
LIN28B and Let-7 in Diffuse Midline Glioma: A Review
by
 , , ,
, , ,
Truman Knowles
1 ,
,
Tina Huang
2,
Jin Qi
2,
Shejuan An
2,
Noah Burket
3,
Scott Cooper
3,
Javad Nazarian
4,5 and
Amanda M. Saratsis
6,* 1
W.M. Keck Science Department, Scripps, Pitzer, and Claremont McKenna Colleges, Claremont, CA 91711, USA
2
Department of Neurosurgery, Northwestern University Feinberg School of Medicine, Chicago, IL 60611, USA
3
Department of Neurosurgery, Indiana University School of Medicine, Indianapolis, IN 46202, USA
4
Department of Pediatrics, Children’s National Hospital, Washington, DC 20010, USA
5
Department of Pediatrics, Zurich Children’s Hospital, 8032 Zurich, Switzerland
6
Department of Neurosurgery, Lutheran General Hospital, Park Ridge, IL 60068, USA
*
Author to whom correspondence should be addressed.
Cancers 2023, 15(12), 3241; https://doi.org/10.3390/cancers15123241
Submission received: 18 April 2023
/
Revised: 12 June 2023
/
Accepted: 17 June 2023
/
Published: 19 June 2023
(This article belongs to the Section Molecular Cancer Biology)
Abstract
:Simple Summary
Diffuse midline glioma (DMG) is a devastating pediatric brain tumor with urgent unmet need for novel treatment modalities. LIN28B RNA binding protein is overexpressed in DMG and suppresses the let-7 family of microRNAs, which in turn suppress a plethora of oncogenes. In the present review, we summarize this LIN28B–let-7–oncogene axis across glioma subtypes and advise future research specific to DMG, offering it as a potential therapeutic vulnerability.
Abstract
Diffuse midline glioma (DMG) is the most lethal of all childhood cancers. DMGs are driven by histone-tail-mutation-mediated epigenetic dysregulation and partner mutations in genes controlling proliferation and migration. One result of this epigenetic and genetic landscape is the overexpression of LIN28B RNA binding protein. In other systems, LIN28B has been shown to prevent let-7 microRNA biogenesis; however, let-7, when available, faithfully suppresses tumorigenic pathways and induces cellular maturation by preventing the translation of numerous oncogenes. Here, we review the current literature on LIN28A/B and the let-7 family and describe their role in gliomagenesis. Future research is then recommended, with a focus on the mechanisms of LIN28B overexpression and localization in DMG.
1. Introduction
Diffuse midline glioma (DMG) is a devastating childhood brain tumor with a median age at diagnosis of 6–7 years [1]. Between 200–300 cases are reported annually in the United States, with a two-year survival rate of less than 10 percent and a median survival of 10 months [2,3,4].
Approximately 80% of DMGs have a mutation in HIST1H3B or H3F3A, which convert position 27 on histone H3.1- or H3.3- from lysine to methionine, respectively (H3K27M mutations) [5,6,7,8]. H3 wild-type (H3WT) DMGs have enhancer of zeste homologs inhibitory protein (EZHIP) overexpression [9]. Both the H3K27M mutations and EZHIP overexpression decrease H3K27 trimethylation (H3K27me3), resulting in dysregulated gene expression, including the upregulation of endogenous retroviruses and LIN28B, an RNA binding protein (RBP) [10,11,12,13,14,15,16,17,18]. LIN28B and its paralog, LIN28A, interact with the let-7 family of microRNAs (miRs)—which includes let-7a/b/c/d/e/f/g/i/miR-98—during development and oncogenesis [19,20,21].
The biogenesis of miRs occur in the following three steps (Figure 1) [22]. First, a gene for a given let-7 family member is transcribed into a primary-let-7 (pri-let-7) transcript; this occurs in the nucleolus, a dense region of the nucleus [20]. Second, pri-let-7 is translocated to the larger, less dense region of the nucleus and processed by the microprocessor complex—which contains two proteins, DROSHA and DGCR8—into a preliminary-let-7 (pre-let-7) form [20]. Third, pre-let-7 is translocated to the cytoplasm and processed by another protein, DICER, into its mature and bioactive form, known simply as let-7 [20].
The biogenesis of the let-7 miR family is suppressed by LIN28A/B. LIN28A works with Musashi1 (MSI1) to sequester pri-let-7 in the nucleus [23]. LIN28A also binds pre-let-7 in the cytoplasm to block processing by DICER, as well as recruits a protein called terminal uridylyltransferase 4/7 (TUT4/7), which adds uridyl groups to the 3′ end of pre-let-7, an action that tags it for degradation [24,25,26,27,28]. In contrast, LIN28B sequesters pri-let-7 transcripts in the nucleolus, preventing them from continuing on in the biogenesis pathway [29].
After their biogenesis, mature miRs direct translational repression of their target gene mRNAs. This is mediated by the RNA-induced silencing complex (RISC). MiRs are first incorporated into RISC, then this miR-loaded RISC uses the miR nucleic acid sequence as a guide, with the functional unit binding to the 3′ untranslated region (UTR) of mRNAs through base pair complementarity [30]. This binding prevents the mRNAs from being translated by the ribosome.
Each of the let-7 family members decrease expression of genes that promote stemness, proliferation, and migration, thereby promoting differentiation programming, while LIN28A/B derepress these genes in a let-7-dependent manner to preserve a pluripotent phenotype [19,20,21]. Both LIN28A/B have also been shown to directly bind certain RNA transcripts, increasing their stability and according function or expression [19,20,21,31,32]. This provides a let-7-independent mechanism of oncogene upregulation. Initially discovered in Caenorhabditis elegans, LIN28A/B and the let-7 family are responsible for the timing of differentiation, with aberrancies in the elegant relative level of each species causing heterochronic developmental defects [33,34,35,36]. Indeed, high LIN28A/B levels and low let-7 levels is appropriate in early developmental contexts, but aberrantly present in many cancers, while the reverse proportion can cause neurodegeneration [37,38]. Interestingly, progressive LIN28A/B downregulation and let-7 family upregulation has similarly been reported in differentiation of oligodendrocyte precursor cells, the cell of origin in DMG, suggesting a conceptual model wherein DMG formation is caused by an arrest of this progression, potentially resulting from the H3K27M mutant epigenetic landscape (Figure 2, [39,40,41,42,43,44]).
Of note, dysregulated expression and function of LIN28A/B and the let-7 family has been characterized in other pediatric non-glioma brain tumors, including embryonal tumor with multilayered rosettes, atypical teratoid rhabdoid tumor, and medulloblastoma [45,46,47,48,49,50,51,52]. For matters of space, however, we focus exclusively on LIN28A/B and the let-7 family in gliomagenesis, specifically DMG.
Our review manuscript makes three novel contributions to the field of DMG. First, it introduces this LIN28B–let-7–oncogene axis, a promising therapeutic vulnerability. Second, it extensively reviews let-dependent and -independent mechanisms of LIN28A/B function across glioma subtypes. Third, it directs future work specific to DMG.
2. LIN28A/B and Let-7 Expression Correlate with Clinical Outcomes in Glioma
One consistent finding in the literature pertained to the expression pattern of LIN28A/B and let-7 family members. In glioma, LIN28A expression is increased, correlating directly with tumor grade and poor survival (Table 1, [32,53,54]). LIN28A expression is also increased in glioma cell lines [32]. Our group and others have shown that LIN28B is expressed in cell lines of diffuse intrinsic pontine glioma (DIPG), the pontine subtype of DMGs (unpublished data, [15,17]). Indeed, RNA-sequencing data shows that LIN28B is overexpressed in H3K27M mutant DMG when compared to H3WT DMG [16].
Of interest is a unique paper by Guo et al. [74] which characterized an LIN28B tumor-specific transcript (LIN28B-TST). The authors showed that LIN28B-TST is encoded upstream of the LIN28B-WT transcript in both low-grade glioma (LGG) and glioblastoma (GBM), providing several more codons to result in additional N-terminal amino acids in the LIN28B-TST protein [74]. An LIN28B-TST was not reported in DMG [74]. A c-Myc-regulated transcription initiation site controls the LIN28B-WT transcript [74]. N-MYC gain-of-function mutation as well as C-MYC overexpression are observed in DMG, and C-MYC downregulation may be seen in response to treatment [75,76]. Such data suggest LIN28B may mediate a c-Myc-driven subset of DMG, as has been characterized in a panel of c-Myc-driven cancer cell lines and in n-Myc-driven neuroblastoma [77,78]. About 20 kilobases (kb) upstream of the LIN28B-WT locus, a nuclear transcription factor Y subunit alpha (NFYA)-regulated alternative transcription initiation (ATI) site controls LIN28B-TST expression [74]. Similar to C-MYC, NFYA is overexpressed in DMG and may drive expression of such an LIN28B-TST [79]. Functionally, additional N-terminal amino acids of the LIN28B-TST protein increase its stability, leading to greater protein product [74]. LIN28B-TST enhances tumorigenicity in an in vivo liver hepatocellular carcinoma (HCC) cell murine model and HCC patients expressing LIN28B-TST have poorer outcomes [74]. In sum, NYFA and c-Myc regulate the transcription of alternative LIN28B transcripts in glioma (Figure 3).
Fitting with this paradigm of LIN28A/B overexpression in glioma, let-7a/b/d/f/g/miR-98 are decreased and correlate inversely with tumor grade (Table 1, [55,56,57,62,68,70,71,72,73]). Correspondingly, let-7a/b/e/f/g and miR-98 are increased in long-term-surviving patients, and let-7f correlates directly with survival [58,68]. Likewise, let-7a/b/d/f/i/miR-98 are decreased in glioma cell lines [57,63,68,71,72,73]. Moreover, in a glioma murine model, let-7a/c/d/e/g/miR-98 are decreased [56].
In a DIPG murine model driven by the expression of H3.3K27M mutant histones, LIN28B expression is increased, as is the expression of insulin-like growth factor mRNA binding protein 2 (IGF2BP2, also known as IMP2) [13]. Interestingly, however, a study investigating glioma stem cells (GSCs) did not detect either LIN28A/B and, in turn, found that let-7 family members were highly expressed [80]. Yet, let-7 targets were still upregulated, suggesting an LIN28A/B-independent mechanism was suppressing let-7 activity [80]. The authors showed IMP2 interacts with Argonaute protein 2 (AGO2), the catalytic component of RISC, at let-7 recognition elements on target mRNAs [80]. This prevented let-7-mediated mRNA translational repression to preserve the GSC phenotype, showing that IMP2 activity may compensate for low LIN28A/B expression (Figure 3, [80]).
3. LIN28A/B-Dependent Mechanisms Regulating Gliomagenesis
3.1. LIN28A Drives Aerobic Glycolysis
LIN28A is an established regulator of cancer metabolism: LIN28A binds to the mRNAs of several glycolytic and insulin signaling genes to increase their translation, thereby increasing glycolysis and glucose uptake, as well as derepresses a let-7 target that inhibits pyruvate entry into the tricarboxylic acid cycle, thereby reenforcing aerobic glycolysis [81]. Fitting with these data, one study of glioma cells showed that LIN28A stabilized the pre-long noncoding RNA (lncRNA) small nucleolar host gene 14 (SNHG14) transcript [32]. Then, SNHG14 recruits staufen double-stranded RNA binding protein 1 (STAU1) to the interferon regulatory factor 6 (IRF6) mRNA 3′ UTR, in turn recruiting UPF1, an RNA helicase [32]. UPF1 mediates IRF6 mRNA degradation, decreasing IRF6 expression [32]. This derepresses IRF6-silenced genes, pyruvate kinase M2 (PKM2) and glucose transporter 1 (GLUT1) [32]. PKM2 and GLUT1 increase aerobic glycolysis, which in turn increases tumor microenvironment (TME) acidification [32]. Consistent with this paradigm, the authors show that decreasing LIN28A or SNHG14, or increasing IRF6, decreases aerobic glycolysis and proliferation [32]. They validate these results using murine subcutaneous and orthotopic xenograft models [32]. Altogether, these data suggest LIN28A changes the metabolic profile of glioma cells (Figure 4).
3.2. LIN28A Inhibits Apoptosis and Drives Proliferation and Migration
Because it derepresses a plethora of oncogenes through let-7 suppression, LIN28A induces a stem-like phenotype characterized by resistance of apoptosis and increased proliferation and migration. Downregulation of LIN28A also increases apoptosis and G1 phase cell count while decreasing S phase cell count and colony formation of glioma cells [54]. This may be related to the findings of one study showing that LIN28A expression correlates directly with stem cell factor expression [53]. These factors were octamer-binding transcription factor (TF) 4 (OCT4), high-mobility AT-hook 2 (HMGA2), and snail family transcriptional repressor 1 (SNAI1) [53]. Another important takeaway from this work was that LIN28A inhibition decreases invasion and growth of glioma cells [53]. Moreover, transduction of LIN28A into these cells decreases let-7b and let-7g expression [53]. Correspondingly, LIN28A transduction into human neural stem cells carrying dominant negative TP53, constitutively active K-RAS, and constitutively active human telomerase reverse transcriptase showed increased growth [53]. The authors validate these results using an orthotopic xenograft intracranial murine model, noting intraparenchymal invasion and a dependency on LIN28A [53]. Taken together with Section 3.1, there are important considerations to LIN28A as a factor sitting at the intersection of a multitude of oncogenic processes.
3.3. LIN28B Inhibits Apoptosis and Drives Proliferation
LIN28B mechanistic studies in DMG are lacking. However, some preliminary data are suggesting that it may resist apoptosis while also driving proliferation. Short hairpin RNA-mediated LIN28B knockdown has been shown to decrease proliferation and increase apoptosis of DIPG cells [15]. An additional study showed that LIN28B knockdown decreases proliferation of human embryonic stem cell-derived neural progenitor cells generated by constitutive activation of PDGFRɑ, TP53 knockdown, and H3.3K27M mutation [82]. Therefore, it is likely that similar to LIN28A, LIN28B increases proliferation and cell viability in glioma.
4. Mechanisms of Glioma Cell Suppression by Let-7
4.1. Let-7 Drives Apoptosis and Inhibits Proliferation and Migration
In line with the conceptual model proposed (Figure 2), the let-7 family performs opposite functions in glioma cells than LIN28A/B, and this is mediated by the translational repression of a plethora of oncogenes (Table 1). Given the intersecting data below, it appears clear that we cannot tease apart these functions independently of each other, likely due to the many targets of let-7 family members.
To begin, neurotensin (NTS) is a peptide neurotransmitter and neuromodulator of the CNS that has been reported as a mediator of diverse pathologies, including Parkinson’s disease, schizophrenia, and various cancers [83]. In glioma, expression of the high-affinity NTS G-protein-coupled receptor 1 (NTSR1) correlates directly with tumor grade and indirectly with survival [84]. Mechanistically, NTSR1 increases C-MYC expression, extracellular signal-regulated kinase 1/2 (ERK1/2) phosphorylation, as well as the proliferation and migration of glioma cells [84,85]. In turn, treatment with the NTSR1 inhibitor, SR48692, decreases growth of a murine glioma model [84,85]. Interestingly, glioma cells treated with SR48692 have increased let-7a-3 expression, as well as activity of the pro-apoptotic protein, caspase 3 [59]. Let-7a-3 binds Bcl-w mRNA 3′ UTR to decrease expression of this anti-apoptotic protein, thereby inducing apoptosis [59,60]. Moreover, SR48692-mediated NTSR1 inhibition decreases C-MYC expression, which decreases LIN28A expression [59]. These results were validated in a murine glioma model [59].
In another study, ultraviolet light-induced apoptosis of glioma cell lines is increased by miR-98 treatment through a mechanism dependent on inhibitor of nuclear kappa-B kinase epsilon (IKBKE, also known as IKKi or IKKε), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), and Bcl-2 (Figure 5, [72]). IKBKE is a protein that mediates several proliferative and migratory signaling pathways in various cancers [86]. IKBKE overexpression has been reported in glioma and NF-κB, a transcription factor well-established to increase a stem cell-phenotype, migration, radiation resistance, aerobic glycolysis, and angiogenesis in GBM [87,88]. NF-κB increases expression of the anti-apoptotic protein, Bcl-2 [87,89]. Of note, miR-98 binds IKBKE mRNA 3′ UTR to decrease IKBKE expression, which decreases NF-κB p50 subunit expression [72]. This leads to a decrease in NF-κB activity, causing a decrease in Bcl-2 expression, ultimately resulting in an increase in caspase 3 activity [72]. Similarly, miR-98 decreases migration of glioma cells by binding to IKBKE mRNA 3′ UTR to decrease IKBKE expression, decreasing NF-κB p65 subunit nuclear translocation, which decreases expression of matrix metalloproteinase (MMP)-9 [71]. MMP-9 expression correlates directly with tumor grade and indirectly with survival in glioma [90]. Mechanistically, MMPs are zinc-dependent endopeptidases that cleave extracellular matrix (ECM) components, thereby remodeling the TME to permit invasion [91]. MMP-9 also functions through a non-canonical mechanism to increase proliferation in glioma [90]. Finally, let-7b/i decreases migration and proliferation of glioma cells by binding to IKBKE mRNA 3′ UTR to decrease IKBKE expression, which increases E-cadherin (E-cad) expression [63]. E-cad is a cell–cell adhesion protein that restricts tumor cell migration and is commonly downregulated in glioma [92]. The let-7 family, therefore, acts through IKBKE to alter apoptotic and migratory signals (Figure 5).
The RAS family is also a target of let-7. RAS family members are membrane-bound guanosine triphosphate (GTP)-activated binary molecular switches that initiate signaling pathways in diverse pathologies, including psychiatric and developmental conditions, as well as various cancers [93]. In DMG, the H3K27M mutation increases RAS activity, driving the RAS pathway component ERK5 to stabilize c-Myc [94]. Treatment with let-7g decreases proliferation and migration of glioma cells by binding to pan-RAS mRNA 3′ UTR, N-RAS mRNA 3′ UTR, and K-RAS mRNA 3′ UTR to decrease pan-RAS, N-RAS, and K-RAS expression, while treatment with let-7a achieves similar results by targeting K-RAS [55,69]. Each of these results were validated in a murine glioma model [55,69]. Therefore, RAS is another target of let-7 in glioma and mediates decreases in proliferation and migration.
Emerging data are shining light on H19 lncRNA in glioma, which, similarly to LIN28A/B, works by suppressing let-7 to derepress let-7 targets. A recent study showed that in DIPG cells, H19 lncRNA expression is increased [61]. Meanwhile, let-7a-5 treatment decreases proliferation [61]. However, H19 decreases let-7a-5 levels via lncRNA:miR complementarity, which increases expression of let-7a-5 targets (Figure 6, [61]). It was shown that sulfatase 2 (SULF2) and oncostatin M receptor (OSMR) expression is increased [61]. SULF2 is expressed in GBM and drives PDGFRɑ activation [95]. OSMR is expressed in GBM, owing in part to hypoxia-induced annexin A2 (ANXA2) activation of STAT3, which drives its expression [96]. Of note, hypoxia is a feature of DMG [97]. OSMR then mediates signaling of the macrophage-associated cytokine oncostatin M to activate signal transducer and activator of transcription 3 (STAT3), leading to the upregulation of genes that increase migration, proliferation, and angiogenesis [96,98,99,100]. OSMR is translocalized to the mitochondrial matrix and interacts with the electron transport chain component NADH ubiquinone oxidoreductase 1/2 (NDUFS1/2) to upregulate oxidative phosphorylation and confer radiation resistance [101]. The same mechanism of H19-mediated let-7 antagonism has also been shown in human embryonic kidney 293 cells [102]. Therefore, more work is warranted to further characterize H19 in glioma and how it blunts let-7-mediated proliferative suppression.
Let-7 also acts through cell-cycle regulators to control proliferative signaling. Treatment with let-7b decreases proliferation of glioma cells by binding to E2F2 mRNA 3′ UTR to decrease E2F2 expression [64]. E2F2 expression correlates directly with tumor grade and indirectly with survival in glioma [103]. E2F2 is a TF that promotes cell-cycle progression [104]. Let-7b decreases cell migration while increasing apoptosis and S phase cell count by binding to cyclin A2 (CCNA2) mRNA 3′ UTR, cyclin B2 (CCNB2) mRNA 3′ UTR, polo like kinase 1 (PLK1) mRNA 3′ UTR, and aurora A kinase (AURKA) mRNA 3′ UTR to decrease CCNA2, CCNB2, PLK1, and AURKA expression [65]. Bioinformatics analyses have revealed that these are “hub” genes which coordinate cell-cycle pathways and can likely serve as biomarkers for GBM, bringing even greater importance to understanding how let-7 interacts with them [65,105,106,107].
HMGA2 is a major target of the LIN28A/B–let-7–oncogene axis, as it changes migratory programming by altering transcriptional dynamics. HMGA2 is a non-histone chromatin-associated protein that binds to the minor groove of DNA, bending it and, thereby, regulating its accessibility during a myriad of processes, most notably transcription [108]. HMGA2 expression correlates directly with tumor grade and indirectly with survival in glioma [109]. Raf-1 kinase inhibitor protein (RKIP) decreases invasion of glioma cells by increasing miR-98 expression, which binds HMGA2 mRNA 3′ UTR to decrease HMGA2 expression [70]. HMGA2 increases glioma cell migration in part by increasing MMP-2 [110,111]. Indeed, one study assessed glioma cell colonies’ migratory rim cells to migration-restricted core cells, showing that let-7a/b/c/d/e/f/g/i are decreased in the migratory population [112]. Together, these data show that HMGA2 is a vital target of let-7 in glioma, especially when taken with the LIN28A data discussed in Section 3.2.
Let-7 also targets pre-B-cell leukemia homeobox 3 (PBX3) in glioma, a protein which mediates transforming growth factor β (TGF-β) signaling, driving the expression of genes that increase migration—N-cadherin (N-cad), zinc finger E-box-binding homeobox 1 (ZEB1), SNAI2, and CD44 [66]. PBX3 activates the MEK/ERK1/2 pathway, which increases c-Myc-mediated LIN28A expression [66]. As such, a positive feedback loop reinforces the migration [66]. Both let-7b and miR-98 decrease migration of glioma cells by binding to PBX3 mRNA 3′ UTR to decrease PBX3 expression [66,73]. All results were validated in murine glioma models [66,73]. These data offer that PBX3 is another important factor to consider in the anti-migratory signaling of let-7.
Another study found that STAT3 mediated the apoptotic effects of a different subset of let-7 family members. STAT3 is the TF component of the canonical Janus kinase (JAK)/STAT signaling pathway, directing expression of genes involved in anti-apoptosis, migration, angiogenesis, and immune suppression in GBM [113]. However, in a PTEN-deficient genetic background—such as that reported in a subset of DMGs—STAT3 may actually be tumor suppressive [114]. Such conflicting data warrant more work to elucidate the context-specific roles of STAT3. Regardless, treatment with let-7a-1/d/f-1 decreases proliferation while increasing apoptosis and autophagy of glioma cells by binding to STAT3 mRNA 3′ UTR to decrease STAT3 expression [57]. In turn, this also causes a decrease in Bcl-2 expression, leading to an increase in c-caspase-3 activity [57]. STAT3 is, therefore, an emerging target of let-7 in glioma and, fitting with its broad biological functions, may be in part responsible for the broad glioma suppressive functions of let-7.
An additional mediator of let-7-driven caspase 3 upregulation is cyclin D1 (CCND1), which phosphorylates and thereby inactivates retinoblastoma (Rb) protein, a tumor suppressor that inhibits G1-S phase progression [115]. CCND1 expression correlates directly with tumor grade, increases proliferation, and contributes to temozolomide resistance [116]. Treatment with let-7b rescues cisplatin sensitivity of cisplatin-resistant glioma cells by binding to CCND1 mRNA 3′ UTR to decrease CCND1 expression, which increases caspase 3 activity, apoptosis, and G1 phase cell count [67]. A partner of cyclin D1 is cyclin E (encoded by CCNE1), which is also responsible for Rb phosphorylation, and thus, cell-cycle stimulation [117]. Treatment with let-7f decreases proliferation while increasing G1 phase cell count and apoptosis of glioma cells by decreasing CCND1, CCNE1, and Bcl-2 expression, increasing P21, P27, and Bax expression, and increasing caspase-3 activity [68]. p21 and p27 block cell-cycle progression by inhibiting the cyclin E:cyclin-dependent kinase 2 complex, while Bax promotes apoptosis by piercing the mitochondrial membrane to release the caspase-3 activator cytochrome-c [118,119,120]. Let-7f also decreases migration and invasion by decreasing MMP-2 and MMP-9 expression [68]. Similar to MMP-9, MMP-2 expression correlates directly with tumor grade and indirectly with survival in glioma [121]. MMP-2 increases invasion by ECM degradation and increases growth by stimulating angiogenesis [122]. Finally, let-7f decreases proliferation, migration, and invasion by binding to periostin mRNA 3′ UTR to decrease periostin expression [68]. Much work has been put into elucidating the roles of periostin, a non-structural ECM protein, in various cancers [123]. As with many oncogenes targeted by the let-7 family, periostin expression correlates directly with tumor grade and indirectly with survival in glioma [124,125]. Mechanistically, periostin recruits M2 tumor-associated macrophages to increase growth, as well as migration, proliferation, and angiogenesis [124,125,126,127,128,129]. The aforementioned targets of let-7f were validated using a murine glioma model [68]. Altogether, let-7 controls apoptotic, proliferative, and migratory signaling in glioma.
4.2. Let-7 Drives Tumor-Suppressive Paracrine Signaling
Mounting evidence suggests that let-7 family members are packaged and exported out of the glioma cell, then function as ligands in tumor-suppressive paracrine signaling. Let-7a/b/e/f/g/miR-98 contain exosome-packaging motifs, and let-7b is enriched in microvesicles isolated from glioma cell media relative to its intracellular level (Figure 6, [58,130]). One study looked at exosomes released by glioma-associated stem cells (GASCs) isolated from LGGs that had undergone anaplastic transformation before four years (labeled aggressive) or after seven years (labeled less aggressive). Let-7a/e/f were downregulated in both sets of GASCs [131]. Let-7d/g/miR-98 were downregulated exclusively in the less aggressive set [131]. Let-7a-3 (a subtype of let-7a) was also downregulated exclusively in the aggressive set [131]. Interestingly, it has also been shown that pre-let-7a-3 and its murine orthologue, pre-let-7c-2, are the only let-7 family members to escape LIN28A/B regulation [132]. Pre-let-7c-2 contains a CUCUG sequence at its short apical stem loop/preE loop junction, which impairs LIN28A/B cold shock domain binding at the preE [132]. This sequence is expected to be conserved in pre-let-7a-3 [132]. This prompts intrigue into future let-7a-3 research.
Another study found that let-7 family members function in a UUGU motif-dependent manner, as toll-like receptor 7 (TLR7) ligands to regulate tumor microglia: treatment with let-7b/e decreases growth of a murine glioma model by increasing microglial infiltration in a TLR7-dependent manner [56]. This same study showed that let-7b increases caspase-3 activation and apoptosis [56]. Let-7b/c/e/f/g increase the release of pro-inflammatory signaling molecules (cytokines), namely TNF-α, IL-6, IL-10, IL-1b, GRO-a, MIP-2, and RANTES [56]. Let-7b/e increase antigen presentation mediated by CD54 and MHC1 [56]. Lastly, let-7b/d/e treatment increases migration of the microglia into the tumor bed, commonly referred to as microglial infiltration [56]. Such preliminary evidence of let-7 as a paracrine factor offers a new and exciting direction for let-7 research in glioma.
5. Future Directions
Across glioma subtypes, LIN28A/B are overexpressed, let-7 family members are underexpressed, and let-7 family members antagonize a plethora of oncogene mRNAs to mediate glioma cell suppression (Figure 2). These steams of evidence should motivate further investigation of the LIN28B–let-7–oncogene axis in DMG.
The mechanisms driving LIN28B overexpression must be elucidated. On the genomic level, the presence of an LIN28B-TST has not been investigated in DMG [74]. If such an LIN28B-TST exists, its mRNA transcript may share the same increased stability, and accordingly, high expression, as in hepatic adenocarcinoma [74]. On the epigenomic level, there are three areas of interest: DNA methylation and histone modifications at the LIN28B locus, as well as regulation of the LIN28B mRNA transcript. Experimentally lowered DNA methylation of the AluJb promoter increases AluJb-LIN28B fusion protein expression and higher DNA methylation reduces its expression in leukemia cells [133]. Similarly, lower DNA methylation at four CpG sites has been observed at the LIN28B promoter in gastric cancer, leading to higher LIN28B expression and increased proliferation and migration [134]. A lack of H3K27me3 and surplus of H3K27ac at the LIN28B locus in H3K27M mutant DIPG cell lines has been reported [135]. The histone deacetlylase (HDAC) sirtuin 6 (SIRT6) has been shown to act on the LIN28B locus to remove K3K9ac/56ac motifs, decreasing LIN28B expression and suppressing pancreatic ductal adenocarcinoma [136]. Of note, SIRT6 is downregulated in GBM cell lines [137]. Together, these data suggest that canonical DNA methylation and histone post-translational modifications contribute to transcriptional regulation of LIN28B expression. Furthermore, the LIN28B mRNA transcript can be stabilized by LIN28B binding, resulting in its enhanced translation [31]. Meanwhile, miR-203 has been shown to repress its translation through LIN28B mRNA 3′ UTR binding in non-small cell lung cancer [138]. Unsurprisingly, miR-203 expression is inversely correlated with tumor grade in glioma [139]. Its downregulation is associated with imatinib-resistance and induction of the epithelial-mesenchymal transition (EMT) in GBM—effects mediated by the corresponding upregulation of another of its targets, SNAI2 [140]. Accordingly, miR-203 is an attractive therapeutic candidate for repressing LIN28B mRNA, and warrants further investigation.
The mechanisms driving LIN28B localization must also be elucidated. LIN28B principally localizes to the nucleolus, its cellular compartment of canonical let-7 antagonism [29]. It has been shown in pancreatic cancer that K-RAS can drive protein kinase C β (PKCβ) to phosphorylate LIN28B at serine 243 (S243), promoting translocation, which leads to decreased let-7i expression and increased expression of the let-7i target, TET3 (Figure 7, [141]). TET3 then catalyzes the conversion of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), resulting in global DNA demethylation which subsequently mediates an increase in LIN28B expression, facilitating a positive feedback loop [141]. Indeed, K-RAS amplification, TET3 overexpression, low 5mC, and high 5hmC have all already been reported in DIPG [75,142,143,144]. PKCβ has been discussed and targeted as a driver of angiogenesis, proliferation, and survival in GBM [145]. Neurofibromatosis 2 (NF2) also regulates LIN28B translocation (Figure 7, [146]). High cell density (cell contact) drives NF2 dephosphorylation at S518, enhancing its association with LIN28B [146]. This sequesters LIN28B in the cytoplasm, leading to higher let-7a/c/g expression [146]. NF2 is glioma-suppressive, increasing large-tumor suppressor signaling and decreasing canonical and non-canonical Wnt signaling [147]. Similarly, NF2 decreases glial cell proliferation by decreasing ErbB2-dependent Src-FAK-paxillin signaling [148]. The NF2 locus is hypermethylated and underexpressed in GBM [147,149]. Altogether, these data suggest that restoration of NF2 expression is a potential strategy for DMG treatment. However, a high level of S518-phosphorylated NF2 correlates with high NOTCH1 and EGFR expression in GBM, promoting proliferation [150]. Thus, both the quantity and relative phosphorylation status of NF2 require consideration. In sum, therapeutic potential lies in decreasing the translocation of LIN28B from the cytoplasm to the nucleolus.
Finally, we must consider the potential for let-7-independent mechanisms of LIN28B expression and its effects in DMG. To the authors’ knowledge, there are currently no publications on let-7-independent mechanisms of LIN28B expression effects in glioma. However, LIN28B has been shown in other cancers to bind the 3′ UTR of oncogene mRNA transcripts, increasing their stability and, accordingly, their translation. In this way, LIN28B promotes transformation and migration in colon cancer through LGR5, PROM1, and CDX2 mRNAs, promotes stemness and EMT in gastric cancer through NRP-1 mRNA, inhibits apoptosis in ovarian cancer through AKT2 mRNA, and promotes migration in neuroblastoma through MYCN-induced mRNAs [151,152,153,154,155]. In addition, LIN28B promotes migration and proliferation in cholangiocarcinoma through TGF-β-induced protein (TGFBI), although a mechanistic relationship between LIN28B and TGFBI mRNA 3′ UTR has not yet been verified [156]. Altogether, searching for let-7-independent mechanisms of LIN28B expression in DMG could be promising, potentially offering additional targets in a wider network of LIN28B-driven gliomagenesis.
6. Conclusions
Here, we provide a detailed review of the expression and function of the LIN28A/B–let-7–oncogene axis in gliomagenesis, including DMG formation. Given these data, additional efforts to elucidate LIN28B expression and localization mechanisms, as well as its functional utility as a therapeutic target, in DMG should be considered.
Author Contributions
Conceptualization, A.M.S. and T.K.; data curation, A.M.S., T.H., J.Q. and S.A.; writing—original draft preparation, T.K.; writing, review and editing, A.M.S., N.B., S.C. and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Institutes of Health, National Institute for Neurological Disorders and Stroke, grant number K08NS097624-01 (A.S.), and the APC was funded by the Pediatric Cancer Research Foundation (A.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
T.K. would like to thank Emily Wiley for her valuable support and mentorship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Srikanthan, D.; Taccone, M.S.; Van Ommeren, R.; Ishida, J.; Krumholtz, S.L.; Rutka, J.T. Diffuse Intrinsic Pontine Glioma: Current Insights and Future Directions. Chin. Neurosurg. J. 2021, 7, 6. [Google Scholar] [CrossRef]
- Jansen, M.H.; Veldhuijzen van Zanten, S.E.; Sanchez Aliaga, E.; Heymans, M.W.; Warmuth-Metz, M.; Hargrave, D.; van der Hoeven, E.J.; Gidding, C.E.; de Bont, E.S.; Eshghi, O.S.; et al. Survival Prediction Model of Children with Diffuse Intrinsic Pontine Glioma Based on Clinical and Radiological Criteria. Neuro-Oncol. 2015, 17, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, K.E. Diffuse Intrinsic Pontine Glioma: Poised for Progress. Front. Oncol. 2012, 2, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, L.M.; Veldhuijzen van Zanten, S.E.M.; Colditz, N.; Baugh, J.; Chaney, B.; Hoffmann, M.; Lane, A.; Fuller, C.; Miles, L.; Hawkins, C.; et al. Clinical, Radiologic, Pathologic, and Molecular Characteristics of Long-Term Survivors of Diffuse Intrinsic Pontine Glioma (DIPG): A Collaborative Report from the International and European Society for Pediatric Oncology DIPG Registries. J. Clin. Oncol. 2018, 36, 1963–1972. [Google Scholar] [CrossRef]
- Lulla, R.R.; Saratsis, A.M.; Hashizume, R. Mutations in Chromatin Machinery and Pediatric High-Grade Glioma. Sci. Adv. 2016, 2, e1501354. [Google Scholar] [CrossRef] [Green Version]
- Castel, D.; Philippe, C.; Calmon, R.; Le Dret, L.; Truffaux, N.; Boddaert, N.; Pagès, M.; Taylor, K.R.; Saulnier, P.; Lacroix, L.; et al. Histone H3F3A and HIST1H3B K27M Mutations Define Two Subgroups of Diffuse Intrinsic Pontine Gliomas with Different Prognosis and Phenotypes. Acta Neuropathol. 2015, 130, 815–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St. Jude Children’s Research Hospital–Washington University Pediatric Cancer Genome Project. Somatic Histone H3 Alterations in Pediatric Diffuse Intrinsic Pontine Gliomas and Non-Brainstem Glioblastomas. Nat. Genet. 2012, 44, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Schwartzentruber, J.; Korshunov, A.; Liu, X.-Y.; Jones, D.T.W.; Pfaff, E.; Jacob, K.; Sturm, D.; Fontebasso, A.M.; Quang, D.-A.K.; Tönjes, M.; et al. Driver Mutations in Histone H3.3 and Chromatin Remodelling Genes in Paediatric Glioblastoma. Nature 2012, 482, 226–231. [Google Scholar] [CrossRef]
- Sievers, P.; Sill, M.; Schrimpf, D.; Stichel, D.; Reuss, D.E.; Sturm, D.; Hench, J.; Frank, S.; Krskova, L.; Vicha, A.; et al. A Subset of Pediatric-Type Thalamic Gliomas Share a Distinct DNA Methylation Profile, H3K27me3 Loss and Frequent Alteration of EGFR. Neuro-Oncol. 2021, 23, 34–43. [Google Scholar] [CrossRef]
- Pasini, D.; Malatesta, M.; Jung, H.R.; Walfridsson, J.; Willer, A.; Olsson, L.; Skotte, J.; Wutz, A.; Porse, B.; Jensen, O.N.; et al. Characterization of an Antagonistic Switch between Histone H3 Lysine 27 Methylation and Acetylation in the Transcriptional Regulation of Polycomb Group Target Genes. Nucleic Acids Res. 2010, 38, 4958–4969. [Google Scholar] [CrossRef]
- Bender, S.; Tang, Y.; Lindroth, A.M.; Hovestadt, V.; Jones, D.T.W.; Kool, M.; Zapatka, M.; Northcott, P.A.; Sturm, D.; Wang, W.; et al. Reduced H3K27me3 and DNA Hypomethylation Are Major Drivers of Gene Expression in K27M Mutant Pediatric High-Grade Gliomas. Cancer Cell 2013, 24, 660–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piunti, A.; Hashizume, R.; Morgan, M.A.; Bartom, E.T.; Horbinski, C.M.; Marshall, S.A.; Rendleman, E.J.; Ma, Q.; Takahashi, Y.-H.; Woodfin, A.R.; et al. Therapeutic Targeting of Polycomb and BET Bromodomain Proteins in Diffuse Intrinsic Pontine Gliomas. Nat. Med. 2017, 23, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Larson, J.D.; Kasper, L.H.; Paugh, B.S.; Jin, H.; Wu, G.; Kwon, C.-H.; Fan, Y.; Shaw, T.I.; Silveira, A.B.; Qu, C.; et al. Histone H3.3 K27M Accelerates Spontaneous Brainstem Glioma and Drives Restricted Changes in Bivalent Gene Expression. Cancer Cell 2019, 35, 140–155.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.U.; Rashoff, A.Q.; Krabbenhoft, S.D.; Hoelper, D.; Do, T.J.; Gibson, T.J.; Lundgren, S.M.; Bondra, E.R.; Deshmukh, S.; Harutyunyan, A.S.; et al. H3 K27M and EZHIP Impede H3K27-Methylation Spreading by Inhibiting Allosterically Stimulated PRC2. Mol. Cell 2020, 80, 726–735.e7. [Google Scholar] [CrossRef]
- Guo, H.; Kaur, H.; Eberhart, C.G.; Raabe, E.H. Abstract 4943: The Stem Cell Factor LIN28B Regulates Proliferation and Apoptosis in Diffuse Intrinsic Pontine Glioma. Cancer Res. 2020, 80 (Suppl. S16), 4943. [Google Scholar] [CrossRef]
- Sanders, L.M.; Cheney, A.; Seninge, L.; van den Bout, A.; Chen, M.; Beale, H.C.; Kephart, E.T.; Pfeil, J.; Learned, K.; Lyle, A.G.; et al. Identification of a Differentiation Stall in Epithelial Mesenchymal Transition in Histone H3–Mutant Diffuse Midline Glioma. GigaScience 2020, 9, giaa136. [Google Scholar] [CrossRef]
- Krug, B.; De Jay, N.; Harutyunyan, A.S.; Deshmukh, S.; Marchione, D.M.; Guilhamon, P.; Bertrand, K.C.; Mikael, L.G.; McConechy, M.K.; Chen, C.C.L.; et al. Pervasive H3K27 Acetylation Leads to ERV Expression and a Therapeutic Vulnerability in H3K27M Gliomas. Cancer Cell 2019, 35, 782–797.e8. [Google Scholar] [CrossRef]
- Lewis, P.W.; Müller, M.M.; Koletsky, M.S.; Cordero, F.; Lin, S.; Banaszynski, L.A.; Garcia, B.A.; Muir, T.W.; Becher, O.J.; Allis, C.D. Inhibition of PRC2 Activity by a Gain-of-Function H3 Mutation Found in Pediatric Glioblastoma. Science 2013, 340, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/Let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Tsialikas, J.; Romer-Seibert, J. LIN28: Roles and Regulation in Development and Beyond. Development 2015, 142, 2397–2404. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, S.R.; Daley, G.Q. Lin28: A MicroRNA Regulator with a Macro Role. Cell 2010, 140, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Kawahara, H.; Okada, Y.; Imai, T.; Iwanami, A.; Mischel, P.S.; Okano, H. Musashi1 Cooperates in Abnormal Cell Lineage Protein 28 (Lin28)-Mediated Let-7 Family MicroRNA Biogenesis in Early Neural Differentiation. J. Biol. Chem. 2011, 286, 16121–16130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 Mediates the Terminal Uridylation of Let-7 Precursor MicroRNA. Mol. Cell 2008, 32, 276–284. [Google Scholar] [CrossRef]
- Heo, I.; Joo, C.; Kim, Y.-K.; Ha, M.; Yoon, M.-J.; Cho, J.; Yeom, K.-H.; Han, J.; Kim, V.N. TUT4 in Concert with Lin28 Suppresses MicroRNA Biogenesis through Pre-MicroRNA Uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, J.E.; Chang, H.-M.; Piskounova, E.; Gregory, R.I. Lin28-Mediated Control of Let-7 MicroRNA Expression by Alternative TUTases Zcchc11 (TUT4) and Zcchc6 (TUT7). RNA 2012, 18, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Yashiro, Y.; Tomita, K. Function and Regulation of Human Terminal Uridylyltransferases. Front. Genet. 2018, 9, 538. [Google Scholar] [CrossRef]
- Yamashita, S.; Nagaike, T.; Tomita, K. Crystal Structure of the Lin28-Interacting Module of Human Terminal Uridylyltransferase That Regulates Let-7 Expression. Nat. Commun. 2019, 10, 1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; LaPierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B Inhibit Let-7 MicroRNA Biogenesis by Distinct Mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and Regulation of the Let-7 MiRNAs and Their Functional Implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Hafner, M.; Max, K.E.A.; Bandaru, P.; Morozov, P.; Gerstberger, S.; Brown, M.; Molina, H.; Tuschl, T. Identification of MRNAs Bound and Regulated by Human LIN28 Proteins and Molecular Requirements for RNA Recognition. RNA 2013, 19, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Liu, X.; Zheng, J.; Song, J.; Liu, Y.; Ruan, X.; Shen, S.; Shao, L.; Yang, C.; Wang, D.; et al. Lin28A Promotes IRF6-Regulated Aerobic Glycolysis in Glioma Cells by Stabilizing SNHG14. Cell Death Dis. 2020, 11, 447. [Google Scholar] [CrossRef]
- Ambros, V.; Horvitz, H.R. Heterochronic Mutants of the Nematode Caenorhabditis Elegans. Science 1984, 226, 409–416. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-Nucleotide Let-7 RNA Regulates Developmental Timing in Caenorhabditis Elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the Sequence and Temporal Expression of Let-7 Heterochronic Regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective Blockade of MicroRNA Processing by Lin28. Science 2008, 320, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ng, S.-B.; Chng, W.-J. LIN28/LIN28B: An Emerging Oncogenic Driver in Cancer Stem Cells. Int. J. Biochem. Cell Biol. 2013, 45, 973–978. [Google Scholar] [CrossRef]
- Shamsuzzama; Kumar, L.; Haque, R.; Nazir, A. Role of MicroRNA Let-7 in Modulating Multifactorial Aspect of Neurodegenerative Diseases: An Overview. Mol. Neurobiol. 2016, 53, 2787–2793. [Google Scholar] [CrossRef]
- Patterson, M.; Gaeta, X.; Loo, K.; Edwards, M.; Smale, S.; Cinkornpumin, J.; Xie, Y.; Listgarten, J.; Azghadi, S.; Douglass, S.M.; et al. Let-7 MiRNAs Can Act through Notch to Regulate Human Gliogenesis. Stem Cell Rep. 2014, 3, 758–773. [Google Scholar] [CrossRef] [Green Version]
- Pooyan, P.; Karamzadeh, R.; Mirzaei, M.; Meyfour, A.; Amirkhan, A.; Wu, Y.; Gupta, V.; Baharvand, H.; Javan, M.; Salekdeh, G.H. The Dynamic Proteome of Oligodendrocyte Lineage Differentiation Features Planar Cell Polarity and Macroautophagy Pathways. GigaScience 2020, 9, giaa116. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, S.; Vitanza, N.A.; Woo, P.J.; Taylor, K.R.; Liu, F.; Zhang, L.; Li, M.; Meng, W.; Ponnuswami, A.; Sun, W.; et al. Transcriptional Dependencies in Diffuse Intrinsic Pontine Glioma. Cancer Cell 2017, 31, 635–652.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filbin, M.G.; Tirosh, I.; Hovestadt, V.; Shaw, M.L.; Escalante, L.E.; Mathewson, N.D.; Neftel, C.; Frank, N.; Pelton, K.; Hebert, C.M.; et al. Developmental and Oncogenic Programs in H3K27M Gliomas Dissected by Single-Cell RNA-Seq. Science 2018, 360, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortin, J.; Tian, R.; Zarrabi, I.; Hill, G.; Williams, E.; Sanchez-Duffhues, G.; Thorikay, M.; Ramachandran, P.; Siddaway, R.; Wong, J.F.; et al. Mutant ACVR1 Arrests Glial Cell Differentiation to Drive Tumorigenesis in Pediatric Gliomas. Cancer Cell 2020, 37, 308–323.e12. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.; Jiang, L.; Samuelsson, E.R.; Marco Salas, S.; Beck, A.; Hack, O.A.; Jeong, D.; Shaw, M.L.; Englinger, B.; LaBelle, J.; et al. The Landscape of Tumor Cell States and Spatial Organization in H3-K27M Mutant Diffuse Midline Glioma across Age and Location. Nat. Genet. 2022, 54, 1881–1894. [Google Scholar] [CrossRef] [PubMed]
- Korshunov, A.; Ryzhova, M.; Jones, D.T.W.; Northcott, P.A.; van Sluis, P.; Volckmann, R.; Koster, J.; Versteeg, R.; Cowdrey, C.; Perry, A.; et al. LIN28A Immunoreactivity Is a Potent Diagnostic Marker of Embryonal Tumor with Multilayered Rosettes (ETMR). Acta Neuropathol. 2012, 124, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Sin-Chan, P.; Mumal, I.; Suwal, T.; Ho, B.; Fan, X.; Singh, I.; Du, Y.; Lu, M.; Patel, N.; Torchia, J.; et al. A C19MC-LIN28A-MYCN Oncogenic Circuit Driven by Hijacked Super-Enhancers Is a Distinct Therapeutic Vulnerability in ETMRs: A Lethal Brain Tumor. Cancer Cell 2019, 36, 51–67.e7. [Google Scholar] [CrossRef] [Green Version]
- Spence, T.; Perotti, C.; Sin-Chan, P.; Picard, D.; Wu, W.; Singh, A.; Anderson, C.; Blough, M.D.; Cairncross, J.G.; Lafay-Cousin, L.; et al. A Novel C19MC Amplified Cell Line Links Lin28/Let-7 to MTOR Signaling in Embryonal Tumor with Multilayered Rosettes. Neuro-Oncol. 2014, 16, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.A.; Kim, S.-K.; Lee, J.Y.; Wang, K.-C.; Lee, C.; Phi, J.H. LIN28B Is Highly Expressed in Atypical Teratoid/Rhabdoid Tumor (AT/RT) and Suppressed through the Restoration of SMARCB1. Cancer Cell Int. 2016, 16, 32. [Google Scholar] [CrossRef] [Green Version]
- Weingart, M.F.; Roth, J.J.; Hutt-Cabezas, M.; Busse, T.M.; Kaur, H.; Price, A.; Maynard, R.; Rubens, J.; Taylor, I.; Mao, X.; et al. Disrupting LIN28 in Atypical Teratoid Rhabdoid Tumors Reveals the Importance of the Mitogen Activated Protein Kinase Pathway as a Therapeutic Target. Oncotarget 2015, 6, 3165–3177. [Google Scholar] [CrossRef] [Green Version]
- Maklad, A.; Sedeeq, M.; Wilson, R.; Heath, J.A.; Gueven, N.; Azimi, I. LIN28 Expression and Function in Medulloblastoma. J. Cell. Physiol. 2023, 238, 533–548. [Google Scholar] [CrossRef]
- Westphal, M.S.; Lee, E.; Schadt, E.E.; Sholler, G.S.; Zhu, J. Identification of Let-7 MiRNA Activity as a Prognostic Biomarker of SHH Medulloblastoma. Cancers 2021, 14, 139. [Google Scholar] [CrossRef]
- Mollashahi, B.; Aghamaleki, F.S.; Movafagh, A. The Roles of MiRNAs in Medulloblastoma: A Systematic Review. J. Cancer Prev. 2019, 24, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Hütt-Cabezas, M.; Orr, B.A.; Weingart, M.; Taylor, I.; Rajan, A.K.D.; Odia, Y.; Kahlert, U.; Maciaczyk, J.; Nikkhah, G.; et al. LIN28A Facilitates the Transformation of Human Neural Stem Cells and Promotes Glioblastoma Tumorigenesis through a Pro-Invasive Genetic Program. Oncotarget 2013, 4, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Qin, R.; Zhou, J.; Chen, C.; Xu, T.; Yan, Y.; Ma, Y.; Zheng, Z.; Shen, Y.; Lu, Y.; Fu, D.; et al. LIN28 Is Involved in Glioma Carcinogenesis and Predicts Outcomes of Glioblastoma Multiforme Patients. PLoS ONE 2014, 9, e86446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-R.; Luo, H.; Li, H.-L.; Cao, L.; Wang, X.-F.; Yan, W.; Wang, Y.-Y.; Zhang, J.-X.; Jiang, T.; Kang, C.-S.; et al. Overexpressed Let-7a Inhibits Glioma Cell Malignancy by Directly Targeting K-Ras, Independently of PTEN. Neuro-Oncol. 2013, 15, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buonfiglioli, A.; Efe, I.E.; Guneykaya, D.; Ivanov, A.; Huang, Y.; Orlowski, E.; Krüger, C.; Deisz, R.A.; Markovic, D.; Flüh, C.; et al. Let-7 MicroRNAs Regulate Microglial Function and Suppress Glioma Growth through Toll-Like Receptor 7. Cell Rep. 2019, 29, 3460–3471.e7. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, Y.; Liu, Q.; Wu, M. MicroRNA Cluster MC-let-7a-1~let-7d Promotes Autophagy and Apoptosis of Glioma Cells by Down-regulating STAT3. CNS Neurosci. Ther. 2020, 26, 319–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen, M.; Johnsen, K.B.; Olesen, P.; Pilgaard, L.; Duroux, M. MicroRNA Expression Signatures and Their Correlation with Clinicopathological Features in Glioblastoma Multiforme. NeuroMolecular Med. 2014, 16, 565–577. [Google Scholar] [CrossRef]
- Dong, Z.; Lei, Q.; Yang, R.; Zhu, S.; Ke, X.-X.; Yang, L.; Cui, H.; Yi, L. Inhibition of Neurotensin Receptor 1 Induces Intrinsic Apoptosis via Let-7a-3p/Bcl-w Axis in Glioblastoma. Br. J. Cancer 2017, 116, 1572–1584. [Google Scholar] [CrossRef] [Green Version]
- Hartman, M.L.; Czyz, M. BCL-w: Apoptotic and Non-Apoptotic Role in Health and Disease. Cell Death Dis. 2020, 11, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Carles, D.; Jackson, H.; Loveson, K.F.; Mackay, A.; Mather, R.L.; Waters, E.; Manzo, M.; Alborelli, I.; Golding, J.; Jones, C.; et al. The Long Non-Coding RNA H19 Drives the Proliferation of Diffuse Intrinsic Pontine Glioma with H3K27 Mutation. Int. J. Mol. Sci. 2021, 22, 9165. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, W.; Ge, C.; Li, X.; Yang, X.; Xiang, Y.; Sun, Z. Decreased Let-7b Is Associated with Poor Prognosis in Glioma. Medicine 2019, 98, e15784. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Hao, S.; Ye, M.; Zhang, A.; Nan, Y.; Wang, G.; Jia, Z.; Yu, K.; Guo, L.; Pu, P.; et al. MicroRNAs Let-7b/i Suppress Human Glioma Cell Invasion and Migration by Targeting IKBKE Directly. Biochem. Biophys. Res. Commun. 2015, 458, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, Y.; Liu, N.; Zhang, D.; Wan, C.; Zhao, S.; Kong, Y.; Yuan, L. Let-7b Inhibits the Malignant Behavior of Glioma Cells and Glioma Stem-like Cells via Downregulation of E2F2. J. Physiol. Biochem. 2016, 72, 733–744. [Google Scholar] [CrossRef]
- Xi, X.; Chu, Y.; Liu, N.; Wang, Q.; Yin, Z.; Lu, Y.; Chen, Y. Joint Bioinformatics Analysis of Underlying Potential Functions of Hsa-Let-7b-5p and Core Genes in Human Glioma. J. Transl. Med. 2019, 17, 129. [Google Scholar] [CrossRef]
- Xu, X.; Bao, Z.; Liu, Y.; Jiang, K.; Zhi, T.; Wang, D.; Fan, L.; Liu, N.; Ji, J. PBX3/MEK/ERK1/2/LIN28/Let-7b Positive Feedback Loop Enhances Mesenchymal Phenotype to Promote Glioblastoma Migration and Invasion. J. Exp. Clin. Cancer Res. 2018, 37, 158. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yan, K.; Fang, J.; Qu, Q.; Zhou, M.; Chen, F. Let-7b Expression Determines Response to Chemotherapy through the Regulation of Cyclin D1 in Glioblastoma. J. Exp. Clin. Cancer Res. 2013, 32, 41. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Han, X.; Xue, H.; Zhang, P.; Guo, X.; Li, T.; Guo, X.; Yuan, G.; Deng, L.; Li, G. Let-7f Inhibits Glioma Cell Proliferation, Migration, and Invasion by Targeting Periostin. J. Cell. Biochem. 2015, 116, 1680–1692. [Google Scholar] [CrossRef]
- Lee, S.-T.; Chu, K.; Oh, H.-J.; Im, W.-S.; Lim, J.-Y.; Kim, S.-K.; Park, C.-K.; Jung, K.-H.; Lee, S.K.; Kim, M.; et al. Let-7 MicroRNA Inhibits the Proliferation of Human Glioblastoma Cells. J. Neurooncol. 2011, 102, 19–24. [Google Scholar] [CrossRef]
- Chen, Z.; Cheng, Q.; Ma, Z.; Xi, H.; Peng, R.; Jiang, B. Overexpression of RKIP Inhibits Cell Invasion in Glioma Cell Lines through Upregulation of MiR-98. BioMed Res. Int. 2013, 2013, 695179. [Google Scholar] [CrossRef]
- Fan, Y.-H.; Ye, M.-H.; Wu, L.; Lv, S.-G.; Wu, M.-J.; Xiao, B.; Liao, C.-C.; Ji, Q.-K.; Chai, Y.; Zhu, X.-G. Overexpression of MiR-98 Inhibits Cell Invasion in Glioma Cell Lines via Downregulation of IKKε. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3593–3604. [Google Scholar]
- Wang, L.; Guo, S.; Zhang, H. MiR-98 Promotes Apoptosis of Glioma Cells via Suppressing IKBKE/NF-ΚB Pathway. Technol. Cancer Res. Treat. 2017, 16, 1226–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Bao, Z.; Liu, Y.; Ji, J.; Liu, N. MicroRNA-98 Attenuates Cell Migration and Invasion in Glioma by Directly Targeting Pre-B Cell Leukemia Homeobox 3. Cell. Mol. Neurobiol. 2017, 37, 1359–1371. [Google Scholar] [CrossRef]
- Guo, W.; Hu, Z.; Bao, Y.; Li, Y.; Li, S.; Zheng, Q.; Lyu, D.; Chen, D.; Yu, T.; Li, Y.; et al. A LIN28B Tumor-Specific Transcript in Cancer. Cell Rep. 2018, 22, 2016–2025. [Google Scholar] [CrossRef] [PubMed]
- Mackay, A.; Burford, A.; Carvalho, D.; Izquierdo, E.; Fazal-Salom, J.; Taylor, K.R.; Bjerke, L.; Clarke, M.; Vinci, M.; Nandhabalan, M.; et al. Integrated Molecular Meta-Analysis of 1000 Pediatric High-Grade and Diffuse Intrinsic Pontine Glioma. Cancer Cell 2017, 32, 520–537.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findlay, I.J.; De Iuliis, G.N.; Duchatel, R.J.; Jackson, E.R.; Vitanza, N.A.; Cain, J.E.; Waszak, S.M.; Dun, M.D. Pharmaco-Proteogenomic Profiling of Pediatric Diffuse Midline Glioma to Inform Future Treatment Strategies. Oncogene 2022, 41, 461–475. [Google Scholar] [CrossRef]
- Molenaar, J.J.; Domingo-Fernández, R.; Ebus, M.E.; Lindner, S.; Koster, J.; Drabek, K.; Mestdagh, P.; van Sluis, P.; Valentijn, L.J.; van Nes, J.; et al. LIN28B Induces Neuroblastoma and Enhances MYCN Levels via Let-7 Suppression. Nat. Genet. 2012, 44, 1199–1206. [Google Scholar] [CrossRef]
- Chang, T.-C.; Zeitels, L.R.; Hwang, H.-W.; Chivukula, R.R.; Wentzel, E.A.; Dews, M.; Jung, J.; Gao, P.; Dang, C.V.; Beer, M.A.; et al. Lin-28B Transactivation Is Necessary for Myc-Mediated Let-7 Repression and Proliferation. Proc. Natl. Acad. Sci. USA 2009, 106, 3384–3389. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N.A.; Klein, R.H.; Kelly, C.; Yee, J.; Knoepfler, P.S. Histone H3.3 K27M Chromatin Functions Implicate a Network of Neurodevelopmental Factors Including ASCL1 and NEUROD1 in DIPG. Epigenet. Chromatin 2022, 15, 18. [Google Scholar] [CrossRef]
- Degrauwe, N.; Schlumpf, T.B.; Janiszewska, M.; Martin, P.; Cauderay, A.; Provero, P.; Riggi, N.; Suvà, M.-L.; Paro, R.; Stamenkovic, I. The RNA Binding Protein IMP2 Preserves Glioblastoma Stem Cells by Preventing Let-7 Target Gene Silencing. Cell Rep. 2016, 15, 1634–1647. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wang, G.; Hao, D.; Liu, X.; Wang, D.; Ning, N.; Li, X. Aberrant Regulation of the LIN28A/LIN28B and Let-7 Loop in Human Malignant Tumors and Its Effects on the Hallmarks of Cancer. Mol. Cancer 2015, 14, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funato, K.; Major, T.; Lewis, P.W.; Allis, C.D.; Tabar, V. Use of Human Embryonic Stem Cells to Model Pediatric Gliomas with H3.3K27M Histone Mutation. Science 2014, 346, 1529–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Q.; Zhou, J.; Yang, W.; Cui, H.; Xu, M.; Yi, L. Oncogenic Role of Neurotensin and Neurotensin Receptors in Various Cancers. Clin. Exp. Pharmacol. Physiol. 2017, 44, 841–846. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Q.; Gong, X.; Xiao, H.; Zhou, J.; Xu, M.; Dai, Y.; Xu, L.; Feng, H.; Cui, H.; Yi, L. Neurotensin Promotes the Progression of Malignant Glioma through NTSR1 and Impacts the Prognosis of Glioma Patients. Mol. Cancer 2015, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Q.; Chen, G.; Zhou, J.; Li, L.; Dong, Z.; Yang, R.; Xu, L.; Cui, H.; Xu, M.; Yi, L. Neurotensin Signaling Stimulates Glioblastoma Cell Proliferation by Upregulating C-Myc and Inhibiting MiR-29b-1 and MiR-129-3p. Neuro-Oncol. 2016, 18, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Wang, X.; Lu, J. Advances in IKBKE as a Potential Target for Cancer Therapy. Cancer Med. 2020, 9, 247–258. [Google Scholar] [CrossRef]
- Guan, H.; Zhang, H.; Cai, J.; Wu, J.; Yuan, J.; Li, J.; Huang, Z.; Li, M. IKBKE Is Over-Expressed in Glioma and Contributes to Resistance of Glioma Cells to Apoptosis via Activating NF-ΚB: IKBKE Inhibits Apoptosis via Activating NF-ΚB. J. Pathol. 2011, 223, 436–445. [Google Scholar] [CrossRef]
- Soubannier, V.; Stifani, S. NF-ΚB Signalling in Glioblastoma. Biomedicines 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youle, R.J.; Strasser, A. The BCL-2 Protein Family: Opposing Activities That Mediate Cell Death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Xue, Q.; Cao, L.; Chen, X.-Y.; Zhao, J.; Gao, L.; Li, S.-Z.; Fei, Z. High Expression of MMP9 in Glioma Affects Cell Proliferation and Is Associated with Patient Survival Rates. Oncol. Lett. 2017, 13, 1325–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egeblad, M.; Werb, Z. New Functions for the Matrix Metalloproteinases in Cancer Progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Noronha, C.; Ribeiro, A.S.; Taipa, R.; Castro, D.S.; Reis, J.; Faria, C.; Paredes, J. Cadherin Expression and EMT: A Focus on Gliomas. Biomedicines 2021, 9, 1328. [Google Scholar] [CrossRef] [PubMed]
- Simanshu, D.K.; Nissley, D.V.; McCormick, F. RAS Proteins and Their Regulators in Human Disease. Cell 2017, 170, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Koncar, R.F.; Dey, B.R.; Stanton, A.-C.J.; Agrawal, N.; Wassell, M.L.; McCarl, L.H.; Locke, A.L.; Sanders, L.; Morozova-Vaske, O.; Myers, M.I.; et al. Identification of Novel RAS Signaling Therapeutic Vulnerabilities in Diffuse Intrinsic Pontine Gliomas. Cancer Res. 2019, 79, 4026–4041. [Google Scholar] [CrossRef]
- Phillips, J.J.; Huillard, E.; Robinson, A.E.; Ward, A.; Lum, D.H.; Polley, M.-Y.; Rosen, S.D.; Rowitch, D.H.; Werb, Z. Heparan Sulfate Sulfatase SULF2 Regulates PDGFRα Signaling and Growth in Human and Mouse Malignant Glioma. J. Clin. Investig. 2012, 122, 911–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Ichikawa, T.; Kurozumi, K.; Otani, Y.; Fujimura, A.; Fujii, K.; Tomita, Y.; Hattori, Y.; Uneda, A.; Tsuboi, N.; et al. Annexin A2–STAT3–Oncostatin M Receptor Axis Drives Phenotypic and Mesenchymal Changes in Glioblastoma. Acta Neuropathol. Commun. 2020, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, Q.; Pierrevelcin, M.; Messe, M.; Lhermitte, B.; Blandin, A.-F.; Papin, C.; Coca, A.; Dontenwill, M.; Entz-Werlé, N. Hypoxia Inducible Factors’ Signaling in Pediatric High-Grade Gliomas: Role, Modelization and Innovative Targeted Approaches. Cancers 2020, 12, 979. [Google Scholar] [CrossRef] [Green Version]
- Natesh, K.; Bhosale, D.; Desai, A.; Chandrika, G.; Pujari, R.; Jagtap, J.; Chugh, A.; Ranade, D.; Shastry, P. Oncostatin-M Differentially Regulates Mesenchymal and Proneural Signature Genes in Gliomas via STAT3 Signaling. Neoplasia 2015, 17, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Chanoch-Myers, R.; Mathewson, N.D.; Myskiw, C.; Atta, L.; Bussema, L.; Eichhorn, S.W.; Greenwald, A.C.; Kinker, G.S.; Rodman, C.; et al. Interactions between Cancer Cells and Immune Cells Drive Transitions to Mesenchymal-like States in Glioblastoma. Cancer Cell 2021, 39, 779–792.e11. [Google Scholar] [CrossRef]
- Mohan, S.; Bonni, A.; Jahani-Asl, A. Targeting OSMR in Glioma Stem Cells. Oncotarget 2017, 8, 16103–16104. [Google Scholar] [CrossRef]
- Sharanek, A.; Burban, A.; Laaper, M.; Heckel, E.; Joyal, J.-S.; Soleimani, V.D.; Jahani-Asl, A. OSMR Controls Glioma Stem Cell Respiration and Confers Resistance of Glioblastoma to Ionizing Radiation. Nat. Commun. 2020, 11, 4116. [Google Scholar] [CrossRef] [PubMed]
- Kallen, A.N.; Zhou, X.-B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.-S.; Zhang, H.; et al. The Imprinted H19 LncRNA Antagonizes Let-7 MicroRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Li, Z.; Wang, M. Expression and Prognostic Role of E2F Transcription Factors in High-grade Glioma. CNS Neurosci. Ther. 2020, 26, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Attwooll, C.; Denchi, E.L.; Helin, K. The E2F Family: Specific Functions and Overlapping Interests. EMBO J. 2004, 23, 4709–4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Sun, C.; Tang, D.; Yang, G.; Zhou, X.; Wang, D. Identification of Key Genes in Glioblastoma-Associated Stromal Cells Using Bioinformatics Analysis. Oncol. Lett. 2016, 11, 3999–4007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zeng, W.; Sun, H.; Huang, F.; Yang, C.; Cai, X.; Lu, Y.; Zeng, J.; Yang, K. Bioinformatical Analysis of Gene Expression Omnibus Database Associates TAF7/CCNB1, TAF7/CCNA2, and GTF2E2/CDC20 Pathways with Glioblastoma Development and Prognosis. World Neurosurg. 2020, 138, e492–e514. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhong, M.; Chen, T.; Zhu, X.; Yang, H.; Lv, K. Gene Regulation Network Analysis Reveals Core Genes Associated with Survival in Glioblastoma Multiforme. J. Cell. Mol. Med. 2020, 24, 10075–10087. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Ditzel, H.J.; Duijf, P.H.G.; Khaze, V.; Gjerstorff, M.F.; Baradaran, B. HMGA2 as a Critical Regulator in Cancer Development. Genes 2021, 12, 269. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, H.; Yu, L. HMGA2 Promotes Glioma Invasion and Poor Prognosis via a Long-range Chromatin Interaction. Cancer Med. 2018, 7, 3226–3239. [Google Scholar] [CrossRef]
- Kaur, H.; Ali, S.Z.; Huey, L.; Hütt-Cabezas, M.; Taylor, I.; Mao, X.; Weingart, M.; Chu, Q.; Rodriguez, F.J.; Eberhart, C.G.; et al. The Transcriptional Modulator HMGA2 Promotes Stemness and Tumorigenicity in Glioblastoma. Cancer Lett. 2016, 377, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Liu, X.; Li, Y.; Cheng, M.; Wang, W.; Tian, K.; Mu, L.; Zeng, T.; Liu, Y.; Jiang, X.; et al. HMGA2 Sustains Self-Renewal and Invasiveness of Glioma-Initiating Cells. Oncotarget 2016, 7, 44365–44380. [Google Scholar] [CrossRef]
- Loftus, J.C.; Ross, J.T.D.; Paquette, K.M.; Paulino, V.M.; Nasser, S.; Yang, Z.; Kloss, J.; Kim, S.; Berens, M.E.; Tran, N.L. MiRNA Expression Profiling in Migrating Glioblastoma Cells: Regulation of Cell Migration and Invasion by MiR-23b via Targeting of Pyk2. PLoS ONE 2012, 7, e39818. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Patel, M.; Ruzevick, J.; Jackson, C.; Lim, M. STAT3 Activation in Glioblastoma: Biochemical and Therapeutic Implications. Cancers 2014, 6, 376–395. [Google Scholar] [CrossRef] [Green Version]
- de la Iglesia, N.; Konopka, G.; Lim, K.-L.; Nutt, C.L.; Bromberg, J.F.; Frank, D.A.; Mischel, P.S.; Louis, D.N.; Bonni, A. Deregulation of a STAT3-Interleukin 8 Signaling Pathway Promotes Human Glioblastoma Cell Proliferation and Invasiveness. J. Neurosci. 2008, 28, 5870–5878. [Google Scholar] [CrossRef]
- Fu, M.; Wang, C.; Li, Z.; Sakamaki, T.; Pestell, R.G. Minireview: Cyclin D1: Normal and Abnormal Functions. Endocrinology 2004, 145, 5439–5447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Dai, D.; Zhou, M.; Li, Z.; Wang, C.; Lu, Y.; Li, Y.; Wang, J. Inhibition of Cyclin D1 Expression in Human Glioblastoma Cells Is Associated with Increased Temozolomide Chemosensitivity. Cell. Physiol. Biochem. 2018, 51, 2496–2508. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.C.; Clurman, B.E. Cyclin E in Normal and Neoplastic Cell Cycles. Oncogene 2005, 24, 2776–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, T.; Dutta, A. P21 in Cancer: Intricate Networks and Multiple Activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Chu, I.M.; Hengst, L.; Slingerland, J.M. The Cdk Inhibitor P27 in Human Cancer: Prognostic Potential and Relevance to Anticancer Therapy. Nat. Rev. Cancer 2008, 8, 253–267. [Google Scholar] [CrossRef]
- Westphal, D.; Dewson, G.; Czabotar, P.E.; Kluck, R.M. Molecular Biology of Bax and Bak Activation and Action. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2011, 1813, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Sincevičiūtė, R.; Vaitkienė, P.; Urbanavičiūtė, R.; Steponaitis, G.; Tamašauskas, A.; Skiriutė, D. MMP2 Is Associated with Glioma Malignancy and Patient Outcome. Int. J. Clin. Exp. Pathol. 2018, 11, 3010–3018. [Google Scholar]
- Yu, C.-F.; Chen, F.-H.; Lu, M.-H.; Hong, J.-H.; Chiang, C.-S. Dual Roles of Tumour Cells-Derived Matrix Metalloproteinase 2 on Brain Tumour Growth and Invasion. Br. J. Cancer 2017, 117, 1828–1836. [Google Scholar] [CrossRef] [Green Version]
- González-González, L.; Alonso, J. Periostin: A Matricellular Protein with Multiple Functions in Cancer Development and Progression. Front. Oncol. 2018, 8, 225. [Google Scholar] [CrossRef] [Green Version]
- Mikheev, A.M.; Mikheeva, S.A.; Trister, A.D.; Tokita, M.J.; Emerson, S.N.; Parada, C.A.; Born, D.E.; Carnemolla, B.; Frankel, S.; Kim, D.-H.; et al. Periostin Is a Novel Therapeutic Target That Predicts and Regulates Glioma Malignancy. Neuro-Oncol. 2015, 17, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, Y.; Jiang, C. Stromal Protein Periostin Identified as a Progression Associated and Prognostic Biomarker in Glioma via Inducing an Invasive and Proliferative Phenotype. Int. J. Oncol. 2013, 42, 1716–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouanouki, A.; Lamy, S.; Annabi, B. Periostin, a Signal Transduction Intermediate in TGF-β-Induced EMT in U-87MG Human Glioblastoma Cells, and Its Inhibition by Anthocyanidins. Oncotarget 2018, 9, 22023–22037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Piao, Y.; Jeong, K.J.; Dong, J.; de Groot, J.F. Periostin (POSTN) Regulates Tumor Resistance to Antiangiogenic Therapy in Glioma Models. Mol. Cancer Ther. 2016, 15, 2187–2197. [Google Scholar] [CrossRef] [Green Version]
- Huizer, K.; Zhu, C.; Chirifi, I.; Krist, B.; Zorgman, D.; van der Weiden, M.; van den Bosch, T.P.P.; Dumas, J.; Cheng, C.; Kros, J.M.; et al. Periostin Is Expressed by Pericytes and Is Crucial for Angiogenesis in Glioma. J. Neuropathol. Exp. Neurol. 2020, 79, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ke, S.Q.; Huang, Z.; Flavahan, W.; Fang, X.; Paul, J.; Wu, L.; Sloan, A.E.; McLendon, R.E.; Li, X.; et al. Periostin Secreted by Glioblastoma Stem Cells Recruits M2 Tumour-Associated Macrophages and Promotes Malignant Growth. Nat. Cell Biol. 2015, 17, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.C.; Eaton, S.A.; Young, P.E.; Lee, M.; Shuttleworth, R.; Humphreys, D.T.; Grau, G.E.; Combes, V.; Bebawy, M.; Gong, J.; et al. Glioma Microvesicles Carry Selectively Packaged Coding and Non-Coding RNAs Which Alter Gene Expression in Recipient Cells. RNA Biol. 2013, 10, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Caponnetto, F.; Dalla, E.; Mangoni, D.; Piazza, S.; Radovic, S.; Ius, T.; Skrap, M.; Di Loreto, C.; Beltrami, A.P.; Manini, I.; et al. The MiRNA Content of Exosomes Released from the Glioma Microenvironment Can Affect Malignant Progression. Biomedicines 2020, 8, 564. [Google Scholar] [CrossRef]
- Triboulet, R.; Pirouz, M.; Gregory, R.I. A Single Let-7 MicroRNA Bypasses LIN28-Mediated Repression. Cell Rep. 2015, 13, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.S.; Shah, N.M.; Du, A.Y.; Dailey, Z.Z.; Pehrsson, E.C.; Godoy, P.M.; Zhang, D.; Li, D.; Xing, X.; Kim, S.; et al. Transposable Elements Drive Widespread Expression of Oncogenes in Human Cancers. Nat. Genet. 2019, 51, 611–617. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Yang, J.; Gu, Y.; Zhang, E.; Yuan, W.; Wang, C.; Jin, G.; Ma, H.; Hu, Z. Hypomethylation-Activated Cancer-Testis Gene LIN28B Promotes Cell Proliferation and Metastasis in Gastric Cancer. Gene 2022, 813, 146115. [Google Scholar] [CrossRef]
- Wang, J.; Huang, T.Y.-T.; Hou, Y.; Bartom, E.; Lu, X.; Shilatifard, A.; Yue, F.; Saratsis, A. Epigenomic Landscape and 3D Genome Structure in Pediatric High-Grade Glioma. Sci. Adv. 2021, 7, eabg4126. [Google Scholar] [CrossRef] [PubMed]
- Kugel, S.; Sebastián, C.; Fitamant, J.; Ross, K.N.; Saha, S.K.; Jain, E.; Gladden, A.; Arora, K.S.; Kato, Y.; Rivera, M.N.; et al. SIRT6 Suppresses Pancreatic Cancer through Control of Lin28b. Cell 2016, 165, 1401–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, D.; Gao, Y.; Cao, Y.; Hao, B. Histone Deacetylase SIRT6 Inhibits Glioma Cell Growth through Down-Regulating NOTCH3 Expression. Acta Biochim. Biophys. Sin. 2018, 50, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liang, H.; Liao, Z.; Wang, Y.; Hu, X.; Chen, X.; Xu, L.; Hu, Z. MiR-203 Enhances Let-7 Biogenesis by Targeting LIN28B to Suppress Tumor Growth in Lung Cancer. Sci. Rep. 2017, 7, 42680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Deng, Y.; Yang, G.; Xie, W. MicroRNA-203 down-Regulation Is Associated with Unfavorable Prognosis in Human Glioma: Prognostic Value of MiR-203 in Glioma. J. Surg. Oncol. 2013, 108, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Bai, Y.; Qiu, S.; Zheng, L.; Huang, L.; Liu, T.; Wang, X.; Liu, Y.; Xu, N.; Yan, X.; et al. MiR-203 Downregulation Is Responsible for Chemoresistance in Human Glioblastoma by Promoting Epithelial-Mesenchymal Transition via SNAI2. Oncotarget 2015, 6, 8914–8928. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, D.; Zhou, M.; Chen, H.; Wang, H.; Min, J.; Chen, J.; Wu, S.; Ni, X.; Zhang, Y.; et al. The KRAS/Lin28B Axis Maintains Stemness of Pancreatic Cancer Cells via the Let-7i/TET3 Pathway. Mol. Oncol. 2021, 15, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, S.; Raabe, E.H.; Haffner, M.C.; Vaghasia, A.; Warren, K.E.; Quezado, M.; Ballester, L.Y.; Nazarian, J.; Eberhart, C.G.; Rodriguez, F.J. Increased 5-Hydroxymethylcytosine and Decreased 5-Methylcytosine Are Indicators of Global Epigenetic Dysregulation in Diffuse Intrinsic Pontine Glioma. Acta Neuropathol. Commun. 2014, 2, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajovic, S.; Siddaway, R.; Bridge, T.; Sheth, J.; Rakopoulos, P.; Kim, B.; Ryall, S.; Agnihotri, S.; Phillips, L.; Yu, M.; et al. Epigenetic Activation of a RAS/MYC Axis in H3.3K27M-Driven Cancer. Nat. Commun. 2020, 11, 6216. [Google Scholar] [CrossRef] [PubMed]
- Paugh, B.S.; Broniscer, A.; Qu, C.; Miller, C.P.; Zhang, J.; Tatevossian, R.G.; Olson, J.M.; Geyer, J.R.; Chi, S.N.; da Silva, N.S.; et al. Genome-Wide Analyses Identify Recurrent Amplifications of Receptor Tyrosine Kinases and Cell-Cycle Regulatory Genes in Diffuse Intrinsic Pontine Glioma. J. Clin. Oncol. 2011, 29, 3999–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- do Carmo, A.; Balça-Silva, J.; Matias, D.; Lopes, M. PKC Signaling in Glioblastoma. Cancer Biol. Ther. 2013, 14, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hikasa, H.; Sekido, Y.; Suzuki, A. Merlin/NF2-Lin28B-Let-7 Is a Tumor-Suppressive Pathway That Is Cell-Density Dependent and Hippo Independent. Cell Rep. 2016, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, Y.-K.I.; Murray, L.B.; Houshmandi, S.S.; Xu, Y.; Gutmann, D.H.; Yu, Q. Merlin Is a Potent Inhibitor of Glioma Growth. Cancer Res. 2008, 68, 5733–5742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houshmandi, S.S.; Emnett, R.J.; Giovannini, M.; Gutmann, D.H. The Neurofibromatosis 2 Protein, Merlin, Regulates Glial Cell Growth in an ErbB2- and Src-Dependent Manner. Mol. Cell. Biol. 2009, 29, 1472–1486. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Tian, X.; Zhang, J.; Huang, Y.; Lin, X.; Chen, L.; Zhang, S. Regulation of Human Glioma Cell Apoptosis and Invasion by MiR-152-3p through Targeting DNMT1 and Regulating NF2: MiR-152-3p Regulate Glioma Cell Apoptosis and Invasion. J. Exp. Clin. Cancer Res. 2017, 36, 100. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, P.A.; Yin, W.; Camacho, L.; Marchetti, D. Oncogenic Role of Merlin/NF2 in Glioblastoma. Oncogene 2015, 34, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- King, C.E.; Wang, L.; Winograd, R.; Madison, B.B.; Mongroo, P.S.; Johnstone, C.N.; Rustgi, A.K. LIN28B Fosters Colon Cancer Migration, Invasion and Transformation through Let-7-Dependent and -Independent Mechanisms. Oncogene 2011, 30, 4185–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Hu, H.; Liu, H. RNA Binding Protein Lin28B Confers Gastric Cancer Cells Stemness via Directly Binding to NRP-1. Biomed. Pharmacother. 2018, 104, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Shen, J.; Peng, D.; He, X.; Xu, C.; Chen, X.; Tanyi, J.L.; Montone, K.; Fan, Y.; Huang, Q.; et al. RNA-Binding Protein LIN28B Inhibits Apoptosis through Regulation of the AKT2/FOXO3A/BIM Axis in Ovarian Cancer Cells. Signal Transduct. Target. Ther. 2018, 3, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missios, P.; da Rocha, E.L.; Pearson, D.S.; Philipp, J.; Aleman, M.M.; Pirouz, M.; Farache, D.; Franses, J.W.; Kubaczka, C.; Tsanov, K.M.; et al. LIN28B Alters Ribosomal Dynamics to Promote Metastasis in MYCN-Driven Malignancy. J. Clin. Investig. 2021, 131, e145142. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Masuike, Y.; Mizuno, R.; Sachdeva, U.M.; Chatterji, P.; Andres, S.F.; Sun, W.; Klein-Szanto, A.J.; Besharati, S.; Remotti, H.E.; et al. LIN28B Induces a Differentiation Program through CDX2 in Colon Cancer. JCI Insight 2021, 6, e140382. [Google Scholar] [CrossRef] [PubMed]
- Puthdee, N.; Sriswasdi, S.; Pisitkun, T.; Ratanasirintrawoot, S.; Israsena, N.; Tangkijvanich, P. The LIN28B/TGF-β/TGFBI Feedback Loop Promotes Cell Migration and Tumour Initiation Potential in Cholangiocarcinoma. Cancer Gene Ther. 2022, 29, 445–455. [Google Scholar] [CrossRef]
Figure 1.
Overview of LIN28A/B activities in the suppression of let-7 miR family member biogenesis. There are two splicing events during let-7 biogenesis: pri-let-7 is spliced by the microprocessor (DROSHA:DGCR8) into pre-let-7 in the nucleus, then pre-let-7 is spliced by DICER in the cytoplasm. Let-7 is then incorporated into RISC to direct translational repression. LIN28A cooperates with MSI1 and TUT4/7 to suppress let-7 biogenesis in the nucleus and cytoplasm, respectively. LIN28B suppresses let-7 biogenesis individually in the nucleolus.
Figure 1.
Overview of LIN28A/B activities in the suppression of let-7 miR family member biogenesis. There are two splicing events during let-7 biogenesis: pri-let-7 is spliced by the microprocessor (DROSHA:DGCR8) into pre-let-7 in the nucleus, then pre-let-7 is spliced by DICER in the cytoplasm. Let-7 is then incorporated into RISC to direct translational repression. LIN28A cooperates with MSI1 and TUT4/7 to suppress let-7 biogenesis in the nucleus and cytoplasm, respectively. LIN28B suppresses let-7 biogenesis individually in the nucleolus.

Figure 2.
A conceptual model for the LIN28B–let-7–oncogene axis in DMG. The H3K27M epigenetic landscape may drive LIN28B expression, which decreases mature let-7 levels, which increases oncogene expression, facilitating DMG formation. This would make the LIN28B–let-7–oncogene axis a mechanistic mediator of H3K27M mutant DMG. Further investigation into the necessity and sufficiency of the H3K27M mutation to promote LIN28B expression in DMG is needed, as it may be the case that even in H3WT tumors LIN28B is upregulated, thereby making H3K27M oncohistones dispensable. DMG, diffuse midline glioma.
Figure 2.
A conceptual model for the LIN28B–let-7–oncogene axis in DMG. The H3K27M epigenetic landscape may drive LIN28B expression, which decreases mature let-7 levels, which increases oncogene expression, facilitating DMG formation. This would make the LIN28B–let-7–oncogene axis a mechanistic mediator of H3K27M mutant DMG. Further investigation into the necessity and sufficiency of the H3K27M mutation to promote LIN28B expression in DMG is needed, as it may be the case that even in H3WT tumors LIN28B is upregulated, thereby making H3K27M oncohistones dispensable. DMG, diffuse midline glioma.

Figure 3.
LIN28B transcriptional dynamics and let-7 functional antagonism in glioma. NFYA and c-Myc drive the transcription of LIN28B-TST and LIN28B-WT, respectively. Meanwhile, IMP2 blocks function of let-7-loaded RISC to derepress let-7 target oncogenes, thereby recapitulating the cellular phenotype induced by let-7-dependent LIN28A/B function. LIN28B-TST, LIN28B tumor-specific transcript. LIN28B-WT, LIN28B wild-type transcript. ONC, oncogene. Grey double helix, DNA. Blue single helix, mRNA. Orange two-subunit structure, ribosome.
Figure 3.
LIN28B transcriptional dynamics and let-7 functional antagonism in glioma. NFYA and c-Myc drive the transcription of LIN28B-TST and LIN28B-WT, respectively. Meanwhile, IMP2 blocks function of let-7-loaded RISC to derepress let-7 target oncogenes, thereby recapitulating the cellular phenotype induced by let-7-dependent LIN28A/B function. LIN28B-TST, LIN28B tumor-specific transcript. LIN28B-WT, LIN28B wild-type transcript. ONC, oncogene. Grey double helix, DNA. Blue single helix, mRNA. Orange two-subunit structure, ribosome.

Figure 4.
Metabolic activities downstream of LIN28A. LIN28A stabilizes SNHG14, driving an STAU1- and UPF1-dependent pathway that downregulates IRF6 expression. This derepresses IRF6-silenced genes PKM2 and GLUT1, resulting in an upregulation of aerobic glycolysis. Grey double helix, DNA. Blue single helix, mRNA. Green single helix, lncRNA. Orange two-subunit structure, ribosome. Pink box, programming.
Figure 4.
Metabolic activities downstream of LIN28A. LIN28A stabilizes SNHG14, driving an STAU1- and UPF1-dependent pathway that downregulates IRF6 expression. This derepresses IRF6-silenced genes PKM2 and GLUT1, resulting in an upregulation of aerobic glycolysis. Grey double helix, DNA. Blue single helix, mRNA. Green single helix, lncRNA. Orange two-subunit structure, ribosome. Pink box, programming.

Figure 5.
Anti-apoptotic and metastatic mechanisms downstream of let-7 target IKBKE in glioma. IKBKE drives NF-κB-mediated MMP-9 and Bcl-2 expression while also downregulating E-cad expression. This inhibits apoptosis and facilitates metastasis. Blue single helix, mRNA. Orange two-subunit structure, ribosome. Pink box, programming.
Figure 5.
Anti-apoptotic and metastatic mechanisms downstream of let-7 target IKBKE in glioma. IKBKE drives NF-κB-mediated MMP-9 and Bcl-2 expression while also downregulating E-cad expression. This inhibits apoptosis and facilitates metastasis. Blue single helix, mRNA. Orange two-subunit structure, ribosome. Pink box, programming.

Figure 6.
Mechanisms of mature let-7 cytoplasmic sequestration or exportation in glioma. Mature let-7a-5 is bound by H19 while several let-7 family members are packaged for export. Each mechanism decreases the amount of let-7 loaded into RISC, thereby decreasing canonical let-7 activity. Green single helix, lncRNA.
Figure 6.
Mechanisms of mature let-7 cytoplasmic sequestration or exportation in glioma. Mature let-7a-5 is bound by H19 while several let-7 family members are packaged for export. Each mechanism decreases the amount of let-7 loaded into RISC, thereby decreasing canonical let-7 activity. Green single helix, lncRNA.

Figure 7.
Potential mechanisms regulating LIN28B nucleolar localization in glioma and cytoplasmic sequestration in glioma. K-RAS drives PKCβ-mediated LIN28B phosphorylation, facilitating its nucleolar translocation. Meanwhile, cell density dephosphorylates NF2, enhancing its association with LIN28B to result in cytoplasmic sequestration. Each mechanism must be verified in glioma.
Figure 7.
Potential mechanisms regulating LIN28B nucleolar localization in glioma and cytoplasmic sequestration in glioma. K-RAS drives PKCβ-mediated LIN28B phosphorylation, facilitating its nucleolar translocation. Meanwhile, cell density dephosphorylates NF2, enhancing its association with LIN28B to result in cytoplasmic sequestration. Each mechanism must be verified in glioma.


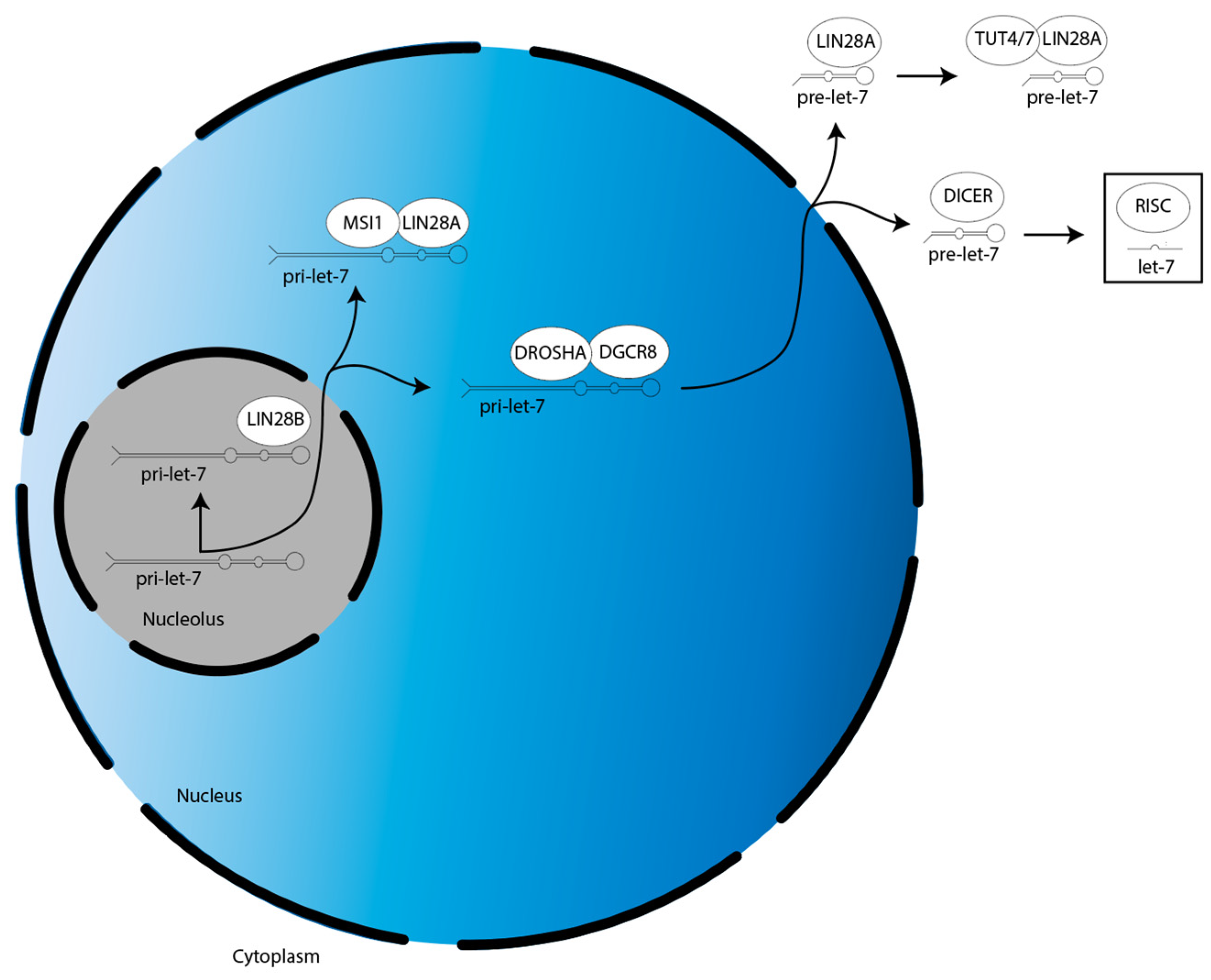{kind=link}
{kind=link}
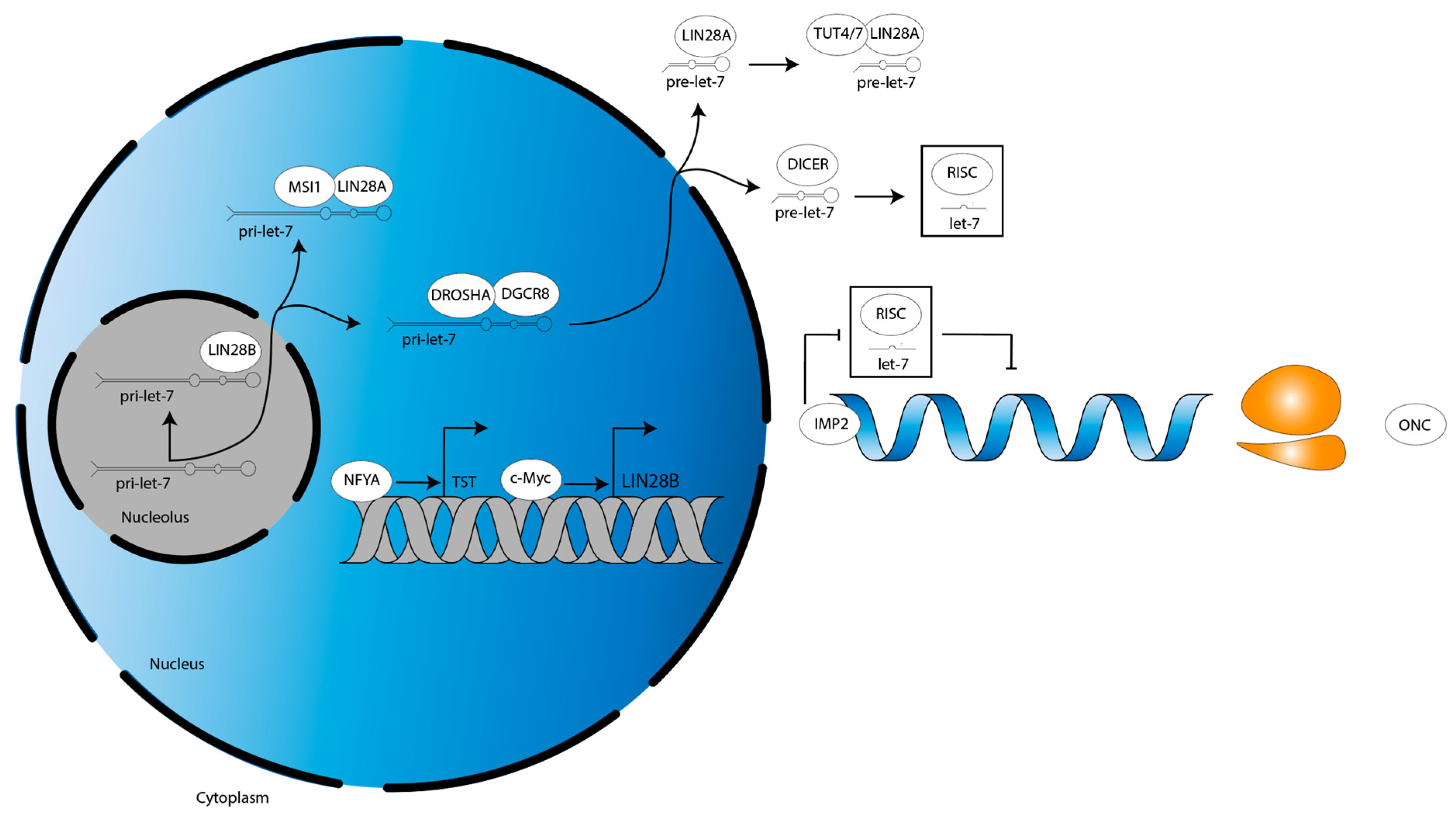{kind=link}
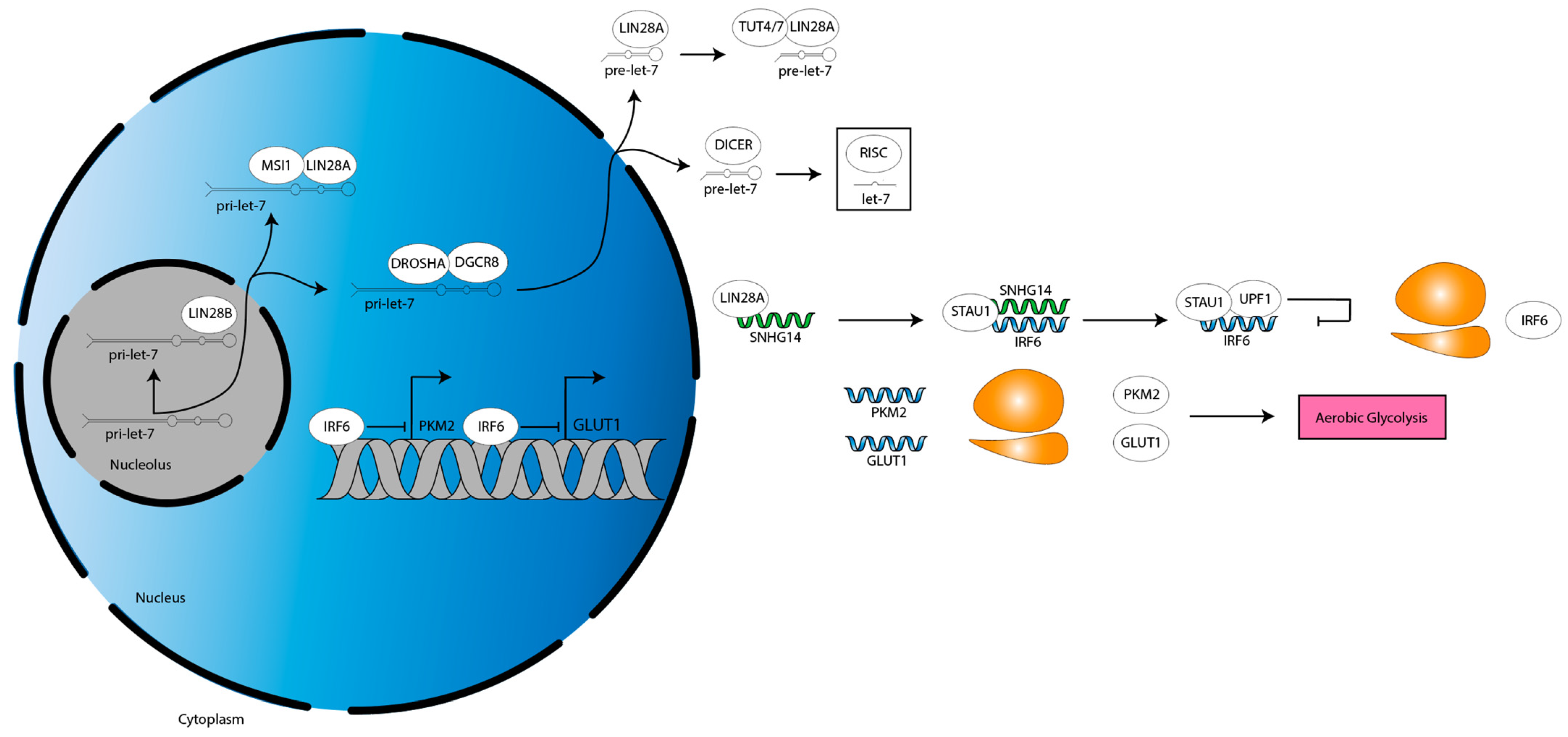{kind=link}
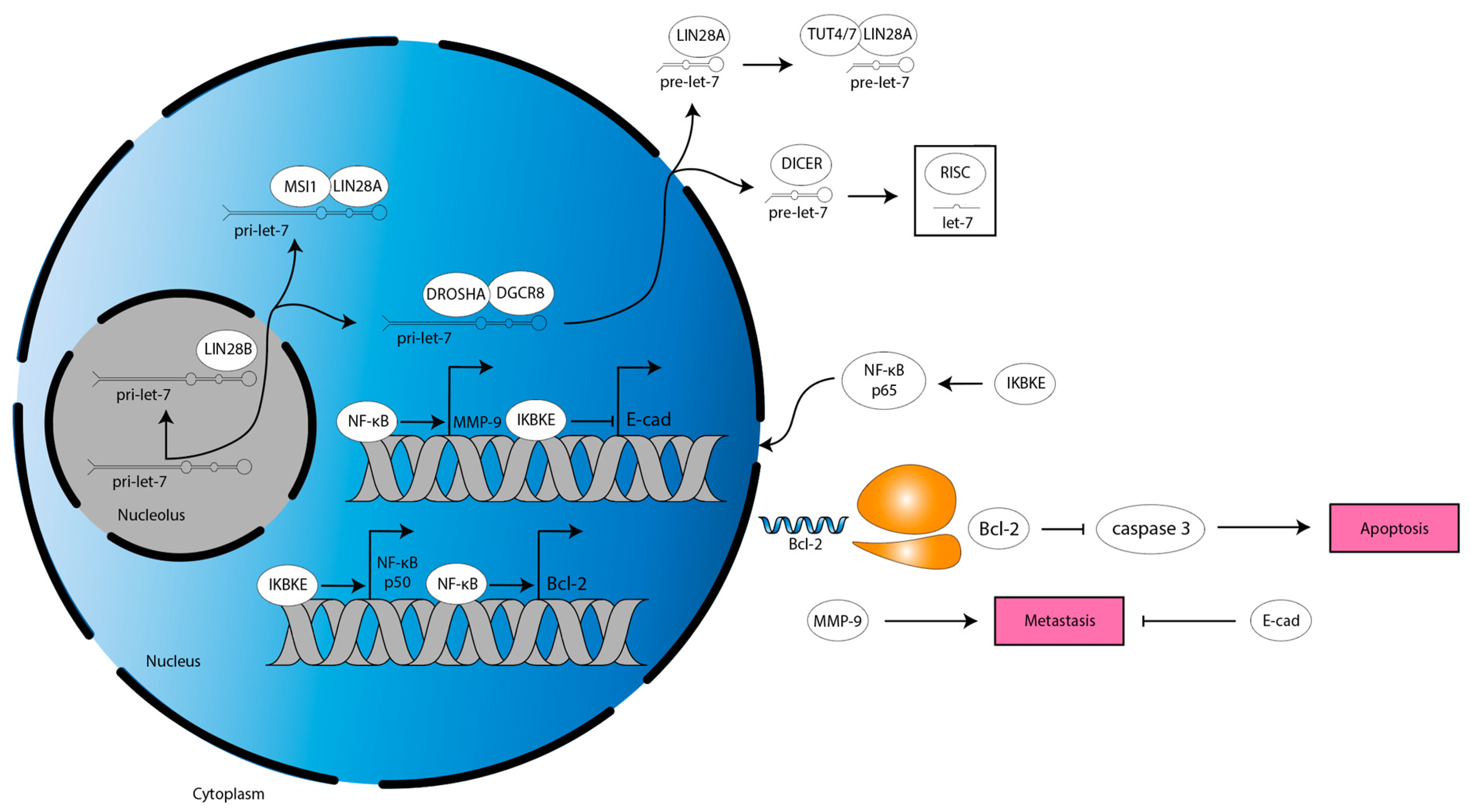{kind=link}
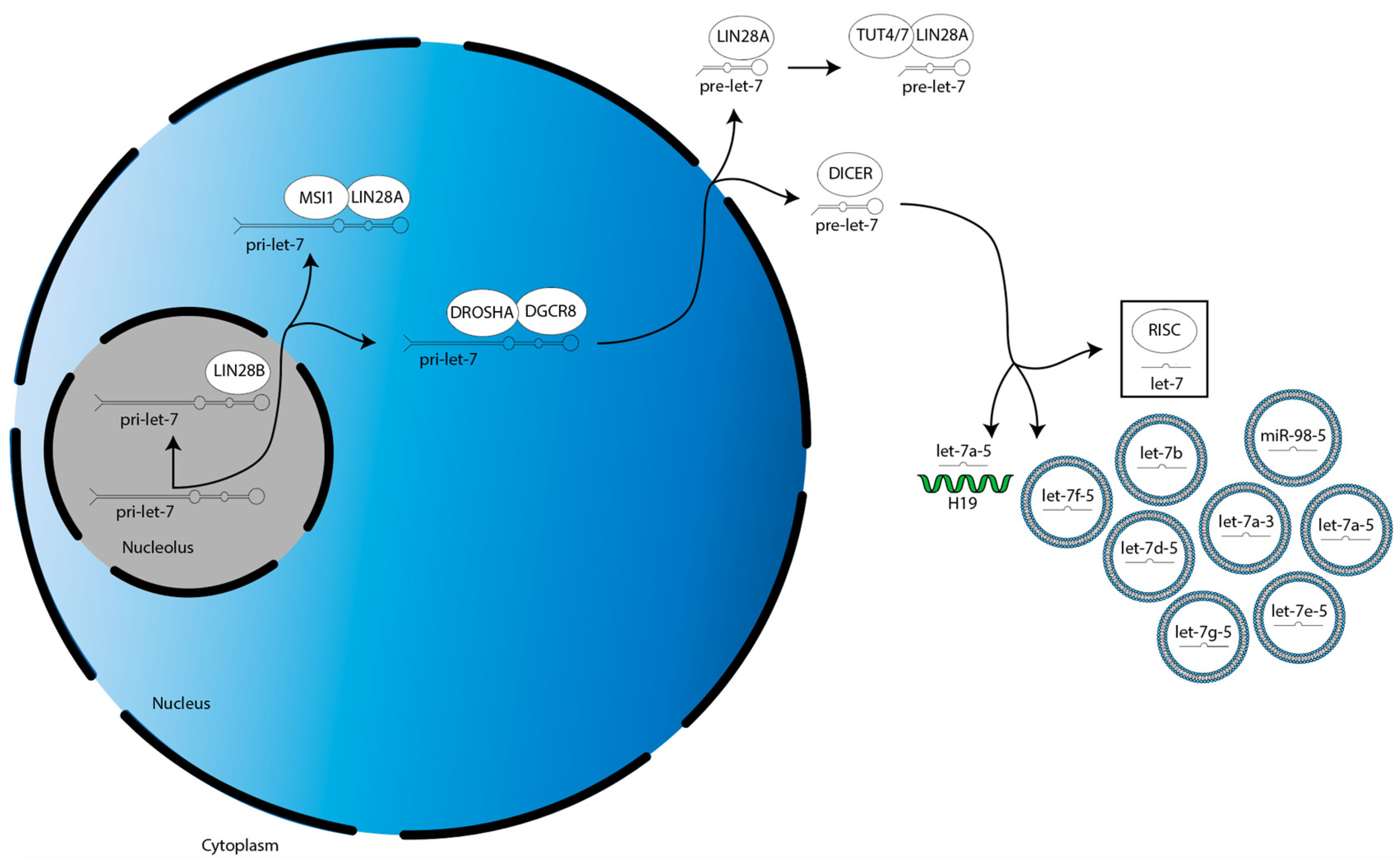{kind=link}
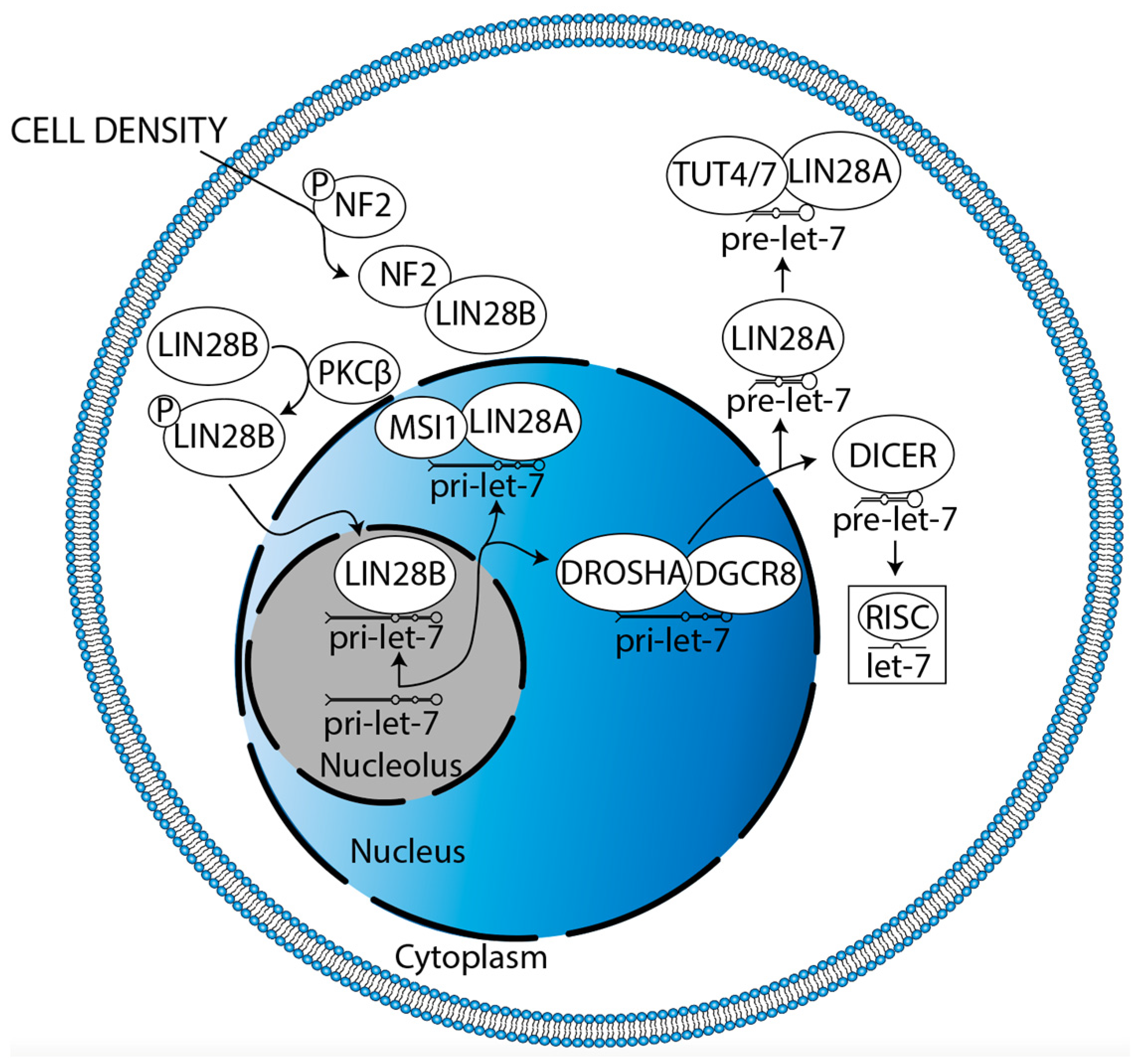{kind=link}
Table 1.
A review of current knowledge on LIN28A/B and let-7 family members in glioma. LIN28A/B are overexpressed in glioma while let-7 family members are underexpressed and mediate the translational repression of a plethora of oncogene mRNAs. ND, no data.
Table 1.
A review of current knowledge on LIN28A/B and let-7 family members in glioma. LIN28A/B are overexpressed in glioma while let-7 family members are underexpressed and mediate the translational repression of a plethora of oncogene mRNAs. ND, no data.
| Species | Expression in Glioma Samples | Expression in Glioma Models | Clinical Correlations | Targets in Glioma | References |
|---|---|---|---|---|---|
| LIN28A | Increased | Increased | Higher grade, poor survival | ND | [32,53,54] |
| LIN28B | Increased | Increased | ND | ND | [13,15,16,17] |
| let-7a | Decreased | Decreased | Lower grade, better survival | Bcl-w, K-RAS, SULF2, OSMR, STAT3 | [55,56,57,58,59,60,61] |
| let-7b | Decreased | Decreased | Lower grade, better survival | IKBKE, E2F2, CCNA2, CCNB2, PLK1, AURKA, PBX3, CCND1 | [58,62,63,64,65,66,67] |
| let-7c | ND | Decreased | ND | ND | [56] |
| let-7d | Decreased | Decreased | ND | STAT3 | [57] |
| let-7e | ND | ND | Lower grade, better survival | ND | [58] |
| let-7f | Decreased | Decreased | Lower grade, better survival | STAT3, CCND1, CCNE1, Bcl-2, MMP-2, MMP-9, periostin | [57,58,68] |
| let-7g | Decreased | Decreased | Lower grade, better survival | pan-RAS, N-RAS, K-RAS | [56,58,69] |
| let-7i | Decreased | Decreased | Lower grade | IKBKE | [63] |
| miR-98 | Decreased | Decreased | Lower grade, better survival | IKBKE, HMGA2, PBX3 | [56,58,70,71,72,73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Knowles, T.; Huang, T.; Qi, J.; An, S.; Burket, N.; Cooper, S.; Nazarian, J.; Saratsis, A.M. LIN28B and Let-7 in Diffuse Midline Glioma: A Review. Cancers 2023, 15, 3241. https://doi.org/10.3390/cancers15123241
AMA Style
Knowles T, Huang T, Qi J, An S, Burket N, Cooper S, Nazarian J, Saratsis AM. LIN28B and Let-7 in Diffuse Midline Glioma: A Review. Cancers. 2023; 15(12):3241. https://doi.org/10.3390/cancers15123241
Chicago/Turabian StyleKnowles, Truman, Tina Huang, Jin Qi, Shejuan An, Noah Burket, Scott Cooper, Javad Nazarian, and Amanda M. Saratsis. 2023. "LIN28B and Let-7 in Diffuse Midline Glioma: A Review" Cancers 15, no. 12: 3241. https://doi.org/10.3390/cancers15123241
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.