curve, was added to each well of a 96-well plate, followed by 98 µL of PBS 1× and 200 µL of Bradford reagent. After a 10 min incubation period, absorbance was measured at 590 nm in the microplate reader (Varioskan Flash-Thermo FisherScientific, Vantaa, Finland).
2.12. Western Blot Analysis
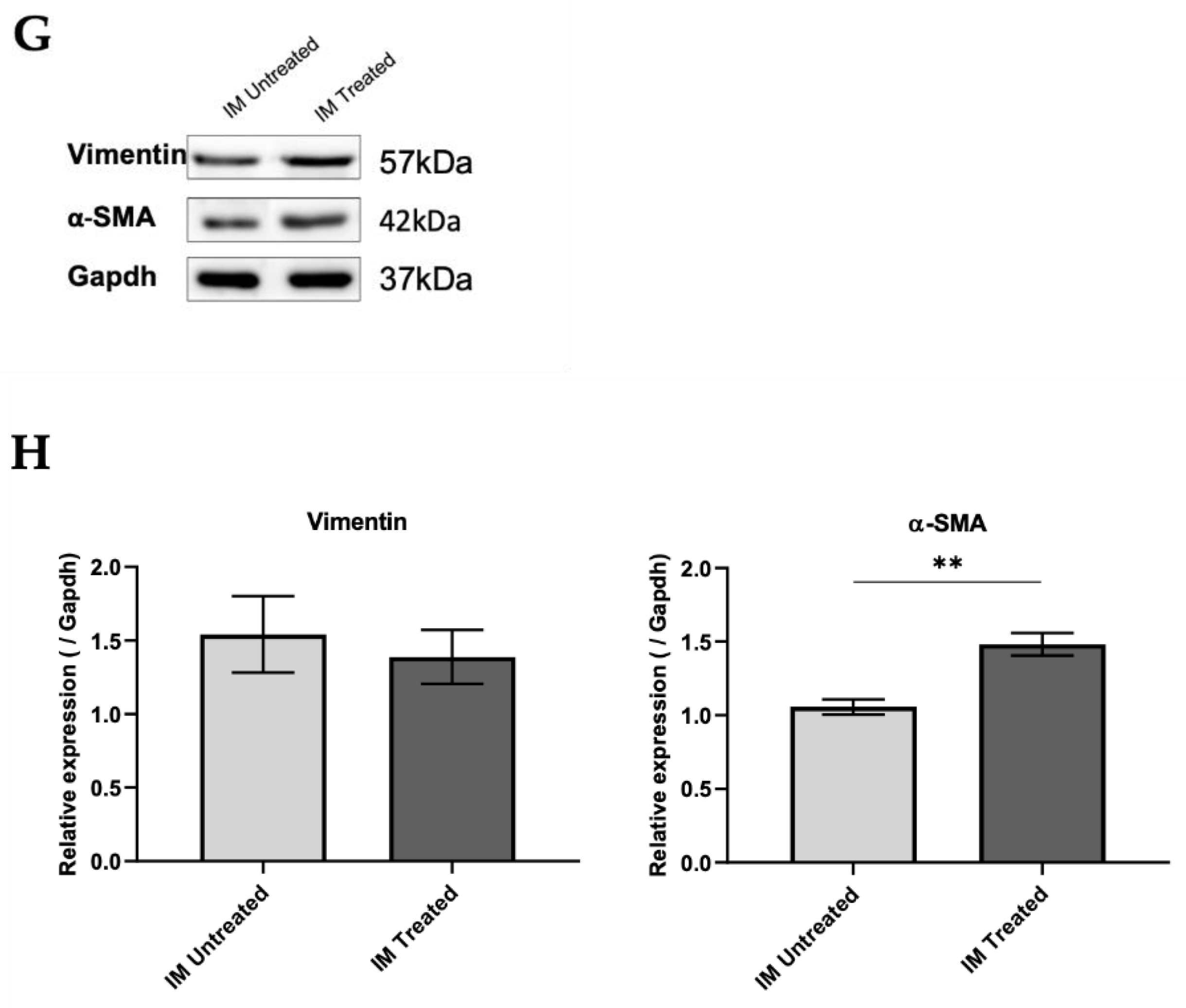
Aliquots of 20 µg of protein from MLg cells were prepared in the proportion of 1:1 with loading buffer (950 μL of Laemmli Sample Buffer 2× (161-0737, Biorad) and 50 μL 2-mercaptoethanol (63689, Millipore)) and denatured over 5 min at 98 °C. Samples were then loaded onto a 10% sodium dodecyl sulphate–polyacrylamide (SDS-PAGE) gel and transferred to Amersham Protran nitrocellulose membranes (GE10600008, Sigma-Aldrich) using the Trans-Blot Turbo Transfer System (BioRad Laboratories, München, Germany). After, membranes were blocked for 1 h with TBS 0.1% Tween containing 5% of BSA. Membranes were incubated overnight at 4 °C with the primary antibodies for vimentin (1:1000, rabbit—#5741, CellSignalling Technology), α-SMA (1:500, mouse—ab32575, Abcam), and Gapdh (1:2500, rabbit—ab9485, Abcam). After incubation, membranes were washed three times in TBS 0.1% Tween and then incubated with a secondary antibody coupled to horseradish peroxidase (HRP, 1:2000, anti-rabbit—7074P2, CellSignalling Technology and 1:3000, anti-mouse—sc-516102, Santa Cruz Biotechnology) diluted in 5% milk for 1 h at RT. Membranes were washed three times with TBS 0.1% Tween, and signal was detected in Sapphire Biomolecular Imager (Azure Biosystems, Dublin, CA, USA) using Western Bright Sirius HRP Substrate Kit (K-12043-D10, Advansta, San Jose, CA, USA). The bands were quantified in ImageJ software, and quantifications were normalized to Gapdh levels.
2.13. Statistical Analysis
Data were analyzed and plotted using GraphPad Prism 6.00 (GraphPad Software, San Diego, CA). All results were expressed as group mean ± standard error of the mean (SEM). A p-value of less than 0.05 was considered a statistically significant difference. For all comparisons, normality was assessed with Shapiro–Wilk test; if normality was not verified, non-parametric tests were used. Comparisons between two groups were performed either with a parametric, unpaired, two-tailed Student’s t-test or a non-parametric, two-tailed Mann–Whitney test. Comparisons between three groups were performed either with parametric one-way ANOVA or a non-parametric Kruskall–Wallis test.
4. Discussion
Pulmonary fibrosis is a common and severe pathological condition, and since there are no effective therapies to halt this condition, the discovery of new treatments is urgently needed. Therefore, a better understanding of macrophages’ signature and functionality in this context is critical to unraveling new therapeutic strategies.
Pulmonary fibrosis is a rapidly progressive condition characterized by the activation of fibroblasts and their differentiation into myofibroblasts. These cells are responsible for the excessive production and consequent accumulation of extracellular matrix [
1,
5,
6]. Our study used the BLM-induced pulmonary fibrosis mouse model. BLM intratracheal administration is one of the best established and most widely used mouse models for studies that mimic human pulmonary fibrosis [
25]. Intratracheal instillation of BLM is associated with the inflammation phase during the first week, reaching a peak at day 7, and a subsequent fibroproliferative phase leading to excessive collagen deposition [
26]. Indeed, several studies demonstrated that this model shares cellular and molecular mechanisms important in the pathogenesis of idiopathic pulmonary fibrosis and other fibrotic interstitial lung human diseases [
27,
28,
29]. We demonstrated in the early stage of this model that IM accumulate in the lung and become phenotypically activated in the disease progression. In vitro, this macrophage population leads to myofibroblast activation by paracrine mechanisms.
Our flow cytometry data in early BLM-induced pulmonary fibrosis showed that the proportion of IM increased throughout the disease and peaked at day 7. On the other hand, the proportion of AM decreased drastically and remained unaltered during these periods. These findings suggest the critical involvement of IM in the fibrogenesis process of the disease and a slightly residual role of the AM population. Moreover, our results show that the population of IM is positive for CD11b, while AM present a low percentage of positive staining. These results indicate that the increase in IM throughout the inflammatory phase of BLM-induced pulmonary fibrosis is due to their monocytic origin. This intensified recruitment and predominance of IM in the lung reinforces the potential involvement of this macrophage population rather than AM in the disease pathogenesis.
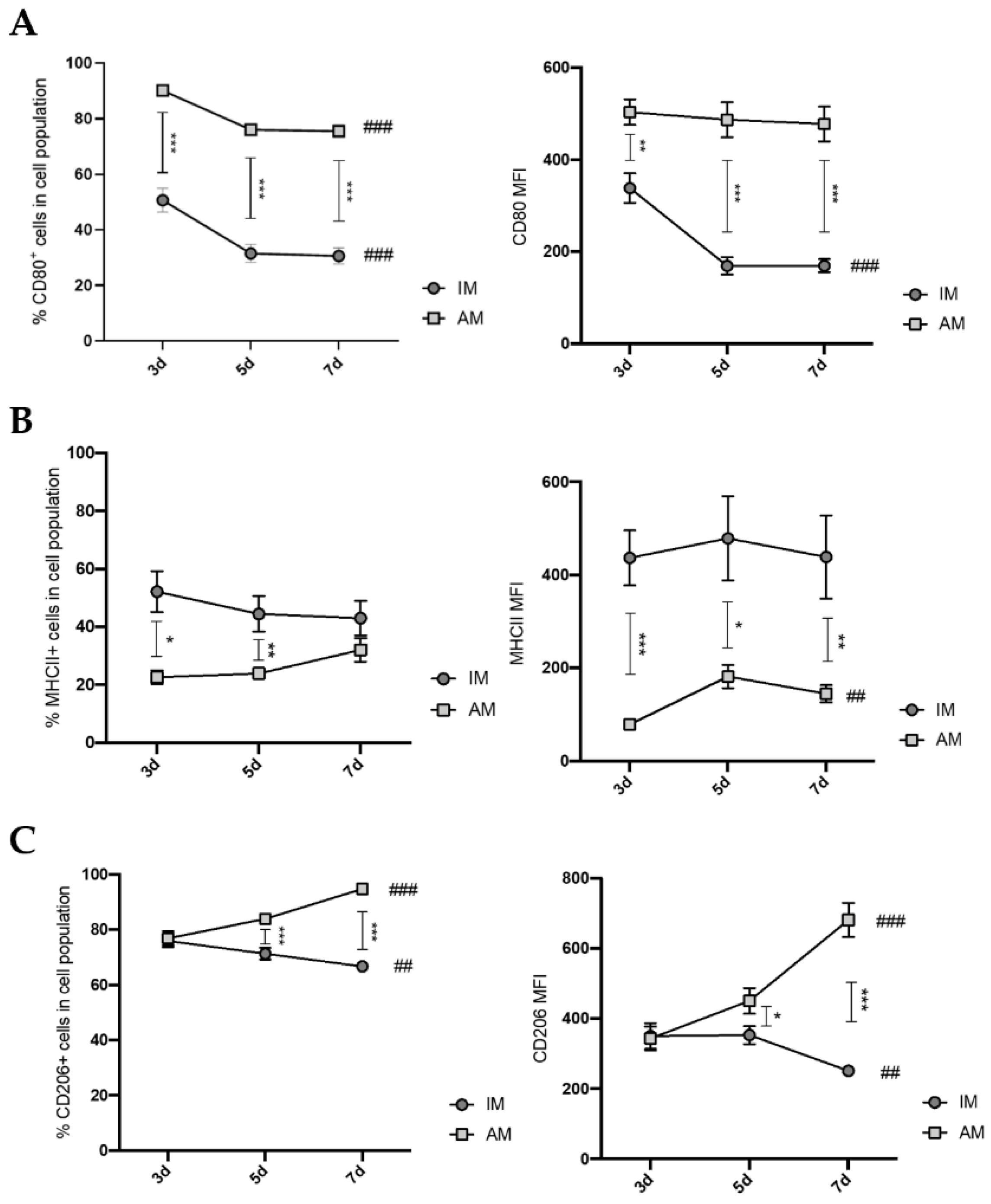
Macrophages present a colossal capacity to modulate their phenotype and function according to the tissue microenvironment. In this way, we investigated the respective activation state of AM and IM 7 days after BLM administration. Our results demonstrated that, until day 7, AM presented increasing expression levels of CD206 and decreasing expression levels of CD80. In addition, at the gene relative expression levels, several M1-, M2-like, and pro-fibrotic factors were simultaneously elevated. Other reports presented the same tendency, such as Misharin et al. [
20], who suggested that, during the early stages of BLM-induced pulmonary fibrosis mouse model, an M1-like response occurs. Instead of its cessation, the M2-like response begins simultaneously in the following stages, and then the M1-like response ceases at later fibrotic stages. This study showed that, in the early stages of BLM-induced pulmonary fibrosis (5 days), AM express higher levels of CD206 and CD80 than the vehicle control, while, in later stages (21 days), AM only express higher levels of CD206 than the vehicle control. Zhang et al. [
30] described that, on day 14 after BLM administration, AM had higher relative expression levels of Arg1 and lower expression levels of Tnf-α than vehicle control. Furthermore, Zhu et al. [
31] stated that, on day 17 after BLM administration, AM expressed higher CD206 levels than the control. These data indicate that, at early stages of BLM-induced lung fibrosis, AM are transient state from an M1-like to an M2-like phenotype.
Regarding IM, our data showed that IM have slightly decreased levels of CD206 and CD80 expression during the inflammatory phase. Additionally, at the gene relative expression levels, several pro-inflammatory factors were reduced, and anti-inflammatory factors were elevated compared with the vehicle control, which is in accordance with previous studies on day 10 after BLM administration. Misharin et al. [
20] showed that, on day 5 after BLM administration, IM have increased CD80 expression and lower CD206 expression compared with the vehicle control, and on day 21, no differences were found in the expression of these markers. Ucero et al. [
32] and Zhang et al. [
33] described that, at day 10 after BLM administration, IM have higher relative expression levels of CD206 and Arg1 compared with the vehicle control, suggesting that, at this phase, IM are activated towards an M2-like phenotype. In line with this, our findings revealed that, at day 7, IM were activated towards an M2-like phenotype.
Since IM accumulate in BLM-treated mice’s lungs, M2-like macrophages are associated with aberrant tissue healing and consequent fibrosis through the secretion of anti-inflammatory and pro-fibrotic factors [
7,
22,
23,
34], we further investigated the role of IM in fibrogenesis using in vitro experiments. We showed that CM from sorted IM of BLM-treated mice promoted fibroblasts proliferation and differentiation into myofibroblasts. Our findings comply with other studies. For instance, Meziani et al. [
18] used sorted IM (F4/80
+ Gr1
−) from radiation-induced lung fibrosis and activated them with interferon-gamma to induce the M1-like phenotype or IL-13/IL-4 to induce the M2 phenotype. After being co-cultured with M2-activated IM, normal fibroblasts overproduce extracellular matrix components and increase their α-SMA expression. On the other hand, co-culturing with M1-activated macrophages does not affect either fibroblasts phenotype or function. Hou et al. [
35] performed co-cultures of lung-resident mesenchymal stem cells (LR-MSCs), with M1 or M2 macrophages polarized, using a mouse macrophage cell line, RAW264.7. This study showed that co-culturing with M2 macrophages induces LR-MSC differentiation, evidenced by the increased α-SMA expression, while M1 macrophages do not alter LR-MSCs. Both studies suggested that M2-like macrophages, rather than M1-like macrophages, induce differentiation into myofibroblasts, because we observed that IM were activated towards an M2-like phenotype at this point in the disease model. Our results showed that the IM population acts as a fibrogenesis promotor in the early stages of BLM-induced pulmonary fibrosis. In contrast, they were able to promote fibroblast activation in vitro, probably through their activated anti-inflammatory and downregulated pro-inflammatory phenotype.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





