
Evaluation of Colon-Specific Plasma Nanovesicles as New Markers of Colorectal Cancer

 , ,
, ,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Reagents and Antibodies
2.3. Extracellular Nanovesicle (ENV) Isolation
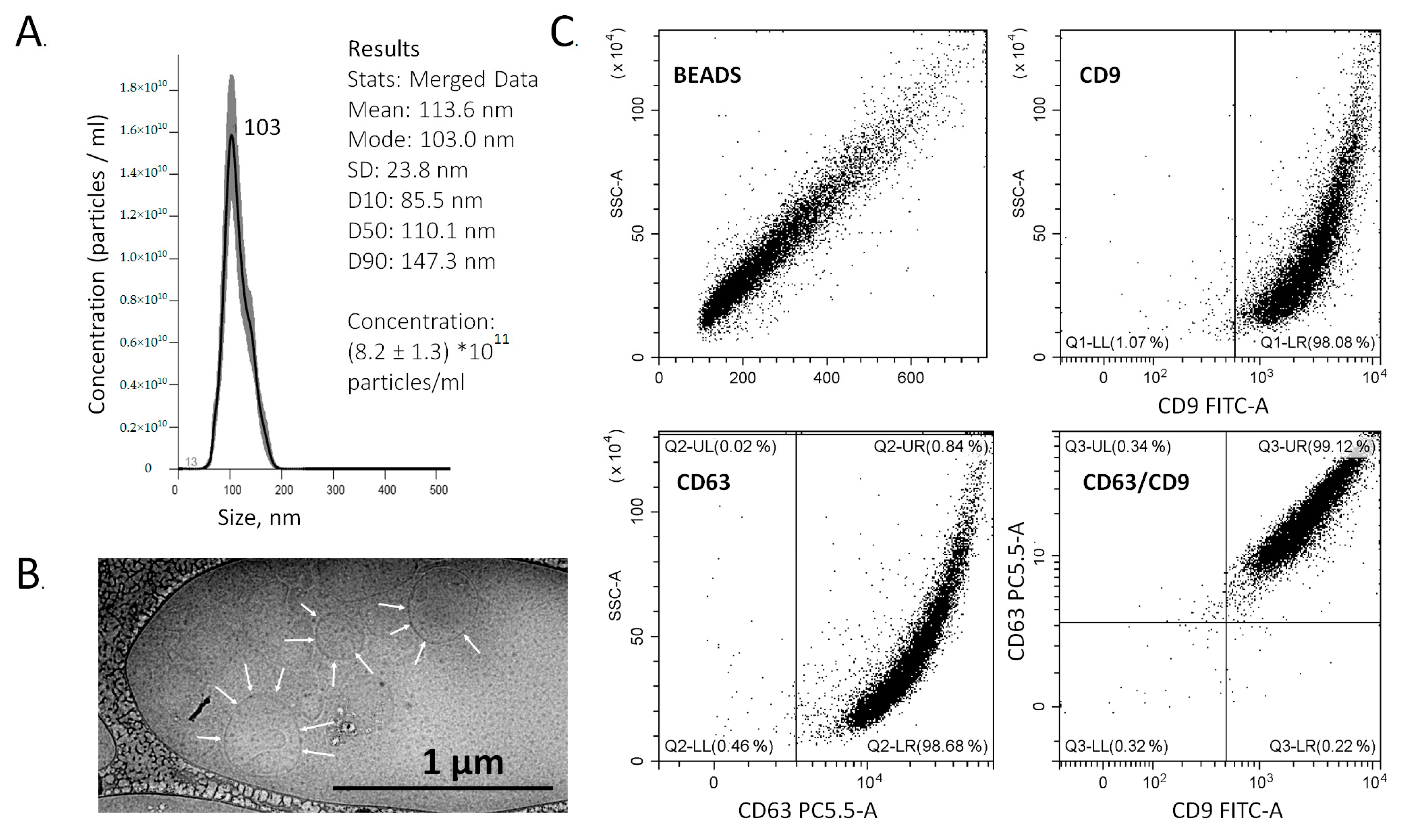
2.4. Nanoparticles Tracking Analysis (NTA)
2.5. Transmission Cryo-Electron Microscopy (Cryo-TEM)
2.6. Analysis of Total ENVs Population by Flow Cytometry
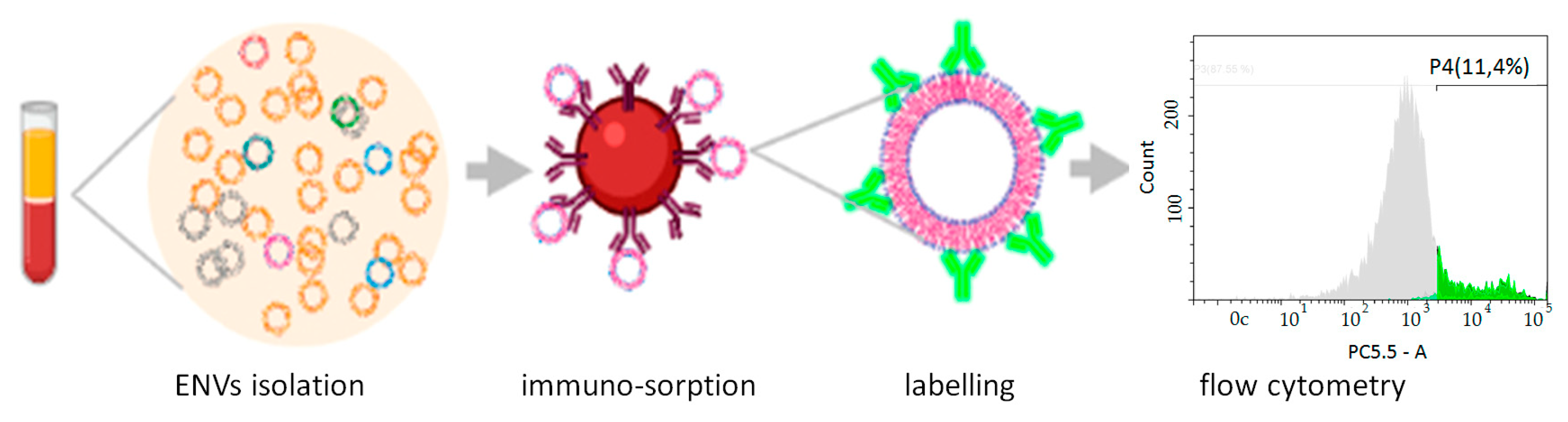
2.7. Preparation of Immune-SPMP and Analysis of ENVs Isolated by Immunosorption
2.8. EV-Track
2.9. Statistical Data Analysis
3. Results
3.1. Isolation and Analysis of the Total ENV Population
3.2. Selection of Potential Surface Markers of Colon Epithelium-Derived ENVs
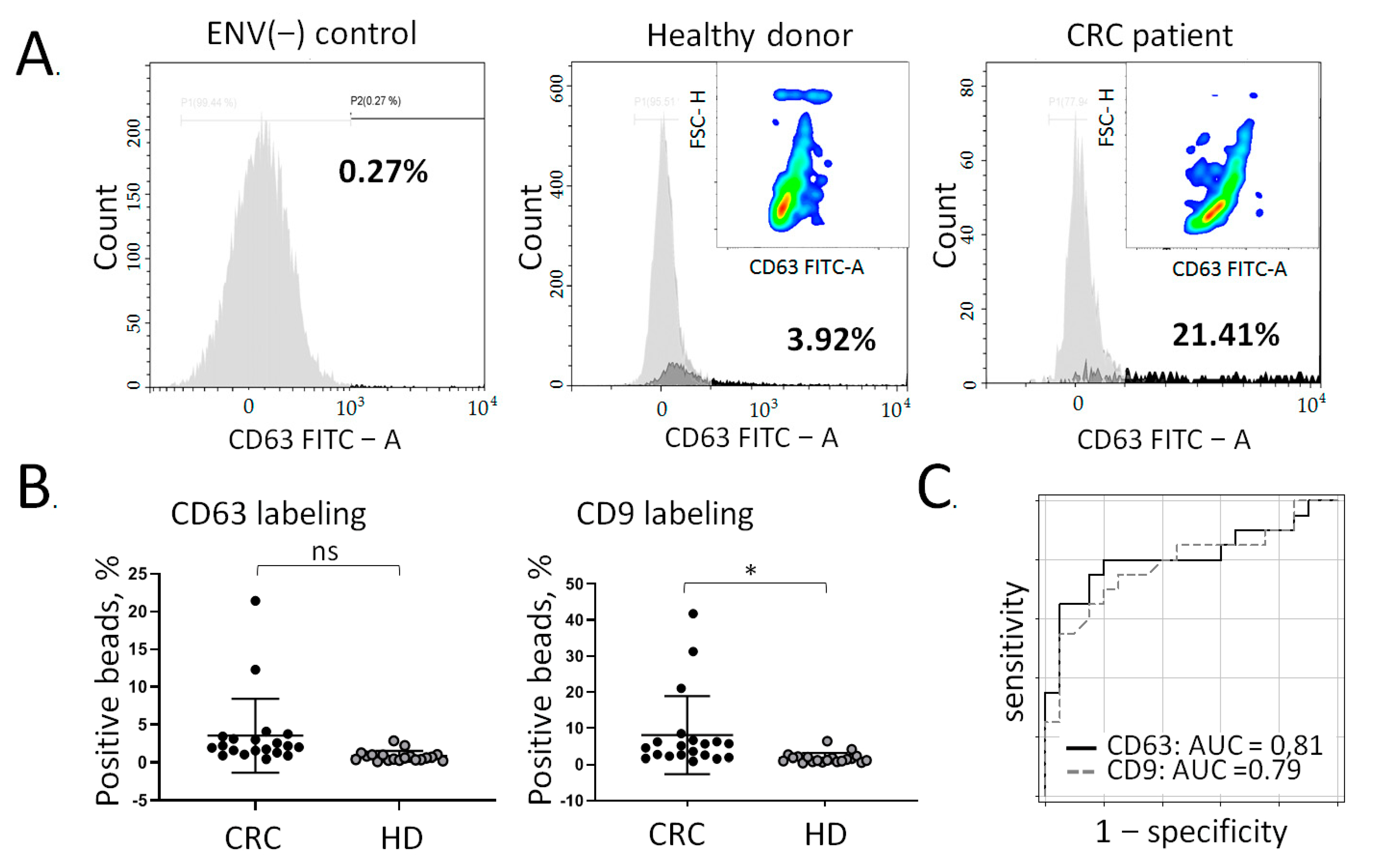
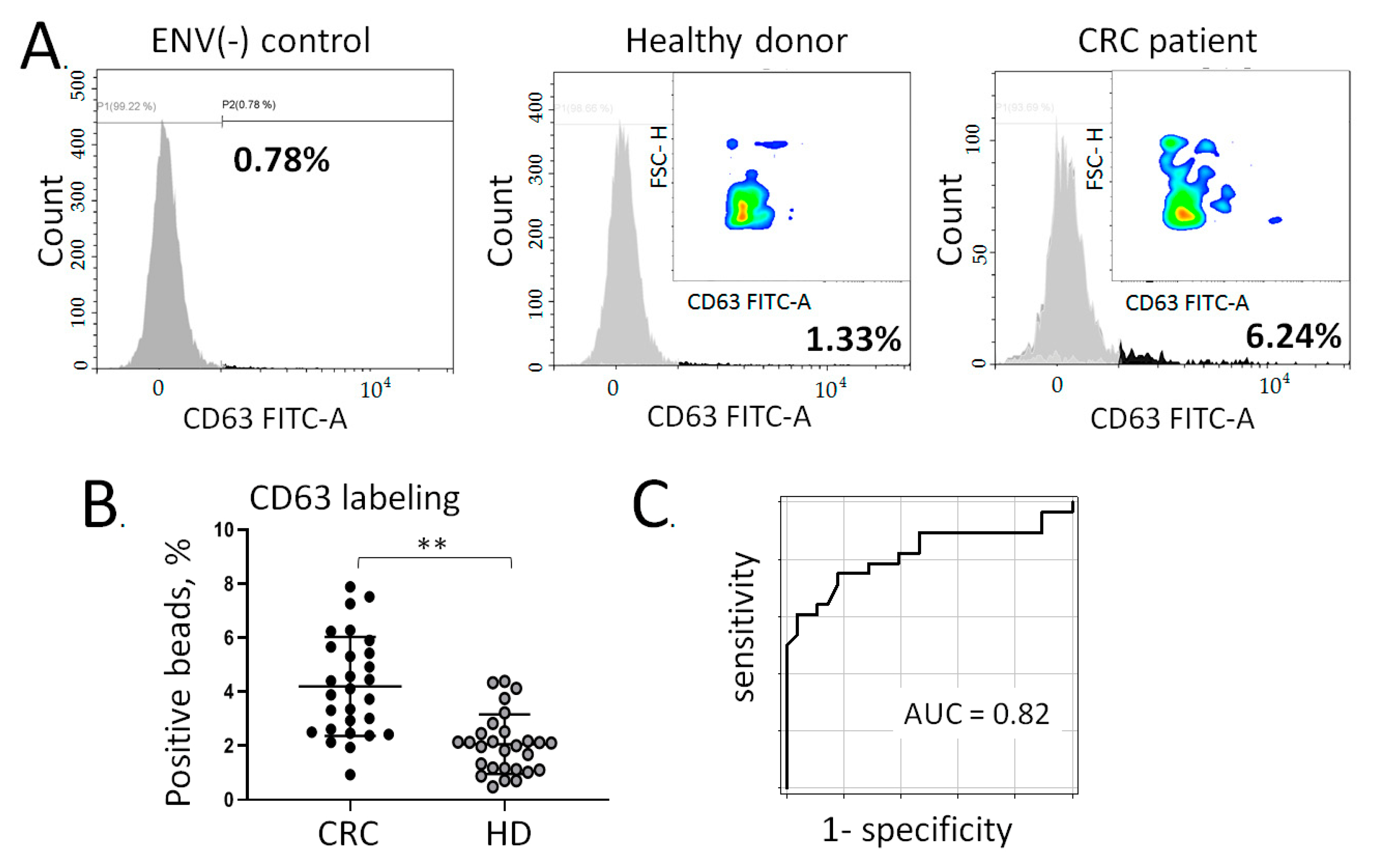
3.3. Quantification of Colon-Specific ENVs in Plasma of CRC Patients
3.4. Multiplex Assessment of Colon-Specific ENVs in Plasma of CRC Patient
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Tschuschke, M.; Kocherova, I.; Bryja, A.; Mozdziak, P.; Angelova Volponi, A.; Janowicz, K.; Sibiak, R.; Piotrowska-Kempisty, H.; Iżycki, D.; Bukowska, D.; et al. Inclusion Biogenesis, Methods of Isolation and Clinical Application of Human Cellular Exosomes. J. Clin. Med. 2020, 9, 436. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, E.M.; Schultz, S.W.; Schink, K.O.; Pedersen, N.M.; Naehse, V.; Carlson, A.; Brech, A.; Stenmark, H.; Raiborg, C. Concerted ESCRT and clathrin recruitment waves define the timing and morphology of intraluminal vesicle formation. Nat. Commun. 2018, 9, 2932. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, O.; Teis, D. The ESCRT machinery. Curr. Biol. 2012, 22, R116–R120. [Google Scholar] [CrossRef] [Green Version]
- Trajkovic, K.; Hsu, C.; Chiantia, S.; Rajendran, L.; Wenzel, D.; Wieland, F.; Simons, M. Ceramide Triggers Budding of Exosome Vesicles into Multivesicular Endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Van Niel, G.; Charrin, S.; Simoes, S.; Romao, M.; Rochin, L.; Saftig, P.; Marks, M.S.; Rubinstein, E.; Raposo, G. The Tetraspanin CD63 Regulates ESCRT-Independent and -Dependent Endosomal Sorting during Melanogenesis. Dev. Cell 2011, 21, 708–721. [Google Scholar] [CrossRef] [Green Version]
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkach, M.; Kowal, J.; Théry, C. Why the need and how to approach the functional diversity of extracellular vesicles. Phil. Trans. R. Soc. B 2017, 373, 20160479. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef]
- Jakobsen, K.R.; Paulsen, B.S.; Bæk, R.; Varming, K.; Sorensen, B.S.; Jørgensen, M.M. Exosomal proteins as potential diagnostic markers in advanced non-small cell lung carcinoma. J. Extracell. Vesicles 2015, 4, 26659. [Google Scholar] [CrossRef]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Nuh Rahbari, N.; et al. Glypican-1 Identifies Cancer Exosomes and Detects Early Pancreatic Cancer. Nature 2015, 523, 177–182. Available online: http://www.nature.com/articles/nature14581 (accessed on 24 June 2015). [CrossRef] [Green Version]
- Theodoraki, M.-N.; Yerneni, S.S.; Hoffmann, T.K.; Gooding, W.E.; Whiteside, T.L. Clinical Significance of PD-L1 + Exosomes in Plasma of Head and Neck Cancer Patients. Clin. Cancer Res. 2018, 24, 896–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Ludwig, S.; Muller, L.; Hong, C.S.; Kirkwood, J.M.; Ferrone, S.; Whiteside, T.L. Immunoaffinity-based isolation of melanoma cell-derived exosomes from plasma of patients with melanoma. J. Extracell. Vesicles 2018, 7, 1435138. [Google Scholar] [CrossRef]
- Czystowska-Kuzmicz, M.; Whiteside, T.L. The potential role of tumor-derived exosomes in diagnosis, prognosis, and response to therapy in cancer. Expert Opin. Biol. Ther. 2020, 21, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, B.; Yue, S.; Galli, U.; Rana, S.; Gross, W.; Müller, M.; Giese, N.A.; Kalthoff, H.; Becker, T.; Büchler, M.W.; et al. Combined evaluation of a panel of protein and miRNA serum-exosome biomarkers for pancreatic cancer diagnosis increases sensitivity and specificity. Int. J. Cancer 2015, 136, 2616–2627. [Google Scholar] [CrossRef]
- Moon, P.G.; Lee, J.E.; Cho, Y.E.; Lee, S.J.; Jung, J.H.; Chae, Y.S.; Bae, H.-I.; Kim, Y.-B.; Kim, I.-S.; Park, H.; et al. Identification of developmental endothelial locus-1 on circulating extracellular vesicles as a novel biomarker for early breast cancer detection. Clin. Cancer Res. 2016, 22, 1757–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Chae, Y.S.; Jung, J.H.; Kim, I.-S.; Park, H.Y.; Baek, M.-C. Fibronectin on circulating extracellular vesicles as a liquid biopsy to detect breast cancer. Oncotarget 2016, 7, 40189–40199. [Google Scholar] [CrossRef] [Green Version]
- Hannafon, B.N.; Trigoso, Y.D.; Calloway, C.L.; Zhao, Y.D.; Lum, D.H.; Welm, A.L.; Zhao, Z.J.; Blick, K.E.; Dooley, W.C.; Ding, W.Q. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016, 18, 90. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Fujita, Y.; Matsuda, Y.; Arai, T.; Horie, K.; Kameyama, K.; Kato, T.; Masunaga, K.; Kasuya, Y.; Tanaka, M.; et al. Gamma-glutamyltransferase activity in exosomes as a potential marker for prostate cancer. BMC Cancer 2017, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bryant, R.J.; Pawlowski, T.; Catto, J.W.F.; Marsden, G.; Vessella, R.L.; Rhees, B.; Kuslich, C.; Visakorpi, T.; Hamd, F.C. Changes in Circulating microRNA Levels Associated with Prostate Cancer. Br. J. Cancer 2012, 106, 768–774. Available online: http://www.nature.com/articles/bjc2011595 (accessed on 12 January 2012). [CrossRef] [Green Version]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Chenga, H.H.; Arroyoa, J.D.; Mereditha, E.K.; Gallichottea, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. USA 2014, 111, 14888–14893. [Google Scholar] [CrossRef] [Green Version]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Srivastava, S.K.; Khushman, M.; Singh, S.; Singh, A.P. Comparative Analysis of exosome Isolation Methods Using Culture Supernatant for Optimum Yield, Purity and Downstream Applications. Sci. Rep. 2019, 9, 5335. Available online: http://www.nature.com/articles/s41598-019-41800-2 (accessed on 29 March 2019). [CrossRef] [Green Version]
- Shtam, T.; Evtushenko, V.; Samsonov, R.; Zabrodskaya, Y.; Kamyshinsky, R.; Zabegina, L.; Verlov, N.; Burdakov, V.; Garaeva, L.; Slyusarenko, M.; et al. Evaluation of Immune and Chemical Precipitation Methods for Plasma Exosome Isolation. PLoS ONE 2020, 15, e0242732. Available online: http://www.ncbi.nlm.nih.gov/pubmed/33232386 (accessed on 24 November 2020). [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Araghi, M.; Soerjomataram, I.; Bardot, A.; Ferlay, J.; Cabasag, C.J.; Morrison, D.S.; Prithwish De, P.; Hanna Tervonen, H.; Walsh, P.M.; Bucher, O.; et al. Changes in Colorectal Cancer Incidence in Seven High-Income Countries: A Population-Based Study. Lancet Gastroenterol. Hepatol. 2019, 4, 511–518. Available online: https://linkinghub.elsevier.com/retrieve/pii/S2468125319301475 (accessed on 16 May 2019). [CrossRef]
- Ogata-Kawata, H.; Izumiya, M.; Kurioka, D.; Honma, Y.; Yamada, Y.; Furuta, K.; Gunji, T.; Ohta, H.; Okamoto, H.; Sonoda, H.; et al. Circulating Exosomal microRNAs as Biomarkers of Colon Cancer. PLoS ONE 2014, 9, e92921. [Google Scholar] [CrossRef] [PubMed]
- Joyce, D.P.; Kerin, M.J.; Dwyer, R.M. Exosome-encapsulated microRNAs as circulating biomarkers for breast cancer. Int. J. Cancer 2016, 139, 1443–1448. [Google Scholar] [CrossRef]
- Yan, S.; Han, B.; Gao, S.; Wang, X.; Wang, Z.; Wang, F.; Jianjun Zhang, J.; Xu, D.; Beicheng Sun, B. Exosome-encapsulated microRNAs as circulating biomarkers for colorectal cancer. Oncotarget 2017, 8, 60149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yan, F.; Zhao, Q.; Zhan, F.; Wang, R.; Wang, L.; Yanqiao Zhang, Y.; Huang, X. Circulating Exosomal miR-125a-3p as a Novel Biomarker for Early-Stage Colon Cancer. Sci. Rep. 2017, 7, 4150. Available online: http://www.nature.com/articles/s41598-017-04386-1 (accessed on 23 June 2017). [CrossRef] [Green Version]
- Yan, S.; Jiang, Y.; Liang, C.; Cheng, M.; Jin, C.; Duan, Q.; Xu, D.; Yang, L.; Zhang, X.; Ren, B.; et al. Exosomal miR-6803-5p as potential diagnostic and prognostic marker in colorectal cancer. J. Cell Biochem. 2018, 119, 4113–4119. [Google Scholar] [CrossRef] [PubMed]
- Karimi, N.; Ali Hosseinpour Feizi, M.; Safaralizadeh, R.; Hashemzadeh, S.; Baradaran, B.; Shokouhi, B.; Shahram, T. Serum Overexpression of miR-301a and miR-23a in Patients with Colorectal Cancer. J. Chin. Med. Assoc. 2019, 82, 215–220. [Google Scholar] [CrossRef]
- Min, L.; Zhu, S.; Chen, L.; Liu, X.; Wei, R.; Zhao, L.; Yang, Y.; Zhang, Z.; Kong, G.; Li, P.; et al. Evaluation of circulating small extracellular vesicles derived miRNAs as biomarkers of early colon cancer: A comparison with plasma total miRNAs. J. Extracell. Vesicles 2019, 8, 1643670. [Google Scholar] [CrossRef] [Green Version]
- Samsonov, R.B.; Tarasov, M.A.; Burdakov, V.S.; Shtam, T.A.; Guljaev, A.M.; Tkachenko, O.B.; Rybakov, E.G.; Filatov, M.V.; Aigner, A.; Malek, A.V. Diagnostic Value of Exosomal miRNA for Colorectal Cancer. Koloproktologia 2018, 2, 25–31. Available online: https://gnck.elpub.ru/jour/article/view/1119 (accessed on 30 May 2018). [CrossRef] [Green Version]
- Liu, T.; Zhang, X.; Gao, S.; Jing, F.; Yang, Y.; Du, L.; Zheng, G.; Li, P.; Li, C.; Wang, C. Exosomal long noncoding RNA CRNDE-h as a novel serum-based biomarker for diagnosis and prognosis of colorectal cancer. Oncotarget 2016, 7, 85551–85563. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Lin, W.; Qi, P.; Xu, M.; Wu, X.; Ni, S.; Huan, D.; Weng, W.; Tan, C.; Sheng, W.; et al. Circulating Long RNAs in Serum Extracellular Vesicles: Their Characterization and Potential Application as Biomarkers for Diagnosis of Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. 2016, 25, 1158–1166. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Duan, W.; Yan, S.; Xie, Y.; Wang, C. Circulating Long Non-Coding RNA Colon Cancer-Associated Transcript 2 Protected by Exosome as a Potential Biomarker for Colorectal Cancer. Biomed. Pharmacother. 2019, 113, 108758. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0753332218382969 (accessed on 4 March 2019). [CrossRef] [PubMed]
- Hu, D.; Zhan, Y.; Zhu, K.; Bai, M.; Han, J.; Si, Y.; Zhang, H.; Kong, D. Plasma Exosomal Long Non-Coding RNAs Serve as Biomarkers for Early Detection of Colorectal Cancer. Cell Physiol. Biochem. 2018, 51, 2704–2715. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, Y.; Xu, L.; Zhan, S.; Xiao, Y.; Gao, Y.; Wub, B.; Gea, W. Protein content and functional characteristics of serum-purified exosomes from patients with colorectal cancer revealed by quantitative proteomics. Int. J. Cancer 2017, 140, 900–913. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Guo, X.; Zhou, L.; Jia, Z.; Peng, Z.; Tang, Y.; Liu, W.; Zhu, B.; Wang, L.; et al. GPC1 exosome and its regulatory miRNAs are specific markers for the detection and target therapy of colorectal cancer. J. Cell Mol. Med. 2017, 21, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ma, L.; Gong, M.; Su, G.; Zhu, S.; Zhang, W.; Wang, S.; Li, Z.; Chen, C.; Li, L.; et al. Protein Profiling and Sizing of Extracellular Vesicles from Colorectal Cancer Patients via Flow Cytometry. ACS Nano 2018, 12, 671–680. [Google Scholar] [CrossRef]
- Scavo, M.P.; Cigliano, A.; Depalo, N.; Fanizza, E.; Bianco, M.G.; Denora, N.; Laquintana, V.; Curri, M.L.; Lorusso, D.; Lotesoriere, C.; et al. Frizzled-10 Extracellular Vesicles Plasma Concentration Is Associated with Tumoral Progression in Patients with Colorectal and Gastric Cancer. J. Oncol. 2019, 2019, 2715968. Available online: https://www.hindawi.com/journals/jo/2019/2715968/ (accessed on 2 June 2019). [CrossRef] [Green Version]
- Xiao, Y.; Zhong, J.; Zhong, B.; Huang, J.; Jiang, L.; Jiang, Y.; Jichao Sun, J.; Dai, L.; Yang, C.; Li, Z.; et al. Exosomes as potential sources of biomarkers in colorectal cancer. Cancer Lett. 2020, 476, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Nazarova, I.; Nikiforova, N.; Sidina, E.; Slyusarenko, M.; Kotova, Z.; Semiglazova, T.; Sluzhev, M.; Semiglazov, V.; Gogolev, A.; Artemyeva, A.; et al. Colorectal Cancer Diagnostics via Detection of Tissue-Specific Extracellular Nano-Vesicles. Koloproktologia 2020, 19, 32–56. Available online: https://www.ruproctology.com/jour/article/view/1607 (accessed on 9 December 2020). [CrossRef]
- Slyusarenko, M.; Nikiforova, N.; Sidina, E.; Nazarova, I.; Egorov, V.; Garmay, Y.; Merdalimova, A.; Yevlampieva, N.; Gorin, D.; Malek, A. Formation and Evaluation of a Two-Phase Polymer System in Human Plasma as a Method for Extracellular Nanovesicle Isolation. Polymers 2021, 13, 458. Available online: https://www.mdpi.com/2073-4360/13/3/458 (accessed on 31 January 2021). [CrossRef]
- The Human Protein Athlas. Available online: www.proteinatlas.org (accessed on 1 December 2003).
- UniProt Consortium. Available online: www.uniprot.org (accessed on 15 December 2004).
- ExoCarta. Available online: www.exocarta.org (accessed on 29 December 2009).
- Compartments. Available online: www.compartments.jensenlab.org (accessed on 28 December 2014).
- Binder, J.X.; Pletscher-Frankild, S.; Tsafou, K.; Stolte, C.; O’Donoghue, S.I.; Schneider, R.; Jensen, L.J. COMPARTMENTS: Unification and visualization of protein subcellular localization evidence. Database 2014, 2014, bau012. [Google Scholar] [CrossRef] [Green Version]
- Habertheuer, A.; Korutla, L.; Rostami, S.; Reddy, S.; Lal, P.; Naji, A.; Vallabhajosyula, P. Donor tissue-specific exosome profiling enables noninvasive monitoring of acute rejection in mouse allogeneic heart transplantation. J. Thorac. Cardiovasc. Surg. 2018, 155, 2479–2489. [Google Scholar] [CrossRef]
- Hu, R.W.; Korutla, L.; Reddy, S.; Harmon, J.; Zielinski, P.D.; Bueker, A.; Molina, M.; Romano, C.; Margulies, K.; McLean, R.; et al. Circulating Donor Heart Exosome Profiling Enables Noninvasive Detection of Antibody-mediated Rejection. Transplant. Direct 2020, 6, e615. [Google Scholar] [CrossRef]
- Taylor, R.M.; Miller, P.R.; Ebrahimi, P.; Polsky, R.; Baca, J.T. Minimally-invasive, microneedle-array extraction of interstitial fluid for comprehensive biomedical applications: Transcriptomics, proteomics, metabolomics, exosome research, and biomarker identification. Lab. Anim. 2018, 52, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Maia, J.; Batista, S.; Couto, N.; Gregório, A.C.; Bodo, C.; Elzanowska, J.; Moraes, M.C.S.; Costa-Silva, B. Employing Flow Cytometry to Extracellular Vesicles Sample Microvolume Analysis and Quality Control. Front. Cell Dev. Biol. 2020, 8, 593750. [Google Scholar] [CrossRef] [PubMed]
- Nizamudeen, Z.; Markus, R.; Lodge, R.; Parmenter, C.; Platt, M.; Chakrabarti, L.; Sottile, V. Rapid and accurate analysis of stem cell-derived extracellular vesicles with super resolution microscopy and live imaging. BBA Mol. Cell Res. 2018, 1865, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, N.; Chumachenko, M.; Nazarova, I.; Zabegina, L.; Slyusarenko, M.; Sidina, E.; Malek, A. CM-Dil Staining and SEC of Plasma as an Approach to Increase Sensitivity of Extracellular Nanovesicles Quantification by Bead-Assisted Flow Cytometry. Membranes 2021, 11, 526. [Google Scholar] [CrossRef]
- Di, H.; Mi, Z.; Sun, Y.; Liu, X.; Liu, X.; Li, A.; Jiang, Y.; Gao, H.; Rong, P.; Liu, D. Nanozyme-assisted sensitive profiling of exosomal proteins for rapid cancer diagnosis. Theranostics 2020, 10, 9303–9314. [Google Scholar] [CrossRef]
- Wang, Q.; Zou, L.; Yang, X.; Liu, X.; Nie, W.; Zheng, Y.; Cheng, Q.; Wang, K. Direct quantification of cancerous exosomes via surface plasmon resonance with dual gold nanoparticle-assisted signal amplification. Biosens. Bioelectron. 2019, 135, 129–136. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Donors (HD) | CRC Patients, IIIb–IV | CRC Patients, II–IIIa | |
|---|---|---|---|
| Number | 50 | 20 | 28 |
| Age, mean (SD) | 50 (6.3) | 53 (9.3) | 51 (5.5) |
| Gender, (m/f) | 36/14 | 13/17 | 15/13 |
| Protein Name | UniProt ID | Colon-Specific Expression 1 | Surface Membrane Localization 2 | Exo Carta ID | CD9 | CD63 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CRC Patients | Health Donors | CRC vs. HD | AUC | CRC Patients | Health Donors | CRC vs. HD | AUC | |||||
| CLRN3 | Q8NCR9 | 27.7 | *** | 119467 | 7.71 | 2.8 | ns | 0.76 | 3.2 | 1.4 | ** | 0.78 |
| GPA33 | Q99795 | 46.9 | ***** | 10223 | 4.8 | 4.1 | ns | 0.54 | 2.1 | 1.5 | ns | 0.67 |
| GCNT3 | O95395 | 37.6 | **** | 9245 | 6.4 | 2.5 | ns | 0.64 | 2.8 | 1.0 | * | 0.71 |
| PIGY | Q3MUY2 | 10.6 | ***** | 84992 | 7.2 | 3.3 | ns | 0.66 | 3.2 | 1.3 | ** | 0.78 |
| REG4 | Q9BYZ8 | 55.5 | *** | 83998 | 7.5 | 2.2 | * | 0.78 | 3.0 | 1.0 | * | 0.74 |
| MEP1A | Q16819 | 100.5 | ***** | 4224 | 8.1 | 2.8 | * | 0.79 | 3.6 | 1.5 | ns | 0.81 |
| LGALS4 | P56470 | 190.6 | **** | 3960 | 5.6 | 2.4 | ns | 0.69 | 2.2 | 1.2 | ns | 0.71 |
| Mucin 12 | Q9UKN1 | 25.2 | ***** | - | 7.3 | 2.7 | * | 0.75 | 3.0 | 1.2 | * | 0.78 |
| PDCD6IP | Q8WUM4 | 36.8 | ***** | 10015 | 5.7 | 4.1 | ns | 0.6 | 2.5 | 1.7 | ns | 0.66 |
| DHRS11 | Q6UWP2 | 38.7 | ** | 360583 | 7.7 | 3.2 | ns | 0.63 | 2.7 | 1.4 | ns | 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarova, I.; Slyusarenko, M.; Sidina, E.; Nikiforova, N.; Semiglazov, V.; Semiglazova, T.; Aigner, A.; Rybakov, E.; Malek, A. Evaluation of Colon-Specific Plasma Nanovesicles as New Markers of Colorectal Cancer. Cancers 2021, 13, 3905. https://doi.org/10.3390/cancers13153905
Nazarova I, Slyusarenko M, Sidina E, Nikiforova N, Semiglazov V, Semiglazova T, Aigner A, Rybakov E, Malek A. Evaluation of Colon-Specific Plasma Nanovesicles as New Markers of Colorectal Cancer. Cancers. 2021; 13(15):3905. https://doi.org/10.3390/cancers13153905
Chicago/Turabian StyleNazarova, Inga, Maria Slyusarenko, Elena Sidina, Nadezhda Nikiforova, Vladislav Semiglazov, Tatiana Semiglazova, Achim Aigner, Evgeny Rybakov, and Anastasia Malek. 2021. "Evaluation of Colon-Specific Plasma Nanovesicles as New Markers of Colorectal Cancer" Cancers 13, no. 15: 3905. https://doi.org/10.3390/cancers13153905