
Localization and Functional Roles of Components of the Translation Apparatus in the Eukaryotic Cell Nucleus

 , , , and
, , , and
Abstract
:1. Introduction
2. Nuclear and Subnuclear Localization of Translation Factors and Its Regulation
3. Regulation of RP Nuclear Localization
4. Roles for CTAs in DNA Repair, Synthesis, and Genome Integrity Control
5. Roles of CTAs in Transcriptional Regulation
6. Role of CTAs in mRNA Processing
7. Role of CTAs in Nuclear mRNA Export
8. Nuclear Localization of some CTAs Is Associated with Oncogenesis
9. Nuclear Translation Hypothesis
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CTA | Component of the translation apparatus |
| NLS | Nuclear localization signal |
| NMD | Nonsense mediated decay |
| NPC | Nuclear pore complex |
| RNAP | RNA polymerase |
| RNP | Ribo-nucleoprotein |
| RP | Ribosomal protein |
| TF | Transcription factor |
References
- Jackson, R.J.; Hellen, C.U.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Lecompte, O.; Ripp, R.; Thierry, J.C.; Moras, D.; Poch, O. Comparative analysis of ribosomal proteins in complete genomes: An example of reductive evolution at the domain scale. Nucleic Acids Res. 2002, 30, 5382–5390. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Yoshihama, M.; Kenmochi, N. RPG: The Ribosomal Protein Gene database. Nucleic Acids Res. 2004, 32, D168–D170. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, J.; Karbstein, K.; Woolford, J.L., Jr. Functions of ribosomal proteins in assembly of eukaryotic ribosomes in vivo. Annu. Rev. Biochem. 2015, 84, 93–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.P. Balanced production of ribosomal proteins. Gene 2007, 401, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Grove, A. Coordination of Ribosomal Protein and Ribosomal RNA Gene Expression in Response to TOR Signaling. Curr. Genom. 2009, 10, 198–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecoraro, A.; Pagano, M.; Russo, G.; Russo, A. Ribosome Biogenesis and Cancer: Overview on Ribosomal Proteins. Int. J. Mol. Sci. 2021, 22, 5496. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R. The economics of ribosome biosynthesis in yeast. Trends Biochem. Sci. 1999, 24, 437–440. [Google Scholar] [CrossRef]
- Petibon, C.; Malik Ghulam, M.; Catala, M.; Abou Elela, S. Regulation of ribosomal protein genes: An ordered anarchy. Wiley Interdiscip. Rev. RNA 2021, 12, e1632. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Yu, J.; Ward, R.; Liu, Y.; Hao, Q.; An, S.; Xu, T. Eukaryotic translation initiation factors as promising targets in cancer therapy. Cell Commun. Signal. 2020, 18, 175. [Google Scholar] [CrossRef] [PubMed]
- Fukao, A.; Tomohiro, T.; Fujiwara, T. Translation Initiation Regulated by RNA-Binding Protein in Mammals: The Modulation of Translation Initiation Complex by Trans-Acting Factors. Cells 2021, 10, 1711. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, G.C. The evolutionary and functional diversity of classical and lesser-known cytoplasmic and organellar translational GTPases across the tree of life. BMC Genom. 2015, 16, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marygold, S.J.; Attrill, H.; Lasko, P. The translation factors of Drosophila melanogaster. Fly 2017, 11, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dever, T.E.; Green, R. The elongation, termination, and recycling phases of translation in eukaryotes. Cold Spring Harb Perspect. Biol. 2012, 4, a013706. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.R.P.; Garland, G.; Poyry, T.; Mead, E.; Vlahov, N.; Sfakianos, A.; Grosso, S.; De-Lima-Hedayioglu, F.; Mallucci, G.R.; von der Haar, T.; et al. Control of translation elongation in health and disease. Dis. Model Mech. 2020, 13, dmm043208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genuth, N.R.; Barna, M. Heterogeneity and specialized functions of translation machinery: From genes to organisms. Nat. Rev. Genet. 2018, 19, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Lejbkowicz, F.; Goyer, C.; Darveau, A.; Neron, S.; Lemieux, R.; Sonenberg, N. A fraction of the mRNA 5′ cap-binding protein, eukaryotic initiation factor 4E, localizes to the nucleus. Proc. Natl. Acad. Sci. USA 1992, 89, 9612–9616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, V.; Zanchin, N.I.; Lunsdorf, H.; Tuite, M.; McCarthy, J.E. Initiation factor eIF-4E of Saccharomyces cerevisiae. Distribution within the cell, binding to mRNA, and consequences of its overproduction. J. Biol. Chem. 1994, 269, 6117–6123. [Google Scholar] [CrossRef]
- Robalino, J.; Joshi, B.; Fahrenkrug, S.C.; Jagus, R. Two zebrafish eIF4E family members are differentially expressed and functionally divergent. J. Biol. Chem. 2004, 279, 10532–10541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuyama, A.; Arai, R.; Yashiroda, Y.; Shirai, A.; Kamata, A.; Sekido, S.; Kobayashi, Y.; Hashimoto, A.; Hamamoto, M.; Hiraoka, Y.; et al. ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2006, 24, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef]
- Lye, C.M.; Naylor, H.W.; Sanson, B. Subcellular localisations of the CPTI collection of YFP-tagged proteins in Drosophila embryos. Development 2014, 141, 4006–4017. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.S.; Cho, E.J.; Choi, E.Y.; Park, O.K. Analysis of the Arabidopsis nuclear proteome and its response to cold stress. Plant J. 2003, 36, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, O.A.; Tomlinson, M.L.; Leader, D.; Shaw, P.; Doonan, J.H. High-throughput protein localization in Arabidopsis using Agrobacterium-mediated transient expression of GFP-ORF fusions. Plant J. 2005, 41, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Goto, C.; Hashizume, S.; Fukao, Y.; Hara-Nishimura, I.; Tamura, K. Comprehensive nuclear proteome of Arabidopsis obtained by sequential extraction. Nucleus 2019, 10, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Ayash, M.; Abukhalaf, M.; Thieme, D.; Proksch, C.; Heilmann, M.; Schattat, M.; Hoehenwarter, W. LC-MS Based Draft Map of the Arabidopsis thaliana Nuclear Proteome and Protein Import in Pattern Triggered Immunity. Front. Plant Sci. 2021, 12, 744103. [Google Scholar] [CrossRef]
- Bohnsack, M.T.; Regener, K.; Schwappach, B.; Saffrich, R.; Paraskeva, E.; Hartmann, E.; Gorlich, D. Exp5 exports eEF1A via tRNA from nuclei and synergizes with oTher. transport pathways to confine translation to the cytoplasm. EMBO J. 2002, 21, 6205–6215. [Google Scholar] [CrossRef] [Green Version]
- Mingot, J.M.; Kostka, S.; Kraft, R.; Hartmann, E.; Gorlich, D. Importin 13: A novel mediator of nuclear import and export. EMBO J. 2001, 20, 3685–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, E.V.; Rakitina, T.V.; Bogatova, O.V.; Ivanova, D.L.; Vorobyeva, E.E.; Lipkin, A.V.; Kostanyan, I.A.; Lipkin, V.M. Novel protein haponin regulates cellular response to oxidative stress. Dokl. Biochem. Biophys. 2011, 440, 225–227. [Google Scholar] [CrossRef]
- Sanvito, F.; Piatti, S.; Villa, A.; Bossi, M.; Lucchini, G.; Marchisio, P.C.; Biffo, S. The beta4 integrin interactor p27(BBP/eIF6) is an essential nuclear matrix protein involved in 60S ribosomal subunit assembly. J. Cell Biol. 1999, 144, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.; Copeland, P.R. The Selenocysteine-Specific Elongation Factor Contains Unique Sequences That Are Required for Both Nuclear Export and Selenocysteine Incorporation. PLoS ONE 2016, 11, e0165642. [Google Scholar] [CrossRef]
- Wuhr, M.; Guttler, T.; Peshkin, L.; McAlister, G.C.; Sonnett, M.; Ishihara, K.; Groen, A.C.; Presler, M.; Erickson, B.K.; Mitchison, T.J.; et al. The Nuclear Proteome of a Vertebrate. Curr. Biol. 2015, 25, 2663–2671. [Google Scholar] [CrossRef] [Green Version]
- Rafiee, M.-R.; Zagalak, J.A.; Sidorov, S.; Steinhauser, S.; Davey, K.; Ule, J.; Luscombe, N.M. Chromatin-contact atlas reveals disorder-mediated protein interactions and moonlighting chromatin-associated RBPs. bioRxiv 2021, 200212. [Google Scholar] [CrossRef]
- Pendle, A.F.; Clark, G.P.; Boon, R.; Lewandowska, D.; Lam, Y.W.; Andersen, J.; Mann, M.; Lamond, A.I.; Brown, J.W.; Shaw, P.J. Proteomic analysis of the Arabidopsis nucleolus suggests novel nucleolar functions. Mol. Biol. Cell 2005, 16, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Lacoux, C.; Wacheul, L.; Saraf, K.; Pythoud, N.; Huvelle, E.; Figaro, S.; Graille, M.; Carapito, C.; Lafontaine, D.L.J.; Heurgue-Hamard, V. The catalytic activity of the translation termination factor methyltransferase Mtq2-Trm112 complex is required for large ribosomal subunit biogenesis. Nucleic Acids Res. 2020, 48, 12310–12325. [Google Scholar] [CrossRef]
- Salsman, J.; Pinder, J.; Tse, B.; Corkery, D.; Dellaire, G. The translation initiation factor 3 subunit eIF3K interacts with PML and associates with PML nuclear bodies. Exp. Cell Res. 2013, 319, 2554–2565. [Google Scholar] [CrossRef]
- Shen, X.; Yang, Y.; Liu, W.; Sun, M.; Jiang, J.; Zong, H.; Gu, J. Identification of the p28 subunit of eukaryotic initiation factor 3(eIF3k) as a new interaction partner of cyclin D3. FEBS Lett. 2004, 573, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Morris-Desbois, C.; Bochard, V.; Reynaud, C.; Jalinot, P. Interaction between the Ret finger protein and the Int-6 gene product and co-localisation into nuclear bodies. J. Cell Sci. 1999, 112 Pt 19, 3331–3342. [Google Scholar] [CrossRef]
- Lai, H.K.; Borden, K.L. The promyelocytic leukemia (PML) protein suppresses cyclin D1 protein production by altering the nuclear cytoplasmic distribution of cyclin D1 mRNA. Oncogene 2000, 19, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, N.; Spahr, C.S.; Patterson, S.D.; Bubulya, P.; Neuwald, A.F.; Spector, D.L. Proteomic analysis of interchromatin granule clusters. Mol. Biol. Cell 2004, 15, 3876–3890. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, R.; Henzing, A.J.; Earnshaw, W.C. Novel components of human mitotic chromosomes identified by proteomic analysis of the chromosome scaffold fraction. Chromosoma 2005, 113, 385–397. [Google Scholar] [CrossRef]
- Takata, H.; Nishijima, H.; Ogura, S.; Sakaguchi, T.; Bubulya, P.A.; Mochizuki, T.; Shibahara, K. Proteome analysis of human nuclear insoluble fractions. Genes Cells 2009, 14, 975–990. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, M.; Blalock, W.L.; Bavelloni, A.; Faenza, I.; D’Angelo, A.; Maraldi, N.M.; Cocco, L. Phosphoinositide-specific phospholipase C beta 1b (PI-PLCbeta1b) interactome: Affinity purification-mass spectrometry analysis of PI-PLCbeta1b with nuclear protein. Mol. Cell Proteom. 2013, 12, 2220–2235. [Google Scholar] [CrossRef] [Green Version]
- Guruharsha, K.G.; Rual, J.F.; Zhai, B.; Mintseris, J.; Vaidya, P.; Vaidya, N.; Beekman, C.; Wong, C.; Rhee, D.Y.; Cenaj, O.; et al. A protein complex network of Drosophila melanogaster. Cell 2011, 147, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Sesen, J.; Casaos, J.; Scotland, S.J.; Seva, C.; Eisinger-Mathason, T.S.; Skuli, N. The Bad, the Good and eIF3e/INT6. Front. BioSci. (Landmark Ed) 2017, 22, 1–20. [Google Scholar] [CrossRef]
- Hoareau Alves, K.; Bochard, V.; Rety, S.; Jalinot, P. Association of the mammalian proto-oncoprotein Int-6 with the three protein complexes eIF3, COP9 signalosome and 26S proteasome. FEBS Lett. 2002, 527, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Yen, H.C.; Gordon, C.; Chang, E.C. Schizosaccharomyces pombe Int6 and Ras homologs regulate cell division and mitotic fidelity via the proteasome. Cell 2003, 112, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Sha, Z.; Brill, L.M.; Cabrera, R.; Kleifeld, O.; Scheliga, J.S.; Glickman, M.H.; Chang, E.C.; Wolf, D.A. The eIF3 interactome reveals the translasome, a supercomplex linking protein synthesis and degradation machineries. Mol. Cell 2009, 36, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Yahalom, A.; Kim, T.H.; Winter, E.; Karniol, B.; von Arnim, A.G.; Chamovitz, D.A. Arabidopsis eIF3e (INT-6) associates with both eIF3c and the COP9 signalosome subunit CSN7. J. Biol. Chem. 2001, 276, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Kimball, S.R.; Horetsky, R.L.; Ron, D.; Jefferson, L.S.; Harding, H.P. Mammalian stress granules represent sites of accumulation of stalled translation initiation complexes. Am. J. Physiol. Cell Physiol. 2003, 284, C273–C284. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, E.N.; Owen, C.R.; White, B.C.; Rafols, J.A. Ultrastructural localization of phosphorylated eIF2alpha [eIF2alpha(P)] in rat dorsal hippocampus during reperfusion. Acta Neuropathol. 1999, 98, 493–505. [Google Scholar] [CrossRef]
- Lobo, M.V.; Alonso, F.J.; Rodriguez, S.; Alcazar, A.; Martin, E.; Munoz, F.; Santander, R.G.; Salinas, M.; Fando, J.L. Localization of eukaryotic initiation factor 2 in neuron primary cultuRes. and established cell lines. Histochem. J. 1997, 29, 453–468. [Google Scholar] [CrossRef]
- DeGracia, D.J.; Sullivan, J.M.; Neumar, R.W.; Alousi, S.S.; Hikade, K.R.; Pittman, J.E.; White, B.C.; Rafols, J.A.; Krause, G.S. Effect of brain ischemia and reperfusion on the localization of phosphorylated eukaryotic initiation factor 2 alpha. J. Cereb. Blood Flow Metab. 1997, 17, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Beausoleil, S.A.; Jedrychowski, M.; Schwartz, D.; Elias, J.E.; Villen, J.; Li, J.; Cohn, M.A.; Cantley, L.C.; Gygi, S.P. Large-scale characterization of HeLa cell nuclear phosphoproteins. Proc. Natl. Acad. Sci. USA 2004, 101, 12130–12135. [Google Scholar] [CrossRef] [Green Version]
- Turpaev, K.T. Translation Factor eIF5A, Modification with Hypusine and Role in Regulation of Gene Expression. eIF5A as a Target for Pharmacological Interventions. Biochemistry 2018, 83, 863–873. [Google Scholar] [CrossRef]
- Lee, S.B.; Park, J.H.; Kaevel, J.; Sramkova, M.; Weigert, R.; Park, M.H. The effect of hypusine modification on the intracellular localization of eIF5A. Biochem. Biophys. Res. Commun. 2009, 383, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Aksu, M.; Trakhanov, S.; Gorlich, D. Structure of the exportin Xpo4 in complex with RanGTP and the hypusine-containing translation factor eIF5A. Nat. Commun. 2016, 7, 11952. [Google Scholar] [CrossRef] [Green Version]
- Ren, B.; Chen, Q.; Hong, S.; Zhao, W.; Feng, J.; Feng, H.; Zuo, J. The Arabidopsis eukaryotic translation initiation factor eIF5A-2 regulates root protoxylem development by modulating cytokinin signaling. Plant Cell 2013, 25, 3841–3857. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, M.T.; Lampi, Y.; Wang, T.W.; Liu, Z.; Thompson, J.E. Eukaryotic translation initiation factor 5A is involved in pathogen-induced cell death and development of disease symptoms in Arabidopsis. Plant Physiol. 2008, 148, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Lebska, M.; Ciesielski, A.; Szymona, L.; Godecka, L.; Lewandowska-Gnatowska, E.; Szczegielniak, J.; Muszynska, G. Phosphorylation of maize eukaryotic translation initiation factor 5A (eIF5A) by casein kinase 2: Identification of phosphorylated residue and influence on intracellular localization of eIF5A. J. Biol. Chem. 2010, 285, 6217–6226. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska-Gnatowska, E.; Szymona, L.; Lebska, M.; Szczegielniak, J.; Muszynska, G. Phosphorylation of maize eukaryotic translation initiation factor on Ser2 by catalytic subunit CK2. Mol. Cell Biochem. 2011, 356, 241–244. [Google Scholar] [CrossRef]
- Jao, D.L.; Yu Chen, K. Subcellular localization of the hypusine-containing eukaryotic initiation factor 5A by immunofluorescent staining and green fluorescent protein tagging. J. Cell Biochem. 2002, 86, 590–600. [Google Scholar] [CrossRef]
- Parreiras, E.S.L.T.; Gomes, M.D.; Oliveira, E.B.; Costa-Neto, C.M. The N-terminal region of eukaryotic translation initiation factor 5A signals to nuclear localization of the protein. Biochem. Biophys. Res. Commun. 2007, 362, 393–398. [Google Scholar] [CrossRef]
- Wu, G.Q.; Xu, Y.M.; Lau, A.T.Y. Recent insights into eukaryotic translation initiation factors 5A1 and 5A2 and their roles in human health and disease. Cancer Cell Int. 2020, 20, 142. [Google Scholar] [CrossRef]
- Dostie, J.; Ferraiuolo, M.; Pause, A.; Adam, S.A.; Sonenberg, N. A novel shuttling protein, 4E-T, mediates the nuclear import of the mRNA 5′ cap-binding protein, eIF4E. EMBO J. 2000, 19, 3142–3156. [Google Scholar] [CrossRef] [Green Version]
- Kamenska, A.; Lu, W.T.; Kubacka, D.; Broomhead, H.; Minshall, N.; Bushell, M.; Standart, N. Human 4E-T represses translation of bound mRNAs and enhances microRNA-mediated silencing. Nucleic Acids Res. 2014, 42, 3298–3313. [Google Scholar] [CrossRef]
- Marnef, A.; Weil, D.; Standart, N. RNA-related nuclear functions of human Pat1b, the P-body mRNA decay factor. Mol. Biol. Cell 2012, 23, 213–224. [Google Scholar] [CrossRef]
- Huang, J.H.; Ku, W.C.; Chen, Y.C.; Chang, Y.L.; Chu, C.Y. Dual mechanisms regulate the nucleocytoplasmic localization of human DDX6. Sci. Rep. 2017, 7, 42853. [Google Scholar] [CrossRef] [Green Version]
- Zappavigna, V.; Piccioni, F.; Villaescusa, J.C.; Verrotti, A.C. Cup is a nucleocytoplasmic shuttling protein that interacts with the eukaryotic translation initiation factor 4E to modulate Drosophila ovary development. Proc. Natl. Acad. Sci. USA 2004, 101, 14800–14805. [Google Scholar] [CrossRef] [Green Version]
- Desbois, C.; Rousset, R.; Bantignies, F.; Jalinot, P. Exclusion of Int-6 from PML nuclear bodies by binding to the HTLV-I Tax oncoprotein. Science 1996, 273, 951–953. [Google Scholar] [CrossRef]
- Toro-Ascuy, D.; Rojas-Araya, B.; Garcia-de-Gracia, F.; Rojas-Fuentes, C.; Pereira-Montecinos, C.; Gaete-Argel, A.; Valiente-Echeverria, F.; Ohlmann, T.; Soto-Rifo, R. A Rev-CBP80-eIF4AI complex drives Gag synthesis from the HIV-1 unspliced mRNA. Nucleic Acids Res. 2018, 46, 11539–11552. [Google Scholar] [CrossRef] [Green Version]
- Etchison, D.; Etchison, J.R. Monoclonal antibody-aided characterization of cellular p220 in uninfected and poliovirus-infected HeLa cells: Subcellular distribution and identification of conformers. J. Virol. 1987, 61, 2702–2710. [Google Scholar] [CrossRef] [Green Version]
- Cakmakci, N.G.; Lerner, R.S.; Wagner, E.J.; Zheng, L.; Marzluff, W.F. SLIP1, a factor required for activation of histone mRNA translation by the stem-loop binding protein. Mol. Cell Biol. 2008, 28, 1182–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, M.; Tanaka, T.; Shigeta, M.; Chuma, S.; Saga, Y.; Saitou, M. Functional reconstruction of NANOS3 expression in the germ cell lineage by a novel transgenic reporter reveals distinct subcellular localizations of NANOS3. Reproduction 2010, 139, 381–393. [Google Scholar] [CrossRef] [Green Version]
- Julaton, V.T.; Reijo Pera, R.A. NANOS3 function in human germ cell development. Hum. Mol. Genet. 2011, 20, 2238–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Zhong, Y.; Molinar-Inglis, O.; Kunkel, M.T.; Chen, M.; Sun, T.; Zhang, J.; Shyy, J.Y.; Trejo, J.; Newton, A.C.; et al. Location-specific inhibition of Akt reveals regulation of mTORC1 activity in the nucleus. Nat. Commun. 2020, 11, 6088. [Google Scholar] [CrossRef]
- Schule, M.; Butto, T.; Dewi, S.; Schlichtholz, L.; Strand, S.; Gerber, S.; Endres, K.; Schweiger, S.; Winter, J. mTOR Driven Gene Transcription Is Required for Cholesterol Production in Neurons of the Developing Cerebral Cortex. Int. J. Mol. Sci. 2021, 22, 6034. [Google Scholar] [CrossRef]
- Blalock, W.L.; Bavelloni, A.; Piazzi, M.; Tagliavini, F.; Faenza, I.; Martelli, A.M.; Follo, M.Y.; Cocco, L. Multiple forms of PKR present in the nuclei of acute leukemia cells represent an active kinase that is responsive to stress. Leukemia 2011, 25, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, K.A.; Gallagher, J.E.; Mitchell, B.M.; Granneman, S.; Baserga, S.J. The small-subunit processome is a ribosome assembly intermediate. Eukaryot Cell 2004, 3, 1619–1626. [Google Scholar] [CrossRef] [Green Version]
- Kressler, D.; Bange, G.; Ogawa, Y.; Stjepanovic, G.; Bradatsch, B.; Pratte, D.; Amlacher, S.; Strauss, D.; Yoneda, Y.; Katahira, J.; et al. Synchronizing nuclear import of ribosomal proteins with ribosome assembly. Science 2012, 338, 666–671. [Google Scholar] [CrossRef] [Green Version]
- Robledo, S.; Idol, R.A.; Crimmins, D.L.; Ladenson, J.H.; Mason, P.J.; Bessler, M. The role of human ribosomal proteins in the maturation of rRNA and ribosome production. RNA 2008, 14, 1918–1929. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, T.; Hayano, T.; Yanagida, M.; Takahashi, N.; Nishimoto, T. NOP132 is required for proper nucleolus localization of DEAD-box RNA helicase DDX47. Nucleic Acids Res. 2006, 34, 4593–4608. [Google Scholar] [CrossRef] [Green Version]
- Fong, K.W.; Li, Y.; Wang, W.; Ma, W.; Li, K.; Qi, R.Z.; Liu, D.; Songyang, Z.; Chen, J. Whole-genome screening identifies proteins localized to distinct nuclear bodies. J. Cell Biol. 2013, 203, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, E.; Parisot, P.; Pinto-Monteiro, C.; de Walque, R.; De Vleeschouwer, C.; Lafontaine, D.L. Involvement of human ribosomal proteins in nucleolar structure and p53-dependent nucleolar stress. Nat. Commun. 2016, 7, 11390. [Google Scholar] [CrossRef]
- Melnikov, S.; Ben-Shem, A.; Yusupova, G.; Yusupov, M. Insights into the origin of the nuclear localization signals in conserved ribosomal proteins. Nat. Commun. 2015, 6, 8382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelter, P.; Huber, F.M.; Kunze, R.; Flemming, D.; Hoelz, A.; Hurt, E. Coordinated Ribosomal L4 Protein Assembly into the Pre-Ribosome Is Regulated by Its Eukaryote-Specific Extension. Mol. Cell 2015, 58, 854–862. [Google Scholar] [CrossRef] [Green Version]
- Claussen, M.; Rudt, F.; Pieler, T. Functional modules in ribosomal protein L5 for ribonucleoprotein complex formation and nucleocytoplasmic transport. J. Biol. Chem. 1999, 274, 33951–33958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahama, M.; Hara, Y.; Seki, A.; Yamazoe, T.; Kawate, Y.; Shinohara, T.; Hatsuzawa, K.; Tani, K.; Tagaya, M. NVL2 is a nucleolar AAA-ATPase that interacts with ribosomal protein L5 through its nucleolar localization sequence. Mol. Biol. Cell 2004, 15, 5712–5723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plafker, S.M.; Macara, I.G. Ribosomal protein L12 uses a distinct nuclear import pathway mediated by importin 11. Mol. Cell Biol. 2002, 22, 1266–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakel, S.; Gorlich, D. Importin beta, transportin, RanBP5 and RanBP7 mediate nuclear import of ribosomal proteins in mammalian cells. EMBO J. 1998, 17, 4491–4502. [Google Scholar] [CrossRef] [Green Version]
- Mitterer, V.; Gantenbein, N.; Birner-Gruenberger, R.; Murat, G.; Bergler, H.; Kressler, D.; Pertschy, B. Nuclear import of dimerized ribosomal protein Rps3 in complex with its chaperone Yar1. Sci. Rep. 2016, 6, 36714. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.S.; Lam, Y.W.; Leung, A.K.; Ong, S.E.; Lyon, C.E.; Lamond, A.I.; Mann, M. Nucleolar proteome dynamics. Nature 2005, 433, 77–83. [Google Scholar] [CrossRef]
- Kazyken, D.; Kaz, Y.; Kiyan, V.; Zhylkibayev, A.A.; Chen, C.H.; Agarwal, N.K.; dos Sarbassov, D. The nuclear import of ribosomal proteins is regulated by mTOR. Oncotarget 2014, 5, 9577–9593. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Casadevall, R.; Luciani, M.D.; Pezza, A.; Casati, P. New evidence for differential roles of l10 ribosomal proteins from Arabidopsis. Plant Physiol. 2013, 163, 378–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Wang, Y.; Liang, Y.; Zhang, Y.; Bao, S.; Xu, Z. Methylation of ribosomal protein S10 by protein-arginine methyltransferase 5 regulates ribosome biogenesis. J. Biol. Chem. 2010, 285, 12695–12705. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.S.; Jang, C.Y.; Kim, H.D.; Kim, T.S.; Kim, S.; Kim, J. Arginine methylation of ribosomal protein S3 affects ribosome assembly. Biochem. Biophys. Res. Commun. 2009, 385, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Malecki, J.M.; Odonohue, M.F.; Kim, Y.; Jakobsson, M.E.; Gessa, L.; Pinto, R.; Wu, J.; Davydova, E.; Moen, A.; Olsen, J.V.; et al. Human METTL18 is a histidine-specific methyltransferase that targets RPL3 and affects ribosome biogenesis and function. Nucleic Acids Res. 2021, 49, 3185–3203. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.G.; Ireland, J.A.; Prem, S.M.; Chen, A.S.; Ware, V.C. RpL22e, but not RpL22e-like-PA, is SUMOylated and localizes to the nucleoplasm of Drosophila meiotic spermatocytes. Nucleus 2013, 4, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Fok, V.; Mitton-Fry, R.M.; Grech, A.; Steitz, J.A. Multiple domains of EBER 1, an Epstein-Barr virus noncoding RNA, recruit human ribosomal protein L22. RNA 2006, 12, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Toczyski, D.P.; Matera, A.G.; Ward, D.C.; Steitz, J.A. The Epstein-Barr virus (EBV) small RNA EBER1 binds and relocalizes ribosomal protein L22 in EBV-infected human B lymphocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 3463–3467. [Google Scholar] [CrossRef] [Green Version]
- Rajamaki, M.L.; Sikorskaite-Gudziuniene, S.; Sarmah, N.; Varjosalo, M.; Valkonen, J.P.T. Nuclear proteome of virus-infected and healthy potato leaves. BMC Plant Biol. 2020, 20, 355. [Google Scholar] [CrossRef]
- Li, Y.; Dong, W.; Shi, Y.; Deng, F.; Chen, X.; Wan, C.; Zhou, M.; Zhao, L.; Fu, Z.F.; Peng, G. Rabies virus phosphoprotein interacts with ribosomal protein L9 and affects rabies virus replication. Virology 2016, 488, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, M.S.; Zhang, Y. Ribosomal protein S9 is a novel B23/NPM-binding protein required for normal cell proliferation. J. Biol. Chem. 2008, 283, 15568–15576. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, M.S. Elucidation of motifs in ribosomal protein S9 that mediate its nucleolar localization and binding to NPM1/nucleophosmin. PLoS ONE 2012, 7, e52476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba, Q.; Li, X.; Huang, C.; Li, J.; Fu, Y.; Chen, P.; Duan, J.; Hao, M.; Zhang, Y.; Li, J.; et al. BCCIPbeta modulates the ribosomal and extraribosomal function of S7 through a direct interaction. J. Mol. Cell Biol. 2017, 9, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Wyler, E.; Wandrey, F.; Badertscher, L.; Montellese, C.; Alper, D.; Kutay, U. The beta-isoform of the BRCA2 and CDKN1A(p21)-interacting protein (BCCIP) stabilizes nuclear RPL23/uL14. FEBS Lett. 2014, 588, 3685–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamaguchi, N.; Ohdaira, T.; Shinohara, A.; Iwamatsu, A.; Ihara, S.; Fukui, Y. Identification of ribosomal protein S3a as a candidate for a novel PI 3-kinase target in the nucleus. Cytotechnology 2002, 40, 85–92. [Google Scholar] [CrossRef]
- Tsubota, S.I.; Phillips, A.C. Drosophila Enhancer of Rudimentary Homolog, ERH, Is a Binding Partner of RPS3, RPL19, and DDIT4, Suggesting a Mechanism for the Nuclear Localization of ERH. Mol. Biol. Int. 2016, 2016, 8371819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, A.R.; Bann, D.V.; Rice, B.; Pultz, I.S.; Kane, M.; Goff, S.P.; Golovkina, T.V.; Parent, L.J. Nucleolar trafficking of the mouse mammary tumor virus gag protein induced by interaction with ribosomal protein L9. J. Virol. 2013, 87, 1069–1082. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, R.; Scaglioni, P.P.; Bergmann, S.; Horn, H.F.; Vousden, K.H.; Pandolfi, P.P. PML regulates p53 stability by sequestering Mdm2 to the nucleolus. Nat. Cell Biol. 2004, 6, 665–672. [Google Scholar] [CrossRef]
- Chen, W.; Dittmer, D.P. Ribosomal protein S6 interacts with the latency-associated nuclear antigen of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2011, 85, 9495–9505. [Google Scholar] [CrossRef] [Green Version]
- Robles, M.S.; Boyault, C.; Knutti, D.; Padmanabhan, K.; Weitz, C.J. Identification of RACK1 and protein kinase Calpha as integral components of the mammalian circadian clock. Science 2010, 327, 463–466. [Google Scholar] [CrossRef]
- Guo, J.; Wang, S.; Valerius, O.; Hall, H.; Zeng, Q.; Li, J.F.; Weston, D.J.; Ellis, B.E.; Chen, J.G. Involvement of Arabidopsis RACK1 in protein translation and its regulation by abscisic acid. Plant Physiol. 2011, 155, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Yadavilli, S.; Hegde, V.; Deutsch, W.A. Translocation of human ribosomal protein S3 to sites of DNA damage is dependant on ERK-mediated phosphorylation following genotoxic stress. DNA Repair 2007, 6, 1453–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, I.S.; Chung, J.H.; Hahm, S.H.; Park, M.J.; Lee, Y.R.; Ko, S.I.; Kang, L.W.; Kim, T.S.; Kim, J.; Han, Y.S. Ribosomal protein S3 is phosphorylated by Cdk1/cdc2 during G2/M phase. BMB Rep. 2011, 44, 529–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.S.; Kim, H.D.; Kim, J. PKCdelta-dependent functional switch of rpS3 between translation and DNA repair. Biochim. Biophys. Acta 2009, 1793, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.D.; Lee, J.Y.; Kim, J. Erk phosphorylates threonine 42 residue of ribosomal protein S3. Biochem. Biophys. Res. Commun. 2005, 333, 110–115. [Google Scholar] [CrossRef]
- Kim, T.S.; Kim, H.D.; Shin, H.S.; Kim, J. Phosphorylation status of nuclear ribosomal protein S3 is reciprocally regulated by protein kinase C{delta} and protein phosphatase 2A. J. Biol. Chem. 2009, 284, 21201–21208. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Chubatsu, L.S.; Admon, A.; Stahl, J.; Fellous, R.; Linn, S. Implication of mammalian ribosomal protein S3 in the processing of DNA damage. J. Biol. Chem. 1995, 270, 13620–13629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Lee, J.Y.; Kim, J. Characterization of a wide range base-damage-endonuclease activity of mammalian rpS3. Biochem. Biophys. Res. Commun. 2005, 328, 962–967. [Google Scholar] [CrossRef]
- Wilson, D.M., 3rd; Deutsch, W.A.; Kelley, M.R. Drosophila ribosomal protein S3 contains an activity that cleaves DNA at apurinic/apyrimidinic sites. J. Biol. Chem. 1994, 269, 25359–25364. [Google Scholar] [CrossRef]
- Grosheva, A.S.; Zharkov, D.O.; Stahl, J.; Gopanenko, A.V.; Tupikin, A.E.; Kabilov, M.R.; Graifer, D.M.; Karpova, G.G. Recognition but no repair of abasic site in single-stranded DNA by human ribosomal uS3 protein residing within intact 40S subunit. Nucleic Acids Res. 2017, 45, 3833–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, V.; Wang, M.; Deutsch, W.A. Human ribosomal protein S3 interacts with DNA base excision repair proteins hAPE/Ref-1 and hOGG1. Biochemistry 2004, 43, 14211–14217. [Google Scholar] [CrossRef]
- Ko, S.I.; Park, J.H.; Park, M.J.; Kim, J.; Kang, L.W.; Han, Y.S. Human ribosomal protein S3 (hRpS3) interacts with uracil-DNA glycosylase (hUNG) and stimulates its glycosylase activity. Mutat. Res. 2008, 648, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Kim, S.H.; Kim, T.S.; Lee, S.M.; Cho, B.S.; Seo, C.I.; Kim, H.D.; Kim, J. Ribosomal protein S3 associates with the TFIIH complex and positively regulates nucleotide excision repair. Cell Mol. Life Sci. 2021, 78, 3591–3606. [Google Scholar] [CrossRef]
- Patil, A.V.; Hsieh, T.S. Ribosomal Protein S3 Negatively Regulates Unwinding Activity of RecQ-like Helicase 4 through Their Physical Interaction. J. Biol. Chem. 2017, 292, 4313–4325. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Kwon, I.S.; Park, J.; Lee, K.H.; Ahn, Y.; Lee, C.; Kim, J.; Choi, S.Y.; Cho, S.W.; Ahn, J.Y. Ribosomal protein S3, a new substrate of Akt, serves as a signal mediator between neuronal apoptosis and DNA repair. J. Biol. Chem. 2010, 285, 29457–29468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, C.Y.; Lee, J.Y.; Kim, J. RpS3, a DNA repair endonuclease and ribosomal protein, is involved in apoptosis. FEBS Lett. 2004, 560, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Rebollo, M.; Mateo, F.; Franke, K.; Huen, M.S.; Lopitz-Otsoa, F.; Rodriguez, M.S.; Plans, V.; Thomson, T.M. Nucleolar exit of RNF8 and BRCA1 in response to DNA damage. Exp. Cell Res. 2012, 318, 2365–2376. [Google Scholar] [CrossRef]
- Yang, C.; Zang, W.; Ji, Y.; Li, T.; Yang, Y.; Zheng, X. Ribosomal protein L6 (RPL6) is recruited to DNA damage sites in a poly(ADP-ribose) polymerase-dependent manner and regulates the DNA damage response. J. Biol. Chem. 2019, 294, 2827–2838. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; He, H.; Ma, Y.; Xu, J.; Chen, G.; Sun, Y.; Xiong, X. Inactivation of ribosomal protein S27-like impairs DNA interstrand cross-link repair by destabilization of FANCD2 and FANCI. Cell Death Dis. 2020, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Vascotto, C.; Fantini, D.; Romanello, M.; Cesaratto, L.; Deganuto, M.; Leonardi, A.; Radicella, J.P.; Kelley, M.R.; D’Ambrosio, C.; Scaloni, A.; et al. APE1/Ref-1 interacts with NPM1 within nucleoli and plays a role in the rRNA quality control process. Mol. Cell Biol. 2009, 29, 1834–1854. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, D.T.; Deutsch, W.A.; Derda, D.; Kelley, M.R. Drosophila AP3, a presumptive DNA repair protein, is homologous to human ribosomal associated protein P0. Nucleic Acids Res. 1991, 19, 4297. [Google Scholar] [CrossRef] [PubMed]
- Molavi, G.; Samadi, N.; Hosseingholi, E.Z. The roles of moonlight ribosomal proteins in the development of human cancers. J. Cell Physiol. 2019, 234, 8327–8341. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Crescenzi, E.; Sagar, V.; Loreni, F.; Russo, A.; Russo, G. Human rpL3 plays a crucial role in cell response to nucleolar stress induced by 5-FU and L-OHP. Oncotarget 2014, 5, 11737–11751. [Google Scholar] [CrossRef] [Green Version]
- Ting, N.S.; Kao, P.N.; Chan, D.W.; Lintott, L.G.; Lees-Miller, S.P. DNA-dependent protein kinase interacts with antigen receptor response element binding proteins NF90 and NF45. J. Biol. Chem. 1998, 273, 2136–2145. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.; Tomimatsu, N.; Richard, D.J.; Cluet, D.; Burma, S.; Khanna, K.K.; Jalinot, P. INT6/EIF3E interacts with ATM and is required for proper execution of the DNA damage response in human cells. Cancer Res. 2012, 72, 2006–2016. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.; Tomimatsu, N.; Burma, S.; Jalinot, P. INT6/EIF3E Controls the RNF8-Dependent Ubiquitylation Pathway and Facilitates DNA Double-Strand Break Repair in Human Cells. Cancer Res. 2016, 76, 6054–6065. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.; Dellaire, G. The Role of the COP9 Signalosome and Neddylation in DNA Damage Signaling and Repair. Biomolecules 2015, 5, 2388–2416. [Google Scholar] [CrossRef] [Green Version]
- Abdulkina, L.R.; Kobayashi, C.; Lovell, J.T.; Chastukhina, I.B.; Aklilu, B.B.; Agabekian, I.A.; Suescun, A.V.; Valeeva, L.R.; Nyamsuren, C.; Aglyamova, G.V.; et al. Components of the ribosome biogenesis pathway underlie establishment of telomere length set poInt. in Arabidopsis. Nat. Commun. 2019, 10, 5479. [Google Scholar] [CrossRef] [Green Version]
- Buchsbaum, S.; Morris, C.; Bochard, V.; Jalinot, P. Human INT6 interacts with MCM7 and regulates its stability during S phase of the cell cycle. Oncogene 2007, 26, 5132–5144. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.L.; Liu, C.D.; You, R.I.; Ching, Y.H.; Liang, J.; Ke, L.; Chen, Y.L.; Chen, H.C.; Hsu, H.J.; Liou, J.W.; et al. Ribosome Protein L4 is essential for Epstein-Barr Virus Nuclear Antigen 1 function. Proc. Natl. Acad. Sci. USA 2016, 113, 2229–2234. [Google Scholar] [CrossRef] [Green Version]
- Park, B.J.; Kang, J.W.; Lee, S.W.; Choi, S.J.; Shin, Y.K.; Ahn, Y.H.; Choi, Y.H.; Choi, D.; Lee, K.S.; Kim, S. The haploinsufficient tumor suppressor p18 upregulates p53 via interactions with ATM/ATR. Cell 2005, 120, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.M.; Jeon, Y.; Kim, D.; Jang, H.; Bae, J.S.; Park, M.K.; Kim, H.; Kim, S.; Lee, H. AIMP3 depletion causes genome instability and loss of stemness in mouse embryonic stem cells. Cell Death Dis. 2018, 9, 972. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, S.; Oh, Y.; Park, S.; Jeon, Y.; Kim, H.; Lee, H.; Kim, S. AIMP3 Deletion Induces Acute Radiation Syndrome-like Phenotype in Mice. Sci. Rep. 2018, 8, 15025. [Google Scholar] [CrossRef] [Green Version]
- Han, J.M.; Park, B.J.; Park, S.G.; Oh, Y.S.; Choi, S.J.; Lee, S.W.; Hwang, S.K.; Chang, S.H.; Cho, M.H.; Kim, S. AIMP2/p38, the scaffold for the multi-tRNA synthetase complex, responds to genotoxic stresses via p53. Proc. Natl. Acad. Sci. USA 2008, 105, 11206–11211. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.G.; Lee, J.Y.; Lee, J.H.; Cho, H.Y.; Kang, B.S.; Jang, S.Y.; Kim, M.H.; Guo, M.; Han, J.M.; Kim, S.J.; et al. Oncogenic Mutation of AIMP2/p38 Inhibits Its Tumor-Suppressive Interaction with Smurf2. Cancer Res. 2016, 76, 3422–3436. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Liu, B.Q.; Li, H.; McGarrigle, D.; Xing, B.W.; Zhou, M.T.; Wang, Z.; Zhang, J.J.; Huang, X.Y.; Guo, L. C-terminal Src kinase (Csk)-mediated phosphorylation of eukaryotic elongation factor 2 (eEF2) promotes proteolytic cleavage and nuclear translocation of eEF2. J. Biol. Chem. 2014, 289, 12666–12678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rugjee, K.N.; Roy Chaudhury, S.; Al-Jubran, K.; Ramanathan, P.; Matina, T.; Wen, J.; Brogna, S. Fluorescent protein tagging confirms the presence of ribosomal proteins at Drosophila polytene chromosomes. PeerJ 2013, 12, e15. [Google Scholar] [CrossRef] [Green Version]
- Brogna, S.; Sato, T.A.; Rosbash, M. Ribosome components are associated with sites of transcription. Mol. Cell 2002, 10, 93–104. [Google Scholar] [CrossRef]
- De, S.; Varsally, W.; Falciani, F.; Brogna, S. Ribosomal proteins’ association with transcription sites peaks at tRNA genes in Schizosaccharomyces pombe. RNA 2011, 17, 1713–1726. [Google Scholar] [CrossRef] [Green Version]
- Schroder, P.A.; Moore, M.J. Association of ribosomal proteins with nascent transcripts in S. cerevisiae. RNA 2005, 11, 1521–1529. [Google Scholar] [CrossRef] [Green Version]
- Tasheva, E.S.; Roufa, D.J. Regulation of human RPS14 transcription by intronic antisense RNAs and ribosomal protein S14. Genes Dev. 1995, 9, 304–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, W.Y.; Huang, Y.C.; Liu, L.F.; Chang, L.H.; Tam, M.F. Rpl12p affects the transcription of the PHO pathway high-affinity inorganic phosphate transporters and repressible phosphatases. Yeast 2011, 28, 481–493. [Google Scholar] [CrossRef]
- Ni, J.Q.; Liu, L.P.; Hess, D.; Rietdorf, J.; Sun, F.L. Drosophila ribosomal proteins are associated with linker histone H1 and suppress gene transcription. Genes Dev. 2006, 20, 1959–1973. [Google Scholar] [CrossRef] [Green Version]
- Hollmuller, E.; Greiner, K.; Kienle, S.M.; Scheffner, M.; Marx, A.; Stengel, F. Interactome of Site-Specifically Acetylated Linker Histone H1. J. Proteome Res. 2021, 20, 4443–4451. [Google Scholar] [CrossRef] [PubMed]
- Kalashnikova, A.A.; Winkler, D.D.; McBryant, S.J.; Henderson, R.K.; Herman, J.A.; DeLuca, J.G.; Luger, K.; Prenni, J.E.; Hansen, J.C. Linker histone H1.0 interacts with an extensive network of proteins found in the nucleolus. Nucleic Acids Res. 2013, 41, 4026–4035. [Google Scholar] [CrossRef]
- Swenson, J.M.; Colmenares, S.U.; Strom, A.R.; Costes, S.V.; Karpen, G.H. The composition and organization of Drosophila heterochromatin are heterogeneous and dynamic. Elife 2016, 5, e16096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, J.; Eskeland, R.; Raffa, G.D.; Kremmer, E.; Imhof, A. Drosophila HP1c is regulated by an auto-regulatory feedback loop through its binding partner Woc. PLoS ONE 2009, 4, e5089. [Google Scholar] [CrossRef] [Green Version]
- De Mateo, S.; Castillo, J.; Estanyol, J.M.; Ballesca, J.L.; Oliva, R. Proteomic characterization of the human sperm nucleus. Proteomics 2011, 11, 2714–2726. [Google Scholar] [CrossRef]
- Dieci, G.; Ruotolo, R.; Braglia, P.; Carles, C.; Carpentieri, A.; Amoresano, A.; Ottonello, S. Positive modulation of RNA polymerase III transcription by ribosomal proteins. Biochem. Biophys. Res. Commun. 2009, 379, 489–493. [Google Scholar] [CrossRef]
- Dai, M.S.; Sun, X.X.; Lu, H. Ribosomal protein L11 associates with c-Myc at 5 S rRNA and tRNA genes and regulates their expression. J. Biol. Chem. 2010, 285, 12587–12594. [Google Scholar] [CrossRef] [Green Version]
- Hermann-Le Denmat, S.; Sipiczki, M.; Thuriaux, P. Suppression of yeast RNA polymerase III mutations by the URP2 gene encoding a protein homologous to the mammalian ribosomal protein S20. J. Mol. Biol. 1994, 240, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.S.; Arnold, H.; Sun, X.X.; Sears, R.; Lu, H. Inhibition of c-Myc activity by ribosomal protein L11. EMBO J. 2007, 26, 3332–3345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hao, Q.; Liao, J.M.; Liao, P.; Lu, H. Ribosomal protein S14 negatively regulates c-Myc activity. J. Biol. Chem. 2013, 288, 21793–21801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, A.; Russo, G. Ribosomal Proteins Control or Bypass p53 during Nucleolar Stress. Int. J. Mol. Sci. 2017, 18, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, K.; Coutts, M.; Stahl, J.; Sytkowski, A.J. Novel interaction between the transcription factor CHOP (GADD153) and the ribosomal protein FTE/S3a modulates erythropoiesis. J. Biol. Chem. 2000, 275, 7591–7596. [Google Scholar] [CrossRef] [Green Version]
- Kashuba, E.; Yurchenko, M.; Szirak, K.; Stahl, J.; Klein, G.; Szekely, L. Epstein-Barr virus-encoded EBNA-5 binds to Epstein-Barr virus-induced Fte1/S3a protein. Exp. Cell Res. 2005, 303, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Sakamoto, S.; Taniguchi, T. Inhibition of poly(ADP-ribose) polymerase activity by Bcl-2 in association with the ribosomal protein S3a. Biochemistry 2002, 41, 929–934. [Google Scholar] [CrossRef]
- Dionne, K.L.; Bergeron, D.; Landry-Voyer, A.M.; Bachand, F. The 40S ribosomal protein uS5 (RPS2) assembles into an extraribosomal complex with human ZNF277 that competes with the PRMT3-uS5 interaction. J. Biol. Chem. 2019, 294, 1944–1955. [Google Scholar] [CrossRef] [Green Version]
- Tchorzewski, M.; Boldyreff, B.; Grankowski, N. Extraribosomal function of the acidic ribosomal P1-protein YP1alpha from Saccharomyces cerevisiae. Acta Biochim. Pol. 1999, 46, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.; Han, S.; Wu, J.; Zhang, Y.; Bai, M.; Abdullah, S.W.; Sun, S.; Guo, H. Ribosomal Protein L13 Participates in Innate Immune Response Induced by Foot-and-Mouth Disease Virus. Front. Immunol. 2021, 12, 616402. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P. Regulated nuclear trafficking of rpL10A mediated by NIK1 represents a defense strategy of plant cells against virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorzatto, C.; Machado, J.P.; Lopes, K.V.; Nascimento, K.J.; Pereira, W.A.; Brustolini, O.J.; Reis, P.A.; Calil, I.P.; Deguchi, M.; Sachetto-Martins, G.; et al. NIK1-mediated translation suppression functions as a plant antiviral immunity mechanism. Nature 2015, 520, 679–682. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Arese, M.; Gualandris, A.; Rifkin, D.B. Intracellular association of FGF-2 with the ribosomal protein L6/TAXREB107. Biochem. Biophys. Res. Commun. 1998, 252, 524–528. [Google Scholar] [CrossRef]
- Berghofer-Hochheimer, Y.; Zurek, C.; Wolfl, S.; Hemmerich, P.; Munder, T. L7 protein is a coregulator of vitamin D receptor-retinoid X receptor-mediated transactivation. J. Cell Biochem. 1998, 69, 1–12. [Google Scholar] [CrossRef]
- Gray, J.P.; Davis, J.W., 2nd; Gopinathan, L.; Leas, T.L.; Nugent, C.A.; Vanden Heuvel, J.P. The ribosomal protein rpL11 associates with and inhibits the transcriptional activity of peroxisome proliferator-activated receptor-alpha. Toxicol. Sci. 2006, 89, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imafuku, I.; Masaki, T.; Waragai, M.; Takeuchi, S.; Kawabata, M.; Hirai, S.; Ohno, S.; Nee, L.E.; Lippa, C.F.; Kanazawa, I.; et al. Presenilin 1 suppresses the function of c-Jun homodimers via interaction with QM/Jif-1. J. Cell Biol. 1999, 147, 121–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, F.; Anderson, D.E.; Barnitz, R.A.; Snow, A.; Bidere, N.; Zheng, L.; Hegde, V.; Lam, L.T.; Staudt, L.M.; Levens, D.; et al. Ribosomal protein S3: A KH domain subunit in NF-kappaB complexes that mediates selective gene regulation. Cell 2007, 131, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-kappaB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhang, Y.; Li, H.; Zhou, Y.; Zhang, Q.; Chen, R.; Jin, T.; Hu, K.; Li, S.; Wang, Y.; et al. Vitamin D promotes the cisplatin sensitivity of oral squamous cell carcinoma by inhibiting LCN2-modulated NF-kappaB pathway activation through RPS3. Cell Death Dis. 2019, 10, 936. [Google Scholar] [CrossRef]
- Wan, F.; Weaver, A.; Gao, X.; Bern, M.; Hardwidge, P.R.; Lenardo, M.J. IKKbeta phosphorylation regulates RPS3 nuclear translocation and NF-kappaB function during infection with Escherichia coli strain O157:H7. Nat. Immunol. 2011, 12, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.J.; Youn, H.; Seong, K.M.; Jin, Y.W.; Kim, J.; Youn, B. Phosphorylation of ribosomal protein S3 and antiapoptotic TRAF2 protein mediates radioresistance in non-small cell lung cancer cells. J. Biol. Chem. 2013, 288, 2965–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; El Qaidi, S.; Hardwidge, P.R. SseL Deubiquitinates RPS3 to Inhibit Its Nuclear Translocation. Pathogens 2018, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Yadavilli, S.; Mayo, L.D.; Higgins, M.; Lain, S.; Hegde, V.; Deutsch, W.A. Ribosomal protein S3: A multi-functional protein that interacts with both p53 and MDM2 through its KH domain. DNA Repair 2009, 8, 1215–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.H.; Leslie, P.; Zhang, Y. Ribosomal proteins as unrevealed caretakers for cellular stress and genomic instability. Oncotarget 2014, 5, 860–871. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Li, Y.; Dai, M.S.; Sun, X.X. Ribosomal protein L4 is a novel regulator of the MDM2-p53 loop. Oncotarget 2016, 7, 16217–16226. [Google Scholar] [CrossRef]
- Cho, J.; Park, J.; Shin, S.C.; Kim, J.H.; Kim, E.E.; Song, E.J. Ribosomal protein S2 interplays with MDM2 to induce p53. Biochem. Biophys. Res. Commun. 2020, 523, 542–547. [Google Scholar] [CrossRef]
- Cho, J.; Park, J.; Shin, S.C.; Jang, M.; Kim, J.H.; Kim, E.E.; Song, E.J. USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2. Cancers 2020, 12, 1137. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Zhang, J.; Xiao, W.; Zheng, X. Regulation of the HDM2-p53 pathway by ribosomal protein L6 in response to ribosomal stress. Nucleic Acids Res. 2014, 42, 1799–1811. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Poyurovsky, M.V.; Li, Y.; Biderman, L.; Stahl, J.; Jacq, X.; Prives, C. Ribosomal protein S7 is both a regulator and a substrate of MDM2. Mol. Cell 2009, 35, 316–326. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; DeVine, T.; Challagundla, K.B.; Dai, M.S. Interplay between ribosomal protein S27a and MDM2 protein in p53 activation in response to ribosomal stress. J. Biol. Chem. 2011, 286, 22730–22741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, B.; Fang, Z.; Liao, P.; Zhou, X.; Xiong, J.; Zeng, S.; Lu, H. Cancer-mutated ribosome protein L22 (RPL22/eL22) suppresses cancer cell survival by blocking p53-MDM2 circuit. Oncotarget 2017, 8, 90651–90661. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Deisenroth, C.; Zhang, Y. RP-MDM2-p53 Pathway: Linking Ribosomal Biogenesis and Tumor Surveillance. Trends Cancer 2016, 2, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.T.; Chen, T.Y.; Weng, C.W.; Yang, C.H.; Tang, M.S. Acrolein preferentially damages nucleolus eliciting ribosomal stress and apoptosis in human cancer cells. Oncotarget 2016, 7, 80450–80464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahata, B.; Sundqvist, A.; Xirodimas, D.P. Recruitment of RPL11 at promoter sites of p53-regulated genes upon nucleolar stress through NEDD8 and in an Mdm2-dependent manner. Oncogene 2012, 31, 3060–3071. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, A.; Liu, G.; Mirsaliotis, A.; Xirodimas, D.P. Regulation of nucleolar signalling to p53 through NEDDylation of L11. EMBO Rep. 2009, 10, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Ebina, M.; Tsuruta, F.; Katoh, M.C.; Kigoshi, Y.; Someya, A.; Chiba, T. Myeloma overexpressed 2 (Myeov2) regulates L11 subnuclear localization through Nedd8 modification. PLoS ONE 2013, 8, e65285. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.S.; Challagundla, K.B.; Sun, X.X.; Palam, L.R.; Zeng, S.X.; Wek, R.C.; Lu, H. Physical and functional interaction between ribosomal protein L11 and the tumor suppressor ARF. J. Biol. Chem. 2012, 287, 17120–17129. [Google Scholar] [CrossRef] [Green Version]
- Kayama, K.; Watanabe, S.; Takafuji, T.; Tsuji, T.; Hironaka, K.; Matsumoto, M.; Nakayama, K.I.; Enari, M.; Kohno, T.; Shiraishi, K.; et al. GRWD1 negatively regulates p53 via the RPL11-MDM2 pathway and promotes tumorigenesis. EMBO Rep. 2017, 18, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Z.; Cao, B.; Liao, J.M.; Deng, J.; Plummer, K.D.; Liao, P.; Liu, T.; Zhang, W.; Zhang, K.; Li, L.; et al. SPIN1 promotes tumorigenesis by blocking the uL18 (universal large ribosomal subunit protein 18)-MDM2-p53 pathway in human cancer. Elife 2018, 7, e31275. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Li, L.; Lou, H.; Sun, H.; Ngai, S.M.; Shao, G.; Tang, J. The ribosomal protein S26 regulates p53 activity in response to DNA damage. Oncogene 2014, 33, 2225–2235. [Google Scholar] [CrossRef] [Green Version]
- Llanos, S.; Serrano, M. Depletion of ribosomal protein L37 occurs in response to DNA damage and activates p53 through the L11/MDM2 pathway. Cell Cycle 2010, 9, 4005–4012. [Google Scholar] [CrossRef]
- Lindstrom, M.S.; Nister, M. Silencing of ribosomal protein S9 elicits a multitude of cellular responses inhibiting the growth of cancer cells subsequent to p53 activation. PLoS ONE 2010, 5, e9578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Zhao, Y.; He, H.; Sun, Y. Ribosomal protein S27-like and S27 interplay with p53-MDM2 axis as a target, a substrate and a regulator. Oncogene 2011, 30, 1798–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecoraro, A.; Carotenuto, P.; Russo, G.; Russo, A. Ribosomal protein uL3 targets E2F1 and Cyclin D1 in cancer cell response to nucleolar stress. Sci. Rep. 2019, 9, 15431. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Hao, Q.; Zhang, Q.; Liao, J.M.; Ke, J.W.; Liao, P.; Cao, B.; Lu, H. Ribosomal proteins L11 and L5 activate TAp73 by overcoming MDM2 inhibition. Cell Death Differ. 2015, 22, 755–766. [Google Scholar] [CrossRef]
- Gao, M.; Li, X.; Dong, W.; Jin, R.; Ma, H.; Yang, P.; Hu, M.; Li, Y.; Hao, Y.; Yuan, S.; et al. Ribosomal protein S7 regulates arsenite-induced GADD45alpha expression by attenuating MDM2-mediated GADD45alpha ubiquitination and degradation. Nucleic Acids Res. 2013, 41, 5210–5222. [Google Scholar] [CrossRef]
- Coleno-Costes, A.; Jang, S.M.; de Vanssay, A.; Rougeot, J.; Bouceba, T.; Randsholt, N.B.; Gibert, J.M.; Le Crom, S.; Mouchel-Vielh, E.; Bloyer, S.; et al. New partners in regulation of gene expression: The enhancer of Trithorax and Polycomb Corto interacts with methylated ribosomal protein l12 via its chromodomain. PLoS Genet. 2012, 8, e1003006. [Google Scholar] [CrossRef]
- Grunchec, H.; Deraze, J.; Dardalhon-Cumenal, D.; Ribeiro, V.; Coleno-Costes, A.; Dias, K.; Bloyer, S.; Mouchel-Vielh, E.; Peronnet, F.; Thomassin, H. Single amino-acid mutation in the Drosophila melanogaster ribosomal protein uL11: An insight in its transcriptional activity. bioRxiv 2021, 456895. [Google Scholar] [CrossRef]
- Frolov, M.V.; Birchler, J.A. Mutation in P0, a dual function ribosomal protein/apurinic/apyrimidinic endonuclease, modifies gene expression and position effect variegation in Drosophila. Genetics 1998, 150, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Kim, S.; Shin, Y.J.; Hur, Y.S.; Kim, W.Y.; Lee, M.S.; Cheon, C.I.; Verma, D.P. Ribosomal protein S6, a target of rapamycin, is involved in the regulation of rRNA genes by possible epigenetic changes in Arabidopsis. J. Biol. Chem. 2014, 289, 3901–3912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, O.; Kim, S.; Shin, Y.J.; Kim, W.Y.; Koh, H.J.; Cheon, C.I. Identification of nucleosome assembly protein 1 (NAP1) as an interacting partner of plant ribosomal protein S6 (RPS6) and a positive regulator of rDNA transcription. Biochem. Biophys. Res. Commun. 2015, 465, 200–205. [Google Scholar] [CrossRef]
- Proshkin, S.A.; Shematorova, E.K.; Souslova, E.A.; Proshkina, G.M.; Shpakovski, G.V. A minor isoform of the human RNA polymerase II subunit hRPB11 (POLR2J) interacts with several components of the translation initiation factor eIF3. Biochemistry 2011, 76, 976–980. [Google Scholar] [CrossRef]
- Harel-Sharvit, L.; Eldad, N.; Haimovich, G.; Barkai, O.; Duek, L.; Choder, M. RNA polymerase II subunits link transcription and mRNA decay to translation. Cell 2010, 143, 552–563. [Google Scholar] [CrossRef] [Green Version]
- Moller, A.; Xie, S.Q.; Hosp, F.; Lang, B.; Phatnani, H.P.; James, S.; Ramirez, F.; Collin, G.B.; Naggert, J.K.; Babu, M.M.; et al. Proteomic analysis of mitotic RNA polymerase II reveals novel interactors and association with proteins dysfunctional in disease. Mol. Cell Proteom. 2012, 11, M111011767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smekalova, E.M.; Gerashchenko, M.V.; O’Connor, P.B.F.; Whittaker, C.A.; Kauffman, K.J.; Fefilova, A.S.; Zatsepin, T.S.; Bogorad, R.L.; Baranov, P.V.; Langer, R.; et al. In Vivo RNAi-Mediated eIF3m Knockdown Affects Ribosome Biogenesis and Transcription but Has Limited Impact on mRNA-Specific Translation. Mol. Ther. Nucleic Acids 2020, 19, 252–266. [Google Scholar] [CrossRef]
- Seither, P.; Iben, S.; Thiry, M.; Grummt, I. PAF67, a novel protein that is associated with the initiation-competent form of RNA polymerase I. Biol. Chem. 2001, 382, 1163–1170. [Google Scholar] [CrossRef]
- Yuan, X.; Zhao, J.; Zentgraf, H.; Hoffmann-Rohrer, U.; Grummt, I. Multiple interactions between RNA polymerase I, TIF-IA and TAF(I) subunits regulate preinitiation complex assembly at the ribosomal gene promoter. EMBO Rep. 2002, 3, 1082–1087. [Google Scholar] [CrossRef] [Green Version]
- DuRose, J.B.; Scheuner, D.; Kaufman, R.J.; Rothblum, L.I.; Niwa, M. Phosphorylation of eukaryotic translation initiation factor 2alpha coordinates rRNA transcription and translation inhibition during endoplasmic reticulum stress. Mol. Cell Biol. 2009, 29, 4295–4307. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, C.C.; Mata, J.; Crane, R.F.; Thomas, B.; Akoulitchev, A.; Bahler, J.; Norbury, C.J. Activation of AP-1-dependent transcription by a truncated translation initiation factor. Eukaryot Cell 2005, 4, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- Esteves, P.; Dard, L.; Brillac, A.; Hubert, C.; Sarlak, S.; Rousseau, B.; Dumon, E.; Izotte, J.; Bonneu, M.; Lacombe, D.; et al. Nuclear control of lung cancer cells migration, invasion and bioenergetics by eukaryotic translation initiation factor 3F. Oncogene 2020, 39, 617–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Feng, Y.; Goulet, A.C.; Vaillancourt, R.R.; Sachs, N.A.; Hershey, J.W.; Nelson, M.A. The p34cdc2-related cyclin-dependent kinase 11 interacts with the p47 subunit of eukaryotic initiation factor 3 during apoptosis. J. Biol. Chem. 2003, 278, 5062–5071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Hershey, J.W.; Nelson, M.A. Phosphorylation of the eukaryotic initiation factor 3f by cyclin-dependent kinase 11 during apoptosis. FEBS Lett. 2009, 583, 971–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daxinger, L.; Oey, H.; Apedaile, A.; Sutton, J.; Ashe, A.; Whitelaw, E. A forward genetic screen identifies eukaryotic translation initiation factor 3, subunit H (eIF3h), as an enhancer of variegation in the mouse. G3 2012, 2, 1393–1396. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fu, L.; Li, J.B.; Qin, Y.; Zeng, T.T.; Zhou, J.; Zeng, Z.L.; Chen, J.; Cao, T.T.; Ban, X.; et al. Increased expression of EIF5A2, via hypoxia or gene amplification, contributes to metastasis and angiogenesis of esophageal squamous cell carcinoma. Gastroenterology 2014, 146, 1701–1713. [Google Scholar] [CrossRef]
- Sanders, J.; Brandsma, M.; Janssen, G.M.; Dijk, J.; Moller, W. Immunofluorescence studies of human fibroblasts demonstrate the presence of the complex of elongation factor-1 beta gamma delta in the endoplasmic reticulum. J. Cell Sci. 1996, 109 Pt 5, 1113–1117. [Google Scholar] [CrossRef]
- Vera, M.; Pani, B.; Griffiths, L.A.; Muchardt, C.; Abbott, C.M.; Singer, R.H.; Nudler, E. The translation elongation factor eEF1A1 couples transcription to translation during heat shock response. Elife 2014, 16, 3164. [Google Scholar] [CrossRef]
- Wu-Baer, F.; Lane, W.S.; Gaynor, R.B. Identification of a group of cellular cofactors that stimulate the binding of RNA polymerase II and TRP-185 to human immunodeficiency virus 1 TAR RNA. J. Biol. Chem. 1996, 271, 4201–4208. [Google Scholar] [CrossRef] [Green Version]
- Billaut-Mulot, O.; Fernandez-Gomez, R.; Loyens, M.; Ouaissi, A. Trypanosoma cruzi elongation factor 1-alpha: Nuclear localization in parasites undergoing apoptosis. Gene 1996, 174, 19–26. [Google Scholar] [CrossRef]
- Zolotarev, N.A.; Maksimenko, O.G.; Shidlovskii, Y.V.; Georgiev, P.G.; Bonchuk, A.N. [Translation elongation factor EF1α1 interacts with ZAD domains of transcription factors from Drosophila melanogaster]. Mol. Biol. 2016, 50, 1014–1019. [Google Scholar] [CrossRef]
- Maruyama, T.; Nara, K.; Yoshikawa, H.; Suzuki, N. Txk, a member of the non-receptor tyrosine kinase of the Tec family, forms a complex with poly(ADP-ribose) polymerase 1 and elongation factor 1alpha and regulates interferon-gamma gene transcription in Th1 cells. Clin. Exp. Immunol. 2007, 147, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Gangwani, L.; Mikrut, M.; Galcheva-Gargova, Z.; Davis, R.J. Interaction of ZPR1 with translation elongation factor-1alpha in proliferating cells. J. Cell Biol. 1998, 143, 1471–1484. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Gangwani, L.; Davis, R.J.; Lambright, D.G. Structural insights into the interaction of the evolutionarily conserved ZPR1 domain tandem with eukaryotic EF1A, receptors, and SMN complexes. Proc. Natl. Acad. Sci. USA 2007, 104, 13930–13935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzi, M.; Bavelloni, A.; Faenza, I.; Blalock, W.; Urbani, A.; D’Aguanno, S.; Fiume, R.; Ramazzotti, G.; Maraldi, N.M.; Cocco, L. eEF1A phosphorylation in the nucleus of insulin-stimulated C2C12 myoblasts: Ser(5)(3) is a novel substrate for protein kinase C betaI. Mol. Cell Proteom. 2010, 9, 2719–2728. [Google Scholar] [CrossRef] [Green Version]
- Negrutskii, B. Non-translational Connections of eEF1B in the Cytoplasm and Nucleus of Cancer Cells. Front. Mol. Biosci. 2020, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbi, N.; Batassa, E.M.; Pisani, C.; Onori, A.; Di Certo, M.G.; Strimpakos, G.; Fanciulli, M.; Mattei, E.; Passananti, C. The eEF1gamma subunit contacts RNA polymerase II and binds vimentin promoter region. PLoS ONE 2010, 5, e14481. [Google Scholar] [CrossRef] [Green Version]
- Pisani, C.; Onori, A.; Gabanella, F.; Delle Monache, F.; Borreca, A.; Ammassari-Teule, M.; Fanciulli, M.; Di Certo, M.G.; Passananti, C.; Corbi, N. eEF1Bgamma binds the Che-1 and TP53 gene promoters and their transcripts. J. Exp. Clin. Cancer Res. 2016, 35, 146. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Schlierf, M.; Gaspar, A.C.; Dreux, C.; Kpebe, A.; Chaney, L.; Mathieu, A.; Hitte, C.; Gremy, O.; Sarot, E.; et al. Drosophila translational elongation factor-1gamma is modified in response to DOA kinase activity and is essential for cellular viability. Genetics 2010, 184, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Kaitsuka, T.; Tomizawa, K.; Matsushita, M. Transformation of eEF1Bdelta into heat-shock response transcription factor by alternative splicing. EMBO Rep. 2011, 12, 673–681. [Google Scholar] [CrossRef] [Green Version]
- In, S.; Kim, Y.I.; Lee, J.E.; Kim, J. RNF20/40-mediated eEF1BdeltaL monoubiquitylation stimulates transcription of heat shock-responsive genes. Nucleic Acids Res. 2019, 47, 2840–2855. [Google Scholar] [CrossRef] [PubMed]
- Wanzel, M.; Russ, A.C.; Kleine-Kohlbrecher, D.; Colombo, E.; Pelicci, P.G.; Eilers, M. A ribosomal protein L23-nucleophosmin circuit coordinates Mizl function with cell growth. Nat. Cell Biol. 2008, 10, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Y.; Qu, Y.; Zhang, Q.; Wei, M.; Liu, C.X.; Chen, X.H.; Yan, M.; Zhu, Z.G.; Liu, B.Y.; Chen, G.Q.; et al. Knockdown of metallopanstimulin-1 inhibits NF-kappaB signaling at different levels: The role of apoptosis induction of gastric cancer cells. Int. J. Cancer 2012, 130, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.Q.; Li, C.; Xu, J.D.; Zhao, X.F.; Wang, J.X. RPS27, a sORF-Encoded Polypeptide, Functions Antivirally by Activating the NF-kappaB Pathway and Interacting With Viral Envelope Proteins in Shrimp. Front. Immunol. 2019, 10, 2763. [Google Scholar] [CrossRef]
- Lim, K.H.; Kim, K.H.; Choi, S.I.; Park, E.S.; Park, S.H.; Ryu, K.; Park, Y.K.; Kwon, S.Y.; Yang, S.I.; Lee, H.C.; et al. RPS3a over-expressed in HBV-associated hepatocellular carcinoma enhances the HBx-induced NF-kappaB signaling via its novel chaperoning function. PLoS ONE 2011, 6, e22258. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Xu, S.; Zhang, X.; He, J.; Yan, D.; Yang, Z.; Xiao, S. Ribosomal protein RPL41 induces rapid degradation of ATF4, a transcription factor critical for tumour cell survival in stress. J. Pathol. 2011, 225, 285–292. [Google Scholar] [CrossRef]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Xu, S.; Wu, X.; Zhang, X.; Chen, C.; Chen, H.; She, F. CagA orchestrates eEF1A1 and PKCdelta to induce interleukin-6 expression in Helicobacter pylori-infected gastric epithelial cells. Gut Pathog. 2020, 12, 31. [Google Scholar] [CrossRef]
- Chen, L.; Uchida, K.; Endler, A.; Shibasaki, F. Mammalian tumor suppressor Int6 specifically targets hypoxia inducible factor 2 alpha for degradation by hypoxia- and pVHL-independent regulation. J. Biol. Chem. 2007, 282, 12707–12716. [Google Scholar] [CrossRef] [Green Version]
- Moretti, J.; Chastagner, P.; Gastaldello, S.; Heuss, S.F.; Dirac, A.M.; Bernards, R.; Masucci, M.G.; Israel, A.; Brou, C. The translation initiation factor 3f (eIF3f) exhibits a deubiquitinase activity regulating Notch activation. PLoS Biol. 2010, 8, e1000545. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Zhou, H.; Ponzoni, L.; Luo, A.; Zhu, R.; He, M.; Huang, Y.; Guan, K.L.; Bahar, I.; Liu, Z.; et al. EIF3H Orchestrates Hippo Pathway-Mediated Oncogenesis via Catalytic Control of YAP Stability. Cancer Res. 2020, 80, 2550–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Zhu, R.; Luo, A.; Zhou, H.; Ding, F.; Yang, H.; Liu, Z. EIF3H promotes aggressiveness of esophageal squamous cell carcinoma by modulating Snail stability. J. Exp. Clin. Cancer Res. 2020, 39, 175. [Google Scholar] [CrossRef] [PubMed]
- Khacho, M.; Mekhail, K.; Pilon-Larose, K.; Pause, A.; Cote, J.; Lee, S. eEF1A is a novel component of the mammalian nuclear protein export machinery. Mol. Biol. Cell 2008, 19, 5296–5308. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ng, N.K.; Koon, A.C.; Chan, H.Y. Expanded polyalanine tracts function as nuclear export signals and promote protein mislocalization via eEF1A1 factor. J. Biol. Chem. 2017, 292, 5784–5800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, M.; Vaquie, A.; Nocera, G.; Heller, M.; Stumpe, M.; Siva Sankar, D.; Dengjel, J.; Meijer, D.; Yamaguchi, T.; Matthias, P.; et al. EEF1A1 deacetylation enables transcriptional activation of remyelination. Nat. Commun. 2020, 11, 3420. [Google Scholar] [CrossRef]
- Mingot, J.M.; Vega, S.; Cano, A.; Portillo, F.; Nieto, M.A. eEF1A mediates the nuclear export of SNAG-containing proteins via the Exportin5-aminoacyl-tRNA complex. Cell Rep. 2013, 5, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calado, A.; Treichel, N.; Muller, E.C.; Otto, A.; Kutay, U. Exportin-5-mediated nuclear export of eukaryotic elongation factor 1A and tRNA. EMBO J. 2002, 21, 6216–6224. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Hopper, A.K. In vivo biochemical analyses reveal distinct roles of beta-importins and eEF1A in tRNA subcellular traffic. Genes Dev. 2015, 29, 772–783. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Shi, Y.; Truong, L.N.; Fisch, K.M.; Xu, T.; Gardiner, E.; Fu, G.; Hsu, Y.O.; Kishi, S.; Su, A.I.; et al. Oxidative stress diverts tRNA synthetase to nucleus for protection against DNA damage. Mol. Cell 2014, 56, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Cui, H.; Shi, Y.; Fu, G.; Rauniyar, N.; Yates, J.R.; Yang, X.-L. Nucleus translocation of tRNA synthetase mediates late integrated stress response. bioRxiv 2020, 138792. [Google Scholar] [CrossRef]
- Cao, X.; Li, C.; Xiao, S.; Tang, Y.; Huang, J.; Zhao, S.; Li, X.; Li, J.; Zhang, R.; Yu, W. Acetylation promotes TyrRS nuclear translocation to prevent oxidative damage. Proc. Natl. Acad. Sci. USA 2017, 114, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Bervoets, S.; Wei, N.; Erfurth, M.L.; Yusein-Myashkova, S.; Ermanoska, B.; Mateiu, L.; Asselbergh, B.; Blocquel, D.; Kakad, P.; Penserga, T.; et al. Transcriptional dysregulation by a nucleus-localized aminoacyl-tRNA synthetase associated with Charcot-Marie-Tooth neuropathy. Nat. Commun. 2019, 10, 5045. [Google Scholar] [CrossRef] [PubMed]
- Harb, M.; Becker, M.M.; Vitour, D.; Baron, C.H.; Vende, P.; Brown, S.C.; Bolte, S.; Arold, S.T.; Poncet, D. Nuclear localization of cytoplasmic poly(A)-binding protein upon rotavirus infection involves the interaction of NSP3 with eIF4G and RoXaN. J. Virol. 2008, 82, 11283–11293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Bhattacharjee, R.B.; Bag, J. Expression of poly(A)-binding protein is upregulated during recovery from heat shock in HeLa cells. FEBS J. 2009, 276, 552–570. [Google Scholar] [CrossRef]
- Salaun, C.; MacDonald, A.I.; Larralde, O.; Howard, L.; Lochtie, K.; Burgess, H.M.; Brook, M.; Malik, P.; Gray, N.K.; Graham, S.V. Poly(A)-binding protein 1 partially relocalizes to the nucleus during herpes simplex virus type 1 infection in an ICP27-independent manner and does not inhibit virus replication. J. Virol. 2010, 84, 8539–8548. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Glaunsinger, B.A. Aberrant herpesvirus-induced polyadenylation correlates with cellular messenger RNA destruction. PLoS Biol. 2009, 7, e1000107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrikova, E.; Shveygert, M.; Walters, R.; Gromeier, M. Herpes simplex virus proteins ICP27 and UL47 associate with polyadenylate-binding protein and control its subcellular distribution. J. Virol. 2010, 84, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Massimelli, M.J.; Majerciak, V.; Kruhlak, M.; Zheng, Z.M. Interplay between polyadenylate-binding protein 1 and Kaposi’s sarcoma-associated herpesvirus ORF57 in accumulation of polyadenylated nuclear RNA, a viral long noncoding RNA. J. Virol. 2013, 87, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Park, R.; El-Guindy, A.; Heston, L.; Lin, S.F.; Yu, K.P.; Nagy, M.; Borah, S.; Delecluse, H.J.; Steitz, J.; Miller, G. Nuclear translocation and regulation of intranuclear distribution of cytoplasmic poly(A)-binding protein are distinct processes mediated by two Epstein Barr virus proteins. PLoS ONE 2014, 9, e92593. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.; Vende, P.; Charpilienne, A.; Gratia, M.; Laroche, C.; Poncet, D. Rotavirus Infection Alters Splicing of the Stress-Related Transcription Factor XBP1. J. Virol. 2019, 93, e01739-18. [Google Scholar] [CrossRef] [Green Version]
- Blakqori, G.; van Knippenberg, I.; Elliott, R.M. Bunyamwera orthobunyavirus S-segment untranslated regions mediate poly(A) tail-independent translation. J. Virol. 2009, 83, 3637–3646. [Google Scholar] [CrossRef] [Green Version]
- Kanno, T.; Sato, Y.; Sata, T.; Katano, H. Expression of Kaposi’s sarcoma-associated herpesvirus-encoded K10/10.1 protein in tissues and its interaction with poly(A)-binding protein. Virology 2006, 352, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Khaperskyy, D.A.; Emara, M.M.; Johnston, B.P.; Anderson, P.; Hatchette, T.F.; McCormick, C. Influenza a virus host shutoff disables antiviral stress-induced translation arrest. PLoS Pathog. 2014, 10, e1004217. [Google Scholar] [CrossRef] [Green Version]
- Montero, H.; Rojas, M.; Arias, C.F.; Lopez, S. Rotavirus infection induces the phosphorylation of eIF2alpha but prevents the formation of stress granules. J. Virol. 2008, 82, 1496–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchemin, C.; Laliberte, J.F. The poly(A) binding protein is internalized in virus-induced vesicles or redistributed to the nucleolus during turnip mosaic virus infection. J. Virol. 2007, 81, 10905–10913. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.A.; Luo, J.M.; Chiang, M.H.; Fang, K.Y.; Li, C.H.; Chen, C.W.; Wang, Y.S.; Chang, C.Y. Nervous Necrosis Virus Coat Protein Mediates Host Translation Shutoff through Nuclear Translocalization and Degradation of Polyadenylate Binding Protein. J. Virol. 2021, 95, e0236420. [Google Scholar] [CrossRef]
- Copeland, A.M.; Altamura, L.A.; Van Deusen, N.M.; Schmaljohn, C.S. Nuclear relocalization of polyadenylate binding protein during rift valley fever virus infection involves expression of the NSs gene. J. Virol. 2013, 87, 11659–11669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borah, S.; Darricarrere, N.; Darnell, A.; Myoung, J.; Steitz, J.A. A viral nuclear noncoding RNA binds re-localized poly(A) binding protein and is required for late KSHV gene expression. PLoS Pathog. 2011, 7, e1002300. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.R.; Glaunsinger, B.A. Nuclear import of cytoplasmic poly(A) binding protein restricts gene expression via hyperadenylation and nuclear retention of mRNA. Mol. Cell Biol. 2010, 30, 4996–5008. [Google Scholar] [CrossRef] [Green Version]
- Burgess, H.M.; Richardson, W.A.; Anderson, R.C.; Salaun, C.; Graham, S.V.; Gray, N.K. Nuclear relocalisation of cytoplasmic poly(A)-binding proteins PABP1 and PABP4 in response to UV irradiation reveals mRNA-dependent export of metazoan PABPs. J. Cell Sci. 2011, 124, 3344–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonina, E.; Stauber, R.; Pavlakis, G.N. The human poly(A)-binding protein 1 shuttles between the nucleus and the cytoplasm. J. Biol. Chem. 1998, 273, 13015–13021. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, N.; Lejeune, F.; Maquat, L.E. Evidence that poly(A) binding protein C1 binds nuclear pre-mRNA poly(A) tails. Mol. Cell Biol. 2006, 26, 3085–3097. [Google Scholar] [CrossRef] [Green Version]
- Burgess, H.M.; Gray, N.K. An integrated model for the nucleo-cytoplasmic transport of cytoplasmic poly(A)-binding proteins. Commun. Integr Biol. 2012, 5, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.; Taylor, M.S.; O’Donnell, K.A.; Boeke, J.D. Poly(A) binding protein C1 is essential for efficient L1 retrotransposition and affects L1 RNP formation. Mol. Cell Biol. 2012, 32, 4323–4336. [Google Scholar] [CrossRef] [Green Version]
- Rivas, H.G.; Schmaling, S.K.; Gaglia, M.M. Shutoff of Host Gene Expression in Influenza A Virus and Herpesviruses: Similar Mechanisms and Common Themes. Viruses 2016, 8, 102. [Google Scholar] [CrossRef]
- Kumar, G.R.; Shum, L.; Glaunsinger, B.A. Importin alpha-mediated nuclear import of cytoplasmic poly(A) binding protein occurs as a direct consequence of cytoplasmic mRNA depletion. Mol. Cell Biol. 2011, 31, 3113–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbertson, S.; Federspiel, J.D.; Hartenian, E.; Cristea, I.M.; Glaunsinger, B. Changes in mRNA abundance drive shuttling of RNA binding proteins, linking cytoplasmic RNA degradation to transcription. Elife 2018, 7, e37663. [Google Scholar] [CrossRef] [PubMed]
- Hartenian, E.; Glaunsinger, B.A. Feedback to the central dogma: Cytoplasmic mRNA decay and transcription are interdependent processes. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 385–398. [Google Scholar] [CrossRef] [Green Version]
- Duncan-Lewis, C.; Hartenian, E.; King, V.; Glaunsinger, B.A. Cytoplasmic mRNA decay represses RNA polymerase II transcription during early apoptosis. Elife 2021, 10, e58342. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.W.; Wang, W.T.; Tsai, T.Y.; Kuo, C.Y.; Li, H.K.; Wu Lee, Y.H. Critical roles of RNA helicase DDX3 and its interactions with eIF4E/PABP1 in stress granule assembly and stress response. Biochem. J. 2012, 441, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copsey, A.C.; Cooper, S.; Parker, R.; Lineham, E.; Lapworth, C.; Jallad, D.; Sweet, S.; Morley, S.J. The helicase, DDX3X, interacts with poly(A)-binding protein 1 (PABP1) and caprin-1 at the leading edge of migrating fibroblasts and is required for efficient cell spreading. Biochem. J. 2017, 474, 3109–3120. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.J.; Kantidakis, T.; Sabe, H.; Critchley, D.R.; Norman, J.C. Interaction of paxillin with poly(A)-binding protein 1 and its role in focal adhesion turnover and cell migration. Mol. Cell Biol. 2005, 25, 3763–3773. [Google Scholar] [CrossRef] [Green Version]
- Eisermann, K.; Dar, J.A.; Dong, J.; Wang, D.; Masoodi, K.Z.; Wang, Z. Poly (A) Binding Protein Cytoplasmic 1 Is a Novel Co-Regulator of the Androgen Receptor. PLoS ONE 2015, 10, e0128495. [Google Scholar] [CrossRef] [Green Version]
- Sen, A.; De Castro, I.; Defranco, D.B.; Deng, F.M.; Melamed, J.; Kapur, P.; Raj, G.V.; Rossi, R.; Hammes, S.R. Paxillin mediates extranuclear and intranuclear signaling in prostate cancer proliferation. J. Clin. Investig. 2012, 122, 2469–2481. [Google Scholar] [CrossRef] [PubMed]
- Kamiyama, T.; Sun, W.; Tani, N.; Nakamura, A.; Niwa, R. Poly(A) Binding Protein Is Required for Nuclear Localization of the Ecdysteroidogenic Transcription Factor Molting Defective in the Prothoracic Gland of Drosophila melanogaster. Front. Genet. 2020, 11, 636. [Google Scholar] [CrossRef]
- Brune, C.; Munchel, S.E.; Fischer, N.; Podtelejnikov, A.V.; Weis, K. Yeast poly(A)-binding protein Pab1 shuttles between the nucleus and the cytoplasm and functions in mRNA export. RNA 2005, 11, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Minvielle-Sebastia, L.; Preker, P.J.; Wiederkehr, T.; Strahm, Y.; Keller, W. The major yeast poly(A)-binding protein is associated with cleavage factor IA and functions in premessenger RNA 3′-end formation. Proc. Natl. Acad. Sci. USA 1997, 94, 7897–7902. [Google Scholar] [CrossRef] [Green Version]
- Khaleghpour, K.; Svitkin, Y.V.; Craig, A.W.; DeMaria, C.T.; Deo, R.C.; Burley, S.K.; Sonenberg, N. Translational repression by a novel partner of human poly(A) binding protein, Paip2. Mol. Cell 2001, 7, 205–216. [Google Scholar] [CrossRef]
- Kachaev, Z.M.; Lebedeva, L.A.; Kozlov, E.N.; Toropygin, I.Y.; Schedl, P.; Shidlovskii, Y.V. Paip2 is localized to active promoters and loaded onto nascent mRNA in Drosophila. Cell Cycle 2018, 17, 1708–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachaev, Z.M.; Lebedeva, L.A.; Shaposhnikov, A.V.; Moresco, J.J.; Yates, J.R., 3rd; Schedl, P.; Shidlovskii, Y.V. Paip2 cooperates with Cbp80 at an active promoter and participates in RNA Polymerase II phosphorylation in Drosophila. FEBS Lett. 2019, 593, 1102–1112. [Google Scholar] [CrossRef]
- Kachaev, Z.M.; Lebedeva, L.A.; Kozlov, E.N.; Shidlovskii, Y.V. Interplay of mRNA capping and transcription machineries. BioSci. Rep. 2020, 40, BSR20192825. [Google Scholar] [CrossRef]
- Wool, I.G. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Lindstrom, M.S. Emerging functions of ribosomal proteins in gene-specific transcription and translation. Biochem. Biophys. Res. Commun. 2009, 379, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R.; McIntosh, K.B. How common are extraribosomal functions of ribosomal proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Malygin, A.A.; Parakhnevitch, N.M.; Ivanov, A.V.; Eperon, I.C.; Karpova, G.G. Human ribosomal protein S13 regulates expression of its own gene at the splicing step by a feedback mechanism. Nucleic Acids Res. 2007, 35, 6414–6423. [Google Scholar] [CrossRef] [Green Version]
- Petibon, C.; Parenteau, J.; Catala, M.; Elela, S.A. Introns regulate the production of ribosomal proteins by modulating splicing of duplicated ribosomal protein genes. Nucleic Acids Res. 2016, 44, 3878–3891. [Google Scholar] [CrossRef] [Green Version]
- Fewell, S.W.; Woolford, J.L., Jr. Ribosomal protein S14 of Saccharomyces cerevisiae regulates its expression by binding to RPS14B pre-mRNA and to 18S rRNA. Mol. Cell Biol. 1999, 19, 826–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Paulovich, A.G.; Woolford, J.L., Jr. Feedback inhibition of the yeast ribosomal protein gene CRY2 is mediated by the nucleotide sequence and secondary structure of CRY2 pre-mRNA. Mol. Cell Biol. 1995, 15, 6454–6464. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Malygin, A.A.; Karpova, G.G. Human ribosomal protein S26 suppresses the splicing of its pre-mRNA. Biochim. Biophys. Acta 2005, 1727, 134–140. [Google Scholar] [CrossRef]
- Cuccurese, M.; Russo, G.; Russo, A.; Pietropaolo, C. Alternative splicing and nonsense-mediated mRNA decay regulate mammalian ribosomal gene expression. Nucleic Acids Res. 2005, 33, 5965–5977. [Google Scholar] [CrossRef] [Green Version]
- Bozzoni, I.; Fragapane, P.; Annesi, F.; Pierandrei-Amaldi, P.; Amaldi, F.; Beccari, E. Expression of two Xenopus laevis ribosomal protein genes in injected frog oocytes. A specific splicing block interfeRes. with the L1 RNA maturation. J. Mol. Biol. 1984, 180, 987–1005. [Google Scholar] [CrossRef]
- Takei, S.; Togo-Ohno, M.; Suzuki, Y.; Kuroyanagi, H. Evolutionarily conserved autoregulation of alternative pre-mRNA splicing by ribosomal protein L10a. Nucleic Acids Res. 2016, 44, 5585–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrovich, Q.M.; Anderson, P. Unproductively spliced ribosomal protein mRNAs are natural targets of mRNA surveillance in C. elegans. Genes Dev. 2000, 14, 2173–2184. [Google Scholar] [CrossRef] [Green Version]
- Gabunilas, J.; Chanfreau, G. Splicing-Mediated Autoregulation Modulates Rpl22p Expression in Saccharomyces cerevisiae. PLoS Genet. 2016, 12, e1005999. [Google Scholar] [CrossRef] [PubMed]
- Vilardell, J.; Warner, J.R. Regulation of splicing at an intermediate step in the formation of the spliceosome. Genes Dev. 1994, 8, 211–220. [Google Scholar] [CrossRef] [Green Version]
- White, S.A.; Hoeger, M.; Schweppe, J.J.; Shillingford, A.; Shipilov, V.; Zarutskie, J. Internal loop mutations in the ribosomal protein L30 binding site of the yeast L30 RNA transcript. RNA 2004, 10, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Eng, F.J.; Warner, J.R. Structural basis for the regulation of splicing of a yeast messenger RNA. Cell 1991, 65, 797–804. [Google Scholar] [CrossRef]
- Dabeva, M.D.; Warner, J.R. Ribosomal protein L32 of Saccharomyces cerevisiae regulates both splicing and translation of its own transcript. J. Biol. Chem. 1993, 268, 19669–19674. [Google Scholar] [CrossRef]
- Parenteau, J.; Durand, M.; Morin, G.; Gagnon, J.; Lucier, J.F.; Wellinger, R.J.; Chabot, B.; Elela, S.A. Introns within ribosomal protein genes regulate the production and function of yeast ribosomes. Cell 2011, 147, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenteau, J.; Maignon, L.; Berthoumieux, M.; Catala, M.; Gagnon, V.; Abou Elela, S. Introns are mediators of cell response to starvation. Nature 2019, 565, 612–617. [Google Scholar] [CrossRef]
- Russo, A.; Siciliano, G.; Catillo, M.; Giangrande, C.; Amoresano, A.; Pucci, P.; Pietropaolo, C.; Russo, G. hnRNP H1 and intronic G runs in the splicing control of the human rpL3 gene. Biochim. Biophys. Acta 2010, 1799, 419–428. [Google Scholar] [CrossRef]
- Russo, A.; Catillo, M.; Esposito, D.; Briata, P.; Pietropaolo, C.; Russo, G. Autoregulatory circuit of human rpL3 expression requiRes. hnRNP H1, NPM and KHSRP. Nucleic Acids Res. 2011, 39, 7576–7585. [Google Scholar] [CrossRef]
- Zhang, Y.; O’Leary, M.N.; Peri, S.; Wang, M.; Zha, J.; Melov, S.; Kappes, D.J.; Feng, Q.; Rhodes, J.; Amieux, P.S.; et al. Ribosomal Proteins Rpl22 and Rpl22l1 Control Morphogenesis by Regulating Pre-mRNA Splicing. Cell Rep. 2017, 18, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Abrhamova, K.; Nemcko, F.; Libus, J.; Prevorovsky, M.; Halova, M.; Puta, F.; Folk, P. Introns provide a platform for intergenic regulatory feedback of RPL22 paralogs in yeast. PLoS ONE 2018, 13, e0190685. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Crutchley, J.; Zhang, D.; Owzar, K.; Kastan, M.B. Identification of a DNA Damage-Induced Alternative Splicing Pathway That Regulates p53 and Cellular Senescence Markers. Cancer Discov. 2017, 7, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Gopanenko, A.V.; Malygin, A.A.; Tupikin, A.E.; Laktionov, P.P.; Kabilov, M.R.; Karpova, G.G. Human ribosomal protein eS1 is engaged in cellular events related to processing and functioning of U11 snRNA. Nucleic Acids Res. 2017, 45, 9121–9137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Liu, K.; Zhang, S.; Wang, X.; Rogers, K.; Ren, G.; Zhang, C.; Yu, B. STV1, a ribosomal protein, binds primary microRNA transcripts to promote their interaction with the processing complex in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Dostie, J.; Lejbkowicz, F.; Sonenberg, N. Nuclear eukaryotic initiation factor 4E (eIF4E) colocalizes with splicing factors in speckles. J. Cell Biol. 2000, 148, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Graham, P.L.; Yanowitz, J.L.; Penn, J.K.; Deshpande, G.; Schedl, P. The translation initiation factor eIF4E regulates the sex-specific expression of the master switch gene Sxl in Drosophila melanogaster. PLoS Genet. 2011, 7, e1002185. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.R.; Delaleau, M.; Borden, K.L.B. Nuclear eIF4E Stimulates 3′-End Cleavage of Target RNAs. Cell Rep. 2019, 27, 1397–1408.e4. [Google Scholar] [CrossRef] [Green Version]
- Culjkovic-Kraljacic, B.; Skrabanek, L.; Revuelta, M.V.; Gasiorek, J.; Cowling, V.H.; Cerchietti, L.; Borden, K.L.B. The eukaryotic translation initiation factor eIF4E elevates steady-state m(7)G capping of coding and noncoding transcripts. Proc. Natl. Acad. Sci. USA 2020, 117, 26773–26783. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Di Giammartino, D.C.; Taylor, D.; Sarkeshik, A.; Rice, W.J.; Yates, J.R., 3rd; Frank, J.; Manley, J.L. Molecular architecture of the human pre-mRNA 3′ processing complex. Mol. Cell 2009, 33, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Gross, S.; Moore, C. Five subunits are required for reconstitution of the cleavage and polyadenylation activities of Saccharomyces cerevisiae cleavage factor I. Proc. Natl. Acad. Sci. USA 2001, 98, 6080–6085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, M.M.; Zhao, J.; Moore, C.L. Purification of the Saccharomyces cerevisiae cleavage/polyadenylation factor I. Separation into two components that are required for both cleavage and polyadenylation of mRNA 3′ ends. J. Biol. Chem. 1996, 271, 27167–27175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.; Wittmann, J.; Jack, H.M.; Jalinot, P. Human INT6/eIF3e is required for nonsense-mediated mRNA decay. EMBO Rep. 2007, 8, 596–602. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ramirez, J.; Franco, M.; Lectez, B.; Gonzalez, M.; Barrio, R.; Mayor, U. Ube3a, the E3 ubiquitin ligase causing Angelman syndrome and linked to autism, regulates protein homeostasis through the proteasomal shuttle Rpn10. Cell Mol. Life Sci. 2014, 71, 2747–2758. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Parisky, K.; Celotto, A.M.; Reenan, R.A.; Graveley, B.R. Identification of alternative splicing regulators by RNA interference in Drosophila. Proc. Natl. Acad. Sci. USA 2004, 101, 15974–15979. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Yan, C.; Zhang, X.; Lei, J.; Shi, Y. Structure of a human catalytic step I spliceosome. Science 2018, 359, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Kanellis, D.C.; Espinoza, J.A.; Zisi, A.; Sakkas, E.; Bartkova, J.; Katsori, A.M.; Bostrom, J.; Dyrskjot, L.; Broholm, H.; Altun, M.; et al. The exon-junction complex helicase eIF4A3 controls cell fate via coordinated regulation of ribosome biogenesis and translational output. Sci. Adv. 2021, 7, eabf7561. [Google Scholar] [CrossRef]
- Koroleva, O.A.; Calder, G.; Pendle, A.F.; Kim, S.H.; Lewandowska, D.; Simpson, C.G.; Jones, I.M.; Brown, J.W.; Shaw, P.J. Dynamic behavior of Arabidopsis eIF4A-III, putative core protein of exon junction complex: Fast relocation to nucleolus and splicing speckles under hypoxia. Plant Cell 2009, 21, 1592–1606. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Chen, T.; Qin, T.; Ding, F.; Wang, Z.; Chen, H.; Xiong, L. The RNA Polymerase II C-Terminal Domain Phosphatase-Like Protein FIERY2/CPL1 Interacts with eIF4AIII and Is Essential for Nonsense-Mediated mRNA Decay in Arabidopsis. Plant Cell 2016, 28, 770–785. [Google Scholar] [CrossRef] [Green Version]
- McKendrick, L.; Thompson, E.; Ferreira, J.; Morley, S.J.; Lewis, J.D. Interaction of eukaryotic translation initiation factor 4G with the nuclear cap-binding complex provides a link between nuclear and cytoplasmic functions of the m(7) guanosine cap. Mol. Cell Biol. 2001, 21, 3632–3641. [Google Scholar] [CrossRef] [Green Version]
- Kafasla, P.; Barrass, J.D.; Thompson, E.; Fromont-Racine, M.; Jacquier, A.; Beggs, J.D.; Lewis, J. Interaction of yeast eIF4G with spliceosome components: Implications in pre-mRNA processing events. RNA Biol. 2009, 6, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Sun, F.; Handel, M.A. Nuclear localization of EIF4G3 suggests a role for the XY body in translational regulation during spermatogenesis in mice. Biol. Reprod. 2018, 98, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, M.; Lasko, P.; Richter, J.D. Cytoplasmic polyadenylation element binding proteins in development, health, and disease. Annu. Rev. Cell Dev. Biol. 2014, 30, 393–415. [Google Scholar] [CrossRef]
- Kozlov, E.; Shidlovskii, Y.V.; Gilmutdinov, R.; Schedl, P.; Zhukova, M. The role of CPEB family proteins in the nervous system function in the norm and pathology. Cell BioSci. 2021, 11, 64. [Google Scholar] [CrossRef]
- Kan, M.C.; Oruganty-Das, A.; Cooper-Morgan, A.; Jin, G.; Swanger, S.A.; Bassell, G.J.; Florman, H.; van Leyen, K.; Richter, J.D. CPEB4 is a cell survival protein retained in the nucleus upon ischemia or endoplasmic reticulum calcium depletion. Mol. Cell Biol. 2010, 30, 5658–5671. [Google Scholar] [CrossRef] [Green Version]
- Ernoult-Lange, M.; Wilczynska, A.; Harper, M.; Aigueperse, C.; Dautry, F.; Kress, M.; Weil, D. Nucleocytoplasmic traffic of CPEB1 and accumulation in Crm1 nucleolar bodies. Mol. Biol. Cell 2009, 20, 176–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Evans, V.; Shen, S.; Xing, Y.; Richter, J.D. The nuclear experience of CPEB: Implications for RNA processing and translational control. RNA 2010, 16, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bava, F.A.; Eliscovich, C.; Ferreira, P.G.; Miñana, B.; Ben-Dov, C.; Guigó, R.; Valcárcel, J.; Méndez, R. CPEB1 coordinates alternative 3′-UTR formation with translational regulation. Nature 2013, 495, 121–125. [Google Scholar] [CrossRef]
- Qu, W.; Jin, H.; Chen, B.P.; Liu, J.; Li, R.; Guo, W.; Tian, H. CPEB3 regulates neuron-specific alternative splicing and involves neurogenesis gene expression. Aging 2020, 13, 2330–2347. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.W.; Lai, Y.T.; Lu, Y.L.; Lin, C.L.; Mai, W.; Huang, Y.S. NMDAR signaling facilitates the IPO5-mediated nuclear import of CPEB3. Nucleic Acids Res. 2012, 40, 8484–8498. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.C.; Lai, Y.T.; Huang, H.Y.; Huang, H.D.; Huang, Y.S. A novel role of CPEB3 in regulating EGFR gene transcription via association with Stat5b in neurons. Nucleic Acids Res. 2010, 38, 7446–7457. [Google Scholar] [CrossRef]
- Culjkovic, B.; Topisirovic, I.; Skrabanek, L.; Ruiz-Gutierrez, M.; Borden, K.L. eIF4E promotes nuclear export of cyclin D1 mRNAs via an element in the 3′UTR. J. Cell Biol. 2005, 169, 245–256. [Google Scholar] [CrossRef]
- Culjkovic-Kraljacic, B.; Baguet, A.; Volpon, L.; Amri, A.; Borden, K.L. The oncogene eIF4E reprograms the nuclear pore complex to promote mRNA export and oncogenic transformation. Cell Rep. 2012, 2, 207–215. [Google Scholar] [CrossRef]
- Culjkovic, B.; Topisirovic, I.; Skrabanek, L.; Ruiz-Gutierrez, M.; Borden, K.L. eIF4E is a central node of an RNA regulon that governs cellular proliferation. J. Cell Biol. 2006, 175, 415–426. [Google Scholar] [CrossRef]
- Topisirovic, I.; Siddiqui, N.; Lapointe, V.L.; Trost, M.; Thibault, P.; Bangeranye, C.; Pinol-Roma, S.; Borden, K.L. Molecular dissection of the eukaryotic initiation factor 4E (eIF4E) export-competent RNP. EMBO J. 2009, 28, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Volpon, L.; Culjkovic-Kraljacic, B.; Sohn, H.S.; Blanchet-Cohen, A.; Osborne, M.J.; Borden, K.L.B. A biochemical framework for eIF4E-dependent mRNA export and nuclear recycling of the export machinery. RNA 2017, 23, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, N.; Sharma, M.; Kentsis, A.; Perez, J.M.; Strudwick, S.; Borden, K.L. PML RING suppresses oncogenic transformation by reducing the affinity of eIF4E for mRNA. EMBO J. 2001, 20, 4547–4559. [Google Scholar] [CrossRef] [Green Version]
- Topisirovic, I.; Kentsis, A.; Perez, J.M.; Guzman, M.L.; Jordan, C.T.; Borden, K.L. Eukaryotic translation initiation factor 4E activity is modulated by HOXA9 at multiple levels. Mol. Cell Biol. 2005, 25, 1100–1112. [Google Scholar] [CrossRef] [Green Version]
- Rosorius, O.; Reichart, B.; Kratzer, F.; Heger, P.; Dabauvalle, M.C.; Hauber, J. Nuclear pore localization and nucleocytoplasmic transport of eIF-5A: Evidence for direct interaction with the export receptor CRM1. J. Cell Sci. 1999, 112 Pt 14, 2369–2380. [Google Scholar] [CrossRef]
- Elfgang, C.; Rosorius, O.; Hofer, L.; Jaksche, H.; Hauber, J.; Bevec, D. Evidence for specific nucleocytoplasmic transport pathways used by leucine-rich nuclear export signals. Proc. Natl. Acad. Sci. USA 1999, 96, 6229–6234. [Google Scholar] [CrossRef] [Green Version]
- Ruhl, M.; Himmelspach, M.; Bahr, G.M.; Hammerschmid, F.; Jaksche, H.; Wolff, B.; Aschauer, H.; Farrington, G.K.; Probst, H.; Bevec, D.; et al. Eukaryotic initiation factor 5A is a cellular target of the human immunodeficiency virus type 1 Rev. activation domain mediating trans-activation. J. Cell Biol. 1993, 123, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Bevec, D.; Jaksche, H.; Oft, M.; Wohl, T.; Himmelspach, M.; Pacher, A.; Schebesta, M.; Koettnitz, K.; Dobrovnik, M.; Csonga, R.; et al. Inhibition of HIV-1 replication in lymphocytes by mutants of the Rev. cofactor eIF-5A. Science 1996, 271, 1858–1860. [Google Scholar] [CrossRef]
- Katahira, J.; Ishizaki, T.; Sakai, H.; Adachi, A.; Yamamoto, K.; Shida, H. Effects of translation initiation factor eIF-5A on the functioning of human T-cell leukemia virus type I Rex and human immunodeficiency virus Rev. inhibited trans dominantly by a Rex mutant deficient in RNA binding. J. Virol. 1995, 69, 3125–3133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balukoff, N.C.; Ho, J.J.D.; Theodoridis, P.R.; Wang, M.; Bokros, M.; Llanio, L.M.; Krieger, J.R.; Schatz, J.H.; Lee, S. A translational program that suppresses metabolism to shield the genome. Nat. Commun. 2020, 11, 5755. [Google Scholar] [CrossRef]
- Gobert, A.P.; Finley, J.L.; Latour, Y.L.; Asim, M.; Smith, T.M.; Verriere, T.G.; Barry, D.P.; Allaman, M.M.; Delagado, A.G.; Rose, K.L.; et al. Hypusination Orchestrates the Antimicrobial Response of Macrophages. Cell Rep. 2020, 33, 108510. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Ogihara, T.; Trace, A.P.; Tersey, S.A.; Robbins, R.D.; Chakrabarti, S.K.; Nunemaker, C.S.; Stull, N.D.; Taylor, C.A.; Thompson, J.E.; et al. The unique hypusine modification of eIF5A promotes islet beta cell inflammation and dysfunction in mice. J. Clin. Investig. 2010, 120, 2156–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, M.; Rosorius, O.; Kratzer, F.; Bevec, D.; Kuhnt, C.; Steinkasserer, A.; Schuler, G.; Hauber, J. Inhibition of CD83 cell surface expression during dendritic cell maturation by interference with nuclear export of CD83 mRNA. J. Exp. Med. 2000, 191, 1581–1590. [Google Scholar] [CrossRef] [Green Version]
- Bohm, M.; Sawicka, K.; Siebrasse, J.P.; Brehmer-Fastnacht, A.; Peters, R.; Klempnauer, K.H. The transformation suppressor protein Pdcd4 shuttles between nucleus and cytoplasm and binds RNA. Oncogene 2003, 22, 4905–4910. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.T.; Rane, C.K.; Zito, C.R.; Weiss, S.A.; Jessel, S.; Lucca, L.; Lu, B.Y.; Oria, V.O.; Adeniran, A.; Chiang, V.L.; et al. Clinical Significance of PDCD4 in Melanoma by Subcellular Expression and in Tumor-Associated Immune Cells. Cancers 2021, 13, 1049. [Google Scholar] [CrossRef]
- Sasaki, M.; Kawahara, K.; Nishio, M.; Mimori, K.; Kogo, R.; Hamada, K.; Itoh, B.; Wang, J.; Komatsu, Y.; Yang, Y.R.; et al. Regulation of the MDM2-P53 pathway and tumor growth by PICT1 via nucleolar RPL11. Nat. Med. 2011, 17, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Han, L.; Tsai, H.; Wang, Z.; Wu, Y.; Duo, Y.; Cao, W.; Chen, L.; Tan, Z.; Xu, N.; et al. PICT-1 is a key nucleolar sensor in DNA damage response signaling that regulates apoptosis through the RPL11-MDM2-p53 pathway. Oncotarget 2016, 7, 83241–83257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, K.; Kumazawa, T.; Kuroda, T.; Katagiri, N.; Tsuchiya, M.; Goto, N.; Furumai, R.; Murayama, A.; Yanagisawa, J.; Kimura, K. Perturbation of ribosome biogenesis drives cells into senescence through 5S RNP-mediated p53 activation. Cell Rep. 2015, 10, 1310–1323. [Google Scholar] [CrossRef] [Green Version]
- Fregoso, O.I.; Das, S.; Akerman, M.; Krainer, A.R. Splicing-factor oncoprotein SRSF1 stabilizes p53 via RPL5 and induces cellular senescence. Mol. Cell 2013, 50, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zhang, J.; Gu, Y.; Gan, X.; Gan, Y.; Zheng, W.; Kim, B.W.; Xu, X.; Lu, X.; Dong, Q.; et al. Identification of a novel RNA giant nuclear body in cancer cells. Oncotarget 2016, 7, 4724–4734. [Google Scholar] [CrossRef]
- Lobo, M.V.; Martin, M.E.; Perez, M.I.; Alonso, F.J.; Redondo, C.; Alvarez, M.I.; Salinas, M. Levels, phosphorylation status and cellular localization of translational factor eIF2 in gastrointestinal carcinomas. HistoChem. J. 2000, 32, 139–150. [Google Scholar] [CrossRef]
- Tejada, S.; Lobo, M.V.; Garcia-Villanueva, M.; Sacristan, S.; Perez-Morgado, M.I.; Salinas, M.; Martin, M.E. Eukaryotic initiation factors (eIF) 2alpha and 4E expression, localization, and phosphorylation in brain tumors. J. HistoChem. CytoChem. 2009, 57, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, S.J.; Norbury, C.J. Cell cycle-related variation in subcellular localization of eIF3e/INT6 in human fibroblasts. Cell Prolif. 2004, 37, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Pisera, A.; Campo, A.; Campo, S. Structure and functions of the translation initiation factor eIF4E and its role in cancer development and treatment. J. Genet. Genom. 2018, 45, 13–24. [Google Scholar] [CrossRef]
- ZHOU, H.; JIA, X.; YANG, F. Elevated nuclear phospho-eIF4E body levels are associated with tumor progression and poor prognosis for acute myeloid leukemia. Biocell 2021, 45, 711–722. [Google Scholar] [CrossRef]
- Volpon, L.; Culjkovic-Kraljacic, B.; Osborne, M.J.; Ramteke, A.; Sun, Q.; Niesman, A.; Chook, Y.M.; Borden, K.L. Importin 8 mediates m7G cap-sensitive nuclear import of the eukaryotic translation initiation factor eIF4E. Proc. Natl. Acad. Sci. USA 2016, 113, 5263–5268. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.F.; Zhou, H. Phospho-eIF4E: A new target for acute myeloid leukemia. Curr. Protein Pept. Sci. 2021, 22, 328–335. [Google Scholar] [CrossRef]
- Chen, G.; Gharib, T.G.; Thomas, D.G.; Huang, C.C.; Misek, D.E.; Kuick, R.D.; Giordano, T.J.; Iannettoni, M.D.; Orringer, M.B.; Hanash, S.M.; et al. Proteomic analysis of eIF-5A in lung adenocarcinomas. Proteomics 2003, 3, 496–504. [Google Scholar] [CrossRef]
- Ning, L.; Wang, L.; Zhang, H.; Jiao, X.; Chen, D. Eukaryotic translation initiation factor 5A in the pathogenesis of cancers. Oncol. Lett. 2020, 20, 81. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.A.; Sun, Z.; Cliche, D.O.; Ming, H.; Eshaque, B.; Jin, S.; Hopkins, M.T.; Thai, B.; Thompson, J.E. Eukaryotic translation initiation factor 5A induces apoptosis in colon cancer cells and associates with the nucleus in response to tumour necrosis factor alpha signalling. Exp. Cell Res. 2007, 313, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Moghanibashi, M.; Rastgar Jazii, F.; Soheili, Z.S.; Zare, M.; Karkhane, A.; Parivar, K.; Mohamadynejad, P. Esophageal cancer alters the expression of nuclear pore complex binding protein Hsc70 and eIF5A-1. Funct. Integr. Genomics 2013, 13, 253–260. [Google Scholar] [CrossRef]
- Khosravi, S.; Martinka, M.; Zhou, Y.; Ong, C.J. Prognostic significance of the expression of nuclear eukaryotic translation initiation factor 5A2 in human melanoma. Oncol Lett. 2016, 12, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, S.; Wong, R.P.; Ardekani, G.S.; Zhang, G.; Martinka, M.; Ong, C.J.; Li, G. Role of EIF5A2, a downstream target of Akt, in promoting melanoma cell invasion. Br. J. Cancer 2014, 110, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Dapas, B.; Tell, G.; Scaloni, A.; Pines, A.; Ferrara, L.; Quadrifoglio, F.; Scaggiante, B. Identification of different isoforms of eEF1A in the nuclear fraction of human T-lymphoblastic cancer cell line specifically binding to aptameric cytotoxic GT oligomers. Eur. J. Biochem. 2003, 270, 3251–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, L.D.; Corydon, T.J.; Rankel, L.; Nielsen, K.M.; Fuchtbauer, E.M.; Knudsen, C.R. An eEF1A1 truncation encoded by PTI-1 exerts its oncogenic effect inside the nucleus. Cancer Cell Int. 2014, 14, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veremieva, M.; Kapustian, L.; Khoruzhenko, A.; Zakharychev, V.; Negrutskii, B.; El’skaya, A. Independent overexpression of the subunits of translation elongation factor complex eEF1H in human lung cancer. BMC Cancer 2014, 14, 913. [Google Scholar] [CrossRef] [Green Version]
- Allfrey, V.G. Amino acid incorporation by isolated thymus nuclei. I. the role of desoxyribonucleic acid in protein synthesis. Proc. Natl. Acad. Sci. USA 1954, 40, 881–885. [Google Scholar] [CrossRef] [Green Version]
- Allfrey, V.G.; Mirsky, A.E. Protein synthesis in isolated cell nuclei. Nature 1955, 176, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Iborra, F.J.; Jackson, D.A.; Cook, P.R. Coupled transcription and translation within nuclei of mammalian cells. Science 2001, 293, 1139–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlberg, J.E.; Lund, E.; Goodwin, E.B. Nuclear translation: What is the evidence? RNA 2003, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hentze, M.W. Protein synthesis. Believe it or not-translation in the nucleus. Science 2001, 293, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Nickless, A.; Bailis, J.M.; You, Z. Control of gene expression through the nonsense-mediated RNA decay pathway. Cell BioSci. 2017, 7, 26. [Google Scholar] [CrossRef]
- Al-Jubran, K.; Wen, J.; Abdullahi, A.; Roy Chaudhury, S.; Li, M.; Ramanathan, P.; Matina, A.; De, S.; Piechocki, K.; Rugjee, K.N.; et al. Visualization of the joining of ribosomal subunits reveals the presence of 80S ribosomes in the nucleus. RNA 2013, 19, 1669–1683. [Google Scholar] [CrossRef] [Green Version]
- David, A.; Dolan, B.P.; Hickman, H.D.; Knowlton, J.J.; Clavarino, G.; Pierre, P.; Bennink, J.R.; Yewdell, J.W. Nuclear translation visualized by ribosome-bound nascent chain puromycylation. J. Cell Biol. 2012, 197, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Apcher, S.; Millot, G.; Daskalogianni, C.; Scherl, A.; Manoury, B.; Fahraeus, R. Translation of pre-spliced RNAs in the nuclear compartment generates peptides for the MHC class I pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 17951–17956. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.P.; Malbert-Colas, L.; Lista, M.J.; Daskalogianni, C.; Apcher, S.; Pla, M.; Findakly, S.; Blondel, M.; Fahraeus, R. Nuclear processing of nascent transcripts determines synthesis of full-length proteins and antigenic peptides. Nucleic Acids Res. 2019, 47, 3086–3100. [Google Scholar] [CrossRef] [PubMed]
- Duvallet, E.; Boulpicante, M.; Yamazaki, T.; Daskalogianni, C.; Prado Martins, R.; Baconnais, S.; Manoury, B.; Fahraeus, R.; Apcher, S. Exosome-driven transfer of tumor-associated Pioneer Translation Products (TA-PTPs) for the MHC class I cross-presentation pathway. Oncoimmunology 2016, 5, e1198865. [Google Scholar] [CrossRef] [Green Version]
- Baboo, S.; Bhushan, B.; Jiang, H.; Grovenor, C.R.; Pierre, P.; Davis, B.G.; Cook, P.R. Most human proteins made in both nucleus and cytoplasm turn over within minutes. PLoS ONE 2014, 9, e99346. [Google Scholar] [CrossRef] [Green Version]
- Boulpicante, M.; Darrigrand, R.; Pierson, A.; Salgues, V.; Rouillon, M.; Gaudineau, B.; Khaled, M.; Cattaneo, A.; Bachi, A.; Cascio, P.; et al. Tumors escape immunosurveillance by overexpressing the proteasome activator PSME3. Oncoimmunology 2020, 9, 1761205. [Google Scholar] [CrossRef] [PubMed]
- Pierson, A.; Darrigrand, R.; Rouillon, M.; Boulpicante, M.; Renko, Z.D.; Garcia, C.; Ghosh, M.; Laiguillon, M.-C.; Lobry, C.; Alami, M.; et al. Splicing inhibition enhances the antitumor immune response through increased tumor antigen presentation and altered MHC-I immunopeptidome. bioRxiv 2019, 512681. [Google Scholar] [CrossRef]
- Theodoridis, P.R.; Bokros, M.; Marijan, D.; Balukoff, N.C.; Wang, D.; Kirk, C.C.; Budine, T.D.; Goldsmith, H.D.; Wang, M.; Audas, T.E.; et al. Local translation in nuclear condensate amyloid bodies. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Lashkevich, K.A.; Dmitriev, S.E. mRNA Targeting, Transport and Local Translation in Eukaryotic Cells: From the Classical View to a Diversity of New Concepts. Mol. Biol. 2021, 55, 507–537. [Google Scholar] [CrossRef]
- Bernhofer, M.; Goldberg, T.; Wolf, S.; Ahmed, M.; Zaugg, J.; Boden, M.; Rost, B. NLSdb-major update for database of nuclear localization signals and nuclear export signals. Nucleic Acids Res. 2018, 46, D503–D508. [Google Scholar] [CrossRef] [PubMed]
- Nalabothula, N.; Indig, F.E.; Carrier, F. The Nucleolus Takes Control of Protein Trafficking Under Cellular Stress. Mol. Cell Pharmacol 2010, 2, 203–212. [Google Scholar]
- Li, S. Regulation of Ribosomal Proteins on Viral Infection. Cells 2019, 8, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, P.P.; Topisirovic, I. Signaling Pathways Involved in the Regulation of mRNA Translation. Mol. Cell Biol. 2018, 38, e00070-18. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.; Lan, T.; Mo, B. Extraribosomal Functions of Cytosolic Ribosomal Proteins in Plants. Front. Plant. Sci. 2021, 12, 607157. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Tang, N.; Yang, J.; Chen, C.; Liu, S.; Cheng, C.; Wang, Y.; Guo, Y.-n.; Wang, H.; Xiang, M.; et al. Decoding regulatory specificity of human ribosomal proteins. bioRxiv 2021, 437302. [Google Scholar] [CrossRef]
- Du, X.; Xiao, R. An emerging role of chromatin-interacting RNA-binding proteins in transcription regulation. Essays Biochem. 2020, 64, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Razin, S.V.; Gavrilov, A.A. Non-coding RNAs in chromatin folding and nuclear organization. Cell Mol. Life Sci. 2021, 78, 5489–5504. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Lim, D.H.; Fu, X.D. Mechanistic Dissection of RNA-Binding Proteins in Regulated Gene Expression at Chromatin Levels. Cold Spring Harb. Symp. Quant. Biol. 2019, 84, 55–66. [Google Scholar] [CrossRef]
- Balcerak, A.; Trebinska-Stryjewska, A.; Konopinski, R.; Wakula, M.; Grzybowska, E.A. RNA-protein interactions: Disorder, moonlighting and junk contribute to eukaryotic complexity. Open Biol. 2019, 9, 190096. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, X.; Cheng, Z.; Zhu, Q. AGO2 and its partners: A silencing complex, a chromatin modulator, and new features. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 33–53. [Google Scholar] [CrossRef]
- Sangermano, F.; Delicato, A.; Calabro, V. Y box binding protein 1 (YB-1) oncoprotein at the hub of DNA proliferation, damage and cancer progression. Biochimie 2020, 179, 205–216. [Google Scholar] [CrossRef]
- Kretov, D.A.; Mordovkina, D.A.; Eliseeva, I.A.; Lyabin, D.N.; Polyakov, D.N.; Joshi, V.; Desforges, B.; Hamon, L.; Lavrik, O.I.; Pastre, D.; et al. Inhibition of Transcription Induces Phosphorylation of YB-1 at Ser102 and Its Accumulation in the Nucleus. Cells 2019, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Mordovkina, D.A.; Kim, E.R.; Buldakov, I.A.; Sorokin, A.V.; Eliseeva, I.A.; Lyabin, D.N.; Ovchinnikov, L.P. Transportin-1-dependent YB-1 nuclear import. Biochem. Biophys. Res. Commun. 2016, 480, 629–634. [Google Scholar] [CrossRef]
- Zanzoni, A.; Ribeiro, D.M.; Brun, C. Understanding protein multifunctionality: From short linear motifs to cellular functions. Cell Mol. Life Sci. 2019, 76, 4407–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Jeffery, C.J. Moonlighting Proteins in the Fuzzy Logic of Cellular Metabolism. Molecules 2020, 25, 3440. [Google Scholar] [CrossRef]
- Espinosa-Cantu, A.; Cruz-Bonilla, E.; Noda-Garcia, L.; DeLuna, A. Multiple Forms of Multifunctional Proteins in Health and Disease. Front. Cell Dev. Biol. 2020, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Franco-Serrano, L.; Huerta, M.; Hernandez, S.; Cedano, J.; Perez-Pons, J.; Pinol, J.; Mozo-Villarias, A.; Amela, I.; Querol, E. Multifunctional Proteins: Involvement in Human Diseases and Targets of Current Drugs. Protein J. 2018, 37, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, V.V. Protein multi-functionality: Introduction. Cell Mol. Life Sci. 2019, 76, 4405–4406. [Google Scholar] [CrossRef] [PubMed]
- Slobodin, B.; Dikstein, R. So close, no matter how far: Multiple paths connecting transcription to mRNA translation in eukaryotes. EMBO Rep. 2020, 21, e50799. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.A.; Bailey-Serres, J. Integrative analysis from the epigenome through translation exposes patterns of dominant nuclear regulation during transient stress. Plant Cell 2019, 31, 2573–2595. [Google Scholar] [CrossRef] [Green Version]
- Maciag, K.; Altschuler, S.J.; Slack, M.D.; Krogan, N.J.; Emili, A.; Greenblatt, J.F.; Maniatis, T.; Wu, L.F. Systems-level analyses identify extensive coupling among gene expression machines. Mol. Syst. Biol. 2006, 2, 2006.0003. [Google Scholar] [CrossRef] [Green Version]


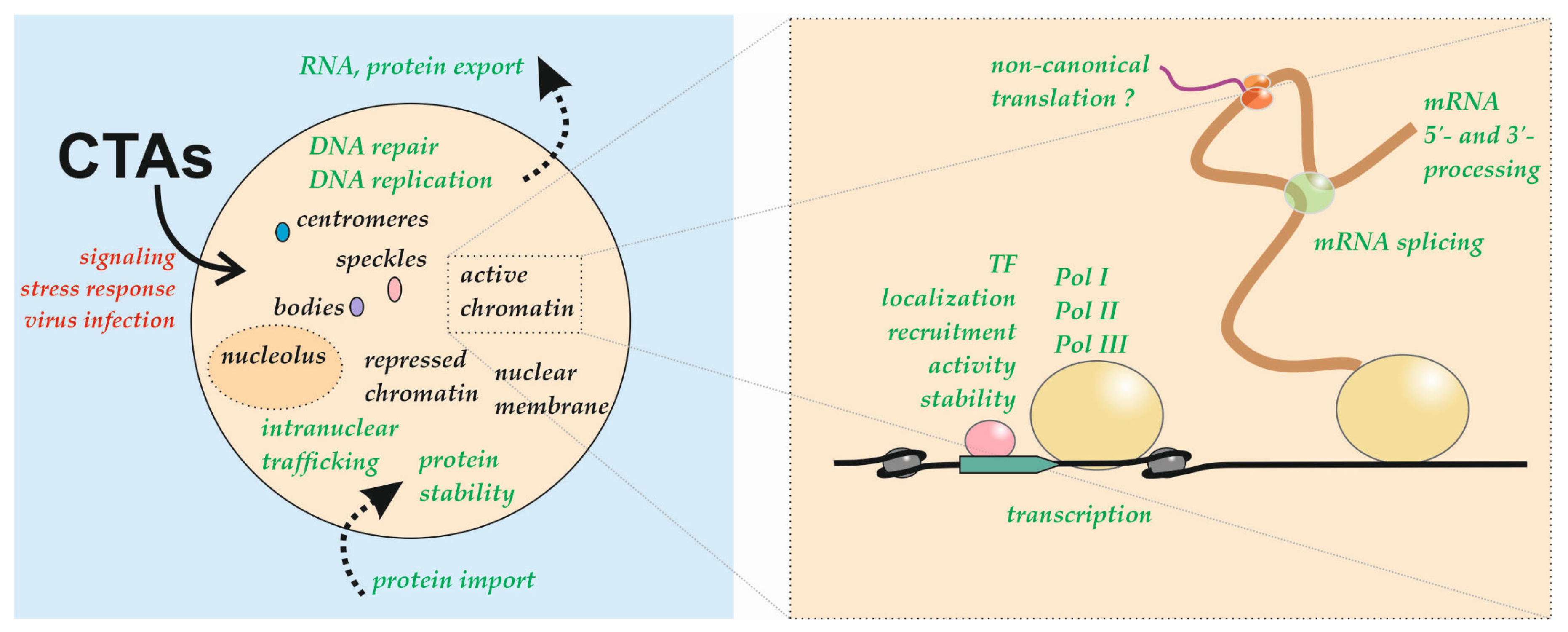{kind=link}
| Protein (RP Names are Indicated According to [3]) | Nuclear Functions (Pooled Data for All Species) | Species in which Nuclear Localization Was Detected | References |
|---|---|---|---|
| RPSA | Controls RNF8 and BRCA1 trafficking Localized at active loci on chromosomes Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,129,149,150] |
| RPS2 | Binds Mdm2 and ZNF277 Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals and flies | [33,40,42,149,150,157,158,160,170,188,189,216] |
| RPS3 | Component of DNA damage response pathway Endonuclease activity Interacts with OGG1, APE/Ref-1, UNG, TFIIH, and RECQL4 repair proteins Induction of proapoptotic genes Essential subunit of NF-κB Binds Mdm2, p53, E2F1, and ERH Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,54,96,108,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,134,158,179,181,182,183,184,185,186,216,294] |
| RPS3A | Binds CHOP, EBNA5, and PARP Interacts with the U5 and U11 snRNAs Binds phosphatidylinositol trisphosphate Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, flies, and plants | [25,26,33,40,41,42,107,157,158,160,167,168,169,216,326] |
| RPS4 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,40,41,42,149,150,158,161,216] |
| RPS5 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,40,42,149,150,158,161,216] |
| RPS6 | Binds and downregulates promoters of rRNA genes Interacts with NAP1 Interacts with LANA Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals and plants | [25,26,33,40,41,42,111,158,160,212,213,216] |
| RPS7 | Binds Mdm2, GADD45α, and BCCIPβ Associated with nascent mRNA Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, yeasts, and plants | [25,26,33,40,42,105,152,160,186,191,208] |
| RPS7A | Unknown | Plants | [26] |
| RPS8 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction Found in the chromosome scaffold fraction | Mammals, flies, yeasts, and plants | [25,26,33,40,41,42,149,150,157,158,161,216] |
| RPS9 | Localized at active loci on chromosomes Suppression of splicing of its own pre-mRNA Interacts with nucleophosmin Associated with TFIIIE Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, flies, and yeasts | [33,40,42,103,104,149,150,157,158,160,161,204,216,306] |
| RPS10 | Associated with HP1a Chromatin-associated Component of interchromatin granule clusters Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,40,41,42,95,158] |
| RPS11 | Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [25,26,33,149,150,158,216] |
| RPS12 | Localized at active loci on chromosomes Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [25,26,33,149,150,216] |
| RPS13 | Suppression of splicing of its own pre-mRNA Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,149,150,157,158,216,305] |
| RPS14 | Recruited to DNA damage sites Binds Mdm2 and c-myc Localized at active loci on chromosomes Inhibits transcription of its own gene Suppression of splicing of its own pre-mRNA Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and yeasts | [33,40,42,130,149,150,153,158,165,186,216,307,308] |
| RPS15 | Binds Mdm2 Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,149,150,157,158,186,216] |
| RPS15A | Associated with RNAP II | Humans and plants | [25,26,216] |
| RPS16 | Associated with HP1a Associated with RNAP II Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,40,42,158,216] |
| RPS17 | Associated with HP1a Found in the insoluble nuclear fraction | Humans, flies, and plants | [25,26,42,158] |
| RPS18 | Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,42,149,150,158,216] |
| RPS19 | Associated with HP1a Found in the insoluble nuclear fraction | Humans, flies, and plants | [25,26,42,158] |
| RPS20 | Binds Mdm2 Involved in RNAP III transcription Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and yeasts | [33,42,158,163,186,216] |
| RPS21 | Chromatin-associated Found in the insoluble nuclear fraction | Mammals and plants | [26,33,42] |
| RPS23 | Associated with HP1a Associated with histone H1 Chromatin-associated | Mammals, flies, and plants | [25,33,157,158] |
| RPS24 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,149,150,157,158,216] |
| RPS25 | Binds Mdm2 Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, flies, yeasts, and plants | [25,26,33,42,158,160,161,186,216] |
| RPS26 | Interacts with Mdm2, p53, and p300 Suppression of splicing of its own pre-mRNA Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [25,26,33,158,186,202,216,309] |
| RPS27 | Binds FANCD2 and FANCI Binds Mdm2 Associated with RNAP II Chromatin-associated | Mammals and plants | [26,33,131,186,216] |
| RPS27A | Binds Mdm2 Chromatin-associated | Mammals and plants | [25,33,192,205] |
| RPS28 | Chromatin-associated Associated with RNAP II Component of interchromatin granule clusters | Mammals and plants | [26,33,40,216] |
| RPS29 | Associated with HP1a | Flies | [158] |
| RPS30 | Localized at active loci on chromosomes | Flies and plants | [26,149,150] |
| RPLP0 | Interacts with APE1/Ref-1 Endonuclease activity Modifies position effect variegation Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals and flies | [33,40,42,54,132,133,157,158,211,216] |
| RPLP1 | Intrinsic transactivation potential Associated with HP1a Chromatin-associated Component of interchromatin granule clusters | Mammals, flies, yeasts, and plants | [25,26,33,40,158,171] |
| RPLP2 | Intrinsic transactivation potential Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,42,158,171] |
| RPL3 | Control of DNA repair Binds Sp1 Downregulates E2F1-dependent promoters by sequestering PARP1 Controls alternative splicing of its own pre-mRNA Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, flies, and plants | [26,33,41,42,97,135,157,158,160,166,206,216,310,321,322] |
| RPL4 | Suppression of splicing of its own pre-mRNA Binds Mdm2, EBNA1 Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the chromosome scaffold fraction Found in the insoluble nuclear fraction | Mammals, frogs, flies, and plants | [26,33,40,41,42,142,157,158,187,216,311] |
| RPL5 | Determines nucleolar localization of the NVL2 Interacts with splicing factor SRSF1 Binds Mdm2 and SPIN1 Induces transcriptional activity of TAp73 Participates in telomere length set point Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction Localized at active loci on chromosomes | Mammals, flies, and plants | [25,26,33,42,87,88,140,149,150,157,158,166,186,195,201,207,216,375] |
| RPL6 | Interacts with histone H2A/H2AX Recruited to DNA damage sites, mediates recruitment of repair proteins Binds Mdm2 Mediates DNA binding of Tax Associated with TFIIIE Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,42,130,157,158,161,175,190,216] |
| RPL7 | Associated with centromeres Localized at active loci on chromosomes Counteracts the binding of VDR-RXR to DNA Associated with nascent mRNA Interacts with histone H1 Associated with TFIIIE Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [26,33,42,149,150,151,152,155,157,158,161,176,216] |
| RPL7A | Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,40,42,149,150,157,158,216] |
| RPL8 | Recruited to DNA damage sites Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters | Mammals and flies | [33,40,130,149,150,157,158,216] |
| RPL9 | Participates in intranuclear traffic of Gag protein Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction Found in nuclei of sperm | Mammals, flies, and plants | [26,33,42,102,109,158,160,216] |
| RPL10 | Suppression of the binding of c-Jun homodimer to DNA Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals and plants | [25,26,33,42,94,95,157,178,216] |
| RPL10A | Interacts with LIMYB and downregulates expression of RP genes Suppression of splicing of its own pre-mRNA Associated with HP1a Associated with RNAP II Found in the insoluble nuclear fraction | Humans, flies, and plants | [42,158,173,174,216,312] |
| RPL11 | Binds c-myc, Mdm2, ARF, GRWD1, PML, and PICT1 Induces transcriptional activity of TAp73 Counteracts binding of PPARα DNA Stimulates p53-mediated transcription Induces apoptosis Nucleolar localization promotes tumor progression Association with centromeres Localized at active loci on chromosomes Required for integrity of the nucleolar structure Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,42,84,110,149,150,151,158,162,164,166,177,186,195,196,199,200,207,216,372,373,374] |
| RPL12 | Localized at active loci on chromosomes Interacts with Corto Required for transcription of the PHO pathway genes Suppression of splicing of its own pre-mRNA Associated with HP1a Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, worms, yeasts, and plants | [26,33,40,42,149,150,154,157,158,209,210,313] |
| RPL13 | Promotes activity of NF-κB and IFN-β promoters Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,40,42,158,161,172,216] |
| RPL13A | Associated with HP1a Associated with RNAP II Found in the insoluble nuclear fraction | Humans and flies | [42,158,216] |
| RPL14 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [25,26,33,40,42,149,150,158,161,216] |
| RPL15 | Required for integrity of the nucleolar structure Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,40,42,84,149,150,158,216] |
| RPL17 | Localized at active loci on chromosomes Associated with HP1a Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,42,149,150,157,158] |
| RPL18 | Localized at active loci on chromosomes Associated with HP1a Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,149,150,157,158] |
| RPL18A | Associated with HP1a Associated with RNAP II Found in the insoluble nuclear fraction | Humans, flies, and plants | [25,26,42,158,216] |
| RPL19 | Interacts with ERH Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,40,42,108,158,216] |
| RPL21 | Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,157,158,216] |
| RPL22 | Binds Mdm2 Participates in alternative splicing of smad2 pre-mRNA Suppression of splicing of paralog pre-mRNA Interacts with histone H1 Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated | Mammals, fish, flies, and plants | [25,26,33,98,99,100,155,157,158,193,216,314,323,324] |
| RPL23 | Binds Mdm2 and nucleophosmin Associated with HP1a Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Humans, flies, and plants | [25,26,33,40,42,106,158,186,242] |
| RPL23A | Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,33,40,42,151,158,216] |
| RPL24 | Binds pre-miRNA and facilitates its processing Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,158,216,327] |
| RPL26 | Participates in alternative splicing of p53 pre-mRNA Binds Mdm2 Associated with nascent mRNA Associated with TFIIIE Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and yeasts | [33,42,152,158,161,186,325] |
| RPL27 | Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,158] |
| RPL27A | Associated with HP1a Associated with histone H1 Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals and flies | [33,40,42,157,158] |
| RPL28 | Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [25,26,33,88,149,150,158,216] |
| RPL29 | Associated with HP1a Associated with RNAP II | Humans, flies, and plants | [25,26,158,216] |
| RPL30 | Suppression of splicing of its own pre-mRNA Associated with HP1a Associated with RNAP II Associated with histone H1 Chromatin-associated | Mammals, flies, yeasts, and plants | [25,26,33,157,158,216,315,316] |
| RPL31 | Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,40,42,158,216] |
| RPL32 | Suppression of splicing of its own pre-mRNA Localized at active loci on chromosomes Associated with HP1a Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,149,150,158,216,317,318] |
| RPL34 | Localized at active loci on chromosomes Associated with nascent mRNA Associated with HP1a Associated with RNAP II | Humans, flies, yeasts, and plants | [25,26,149,150,152,158,216] |
| RPL35 | Associated with HP1a Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [26,33,40,42,158,216] |
| RPL35A | Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [26,33,158,216] |
| RPL36 | Localized at active loci on chromosomes Associated with TFIIIE Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, yeasts, and plants | [25,26,33,149,150,158,161,216] |
| RPL36A | Chromatin-associated | Mammals | [33] |
| RPL37 | Binds Mdm2 | Humans | [186,203] |
| RPL37A | Associated with HP1a Chromatin-associated | Mammals and flies | [33,158] |
| RPL38 | Chromatin-associated | Mammals and plants | [26,33] |
| RPL39 | Localized at active loci on chromosomes | Flies and plants | [26,149,150] |
| RPL41 | Induces export of ATF4 from nuclei | Humans | [246] |
| eIF1 | Chromatin-associated | Mammals | [27,33] |
| eIF1A | Chromatin-associated | Mammals, flies, and yeasts | [20,22,27,28,33] |
| eIF2 (S1–S3 subunits) | Stabilization of DNA-PKcs-Ku complex Controls RRN3/TIF-IA and RNAP I activity Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, yeasts, and plants | [20,23,25,26,33,50,51,52,53,136,158,216,220] |
| eIF2A | Associated with mRNA 3′-processing complex Chromatin-associated | Mammals | [33,332] |
| eIF2B (1–5 subunits) | Chromatin-associated | Mammals and yeasts | [20,33,54] |
| eIF3a | Interacts with RNAP II Associated with mRNA 3′-processing complex Associated with histone H1 Chromatin-associated | Mammals and plants | [26,33,157,214,332] |
| eIF3b | Associated with mRNA 3′-processing complex Associated with histone H1 Chromatin-associated | Mammals and plants | [26,33,54,157,332] |
| eIF3c | Interacts with RNAP II Interacts with the nuclear COP9 signalosome Chromatin-associated | Mammals, yeasts, and plants | [26,33,49,54,215] |
| eIF3d | Associated with HP1a Chromatin-associated | Mammals, flies, yeasts, and plants | [20,26,33,158] |
| eIF3e | Participates in DNA repair Interacts with ATM Promotes loading of the RAD51 Interacts with Ub-MCM7 Interacts with the nuclear COP9 signalosomeProper accumulation of proteasome in the nucleus Interacts with Rfp, HIF-2α Increases the transcriptional activity of Pap1 Colocalized in certain PML nuclear bodies Copurifies with CBP80 Associated with mRNA 3′-processing complex Chromatin-associated | Mammals and plants | [26,33,38,45,46,47,48,70,137,138,141,221,249,332,335,379] |
| eIF3f | Cooperates with STAT3 and other TFs Associated with mRNA 3′-processing complex Colocalized in nucleus with CDK11 Chromatin-associated | Mammals and plants | [24,25,26,33,157,222,223,224,332] |
| eIF3g | Chromatin-associated | Mammals, yeasts, and plants | [20,25,26,33,54] |
| eIF3h | Enhancer of variegation function Associated with mRNA 3′-processing complex Chromatin-associated | Mammals and plants | [26,33,225,332] |
| eIF3i | Interacts with RNAP II Associated with mRNA 3′-processing complex Associated with histone H1 Chromatin-associated | Mammals and yeasts | [20,33,157,214,332] |
| eIF3j | Interacts with RNAP II | Yeasts | [215] |
| eIF3k | Colocalized with PML bodies Interacts with cyclin D3 | Humans and plants | [26,36,37] |
| eIF3l | Cofactor of RNAP I Associated with histone H1 Chromatin-associated | Mammals | [33,157,218,219] |
| eIF3m | Interacts with RNAP II Chromatin-associated | Mammals | [33,214] |
| eIF4E | Defines specific pathway of mRNA export Binds LRPPRC Interacts with PML Interacts with factors of mRNA 3′-processing Participates in mRNA biogenesis Stimulates mRNA capping and alternative splicing Associated with splicing factors Participates in cancer cell transformation | Humans, flies, frogs, yeasts, and plants | [17,18,19,24,25,26,32,39,65,66,329,330,331,354,355,356,357,358,359,380,381,382,383] |
| eIF4A | Associated with mRNA 3′-processing complex Interacts with CBP80 Presumable role in splicing Associated with HP1a and HP1c Associated with RNAP II Chromatin-associated Component of interchromatin granule clusters Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,34,40,42,71,158,159,216,332,336,337] |
| eIF4G | Associated with mRNA 3′-processing complex Interacts with the nuclear CBC and splicing factors Localized in XY body of spermatocytes Associated with HP1a Associated with RNAP II Chromatin-associated | Mammals, flies, and plants | [25,26,33,54,72,158,216,332,342,343,344] |
| eIF4B | Associated with HP1a Chromatin-associated | Mammals, flies, and plants | [26,33,54,158] |
| eIF4H | Chromatin-associated | Mammals | [33] |
| eIF5 | Chromatin-associated | Mammals and plants | [24,33] |
| eIF5B | Active transcription sites Associated with RNAP II Associated with histone H1 | Humans, flies, and plants | [26,54,150,157,216] |
| eIF6 | Participates in ribosome biogenesis Associated with RNAP II Chromatin-associated Found in the insoluble nuclear fraction | Mammals, frogs, yeasts, and plants | [20,21,25,26,27,30,32,33,34,42,106,216] |
| eEF1A | Stimulates the recruitment of HSF1 to DNA and activity of HSP70 Recruited to IFN-γ promoter and stimulates transcription Interacts with ZPR1, RNAP II, Zw5, ZIPIC, and Grau Promotes nuclear export of Sox10 and Snail Stabilizes RNAP II-TAR RNA interaction Associated with mRNA 3′-processing complex Defines a specific protein export pathway mRNA export, tRNA re-export Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Humans, flies, yeasts, and plants | [20,24,27,33,34,42,158,216,227,228,229,230,231,232,233,234,235,253,254,255,256,257,258,332,333,334,390,391,392] |
| eEF1Bβ | Chromatin-associated Putative role in transcription, splicing, and DNA damage response | Mammals and plants | [25,26,33,236,392] |
| eEF1Bγ | Putative role in splicing and control of mRNA stability Binds RNAP II and recruited to several promoters Associated with mRNA 3′-processing complex Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,236,237,238,239,332,392] |
| eEF1Bδ | Chromatin-associated | Mammals and plants | [26,33] |
| eEF2 | Induces nuclear morphological changes and aneuploidy Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, and plants | [25,26,33,42,148,158] |
| eIF5A | Induction of apoptosis Associated with intranuclear filaments of NPC Binds the promoter of the HIF1α and activates its transcription Part of a specific RNA nuclear export pathway Associated with oncogenesis Associated with HP1a Chromatin-associated Found in the insoluble nuclear fraction | Mammals, flies, yeasts, and plants | [20,25,26,33,42,55,56,57,58,59,60,61,62,63,64,158,226,361,362,363,364,365,366,367,368,369,384,385,386,387,388,389] |
| eEFSec | Unknown | Humans and frogs | [31,32] |
| eRF1 | The quality control mechanism of maturing ribosomes Chromatin-associated | Mammals, yeasts, and plants | [20,25,35] |
| eRF3 | Found at active transcription sites Chromatin-associated | Mammals and flies | [150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachaev, Z.M.; Ivashchenko, S.D.; Kozlov, E.N.; Lebedeva, L.A.; Shidlovskii, Y.V. Localization and Functional Roles of Components of the Translation Apparatus in the Eukaryotic Cell Nucleus. Cells 2021, 10, 3239. https://doi.org/10.3390/cells10113239
Kachaev ZM, Ivashchenko SD, Kozlov EN, Lebedeva LA, Shidlovskii YV. Localization and Functional Roles of Components of the Translation Apparatus in the Eukaryotic Cell Nucleus. Cells. 2021; 10(11):3239. https://doi.org/10.3390/cells10113239
Chicago/Turabian StyleKachaev, Zaur M., Sergey D. Ivashchenko, Eugene N. Kozlov, Lyubov A. Lebedeva, and Yulii V. Shidlovskii. 2021. "Localization and Functional Roles of Components of the Translation Apparatus in the Eukaryotic Cell Nucleus" Cells 10, no. 11: 3239. https://doi.org/10.3390/cells10113239