
The Role of Symbiont-Targeted Strategies in the Management of Pentatomidae and Tephritidae Pests under an Integrated Vision


Department of Agricultural, Forest and Food Sciences, University of Torino, 10095 Grugliasco, TO, Italy
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(3), 868; https://doi.org/10.3390/agronomy13030868
Submission received: 11 January 2023
/
Revised: 10 March 2023
/
Accepted: 14 March 2023
/
Published: 16 March 2023
(This article belongs to the Special Issue Microbiome Innovation in Agriculture: Microbial Pest Control)
Abstract
:The interaction between insects and gut bacterial symbionts is, nowadays, regarded as an important element in the implementation of pest management, in consideration of the urgent need for sustainable alternatives to insecticide use. In this framework, a major tool is symbiotic control, with the main ready-to-use application represented by the interruption of obligate symbioses. Two insect families, namely Pentatomidae in the Hemiptera order and Tephritidae in Diptera, have been indicated as outstanding targets for symbiont-oriented control tactics. An important advantage of interrupting obligate symbioses is the target shift from insect to bacteria, which avoids insecticide use; however, the compatibility between this approach and other pest/disease management strategies is crucial to design low-impact pest control programs. Here we present the state of knowledge regarding the integration of symbiont manipulation in sustainable plant protection plans. Research assessing the potential for multitarget applications is reported, as well as studies on the impact of symbiont interruption on nontarget species. Besides symbiont-targeted pest control, another relevant outcome of symbiont manipulation is the restoration of microbial perturbation in mass-reared insects used in pest control programs, which is a required step to allow the success of other tactics, such as the Sterile Insect Technique. Despite the potential contribution that symbiont-targeted strategies may offer to integrated pest management, we point out that operational caveats may emerge in symbiont-oriented control in relation to the target extension on the label directions and to the number of required treatments. Future work is needed to increase the target range and the number of tested formulations exploiting the interruption of bacterial symbioses. This will also require assessment of the effect of different products on beneficial organisms, including biological control agents. Finally, the authorization of formulates for symbiotic control should be taken into consideration by the regulatory bodies, to really promote new readily available control options.
1. Introduction
The need for a more sustainable view of agriculture is a global priority, as expressed by the United Nations Sustainable Development Goals. The European Commission supports sustainable growth strategies with the adoption of the “European Green Deal” to promote solutions capable of preserving human health and environment quality, whilst also allowing economic development [1]. In this context, the Farm to Fork Strategy aims to promote novel and low-impact techniques to achieve sustainable food systems [2]. One of the major threats to the reduction of chemical agricultural input is the constant increase of crop vulnerability to pathogens and pests, which is exacerbated by climate change and global trade [3].
Recently, bacterial symbioses in insects have been regarded as powerful sources of sustainable options to achieve insecticide-free pest management [4]. Indeed, the wide range of symbiotic associations involving insects and bacteria is now largely recognized, as well as their powerful role in determining a huge increase in the metabolic and ecological capabilities of insects [5]. A major function of bacterial symbionts is nutrient provisioning, which is regarded as a key element in determining the insects’ capability to use diets of low or unbalanced nutritional composition, such as in sugar-feeding species [6]. The order Hemiptera includes a large number of species involved in strict relationships with obligate bacteria, whose contribution to nutrient supply integrates the host-feeding capability on plant saps [6]. However, many holometabolous insects have also been shown to rely on bacterial symbionts. For example, the gut microbiota of tephritid flies is thought to support the host when feeding on nitrogen-poor sources, such as honeydew [7,8]. Along with providing an important increase in knowledge about the metabolic processes for which insects benefit from symbiotic associations, the awareness of the role played by gut bacterial partners can be exploited for pest management, according to the symbiotic control techniques [9]. One of the most promising strategies to apply symbiotic control relies on the elimination of obligate symbionts to impair insect fitness. Conversely, the preservation of gut microbiota is crucial for maintaining the health and competitivity of mass-reared insects to be used in pest control programs, like those involving the Sterile Insect technique (SIT).
Insects that are especially susceptible to symbiont-targeted control are those species where obligate symbionts are extracellularly located and undergo an environmental route to be transmitted from the mother to the offspring (vertical transmission). In the species that exhibit this trait, symbiont interruption is favored as it is possible to prevent transgenerational symbiont transfer, rather than curing an existing symbiont infection. Examples of insects that have been proposed as the target for symbiotic control include stink bugs (Hemiptera: Pentatomidae) [4] and fruit flies (Diptera: Tephritidae) [10,11]. Recent work demonstrated the efficacy of symbiotic control against representatives of both families [12,13,14,15], emphasizing the beneficial outcomes of a target shift from insect to bacteria, with the consequent reduction of insecticide use [16,17]. Several substances with low environmental impact (e.g., naturally derived biostimulants and microbial biocontrol agents) showed a noticeable bactericidal effect against symbionts in their environmental stage [12,13,14,15], highlighting the potential environmental sustainability of symbiotic control. To maintain the low-impact status of pest control programs exploiting symbiotic control, it is essential to take into consideration the interplay between this method and other strategies. Indeed, the combination of control tactics is considered a key approach to improve plant protection programs, by promoting synergistic effects and limiting antagonistic interactions [18]; this will be especially relevant when combining non-chemical tools, due to their general lower persistence and lower knock-down effect. Biological control is one of the better-established methods, and it is widely used in plant protection programs oriented towards preserving the sustainability of agroecosystems [19]. A main issue to be addressed, when assessing the compatibility between the perturbation of symbiont transmission and biological control, is the effect of symbiont-targeting products on the performance of biocontrol agents attacking the egg. Biological control of tephritid fruit flies has been deeply investigated in several pest species; successful suppression was reported especially using parasitoids, including species that target the eggs [20]. Moreover, the most common and effective biocontrol agents against pentatomids are egg parasitoids [21]. Besides the occurrence of possible non-target effects on biocontrol agents, the effects related to the introduction of antimicrobial substances should be investigated in the entire agroecosystem. A different facet of the role of symbiotic control within the overall pest management plans is the potential multi-target effect. This may be obtained because symbiont-targeting products are antimicrobials [4,15]. Therefore, along with potentially preventing symbiont acquisition in distantly related insects [22], symbiont-killing products may allow the concurrent management of bacterial and fungal pathogens. Finally, bacterial symbionts may have a role in enhancing SIT. This approach has been proposed for both Tephritidae and Pentatomidae [23,24,25,26], even though the application on the latter family is not yet operative. SIT is applied by releasing in-field males of the target species which have been sterilized by irradiation; however, such a procedure is costly for insect performance. Although nothing is known regarding the effect of irradiation on the bacterial symbionts of pentatomids, significant damage was reported on the gut community structure of fruit flies after irradiation [27].
This review aims to examine the most recent work regarding the interaction between symbiont manipulation in the families Pentatomidae and Tephritidae and other pest control strategies, to maximize the outcome of integrated pest management (IPM) protocols that include the incorporation of symbiont-targeted measures (Figure 1). Symbiont-oriented methods may deal with offspring production, post-natal success, or juvenile fitness. Moreover, they may display a multi-target activity, but they must be respectful of non-target species. The shift of target that results from controlling insects through bacteria-oriented methods may entail some application caveats due to regulatory issues that are still to be solved. Symbiont-targeted strategies may also enhance, in a direct or indirect way, the outcome of other pest management methods.
2. The Importance of Gut Symbionts in Pentatomidae and Tephritidae
Insect species in the family Pentatomidae host obligate gut bacteria belonging to the genus Pantoea [17]. These symbionts are located in highly specialized structures, namely the crypts, in the terminal part of the midgut [28] (Figure 2A). The crypts are not in direct connection with the main gastric regions. Symbiont colonization of newly emerging nymphs starts in the early developmental phases at the first instar; before the host reaches the adult stage, intestinal wall development produces crypt isolation from the other gut regions [4]. Such a process guarantees symbiont protection from adverse factors in the host gut, despite the extracellular localization. The major role of obligate symbionts of pentatomids is nutrient provision, as widely documented for the bacterium associated with the brown marmorated stink bug Halyomorpha halys (Stål), named ‘Candidatus Pantoea carbekii’ [29]. Genomic investigations of this bacterium showed it has the potential to provide essential amino acids and vitamins [30]. However, the intensity of the relation between pentatomid species and their symbionts showed high variability, suggesting that gut bacteria of these insects are undergoing a transition from the facultative to the obligate status. For example, the experimental elimination of symbionts is lethal for H. halys, whereas the same process only moderately affects the growth of the southern green stink bug Nezara viridula L., with different outcomes observed in different populations [4]. Beside their nutritional role, very little is known regarding the possible additional benefits provided by Pantoea symbionts to Pentatomidae. No direct protective functions have been described for the symbionts of Pentatomidae so far; however, in the related Riptortus pedestris Fabricius (Alydidae), a fenitrothion-degrading symbiont in the genus Burkholderia provides insecticide resistance to the host [31]. Another aspect that is still poorly understood is the interactions occurring between primary symbionts of Pentatomidae and the complex microbiota residing in other gut compartments to establish symbiotic homeostasis [4]. However, a balanced microbial community is likely to be necessary to preserve the insect’s immune competence, allowing the full completion of digestive functions integrated by the obligate bacterial partners.
In tephritid flies, gut symbionts are located extracellularly in the intestine lumen or in the oesophageal bulb (Figure 2A), a unique organ where obligate symbionts multiply prior to being released into the digestive tract [32]. The main bacterial family that has been found associated with the gut of fruit flies is Enterobacteriaceae, with the most representative genera being Klebsiella, Enterobacter, Citrobacter, Serratia, Pantoea, and Erwinia [32]. Additionally, lactic acid bacteria and acetic acid bacteria have been found in fruit fly species, due to the acidic conditions of their midgut [33,34]. The gut microbiota of teprhitid fruit flies has been proven to enhance host performance, increasing insect size, longevity, and fecundity. Beneficial effects are provided through direct nutrient provisioning or by increasing the availability of nutrients [32]; in this case, the associations are often obligate. For example, the symbiont of Bactrocera oleae (Rossi), ‘Candidatus Erwinia dacicola’, plays an essential role in the host’s nitrogen metabolism [10]; whereas, the microbiota of Bactrocera dorsalis (Hendel) supports the degradation of complex polysaccharides into simple sugars [35]. Moreover, some gut symbionts of tephritids display many defensive traits, including the limitation of pathogen growth [36] and the detoxification of plant secondary metabolites or other xenobiotic compounds, such as insecticides [32]. For example, a gut bacterium found in B. dorsalis, namely Citrobacter freundii, is capable of degrading Trichlorphon, providing host resistance to this molecule [37]. Another defensive function of the gut microbiota of Teprhitidae is exerted by promoting the host’s immune homeostasis [32]. Since the immune system modulation is often maintained by a balanced microbial community, rather than by a single symbiont [38], even fly species that do not rely on an obligate symbiont can be seriously hampered by the perturbation of their microbiota. Still, the gut microbiota of tephritids is not static but rather shows high plasticity, which allows the host to adapt to different environments [39] and facilitates their invasive potential [40].
3. Extracellular Routes for Symbiont Vertical Transmission
The transgenerational transfer is a crucial step for the maintenance of obligate symbioses between insects and bacteria. The most studied obligate (primary) symbionts are found in the order Hemiptera and are thought to be responsible for the capability of members of this order to feed on plant saps, since they supply nutrients missing from the hosts’ diet [41]. These symbionts are mainly intracellularly located in specific insect organs, namely the bacteriomes; their vertical transmission is guaranteed by the migration of bacteria into the eggs during maturation in the ovaries [42]. For example, the aphid primary symbiont Buchnera aphidicola is transferred by exocytosis from maternal cells to the hemolymph, to immediately colonize the embryo by endocytosis [43]. The transovarial transmission and intracellular localization are derived from a long coevolutionary history, resulting in high functional and genetic specialization with strong genomic erosion. This specialization leads to a noticeable limitation in the risk of losing the obligate partner, as they are hardly altered by external factors. In contrast, obligate symbionts that have been more recently acquired are often extracellularly located and transmitted [42]. The extracellular status is also widespread among facultative symbionts [44]. The latter group may have a relevant nutritional impact on the host and obligate symbionts by supporting the degradation of complex or toxic substrates, hence, allowing the expansion of consumed plants. The transmission pathways of extracellular symbionts are highly diversified across insect families. The most common, especially among facultative associates, is the horizontal transfer (e.g., by sharing a common feeding substrate, by mating, or via trophallaxis) [45]; however, routes for vertical transmission have also been reported in the orders Blattodea, Hemiptera, Thysanoptera, Isoptera, Coleoptera, Diptera, and Hymenoptera [42]. Environmental acquisition is a widespread way to acquire stably associated partners, including obligate symbionts, as reported for Lygaeoidea and Coreoidea hemipterans [46]. Coprophagy allows the transfer of symbionts that are located in the gut lumen and excreted in feces, by exploiting the fecal probing of newborns [47]. This mechanism was reported for wood-feeding insects such as termites or cockroaches, as well as for the hematophagous heteropterans of the genus Rhodnius [47]. Moreover, different strategies have been developed by insects to inoculate symbiotic bacteria upon or close to eggs to favor the ingestion by the new generation. Egg smearing is a widespread way to allow the bacterial transfer to the progeny in distantly related orders, including many economically relevant Hemiptera, Coleoptera, and Diptera [42], such as the families Pentatomidae and Tephritidae (Table 1). Alternatively, the mother lays symbiont-containing capsules or jelly secretions close to the eggs, which are promptly ingested by the newborns [48]. Such a behavior is observed in Plataspidae and Urostylidae stink bugs, as well as in the tortoise leaf beetle Cassida rubiginosa Müller [17,49]. A shared trait of these strategies is the post-natal acquisition by each generation (differently from transovarial transmission), with the consequent need for bacteria to undergo an environmental phase that exposes them to external alterations. From an evolutionary point of view, these strategies appear as subsequent transitory steps before intracellular transmission (with environmental acquisition being the less evolved route and egg-related secretions being the most evolved ones) [42]. The extracellular route for vertical transmission can involve a single obligate symbiont, as reported for several species [50,51,52]. However, complex bacterial consortia are also vertically transferred; such is true for tephritid flies, where the interexchange between groups of bacterial taxa have been demonstrated, maintaining the same functions that allow the host to adapt to different diets according to the acquired symbionts [27]. The dramatic outcome of interrupting the acquisition of obligate symbionts by the new generation is evident, with symbiont-deprived juveniles showing markedly impaired development [47] (Figure 2B,C); however, inducing a dysbiotic condition is generally deleterious for insects whose microbiota is not dominated by a primary symbiont [27,53]. For example, in B. dorsalis, antibiotic treatment and egg disinfection, causing a significant alteration of the gut microbiota, resulted in reduced longevity when flies were reared on a nutrient-poor diet [8]. The gut dysbiosis may enhance (or be enhanced by) the sublethal effects of insect exposure to stressors (e.g., nutrient scarcity, contact with xenobiotics, radiation), leading to long-term survival reduction or other fitness defects [54,55] (Figure 2C,D).
4. Symbiotic Control through Symbiosis Disruption
Symbiotic control refers to all insect pest control methods founded upon the manipulation of microbial associates [9]. It was originally considered to correspond to paratransgenesis, the genetic manipulation of insect symbionts to alter insect traits [71], but it now includes strategies that avoid the use of genetically modified symbionts. Control methods that can be applied include the establishment of heterologous associations (i.e., the introduction of a new symbiotic strain to transfer desired traits), and the perturbation of beneficial symbioses (e.g., elimination of obligate symbionts, alteration of the microbiota balance) to create a dysbiotic status [9]. Symbiotic control strategies have been proposed to contain several insects of medical concern, such as vectors of disease agents, as well as many agricultural pests. In the latter group, recent attention was given to control methods exploiting the interruption of the vertical transmission of obligate symbionts, with particular efforts made on tephritid fruit flies and pentatomid stink bugs (Table 2). The detrimental effect derived from symbiont deprivation in pentatomid nymphs was indicated by several authors [4]. Impairing symbiont acquisition by neonate nymphs is relatively easy in this group of insects, as egg masses are highly exposed to environmental perturbation; however, only a few reports of symbiotic control protocols that are ready for field application have been produced, and they exclusively address H. halys (Table 2). Containment of this species was proposed using commercial products with antimicrobial activity, including chemical bactericides and fungicides, copper-based fertilizers, and microbial biopesticides [13,14,72]. The control of Tephritidae is complicated by the more complex gut microbiota associated with this family, since only the olive fruit fly B. oleae is dominated by a single obligate symbiont, namely ‘Ca. Erwinia dacicola’ [50]. The pest control potential for impairing the larval acquisition of ‘Ca. E. dacicola’ has been widely investigated [12,15]; however, the oviposition behavior of B. oleae, which places eggs endophytically in the olive pulp, hampers the efficiency of symbiotic control strategies. Several antimicrobial formulations, such as fungicides and plant biostimulants, revealed effective containment of B. oleae field populations [11,22,73,74] (Table 2). However, they mostly showed oviposition deterrence, whereas their anti-symbiont effect has been suggested but is still little explored.
Symbiont-targeted control has also been proposed for insects that do not rely on a single obligate symbiont, through the alteration of gut microbial balance by delivering substances that kill facultative symbionts. For example, the use of antibiotics have been suggested to control the fall armyworm Spodoptera frugiperda (Smith) [75]; nevertheless, at present, the use of antibiotics against agricultural pests is highly controversial [76]. As for pentatomids and teprhitids, no examples of field containment performed through the perturbation of facultative symbioses are available; this field deserves future attention.
5. Symbiont-Targeted Strategies for Multiple Pest Control
A main strength of symbiotic control is the combination of high selectivity (due to the specific symbiotic target) with a potential multi-target effect, as all pest species that undergo the same vertical transmission process may be contained. This is especially valuable for pentatomids, as many species commonly infest the same crop. For example, in hazelnut groves of northern Italy and Georgia, which were recently invaded by H. halys, other pentatomids such as Palomena prasina L. and Nezara viridula L. were also consistently found; additionally, they were both shown to lay eggs on the crop at the same time as H. halys [81]. Since the vertical transmission of obligate Pantoea spp. symbionts through egg smearing is a conserved trait in Pentatomidae [4], the application of symbiotic control treatments is expected to produce a similar outcome on different species. However, little work has been conducted to assess the effect of symbiotic control against pentatomids other than H. halys [4], and comprehensive field studies are required to estimate the effectiveness of this technique on a complex pest community.
The possibility of containing multiple insect pests at the same time through symbiotic control performed against H. halys is facilitated by the wide occurrence of this broad-range species in many crops, where it could likely co-exist with other potential targets. The pest has been also documented to attack olive trees, where it is considered a serious threat in invaded areas [82]. Therefore, the possible combined effect of symbiotic control against H. halys and B. oleae may be obtained, as the ovipositional period of the two pests is largely overlapped. Furthermore, insect containment can be regarded as a possible accessory effect of treatments that are already applied to control olive pathogens or as biostimulants [80]. Indeed, olive is susceptible to many bacterial and fungal pathogens (e.g., the Gram-negative bacterium Pseudomonas savastanoi which causes olive knot disease; the pathogenic fungi Spilocaea oleaginea which causes olive leaf spot; or Colletotrichum acutatum and Colletotrichum gloeosporioides, the agents of anthracnose disease [83,84]). Such pathogens may require treatments with copper or other fungicides (including biopesticides such as Trichoderma or Bacillus strains) [84,85]. Summer treatments performed for the symbiotic control of B. oleae and/or H. halys may also prove effective in limiting fungal pathogens, although the possible combined effect is yet to be explored. For example, the commercial biofungicide-bactericide Amylo-X®, containing the biocontrol agent (BCA) Bacillus velezensis, was shown to induce significant mortality on H. halys neonates after egg mass treatment, due to the missed acquisition of the symbiont ‘Ca. Pantoea carbekii’ [14]. Interestingly, nymphal mortality was also reported after direct or residual contact; this suggests that the BCA may display additional (yet uncharacterized) antagonistic activity against the insect, opening a novel field of multiple pesticidal activity. Such a process may take place in a similar way for biopesticides containing antifungal Trichoderma strains, for which insecticidal properties have been demonstrated [86]. However, whether B. velezensis could be included in olive protection protocols, targeting both plant pathogens and insect pests, still needs to be demonstrated. Beside the field efficacy of the proposed approach, a further important issue that must be considered is the application timing, since microbial biopesticides generally show low persistence and the optimal treatment time may not coincide for different targets.
6. Symbiont-Targeted Strategies and Biological Control
One of the major challenges in the next generation of pest control is minimizing the side effects on non-target fauna, including beneficial insects such as pollinators and natural enemies [87,88]. Besides the obvious injury related to broad-spectrum insecticides, many low-impact molecules or organisms, including those that do not specifically target insects, also show lethal or sub-lethal effects on beneficials [89,90]. Therefore, to include symbiotic control in IPM programs, it is crucial to include a specific risk assessment with respect to the effect on non-target insects. Several formulations of copper-based fungicides have been tested for their side effects on the main parasitoids of B. oleae, such as Psyttalia concolor Szépligeti, Aphidius gifuensis Ashmead, and Aphidius colemani Viereck; they were mostly non-harmful or slightly harmful for parasitoid females, and no detriments were recorded after the treatment of parasitized pupae [91]. Moreover, treatment with copper-based products did not alter the total arthropod diversity and abundance, when considering phytophagous, predators, and parasitoids [92]. Therefore, control strategies based on these formulations (including symbiotic control) are considered compatible with parasitoid BCAs. On the other hand, nothing is known about the possible effects of fungicidal anti-symbiont treatments on the bacterial and fungal microbiota of parasitoids or other beneficial insects. A sublethal impairment of insect-symbionts associations in non-target species may be detrimental for the agroecosystem food webs. Furthermore, if a few symbiont-deprived target insects survive the treatment they may become the prey for other animals, possibly transferring persistent antimicrobial molecules throughout the food chain and threatening potential for the development of resistance mechanisms in the environment. Therefore, we suggest that this aspect should be included in a comprehensive risk assessment of symbiont-targeted strategies.
The short-term impact on egg parasitism of the symbiont-targeted treatment of eggs was tested using the micronutrient biocomplex Dentamet® [93], which was previously shown to produce high nymphal mortality in H. halys [13]. The effect was evaluated considering the parasitoid species introduced in Italy attacking H. halys (namely Trissolcus japonicus (Ashmead) and Trissolcus mitsukurii (Ashmead)), as well as native wasps (Anastatus bifasciatus (Geoffroy), Ooencyrtus telenomicida (Vassiliev), and Trissolcus kozlovi Rjachovskij) [93]. Exposing H. halys egg masses to Dentamet® showed no direct detrimental effect on female wasps; besides, it did not affect emergence percentages of any parasitoid species, with the only exception of T. mitsukurii. Additionally, T. japonicus did not show any preference when exposed to treated and untreated egg masses in paired choice tests, whereas T. mitsukurii preferred untreated eggs. Therefore, the combination of symbiotic control and biological control is regarded as a safe option for crops subject to H. halys attacks. On the other hand, treatment with the biocomplex reduced the non-reproductive effects on native wasp species [93]. The reduction of these effects may be of little concern due to the low parasitization efficiency of non-coevolved parasitoid species on H. halys egg masses; however, the possible emergence of long-term, sub-lethal alterations of their performance should not be overlooked and should be extended to other products for a deeper risk assessment, once the direct toxicity to beneficial insects is excluded.
7. Symbiont-Targeted Strategies to Enhance SIT
The manipulation of the gut microbiota has the potential to provide significant benefits to the implementation of SIT programs, with special regard to the enhancement of insect rearing before field release. A number of reports have been produced describing an alteration of the gut bacterial community in lab-reared insects [94]. Relevant changes are observed in response to rearing on artificial diets, with a reported simplification of the microbiota [95,96,97]; likewise, the irradiation produced a dramatic perturbation of bacterial community [98,99,100]. The microbiota alteration can result in impaired fitness, with lower male competition after the field release [94]. Bacterial supplements have been proposed to restore the gut microbiota homeostasis of reared tephritid flies used for SIT; however, different studies produced inconsistent results [27]. Reports were produced showing that the delivered microbial consortia did not cause any impact on the fly fitness [101], while others showed a positive or negative outcome for the host [27,102,103,104]. This variability is likely to reflect the complexity of interactions between different fly hosts and their gut microbiota, and is also related to the supplemented insect stage and/or the type of diet. For example, the administration of Pseudomonas putida to B. oleae was beneficial for the female fecundity when they were reared on a diet containing only sugar, whereas it had no effect when the insects were fed with a complete diet [105]. A more significant benefit is obtained by supplementing flies with bacteria in the larval diet. In Ceratitis capitata Wiedemann, provisioning a bacterial cocktail to larvae induced several fitness improvements, including weight and size increase, and reduced developmental time [106,107]. In B. dorsalis, the delivery of a symbiotic strain of Klebsiella oxytoca restored the host’s ecological fitness after irradiation [108]. Therefore, the development of larval supplements is regarded as a major improvement for tephritid mass rearing in ameliorate SIT programs [27].
8. Potential Roles of Symbiont-Targeted Strategies in Pest Control Implementation
Under the perspective of integrating symbiotic control with other strategies for pest containment in the field, an outstanding possibility to be explored is related to the sub-lethal effects that have been recorded in several symbiont-deprived insects [8,32,67,109]. The reduced fitness of dysbiotic insects has been proven to increase their susceptibility to stress-inducing agents [110,111,112]. No studies have tackled the effects of exposing dysbiotic insects to low doses of chemical insecticides, even though the correlation between several active substances and microbiota perturbation has been widely documented [55,113,114]. The impaired health status of insects after exposure to symbiont-killing treatments may produce a synergistic effect with subsequent insecticide delivery, and may support the reduction of the application dose. Similarly, a synergistic interaction between SIT and symbiotic control may be supposed, as irradiation may increase insect susceptibility to dysbiotic effects. However, the biological traits induced by a mixed approach are still unknown. For example, whether irradiation alters the symbiotic colonization by obligate symbionts is still undetermined. Greater knowledge on this aspect would support further investigation as to whether susceptibility to the detrimental effect of bacterial perturbation on a new generation is augmented after parental irradiation. If so, a strategy combining SIT and symbiotic control may be useful when only partial sterility can be obtained, as observed in the case of stink bugs [23,25]. The outcome of a combined strategy would be also ameliorated as symbiotic control may compensate for the reduced competitive fitness of irradiated males, which has often been observed in both fruit flies and stink bugs [115,116,117]. Indeed, if the wild males would outcompete irradiated ones in mating in the field, symbiotic control may target their newly emerged offspring. Since the use of symbiont-targeted strategies for pest control is currently increasing in relevance, a comprehensive study of possible synergistic effects with other approaches could provide important information to remodulate plant protection plans to achieve pesticide reduction.
Another symbiotic target that could be relevant to increase insect susceptibility to chemical control is represented by the facultative associates that induce insecticide resistance [118]. The modulation of insect gut microbiota to promote insecticide susceptibility has been proposed as a novel approach for enhanced pest control [119]. However, at present, no studies have addressed the real outcome of combining anti-symbiont and insecticides in terms of increased containment of field populations; besides, several parameters of insect associations with pesticide-degrading bacteria are still to be uncovered, including their incidence in many pest species and populations.
9. Operational Constraints in Symbiotic Control Programs
Although symbiotic control proved extremely valuable within IPM programs combining different strategies, it still shows important applicative limitations. A main concern is related to the total number of treatments that may be necessary to contain many targets in the same crop. For example, fungicides are mainly applied in autumn and spring in olive groves against different pathogens, whereas the main oviposition period for both B. oleae and H. halys is in summer. A preventive effect of fungicidal treatments against B. oleae has been suggested given a reported oviposition deterrence [22]; nevertheless, the persistence of anti-symbiont activity is less likely to be maintained, especially considering biofungicides or other low-impact molecules. Therefore, although the multi-target effect is mostly expected to allow an overall input reduction, the required number of treatments may be increased sporadically by the need to cover the whole pest flight season, with several adverse implications. First, the use of many active substances is subject to regulatory restrictions, in terms of the number of treatments and doses. For example, the use of copper-based products is limited by an annual threshold for the amount of active substance [120]; furthermore, it must be pointed out that copper hydroxide and copper oxychloride are included in the EU candidate for substitution list (Reg. (EC) No 1107/2009). Hence, a correct rotation of active substances with different modes of action must be conducted; however, the success of such plans may be hampered by the still small number of products that have been deemed effective for symbiotic control. Additionally, most products whose effectiveness has already been demonstrated are commercialized only in restricted geographical areas, due to authorization limitations. Moreover, even where these products are authorized, they are not registered for use against insects as they are not insecticides; therefore, the extension of authorized targets in the label directions of each product is necessary to justify an increased number of applications. This process is hampered by slow registration and authorization procedures, which are often unsuitable to define the complexity of multi-purpose approaches (e.g., different effects on different targets) [121]. The limitation may be even larger for those products that are registered as biostimulants, since in most countries they are subject to different regulations than plant protection products [122]. Finally, even if the mentioned restraints are overcome, the increased cost of pest management strategies requiring the repetition of numerous treatments to cover all possible targets may become economically inconvenient.
10. Conclusive Remarks
The increase in knowledge on the effectiveness of symbiotic control protocols, based on the disruption of obligate bacterial associations in insects, largely supports the introduction of this approach into integrated pest control programs. Symbiont-targeted tactics are sustainable control measures that are well-fitted to keep pest-related damages under the economic threshold and to, simultaneously, exploit the ecosystem services derived from enhanced agro-biodiversity. This research field is still taking its first steps, and many aspects should be further clarified to assess its real target range. Additionally, the compatibility of different symbiont-oriented products with other strategies and with non-target organisms should be further investigated for the practical design of rational plant protection plans. Finally, the legislative framework that regulates the authorization of formulates should consider the increased complexity of the newest proposed strategies, including a still underexplored, multipurpose potential.
Author Contributions
E.G.: Writing original draft preparation; Literature search and figure making; A.A.: Writing—review and editing. All authors made direct and intellectual contributions to the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Delgado, L.; Garino, C.; Moreno, F.J.; Zagon, J.; Broll, H. Sustainable food systems: EU regulatory framework and contribution of insects to the Farm-To-Fork strategy. Food Rev. Int. 2022, 105, 104760. [Google Scholar] [CrossRef]
- Purnhagen, K.P.; Clemens, S.; Eriksson, D.; Fresco, L.O.; Tosun, J.; Qaim, M.; Visser, R.G.F.; Weber, A.P.M.; Wesseler, J.H.H.; Zilberman, D. Europe’s Farm to Fork Strategy and its commitment to biotechnology and organic farming: Conflicting or complementary goals? Trends Plant Sci. 2021, 26, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Hatt, S.; Osawa, N. Beyond “greening”: Which paradigms shape sustainable pest management strategies in the European Union? BioControl 2019, 64, 343–355. [Google Scholar] [CrossRef]
- Gonella, E.; Orrù, B.; Marasco, R.; Daffonchio, D.; Alma, A. Disruption of host-symbiont associations for the symbiotic control and management of pentatomid agricultural pest—A review. Front. Microbiol. 2020, 11, 547031. [Google Scholar] [CrossRef] [PubMed]
- Coolen, S.; Magda, R.D.; Welte, C.U. The secret life of insect-associated microbes and how they shape insect-plant interactions. FEMS Microbiol. Ecol. 2022, 98, fiac083. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yosef, M.; Aharon, Y.; Jurkevitch, E.; Yuval, B. Give us the tools and we will do the job: Symbiotic bacteria affect olive fly fitness in a diet-dependent fashion. Proc. R. Soc. B Biol. Sci. 2010, 277, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Akami, M.; Tamgue, O.; Ren, X.; Wang, Y.; Qi, X.; Luther, K.M.M.; Ngane, R.A.N.; Niu, C.-Y. Effects of symbiotic population impairment on microbiome composition and longevity of Bactrocera dorsalis. R. Soc. Open Sci. 2021, 8, 211104. [Google Scholar] [CrossRef]
- Arora, A.K.; Douglas, A.E. Hype or opportunity? Using microbial symbionts in novel strategies for insect pest control. J. Insect Physiol. 2017, 103, 10–17. [Google Scholar] [CrossRef]
- Noman, M.S.; Liu, L.; Bai, Z.; Li, Z. Tephritidae bacterial symbionts: Potentials for pest management. Bull. Entomol. Res. 2020, 110, 1–14. [Google Scholar] [CrossRef]
- Bigiotti, G.; Sacchetti, P.; Pastorelli, R.; Lauzon, C.R.; Belcari, A. Bacterial symbiosis in Bactrocera oleae, an Achilles’ heel for its pest control. Insect Sci. 2021, 28, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Bigiotti, G.; Pastorelli, R.; Belcari, A.; Sacchetti, P. Symbiosis interruption in the olive fly: Effect of copper and propolis on Candidatus Erwinia dacicola. J. Appl. Entomol. 2019, 143, 357–364. [Google Scholar] [CrossRef]
- Gonella, E.; Orrù, B.; Alma, A. Egg masses treatment with micronutrient fertilizers has a suppressive effect on newly-emerged nymphs of the brown marmorated stink bug Halyomorpha halys. Entomol. Gen. 2019, 39, 231–238. [Google Scholar] [CrossRef]
- Gonella, E.; Orrù, B.; Alma, A. Symbiotic control of Halyomorpha halys using a microbial biopesticide. Entomol. Gen. 2022, 42, 901–909. [Google Scholar] [CrossRef]
- Sinno, M.; Bézier, A.; Vinale, F.; Giron, D.; Laudonia, S.; Garonna, A.P.; Pennacchio, F. Symbiosis disruption in the olive fruit fly, Bactrocera oleae (Rossi), as a potential tool for sustainable control. Pest Manag. Sci. 2020, 76, 3199–3207. [Google Scholar] [CrossRef]
- Nobre, T. Symbiosis in sustainable agriculture: Can olive fruit fly bacterial microbiome be useful in pest management? Microorganisms 2019, 7, 238. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.; Wu, W.; Sun, Z.; Chen, J.; Li, H. The gut microbiota of the insect infraorder Pentatomomorpha (Hemiptera: Heteroptera) for the light of ecology and evolution. Microorganisms 2021, 9, 464. [Google Scholar] [CrossRef]
- Suckling, D.M.; Tobin, P.C.; McCullough, D.G.; Herms, D.A. Combining tactics to exploit Allee effects for eradication of alien insect populations. J. Econ. Entomol. 2012, 105, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Maurya, R.P.; Koranga, R.; Samal, I.; Chaudhary, D.; Paschapur, A.U.; Sreedhar, M.; Manimala, R.N. Biological control: A global perspective. Int. J. Trop. Insect Sci. 2022, 42, 3203–3220. [Google Scholar] [CrossRef]
- Dias, N.P.; Montoya, P.; Nava, D.E. A 30-year systematic review reveals success in tephritid fruit fly biological control research. Entomol. Exp. Appl. 2022, 170, 370–384. [Google Scholar] [CrossRef]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G.; et al. Biological control of invasive stink bugs: Review of global state and future prospects. Entomol. Exp. Appl. 2021, 169, 28–51. [Google Scholar] [CrossRef]
- Checchia, I.; Perin, C.; Mori, N.; Mazzon, L. Oviposition deterrent activity of fungicides and low-risk substances for the integrated management of the olive fruit fly Bactrocera oleae (Diptera, Tephritidae). Insects 2022, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Horrocks, K.J.; Welsh, T.; Carpenter, J.E.; Suckling, D.M. Egg sterilisation of irradiated Nezara viridula (Hemiptera: Pentatomidae). Insects 2020, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Nguyen, H.N.; Stringer, L.D.; Hong, K.-J. Influence of irradiation on the biology of the brown marmorated stink bug (Hemiptera: Pentatomidae). J. Econ. Entomol. 2021, 114, 1211–1218. [Google Scholar] [CrossRef]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of gamma irradiation on the fecundity, fertility, and longevity of the invasive stink bug pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Chapman, T.A.; Shuttleworth, L.A.; Riegler, M.; Reynolds, O.L. Tephritid-microbial interactions to enhance fruit fly performance in sterile insect technique programs. BMC Micriobiol. 2019, 19, 287. [Google Scholar] [CrossRef] [Green Version]
- Ohbayashi, T.; Takeshita, K.; Kitagawa, W.; Nikoh, N.; Koga, R.; Meng, X.Y.; Tago, K.; Hori, T.; Hayatsu, M.; Asano, K.; et al. Insect’s intestinal organ for symbiont sorting. Proc. Natl. Acad. Sci. USA 2015, 112, E5179–E5188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, R.; Michel, A.P.; Sabree, Z.L. The crypt-dwelling primary bacterial symbiont of the polyphagous pentatomid pest Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2014, 43, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, L.J.; Meulia, T.; Sabree, Z.L. Habitat visualization and genomic analysis of ‘Candidatus Pantoea carbekii’, the primary symbiont of the brown marmorated stink bug. Genome Biol. Evol. 2015, 7, 620–635. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.F.; Yao, Z.; Bai, S.; Cai, Z.; Zhang, H. Tephritidae fruit fly gut microbiome diversity, function and potential for applications. Bull. Entomol. Res. 2020, 110, 423–437. [Google Scholar] [CrossRef]
- Kounatidis, I.; Crotti, E.; Sapountzis, P.; Sacchi, L.; Rizzi, A.; Chouaia, B.; Bandi, C.; Alma, A.; Daffonchio, D.; Mavragani-Tsipidou, P.; et al. Acetobacter tropicalis is a major symbiont of the olive fruit fly (Bactrocera oleae). Appl. Environ. Microbiol. 2009, 75, 3281–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaochan, N.; Drew, D.; Chinajariyawong, A.; Sunpapao, A.; Pornsuriya, C. Gut bacterial community structure of two Australian tropical fruit fly species (Diptera: Tephritidae). Songklanakarin J. Sci. Technol. 2015, 37, 617–624. [Google Scholar]
- Saha, P.; Ray, R.R. Production of polysaccharide degrading enzymes by the gut microbiota of Leucinodes Orbonalis and Bactrocera dorsalis. J. Entomol. Zool. Stud. 2015, 3, 122–125. [Google Scholar]
- Ben Ami, E.; Yuval, B.; Jurkevitch, E. Manipulation of the microbiota of mass-reared Mediterranean fruit flies Ceratitis capitata (Diptera: Tephritidae) improves sterile male sexual performance. ISME J. 2010, 4, 28–37. [Google Scholar]
- Cheng, D.; Guo, Z.; Riegler, M.; Xi, Z.; Liang, G.; Xu, Y. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.H.; Kim, S.H.; Lee, H.Y.; Bai, J.Y.; Nam, Y.D.; Bae, J.W.; Lee, D.G.; Shin, S.C.; Ha, E.M.; Lee, W.J. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behar, A.; Yuval, B.; Jurkevitch, E. Community structure of the Mediterranean fruit fly microbiota: Seasonal and spatial sources of variation. Isr. J. Ecol. Evol. 2008, 54, 181–191. [Google Scholar] [CrossRef]
- Hafsi, A.; Delatte, H. Enterobactereaceae symbiont as facilitators of biological invasion: Review on Tephritidae fruit flies. Biol. Invasions 2022, 1–16. [Google Scholar] [CrossRef]
- Baumann, P. Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. R. Soc. B 2015, 282, 20142957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, R.; Meng, X.Y.; Tsuchida, T.; Fukatsu, T. Cellular mechanism for selective vertical transmission of an obligate insect symbiont at the bacteriocyte embryo interface. Proc. Natl. Acad. Sci. USA 2012, 109, E1230–E1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudakaran, S.; Kost, C.; Kaltenpoth, M. Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol. 2017, 25, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. An ancient but promiscuous host-symbiont association between Burkholderia gut symbionts and their heteropteran hosts. ISME J. 2011, 5, 446–460. [Google Scholar] [CrossRef] [Green Version]
- Ohbayashi, T.; Mergaert, P.; Kikuchi, Y. Host-symbiont specificity in insects: Underpinning mechanisms and evolution. Adv. Insect Physiol. 2020, 58, 27–62. [Google Scholar]
- Kaiwa, N.; Hosokawa, T.; Nikoh, N.; Tanahashi, M.; Moriyama, M.; Meng, X.Y.; Maeda, T.; Yamaguchi, K.; Shigenobu, S.; Ito, M.; et al. Symbiont-supplemented maternal investment underpinning host’s ecological adaptation. Curr. Biol. 2014, 24, 2465–2470. [Google Scholar] [CrossRef] [Green Version]
- Salem, H.; Bauer, E.; Kirsch, R.; Berasategui, A.; Cripps, M.; Weiss, B.; Koga, R.; Fukumori, K.; Vogel, H.; Fukatsu, T.; et al. Drastic genome reduction in an herbivore’s pectinolytic symbiont. Cell 2017, 171, 1520–1531. [Google Scholar] [CrossRef] [Green Version]
- Capuzzo, C.; Firrao, G.; Mazzon, L.; Squartini, A.; Girolami, V. “Candidatus Erwinia dacicola”, a coevolved symbiotic bacterium of the olive fly Bactrocera oleae (Gmelin). Int. J. Syst. Evol. Microbiol. 2005, 55, 1641–1647. [Google Scholar] [CrossRef]
- Taylor, C.M.; Coffey, P.L.; DeLay, B.D.; Dively, G.P. The importance of gut symbionts in the development of the brown marmorated stink bug, Halyomorpha halys (Stål). PLoS ONE 2014, 9, e90312. [Google Scholar] [CrossRef]
- Pons, I.; González Porras, M.Á.; Breitenbach, N.; Berger, J.; Hipp, K.; Salem, H. For the road: Calibrated maternal investment in light of extracellular symbiont transmission. Proc. R. Soc. B 2022, 289, 20220386. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cai, M.; Zheng, X.; Xie, X.; Zhu, Y.; Long, Y. Impact of disinfectants on the intestinal bacterial symbionts and immunity of silkworm (Bombyx mori L.). Environ. Sci. Pollut. Res. 2022, 29, 79545–79554. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Raychoudhury, R.; Cai, Y.; Sun, Y.; Lietze, V.-U.; Peterson, B.F.; Scharf, M.E.; Boucias, D.G. Molecular signatures of nicotinoid-pathogen synergy in the termite gut. PLoS ONE 2015, 10, e0123391. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Yu, J.; Qin, Z.; Liu, X.; Zhao, X.; Hu, X.; Yu, R.; Wang, Q.; Yang, J.; Shi, Y.; et al. Guadipyr, a new insecticide, induces microbiota dysbiosis and immune disorders in the midgut of silkworms (Bombyx mori). Environ. Pollut. 2021, 286, 117531. [Google Scholar] [CrossRef] [PubMed]
- Viale, E.; Martinez-Sañudo, I.; Brown, J.M.; Simonato, M.; Girolami, V.; Squartini, A.; Bressan, A.; Faccoli, M.; Mazzon, L. Pattern of association between endemic Hawaiian fruit flies (Diptera, Tephritidae) and their symbiotic bacteria: Evidence of cospeciation events and proposal of “Candidatus Stammerula trupaneae”. Mol. Phylogenet. Evol. 2015, 90, 67–79. [Google Scholar] [CrossRef]
- Selivon, D.; Morgante, J.S.; Ribeiro, A.F.; Perondini, A.L.P. Extrusion of masses of yolk during embryonary development of the fruit fly Anastrepha fraterculus. Invertebr. Reprod. Dev. 1996, 29, 1–7. [Google Scholar] [CrossRef]
- Salgueiro, J.; Nussenbaum, A.L.; Milla, F.H.; Asimakis, E.; Goane, L.; Ruiz, M.J.; Bachmann, G.E.; Vera, M.T.; Stathopoulou, P.; Bourtzis, K.; et al. Analysis of the gut bacterial community of wild larvae of Anastrepha fraterculus sp. 1: Effect of host fruit, environment, and prominent stable associations of the genera Wolbachia, Tatumella, and Enterobacter. Front. Microbiol. 2022, 13, 822990. [Google Scholar] [CrossRef]
- Guo, Z.; Lu, Y.; Yang, F.; Zeng, L.; Liang, G.; Xu, Y. Transmission modes of a pesticide-degrading symbiont of the oriental fruit fly Bactrocera dorsalis (Hendel). Appl. Microbiol. Biotechnol. 2017, 101, 8543–8556. [Google Scholar] [CrossRef]
- Estes, A.M.; Hearn, D.J.; Bronstein, J.L.; Pierson, E.A. The olive fly endosymbiont, “Candidatus Erwinia dacicola,” switches from an intracellular existence to an extracellular existence during host insect development. Appl. Environ. Microbiol. 2009, 75, 7097–7106. [Google Scholar] [CrossRef] [Green Version]
- Kashkouli, M.; Fathipour, Y.; Mehrabadi, M. Heritable gammaproteobacterial symbiont improves the fitness of Brachynema germari Kolenati (Hemiptera: Pentatomidae). Environ. Entomol. 2019, 48, 1079–1087. [Google Scholar] [CrossRef]
- Mazzon, L.; Piscedda, A.; Simonato, M.; Martinez-Sañudo, I.; Squartini, A.; Girolami, V. Presence of specific symbiotic bacteria in flies of the subfamily Tephritinae (Diptera Tephritidae) and their phylogenetic relationships: Proposal of ‘Candidatus Stammerula tephritidis’. Int. J. Syst. Evol. Microbiol. 2008, 58, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Lauzon, C.R.; Mccombs, S.D.; Potter, S.E.; Peabody, N.C. Establishment and vertical passage of Enterobacter (Pantoea) agglomerans and Klebsiella pneumoniae through all life stages of the Mediterranean fruit fly (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2009, 102, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Otero-Bravo, A.; Goffredi, S.; Sabree, Z.L. Cladogenesis and genomic streamlining in extracellular endosymbionts of tropical stink bugs. Genome Biol. Evol. 2018, 10, 680–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, Y.; Fukatsu, T. Diversity of Wolbachia endosymbionts in heteropteran bugs. Appl. Environ. Microbiol. 2003, 69, 6082–6090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamipour, N.; Fathipour, Y.; Mehrabadi, M. Removal of gut symbiotic bacteria negatively affects life history traits of the shield bug, Graphosoma lineatum. Ecol. Evol. 2021, 11, 2515–2523. [Google Scholar] [CrossRef]
- Prado, S.S.; Almeida, P.P. Role of symbiotic gut bacteria in the development of Acrosternum hilare and Murgantia histrionica. Entomol. Exp. Appl. 2009, 132, 21–29. [Google Scholar] [CrossRef]
- Geerinck, M.W.J.; Van Hee, S.; Gloder, G.; Crauwels, S.; Colazza, S.; Jacquemyn, H.; Cusumano, A.; Lievens, B. Diversity and composition of the microbiome associated with eggs of the Southern green stinkbug, Nezara viridula (Hemiptera: Pentatomidae). MicrobiologyOpen 2022, 11, e1337. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Ishii, Y.; Nikoh, N.; Fujie, M.; Satoh, N.; Fukatsu, T. Obligate bacterial mutualists evolving from environmental bacteria in natural insect populations. Nat. Microbiol. 2016, 1, 15011. [Google Scholar] [CrossRef] [PubMed]
- Bistolas, K.S.I.; Sakamoto, R.I.; Fernandes, J.A.M.; Goffredi, S.K. Symbiont polyphyly, co-evolution, and necessity in pentatomid stinkbugs from Costa Rica. Front. Microbiol. 2014, 5, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, A.E. Symbiotic microorganisms: Untapped resources for insect pest control. Trends Biotechnol. 2007, 25, 338–342. [Google Scholar] [CrossRef]
- Taylor, C.M.; Johnson, V.; Dively, G. Assessing the use of antimicrobials to sterilize brown marmorated stink bug egg masses and prevent symbiont acquisition. J. Pest Sci. 2017, 90, 1287–1294. [Google Scholar] [CrossRef]
- Belcari, A.; Sacchetti, P.; Rosi, M.C.; Del Pianta, R. Control of the olive fly (Bactrocera oleae) through the use of copper products in Central Italy. IOBC-WPRS Bull. 2005, 28, 45–48. [Google Scholar]
- Caleca, V.; Lo Verde, G.; Lo Verde, V.; Palumbo Piccionello, M.; Rizzo, R. Control of Bactrocera oleae and Ceratitis capitata in organic orchards: Use of clays and copper products. Acta Hortic. 2010, 873, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhou, H.; Lai, Y.; Chen, Q.; Yu, X.Q.; Wang, X. Gut microbiota dysbiosis influences metabolic homeostasis in Spodoptera frugiperda. Front. Microbiol. 2021, 12, 727434. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Mojdehi, M.R.A.; Keyhanian, A.A.; Rafiei, B. Application of oviposition deterrent compounds for the control of olive fruit fly, Bactrocera oleae Rossi. (Dip. Tephritidae) control. Int. J. Trop. Insect Sci. 2022, 42, 63–70. [Google Scholar] [CrossRef]
- Gonçalves, F.; Torres, L. Effect of copper oxychloride on the olive infestation by Bactrocera oleae in Northeastern Portugal. Acta Hortic. 2012, 949, 333–340. [Google Scholar] [CrossRef]
- Tsolakis, H.; Ragusa, E.; Tarantino, P. Control of Bactrocera oleae by low environmental impact methods: NPC methodology to evaluate the efficacy of lure-and-kill method and copper hydroxide treatments. Bull. Insectol. 2011, 64, 1–8. [Google Scholar]
- Daher, E.; Cinosi, N.; Chierici, E.; Rondoni, G.; Famiani, F.; Conti, E. Field and laboratory efficacy of low-impact commercial products in preventing olive fruit fly, Bactrocera oleae, infestation. Insects 2022, 13, 213. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- Zapponi, L.; Morten, M.; Chiesa, S.G.; Angeli, G.; Borri, G.; Mazzoni, V.; Sofia, M.; Anfora, G. Brown marmorated stink bug (Halyomorpha halys) feeding damage determines early drop in olive crops. J. Appl. Entomol. 2022, 146, 791–795. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Faedda, R.; Sinatra, F.; Agosteo, G.E.; Schena, L.; Frisullo, S.; Magnano di San Lio, G. Olive Anthracnose. J. Plant Pathol. 2012, 94, 29–44. [Google Scholar]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The role of microbial inoculants on plant protection, growth stimulation, and crop productivity of the olive tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef]
- Kir, A.; Cetinel, B.; Sevim, D.; Gungor, F.O.; Rayns, F.; Touliatos, D.; Schmutz, U. Agroecological screening of copper alternatives for the conservation of soil health in organic olive production. Agronomy 2022, 12, 1712. [Google Scholar] [CrossRef]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Cappa, F.; Baracchi, D.; Cervo, R. Biopesticides and insect pollinators: Detrimental effects, outdated guidelines, and future directions. Sci. Total Environ. 2022, 837, 155714. [Google Scholar] [CrossRef]
- Serrão, J.E.; Plata-Rueda, A.; Martínez, L.C.; Zanucio, J.C. Side-effects of pesticides on non-target insects in agriculture: A mini-review. Sci. Nat. 2022, 109, 17. [Google Scholar] [CrossRef] [PubMed]
- Michaud, J.P.; Grant, A.K. Sub-lethal effects of a copper sulfate fungicide on development and reproduction in three coccinellid species. J. Insect Sci. 2003, 3, 1–6. [Google Scholar]
- Sabo, R.; Kopčáková, A.; Hamarová, Ľ.; Cingeľová Maruščáková, I.; Mudroňová, D.; Sabová, L.; Javorský, P.; Legáth, J. Sublethal effects of commercial plant protection product containing spores Bacillus amyloliquefaciens QST 713 (formerly subtilis) on winter adult honeybees. Apidologie 2020, 51, 226–239. [Google Scholar] [CrossRef]
- Pinheiro, L.A.; Dáder, B.; Wanumen, A.C.; Pereira, J.A.; Santos, S.A.P.; Medina, P. Side effects of pesticides on the olive fruit fly parasitoid Psyttalia concolor (Szépligeti): A review. Agronomy 2020, 10, 1755. [Google Scholar] [CrossRef]
- González-Núñez, M.; Pascual, S.; Cobo, A.; Seris, E.; Cobos, G.; Fernández, C.E.; Sánchez-Ramos, I. Copper and kaolin sprays as tools for controlling the olive fruit fly. Entomol. Gen. 2021, 41, 97–110. [Google Scholar] [CrossRef]
- Orrù, B.; Moraglio, S.T.; Tortorici, F.; Gonella, E.; Tavella, L.; Alma, A. No adverse effects of symbiotic control on the parasitism of Halyomorpha halys by egg parasitoids. J. Pest Sci. 2022, 1–12. [Google Scholar] [CrossRef]
- Cáceres, C.; Tsiamis, G.; Yuval, B.; Jurkevitch, E.; Bourtzis, K. Joint FAO/IAEA coordinated research project on “Use of symbiotic bacteria to reduce mass-rearing costs and increase mating success in selected fruit pests in support of SIT application”. BMC Microbiol. 2019, 19, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadapad, A.B.; Shettigar, S.K.G.; Hire, R.S. Bacterial communities in the gut of wild and mass-reared Zeugodacus cucurbitae and Bactrocera dorsalis revealed by metagenomic sequencing. BMC Microbiol. 2019, 19 (Suppl. 1), 282. [Google Scholar] [CrossRef] [PubMed]
- Majumder, R.; Sutcliffe, B.; Adnan, S.M.; Mainali, B.; Dominiak, B.C.; Taylor, P.W.; Chapman, T.A. Artificial larval diet mediates the microbiome of Queensland fruit fly. Front. Microbiol. 2020, 11, 576156. [Google Scholar] [CrossRef] [PubMed]
- Bel Mokhtar, N.; Catalá-Oltra, M.; Stathopoulou, P.; Asimakis, E.; Remmal, I.; Remmas, N.; Maurady, A.; Britel, M.R.; García de Oteyza, J.; Tsiamis, G.; et al. Dynamics of the gut bacteriome during a laboratory adaptation process of the Mediterranean fruit fly, Ceratitis capitata. Front. Microbiol. 2022, 13, 919760. [Google Scholar] [CrossRef]
- Asimakis, E.D.; Khan, M.; Stathopoulou, P.; Cáceres, C.; Bourtzis, K.; Tsiamis, G. The effect of diet and radiation on the bacterial symbiome of the melon fly, Zeugodacus cucurbitae (Coquillett). BMC Biotechnol. 2019, 19 (Suppl. 2), 88. [Google Scholar] [CrossRef] [Green Version]
- Woruba, D.N.; Morrow, J.L.; Reynolds, O.L.; Chapman, T.A.; Collins, D.P.; Riegler, M. Diet and irradiation effects on the bacterial community composition and structure in the gut of domesticated teneral and mature Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). BMC Microbiol. 2019, 19 (Suppl. 1), 281. [Google Scholar] [CrossRef] [Green Version]
- Roque-Romero, L.; Guillén-Navarro, K.; Zarza, E.; Montoya, P.; Liedo, P. Bacterial diversity associated with Anastrepha obliqua males change under mass-rearing conditions and with irradiation. Curr. Microbiol. 2023, 80, 26. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Khan, M.A.M.; Collins, D.; Osborne, T.; Reynolds, O.L. Wild bacterial probiotics fed to larvae of mass-reared Queensland fruit fly [Bactrocera tryoni (Froggatt)] do not impact long-term survival, mate selection, or locomotor activity. Insect Sci. 2020, 27, 745–755. [Google Scholar] [CrossRef]
- Koskinioti, P.; Ras, E.; Augustinos, A.A.; Beukeboom, L.W.; Mathiopoulos, K.D.; Cáceres, C.; Bourtzis, K. Manipulation of insect gut microbiota towards the improvement of Bactrocera oleae artificial rearing. Entomol. Exp. Appl. 2020, 168, 523–540. [Google Scholar] [CrossRef]
- Msaad Guerfali, M.; Charaabi, K.; Hamden, H.; Djobbi, W.; Fadhl, S.; Mosbah, A.; Cherif, A. Probiotic based-diet effect on the immune response and induced stress in irradiated mass reared Ceratitis capitata males (Diptera: Tephritidae) destined for the release in the sterile insect technique programs. PLoS ONE 2021, 16, e0257097. [Google Scholar] [CrossRef]
- Zhang, Q.; Cai, P.; Wang, B.; Liu, X.; Lin, J.; Hua, R.; Zhang, H.; Yi, C.; Song, X.; Ji, Q.; et al. Manipulation of gut symbionts for improving the sterile insect technique: Quality parameters of Bactrocera dorsalis (Diptera: Tephritidae) genetic sexing strain males after feeding on bacteria-enriched diets. J. Econ. Entomol. 2021, 114, 560–570. [Google Scholar] [CrossRef]
- Sacchetti, P.; Ghiardi, B.; Granchietti, A.; Stefanini, F.M.; Belcari, A. Development of probiotic diets for the olive fly: Evaluation of their effects on fly longevity and fecundity. Ann. Appl. Biol. 2014, 164, 138–150. [Google Scholar] [CrossRef]
- Hamden, H.; Guerfali, M.M.; Fadhl, S.; Saidi, M.; Chevrier, C. Fitness improvement of mass-reared sterile males Ceratitis capitata (Vienna 8 strain) (Diptera: Tephritidae) after gut enrichment with probiotics. J. Econ. Entomol. 2013, 106, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Augustinos, A.A.; Kyritis, G.A.; Papadopoulos, N.T.; Abd-Alla, A.M.M.; Cáceres, C.; Bourtzis, K. Exploitation of the medfly gut microbiota for the enhancement of sterile insect technique: Use of Enterobacter sp. in larval diet-based probiotic applications. PLoS ONE 2015, 10, e0136459. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Yao, Z.; Li, Y.; Xi, Z.; Bourtzis, K.; Zhao, Z.; Bai, S.; Zhang, H. Intestinal probiotics restore the ecological fitness decline of Bactrocera dorsalis by irradiation. Evol. Appl. 2018, 11, 1946–1963. [Google Scholar] [CrossRef] [PubMed]
- Kashkouli, M.; Fathipour, Y.; Mehrabadi, M. Potential management tactics for pistachio stink bugs, Brachynema germari, Acrosternum heegeri and Acrosternum arabicum (Hemiptera: Pentatomidae): High temperature and chemical surface sterilants leading to symbiont suppression. J. Econom. Entomol. 2019, 112, 244–254. [Google Scholar] [CrossRef]
- Cariveau, D.; Powell, J.E.; Koch, H.; Winfree, R.; Moran, N.A. Variation in gut microbial communities and its association with pathogen infection in wild bumble bees (Bombus). ISME J. 2014, 8, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.W.; Rodrigues, P.A.P.; Oliver, R.; Mott, B.M.; Anderson, K.E. Diet-related gut bacterial dysbiosis correlates with impaired development, increased mortality and Nosema disease in the honeybee (Apis mellifera). Mol. Ecol. 2016, 25, 5439–5450. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zheng, L.; Tan, M.; Li, Y.; Xu, J.; Yan, S.; Jiang, D. Cd exposure-triggered susceptibility to Bacillus thuringiensis in Lymantria dispar involves in gut microbiota dysbiosis and hemolymph metabolic disorder. Ecotoxicol. Environ. Saf. 2022, 241, 113763. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-induced pathogen susceptibility is mitigated by Lactobacillus plantarum immune stimulation in a Drosophila melanogaster model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef]
- Li, M.; Liu, T.; Yang, T.; Zhu, J.; Zhou, Y.; Wang, M.; Wang, Q. Gut microbiota dysbiosis involves in host non-alcoholic fatty liver disease upon pyrethroid pesticide exposure. Environ. Sci. Ecotechnol. 2022, 11, 100185. [Google Scholar] [CrossRef]
- Estes, A.M.; Nestel, D.; Belcari, A.; Jessup, A.; Rempoulakis, P.; Economopoulos, A.P. A basis for the renewal of sterile insect technique for the olive fly, Bactrocera oleae (Rossi). J. Appl. Entomol. 2012, 136, 1–16. [Google Scholar] [CrossRef]
- Suckling, D.M.; Cristofaro, M.; Roselli, G.; Levy, M.C.; Cemmi, A.; Mazzoni, V.; Stringer, L.D.; Zeni, V.; Ioriatti, C.; Anfora, G. The competitive mating of irradiated brown marmorated stink bugs, Halyomorpha halys, for the sterile insect technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [Green Version]
- Barud, F.J.; Gómez, M.P.; Ruiz, M.J.; Bachmann, G.E.; Goane, L.; Segura, D.F.; Lara, N.; Murúa, F.; Asfennato, A.; Gómez, E.; et al. Sexual competitiveness of sterile Ceratitis capitata males exposed to essential oils from non-host plant species native to Argentina. Entomol. Exp. Appl. 2022, 171, 146–155. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; Khan, M.M.; Bamisile, B.S.; Hafeez, M.; Qasim, M.; Rasheed, M.T.; Rasheed, M.A.; Ahmad, S.; Shahid, M.I.; Xu, Y. Role of insect gut microbiota in pesticide degradation: A review. Front. Microbiol. 2022, 13, 870462. [Google Scholar] [CrossRef]
- Blanton, A.G.; Peterson, B.F. Symbiont-mediated insecticide detoxification as an emerging problem in insect pests. Front. Microbiol. 2020, 11, 547108. [Google Scholar] [CrossRef] [PubMed]
- Tamm, L.; Thuerig, B.; Apostolov, S.; Blogg, H.; Borgo, E.; Corneo, P.E.; Fittje, S.; de Palma, M.; Donko, A.; Experton, C.; et al. Use of copper-based fungicides in organic agriculture in twelve European countries. Agronomy 2022, 12, 673. [Google Scholar] [CrossRef]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2022, 1–15. [Google Scholar] [CrossRef]
- Sundh, I.; Del Giudice, T.; Cembalo, L. Reaping the benefits of microorganisms in cropping systems: Is the regulatory policy adequate? Microorganisms 2021, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
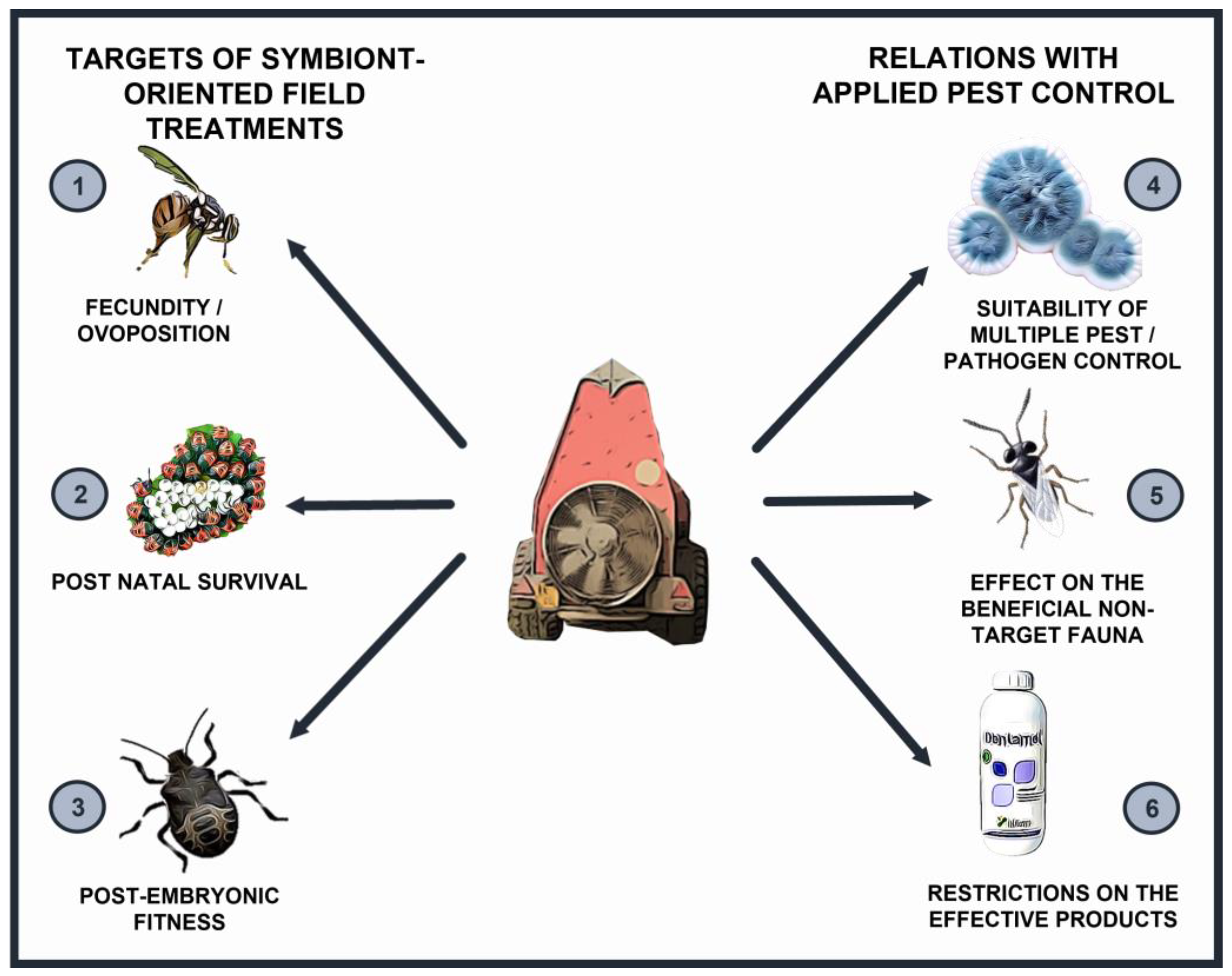
Figure 1.
Main operational components of symbiotic control applications in integrated pest management. The targets of symbiont manipulation include: (1) Altering the oviposition potential or affect the female fecundity; (2) Hampering the survival of the newly-hatched generation; (3) Affecting the juvenile fitness at later stages. To incorporate symbiont-targeted control in pest management programs, its compatibility with other strategies must be assessed: the use of antimicrobials may result in multiple activity against pests and pathogens (4). In turn, an anti-symbiont product must avoid detrimental interaction with the resident non-target fauna (5). Finally, symbiont-oriented approaches may require the extension of treatment numbers and periods, with environmental and economic costs, and the shift of target may be not included in the label directions, resulting in meaningful regulatory constraints (6).
Figure 1.
Main operational components of symbiotic control applications in integrated pest management. The targets of symbiont manipulation include: (1) Altering the oviposition potential or affect the female fecundity; (2) Hampering the survival of the newly-hatched generation; (3) Affecting the juvenile fitness at later stages. To incorporate symbiont-targeted control in pest management programs, its compatibility with other strategies must be assessed: the use of antimicrobials may result in multiple activity against pests and pathogens (4). In turn, an anti-symbiont product must avoid detrimental interaction with the resident non-target fauna (5). Finally, symbiont-oriented approaches may require the extension of treatment numbers and periods, with environmental and economic costs, and the shift of target may be not included in the label directions, resulting in meaningful regulatory constraints (6).

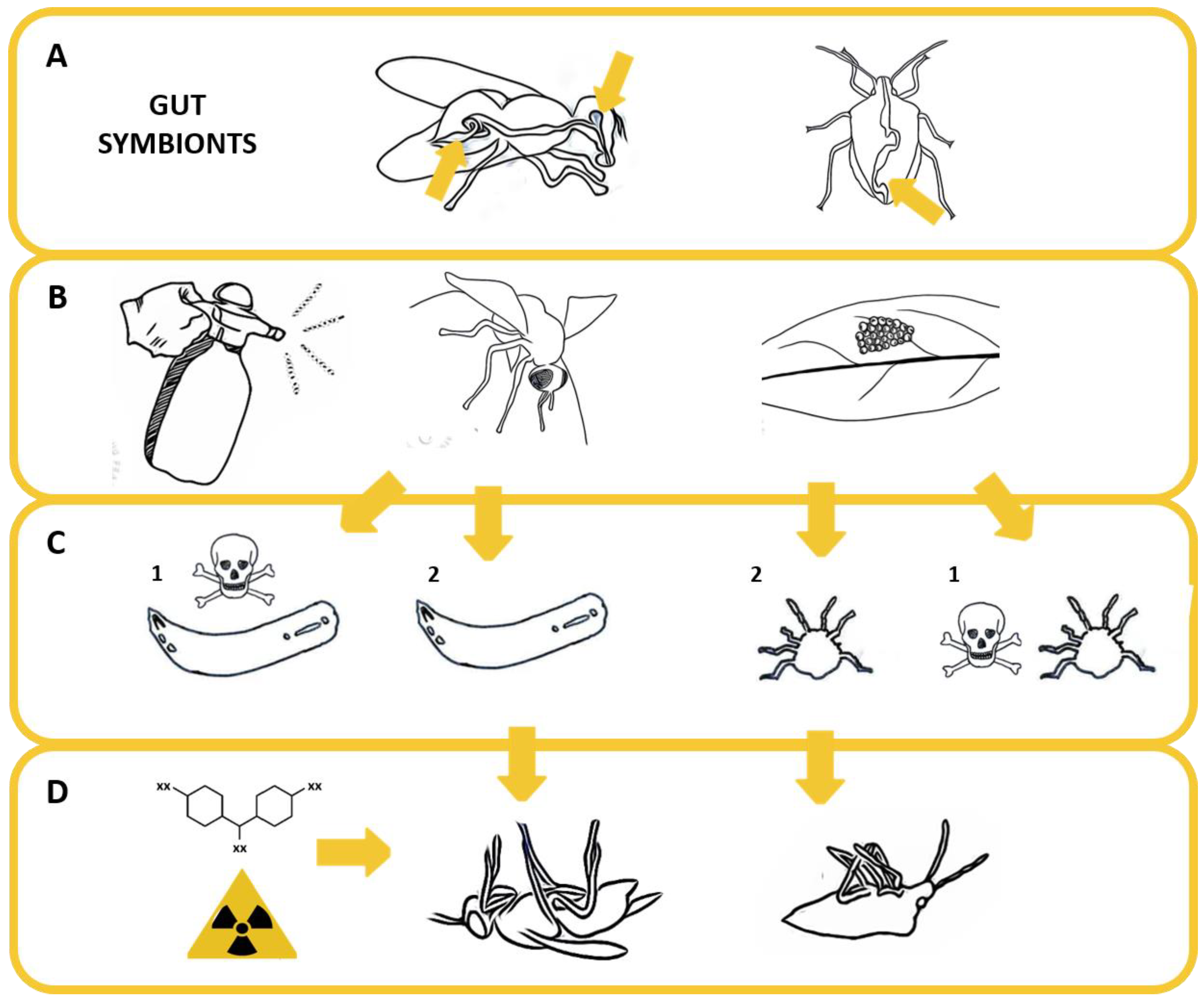
Figure 2.
Importance of gut symbiotic bacteria of Pentatomidae and Tephritidae and effect of symbiotic perturbation. (A) The obligate gut symbionts of tephritid flies are located in the gut lumen, in the oesophageal bulb in the insect head or in rectal crypts. In pentatomid stink bugs, obligate symbionts are hosted in crypts located in the terminal part of midgut. (B) Experimental exposure to bactericidal agents may affect the vertical transmission of obligate symbionts (and/or other symbiotic bacteria) during or after oviposition, resulting in (C) an immediate lethal effect on newborns (1), or defective survival (2). (D) In turn, harm related to the induction of dysbiosis after the interruption of symbiont vertical transmission may synergize the effect of other stressors, inducing long-term mortality.
Figure 2.
Importance of gut symbiotic bacteria of Pentatomidae and Tephritidae and effect of symbiotic perturbation. (A) The obligate gut symbionts of tephritid flies are located in the gut lumen, in the oesophageal bulb in the insect head or in rectal crypts. In pentatomid stink bugs, obligate symbionts are hosted in crypts located in the terminal part of midgut. (B) Experimental exposure to bactericidal agents may affect the vertical transmission of obligate symbionts (and/or other symbiotic bacteria) during or after oviposition, resulting in (C) an immediate lethal effect on newborns (1), or defective survival (2). (D) In turn, harm related to the induction of dysbiosis after the interruption of symbiont vertical transmission may synergize the effect of other stressors, inducing long-term mortality.


{kind=link}
{kind=link}
Table 1.
Extracellularly-inherited symbiotic bacteria in the insect families Pentatomidae and Tephritidae.
Table 1.
Extracellularly-inherited symbiotic bacteria in the insect families Pentatomidae and Tephritidae.
| Host Species | Host Family | Symbiont Species | Obligate/Facultative Association | Type of Egg Smearing Transmission * | Reference |
|---|---|---|---|---|---|
| Acanthiophilus helianthi. | Tephritidae | ‘Ca. Stammerula sp.’ | Obligate | Ovopositional smearing | [56] |
| Acrosternum spp. | Pentatomidae | Pantoea sp. | Obligate | Ovopositional smearing | [4] |
| Anastrepha fraterculus | Tephritidae | N.D. | N.D. | Ovopositional smearing | [57,58] |
| Bactrocera dorsalis | Tephritidae | Citrobacter sp. | Facultative | Ovopositional smearing | [59] |
| Bactrocera oleae | Tephritidae | ‘Ca. Erwinia dacicola’ | Obligate | Ovopositional smearing | [60] |
| Brachynema germari | Pentatomidae | Pantoea sp. | Obligate | Ovopositional smearing | [61] |
| Campiglossa spp. | Tephritidae | ‘Ca. Stammerula sp.’ | Obligate | Ovopositional smearing | [62] |
| Capitites ramulosa | Tephritidae | ‘Ca. Stammerula sp.’ | Obligate | Ovopositional smearing | [56] |
| Ceratitis capitata | Tephritidae | Enterobacter (Pantoea) agglomerans and Klebsiella pneumoniae | Facultative | Pre-ovopositional smearing | [63] |
| Dioxyna bidentis | Tephritidae | N.D. | Obligate | Ovopositional smearing | [56] |
| Edessa spp. | Pentatomidae | ‘Ca. Pantoea edessiphila’ | Obligate | Ovoposi-tional smearing | [64] |
| Eurydema spp. | Pentatomidae | N.D. | Obligate | Ovopositional smearing | [65] |
| Graphosoma lineatum | Pentatomidae | Pantoea sp. | Obligate | Ovopositional smearing | [66] |
| Halyomorpha halys | Pentatomi-dae | ‘Ca. Pantoea carbekii’ | Obligate | Ovopositional smearing | [51] |
| Murgantia histrionica | Pentatomidae | N.D. | Facultative | Ovopositional smearing | [67] |
| Nezara viridula | Pentatomidae | Pantoea sp. | Facultative | Ovopositional smearing | [68] |
| Nezara viridula | Pentatomidae | Sodalis sp. | Facultative | Ovopositional smearing | [68] |
| Noeeta spp. | Tephritidae | N.D. | Obligate | Ovopositional smearing | [56] |
| Oxyna flavipennis | Tephritidae | ‘Ca. Stammerula sp.’ | Obligate | Ovopositional smearing | [56] |
| Plautia stali | Pentatomidae | Pantoea spp. | Obligate | Ovopositional smearing | [69] |
| Sibaria englemani | Pentatomidae | Pantoea sp. | N.D. | Ovopositional smearing | [70] |
| Sphenella marginata | Tephritidae | ‘Ca. Stammerula sp.’ | Obligate | Ovopositional smearing | [56] |
| Tephritis spp. | Tephritidae | ‘Ca. Stammerula tephritidis’ | Obligate | Ovopositional smearing | [62] |
| Trupanea spp. | Tephritidae | ‘Ca. Stammerula trupaneae’ | Obligate | Ovopositional smearing | [56] |
* Ovopositional smearing: the mother covers eggs with symbiont-containing secretes during ovoposition; pre-ovopositional smearing: symbiont cells reach ovaric eggs and cover the egg chorion before egg laying.
Table 2.
List of active substances or microbial pest control products that have been shown to contain insect populations through symbiont suppression.
Table 2.
List of active substances or microbial pest control products that have been shown to contain insect populations through symbiont suppression.
| Active Substances or Microbial Biocontrol Agent | Trade Name and Manufacturer | Main Product Use | Target Species for Symbiotic Control (Insect, Symbiont) | Laboratory/ Field Validation | Symbiont Elimination Demonstrated (D)/Suggested (S) | References |
|---|---|---|---|---|---|---|
| Mixture (surfactant) | Naiad (Naiad Company, Inc.) | Wetting agent | H. halys, ‘Ca. Pantoea carbekii’ | Laboratory (field validation failed in containing insects) | D | [72] |
| Copper hydroxide | Keos® (Green Ravenna Srl) | Biostimulant | H. halys, ‘Ca. Pantoea carbekii’ | Laboratory | S | [13] |
| Manganese and copper biocomplex in water solution | Bio-D® (Diachem S.p.A.) | Biostimulant | H. halys, ‘Ca. Pantoea carbekii’ | Laboratory | D | [13] |
| Propolis | Propoli® (Serbios s.r.l.) | Biostimulant | B. oleae, ‘Ca. Erwinia dacicola’ | Field | D | [12,22] |
| Tannins | Distillato di Legno (BioDea) | Biostimulant | B. oleae, ‘Ca. Erwinia dacicola’ | Field | S | [22] |
| Zinc and copper biocomplex in water solution | Dentamet® (Diachem S.p.A.) | Biostimulant | H. halys, ‘Ca. Pantoea carbekii’; B. oleae, ‘Ca. Erwinia dacicola’ | Laboratory (H. halys), field (B. oleae) | D (H. halys); S (B. oleae) | [13,22] |
| Copper hydroxide + kaolin | Kaolin (SEPIDAN, WP®), Copper oxychloride (Oxifam®) | Fungicide (copper oxychloride), biostimulant (kaolin) | B. oleae, ‘Ca. Erwinia dacicola’ | Field | S | [77] |
| Copper hydroxide | Gafex® (Bayer CropScience); Copper idro 40 WDG® (SARIAF SA) | Fungicide | B. oleae, ‘Ca. Erwinia dacicola’ | Field | S | [78,79] |
| Copper hydroxide, calcium hydroxide | Manisol (Manica S.p.A,) | Fungicide | B. oleae, ‘Ca. Erwinia dacicola’ | Field | D | [12] |
| Copper oxychloride | Cupravit Blu 35 WG® (Bayer Cropscience S.r.l.); Neoram® (Isagro S.p.A.) | Fungicide | B. oleae, ‘Ca. Erwinia dacicola’ | Field | S | [22,80] |
| Copper oxychloride | N.A. | - | B. oleae, ‘Ca. Erwinia dacicola’ | Laboratory | D | [15] |
| Copper sulfate (Bordeaux mixture) | Poltiglia Caffaro 20 DF NEW (Isagro and Sumitomo Chemicals) | Fungicide | B. oleae, ‘Ca. Erwinia dacicola’ | Field | D | [12] |
| Dodine | Syllit® 544 SC (ARYSTA LifeScience Italia S.r.l.) | Fungicide | B. oleae, ‘Ca. Erwinia dacicola’ | Field | S | [22] |
| Bacillus velezensis (formerly amyloliquefaciens) D747 | Amylo-X (CBC (Europe) S.r.l.) | Fungicide, bactericide | H. halys, ‘Ca. Pantoea carbekii’ | Laboratory | D | [14] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gonella, E.; Alma, A. The Role of Symbiont-Targeted Strategies in the Management of Pentatomidae and Tephritidae Pests under an Integrated Vision. Agronomy 2023, 13, 868. https://doi.org/10.3390/agronomy13030868
AMA Style
Gonella E, Alma A. The Role of Symbiont-Targeted Strategies in the Management of Pentatomidae and Tephritidae Pests under an Integrated Vision. Agronomy. 2023; 13(3):868. https://doi.org/10.3390/agronomy13030868
Chicago/Turabian StyleGonella, Elena, and Alberto Alma. 2023. "The Role of Symbiont-Targeted Strategies in the Management of Pentatomidae and Tephritidae Pests under an Integrated Vision" Agronomy 13, no. 3: 868. https://doi.org/10.3390/agronomy13030868
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.