
The Role of Tumor-Associated Macrophages in Hematologic Malignancies



Unit of Hematology, Azienda Ospedaliera Universitaria Senese, University of Siena, 53100 Siena, Italy
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(14), 3597; https://doi.org/10.3390/cancers13143597
Submission received: 29 June 2021
/
Revised: 11 July 2021
/
Accepted: 15 July 2021
/
Published: 18 July 2021
(This article belongs to the Special Issue Targeting Innate Immunity to Treat Cancer)
Abstract
:Simple Summary
Tumor-associated macrophages (TAM) represent a leading component of the tumor microenvironment in hematologic malignancies. TAM could display antitumor activity or, conversely, could contribute to tumor growth and survival, depending on their polarization. TAM are polarized towards form M1, with a pro-inflammatory phenotype and an antineoplastic activity, or M2, with an alternately activated phenotype, associated with a poor outcome in patients presenting with leukemia, lymphoma or multiple myeloma. The molecular mechanisms of TAM in different types of hematologic malignancies are different due to the peculiar microenvironment of each disease. TAM could contribute to tumor progression, reduced apoptosis and angiogenesis; a different TAM polarization could explain a reduced treatment response in patients with a similar disease subtype. The aim of our review is to better define the role of TAM in patients with leukemia, lymphoma or multiple myeloma. Finally, we would like to focus on TAM as a possible target for antineoplastic therapy.
Abstract
The tumor microenvironment includes dendritic cells, T-cytotoxic, T-helper, reactive B-lymphoid cells and macrophages; these reactive cells could interplay with malignant cells and promote tumor growth and survival. Among its cellular components, tumor-associated macrophages (TAM) represent a component of the innate immune system and play an important role, especially in hematologic malignancies. Depending on the stimuli that trigger their activation, TAM are polarized towards form M1, contributing to antitumor responses, or M2, associated with tumor progression. Many studies demonstrated a correlation between TAM, disease progression and the patient’s outcome in lymphoproliferative neoplasms, such as Hodgkin lymphoma (HL) and non-Hodgkin lymphoma (NHL), even if with conflicting results. A critical hurdle to overcome is surely represented by the heterogeneity in the choice of the optimal markers and methods used for TAM analysis (gene-expression profile vs. immunohistochemistry, CD163vs. CD68vs. CD163/CD68 double-positive cells). TAM have been recently linked to the development and progression of multiple myeloma and leukemia, with a critical role in the homing of malignant cells, drug resistance, immune suppression and angiogenesis. As such, this review will summarize the role of TAM in different hematologic malignancies, focusing on the complex interplay between TAM and tumor cells, the prognostic value of TAM and the possible TAM-targeted therapeutic strategies.
1. Introduction
The innate immune system defends the host from infections and non-self antigens in a non-specific manner; thus, it could be involved in the identification and destruction of neoplastic cells [1,2,3]. It includes natural physical barriers, the complement system, natural killer (NK) cells, mast cells, dendritic cells, neutrophils, eosinophils, basophils, monocytes and macrophages. Macrophages represent a leading component of the innate immune response, are involved in the inflammation, tissue repair and phagocytosis of pathogens and can release pro-inflammatory cytokines and chemokines [4,5,6]. Phagocytosis is a mechanism leading to the destruction of foreign antigens; macrophages can secrete cytokines that promote the recruitment of leukocytes from the circulating blood to the sites of inflammation. The antitumor effect of macrophages includes the killing of cancer cells through both their own Fc receptors that bind antibodies on the surface of cancer cells and the production of nitric oxide (NO) and tumor necrosis factor (TNF). After phagocytosis, macrophages can expose tumor antigens on their surface, thus permitting the antigen recognition by T-lymphoid cells [4,5,6].
Since the 1990s, immunohistochemical (IHC) analyses have shown many macrophages in the tumor microenvironment (TME) [7,8]. This infiltration could be induced by tumor cells by secreting molecules such as chemokine ligands with a C-C motif (CCL) or C-X3-C motif (CXCL) and granulocyte–macrophage colony-stimulating factor (GM-CSF) [9,10,11,12,13]. A high number of macrophages has been frequently observed in both solid and hematological malignancies, and it was first considered as a mechanism involved in anticancer surveillance [14,15]. However, several reports demonstrated that the so-called tumor-associated macrophages (TAM) could conversely contribute to oncogenesis and neoplastic progression as “bad guys”, by promoting genetic instability and angiogenesis, while reducing the immune response and apoptosis [4,16,17,18].
TAM are characterized by a wide morphological, phenotypical and functional heterogeneity [4]. Complex interactions have been reported between reactive cells, including TAM and malignant cells in patients with leukemia, Hodgkin lymphoma (HL), non-Hodgkin lymphoma (NHL) or multiple myeloma (MM); as a final result, TAM could acquire an immunosuppressive and oncogenic phenotype [19]. Macrophages are characterized by remarkable plasticity and were initially divided into six subtypes (immunosuppressive, angiogenic, metastasis-associated, invasive, activated and perivascular), depending on the stimuli that trigger their activation [20]. However, in the currently accepted classification, even if oversimplified, TAM have been divided into M1 (classically activated) and M2 (alternatively activated), probably representing the 2 extremities of a continuous spectrum [21,22,23]. Due to this different polarization, which occurs in the tissue, M1 TAM could provoke a Th-1 response and play an antineoplastic effect leading to tumor suppression, while M2 TAM, with a low antigen-presenting capacity, could promote tumor growth and survival by inducing angiogenesis and immunosuppression [24]. In particular, as shown in Figure 1 and Figure 2, the M1 subtype, triggered by GM-CSF, bacterial products and interferon-γ, could secrete pro-inflammatory molecules, such as interleukin (IL)-1, IL-6, IL-12, IL-23, TNF-α, NO, CXCL9, CXCL10, CXCL11 and reactive oxygen species; conversely, the M2 subtype is activated by IL-4, IL-10, IL-13 and express anti-inflammatory molecules, such as IL-10, tumor growth factor (TGF-β), CCL17, CCL18, CCL22, mannose receptor C type 1 (CD206), class A scavenger receptor (CD204) and hemoglobin scavenger receptor (CD163), which are currently used as M2-associated markers [21,22,23,24].
In order to exert their action within the tumor microenvironment, TAM have to be recruited and then polarized into a tumor-promoting phenotype. TAM recruitment is mediated by chemokines, such as CCL2, CCL5, CCL7 and CXCL1, macrophages(M) CSF and vascular endothelial growth factor (VEGF) [25,26,27,28,29,30,31]. At the tumor site, a complex interplay occurs between TAM, malignant cells and stromal cells, in which stroma-derived and tumor-derived factors, such as IL4, IL-10, IL-13 and TGF-β, cause a shift towards M2 polarization [32,33,34,35]. Once recruited and polarized, TAM could contribute to tumor progression, metastasis and chemo resistance by producing matrix remodeling molecules and impairing innate or adaptive immune cells functionality [36,37,38].
This review summarizes the publications associated to the role of TAM in hematological malignancies, including HL, NHL, MM and leukemia, with a particular focus on M2, which are mainly related to tumor progression and treatment response.
2. Materials and Methods
We performed a computerized search in MEDLINE in order to find full-text publications, written in English, focusing on relationship between TAM and hematologic malignancies. The key terms to search included “tumor-associated macrophages OR TAM OR M1 macrophages OR M2 macrophages OR CD68 OR CD163 OR CD204 OR CD206 AND hematologic malignancies OR Hodgkin lymphoma OR Hodgkin disease OR non-Hodgkin lymphoma OR multiple myeloma OR acute myeloid leukemia OR acute lymphoblastic leukemia OR acute T-cell lymphoma/leukemia OR chronic lymphocytic leukemia”. We have also searched in the reference list of selected articles in order to perform a more comprehensive research. We included retrospective studies but excluded conference abstracts and case reports. For each study, we have extrapolated: disease, methods of TAM determination, TAM markers, the antibodies and the choice of cut-off for high vs. low infiltration, patient number, treatment schedule and the relationship between TAM and disease outcome, especially progression-free survival (PFS) and overall survival (OS).
3. TAM Role in Leukemia
Due to the growing evidence that leukemic cells interact with bone marrow (BM) reactive cells, including macrophages, the concept of leukemia-associated macrophages has been formulated [39,40]. A possible correlation between an immunosuppressive microenvironment and leukemia progression has been hypothesized for acute lymphoblastic leukemia (ALL), adult T-cell leukemia/lymphoma (ATLL), acute myeloid leukemia (AML) and chronic lymphocytic leukemia (CLL) [41]. The pathophysiology could be linked at least in part to an immune escape, due to a reduced ability of macrophages to recognize antigens and to perform their phagocytosis [39,40,41,42,43].
3.1. Acute Lymphoblastic Leukemia
ALL is a rare and aggressive disease, often characterized by a poor outcome despite high-dose therapy and allogeneic stem cell transplantation (allo-SCT) [44]. Even if there has been an undeniable therapeutic advance in the last few years, OS remains significantly reduced compared to the age-matched general population. A higher white blood cells (WBC) count at diagnosis (reflecting an elevated tumor burden), an older age and comorbidities (related to a reduced treatment tolerability) are recognized as prognostic factors for a lower complete remission (CR) and shorter duration of response (DOR) [44]. However, many other immunophenotypic, cytogenetic and molecular disease-related factors have been studied and there is a growing interest about a possible prognostic relevance of an immunosuppressive microenvironment [45,46,47,48].
3.1.1. Pre-Clinical Studies
In a mouse model of T-ALL with Notch1 overexpression, a different gene expression profile (GEP) between BM and splenic macrophages was observed [49]. After coculture, splenic macrophages caused greater leukemic cells stimulation compared to BM macrophages [49]. Moreover, ALL cells could release the bone morphogenetic protein 4 (BMP4), as a mechanism to induce the M2 polarization of TAM, which are in turn capable of producing CCL2, IL-6 and IL-10 [50]. As discovered recently, the deletion of the stromal interaction molecule (Stim) 1 and 2 in ALL cells could reduce TAM infiltration and favor a shift towards an M1 polarization through IFN-γ [51].
The concept of a BM niche as a sanctuary supporting ALL progression was recently shared, in which TAM acquire immunosuppressive properties from the leukemic cells, such as a reduction in their phagocytic activity by the interaction of signal regulatory protein (SIRP)α with CD47 molecule located on the leukemic cells surface [52].
3.1.2. Clinical Studies
The results obtained from pre-clinical studies helped to develop clinical studies in ALL patients; however, the hypothesized prognostic role of TAM in ALL has not been demonstrated in clinical studies, even if macrophages count in patients’ samples was increased compared to healthy subjects. In a retrospective study, BM biopsies of 52 B-ALL patients and 14 healthy controls were analyzed [53]. In B-ALL BM, a decreased proportion of M1 TAM and CD27-positive T-cells was reported, while M2 TAM and myeloid-derived suppressor cells (MDSC) were increased. The author suggested the reactive BM microenvironment in ALL cases significantly differs from healthy controls [53]. In another study, 97 BM samples of acute leukemia patients (26/97 with ALL) were compared to 30 with iron-deficiency anemia (IDA), as healthy control group [47]. In leukemic BM, CD68-positive, CD163-positive and CD206-positive macrophages count was significantly increased compared to IDA and significantly decreased after therapy in patients achieving a CR, even if it remained higher than in the control group. If we consider CD68 as a pan-macrophage marker, CD163-positive or CD206-positive/CD68-positive ratio was increased in the BM leukemic samples compared to the control group, further supporting an M2 polarization [47].
3.2. Adult T-Cell Leukemia/Lymphoma
ATLL represents an uncommon neoplasm, linked to the human T-lymphotropic virus type 1 (HTLV-1) and characterized by dismal prognosis [54]. The incidence is higher in Japan, the Caribbean and sub-Saharan Africa, with older age at the onset in cases outside of Japan. Anthracyclines-containing regimens demonstrated limited efficacy, and a significant proportion of patients were refractory or relapsed after first-line therapy [54].
3.2.1. Pre-Clinical Studies
In ATLL lines, a significant cell activation was noted by direct coculture with M2 TAM. The authors showed some soluble factors were implicated in this interaction between M2 TAM and tumor cells, such as TNF-α, C5a and IL-6 [55]. Moreover, a higher CD163 expression was induced by direct TAM–ATLL cells interaction and ATLL cells proliferation was reduced if cocultured with CD163-silenced macrophages [55].
The number of CD204-positive TAM was associated with malignant cells proliferation, measured according to the Ki-67 labeling index, giving a novel insight into ATLL pathophysiology [56].
3.2.2. Clinical Studies
The role of TAM in ATLL showed in pre-clinical studies was confirmed and a significant correlation between CD163-positive TAM and poor prognosis was reported in 58 ATLL patients, in which double-immunostaining demonstrated CD163-positive TAM also expressed a CD68 marker [55]. In the univariate analysis, a higher number of both CD68-positive and CD163-positive cells was significantly associated with a dismal disease outcome; however, in the multivariate analysis, only an elevated CD163 expression confirmed a prognostic correlation [55]. Interestingly, CD206-positive macrophages showed an overlap in ATLL tissues with CD163-positive elements, but not with the CD204-positive population, further reaffirming a remarkable plasticity [56].
3.3. Acute Myeloid Leukemia
AML represents an aggressive malignancy due to the BM clonal expansion of myeloid precursors. Circulating tumor cells (blasts) in the peripheral blood occur together with anemia, neutropenia and thrombocytopenia [57]. Adverse prognostic factors include an advanced age, high WBC count at diagnosis, history of myelodysplastic syndrome (MDS) or chronic myeloproliferative neoplasm (MPN), cytogenetic alterations such as deletion of chromosome 5 and/or 7 and molecular aberrations such as Fms-related receptor tyrosine kinase 3 (FLT3) mutations [57].
3.3.1. Pre-Clinical Studies
Several reports showed TAM could influence AML cells survival and drug resistance. The frequency of BM CD163/CD206 double-positive M2 TAM was analyzed and was increased in AML patients compared to healthy donors [58]. AML cells could polarize BM and splenic TAM towards a pro-leukemic phenotype in mouse models. The growth factor independence 1 (Gfi1), a transcriptional repressor involved in myeloid and lymphoid hematopoiesis, plays a fundamental role in TAM polarization. Gfi-deficient mice showed an increased production of pro-inflammatory molecules, thus Gfi1 could enhance in vitro M2 polarization induced by IL-4 and reduce M1 polarization induced by LPS [58].
Similarly, peritoneal macrophages in mixed lineage leukemia (MLL)-AF9-induced AML mouse models showed a prevalent M2 TAM phenotype. The results of this study suggested the AML microenvironment significantly influenced the morphology, killing and phagocytic function of peritoneal macrophages; peritoneal TAM could acquire an M2 phenotype with different GEP compared to normal macrophages [59]. Consequently, a reprogramming towards an M1 polarization could exert an immunotherapeutic effect against AML. Another study investigated the molecular basis for different TAM subtypes; in leukemia models, BM TAM showed M1 characteristics while splenic TAM had more M2 characteristics [60]. An interferon regulatory factor 7 (IRF7) gene could contribute to M1 TAM polarization through SAPK/JNK pathway activation and showed a higher expression in BM TAM compared to splenic TAM [60].
3.3.2. Clinical Studies
Pre-clinical studies suggested the capability of M2 TAM to influence AML progression and drug resistance, and this was confirmed by clinical data. Due to the fact that AML cells could polarize TAM towards a pro-tumor subtype, it seems necessary to eliminate these TAM subsets by depletion and/or reprogramming towards an antitumor subtype. It could have a prognostic relevance, as shown in an interesting study, in which the clinical impact of TAM in AML patients was investigated using an open database called BloodSpot [60]. A higher expression of CD163 was associated with poor prognosis in human AML, while no correlation with survival was noted for CD68, further suggesting that M2 TAM could be related to a dismal disease outcome, rather than total macrophages count.
In the above-mentioned study including both ALL and AML, CD163/CD206 double-positive TAM had an increased expression in leukemic samples than in healthy donors [47]. The M2 marker CD206 was identified as a novel prognostic factor for AML patients using the algorithm CIBERSORT [43]. The authors confirmed an increased M2 TAM frequency compared to healthy donors;this was correlated with a poor outcome. Specifically, high CD206 expression was associated with reduced event free survival (EFS) and OS in AML cohorts. Interestingly, among 175 AML cases presenting with intermediate-risk cytogenetics, 3-y EFS and OS for patients with low and high CD206 expression were 47% vs. 25% and 56% vs. 32%, respectively (p < 0.001) [43].
Lastly, a low level of monocytic leukemia zincfinger protein (MOZ) was associated with reduced M1 polarization and poor prognosis in AML cases, and miR-223, capable of suppressing M1 polarization, was involved in MOZ regulation [61].
3.4. Chronic Lymphocytic Leukemia
CLL is the most common leukemia subtype in western countries and in elderly patients and is characterized by highly variable clinical course and prognosis. Approximately one-third of CLL patients do not require any therapy and have a normal lifespan. However, a not negligible proportion exhibit aggressive behavior that requires treatment at diagnosis, or develop a clinical progression after an initial indolent phase [62]. Biological variables, such as cytogenetic aberrations and immunoglobulin heavy chain variable region genes (IGHV) mutational status, represent the most important factors to consider before treatment choice [63]. The highest risk group is represented by CLL harboring TP53 mutations and/or del(17p), in which targeted therapy is strongly recommended as first-line regimen [64,65].
3.4.1. Pre-Clinical Studies
The first identification of reactive cells in the CLL microenvironment that could promote CLL survival in vitro was performed more than 20 years ago, with the identification of the so-called nurse-like cells (NLC) [66]. Subsequent investigations showed NLC were CLL-associated macrophages and were therefore considered as TAM [67,68]. NLC present a prevalent M2 polarization, with high CD163 and CD206 expression [69,70,71]. NLC functions include recruiting cells, enhancing CLL proliferation and inhibiting the apoptotic pathways [66,67,68,69,70,71]. Specifically, NLC can support CLL cells growth and survival through CXCL12, CXCL13 and vascular cell adhesion molecule 1 [66,67]. In a mouse model, the migration inhibitory factor was identified as a cofactor for disease pathogenesis by favoring macrophages accumulation in the organs involved by CLL [69,70]. An in vivo deep crosstalk between NLC and CLL cells was discovered and a macrophage depletion could reduce CLL engraftment and prolong mouse survival [72]. Antineoplastic activity could be based on interfering between leukemic cells and NLC interactions by an IFN-mediated NLC reprogramming or by the inhibition of the CSF1 receptor [73,74].
3.4.2. Clinical Studies
Despite the findings of pre-clinical data about the NLC support of CLL cells, a prognostic role for TAM/NLC has not been demonstrated in a clinical cohort to date, even if a TAM-based clinical approach is under investigation [75,76].
Ibrutinib, a Bruton tyrosine kinase (BTK) inhibitor, is widely used for CLL therapy, both as first-line regimen and for relapsed/refractory (R/R) cases [77,78]. Ibrutinib could inhibit the macrophage production of CXCL13 and directly modify NLC phenotype and functions by targeting BTK expressed on NLC [79]. Surprisingly, ibrutinib could favor M2 polarization and could represent a possible explanation for reduced drug efficacy [80,81].
4. TAM in Multiple Myeloma
MM is the second most common hematological plasma cells malignancy and accounts for 1–2% of all cancers, with an increasing annual incidence of approximately 4.5–6.0 new cases/100,000 inhabitants in Europe [84]. In the last few years, the use of novel agents has given an unprecedented increase of response and survival improvement; however, most patients relapse, need multiple lines of treatment and eventually die from the disease or its complications [85].
4.1. Pre-Clinical Studies
The interaction between MM plasma cells and TME is well established, including osteoblasts, osteoclasts, mesenchymal stem cells (MSC) and TAM [86]. TME influences the homing, survival, proliferation, drug resistance and immune escape of MM plasma cells, which in turn can recruit macrophages through chemotactic molecules and alter the BM TME by suppressing cytotoxic effects of reactive immune cells [86]. Specifically, TAM carry out a fundamental role in MM pathogenesis, including promoting BM plasma cells homing and proliferation, angiogenesis and the so-called vasculogenic mimicry, further supporting MM immune evasion and progression [87,88,89,90,91,92,93].
MM plasma cells in vitro could favor an M2 TAM polarization by upregulating CD206 expression of cocultured macrophages [94]. In MM cell lines, CD68-positive TAM represent an important component of TME and could inhibit the drug-induced apoptosis of neoplastic cells by the cleavage of caspase-3 and poly-ADP ribose polymerase (PARP) [92,95]. Moreover, P-selectin glycoprotein ligand-1 (PSGL-1) and intracellular adhesion molecule-1 (ICAM-1) on the MM cells surface could induce TAM activation and were involved in TAM-induced chemo resistance through the Src, Erk1/2 and c-myc pathway [96]. In addition, together with MSC, TAM could support MM cells survival and proliferation through IL-6 and IL-10 [96]. An early in vitro study showed a coculture with macrophages could increase the MM cells growth rate, and ex vivo matured human macrophages could increase human MM cells proliferation by producing IL-6, IL-10, IL-12 and VEGF [96,97,98].
Due to the TAM production of insulin growth factor (IGF)-1, clodronateliposome-mediated macrophage depletion was capable of inhibiting MM development in vitro and in vivo by influencing plasma cells migration and homing to the BM [99]. As demonstrated in several reports, during progression from monoclonal gammopathy of undetermined significance (MGUS) to MM, M2 TAM could also favor the angiogenic switch by secreting VEGF [87,88]. In a recent paper, in mouse models, BMI1, a polycombgroup protein, showed the capability to modulate the pro-myeloma functions of TAM, which showed higher BMI1 levels compared to normal macrophages. In a BMI1 knockout model, a reduced TAM proliferation was reported, together with a reduced expression of angiogenic factors [93].
De Beule and colleagues showed the pro-tumor effect of TAM was correlated with Stat3 pathway activation in 5T33MM cells. Interestingly, AZD1480, an ATP-competitive Janus kinase (JAK)2 inhibitor, could inhibit this effect and resensitize MM cells to bortezomib [100,101]. Lastly, micro (mi)RNA were investigated as potential factors contributing to MM molecular pathogenesis; in this field, exosome-derived miR-let-7c and miR-214 were recently involved in M2 TAM polarization and angiogenesis promotion in BM TME [102,103].
4.2. Clinical Studies
Pivotal clinical studies confirmed both the promising pre-clinical data and the prognostic relevance of both total and M2 TAM, in which CD68/CD163 double-positive M2 TAM were associated with an increased micro-vessel density and reduced survival, independently of the tumor stage [104,105,106,107,108].
In the BM of MM patients with an active disease, CD206-positive M2 TAM were increased compared to healthy subjects or patients presenting with MGUS [94,100].
CD163 expression detected by IHC was assessed in 198 MM patients treated with bortezomib-based regimens. A high CD163-positive M2 TAM expression at diagnosis, with the used cutoff of >55/high power field, was associated with a lower CR rate and worse PFS and OS, and its value was confirmed in a multivariate analysis [104]. Furthermore, an elevated level of soluble M2 TAM markers CD163 and CD206 was associated with a reduced OS, while a higher M1 density was correlated with an OS improvement [91,105].
In a retrospective study, 68 MM patients were enrolled, and TAM were assessed with anti-CD68 and anti CD163 antibodies [106]. An elevated CD68-positive and CD163-positive TAM expression had a detrimental effect on 6-y OS in a multivariate analysis. As a complementary finding, an elevated microvessel density was associated with an increased CD163-positive TAM number, further suggesting M2 TAM could have an adverse prognostic role [106].
In a relevant study, the polarized functional status of BM TAM by CD68, inducible NO synthase (iNOS) and CD163 IHC staining was investigated in 240 MM patients [107]. A reduced overall response rate (ORR) was observed in patients presenting with a high CD68-postive and CD163-positive TAM density; however, only high CD163 expression was associated with inferior PFS and OS. CD163 and iNOS were identified as independent prognostic factors and were combined with the international staging system (ISS) in order to generate a new prognostic score [107].
As angiogenesis induction is one of the mechanisms through which M2 TAM favor MM progression, M2 TAM infiltration and the correlation with the pro-angiogenic factor CD147 were evaluated in a spectrum ranging from MGUS to relapsed MM [108]. CD163 was used as M2 marker and the used cutoff for M2 infiltration was 100 per core. The authors showed a significant OS reduction for relapsed MM patients with high M2 expression (32 vs. 6 months, p = 0.02), further suggesting a prognostic role for CD163-positive TAM in MM [108].
Andersen and colleagues evaluated CD163 as a soluble protein in 104 blood samples and 17 BM samples in newly diagnosed MM cases [91]. CD163 expression was higher in BM compared to blood samples and was associated with a higher ISS stage and other adverse prognostic factors. An elevated CD163 expression, with the suggested cutoff of 1.8 mg/L, was correlated with a poor outcome, further indicating TAM could influence MM growth and progression [91].
5. TAM in Hodgkin lymphoma
Classic Hodgkin lymphoma (cHL) is a highly curable lymphoid malignancy that mostly affects young adults.
CHL is characterized by good prognosis in most cases, in which neoplastic cell, called Reed–Sternberg (R–S) cells, are B-cells with a defective phenotype [109]. In HL samples, R–S cells represent a minority of total cellularity and are surrounded by a reactive TME, including eosinophils, basophils, dendritic cells, plasma cells and TAM [109]. First-line therapy is represented by an ABVD regimen (doxorubicin, bleomycin, vinblastine and dacarbazine) for a total of 4–6 cycles, followed by radiation therapy as consolidation for early-stage disease. Despite encouraging results, approximately 20% of total cases experience R/R disease and none of the published prognostic factors at diagnosis can definitely identify patients at high risk of treatment failure [109].
5.1. Pre-Clinical Studies
Due to the peculiar HL biology, a possible prognostic role of TAM was first suggested in 1985 [110]. After a direct interaction with HL cells, TAM could induce in vitro HL cells proliferation through STAT3 pathway activation [111]. The signaling events and the mechanism of M2 TAM polarization in HL is not well understood and further research is needed in this field.
Programmed death protein 1 (PD-1) and its ligands (PD-L1 and PD-L2) represent fundamental molecules for the immunoevasion of R–S cells in HL. PD-L1 is expressed by R–S cells and TAM in the TME, while PD-1 is expressed by reactive T-cells. Carey and colleagues found PD-L1-positive TAMs could surround R–S cells and could especially interact with PD-1-positive/CD4-positive T-cells, further suggesting CD4-positive T-cells represent the target of the PD-1 blockade [112].
Interestingly, the CD163-positive monocytes population expressing PD-L1 was more elevated in HL patients compared to diffuse large B-cell lymphoma (DLBCL) [113]. Consistently, in HL, CD163/PD-L1/PD-L2 gene expression was more elevated compared to DLBCL. The phenotype could revert to normal after the ABVD regimen and the monocytes depletion from the baseline blood samples of HL patients could favor CD3-negative/CD56hi/CD16-negative NK-cells activation [113].
Recent reports demonstrated the role of PI3K inhibition on M2 TAM polarization, further suggesting the PI3K-Akt pathway is involved in HL pathogenesis, and its blockade could favor a switch towards an M1 polarization and lead to tumor regression [114].
5.2. Clinical Studies
Unlike in leukemia, for HL, pre-clinical studies are reduced compared to clinical data and there is not a strong correlation between pre-clinical and clinical findings. TAM in HL, determined both by GEP and by IHC using the markers CD68, emerged as a relevant prognostic factor of disease-specific survival and PFS in the pivotal paper by Steidl and colleagues, outperforming the international prognostic score (IPS) [115]. These macrophages could be considered as “bad guys”, with a pro-tumor activity and a possible inhibition of drug-mediated cell death, leading to a reduced treatment response, disease progression and reduced survival [115]. Since then, many subsequent reports were published, but showed conflicting results about a definitive association between TAM and survival, as illustrated in Table 1 [115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130].
Azambuja and colleagues showed a lack of IHC reproducibility compared to GEP, while other reports suggested CD163 could represent a more appropriate marker to detect TAM infiltration compared to CD68, that could stain myelomonocytic cells, fibroblasts and endothelial cells [115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130].
It was also speculated that TAM could represent an expression of reactive TME detected by 18F-FDG PET [117,118,119,120]. A correlation between high TAM infiltration at diagnosis (CD68 > 25%), PET positivity (Deauville score 4–5) after two ABVD cycles and reduced PFS and OS has been observed in a small study; however, these findings were not confirmed by three larger studies performed in recent years [117,118,119,120].
Recent reports demonstrated HL patients with the highest M2 TAM count, measured using CD163 or CD68/CD163 as M2 polarization markers, had a significantly reduced disease-free survival (DFS), PFS and OS, further confirming M2 TAM could support tumor progression and immune escape [122,123,128,129].
Even if a metanalysis confirmed the prognostic value of both CD68 and CD163 in HL cases, the main hurdles to overcome are represented by the choice of TAM markers (CD68 vs. CD163 vs. CD68/CD163 double-positive), the different cutoffs used to define a high TAM expression and the different monoclonal antibodies used for IHC (KP-1 or PGM-1 for CD68 evaluation) [101,131].
6. TAM in Non-Hodgkin Lymphoma
DLBCL and follicular lymphoma (FL) represent the most common aggressive and indolent NHL subtype, respectively. FL has an excellent prognosis and therapy should be reserved to patients with B symptoms, cytopenia, bulky disease or rapidly progressive disease. First-line treatments include obinutuzumab (O) or rituximab (R) in association with cyclophosphamide, doxorubicin, vincristine and prednisone (CHOP) or bendamustine (B) [132]. Conversely, DLBCL is an aggressive disease and a significant proportion of cases are refractory or relapse early after R-CHOP is used as first-line therapy [133].
6.1. Pre-Clinical Studies
TAM M2 polarization could be induced by apoptotic NHL cells, and M2 TAM have a reduced galectin-3 expression, a glycoprotein involved in the clearance of apoptotic cells progression, finally leading to NHL progression [134]. Even if the prognostic value of CD163 as an M2 marker is debated in FL, a high CD163-positive TAM density was associated with neovascularization located in the interfollicular area [135]. Early in vitro studies demonstrated macrophages could induce NHL cells proliferation after a direct contact. In this field, TAM could produce cytokines such as C5a, IL-6 and TNF-α, that in turn activate Stat3 and NF-kB pathways [136]. Lastly, in DLBCL, M2 TAM could contribute to extracellular matrix remodeling through legumain, that leads to fibronectin degradation and enhances local angiogenesis [137].
6.2. Clinical Studies
As for HL, there are limited pre-clinical studies for NHL cases and clinical data showed conflicting results compared to pre-clinical findings. Pivotal GEP-based studies demonstrated that TME has a prognostic relevance in DLBCL. A stromal-1 signature, characterized by genes expressed by components of an extracellular matrix, was associated with a good outcome, while a stromal-2 signature was characterized by a dismal prognosis, mainly due to increased angiogenesis [138]. Moreover, several studies showed TAM CD68-positive content was associated with an increased survival in DLBCL, while an elevation of CD163/CD68 ratio, suggestive of M2 polarization, was predictive of a dismal outcome, as illustrated in Table 2 [139,140,141,142,143,144,145,146,147,148]. Riihijarvi and colleagues described the outcome correlation of GEP-assessed CD68 mRNA levels and IHC CD68 protein expression. A cohort of 59 cases was investigated using the anti-CD68 KP1 antibody and an arbitrary cut-off choice; CD68 was associated with a good prognosis in patients receiving rituximab in combination with chemotherapy, while the disease outcome was poor in patients receiving only chemotherapy [139]. In this paper, CD163 expression in GEP and IHC did not demonstrate any prognostic value. Another study investigated 165 patients treated with R-CHOP, in which high CD68 expression was associated with an improved OS, while both PFS and OS were decreased in patients presenting with an elevated CD163/CD68 ratio [140]. Conversely, the other report did not observe any correlation between CD68 and survival.
As suggested for HL, a wide heterogeneity in the choice of IHC antibodies, TAM markers, scoring methods (manual or automated) and threshold could generate these conflicting findings. As suggested by a recent meta-analysis of 11 studies, the highest hazard ratio for survival is reached by CD163/CD68 TAM ratio, further confirming M2 TAM represent a robust outcome predictor for NHL patients, including DLBCL and FL [149].
In FL, high CD68-positive TAM expression was correlated with a poor clinical outcome in several reports, but other studies showed opposite findings [150,151,152,153,154]. Interestingly, a high CD163 TAM count was correlated with increased angiogenesis and microvascular density, which were in turn linked to a poor disease outcome.
7. Macrophages and Bioenergetic Modifications
One of the major aspects of the tumor–stromal interaction that was observed in solid and hematologic malignancies is represented by the bioenergetics dependence–independenceinteraction. In this field, a reverse Warburg effect, metabolic coupling and hypoxic induction, and even a mitochondrial transfer, have been suggested as possible mechanisms of interaction between tumor cells and stromal cells in leukemia, lymphoma and MM. Although the reactive cells involved in this crosstalk that finally supports tumor growth and survival have been generally identified as stromal elements, we have to mention a possible relationship between macrophages and bioenergetics modifications. In the Philadelphia (Ph) chromosome-positive leukemic cells, in which the BCR–ABL oncogene is constitutively expressed as a fusion protein with tyrosine kinase activity, the therapeutic concentration of the tyrosine kinase inhibitor imatinib reduced glucose uptake by suppressing glycolytic cell activity [155]. Furthermore, the mitochondrial Krebs cycle activity was improved, finally resulting in an increased energy state and apoptosis. The author suggested imatinib, without direct cytocidal activity, could reverse the Warburg effect in BCR–ABL-positive cells by switching from glycolysis to mitochondrial glucose metabolism [155]. The Warburg effect was demonstrated in leukemic–stromal cocultures and was mediated by mitochondrial uncoupling associated with the uncoupling protein activation 2 (UCP2). Under exposure to normal oxygen concentration, a significant interaction between leukemic cells and BM-derived MSC was reported, which promoted the accumulation of lactate in the culture medium without glucose consumption, with a subsequent reduction of mitochondrial membrane potential in both cell types. Interestingly, in leukemic cells, this reduction was mediated by mitochondrial uncoupling and was associated with increased UCP2 expression [156].
Overall, in hematologic malignancies, there is a strong association between cellular metabolism, mitochondrial bioenergetics and the interaction with supportive TME, finally contributing to drug resistance. Lymphoma and CLL cells could increase mitochondrial biogenesis with the aim of adapting to oxidative stress. Conversely, in MM, changes in bioenergetics represent an adaptive response to drug-induced stress [157]. In the BM niche, a reverse Warburg effect has been recently demonstrated, in which metabolic changes occurring in stromal cells could provide support to adjacent tumor cells. Interestingly, an increased mitochondrial biogenesis of tumor cells due to the acquisition of new mitochondria transferred by MSC could increase adenosine triphosphate production by oxidative phosphorylation and could mediate drug resistance [157]. This field could represent a possible rationale for a future target therapy for R/R patients with hematologic malignancies, who represent unmet medical need.
8. New Perspectives and Possible TAM-Related Treatment Approach
Since TAM are relevant for cancer progression, they could represent a target for immunotherapy. The aim to treat hematologic malignancies using M2 TAM as a target is under investigation with at least four research fields, including the blockade of monocyte recruitment, TAM depletion, TAM reprogramming into M1-phenotype and molecular signaling modification, as illustrated in Table 3.
Since the CCL2–CCR2 signaling axis is involved in monocytes trafficking, its inactivation in solid neoplasms significantly reduced tumor growth. Trabectedin, a DNA-binding sea squirt-derived compound, approved to treat solid tumors, could kill monocytes and macrophages and exert an antiangiogenic activity by inhibiting CCL2 and VEGF production [158,159]. In CLL mouse models, trabectedin showed an antileukemic role by reducing TAM recruitment, directly depleting TAM and increasing the memory T-cells count [158]. Due to a triggered apoptosis in MM cell lines, cell cycle arrest, VEGF depletion and NK cells upregulation, a possible role has been proposed for trabectedin in MM [159].
CSF-1R signaling is another important pathway for TAM recruitment and differentiation [74,160,161,162]. It was demonstrated that CSF-1R inhibition could block TAM polarization in T-ALL mouse models [160]. A treatment combination between vincristine and a CSF-1R inhibitor could improve survival in leukemic T-ALL mice compared to vincristine as a singleagent [160]. These findings were confirmed in CLL, in which pacritinib, a JAK2/FLT3 double inhibitor, blocked CSF-1R and was associated with TAM depletion and an improved survival [74]. The antineoplastic effect of CSF-1R inhibition was recently observed in ATLL, in which it could reduce tumor growth and sensitize malignant cells to chemotherapy [161]. Interestingly, idelalisib, a PI3K inhibitor approved for CLL and FL therapy, could block the CSF-1-mediated TAM spreading [162].
Since CXCR4/CXCL12 axis could promote M2 TAM polarization, it has been speculated that its inhibition could permit TAM reprogramming towards a M1-phenotype. Interestingly, in recent preclinical studies, the addition of the CXCR4 inhibitor plerixafor, currently used to mobilize stem cells, with ruxolitinib and venetoclax improved survival in T-ALL [163].
Ph chromosome-positive B-ALL cells could be reprogrammed in vitro into normal macrophage-like cells after exposure to myeloid differentiation-promoting cytokines. This lineage reprogramming could eliminate the leukemogenicity of Ph-positive B-ALL cells, thus representing a promising future treatment strategy [164].
Biphosphonates, currently approved for MM with bone lesions, could deplete TAM and abrogate MM establishment in vivo [99]. In hepatocellular carcinoma, the transformation into M1 pro-inflammatory TAM could be induced by sorafenib, an FLT-3 inhibitor, arguing it could represent a complementary mechanism of action for the drug when used to treat FLT3-positive AML [171]. The intermediate CD40 agonist ChiLob 7/4 and the weak agonist dacetuzumab, which were capable of inducing pro-inflammatory cytokines, gave encouraging results in pivotal trials, including hematologic malignancies [165]. M1 TAM polarization could represent a possible mechanism of action of IFN-α, giving an explanation of the reported efficacy as a preemptive therapy for ALL patients after allo-SCT [166]. Artesunate (ART) induced an increase in inflammatory monocytes in vitro, while it reduced macrophage expression of CD206 and CD163. After contact with monocytes reprogrammed by ART, the in vitro apoptosis of leukemic cells was increased, due to ART changing the monocyte phenotype by JAK2/STAT3 downregulation [167].
Lenalidomide, an antineoplastic agent with a pleiotropic effect, could influence TME through the improvement of T- and NK-cells function in combination with a significant reduction of angiogenesis [168]. Moreover, in mouse MM models, lenalidomide could permit a M2 TAM depletion and a reduction in IL-10 production [172].
Cladribine, a purine analog, currently approved to treat hairy cell leukemia with variable treatment modalities, inhibits in vitro the secretion of pro-inflammatory cytokines and phagocytosis in human M1 macrophages activated by LPS [173,174].
The most promising research field is probably represented by the usage of CD47, a glycoprotein commonly expressed in both myeloid and lymphoid malignancies, as a treatment target. The CD47 binding to its receptor SIRPα inhibits macrophage phagocytosis and favors the immune escape of tumor cells. Preclinical studies demonstrated CD47 inhibition reduced ALL and MM cells growth and improved T-cell-mediated cytotoxicity [169].
The pivotal phase I study by Advani and colleagues investigated the Hu5F9-G4 antibody and the possible synergism with rituximab in 22 R/R DLBCL or FL cases (95% were rituximab-refractory). Due to the rarity of dose-limiting effects, a phase II study was initiated; ORR was 50% (CR rate was 36%) [170]. After a median follow-up of 6.2 months and 8.1 months for DLBCL and FL, respectively, the treatment response was maintained in 91% of responsive patients. These findings demonstrated a promising antineoplastic activity by blocking the so-called CD47 “don’t eat me” signal [170].
9. Conclusions
Targeting the innate immunity to treating hematologic malignancies represents an attractive strategy, due to the peculiar TME of leukemia, myeloma and lymphoma and the well-established crosstalk between neoplastic and reactive cells. We have summarized the critical TAM role in tumor growth and progression, especially for M2 TAM, which is characterized by an immunosuppressive phenotype. We can suggest that high baseline M2 TAM content, especially in association with clinical prognostic factors, could contribute to the identification of patients characterized by high-risk disease at diagnosis. A TAM-target strategy could act in association with chemoimmunotherapy and reduce drug resistance; thus, we suggest investigating this promising approach in future studies focused on poor-risk patients.
Author Contributions
Conceptualization, E.C. and A.F.; validation, A.G., A.S. and M.B.; data curation, E.C. and A.S.; writing—original draft preparation, E.C. and A.F.; writing—review and editing, A.G. and M.B.; supervision, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- St-Pierre, F.; Bhatia, S.; Chandra, S. Harnessing Natural Killer Cells in Cancer Immunotherapy: A Review of Mechanisms and Novel Therapies. Cancers 2021, 13, 1988. [Google Scholar] [CrossRef] [PubMed]
- Djureinovic, D.; Wang, M.; Kluger, H.M. Agonistic CD40 Antibodies in Cancer Treatment. Cancers 2021, 13, 1302. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.; Kluger, H. Targeting Innate Immunity to Treat Cancer. Cancers 2020, 12, 2723. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, L.A.; Wilgus, T.A.; Koh, T.J. Macrophages in Healing Wounds: Paradoxes and Paradigms. Int. J. Mol. Sci. 2021, 22, 950. [Google Scholar] [CrossRef] [PubMed]
- Blériot, C.; Chakarov, S.; Ginhoux, F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity 2020, 52, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Heusinkveld, M.; van der Burg, S.H. Identification and manipulation of tumor associated macrophages in human cancers. J. Transl. Med. 2011, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Komohara, Y.; Takaishi, K.; Katabuchi, H.; Takeya, M. Detection of M2 macrophages and colony-stimulating factor 1 expression in serous and mucinous ovarian epithelial tumors. Pathol. Int. 2009, 59, 300–305. [Google Scholar] [CrossRef]
- Robinson, A.; Han, C.Z.; Glass, C.K.; Pollard, J.W. Monocyte Regulation in Homeostasis and Malignancy. Trends Immunol. 2021, 42, 104–119. [Google Scholar] [CrossRef]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: Potential targets of anti-cancer therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef]
- Mantovani, A.; Schioppa, T.; Porta, C.; Allavena, P.; Sica, A. Role of tumor-associated macrophages in tumor progression and invasion. Cancer Metastasis Rev. 2006, 25, 315–322. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Unver, N. Macrophage chemoattractants secreted by cancer cells: Sculptors of the tumor microenvironment and another crucial piece of the cancer secretome as a therapeutic target. Cytokine Growth Factor Rev. 2019, 50, 13–18. [Google Scholar] [CrossRef]
- Locati, M.; Mantovani, A.; Sica, A. Macrophage activation and polarization as an adaptive component of innate immunity. Adv. Immunol. 2013, 120, 163–184. [Google Scholar] [PubMed]
- Petty, A.J.; Yang, Y. Tumor-associated macrophages: Implications in cancer immunotherapy. Immunotherapy 2017, 9, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, C.; Munera-Maravilla, E.; Lodewijk, I.; Suarez-Cabrera, C.; Karaivanova, V.; Ruiz-Palomares, R.; Paramio, J.M.; Dueñas, M. Macrophage polarization as a novel weapon in conditioning tumor microenvironment for bladder cancer: Can we turn demons into gods? Clin. Transl. Oncol. 2019, 21, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.B.; Netea, M.G.; Kater, A.P.; van der Velden, W.J. ‘Trained immunity’: Consequences for lymphoid malignancies. Haematologica 2016, 101, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Ponzetta, A.; Inforzato, A.; Jaillon, S. Innate immunity, inflammation and tumour progression: Double-edged swords. J. Intern. Med. 2019, 285, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Komohara, Y.; Niino, D.; Ohnishi, K.; Ohshima, K.; Takeya, M. Role of tumor-associated macrophages in hematological malignancies. Pathol. Int. 2015, 65, 170–176. [Google Scholar] [CrossRef]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Mills, C.D. Anatomy of a discovery: m1 and m2 macrophages. Front. Immunol. 2015, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, T.D.; Prieto-Dominguez, N.; Gowda, P.S.; Ubil, E. Mechanisms of Macrophage Plasticity in the Tumor Environment: Manipulating Activation State to Improve Outcomes. Front. Immunol. 2021, 12, 642285. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Schmidt, S.V.; Sander, J.; Draffehn, A.; Krebs, W.; Quester, I.; De Nardo, D.; Gohel, T.D.; Emde, M.; Schmidleithner, L.; et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity 2014, 40, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, T.; Qian, B.Z.; Soong, D.; Cassetta, L.; Noy, R.; Sugano, G.; Kato, Y.; Li, J.; Pollard, J.W. CCL2-induced chemokine cascade promotes breast cancer metastasis by enhancing retention of metastasis-associated macrophages. J. Exp. Med. 2015, 212, 1043–1059. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Tsai, Y.; Yang, L.; Chuang, K.H.; Keng, P.C.; Lee, S.O.; Chen, Y. IL-6 Mediates Macrophage Infiltration after Irradiation via Up-regulation of CCL2/CCL5 in Non-small Cell Lung Cancer. Radiat. Res. 2017, 187, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Imamichi, T.; Weiss, J.M.; Sato, M.; Li, L.; Matsukawa, A.; Wang, J.M. Induction of Monocyte Chemoattractant Proteins in Macrophages via the Production of Granulocyte/Macrophage Colony-Stimulating Factor by Breast Cancer Cells. Front. Immunol. 2016, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.A.; Yang, B.; Tang, T.; Yang, Y.; Zhang, D.; Xiao, H.; Xu, J.; Wang, L.; Lin, L.; Jiang, J. Correlation of APE1 with VEGFA and CD163+ macrophage infiltration in bladder cancer and their prognostic significance. Oncol. Lett. 2020, 20, 2881–2887. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Matsui, S.; Touma, M.; Wu, Q.; Sugimoto, K. MicroRNA-342 inhibits tumor growth via targeting chemokine CXCL12 involved in macrophages recruitment/activation. Genes Cells 2018, 23, 1009–1022. [Google Scholar] [CrossRef] [Green Version]
- Xuan, W.; Qu, Q.; Zheng, B.; Xiong, S.; Fan, G.H. The chemotaxis of M1 and M2 macrophages is regulated by different chemokines. J. Leukoc. Biol. 2015, 97, 61–69. [Google Scholar] [CrossRef]
- Linde, N.; Lederle, W.; Depner, S.; van Rooijen, N.; Gutschalk, C.M.; Mueller, M.M. Vascular endothelial growth factor-induced skin carcinogenesis depends on recruitment and alternative activation of macrophages. J. Pathol. 2012, 227, 17–28. [Google Scholar] [CrossRef]
- Molawi, K.; Sieweke, M.H. Transcriptional control of macrophage identity, self-renewal, and function. Adv. Immunol. 2013, 120, 269–300. [Google Scholar] [PubMed]
- Laoui, D.; Van Overmeire, E.; De Baetselier, P.; Van Ginderachter, J.A.; Raes, G. Functional relationship between tumor-associated macrophages and macrophage colony-stimulating factor as contributors to cancer progression. Front. Immunol. 2014, 5, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, D.; Shi, W.; Yi, S.J.; Chen, H.; Groffen, J.; Heisterkamp, N. TGFbeta signaling plays a critical role in promoting alternative macrophage activation. BMC Immunol. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, I.L.; Arifa, R.D.N.; Rallapalli, H.; Kana, V.; Lao, Z.; Sanghrajka, R.M.; SumruBayin, N.; Tanne, A.; Wojcinski, A.; Korshunov, A.; et al. CSF1R inhibition depletes tumor-associated macrophages and attenuates tumor progression in a mouse sonic Hedgehog-Medulloblastoma model. Oncogene 2021, 40, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Schiarea, S.; Liguori, M.; Fabbri, M.; Pesce, S.; Zammataro, L.; Pasqualini, F.; Nebuloni, M.; Chiabrando, C.; Mantovani, A.; et al. Tumor-conditioned macrophages secrete migration-stimulating factor: A new marker for M2-polarization, influencing tumor cell motility. J. Immunol. 2010, 185, 642–652. [Google Scholar] [CrossRef] [Green Version]
- Colegio, O.R.; Chu, N.Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef]
- Maller, O.; Drain, A.P.; Barrett, A.S.; Borgquist, S.; Ruffell, B.; Zakharevich, I.; Pham, T.T.; Gruosso, T.; Kuasne, H.; Lakins, J.N.; et al. Tumour-associated macrophages drive stromal cell-dependent collagen crosslinking and stiffening to promote breast cancer aggression. Nat. Mater. 2021, 20, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, N.; Safari, Z.; Pornour, M.; Amirizadeh, N.; Forouzandeh Moghadam, M.; Sadeghizadeh, M. Alteration of cellular and immune-related properties of bone marrow mesenchymal stem cells and macrophages by K562 chronic myeloid leukemia cell derived exosomes. J. Cell. Physiol. 2019, 234, 3697–3710. [Google Scholar] [CrossRef]
- Que, Y.; Li, H.; Lin, L.; Zhu, X.; Xiao, M.; Wang, Y.; Zhu, L.; Li, D. Study on the Immune Escape Mechanism of Acute Myeloid Leukemia with DNMT3A Mutation. Front. Immunol. 2021, 12, 653030. [Google Scholar] [CrossRef]
- Jarosz-Biej, M.; Kamińska, N.; Matuszczak, S.; Cichoń, T.; Pamuła-Piłat, J.; Czapla, J.; Smolarczyk, R.; Skwarzyńska, D.; Kulik, K.; Szala, S. M1-like macrophages change tumor blood vessels and microenvironment in murine melanoma. PLoS ONE 2018, 13, e0191012. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.J.; Gu, Y.; Wang, C.Z.; Jin, Y.; Wen, X.M.; Ma, J.C.; Tang, L.J.; Mao, Z.W.; Qian, J.; Lin, J. The M2 macrophage marker CD206: A novel prognostic indicator for acute myeloid leukemia. Oncoimmunology 2019, 9, 1683347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelzer, D.; Bassan, R.; Dombret, H.; Fielding, A.; Ribera, J.M.; Buske, C.; ESMO Guidelines Committee. Acute lymphoblastic leukaemia in adult patients: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2016, 27, v69–v82. [Google Scholar] [CrossRef] [PubMed]
- Simioni, C.; Bergamini, F.; Ferioli, M.; Rimondi, E.; Caruso, L.; Neri, L.M. New biomarkers and therapeutic strategies in acute lymphoblastic leukemias: Recent advances. Hematol. Oncol. 2020, 38, 22–33. [Google Scholar] [CrossRef]
- Zappone, E.; Cencini, E.; Defina, M.; Sicuranza, A.; Gozzetti, A.; Ciofini, S.; Raspadori, D.; Mecacci, B.; Bocchia, M. Venetoclax in association with decitabine as effective bridge to transplant in a case of relapsed early T-cell lymphoblastic leukemia. Clin. Case Rep. 2020, 8, 2000–2002. [Google Scholar] [CrossRef] [PubMed]
- Song, J.X.; Wen, Y.; Li, R.W.; Dong, T.; Tang, Y.F.; Zhang, J.J.; Sa, Y.L. Phenotypic characterization of macrophages in the BMB sample of human acute leukemia. Ann. Hematol. 2020, 99, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Dander, E.; Fallati, A.; Gulić, T.; Pagni, F.; Gaspari, S.; Silvestri, D.; Cricrì, G.; Bedini, G.; Portale, F.; Buracchi, C.; et al. Monocyte-macrophage polarization and recruitment pathways in the tumour microenvironment of B-cell acute lymphoblastic leukaemia. Br. J. Haematol. 2021, in press. [Google Scholar] [CrossRef]
- Chen, S.Y.; Yang, X.; Feng, W.L.; Liao, J.F.; Wang, L.N.; Feng, L.; Lin, Y.M.; Ren, Q.; Zheng, G.G. Organ-specific microenvironment modifies diverse functional and phenotypic characteristics of leukemia-associated macrophages in mouse T cell acute lymphoblastic leukemia. J. Immunol. 2015, 194, 2919–2929. [Google Scholar] [CrossRef] [Green Version]
- Valencia, J.M.; Fernández-Sevilla, L.; Fraile-Ramos, A.; Sacedón, R.; Jiménez, E.; Vicente, A.; Varas, A. Acute Lymphoblastic Leukaemia Cells Impair Dendritic Cell and Macrophage Differentiation: Role of BMP4. Cells 2019, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Saint Fleur-Lominy, S.; Maus, M.; Vaeth, M.; Lange, I.; Zee, I.; Suh, D.; Liu, C.; Wu, X.; Tikhonova, A.; Aifantis, I.; et al. STIM1 and STIM2 Mediate Cancer-Induced Inflammation in T Cell Acute Lymphoblastic Leukemia. Cell Rep. 2018, 24, 3045–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Jan, M.; Weissman-Tsukamoto, R.; Zhao, F.; Park, C.Y.; Weissman, I.L.; Majeti, R. Therapeutic antibody targeting of CD47 eliminates human acute lymphoblastic leukemia. Cancer Res. 2011, 71, 1374–1384. [Google Scholar] [CrossRef] [Green Version]
- Hohtari, H.; Brück, O.; Blom, S.; Turkki, R.; Sinisalo, M.; Kovanen, P.E.; Kallioniemi, O.; Pellinen, T.; Porkka, K.; Mustjoki, S. Immune cell constitution in bone marrow microenvironment predicts outcome in adult ALL. Leukemia 2019, 33, 1570–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukasaki, K.; Marçais, A.; Nasr, R.; Kato, K.; Fukuda, T.; Hermine, O.; Bazarbachi, A. Diagnostic Approaches and Established Treatments for Adult T Cell Leukemia Lymphoma. Front. Microbiol. 2020, 11, 1207. [Google Scholar] [CrossRef]
- Komohara, Y.; Niino, D.; Saito, Y.; Ohnishi, K.; Horlad, H.; Ohshima, K.; Takeya, M. Clinical significance of CD163+ tumor-associated macrophages in patients with adult T-cell leukemia/lymphoma. Cancer Sci. 2013, 104, 945–951. [Google Scholar] [CrossRef]
- Saito, Y.; Komohara, Y.; Niino, D.; Horlad, H.; Ohnishi, K.; Takeya, H.; Kawaguchi, H.; Shimizu, H.; Ohshima, K.; Takeya, M. Role of CD204-positive tumor-associated macrophages in adult T-cell leukemia/lymphoma. J. Clin. Exp. Hematop. 2014, 54, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollyea, D.A.; Bixby, D.; Perl, A.; Bhatt, V.R.; Altman, J.K.; Appelbaum, F.R.; de Lima, M.; Fathi, A.T.; Foran, J.M.; Gojo, I.; et al. NCCN Guidelines Insights: Acute Myeloid Leukemia, Version 2. 2021. J. Natl. Compr. Cancer Netw. 2021, 19, 16–27. [Google Scholar] [CrossRef]
- Al-Matary, Y.S.; Botezatu, L.; Opalka, B.; Hönes, J.M.; Lams, R.F.; Thivakaran, A.; Schütte, J.; Köster, R.; Lennartz, K.; Schroeder, T.; et al. Acute myeloid leukemia cells polarize macrophages towards a leukemia supporting state in a Growth factor independence 1 dependent manner. Haematologica 2016, 101, 1216–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wang, R.; Feng, W.; Yang, F.; Wang, L.; Yang, X.; Ren, L.; Zheng, G. Peritoneal resident macrophages in mice with MLL-AF9-induced acute myeloid leukemia show an M2-like phenotype. Ann. Transl. Med. 2021, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, W.; Wang, R.; Yang, F.; Wang, L.; Chen, S.; Ru, Y.; Cheng, T.; Zheng, G. Repolarizing heterogeneous leukemia-associated macrophages with more M1 characteristics eliminates their pro-leukemic effects. Oncoimmunology 2017, 7, e1412910. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J.; Qian, L.; Miao, Y.; Song, W.; Liu, H.; Li, R. MOZ Forms an Autoregulatory Feedback Loop with miR-223 in AML and Monocyte/Macrophage Development. iScience 2019, 11, 189–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorst, B.; Robak, T.; Montserrat, E.; Ghia, P.; Niemann, C.U.; Kater, A.P.; Gregor, M.; Cymbalista, F.; Buske, C.; Hillmen, P.; et al. Chronic lymphocytic leukaemia: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2021, 32, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Chiorazzi, N.; Stevenson, F.K. Celebrating 20 Years of IGHV Mutation Analysis in CLL. Hemasphere 2020, 4, e334. [Google Scholar] [CrossRef] [PubMed]
- Morabito, F.; Gentile, M.; Monti, P.; Recchia, A.G.; Menichini, P.; Skafi, M.; Atrash, M.; De Luca, G.; Bossio, S.; Al-Janazreh, H.; et al. TP53 dysfunction in chronic lymphocytic leukemia: Clinical relevance in the era of B-cell receptors and BCL-2 inhibitors. Expert Opin. Investig. Drugs 2020, 29, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Cencini, E.; Sozzi, E.; Sicuranza, A.; Fabbri, A.; Raspadori, D.; Lauria, F.; Forconi, F. A pilot monocentric analysis of efficacy and safety of Fludarabine-Campath combination (Flucam) as first line treatment in elderly patients with chronic lymphocytic leukaemia and Tp53 disfunction. Br. J. Haematol. 2011, 154, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Tsukada, N.; Burger, M.; Zvaifler, N.J.; Dell’Aquila, M.; Kipps, T.J. Blood-derived nurse-like cells protect chronic lymphocytic leukemia B cells from spontaneous apoptosis through stromal cell-derived factor-1. Blood 2000, 96, 2655–2663. [Google Scholar] [CrossRef]
- Tsukada, N.; Burger, J.A.; Zvaifler, N.J.; Kipps, T.J. Distinctive features of “nurselike” cells that differentiate in the context of chronic lymphocytic leukemia. Blood 2002, 99, 1030–1037. [Google Scholar] [CrossRef]
- Boissard, F.; Fournié, J.J.; Laurent, C.; Poupot, M.; Ysebaert, L. Nurse like cells: Chronic lymphocytic leukemia associated macrophages. Leuk. Lymphoma 2015, 56, 1570–1572. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, P.; Pietra, G.; Travaini, G.; Quarto, R.; Shyti, G.; Benelli, R.; Ottaggio, L.; Mingari, M.C.; Zupo, S.; Cutrona, G.; et al. Chronic lymphocytic leukemia nurse-like cells express hepatocyte growth factor receptor (c-MET) and indoleamine 2,3-dioxygenase and display features of immunosuppressive type 2 skewed macrophages. Haematologica 2014, 99, 1078–1087. [Google Scholar] [CrossRef] [Green Version]
- Hanna, B.S.; McClanahan, F.; Yazdanparast, H.; Zaborsky, N.; Kalter, V.; Rößner, P.M.; Benner, A.; Dürr, C.; Egle, A.; Gribben, J.G.; et al. Depletion of CLL-associated patrolling monocytes and macrophages controls disease development and repairs immune dysfunction in vivo. Leukemia 2016, 30, 570–579. [Google Scholar] [CrossRef]
- Audrito, V.; Serra, S.; Brusa, D.; Mazzola, F.; Arruga, F.; Vaisitti, T.; Coscia, M.; Maffei, R.; Rossi, D.; Wang, T.; et al. Extracellular nicotinamide phosphoribosyltransferase (NAMPT) promotes M2 macrophage polarization in chronic lymphocytic leukemia. Blood 2015, 125, 111–123. [Google Scholar] [CrossRef]
- Galletti, G.; Scielzo, C.; Barbaglio, F.; Rodriguez, T.V.; Riba, M.; Lazarevic, D.; Cittaro, D.; Simonetti, G.; Ranghetti, P.; Scarfò, L.; et al. Targeting Macrophages Sensitizes Chronic Lymphocytic Leukemia to Apoptosis and Inhibits Disease Progression. Cell Rep. 2016, 14, 1748–1760. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.; Fatehchand, K.; Elavazhagan, S.; Reader, B.F.; Ren, L.; Mo, X.; Byrd, J.C.; Tridandapani, S.; Butchar, J.P. Reprogramming Nurse-like Cells with Interferon gamma to Interrupt Chronic Lymphocytic Leukemia Cell Survival. J. Biol. Chem. 2016, 291, 14356–14362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polk, A.; Lu, Y.; Wang, T.; Seymour, E.; Bailey, N.G.; Singer, J.W.; Boonstra, P.S.; Lim, M.S.; Malek, S.; Wilcox, R.A. Colony-Stimulating Factor-1 Receptor Is Required for Nurse-like Cell Survival in Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2016, 22, 6118–6128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissard, F.; Laurent, C.; Ramsay, A.G.; Quillet-Mary, A.; Fournié, J.J.; Poupot, M.; Ysebaert, L. Nurse-like cells impact on disease progression in chronic lymphocytic leukemia. Blood Cancer J. 2016, 6, e381. [Google Scholar] [CrossRef] [Green Version]
- Fiorcari, S.; Maffei, R.; Atene, C.G.; Potenza, L.; Luppi, M.; Marasca, R. Nurse-Like Cells and Chronic Lymphocytic Leukemia B Cells: A Mutualistic Crosstalk inside Tissue Microenvironments. Cells 2021, 10, 217. [Google Scholar] [CrossRef]
- Woyach, J.A.; Ruppert, A.S.; Heerema, N.A.; Zhao, W.; Booth, A.M.; Ding, W.; Bartlett, N.L.; Brander, D.M.; Barr, P.M.; Rogers, K.A.; et al. Ibrutinib Regimens versus Chemoimmunotherapy in Older Patients with Untreated CLL. N. Engl. J. Med. 2018, 379, 2517–2528. [Google Scholar] [CrossRef]
- Byrd, J.C.; Brown, J.R.; O’Brien, S.; Barrientos, J.C.; Kay, N.E.; Reddy, N.M.; Coutre, S.; Tam, C.S.; Mulligan, S.P.; Jaeger, U.; et al. RESONATE Investigators. Ibrutinib versus ofatumumab in previously treated chronic lymphoid leukemia. N. Engl. J. Med. 2014, 371, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Fiorcari, S.; Maffei, R.; Audrito, V.; Martinelli, S.; Ten Hacken, E.; Zucchini, P.; Grisendi, G.; Potenza, L.; Luppi, M.; Burger, J.A.; et al. Ibrutinib modifies the function of monocyte/macrophage population in chronic lymphocytic leukemia. Oncotarget 2016, 7, 65968–65981. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Beckwith, K.; Do, P.; Mundy, B.L.; Gordon, A.; Lehman, A.M.; Maddocks, K.J.; Cheney, C.; Jones, J.A.; Flynn, J.M.; et al. Ibrutinib treatment improves T cell number and function in CLL patients. J. Clin. Investig. 2017, 127, 3052–3064. [Google Scholar] [CrossRef]
- Boissard, F.; Fournié, J.J.; Quillet-Mary, A.; Ysebaert, L.; Poupot, M. Nurse-like cells mediate ibrutinib resistance in chronic lymphocytic leukemia patients. Blood Cancer J. 2015, 5, e355. [Google Scholar] [CrossRef] [Green Version]
- Fiorcari, S.; Martinelli, S.; Bulgarelli, J.; Audrito, V.; Zucchini, P.; Colaci, E.; Potenza, L.; Narni, F.; Luppi, M.; Deaglio, S.; et al. Lenalidomide interferes with tumor-promoting properties of nurse-like cells in chronic lymphocytic leukemia. Haematologica 2015, 100, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Schulz, A.; Dürr, C.; Zenz, T.; Döhner, H.; Stilgenbauer, S.; Lichter, P.; Seiffert, M. Lenalidomide reduces survival of chronic lymphocytic leukemia cells in primary cocultures by altering the myeloid microenvironment. Blood 2013, 121, 2503–2511. [Google Scholar] [CrossRef] [PubMed]
- Gozzetti, A.; Candi, V.; Papini, G.; Bocchia, M. Therapeutic advancements in multiple myeloma. Front. Oncol. 2014, 4, 241. [Google Scholar] [CrossRef] [PubMed]
- Gozzetti, A.; Raspadori, D.; Bacchiarri, F.; Sicuranza, A.; Pacelli, P.; Ferrigno, I.; Tocci, D.; Bocchia, M. Minimal Residual Disease in Multiple Myeloma: State of the Art and Applications in Clinical Practice. J. Pers. Med. 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- García-Ortiz, A.; Rodríguez-García, Y.; Encinas, J.; Maroto-Martín, E.; Castellano, E.; Teixidó, J.; Martínez-López, J. The Role of Tumor Microenvironment in Multiple Myeloma Development and Progression. Cancers 2021, 13, 217. [Google Scholar] [CrossRef] [PubMed]
- Scavelli, C.; Nico, B.; Cirulli, T.; Ria, R.; Di Pietro, G.; Mangieri, D.; Bacigalupo, A.; Mangialardi, G.; Coluccia, A.M.; Caravita, T.; et al. Vasculogenic mimicry by bone marrow macrophages in patients with multiple myeloma. Oncogene 2008, 27, 663–674. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A. The role of monocytes-macrophages in vasculogenesis in multiple myeloma. Leukemia 2009, 23, 1535–1536. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zheng, Y.; Li, T.; Wang, Q.; Qian, J.; Lu, Y.; Zhang, M.; Bi, E.; Yang, M.; Reu, F.; et al. Chemokines CCL2, 3, 14 stimulate macrophage bone marrow homing, proliferation, and polarization in multiple myeloma. Oncotarget 2015, 6, 24218–24229. [Google Scholar] [CrossRef] [Green Version]
- Calcinotto, A.; Ponzoni, M.; Ria, R.; Grioni, M.; Cattaneo, E.; Villa, I.; Sabrina Bertilaccio, M.T.; Chesi, M.; Rubinacci, A.; Tonon, G.; et al. Modifications of the mouse bone marrow microenvironment favor angiogenesis and correlate with disease progression from asymptomatic to symptomatic multiple myeloma. Oncoimmunology 2015, 4, e1008850. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.N.; Abildgaard, N.; Maniecki, M.B.; Møller, H.J.; Andersen, N.F. Monocyte/macrophage-derived soluble CD163: A novel biomarker in multiple myeloma. Eur. J. Haematol. 2014, 93, 41–47. [Google Scholar] [CrossRef]
- Zheng, Y.; Cai, Z.; Wang, S.; Zhang, X.; Qian, J.; Hong, S.; Li, H.; Wang, M.; Yang, J.; Yi, Q. Macrophages are an abundant component of myeloma microenvironment and protect myeloma cells from chemotherapy drug-induced apoptosis. Blood 2009, 114, 3625–3628. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Huang, J.; Wang, F.; Ding, H.; Cui, Y.; Yang, Y.; Xu, J.; Luo, H.; Gao, Y.; Pan, L.; et al. BMI1 regulates multiple myeloma-associated macrophage’s pro-myeloma functions. Cell Death Dis. 2021, 12, 495. [Google Scholar] [CrossRef]
- Beider, K.; Bitner, H.; Leiba, M.; Gutwein, O.; Koren-Michowitz, M.; Ostrovsky, O.; Abraham, M.; Wald, H.; Galun, E.; Peled, A.; et al. Multiple myeloma cells recruit tumor-supportive macrophages through the CXCR4/CXCL12 axis and promote their polarization toward the M2 phenotype. Oncotarget 2014, 5, 11283–11296. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Yang, J.; Qian, J.; Qiu, P.; Hanabuchi, S.; Lu, Y.; Wang, Z.; Liu, Z.; Li, H.; He, J.; et al. PSGL-1/selectin and ICAM-1/CD18 interactions are involved in macrophage-induced drug resistance in myeloma. Leukemia 2013, 27, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Denu, R.A.; Dollar, B.A.; Escalante, L.E.; Kuether, J.P.; Callander, N.S.; Asimakopoulos, F.; Hematti, P. Macrophages and mesenchymal stromal cells support survival and proliferation of multiple myeloma cells. Br. J. Haematol. 2012, 158, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Durie, B.G.; Vela, E.E.; Frutiger, Y. Macrophages as an important source of paracrine IL6 in myeloma bone marrow. Curr. Top. Microbiol. Immunol. 1990, 166, 33–36. [Google Scholar]
- Tai, Y.T.; Acharya, C.; An, G.; Moschetta, M.; Zhong, M.Y.; Feng, X.; Cea, M.; Cagnetta, A.; Wen, K.; van Eenennaam, H.; et al. APRIL and BCMA promote human multiple myeloma growth and immunosuppression in the bone marrow microenvironment. Blood 2016, 127, 3225–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opperman, K.S.; Vandyke, K.; Clark, K.C.; Coulter, E.A.; Hewett, D.R.; Mrozik, K.M.; Schwarz, N.; Evdokiou, A.; Croucher, P.I.; Psaltis, P.J.; et al. Clodronate-Liposome Mediated Macrophage Depletion Abrogates Multiple Myeloma Tumor Establishment In Vivo. Neoplasia 2019, 21, 777–787. [Google Scholar] [CrossRef] [PubMed]
- De Beule, N.; De Veirman, K.; Maes, K.; De Bruyne, E.; Menu, E.; Breckpot, K.; De Raeve, H.; Van Rampelbergh, R.; Van Ginderachter, J.A.; Schots, R.; et al. Tumour-associated macrophage-mediated survival of myeloma cells through STAT3 activation. J. Pathol. 2017, 241, 534–546. [Google Scholar] [CrossRef]
- Cencini, E.; Fabbri, A.; Bocchia, M. Prognostic role of M2 tumour-associated macrophages in lymphoproliferative disorders. J. Pathol. 2017, 242, 511–512. [Google Scholar] [CrossRef]
- Tian, X.; Sun, M.; Wu, H.; Chen, C.; Li, H.; Qiu, S.; Wang, T.; Han, J.; Xiao, Q.; Chen, K. Exosome-derived miR-let-7c promotes angiogenesis in multiple myeloma by polarizing M2 macrophages in the bone marrow microenvironment. Leuk. Res. 2021, 105, 106566. [Google Scholar] [CrossRef]
- Gao, Y.; Fang, P.; Li, W.J.; Zhang, J.; Wang, G.P.; Jiang, D.F.; Chen, F.P. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol. Immunol. 2020, 117, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, W.M.; Xia, Z.J.; Liang, Y.; Lu, Y.; Lin, S.X.; Tang, H. High numbers of CD163+ tumor-associated macrophages correlate with poor prognosis in multiple myeloma patients receiving bortezomib-based regimens. J. Cancer 2019, 10, 3239–3245. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.N.; Andersen, N.F.; Rødgaard-Hansen, S.; Hokland, M.; Abildgaard, N.; Møller, H.J. The novel biomarker of alternative macrophage activation, soluble mannose receptor (sMR/sCD206): Implications in multiple myeloma. Leuk. Res. 2015, 39, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Suyanı, E.; Sucak, G.T.; Akyürek, N.; Sahin, S.; Baysal, N.A.; Yağcı, M.; Haznedar, R. Tumor-associated macrophages as a prognostic parameter in multiple myeloma. Ann. Hematol. 2013, 92, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, J.; Zhang, W.; Sun, R.; Liu, T.; Zheng, Y.; Wu, Y. Prognostic value of diametrically polarized tumor-associated macrophages in multiple myeloma. Oncotarget 2017, 8, 112685–112696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchabhai, S.; Kelemen, K.; Ahmann, G.; Sebastian, S.; Mantei, J.; Fonseca, R. Tumor-associated macrophages and extracellular matrix metalloproteinase inducer in prognosis of multiple myeloma. Leukemia 2016, 30, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Momotow, J.; Borchmann, S.; Eichenauer, D.A.; Engert, A.; Sasse, S. Hodgkin Lymphoma-Review on Pathogenesis, Diagnosis, Current and Future Treatment Approaches for Adult Patients. J. Clin. Med. 2021, 10, 1125. [Google Scholar] [CrossRef] [PubMed]
- Ree, H.J.; Kadin, M.E. Macrophage-histiocytes in Hodgkin’s disease. The relation of peanut-agglutinin-binding macrophage-histiocytes to clinicopathologic presentation and course of disease. Cancer 1985, 56, 333–338. [Google Scholar] [CrossRef]
- Holtick, U.; Vockerodt, M.; Pinkert, D.; Schoof, N.; Stürzenhofecker, B.; Kussebi, N.; Lauber, K.; Wesselborg, S.; Löffler, D.; Horn, F.; et al. STAT3 is essential for Hodgkin lymphoma cell proliferation and is a target of tyrphostin AG17 which confers sensitization for apoptosis. Leukemia 2005, 19, 936–944. [Google Scholar] [CrossRef]
- Carey, C.D.; Gusenleitner, D.; Lipschitz, M.; Roemer, M.G.M.; Stack, E.C.; Gjini, E.; Hu, X.; Redd, R.; Freeman, G.J.; Neuberg, D.; et al. Topological analysis reveals a PD-L1-associated microenvironmental niche for Reed-Sternberg cells in Hodgkin lymphoma. Blood 2017, 130, 2420–2430. [Google Scholar] [CrossRef]
- Vari, F.; Arpon, D.; Keane, C.; Hertzberg, M.S.; Talaulikar, D.; Jain, S.; Cui, Q.; Han, E.; Tobin, J.; Bird, R.; et al. Immune evasion via PD-1/PD-L1 on NK cells and monocyte/macrophages is more prominent in Hodgkin lymphoma than DLBCL. Blood 2018, 131, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, S.L.; Careddu, G.; Serio, S.; Consonni, F.M.; Maeda, A.; Viswanadha, S.; Vakkalanka, S.; Castagna, L.; Santoro, A.; Allavena, P. Targeting Cancer Cells and Tumor Microenvironment in Preclinical and Clinical Models of Hodgkin Lymphoma Using the Dual PI3Kδ/γ Inhibitor RP6530. Clin. Cancer Res. 2019, 25, 1098–1112. [Google Scholar] [CrossRef] [Green Version]
- Steidl, C.; Lee, T.; Shah, S.P.; Farinha, P.; Han, G.; Nayar, T.; Delaney, A.; Jones, S.J.; Iqbal, J.; Weisenburger, D.D.; et al. Tumor-associated macrophages and survival in classic Hodgkin’s lymphoma. N. Engl. J. Med. 2010, 362, 875–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azambuja, D.; Natkunam, Y.; Biasoli, I.; Lossos, I.S.; Anderson, M.W.; Morais, J.C.; Spector, N. Lack of association of tumor-associated macrophages with clinical outcome in patients with classical Hodgkin’s lymphoma. Ann. Oncol. 2012, 23, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Cencini, E.; Fabbri, A.; Rigacci, L.; Lazzi, S.; Gini, G.; Cox, M.C.; Mancuso, S.; Abruzzese, E.; Kovalchuk, S.; Goteri, G.; et al. Evaluation of the prognostic role of tumour-associated macrophages in newly diagnosed classical Hodgkin lymphoma and correlation with early FDG-PET assessment. Hematol. Oncol. 2017, 35, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Agur, A.; Amir, G.; Paltiel, O.; Klein, M.; Dann, E.J.; Goldschmidt, H.; Goldschmidt, N. CD68 staining correlates with the size of residual mass but not with survival in classical Hodgkin lymphoma. Leuk. Lymphoma 2015, 56, 1315–1319. [Google Scholar] [CrossRef] [PubMed]
- Cuccaro, A.; Annunziata, S.; Cupelli, E.; Martini, M.; Calcagni, M.L.; Rufini, V.; Giachelia, M.; Bartolomei, F.; Galli, E.; D’Alò, F.; et al. CD68+ cell count, early evaluation with PET and plasma TARC levels predict response in Hodgkin lymphoma. Cancer Med. 2016, 5, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Touati, M.; Delage-Corre, M.; Monteil, J.; Abraham, J.; Moreau, S.; Remenieras, L.; Gourin, M.P.; Dmytruk, N.; Olivrie, A.; Turlure, P.; et al. CD68-positive tumor-associated macrophages predict unfavorable treatment outcomes in classical Hodgkin lymphoma in correlation with interim fluorodeoxyglucose-positron emission tomography assessment. Leuk. Lymphoma 2015, 56, 332–341. [Google Scholar] [CrossRef]
- Kayal, S.; Mathur, S.; Karak, A.K.; Kumar, L.; Sharma, A.; Bakhshi, S.; Raina, V. CD68 tumor-associated macrophage marker is not prognostic of clinical outcome in classical Hodgkin lymphoma. Leuk. Lymphoma 2014, 55, 1031–1037. [Google Scholar] [CrossRef]
- Klein, J.L.; Nguyen, T.T.; Bien-Willner, G.A.; Chen, L.; Foyil, K.V.; Bartlett, N.L.; Duncavage, E.J.; Hassan, A.; Frater, J.L.; Kreisel, F. CD163 immunohistochemistry is superior to CD68 in predicting outcome in classical Hodgkin lymphoma. Am. J. Clin. Pathol. 2014, 141, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Yoon, D.H.; Koh, Y.W.; Kang, H.J.; Kim, S.; Park, C.S.; Lee, S.W.; Suh, C.; Huh, J. CD68 and CD163 as prognostic factors for Korean patients with Hodgkin lymphoma. Eur. J. Haematol. 2012, 88, 292–305. [Google Scholar] [CrossRef]
- Tan, K.L.; Scott, D.W.; Hong, F.; Kahl, B.S.; Fisher, R.I.; Bartlett, N.L.; Advani, R.H.; Buckstein, R.; Rimsza, L.M.; Connors, J.M.; et al. Tumor-associated macrophages predict inferior outcomes in classic Hodgkin lymphoma: A correlative study from the E2496 Intergroup trial. Blood 2012, 120, 3280–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, P.; Clear, A.; Coutinho, R.; Wilson, A.; Matthews, J.; Owen, A.; Shanyinde, M.; Lister, T.A.; Calaminici, M.; Gribben, J.G. Expression of FOXP3, CD68, and CD20 at diagnosis in the microenvironment of classical Hodgkin lymphoma is predictive of outcome. J. Clin. Oncol. 2013, 31, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakovic, L.R.; Mihaljevic, B.S.; Perunicic Jovanovic, M.D.; Bogdanovic, A.D.; Andjelic, B.M.; Bumbasirevic, V.Z. The prognostic relevance of tumor associated macrophages in advanced stage classical Hodgkin lymphoma. Leuk. Lymphoma 2011, 52, 1913–1919. [Google Scholar] [CrossRef]
- Kamper, P.; Bendix, K.; Hamilton-Dutoit, S.; Honoré, B.; Nyengaard, J.R.; d’Amore, F. Tumor-infiltrating macrophages correlate with adverse prognosis and Epstein-Barr virus status in classical Hodgkin’s lymphoma. Haematologica 2011, 96, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Werner, L.; Dreyer, J.H.; Hartmann, D.; Barros, M.H.M.; Büttner-Herold, M.; Grittner, U.; Niedobitek, G. Tumor-associated macrophages in classical Hodgkin lymphoma: Hormetic relationship to outcome. Sci. Rep. 2020, 10, 9410. [Google Scholar] [CrossRef]
- Karihtala, K.; Leivonen, S.K.; Brück, O.; Karjalainen-Lindsberg, M.L.; Mustjoki, S.; Pellinen, T.; Leppä, S. Prognostic Impact of Tumor-Associated Macrophages on Survival Is Checkpoint Dependent in Classical Hodgkin Lymphoma. Cancers 2020, 12, 877. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, O.; El Bastawisy, A.; Allahlobi, N.; Abdellateif, M.S.; Zekri, A.R.N.; Shaarawy, S.; Korany, Z.; Mohanad, M.; Bahnassy, A.A. The role of CD68+ macrophage in classical Hodgkin lymphoma patients from Egypt. Diagn. Pathol. 2020, 15, 10. [Google Scholar] [CrossRef]
- Guo, B.; Cen, H.; Tan, X.; Ke, Q. Meta-analysis of the prognostic and clinical value of tumor-associated macrophages in adult classical Hodgkin lymphoma. BMC Med. 2016, 14, 159. [Google Scholar] [CrossRef] [Green Version]
- Tilly, H.; Gomes da Silva, M.; Vitolo, U.; Jack, A.; Meignan, M.; Lopez-Guillermo, A.; Walewski, J.; André, M.; Johnson, P.W.; Pfreundschuh, M.; et al. Diffuse large B-cell lymphoma (DLBCL): ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Dreyling, M.; Ghielmini, M.; Rule, S.; Salles, G.; Ladetto, M.; Tonino, S.H.; Herfarth, K.; Seymour, J.F.; Jerkeman, M.; ESMO Guidelines Committee. Newly diagnosed and relapsed follicular lymphoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2021, 32, 298–308. [Google Scholar] [CrossRef]
- Voss, J.J.L.P.; Ford, C.A.; Petrova, S.; Melville, L.; Paterson, M.; Pound, J.D.; Holland, P.; Giotti, B.; Freeman, T.C.; Gregory, C.D. Modulation of macrophage antitumor potential by apoptotic lymphoma cells. Cell Death Differ. 2017, 24, 971–983. [Google Scholar] [CrossRef] [Green Version]
- Clear, A.J.; Lee, A.M.; Calaminici, M.; Ramsay, A.G.; Morris, K.J.; Hallam, S.; Kelly, G.; Macdougall, F.; Lister, T.A.; Gribben, J.G. Increased angiogenic sprouting in poor prognosis FL is associated with elevated numbers of CD163+ macrophages within the immediate sprouting microenvironment. Blood 2010, 115, 5053–5056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, B.; Horlad, H.; Saito, Y.; Ohnishi, K.; Fujiwara, Y.; Takeya, M.; Komohara, Y. Role of Stat3 activation in cell-cell interaction between B-cell lymphoma and macrophages: The in vitro study. J. Clin. Exp. Hematop. 2013, 53, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, H.; Shi, Y.; Wang, D.; Gong, J.; Xun, J.; Zhou, S.; Xiang, R.; Tan, X. M2 tumour-associated macrophages contribute to tumour progression via legumain remodelling the extracellular matrix in diffuse large B cell lymphoma. Sci. Rep. 2016, 6, 30347. [Google Scholar] [CrossRef] [PubMed]
- Lenz, G.; Wright, G.; Dave, S.S.; Xiao, W.; Powell, J.; Zhao, H.; Xu, W.; Tan, B.; Goldschmidt, N.; Iqbal, J.; et al. Lymphoma/Leukemia Molecular Profiling Project. Stromal gene signatures in large-B-cell lymphomas. N. Engl. J. Med. 2008, 359, 2313–2323. [Google Scholar] [CrossRef] [Green Version]
- Riihijärvi, S.; Fiskvik, I.; Taskinen, M.; Vajavaara, H.; Tikkala, M.; Yri, O.; Karjalainen-Lindsberg, M.L.; Delabie, J.; Smeland, E.; Holte, H.; et al. Prognostic influence of macrophages in patients with diffuse large B-cell lymphoma: A correlative study from a Nordic phase II trial. Haematologica 2015, 100, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Marchesi, F.; Cirillo, M.; Bianchi, A.; Gately, M.; Olimpieri, O.M.; Cerchiara, E.; Renzi, D.; Micera, A.; Balzamino, B.O.; Bonini, S.; et al. High density of CD68+/CD163+ tumour-associated macrophages (M2-TAM) at diagnosis is significantly correlated to unfavorable prognostic factors and to poor clinical outcomes in patients with diffuse large B-cell lymphoma. Hematol. Oncol. 2015, 33, 110–112. [Google Scholar] [CrossRef]
- Wada, N.; Zaki, M.A.; Hori, Y.; Hashimoto, K.; Tsukaguchi, M.; Tatsumi, Y.; Ishikawa, J.; Tominaga, N.; Sakoda, H.; Take, H.; et al. Osaka Lymphoma Study Group. Tumour-associated macrophages in diffuse large B-cell lymphoma: A study of the Osaka Lymphoma Study Group. Histopathology 2012, 60, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.J.; Go, H.; Paik, J.H.; Kim, T.M.; Heo, D.S.; Kim, C.W.; Jeon, Y.K. An increase of M2 macrophages predicts poor prognosis in patients with diffuse large B-cell lymphoma treated with rituximab, cyclophosphamide, doxorubicin, vincristine and prednisone. Leuk. Lymphoma 2014, 55, 2466–2476. [Google Scholar] [CrossRef] [PubMed]
- Hasselblom, S.; Hansson, U.; Sigurdardottir, M.; Nilsson-Ehle, H.; Ridell, B.; Andersson, P.O. Expression of CD68+ tumor-associated macrophages in patients with diffuse large B-cell lymphoma and its relation to prognosis. Pathol. Int. 2008, 58, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, R.; Clear, A.J.; Mazzola, E.; Owen, A.; Greaves, P.; Wilson, A.; Matthews, J.; Lee, A.; Alvarez, R.; da Silva, M.G.; et al. Revisiting the immune microenvironment of diffuse large B-cell lymphoma using a tissue microarray and immunohistochemistry: Robust semi-automated analysis reveals CD3 and FoxP3 as potential predictors of response to R-CHOP. Haematologica 2015, 100, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Gelvez, J.C.; Salama, M.E.; Perkins, S.L.; Leavitt, M.; Inamdar, K.V. Prognostic Impact of Tumor Microenvironment in Diffuse Large B-Cell Lymphoma Uniformly Treated With R-CHOP Chemotherapy. Am. J. Clin. Pathol. 2016, 145, 514–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gao, K.; Lei, W.; Dong, L.; Xuan, Q.; Feng, M.; Wang, J.; Ye, X.; Jin, T.; Zhang, Z.; et al. Lymphocyte-to-monocyte ratio is associated with prognosis of diffuse large B-cell lymphoma: Correlation with CD163 positive M2 type tumor-associated macrophages, not PD-1 positive tumor-infiltrating lymphocytes. Oncotarget 2017, 8, 5414–5425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cencini, E.; Fabbri, A.; Schiattone, L.; Sicuranza, A.; Mecacci, B.; Granai, M.; Mancini, V.; Lazzi, S.; Bocchia, M.; Leoncini, L. Prognostic impact of tumor-associated macrophages, lymphocyte-to-monocyte and neutrophil-to-lymphocyte ratio in diffuse large B-cell lymphoma. Am. J. Blood Res. 2020, 10, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.C.; Liao, H.; Lin, S.X.; Xia, Y.; Wang, X.X.; Gao, Y.; Lin, Z.X.; Lu, J.B.; Huang, H.Q. High expression of tumor-infiltrating macrophages correlates with poor prognosis in patients with diffuse large B-cell lymphoma. Med. Oncol. 2012, 29, 2317–2322. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, Z.; Liu, J.; Zhu, F.; Wang, Z.; Wang, J.; Zhang, J.; Wang, H.; Zhai, Z. The prognostic value of tumour-associated macrophages in Non-Hodgkin’s lymphoma: A systematic review and meta-analysis. Scand. J. Immunol. 2020, 91, e12814. [Google Scholar] [CrossRef]
- Kridel, R.; Xerri, L.; Gelas-Dore, B.; Tan, K.; Feugier, P.; Vawda, A.; Canioni, D.; Farinha, P.; Boussetta, S.; Moccia, A.A.; et al. The Prognostic Impact of CD163-Positive Macrophages in Follicular Lymphoma: A Study from the BC Cancer Agency and the Lymphoma Study Association. Clin. Cancer Res. 2015, 21, 3428–3435. [Google Scholar] [CrossRef] [Green Version]
- Taskinen, M.; Karjalainen-Lindsberg, M.L.; Nyman, H.; Eerola, L.M.; Leppä, S. A high tumor-associated macrophage content predicts favorable outcome in follicular lymphoma patients treated with rituximab and cyclophosphamide-doxorubicin-vincristine-prednisone. Clin. Cancer Res. 2007, 13, 5784–5789. [Google Scholar] [CrossRef] [Green Version]
- Farinha, P.; Masoudi, H.; Skinnider, B.F.; Shumansky, K.; Spinelli, J.J.; Gill, K.; Klasa, R.; Voss, N.; Connors, J.M.; Gascoyne, R.D. Analysis of multiple biomarkers shows that lymphoma-associated macrophage (LAM) content is an independent predictor of survival in follicular lymphoma (FL). Blood 2005, 106, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Canioni, D.; Salles, G.; Mounier, N.; Brousse, N.; Keuppens, M.; Morchhauser, F.; Lamy, T.; Sonet, A.; Rousselet, M.C.; Foussard, C.; et al. High numbers of tumor-associated macrophages have an adverse prognostic value that can be circumvented by rituximab in patients with follicular lymphoma enrolled onto the GELA-GOELAMS FL-2000 trial. J. Clin. Oncol. 2008, 26, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.B.C.; Mendeville, M.; Redd, R.; Clear, A.J.; Bladergroen, R.; Calaminici, M.; Rosenwald, A.; Hoster, E.; Hiddemann, W.; Gaulard, P.; et al. Prognostic relevance of CD163 and CD8 combined with EZH2 and gain of chromosome 18 in follicular lymphoma: A study by the Lunenburg Lymphoma Biomarker Consortium. Haematologica 2017, 102, 1413–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, S.; Anderson, N.; Hainz, C.; Eckhardt, S.G.; Serkova, N.J. Imatinib (STI571)-mediated changes in glucose metabolism in human leukemia BCR-ABL-positive cells. Clin. Cancer Res. 2004, 10, 6661–6668. [Google Scholar] [CrossRef] [Green Version]
- Samudio, I.; Fiegl, M.; McQueen, T.; Clise-Dwyer, K.; Andreeff, M. The warburg effect in leukemia-stroma cocultures is mediated by mitochondrial uncoupling associated with uncoupling protein 2 activation. Cancer Res. 2008, 68, 5198–5205. [Google Scholar] [CrossRef] [Green Version]
- Barbato, A.; Scandura, G.; Puglisi, F.; Cambria, D.; La Spina, E.; Palumbo, G.A.; Lazzarino, G.; Tibullo, D.; Di Raimondo, F.; Giallongo, C.; et al. Mitochondrial Bioenergetics at the Onset of Drug Resistance in Hematological Malignancies: An Overview. Front. Oncol. 2020, 10, 604143. [Google Scholar] [CrossRef]
- Banerjee, P.; Zhang, R.; Ivan, C.; Galletti, G.; Clise-Dwyer, K.; Barbaglio, F.; Scarfò, L.; Aracil, M.; Klein, C.; Wierda, W.; et al. Trabectedin Reveals a Strategy of Immunomodulation in Chronic Lymphocytic Leukemia. Cancer Immunol. Res. 2019, 7, 2036–2051. [Google Scholar] [CrossRef]
- Cucè, M.; Gallo Cantafio, M.E.; Siciliano, M.A.; Riillo, C.; Caracciolo, D.; Scionti, F.; Staropoli, N.; Zuccalà, V.; Maltese, L.; Di Vito, A.; et al. Trabectedin triggers direct and NK-mediated cytotoxicity in multiple myeloma. J. Hematol. Oncol. 2019, 12, 32. [Google Scholar] [CrossRef]
- Li, K.; Xu, W.; Lu, K.; Wen, Y.; Xin, T.; Shen, Y.; Lv, X.; Hu, S.; Jin, R.; Wu, X. CSF-1R inhibition disrupts the dialog between leukaemia cells and macrophages and delays leukaemia progression. J. Cell. Mol. Med. 2020, 24, 13115–13128. [Google Scholar] [CrossRef]
- Komohara, Y.; Noyori, O.; Saito, Y.; Takeya, H.; Baghdadi, M.; Kitagawa, F.; Hama, N.; Ishikawa, K.; Okuno, Y.; Nosaka, K.; et al. Potential anti-lymphoma effect of M-CSFR inhibitor in adult T-cell leukemia/lymphoma. J. Clin. Exp. Hematop. 2018, 58, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Edwards, V.D.K.; Sweeney, D.T.; Ho, H.; Eide, C.A.; Rofelty, A.; Agarwal, A.; Liu, S.Q.; Danilov, A.V.; Lee, P.; Chantry, D.; et al. Targeting of colony-stimulating factor 1 receptor (CSF1R) in the CLL microenvironment yields antineoplastic activity in primary patient samples. Oncotarget 2018, 9, 24576–24589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, K.L.; Rinella, S.P.; Hess, N.J.; Turicek, D.P.; Kabakov, S.A.; Zhu, F.; Bouchlaka, M.N.; Olson, S.L.; Cho, M.M.; Quamine, A.E.; et al. CXCR4 allows T cell acute lymphoblastic leukemia to escape from JAK1/2 and BCL2 inhibition through CNS infiltration. Leuk. Lymphoma 2021, 62, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- McClellan, J.S.; Dove, C.; Gentles, A.J.; Ryan, C.E.; Majeti, R. Reprogramming of primary human Philadelphia chromosome-positive B cell acute lymphoblastic leukemia cells into nonleukemic macrophages. Proc. Natl. Acad. Sci. USA 2015, 112, 4074–4079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vos, S.; Forero-Torres, A.; Ansell, S.M.; Kahl, B.; Cheson, B.D.; Bartlett, N.L.; Furman, R.R.; Winter, J.N.; Kaplan, H.; Timmerman, J.; et al. A phase II study of dacetuzumab (SGN-40) in patients with relapsed diffuse large B-cell lymphoma (DLBCL) and correlative analyses of patient-specific factors. J. Hematol. Oncol. 2014, 7, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Luo, X.; Zhang, X.; Xu, L.; Wang, Y.; Yan, C.; Chen, H.; Chen, Y.; Han, W.; Wang, F.; et al. Preemptive interferon-alpha treatment could protect against relapse and improve long-term survival of ALL patients after allo-HSCT. Sci. Rep. 2020, 10, 20148. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, R.I.; Olalla Saad, S.T.; Azambuja, J.H. Artesunate Switches Monocytes to an Inflammatory Phenotype with the Ability to Kill Leukemic Cells. Int. J. Mol. Sci. 2021, 22, 608. [Google Scholar] [CrossRef]
- Kriston, C.; Hernádfői, M.; Plander, M.; Márk, Á.; Takács, F.; Czeti, Á.; Szalóki, G.; Szabó, O.; Matolcsy, A.; Barna, G. Lenalidomide abrogates the survival effect of bone marrow stromal cells in chronic lymphocytic leukemia. Hematol. Oncol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.P.; Alizadeh, A.A.; Tang, C.; Myklebust, J.H.; Varghese, B.; Gill, S.; Jan, M.; Cha, A.C.; Chan, C.K.; Tan, B.T.; et al. Anti-CD47 antibody synergizes with rituximab to promote phagocytosis and eradicate non-Hodgkin lymphoma. Cell 2010, 142, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Advani, R.; Flinn, I.; Popplewell, L.; Forero, A.; Bartlett, N.L.; Ghosh, N.; Kline, J.; Roschewski, M.; LaCasce, A.; Collins, G.P.; et al. CD47 Blockade by Hu5F9-G4 and Rituximab in Non-Hodgkin’s Lymphoma. N. Engl. J. Med. 2018, 379, 1711–1721. [Google Scholar] [CrossRef]
- Deng, Y.R.; Liu, W.B.; Lian, Z.X.; Li, X.; Hou, X. Sorafenib inhibits macrophage-mediated epithelial-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2016, 7, 38292–38305. [Google Scholar] [CrossRef] [PubMed]
- Vo, M.C.; Jung, S.H.; Chu, T.H.; Lee, H.J.; Lakshmi, T.J.; Park, H.S.; Kim, H.J.; Rhee, J.H.; Lee, J.J. Lenalidomide and Programmed Death-1 Blockade Synergistically Enhances the Effects of Dendritic Cell Vaccination in a Model of Murine Myeloma. Front. Immunol. 2018, 9, 1370. [Google Scholar] [CrossRef] [PubMed]
- Lauria, F.; Cencini, E.; Forconi, F. Alternative methods of cladribine administration. Leuk. Lymphoma 2011, 52, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, C.B.K.; Rudjord-Levann, A.M.; Gad, M.; Larsen, J.; Sellebjerg, F.; Pedersen, A.E. Cladribine inhibits secretion of pro-inflammatory cytokines and phagocytosis in human monocyte-derived M1 macrophages in-vitro. Int. Immunopharmacol. 2021, 91, 107270. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Tumor-associated macrophages polarization model in hematologic malignancies. TAM, tumor-associated macrophages; GM-CSF, granulocyte and monocyte colony stimulating factor; IFN, interferon; TNF, tumor necrosis factor; LPS, lipopolysaccharide; M-CSF, monocyte colony stimulating factor; IL, interleukin; TNF, tumor necrosis factor; NO, nitric oxide; ROS, reactive oxygen species; CXCL, chemokine ligands with C-X3-C motif; Th, T helper; TGF, tumor growth factor.
Figure 1.
Tumor-associated macrophages polarization model in hematologic malignancies. TAM, tumor-associated macrophages; GM-CSF, granulocyte and monocyte colony stimulating factor; IFN, interferon; TNF, tumor necrosis factor; LPS, lipopolysaccharide; M-CSF, monocyte colony stimulating factor; IL, interleukin; TNF, tumor necrosis factor; NO, nitric oxide; ROS, reactive oxygen species; CXCL, chemokine ligands with C-X3-C motif; Th, T helper; TGF, tumor growth factor.

Figure 2.
Visual mean of tumor-associated macrophages polarization.


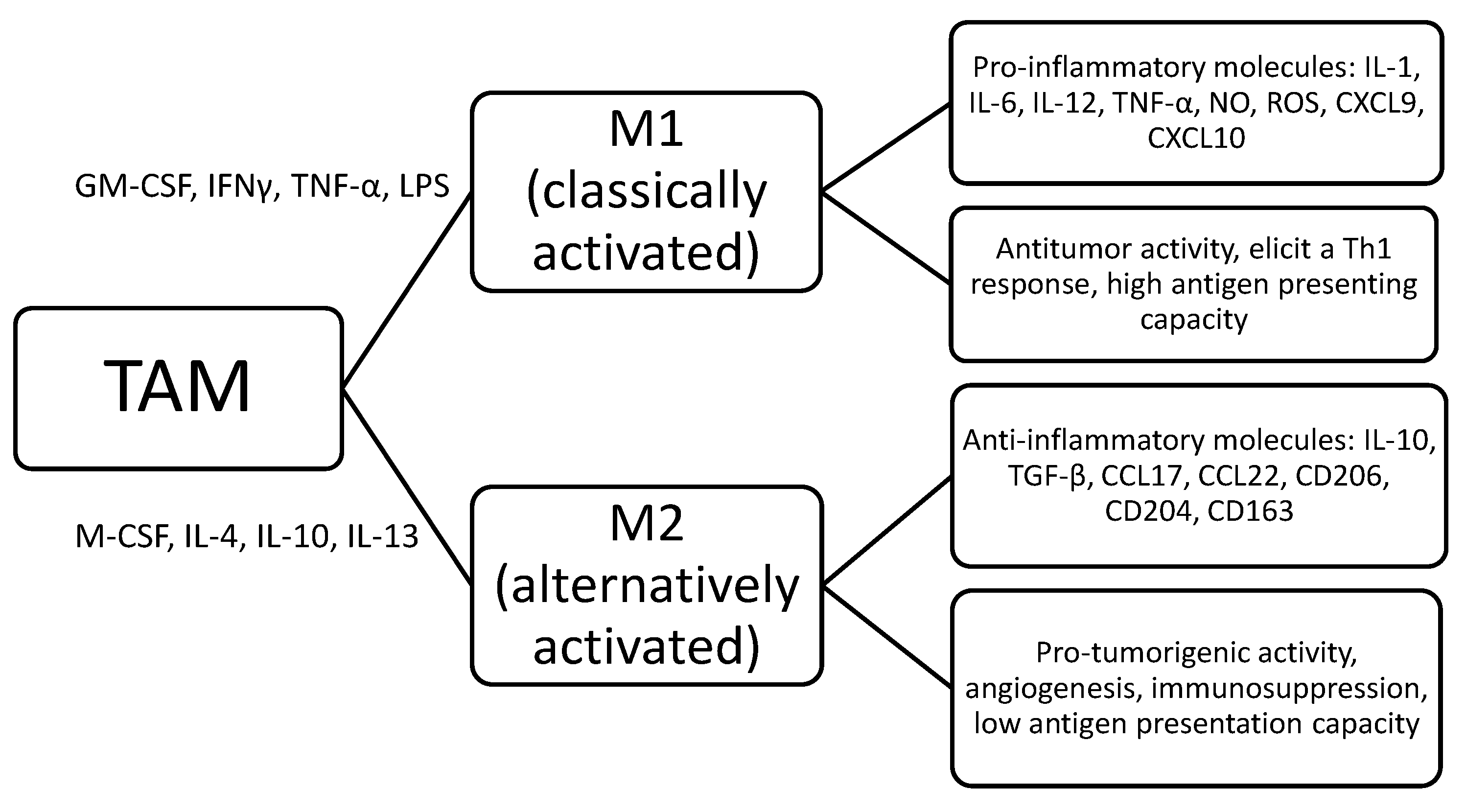{kind=link}
{kind=link}
Table 1.
Clinical studies about prognostic role of tumor-associated macrophages (TAM) in Hodgkin lymphoma.
Table 1.
Clinical studies about prognostic role of tumor-associated macrophages (TAM) in Hodgkin lymphoma.
| Reference | Number of Patients | Marker | Antibody | Cut-Off | Survival Correlation |
|---|---|---|---|---|---|
| Steidl et al. [115] | 166 | CD68 | KP1 | Steidl score (<5%, 5–25%, >25%) | Inferior PFS, DSS |
| Azambuja et al. [116] | 265 | CD68, CD163 | KP1 10D6 | Steidl score | No |
| Cencini et al. [117] | 200 | CD68 | KP1 | Steidl score | No |
| Agur et al. [118] | 98 | CD68 | PGM1 | Steidl score | No |
| Cuccaro et al. [119] | 102 | CD68 | PGM1 | Steidl score | Inferior PFS |
| Touati et al. [120] | 158 | CD68 | PGM1 | <25%, ≥25% | Inferior PFS, OS |
| Kayal et al. [121] | 100 | CD68 | (CD68/G2) | Steidl score <25%, >25%; <12.9%, >12.9% (the quartiles) <18.2%, >18.2% (median) | No No No |
| Klein et al. [122] | 88 | CD68, CD163 | KP1 10D6 | <25%, >25%; <25%, >25% | No Inferior OS |
| Yoon et al. [123] | 144 | CD68, CD163 | KP1 10D6 | <20%, >20% <20%, >20% | Inferior EFS, DSS, OS Inferior EFS, DSS, OS |
| Tan et al. [124] | 287 | CD68, CD163 | KP1 10D6 | <12.7%, >12.7% 16.8% (CD163) | Inferior FFS, OS Inferior FFS, OS |
| Greaves et al. [125] | 90 | CD68 | KP1 | <5%, 5–15%, >15% | Inferior FFTF, OS |
| Jakovic et al. [126] | 52 | CD68 | PGM1 | <25%, ≥25% | Inferior EFS, OS |
| Kamper et al. [127] | 288 | CD68, CD163 | KP1 10D6 | <7.8%, ≥7.8% <21.1%, ≥21.1% | Inferior EFS, OS Inferior EFS, OS |
| Werner et al. [128] | 76 | CD68, CD163 | PGM1 10D6 | <724, 725–937, ≥938 <769, 770–1325, ≥1326 | Inferior DSS Inferior DSS, OS (NS) |
| Karihtala et al. [129] | 130 | CD68, CD163 | Ab955 Ab188571 | Continuous variable Continuous variable | Inferior OS No |
| Mohamed et al. [130] | 81 | CD68 | Ab955 | <20%, 20–50%, >50% | Inferior DFS, OS |
Abbreviations: PFS, progression-free survival; DSS, disease-specific survival; OS, overall survival; EFS, event-free survival; FFS, failure-free survival; FFTF, freedom from treatment failure; NS, not significant.
Table 2.
Clinical studies about prognostic role of tumor-associated macrophages (TAM) in diffuse large B-cell lymphoma.
Table 2.
Clinical studies about prognostic role of tumor-associated macrophages (TAM) in diffuse large B-cell lymphoma.
| Reference | Number of Patients | Marker | Antibody | Cut-Off | Survival Correlation |
|---|---|---|---|---|---|
| Riihijarvi et al. [139] | 59 | CD68, CD163 | KP1 10D6 | 26/hpf Various | Better PFS, OS (R-chemo) No |
| Marchesi et al. [140] | 61 | CD68/CD163 | Doublestain (NS) | double-marked cells/ total macrophages > 0.2 | Inferior DFS, OS |
| Wada et al. [141] | 101 | CD68 CD68/CD163 | PGM1 Doublestain (G2 EnVision) | <average, >average <average, >average | Inferior OS Inferior OS |
| Nam et al. [142] | 109 | CD68 CD163 CD68/CD163 | PGM1 10D6 Doublestain (automated system Ventana) | Best cut-off Best cut-off Best cut-off | Better OS Inferior OS Inferior OS |
| Hasselblom et al. [143] | 176 | CD68 | KP1 | <median, >median | No |
| Coutinho et al. [144] | 161 | CD68 | KP1 | <median, >median | No |
| Gomez-Gelvez et al. [145] | 74 | CD68 | KP1 | <2%, >2% | Inferior EFS |
| Wang et al. [146] | 355 | CD163 | 10D6 | >9.5% | Inferior PFS, OS |
| Cencini et al. [147] | 37 | CD68 CD163 CD68/CD163 | PGM1 10D6 Doublestain (automated system Dako) | Steidl score | No No Inferior PFS (IPI ≥ 2) |
| Cai et al. [148] | 112 | CD68 | KP1 | 200/hpf | Inferior PFS |
Abbreviations: PFS, progression-free survival; OS, overall survival; hpf, high-power field; R, rituximab; DFS, disease-free survival; NS, not significant; EFS, event-free survival; IPI, international prognostic index.
Table 3.
The role of tumor-associated macrophages (TAM) as treatment target in hematologic malignancies.
Table 3.
The role of tumor-associated macrophages (TAM) as treatment target in hematologic malignancies.
| Reference | Treatment | Disease | Mechanism of Action | Type of Study | Results |
|---|---|---|---|---|---|
| Benerjee et al. [158] | Trabectedin | CLL | Macrophage killing due to CCL2-CCR2 signalling axis inhibition, antiangiogenic | Leukemic mouse model Human CLL cells | Apoptosis trigger MDSC and TAM depletion. M1 shift Memory T-cells increase Increased mice survival |
| Cucè et al. [159] | Trabectedin | MM | Macrophage killing due to CCL2-CCR2 signalling axis inhibition, antiangiogenic | Human MM cells | Apoptosis trigger VEGF depletion NK cells upregulation |
| Polk et al. [74] | CSF-1R signalling inhibitor | CLL | CSF-1R signalling Inhibition | Human CLL cells | NLC depletion Reduced CLL cells Viability |
| Li et al. [160] | BLZ-945 | T-ALL | CSF-1R signalling Inhibition | Mouse model | Inhibition of BMDMs viability, LAMs polarization blocking and depletion. Combination of CSF-1R inhibitor and VCR increased the survival of T-ALL mice. |
| Komohara et al. [161] | PLX3397 | ATLL | CSF-1R signalling Inhibition | ATLL cells | Inhibition of ATLL cell proliferation Apoptosis induction |
| Edwards et al. [162] | GW-2580, ARRY-382 | CLL | CSF-1R signaling Inhibition | Human CLL cells | NLC depletion BCR signaling disruption (together with ibrutinib or idelalisib) |
| Walker et al. [163] | CXCR4 inhibitor plerixafor | T-ALL | CXCR4/CXCL12 axis inhibition | In vitro Xenograft model | Improved survival (together with ruxolitinib and venetoclax) M1 shift of TAM |
| McClellan et al. [164] | Exposure to myeloid differentiation promoting cytokines | Ph+ B-ALL | B-ALL blasts reprogram into Macrophage | In vitro Xenograft model | Eliminates B-ALL cells leukemogenicity. |
| De Vos et al. [165] | Dacetuzumab | R/R DLBCL | anti-CD40 mAb | Phase II study, 46 patients | Disease control (CR/PR/SD) 37% |
| Liu et al. [166] | Preemptive IFN-α | ALL | TAM reprogramming | 47 B-ALL 21 T-ALL (MRD+ after allo-SCT) | M1 shift of TAM, 76% of cases became MRD- |
| Mancuso et al. [167] | Artesunate | Leukemia | TAM reprogramming JAK2/STAT3 Downregulation | Human monocytes, leukemic cells | Increase in inflammatory monocytes. After contact with monocytes, in vitro apoptosis of leukemic cells increased |
| Kriston et al. [168] | Lenalidomide | CLL | Abrogation of BMSC survival effect | CLL cells | Abrogation of anti-apoptotic effect of BMSCs on CLL cells. |
| Chao et al. [169] | Anti-CD47 mAb | ALL | Enabled phagocytosis of tumor cells by TAM | Mouse model | Induces remissions in ALL-engrafted mice |
| Advani et al. [170] | Anti-CD47 mAb Hu5F9-G4 | R/R DLBCL or FL | Enabled phagocytosis of tumor cells by TAM | Phase 1b-2 study | ORR 50% CR rate 36% |
Abbreviations: CCL, chemokine ligands with C-C motif; CCR, C-C chemokine receptor; CLL, chronic lymphocytic leukemia; MDSC, myeloid-derived suppressor cells; TAM, tumor-associated macrophages; MM, multiple myeloma; VEGF, vascular endothelial growth factor; NK, natural killer; CSF-1R, colony stimulating factor-1 receptor; NLC, nurse-like cells; ALL, acute lymphoblastic leukemia; BMDM, bone marrow-derived macrophages; LAM, leukemia-associated macrophages; VCR, vincristine; ATLL, adult T-cell lymphoma leukemia; BCR, B-cell receptor; CXCR, chemokine receptor; Ph, Philadelphia; mAb, monoclonal antibody; CR, complete response; PR, partial response; SD, stable disease; IFN, interferon; allo-SCT, allogeneic stem-cell transplantation; MRD, minimal residual disease; JAK, Janus kinase, BMSC, bone marrow stromal cells; DLBCL, diffuse large B-cell lymphoma; FL, follicular lymphoma; ORR, overall response rate.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cencini, E.; Fabbri, A.; Sicuranza, A.; Gozzetti, A.; Bocchia, M. The Role of Tumor-Associated Macrophages in Hematologic Malignancies. Cancers 2021, 13, 3597. https://doi.org/10.3390/cancers13143597
AMA Style
Cencini E, Fabbri A, Sicuranza A, Gozzetti A, Bocchia M. The Role of Tumor-Associated Macrophages in Hematologic Malignancies. Cancers. 2021; 13(14):3597. https://doi.org/10.3390/cancers13143597
Chicago/Turabian StyleCencini, Emanuele, Alberto Fabbri, Anna Sicuranza, Alessandro Gozzetti, and Monica Bocchia. 2021. "The Role of Tumor-Associated Macrophages in Hematologic Malignancies" Cancers 13, no. 14: 3597. https://doi.org/10.3390/cancers13143597
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.