
ERK Phosphorylation Regulates the Aml1/Runx1 Splice Variants and the TRP Channels Expression during the Differentiation of Glioma Stem Cell Lines

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Antibodies
2.3. Western Blot Analysis
2.4. Cell Death Analysis
2.5. Confocal Laser Scanning Microscopy Analysis
2.6. Gene Expression Analysis
2.7. qRT-PCR and PCR Array
2.8. Statistical Analysis
3. Results
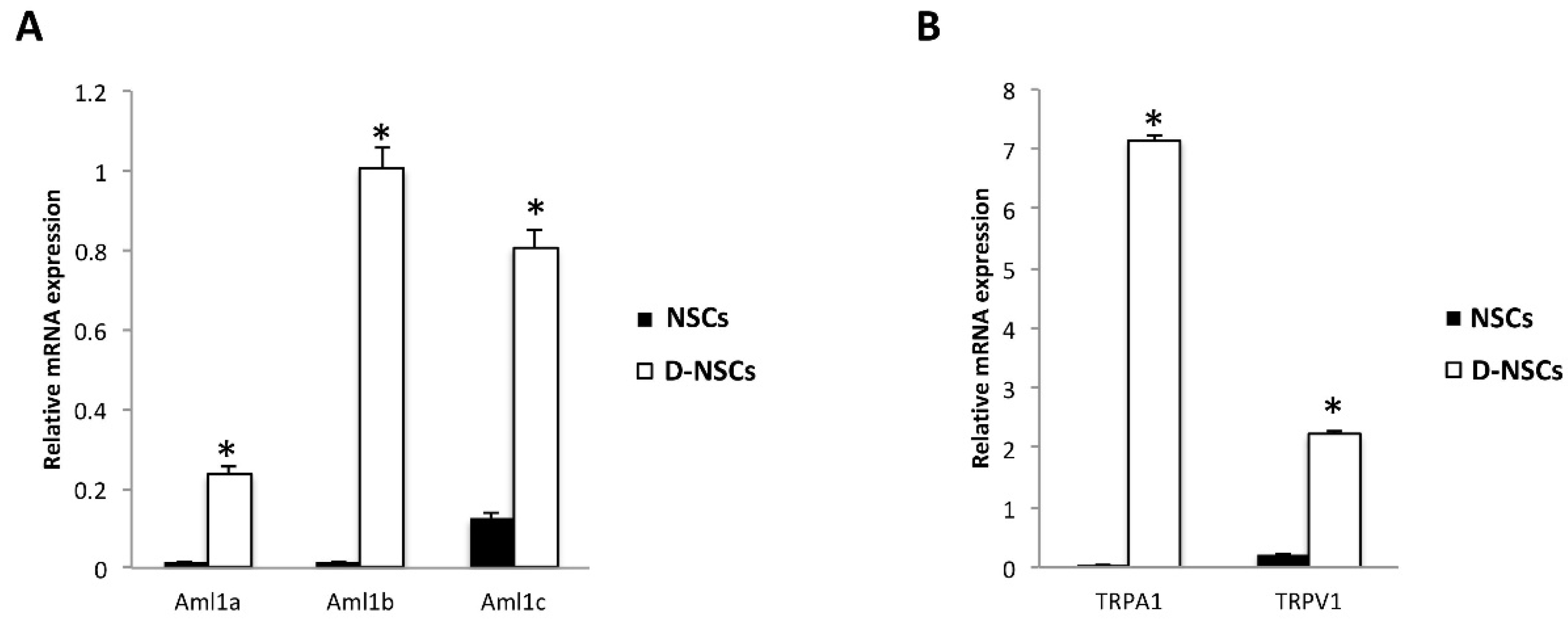
3.1. Expression of the Aml1 Spliced Variants and TRP Channels in NSC and D-NSC
3.2. Overexpression of Aml1b and Aml1c Variants and TRP Channels in Distinct PN- and MES-like GSCs Compared to NSCs
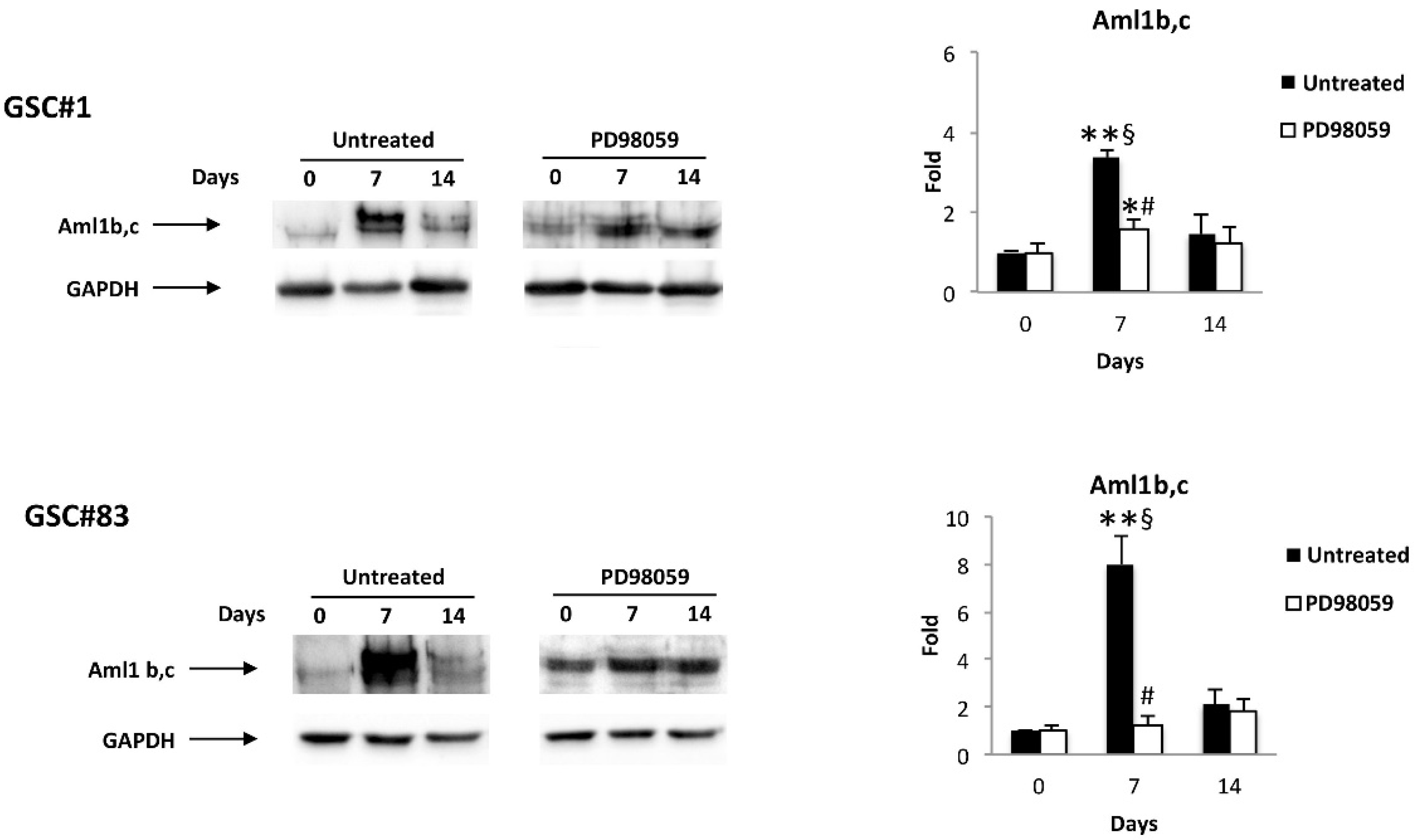
3.3. Differentiation of GSC Lines Increases the Aml1b/Aml1c and TRP Ion Channel mRNA Levels
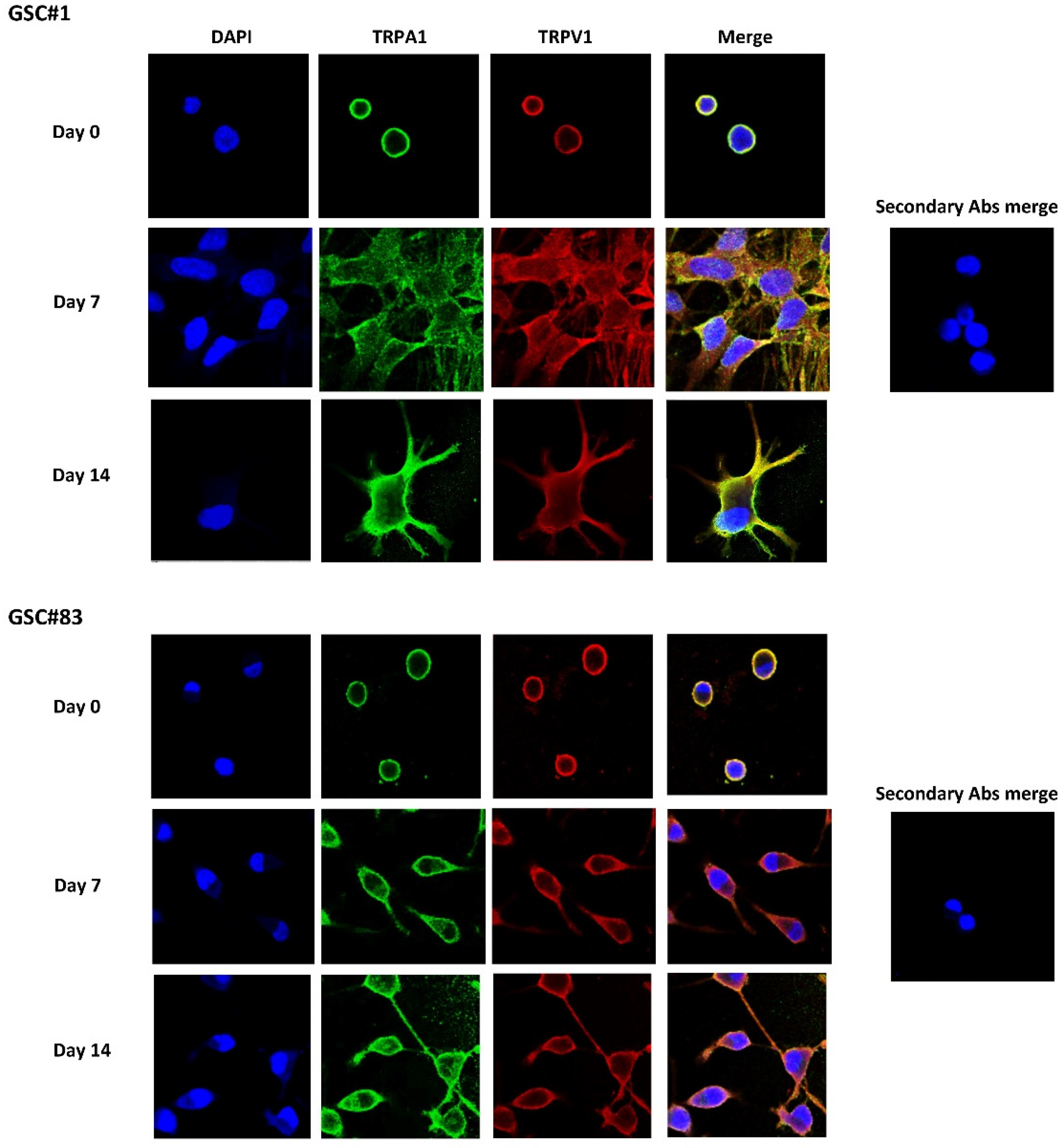
3.4. TRPA1 and TRPV1 Channel Expression in PN- and MES-like GSCs and D-GSC Lines
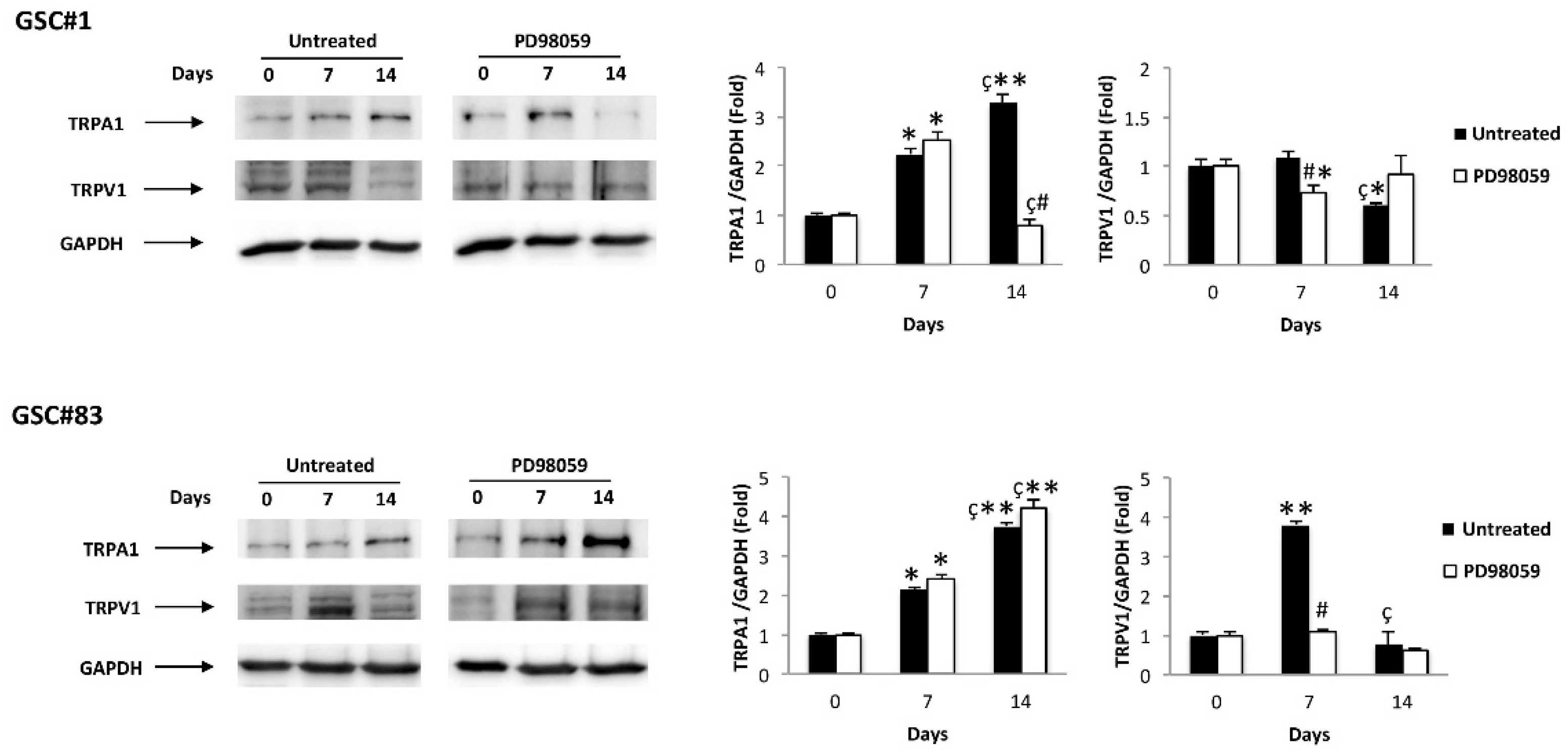
3.5. ERK Phosphorylation Increases the Aml1b and Aml1c Protein Expression and Induces Changes in TRP Channel Expression during GSC Differentiation
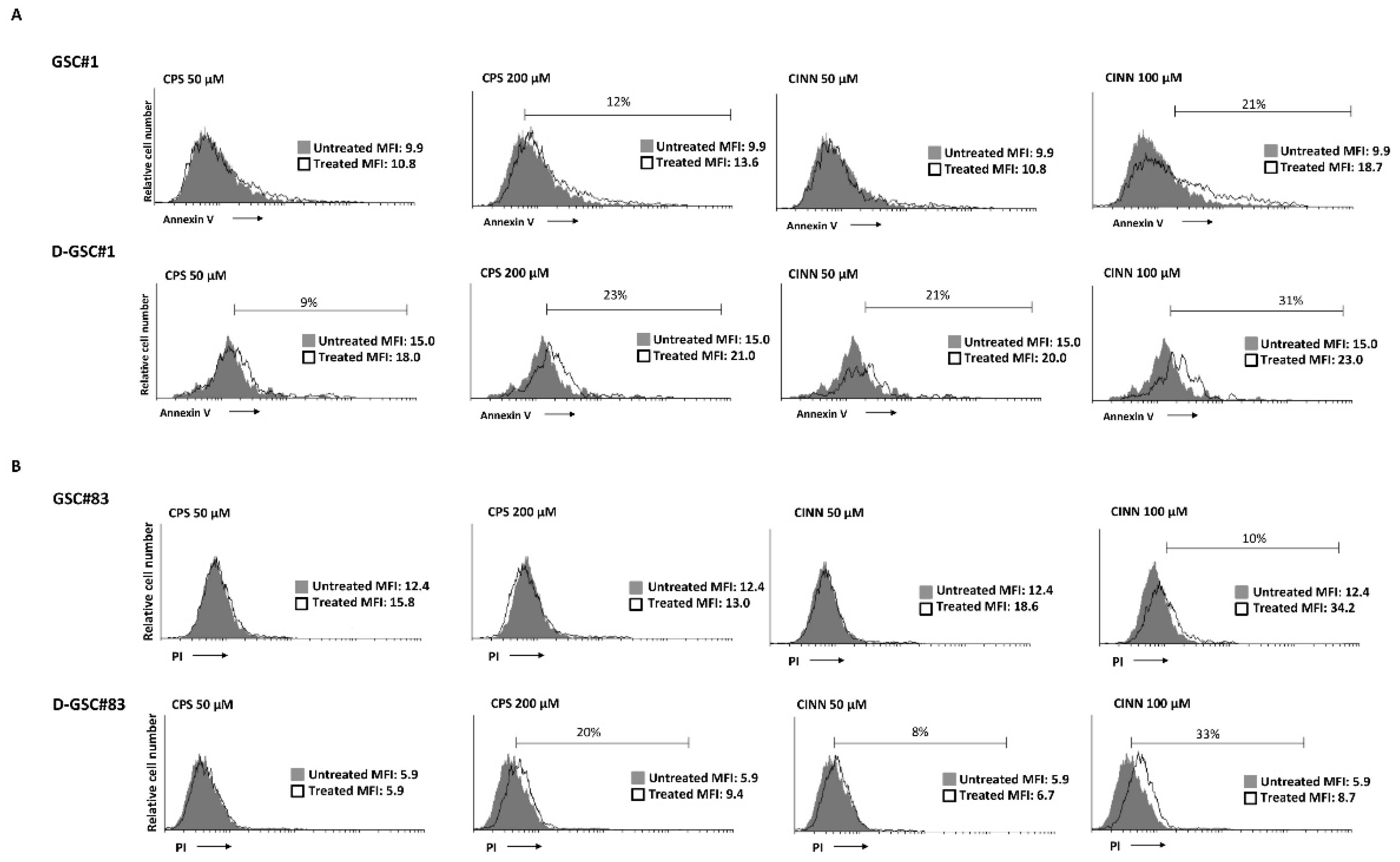
3.6. Specific TRPA1 and TRPV1 Agonists Stimulate Apoptotic/Necrotic Cell Death in PN- and MES-GSCs and D-GSCs
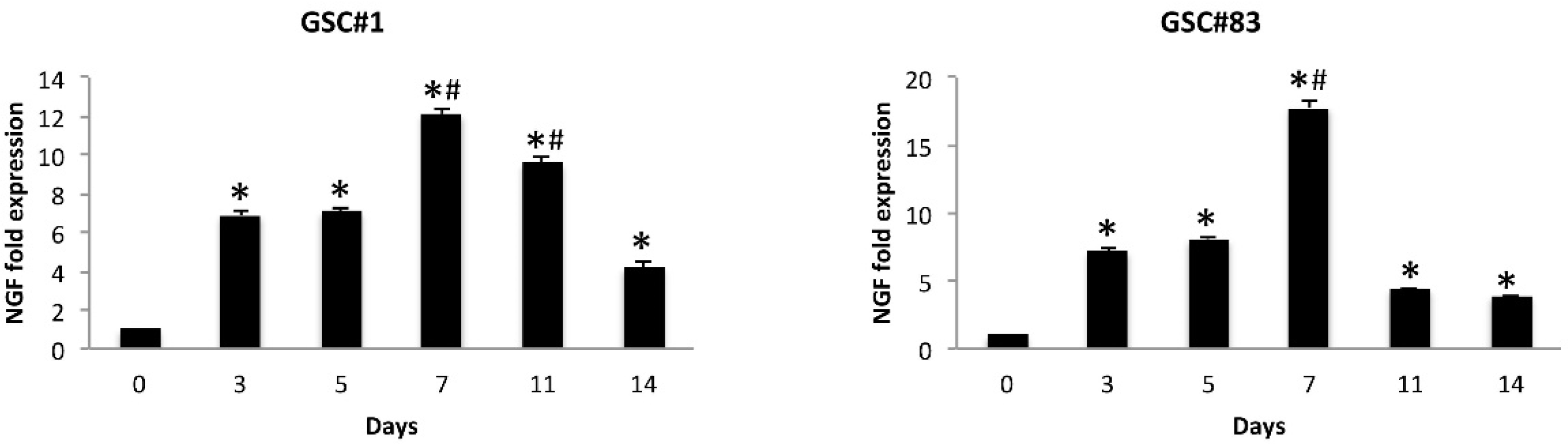
3.7. NGF Levels Increase during GSC Differentiation
3.8. GSC Differentiation Is Accompanied by Changes of Gene Pathways Involved in GSC Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maher, E.A.; Furnari, F.B.; Bachoo, R.M.; Rowitch, D.H.; Louis, D.N.; Cavenee, W.K.; DePinho, R.A. Malignant Glioma: Genetics and Biology of a Grave Matter. Genes Dev. 2001, 15, 1311–1333. [Google Scholar] [CrossRef] [Green Version]
- Sanai, N.; Alvarez-Buylla, A.; Berger, M.S. Neural Stem Cells and the Origin of Gliomas. N. Engl. J. Med. 2005, 353, 811–822. [Google Scholar] [CrossRef]
- Vescovi, A.L.; Galli, R.; Reynolds, B.A. Brain Tumour Stem Cells. Nat. Rev. Cancer 2006, 6, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, T.N.; Kukekov, V.G.; Laywell, E.D.; Suslov, O.N.; Vrionis, F.D.; Steindler, D.A. Human Cortical Glial Tumors Contain Neural Stem-like Cells Expressing Astroglial and Neuronal Markers in Vitro. Glia 2002, 39, 193–206. [Google Scholar] [CrossRef]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous Stem Cells Can Arise from Pediatric Brain Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [Green Version]
- Galli, R.; Binda, E.; Orfanelli, U.; Cipelletti, B.; Gritti, A.; De Vitis, S.; Fiocco, R.; Foroni, C.; Dimeco, F.; Vescovi, A. Isolation and Characterization of Tumorigenic, Stem-like Neural Precursors from Human Glioblastoma. Cancer Res. 2004, 64, 7011–7021. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a Cancer Stem Cell in Human Brain Tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of Human Brain Tumour Initiating Cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Kondo, T.; Setoguchi, T.; Taga, T. Persistence of a Small Subpopulation of Cancer Stem-like Cells in the C6 Glioma Cell Line. Proc. Natl. Acad. Sci. USA 2004, 101, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrawala, L.; Calhoun, T.; Schneider-Broussard, R.; Zhou, J.; Claypool, K.; Tang, D.G. Side Population Is Enriched in Tumorigenic, Stem-like Cancer Cells, Whereas ABCG2+ and ABCG2− Cancer Cells Are Similarly Tumorigenic. Cancer Res. 2005, 65, 6207–6219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirks, P.B. Cancer: Stem Cells and Brain Tumours. Nature 2006, 444, 687–688. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Wan, F.; Farhadi, M.; Ernst, A.; Zeppernick, F.; Tagscherer, K.E.; Ahmadi, R.; Lohr, J.; Dictus, C.; Gdynia, G.; et al. Differentiation Therapy Exerts Antitumor Effects on Stem-like Glioma Cells. Clin. Cancer Res. 2010, 16, 2715–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lathia, J.D.; Mack, S.C.; Mulkearns-Hubert, E.E.; Valentim, C.L.L.; Rich, J.N. Cancer Stem Cells in Glioblastoma. Genes Dev. 2015, 29, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Ricci-Vitiani, L.; Pallini, R.; Biffoni, M.; Todaro, M.; Invernici, G.; Cenci, T.; Maira, G.; Parati, E.A.; Stassi, G.; Larocca, L.M.; et al. Tumour Vascularization via Endothelial Differentiation of Glioblastoma Stem-like Cells. Nature 2010, 468, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Steponaitis, G.; Tamasauskas, A. Mesenchymal and Proneural Subtypes of Glioblastoma Disclose Branching Based on GSC Associated Signature. Int. J. Mol. Sci. 2021, 22, 4964. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, T.; Hildebrandt, J.; Nuebling, G.; Sostak, P.; Straube, A. Glioblastoma-Dependent Differentiation and Angiogenic Potential of Human Mesenchymal Stem Cells in Vitro. J. Neurooncol. 2011, 105, 57–65. [Google Scholar] [CrossRef]
- Mao, P.; Joshi, K.; Li, J.; Kim, S.-H.; Li, P.; Santana-Santos, L.; Luthra, S.; Chandran, U.R.; Benos, P.V.; Smith, L.; et al. Mesenchymal Glioma Stem Cells Are Maintained by Activated Glycolytic Metabolism Involving Aldehyde Dehydrogenase 1A3. Proc. Natl. Acad. Sci. USA 2013, 110, 8644–8649. [Google Scholar] [CrossRef] [Green Version]
- Cooper, L.A.D.; Gutman, D.A.; Chisolm, C.; Appin, C.; Kong, J.; Rong, Y.; Kurc, T.; Van Meir, E.G.; Saltz, J.H.; Moreno, C.S.; et al. The Tumor Microenvironment Strongly Impacts Master Transcriptional Regulators and Gene Expression Class of Glioblastoma. Am. J. Pathol. 2012, 180, 2108–2119. [Google Scholar] [CrossRef] [Green Version]
- Sumazin, P.; Yang, X.; Chiu, H.-S.; Chung, W.-J.; Iyer, A.; Llobet-Navas, D.; Rajbhandari, P.; Bansal, M.; Guarnieri, P.; Silva, J.; et al. An Extensive MicroRNA-Mediated Network of RNA-RNA Interactions Regulates Established Oncogenic Pathways in Glioblastoma. Cell 2011, 147, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Chuang, L.S.H.; Ito, K.; Ito, Y. RUNX Family: Regulation and Diversification of Roles through Interacting Proteins. Int. J. Cancer 2013, 132, 1260–1271. [Google Scholar] [CrossRef]
- Chen, C.-L.; Broom, D.C.; Liu, Y.; de Nooij, J.C.; Li, Z.; Cen, C.; Samad, O.A.; Jessell, T.M.; Woolf, C.J.; Ma, Q. Runx1 Determines Nociceptive Sensory Neuron Phenotype and Is Required for Thermal and Neuropathic Pain. Neuron 2006, 49, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugarte, G.D.; Diaz, E.; Biscaia, M.; Stehberg, J.; Montecino, M.; van Zundert, B. Transcription of the Pain-Related TRPV1 Gene Requires Runx1 and C/EBPβ Factors. J. Cell. Physiol. 2013, 228, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Kamikubo, Y.; Liu, P. Role of RUNX1 in Hematological Malignancies. Blood 2017, 129, 2070–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, N.; Mohammed, Z.M.A.; Nixon, C.; Mason, S.M.; Mallon, E.; McMillan, D.C.; Morris, J.S.; Cameron, E.R.; Edwards, J.; Blyth, K. Expression of RUNX1 Correlates with Poor Patient Prognosis in Triple Negative Breast Cancer. PLoS ONE 2014, 9, e100759. [Google Scholar] [CrossRef]
- Dulak, A.M.; Schumacher, S.E.; Van Lieshout, J.; Imamura, Y.; Fox, C.; Shim, B.; Ramos, A.H.; Saksena, G.; Baca, S.C.; Baselga, J.; et al. Gastrointestinal Adenocarcinomas of the Esophagus, Stomach, and Colon Exhibit Distinct Patterns of Genome Instability and Oncogenesis. Cancer Res. 2012, 72, 4383–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planagumà, J.; Díaz-Fuertes, M.; Gil-Moreno, A.; Abal, M.; Monge, M.; García, A.; Baró, T.; Thomson, T.M.; Xercavins, J.; Alameda, F.; et al. A Differential Gene Expression Profile Reveals Overexpression of RUNX1/AML1 in Invasive Endometrioid Carcinoma. Cancer Res. 2004, 64, 8846–8853. [Google Scholar] [CrossRef] [Green Version]
- Sakakura, C.; Hagiwara, A.; Miyagawa, K.; Nakashima, S.; Yoshikawa, T.; Kin, S.; Nakase, Y.; Ito, K.; Yamagishi, H.; Yazumi, S.; et al. Frequent Downregulation of the Runt Domain Transcription Factors RUNX1, RUNX3 and Their Cofactor CBFB in Gastric Cancer. Int. J. Cancer 2005, 113, 221–228. [Google Scholar] [CrossRef]
- Nabissi, M.; Morelli, M.B.; Amantini, C.; Liberati, S.; Santoni, M.; Ricci-Vitiani, L.; Pallini, R.; Santoni, G. Cannabidiol Stimulates Aml-1a-Dependent Glial Differentiation and Inhibits Glioma Stem-like Cells Proliferation by Inducing Autophagy in a TRPV2-Dependent Manner. Int. J. Cancer 2015, 137, 1855–1869. [Google Scholar] [CrossRef] [Green Version]
- Kleihues, P.; Cavenee, W.K. Pathology & Genetics. Tumours of the Nervous System. In World Health Organisation Classification of Tumours; IARC Press: Edinburgh, UK, 2000; p. 314. [Google Scholar]
- Visconti, P.; Parodi, F.; Parodi, B.; Casarino, L.; Romano, P.; Buccarelli, M.; Pallini, R.; D’Alessandris, Q.G.; Montori, A.; Pilozzi, E.; et al. Short Tandem Repeat Profiling for the Authentication of Cancer Stem-like Cells. Int. J. Cancer 2021, 148, 1489–1498. [Google Scholar] [CrossRef]
- Morelli, M.B.; Nabissi, M.; Amantini, C.; Farfariello, V.; Ricci-Vitiani, L.; di Martino, S.; Pallini, R.; Larocca, L.M.; Caprodossi, S.; Santoni, M.; et al. The Transient Receptor Potential Vanilloid-2 Cation Channel Impairs Glioblastoma Stem-like Cell Proliferation and Promotes Differentiation. Int. J. Cancer 2012, 131, E1067–E1077. [Google Scholar] [CrossRef] [Green Version]
- Günther, H.S.; Schmidt, N.O.; Phillips, H.S.; Kemming, D.; Kharbanda, S.; Soriano, R.; Modrusan, Z.; Meissner, H.; Westphal, M.; Lamszus, K. Glioblastoma-Derived Stem Cell-Enriched Cultures Form Distinct Subgroups According to Molecular and Phenotypic Criteria. Oncogene 2008, 27, 2897–2909. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Mattar, P.; Dixit, R.; Lawn, S.O.; Wilkinson, G.; Kinch, C.; Eisenstat, D.; Kurrasch, D.M.; Chan, J.A.; Schuurmans, C. RAS/ERK Signaling Controls Proneural Genetic Programs in Cortical Development and Gliomagenesis. J. Neurosci. 2014, 34, 2169–2190. [Google Scholar] [CrossRef]
- Cheng, P.; Alberts, I.; Li, X. The Role of ERK1/2 in the Regulation of Proliferation and Differentiation of Astrocytes in Developing Brain. Int. J. Dev. Neurosci. 2013, 31, 783–789. [Google Scholar] [CrossRef]
- Kwon, S.-J.; Kwon, O.-S.; Kim, K.-T.; Go, Y.-H.; Yu, S.; Lee, B.; Miyoshi, H.; Oh, E.; Cho, S.-J.; Cha, H.-J. Role of MEK Partner-1 in Cancer Stemness through MEK/ERK Pathway in Cancerous Neural Stem Cells, Expressing EGFRviii. Mol. Cancer 2017, 16, 140. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious Cold Ion Channel TRPA1 Is Activated by Pungent Compounds and Bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Klesse, L.J.; Meyers, K.A.; Marshall, C.J.; Parada, L.F. Nerve Growth Factor Induces Survival and Differentiation through Two Distinct Signaling Cascades in PC12 Cells. Oncogene 1999, 18, 2055–2068. [Google Scholar] [CrossRef] [Green Version]
- Vaudry, D.; Stork, P.J.S.; Lazarovici, P.; Eiden, L.E. Signaling Pathways for PC12 Cell Differentiation: Making the Right Connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef]
- Holland, E.C. Progenitor Cells and Glioma Formation. Curr. Opin. Neurol. 2001, 14, 683–688. [Google Scholar] [CrossRef]
- Michaud, J.; Simpson, K.M.; Escher, R.; Buchet-Poyau, K.; Beissbarth, T.; Carmichael, C.; Ritchie, M.E.; Schütz, F.; Cannon, P.; Liu, M.; et al. Integrative Analysis of RUNX1 Downstream Pathways and Target Genes. BMC Genom. 2008, 9, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffman, J.A. Runx Transcription Factors and the Developmental Balance between Cell Proliferation and Differentiation. Cell Biol. Int. 2003, 27, 315–324. [Google Scholar] [CrossRef]
- Hong, D.; Fritz, A.J.; Gordon, J.A.; Tye, C.E.; Boyd, J.R.; Tracy, K.M.; Frietze, S.E.; Carr, F.E.; Nickerson, J.A.; Van Wijnen, A.J.; et al. RUNX1-Dependent Mechanisms in Biological Control and Dysregulation in Cancer. J. Cell. Physiol. 2019, 234, 8597–8609. [Google Scholar] [CrossRef] [PubMed]
- Lie-A-Ling, M.; Mevel, R.; Patel, R.; Blyth, K.; Baena, E.; Kouskoff, V.; Lacaud, G. RUNX1 Dosage in Development and Cancer. Mol. Cells 2020, 43, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.; Eldor, A.; Soreq, H. Runx1/AML1 in Leukemia: Disrupted Association with Diverse Protein Partners. Leuk. Res. 2002, 26, 221–228. [Google Scholar] [CrossRef]
- Nanjundan, M.; Nakayama, Y.; Cheng, K.W.; Lahad, J.; Liu, J.; Lu, K.; Kuo, W.-L.; Smith-McCune, K.; Fishman, D.; Gray, J.W.; et al. Amplification of MDS1/EVI1 and EVI1, Located in the 3q26.2 Amplicon, Is Associated with Favorable Patient Prognosis in Ovarian Cancer. Cancer Res. 2007, 67, 3074–3084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozai, D.; Ogawa, N.; Mori, Y. Redox Regulation of Transient Receptor Potential Channels. Antioxid. Redox Signal. 2014, 21, 971–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveci, H.A.; Akyuva, Y.; Nur, G.; Naziroglu, M. Alpha Lipoic Acid Attenuates Hypoxia-Induced Apoptosis, Inflammation and Mitochondrial Oxidative Stress via Inhibition of TRPA1 Channel in Human Glioblastoma Cell Line. Biomed. Pharmacother. 2019, 111, 292–304. [Google Scholar] [CrossRef]
- Borkum, J.M. The Migraine Attack as a Homeostatic, Neuroprotective Response to Brain Oxidative Stress: Preliminary Evidence for a Theory. Headache 2018, 58, 118–135. [Google Scholar] [CrossRef]
- Osterloh, M.; Böhm, M.; Kalbe, B.; Osterloh, S.; Hatt, H. Identification and Functional Characterization of TRPA1 in Human Myoblasts. Pflugers Arch. 2016, 468, 321–333. [Google Scholar] [CrossRef]
- Raghuwanshi, S.; Dahariya, S.; Sharma, D.S.; Kovuru, N.; Sahu, I.; Gutti, R.K. RUNX1 and TGF-β Signaling Cross Talk Regulates Ca2+ Ion Channels Expression and Activity during Megakaryocyte Development. FEBS J. 2020, 287, 5411–5438. [Google Scholar] [CrossRef]
- Berrout, J.; Kyriakopoulou, E.; Moparthi, L.; Hogea, A.S.; Berrout, L.; Ivan, C.; Lorger, M.; Boyle, J.; Peers, C.; Muench, S.; et al. TRPA1-FGFR2 Binding Event Is a Regulatory Oncogenic Driver Modulated by MiRNA-142-3p. Nat. Commun. 2017, 8, 947. [Google Scholar] [CrossRef] [Green Version]
- Stock, K.; Garthe, A.; de Almeida Sassi, F.; Glass, R.; Wolf, S.A.; Kettenmann, H. The Capsaicin Receptor TRPV1 as a Novel Modulator of Neural Precursor Cell Proliferation. Stem Cells 2014, 32, 3183–3195. [Google Scholar] [CrossRef]
- Ramírez-Barrantes, R.; Cordova, C.; Poblete, H.; Muñoz, P.; Marchant, I.; Wianny, F.; Olivero, P. Perspectives of TRPV1 Function on the Neurogenesis and Neural Plasticity. Neural Plast. 2016, 2016, 1568145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Andaloussi-Lilja, J.; Lundqvist, J.; Forsby, A. TRPV1 Expression and Activity during Retinoic Acid-Induced Neuronal Differentiation. Neurochem. Int. 2009, 55, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Stock, K.; Kumar, J.; Synowitz, M.; Petrosino, S.; Imperatore, R.; Smith, E.S.J.; Wend, P.; Purfürst, B.; Nuber, U.A.; Gurok, U.; et al. Neural Precursor Cells Induce Cell Death of High-Grade Astrocytomas through Stimulation of TRPV1. Nat. Med. 2012, 18, 1232–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabissi, M.; Morelli, M.B.; Arcella, A.; Cardinali, C.; Santoni, M.; Bernardini, G.; Santoni, A.; Santoni, G.; Amantini, C. Post-Transcriptional Regulation of 5’-Untranslated Regions of Human Transient Receptor Potential Vanilloid Type-1 (TRPV-1) Channels: Role in the Survival of Glioma Patients. Oncotarget 2016, 7, 81541–81554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Shigesada, K.; Ito, K.; Wee, H.J.; Yokomizo, T.; Ito, Y. Dimerization with PEBP2beta Protects RUNX1/AML1 from Ubiquitin-Proteasome-Mediated Degradation. EMBO J. 2001, 20, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Gao, Y.; Xiao, Z.; Chen, B.; Han, J.; Zhang, J.; Wang, X.; Dai, J. Erk1/2 Promotes Proliferation and Inhibits Neuronal Differentiation of Neural Stem Cells. Neurosci. Lett. 2009, 461, 252–257. [Google Scholar] [CrossRef]
- Chien, C.-S.; Ma, K.-H.; Lee, H.-S.; Liu, P.-S.; Li, Y.-H.; Huang, Y.-S.; Chueh, S.-H. Dual Effect of Capsaicin on Cell Death in Human Osteosarcoma G292 Cells. Eur. J. Pharmacol. 2013, 718, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Donnerer, J.; Liebmann, I.; Schicho, R. ERK and STAT3 Phosphorylation in Sensory Neurons during Capsaicin-Induced Impairment and Nerve Growth Factor Treatment. Pharmacology 2005, 75, 116–121. [Google Scholar] [CrossRef]
- Iwashita, T.; Shimizu, T.; Shibata, M.; Toriumi, H.; Ebine, T.; Funakubo, M.; Suzuki, N. Activation of Extracellular Signal-Regulated Kinase in the Trigeminal Ganglion Following Both Treatment of the Dura Mater with Capsaicin and Cortical Spreading Depression. Neurosci. Res. 2013, 77, 110–119. [Google Scholar] [CrossRef]
- Firner, M.; Greffrath, W.; Treede, R.-D. Phosphorylation of Extracellular Signal-Related Protein Kinase Is Required for Rapid Facilitation of Heat-Induced Currents in Rat Dorsal Root Ganglion Neurons. Neuroscience 2006, 143, 253–263. [Google Scholar] [CrossRef]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhé, V.; Plée-Gautier, E.; Carré, J.-L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in Cutaneous Neurogenic and Chronic Inflammation: Pro-Inflammatory Response Induced by Their Activation and Their Sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Chiba, N.; Chiba, T.; Kambe, T.; Abe, K.; Kawakami, K.; Utsunomiya, I.; Taguchi, K. Transient Receptor Potential Ankyrin 1 That Is Induced in Dorsal Root Ganglion Neurons Contributes to Acute Cold Hypersensitivity after Oxaliplatin Administration. Mol. Pain 2015, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levanon, D.; Groner, Y. Structure and Regulated Expression of Mammalian RUNX Genes. Oncogene 2004, 23, 4211–4219. [Google Scholar] [CrossRef] [Green Version]
- Harrington, A.W.; Ginty, D.D. Long-Distance Retrograde Neurotrophic Factor Signalling in Neurons. Nat. Rev. Neurosci. 2013, 14, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Singer, H.S.; Hansen, B.; Martinie, D.; Karp, C.L. Mitogenesis in Glioblastoma Multiforme Cell Lines: A Role for NGF and Its TrkA Receptors. J. Neurooncol. 1999, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, J.; Xu, B.; Yuan, Z.; Leng, Y.; Min, J.; Lan, X.; Luo, J. Kaempferol Promotes Bone Formation in Part via the MTOR Signaling Pathway. Mol. Med. Rep. 2019, 20, 5197–5207. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cai, J.; Fang, C.; Yang, C.; Zhou, J.; Tan, Y.; Wang, Y.; Li, Y.; Meng, X.; Zhao, K.; et al. Mesenchymal Glioblastoma Constitutes a Major CeRNA Signature in the TGF-β Pathway. Theranostics 2018, 8, 4733–4749. [Google Scholar] [CrossRef]
- Beck, T.N.; Korobeynikov, V.A.; Kudinov, A.E.; Georgopoulos, R.; Solanki, N.R.; Andrews-Hoke, M.; Kistner, T.M.; Pépin, D.; Donahoe, P.K.; Nicolas, E.; et al. Anti-Müllerian Hormone Signaling Regulates Epithelial Plasticity and Chemoresistance in Lung Cancer. Cell Rep. 2016, 16, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Ruan, X.; Ma, J.; Liu, X.; Zheng, J.; Liu, Y.; Liu, L.; Shen, S.; Shao, L.; Wang, D.; et al. DGCR8/ZFAT-AS1 Promotes CDX2 Transcription in a PRC2 Complex-Dependent Manner to Facilitate the Malignant Biological Behavior of Glioma Cells. Mol. Ther. 2020, 28, 613–630. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-W.; Kirtay, M.; Schneble, N.; Yakoub, G.; Ding, M.; Rüdiger, T.; Siniuk, K.; Lu, R.; Jiang, Y.-N.; Li, T.-L.; et al. NBS1 Interacts with Notch Signaling in Neuronal Homeostasis. Nucleic Acids Res. 2020, 48, 10924–10939. [Google Scholar] [CrossRef]
- Mardones, M.D.; Andaur, G.A.; Varas-Godoy, M.; Henriquez, J.F.; Salech, F.; Behrens, M.I.; Couve, A.; Inestrosa, N.C.; Varela-Nallar, L. Frizzled-1 Receptor Regulates Adult Hippocampal Neurogenesis. Mol. Brain 2016, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Nguyen, V.; Clark, K.N.; Zahed, T.; Sharkas, S.; Filipp, F.V.; Boiko, A.D. Down-Regulation of FZD3 Receptor Suppresses Growth and Metastasis of Human Melanoma Independently of Canonical WNT Signaling. Proc. Natl. Acad. Sci. USA. 2019, 116, 4548–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peñuelas, S.; Anido, J.; Prieto-Sánchez, R.M.; Folch, G.; Barba, I.; Cuartas, I.; García-Dorado, D.; Poca, M.A.; Sahuquillo, J.; Baselga, J.; et al. TGF-Beta Increases Glioma-Initiating Cell Self-Renewal through the Induction of LIF in Human Glioblastoma. Cancer Cell 2009, 15, 315–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Tanaka, S.; Jiapaer, S.; Sabit, H.; Tamai, S.; Kinoshita, M.; Nakada, M. RBPJ Contributes to the Malignancy of Glioblastoma and Induction of Proneural-Mesenchymal Transition via IL-6-STAT3 Pathway. Cancer Sci. 2020, 111, 4166–4176. [Google Scholar] [CrossRef] [PubMed]
- Fedele, M.; Cerchia, L.; Pegoraro, S.; Sgarra, R.; Manfioletti, G. Proneural-Mesenchymal Transition: Phenotypic Plasticity to Acquire Multitherapy Resistance in Glioblastoma. Int. J. Mol. Sci. 2019, 20, 2746. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Cai, N.; Zhi, T.; Bao, Z.; Wang, D.; Liu, Y.; Jiang, K.; Fan, L.; Ji, J.; Liu, N. MicroRNA-1179 Inhibits Glioblastoma Cell Proliferation and Cell Cycle Progression via Directly Targeting E2F Transcription Factor 5. Am. J. Cancer Res. 2017, 7, 1680–1692. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | Cell Lines | Aml1b mRNA | Aml1c mRNA | ||
|---|---|---|---|---|---|
| Relative mRNA Expression ± SD | Fold Expression | Relative mRNA Expression ± SD | Fold Expression | ||
| NSC | 0.016 ± 0.001 | 1.00 | 0.124 ± 0.012 | 1.00 | |
| PN-like | GSC#1 | 1.141 ± 0.031 | 70.34 | 0.963 ± 0.089 | 7.77 |
| MES-like | GSC#83 | 2.432 ± 0.101 | 150.23 | 4.626 ± 0.651 | 37.31 |
| PN-like | GSC#23C | 0.721 ± 0.032 | 45.00 | 0.831 ± 0.021 | 6.70 |
| PN-like | GSC#28 | 0.733 ± 0.034 | 45.62 | 0.923 ± 0.023 | 7.44 |
| MES-like | GSC#30 | 1.952 ± 0.102 | 107.56 | 0.322 ± 0.004 | 2.60 |
| PN-like | GSC#68 | 1.721 ± 0.111 | 121.87 | 0.214 ± 0.005 | 1.69 |
| PN-like | GSC#70 | 1.232 ± 0.055 | 78.87 | 0.324 ± 0.013 | 2.61 |
| Phenotype | Cell Lines | TRPA1 mRNA | TRPV1 mRNA | ||
|---|---|---|---|---|---|
| Relative mRNA Expression ± SD | Fold Expression | Relative mRNA Expression ± SD | Fold Expression | ||
| NSC | 0.151 ± 0.014 | 1.00 | 0.554 ± 0.023 | 1.00 | |
| PN-like | GSC#1 | 0.961 ± 0.031 | 6.36 | 0.314 ± 0.089 | 0.57 |
| MES-like | GSC#83 | 8.881 ± 0.101 | 58.81 | 1.774 ± 0.051 | 3.20 |
| PN-like | GSC#23C | 0.071 ± 0.003 | 0.47 | 0.163 ± 0.004 | 0.29 |
| PN-like | GSC#28 | 0.292 ± 0.043 | 1.93 | 0.217 ± 0.023 | 0.39 |
| MES-like | GSC#30 | 0.302 ± 0.003 | 2.00 | 0.015 ± 0.003 | 0.03 |
| PN-like | GSC#68 | 0.452 ± 0.013 | 2.99 | 0.543 ± 0.005 | 0.98 |
| PN-like | GSC#70 | 0.881 ± 0.054 | 5.83 | 0.692 ± 0.022 | 1.25 |
| Phenotype | Cell Lines | Aml1b mRNA | Aml1c mRNA | ||
|---|---|---|---|---|---|
| Relative mRNA Expression ± SD | Fold Expression | Relative mRNA Expression ± SD | Fold Expression | ||
| PN-like | D-GSC#1 | 2.621 ± 0.031 | 2.15 | 2.813 ± 0.089 | 2.26 |
| MES-like | D-GSC#83 | 5.056 ± 2.101 | 2.08 | 10.085 ± 0.651 | 2.18 |
| PN-like | D-GSC#23C | 1.722 ± 0.032 | 2.39 | 1.988 ± 0.021 | 2.38 |
| PN-like | D-GSC#28 | 2.776 ± 0.035 | 3.79 | 3.500 ± 0.023 | 3.23 |
| MES-like | D-GSC#30 | 7.810 ± 0.013 | 4.00 | 0.646 ± 0.004 | 1.99 |
| PN-like | D-GSC#68 | 2.071 ± 0.107 | 1.20 | 0.612 ± 0.005 | 2.85 |
| PN-like | D-GSC#70 | 1.303 ± 0.046 | 1.06 | 0.998 ± 0.013 | 3.08 |
| Phenotype | Cell Lines | TRPA1 mRNA | TRPV1 mRNA | ||
|---|---|---|---|---|---|
| Relative mRNA Expression ± SD | Fold Expression | Relative mRNA Expression ± SD | Fold Expression | ||
| PN-like | D-GSC#1 | 4.622 ± 0.031 | 4.81 | 0.187 ± 0.089 | 0.34 |
| MES-like | D-GSC#83 | 38.851 ± 0.101 | 4.49 | 1.202 ± 0.051 | 0.68 |
| PN-like | D-GSC#23C | 4.994 ± 0.221 | 212.93 | 0.113 ± 0.011 | 0.69 |
| PN-like | D-GSC#28 | 0.122 ± 0.001 | 0.42 | 0.131 ± 0.001 | 0.60 |
| MES-like | D-GSC#30 | 1.143 ± 0.013 | 3.78 | 0.007 ± 0.003 | 0.47 |
| PN-like | D-GSC#68 | 0.292 ± 0.022 | 0.65 | 0.363 ± 0.023 | 0.67 |
| PN-like | D-GSC#70 | 0.227 ± 0.011 | 0.26 | 0.233 ± 0.014 | 0.34 |
| Upregulated Genes | Fold Expression ± SD | Downregulated Genes | Fold Expression ± SD |
|---|---|---|---|
| ACVR1 | 2.54 ± 0.20 | E2F5 | −2.45 ± 0.12 |
| AMHR2 | 3.81 ± 0.10 | FZD2 | −2.86 ± 0.11 |
| BMPR2 | 2.66 ± 0.32 | LIFR | −2.91 ± 0.01 |
| CDX2 | 2.40 ± 0.15 | NOTCH3 | −2.95 ± 0.15 |
| FZD1 | 2.77 ± 0.03 | NOTCH4 | −2.00 ± 0.29 |
| FZD3 | 2.69 ± 0.10 | SMAD3 | −2.15 ± 0.08 |
| SMAD2 | 2.67 ± 0.10 | STAT3 | −5.06 ± 0.50 |
| Upregulated Genes | Fold Expression ± SD | Downregulated Genes | Fold Expression ± SD |
|---|---|---|---|
| ACVR1 | 2.85 ± 0.10 | FGFR4 | −11.64 ± 0.08 |
| AMHR2 | 2.55 ± 0.30 | FZD1 | −10.26 ± 0.01 |
| BMPR1B | 2.90 ± 0.29 | LRP6 | −4.83 ± 0.11 |
| CDX2 | 2.00 ± 0.05 | LTBP2 | −7.75 ± 0.10 |
| FGFR2 | 6.70 ± 0.30 | NFAT5 | −2.24 ± 0.09 |
| FZD9 | 3.00 ± 0.15 | NOTCH4 | −8.84 ± 0.51 |
| IL6ST | 3.34 ± 0.10 | RGMA | −2.64 ± 0.61 |
| LIFR | 2.40 ± 0.12 | SMAD3 | −3.33 ± 0.31 |
| RBPJL | 2.05 ± 0.13 | TGFBR2 | −6.40 ± 0.72 |
| SMAD9 | 4.70 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoni, G.; Nabissi, M.; Amantini, C.; Santoni, M.; Ricci-Vitiani, L.; Pallini, R.; Maggi, F.; Morelli, M.B. ERK Phosphorylation Regulates the Aml1/Runx1 Splice Variants and the TRP Channels Expression during the Differentiation of Glioma Stem Cell Lines. Cells 2021, 10, 2052. https://doi.org/10.3390/cells10082052
Santoni G, Nabissi M, Amantini C, Santoni M, Ricci-Vitiani L, Pallini R, Maggi F, Morelli MB. ERK Phosphorylation Regulates the Aml1/Runx1 Splice Variants and the TRP Channels Expression during the Differentiation of Glioma Stem Cell Lines. Cells. 2021; 10(8):2052. https://doi.org/10.3390/cells10082052
Chicago/Turabian StyleSantoni, Giorgio, Massimo Nabissi, Consuelo Amantini, Matteo Santoni, Lucia Ricci-Vitiani, Roberto Pallini, Federica Maggi, and Maria Beatrice Morelli. 2021. "ERK Phosphorylation Regulates the Aml1/Runx1 Splice Variants and the TRP Channels Expression during the Differentiation of Glioma Stem Cell Lines" Cells 10, no. 8: 2052. https://doi.org/10.3390/cells10082052