
MAP4K4/JNK Signaling Pathway Stimulates Proliferation and Suppresses Apoptosis of Human Spermatogonial Stem Cells and Lower Level of MAP4K4 Is Associated with Male Infertility

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Treatment of Human Testis Tissues
2.2. Culture of a Human SSC Line
2.3. The siRNA Transfection
2.4. RNA Extraction, RT-PCR, and qPCR
2.5. Immunocytochemistry and Immunofluorescence
2.6. Western Blots
2.7. CCK-8 Assay
2.8. EdU Incorporation Assay
2.9. Flow Cytometric Analysis
2.10. TUNEL Assay
2.11. RNA Sequencing
2.12. Identification of Cell Types and Analysis of Clusters
2.13. Statistical Analysis
3. Results
3.1. MAP4K4 was Highly Expressed in Human Spermatogonial Stem Cells
3.2. MAP4K4 Knockdown Inhibited Proliferation of Human SSCs
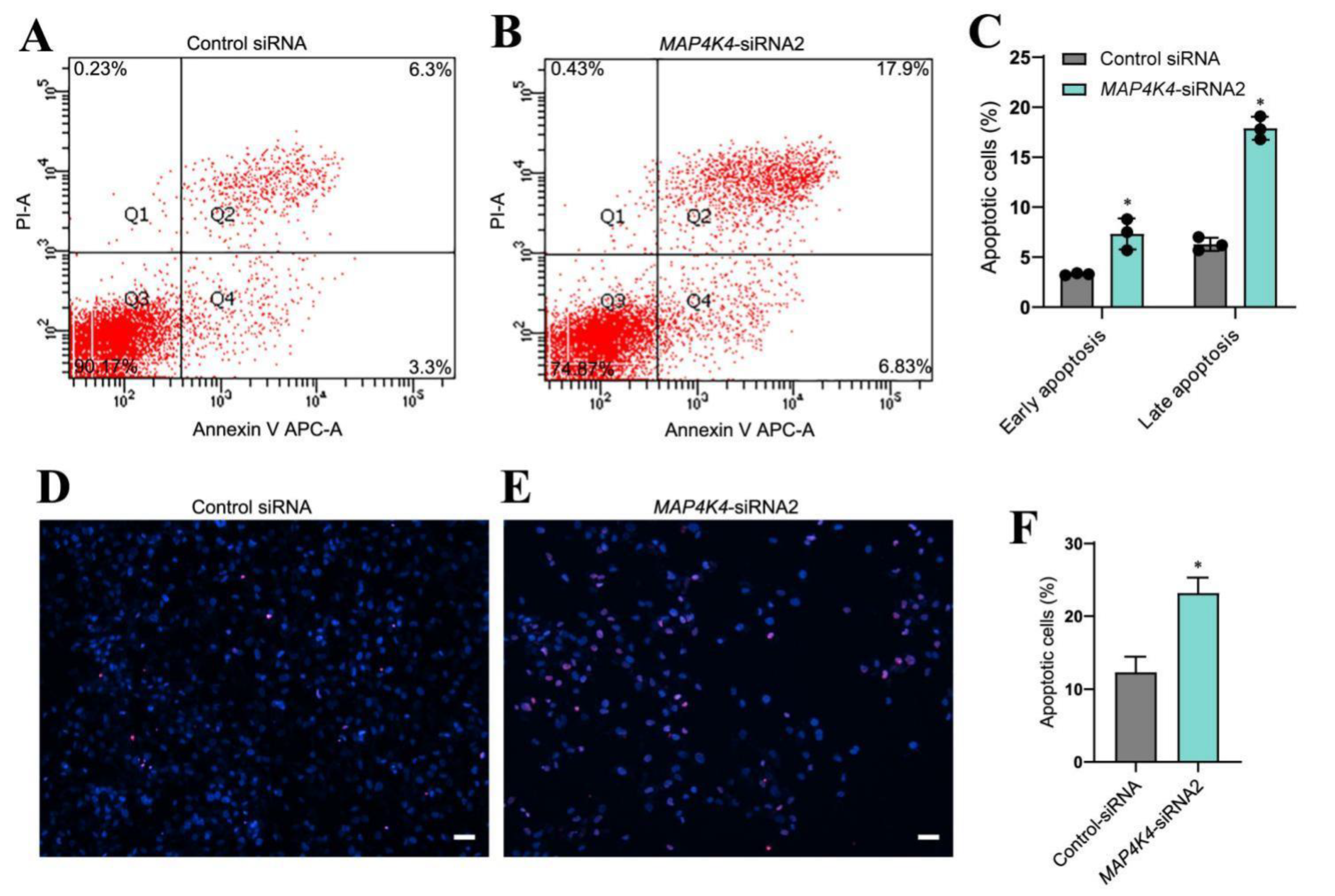
3.3. MAP4K4 knockdown Enhanced Apoptosis of Human SSCs
3.4. MAP4K4 Interacted with JNKs and Mediated JNK Phosphorylation in the Human SSC line
3.5. JNK Phosphorylation Inhibition Affected Proliferation and Apoptosis of the Human SSC Line
3.6. Screening of MAP4K4 Target Genes in Human SSCs
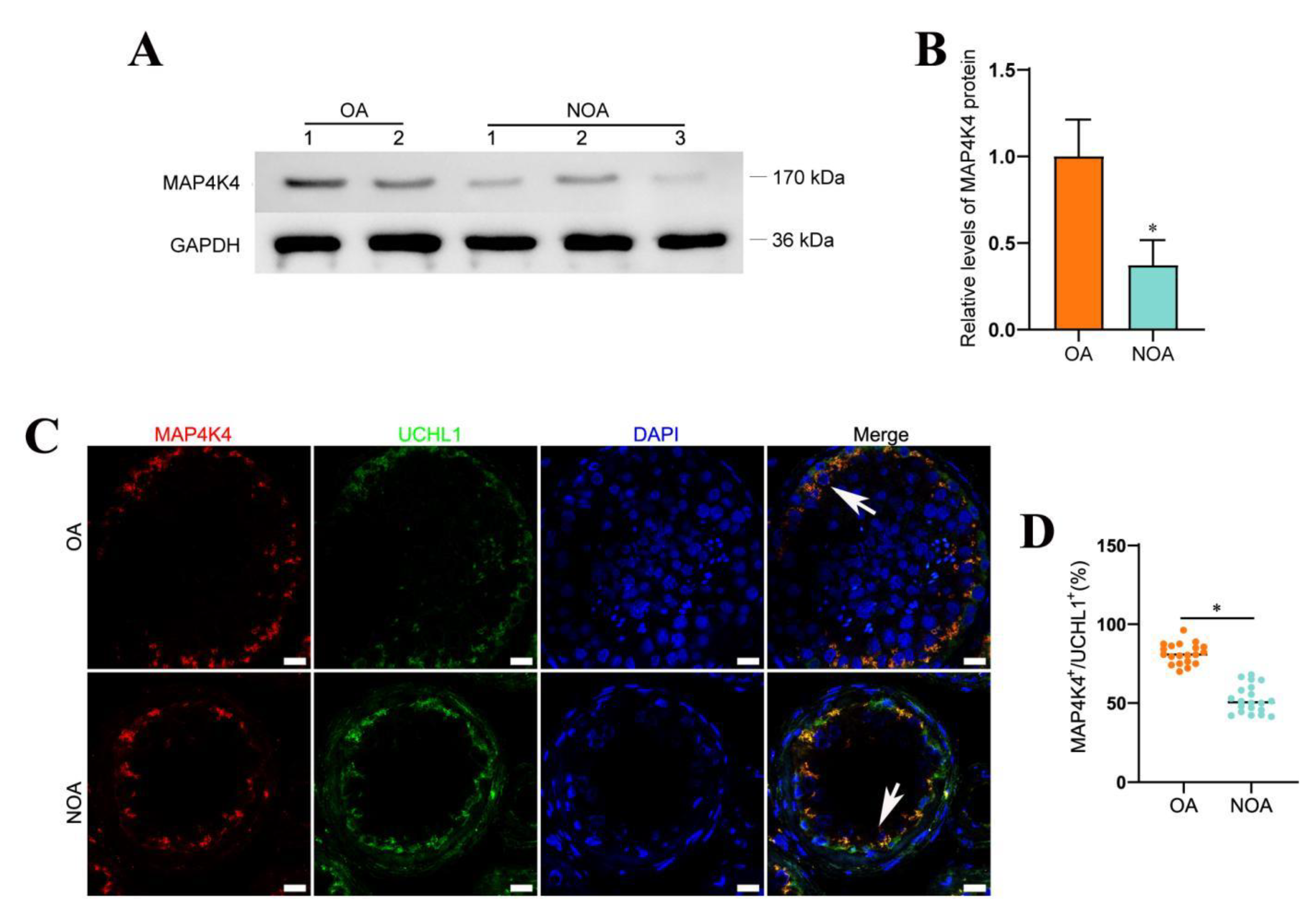
3.7. Decreased Expression Level of MAP4K4 in NOA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lord, T.; Oatley, J.M. A Revised Asingle Model to Explain Stem Cell Dynamics in the Mouse Male Germline. Reproduction 2017, 154, R55–R64. [Google Scholar] [CrossRef] [PubMed]
- Clermont, Y. Renewal of Spermatogonia in Man. Am. J. Anat. 1966, 118, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Simorangkir, D.R.; Marshall, G.R.; Plant, T.M. A Re-Examination of Proliferation and Differentiation of Type A Spermatogonia in the Adult Rhesus Monkey (Macaca mulatta). Hum. Reprod. 2009, 24, 1596–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrieri, C.; Comazzetto, S.; Grover, A.; Morgan, M.; Buness, A.; Nerlov, C.; O’Carroll, D. A Transit-Amplifying Population Underpins the Efficient Regenerative Capacity of the Testis. J. Exp. Med. 2017, 214, 1631–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial Stem Cells and Spermatogenesis in Mice, Monkeys and Men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Sachs, C.; Robinson, B.D.; Andres Martin, L.; Webster, T.; Gilbert, M.; Lo, H.-Y.; Rafii, S.; Ng, C.K.; Seandel, M. Evaluation of Candidate Spermatogonial Markers ID4 and GPR125 in Testes of Adult Human Cadaveric Organ Donors. Andrology 2014, 2, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, M.J.; Kaucher, A.V.; Racicot, K.E.; Oatley, J.M. Inhibitor of DNA Binding 4 Is Expressed Selectively by Single Spermatogonia in the Male Germline and Regulates the Self-Renewal of Spermatogonial Stem Cells in Mice1. Biol. Reprod. 2011, 85, 347–356. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Jiang, J.; Kokkinaki, M.; Golestaneh, N.; Hofmann, M.-C.; Dym, M. Gdnf Upregulates C-Fos Transcription via the Ras/Erk1/2 Pathway to Promote Mouse Spermatogonial Stem Cell Proliferation. Stem Cells 2008, 26, 266–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis Following Male Germ-Cell Transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Lindahl, M.; Hyvönen, M.E.; Parvinen, M.; de Rooij, D.G.; Hess, M.W.; Raatikainen-Ahokas, A.; Sainio, K.; Rauvala, H.; Lakso, M.; et al. Regulation of Cell Fate Decision of Undifferentiated Spermatogonia by GDNF. Science 2000, 287, 1489–1493. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Inoue, K.; Lee, J.; Yoshimoto, M.; Ogonuki, N.; Miki, H.; Baba, S.; Kato, T.; Kazuki, Y.; Toyokuni, S.; et al. Generation of Pluripotent Stem Cells from Neonatal Mouse Testis. Cell 2004, 119, 1001–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth Factors Essential for Self-Renewal and Expansion of Mouse Spermatogonial Stem Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oatley, J.M.; Avarbock, M.R.; Telaranta, A.I.; Fearon, D.T.; Brinster, R.L. Identifying Genes Important for Spermatogonial Stem Cell Self-Renewal and Survival. Proc. Natl. Acad. Sci. USA 2006, 103, 9524–9529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sada, A.; Hasegawa, K.; Pin, P.H.; Saga, Y. NANOS2 Acts Downstream of Glial Cell Line-Derived Neurotrophic Factor Signaling to Suppress Differentiation of Spermatogonial Stem Cells. Stem Cells 2012, 30, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Takashima, S.; Kanatsu-Shinohara, M.; Yi, Z.; Shinohara, T. Cdc42 Is Required for Male Germline Niche Development in Mice. Cell Rep. 2021, 36, 109550. [Google Scholar] [CrossRef] [PubMed]
- Sohni, A.; Tan, K.; Song, H.-W.; Burow, D.; de Rooij, D.G.; Laurent, L.; Hsieh, T.-C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517. [Google Scholar] [CrossRef] [Green Version]
- Rebourcet, D.; Wu, J.; Cruickshanks, L.; Smith, S.E.; Milne, L.; Fernando, A.; Wallace, R.J.; Gray, C.D.; Hadoke, P.W.F.; Mitchell, R.T.; et al. Sertoli Cells Modulate Testicular Vascular Network Development, Structure, and Function to Influence Circulating Testosterone Concentrations in Adult Male Mice. Endocrinology 2016, 157, 2479–2488. [Google Scholar] [CrossRef] [Green Version]
- Soffientini, U.; Rebourcet, D.; Abel, M.H.; Lee, S.; Hamilton, G.; Fowler, P.A.; Smith, L.B.; O’Shaughnessy, P.J. Identification of Sertoli Cell-Specific Transcripts in the Mouse Testis and the Role of FSH and Androgen in the Control of Sertoli Cell Activity. BMC Genom. 2017, 18, 972. [Google Scholar] [CrossRef] [Green Version]
- Cooke, H.J.; Saunders, P.T.K. Mouse Models of Male Infertility. Nat. Rev. Genet. 2002, 3, 790–801. [Google Scholar] [CrossRef]
- Sharma, M.; Braun, R.E. Cyclical Expression of GDNF Is Required for Spermatogonial Stem Cell Homeostasis. Development 2018, 145, dev.151555. [Google Scholar] [CrossRef]
- Ishii, K.; Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. FGF2 Mediates Mouse Spermatogonial Stem Cell Self-Renewal via Upregulation of Etv5 and Bcl6b through MAP2K1 Activation. Development 2012, 139, 1734–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Yuan, Q.; Niu, M.; Wang, H.; Wen, L.; Yao, C.; Hou, J.; Chen, Z.; Fu, H.; Zhou, F.; et al. Efficient Generation of Functional Haploid Spermatids from Human Germline Stem Cells by Three-Dimensional-Induced System. Cell Death Differ. 2018, 25, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Niu, M.; Liu, L.; Zhu, Z.; Wang, X.; Sun, M.; Yuan, Q.; Yang, S.; Zeng, W.; Liu, Y.; et al. Establishment and Characterization of Human Germline Stem Cell Line with Unlimited Proliferation Potentials and No Tumor Formation. Sci. Rep. 2015, 5, 16922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Cui, Y.; Liu, B.; Li, C.; Du, L.; Tang, R.; Qin, L.; Jiang, Y.; Li, J.; Yu, X.; et al. Hsa-MiR-1908-3p Mediates the Self-Renewal and Apoptosis of Human Spermatogonial Stem Cells via Targeting KLF2. Mol. Ther.-Nucleic Acids 2020, 20, 788–800. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.S.; Hong, J.; Sapinoso, L.; Zhou, Y.; Liu, Z.; Micklash, K.; Schultz, P.G.; Hampton, G.M. A Small Interfering RNA Screen for Modulators of Tumor Cell Motility Identifies MAP4K4 as a Promigratory Kinase. Proc. Natl. Acad. Sci. USA 2006, 103, 3775–3780. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Guilherme, A.; Chakladar, A.; Powelka, A.M.; Konda, S.; Virbasius, J.V.; Nicoloro, S.M.C.; Straubhaar, J.; Czech, M.P. An RNA Interference-Based Screen Identifies MAP4K4/NIK as a Negative Regulator of PPARgamma, Adipogenesis, and Insulin-Responsive Hexose Transport. Proc. Natl. Acad. Sci. USA 2006, 103, 2087–2092. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, M.V.; Azimzadeh, O.; Merl-Pham, J.; Verreet, T.; Hauck, S.M.; Benotmane, M.A.; Atkinson, M.J.; Tapio, S. In-Utero Low-Dose Irradiation Leads to Persistent Alterations in the Mouse Heart Proteome. PLoS ONE 2016, 11, e0156952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafat, M.; Har-Vardi, I.; Harlev, A.; Levitas, E.; Zeadna, A.; Abofoul-Azab, M.; Dyomin, V.; Sheffield, V.C.; Lunenfeld, E.; Huleihel, M.; et al. Mutation in TDRD9 Causes Non-Obstructive Azoospermia in Infertile Men. J. Med. Genet. 2017, 54, 633–639. [Google Scholar] [CrossRef]
- Cioppi, F.; Rosta, V.; Krausz, C. Genetics of Azoospermia. Int. J. Mol. Sci. 2021, 22, 3264. [Google Scholar] [CrossRef]
- Du, L.; Chen, W.; Li, C.; Cui, Y.; He, Z. RNF144B Stimulates the Proliferation and Inhibits the Apoptosis of Human Spermatogonial Stem Cells via the FCER2/NOTCH2/HES1 Pathway and Its Abnormality Is Associated with Azoospermia. J. Cell. Physiol. 2022, 237, 3565–3577. [Google Scholar] [CrossRef]
- Niu, L.; Fang, Y.; Yao, X.; Zhang, Y.; Wu, J.; Chen, D.F.; Sun, X. TNFα Activates MAPK and Jak-Stat Pathways to Promote Mouse Müller Cell Proliferation. Exp. Eye Res. 2021, 202, 108353. [Google Scholar] [CrossRef]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic Review of Patient Factors Affecting Adipose Stem Cell Viability and Function: Implications for Regenerative Therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.-W.; Cai, J.; Zhao, X.-L.; Jiang, T.-H.; He, T.-F.; Fu, H.-Q.; Zhu, M.-H.; Zhang, S. ShRNA-Targeted MAP4K4 Inhibits Hepatocellular Carcinoma Growth. Clin. Cancer Res. 2011, 17, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, X.; Lu, D.; Xu, Y.; Mou, W.; Wang, L.; Chen, Y.; Liu, Y.; Li, X.; Li, L.-Y.; et al. SOX2 Regulates Apoptosis through MAP4K4-Survivin Signaling Pathway in Human Lung Cancer Cells. Carcinogenesis 2014, 35, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Wang, B.; Liu, Y.; Zhang, J.; Deng, S.; Qin, Q.; Tian, K.; Li, X.; Zhu, S.; Niu, Y.; et al. MiRNA-141, Downregulated in Pancreatic Cancer, Inhibits Cell Proliferation and Invasion by Directly Targeting MAP4K4. Mol. Cancer Ther. 2013, 12, 2569–2580. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, L.R.; Chapman, K.; Xie, M.; Maifoshie, E.; Jenkins, M.; Golforoush, P.A.; Bellahcene, M.; Noseda, M.; Faust, D.; Jarvis, A.; et al. MAP4K4 Inhibition Promotes Survival of Human Stem Cell-Derived Cardiomyocytes and Reduces Infarct Size In Vivo. Cell Stem Cell 2019, 24, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Watts, M.E.; Rubin, L.L. MAP4K4 Activation Mediates Motor Neuron Degeneration in Amyotrophic Lateral Sclerosis. Cell Rep. 2019, 26, 1143–1156. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, S.; Mana-Capelli, S.; Roth Flach, R.J.; Danai, L.V.; Amcheslavsky, A.; Nie, Y.; Kaneko, S.; Yao, X.; Chen, X.; et al. The Conserved Misshapen-Warts-Yorkie Pathway Acts in Enteroblasts to Regulate Intestinal Stem Cells in Drosophila. Dev. Cell 2014, 31, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Qiu, Y.; Lin, K.C.; Kumar, A.; Placone, J.K.; Fang, C.; Wang, K.-C.; Lu, S.; Pan, M.; Hong, A.W.; et al. RAP2 Mediates Mechanoresponses of the Hippo Pathway. Nature 2018, 560, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Gupta, S.K.; Kim, K.J.; Powers, B.E.; Cerqueira, A.; Wainger, B.J.; Ngo, H.D.; Rosowski, K.A.; Schein, P.A.; Ackeifi, C.A.; et al. A Small Molecule Screen in Stem-Cell-Derived Motor Neurons Identifies a Kinase Inhibitor as a Candidate Therapeutic for ALS. Cell Stem Cell 2013, 12, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Wang, X.; Li, Z.; Gotoh, N.; Chapman, D.; Skolnik, E.Y. Mesodermal Patterning Defect in Mice Lacking the Ste20 NCK Interacting Kinase (NIK). Dev. Camb. Engl. 2001, 128, 1559–1572. [Google Scholar] [CrossRef]
- Vora, N.L.; Powell, B.; Brandt, A.; Strande, N.; Hardisty, E.; Gilmore, K.; Foreman, A.K.M.; Wilhelmsen, K.; Bizon, C.; Reilly, J.; et al. Prenatal Exome Sequencing in Anomalous Fetuses: New Opportunities and Challenges. Genet. Med. 2017, 19, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.-C. NIK Is a New Ste20-Related Kinase That Binds NCK and MEKK1 and Activates the SAPK/JNK Cascade via a Conserved Regulatory Domain. EMBO J. 1997, 16, 1279–1290. [Google Scholar] [CrossRef]
- Yao, Z.; Zhou, G.; Wang, X.S.; Brown, A.; Diener, K.; Gan, H.; Tan, T.-H. A Novel Human STE20-Related Protein Kinase, HGK, That Specifically Activates the c-Jun N-Terminal Kinase Signaling Pathway. J. Biol. Chem. 1999, 274, 2118–2125. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zheng, Y.; Gao, Y.; Lin, Z.; Yang, S.; Wang, T.; Wang, Q.; Xie, N.; Hua, R.; Liu, M.; et al. Single-Cell RNA-Seq Uncovers Dynamic Processes and Critical Regulators in Mouse Spermatogenesis. Cell Res. 2018, 28, 879–896. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. ROS Are Required for Mouse Spermatogonial Stem Cell Self-Renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Wertz, K.; Herrmann, B.G. Large-Scale Screen for Genes Involved in Gonad Development. Mech. Dev. 2000, 98, 51–70. [Google Scholar] [CrossRef]
- Vernon, R.B.; Sage, H. The Calcium-Binding Protein SPARC Is Secreted by Leydig and Sertoli Cells of the Adult Mouse Testis1. Biol. Reprod. 1989, 40, 1329–1340. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, M.C.; Schwarzbauer, J.E. Importance of the Basement Membrane Protein SPARC for Viability and Fertility in Caenorhabditis Elegans. Curr. Biol. 1998, 8, 1285–1288. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, L.; Li, C.; Wu, J.; Zhang, Y.; Pang, K.; Wei, Y.; Du, L.; Liu, M.; Wu, X. SPARC Promotes Self-Renewal of Limbal Epithelial Stem Cells and Ocular Surface Restoration through JNK and P38-MAPK Signaling Pathways. Stem Cells 2020, 38, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Mao, M.; Thedens, D.R.; Chang, B.; Harris, B.S.; Zheng, Q.Y.; Johnson, K.R.; Donahue, L.R.; Anderson, M.G. The Podosomal-Adaptor Protein SH3PXD2B Is Essential for Normal Postnatal Development. Mamm. Genome 2009, 20, 462–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.; O’Dell Bunch, D.; Faure, J.-E.; Goulding, E.H.; Eddy, E.M.; Primakoff, P.; Myles, D.G. Fertilization Defects in Sperm from Mice Lacking Fertilin β. Science 1998, 281, 1857–1859. [Google Scholar] [CrossRef] [PubMed]
- Blobel, C.P.; Wolfsberg, T.G.; Turck, C.W.; Myles, D.G.; Primakoff, P.; White, J.M. A Potential Fusion Peptide and an Integrin Ligand Domain in a Protein Active in Sperm–Egg Fusion. Nature 1992, 356, 248–252. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.T. MAPK Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.; Allen, W.E.; Sawyer, C.; Vanhaesebroeck, B.; Trimble, E.R. Glucose-Potentiated Chemotaxis in Human Vascular Smooth Muscle Is Dependent on Cross-Talk Between the PI3K and MAPK Signaling Pathways. Circ. Res. 2004, 95, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Tolcher, A.W.; Peng, W.; Calvo, E. Rational Approaches for Combination Therapy Strategies Targeting the MAP Kinase Pathway in Solid Tumors. Mol. Cancer Ther. 2018, 17, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Liu, Y.; Kaneto, H.; Fanburg, B.L. JNK Regulates Serotonin-Mediated Proliferation and Migration of Pulmonary Artery Smooth Muscle Cells. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2010, 298, L863–L869. [Google Scholar] [CrossRef] [Green Version]
- Vlotides, G.; Cruz-Soto, M.; Rubinek, T.; Eigler, T.; Auernhammer, C.J.; Melmed, S. Mechanisms for Growth Factor-Induced Pituitary Tumor Transforming Gene-1 Expression in Pituitary Folliculostellate TtT/GF Cells. Mol. Endocrinol. 2006, 20, 3321–3335. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Zhang, W.; Yuan, Q.; Niu, M.; Zhou, F.; Qiu, Q.; Mao, G.; Wang, H.; Wen, L.; Sun, M.; et al. PAK1 Promotes the Proliferation and Inhibits Apoptosis of Human Spermatogonial Stem Cells via PDK1/KDR/ZNF367 and ERK1/2 and AKT Pathways. Mol. Ther.-Nucleic Acids 2018, 12, 769–786. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.-Y.; Liang, Y.-C.; Tan, T.-H.; Chuang, H.-C.; Lin, Y.-J.; Lin, J.-C. RBM4a-SRSF3-MAP4K4 Splicing Cascade Constitutes a Molecular Mechanism for Regulating Brown Adipogenesis. Int. J. Mol. Sci. 2018, 19, 2646. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, C.; Chen, W.; Cui, Y.; He, Z. MAP4K4/JNK Signaling Pathway Stimulates Proliferation and Suppresses Apoptosis of Human Spermatogonial Stem Cells and Lower Level of MAP4K4 Is Associated with Male Infertility. Cells 2022, 11, 3807. https://doi.org/10.3390/cells11233807
Wan C, Chen W, Cui Y, He Z. MAP4K4/JNK Signaling Pathway Stimulates Proliferation and Suppresses Apoptosis of Human Spermatogonial Stem Cells and Lower Level of MAP4K4 Is Associated with Male Infertility. Cells. 2022; 11(23):3807. https://doi.org/10.3390/cells11233807
Chicago/Turabian StyleWan, Cailin, Wei Chen, Yinghong Cui, and Zuping He. 2022. "MAP4K4/JNK Signaling Pathway Stimulates Proliferation and Suppresses Apoptosis of Human Spermatogonial Stem Cells and Lower Level of MAP4K4 Is Associated with Male Infertility" Cells 11, no. 23: 3807. https://doi.org/10.3390/cells11233807