
Modulation of Skin Inflammatory Responses by Aluminum Adjuvant
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Antibodies and Dyes
2.2. Animal Cares and Animal Models
2.3. Histology and Immunohistochemistry (IHC)
2.4. Flow Cytometry and Analysis (FACS)
2.5. Quantitative Reverse Transcription-Quantitative PCR (qRT-PCR) Analyses
2.6. In Vitro T Cell Differentiation
2.7. Neutrophil and Macrophage Cultures
2.8. Statistics
3. Results
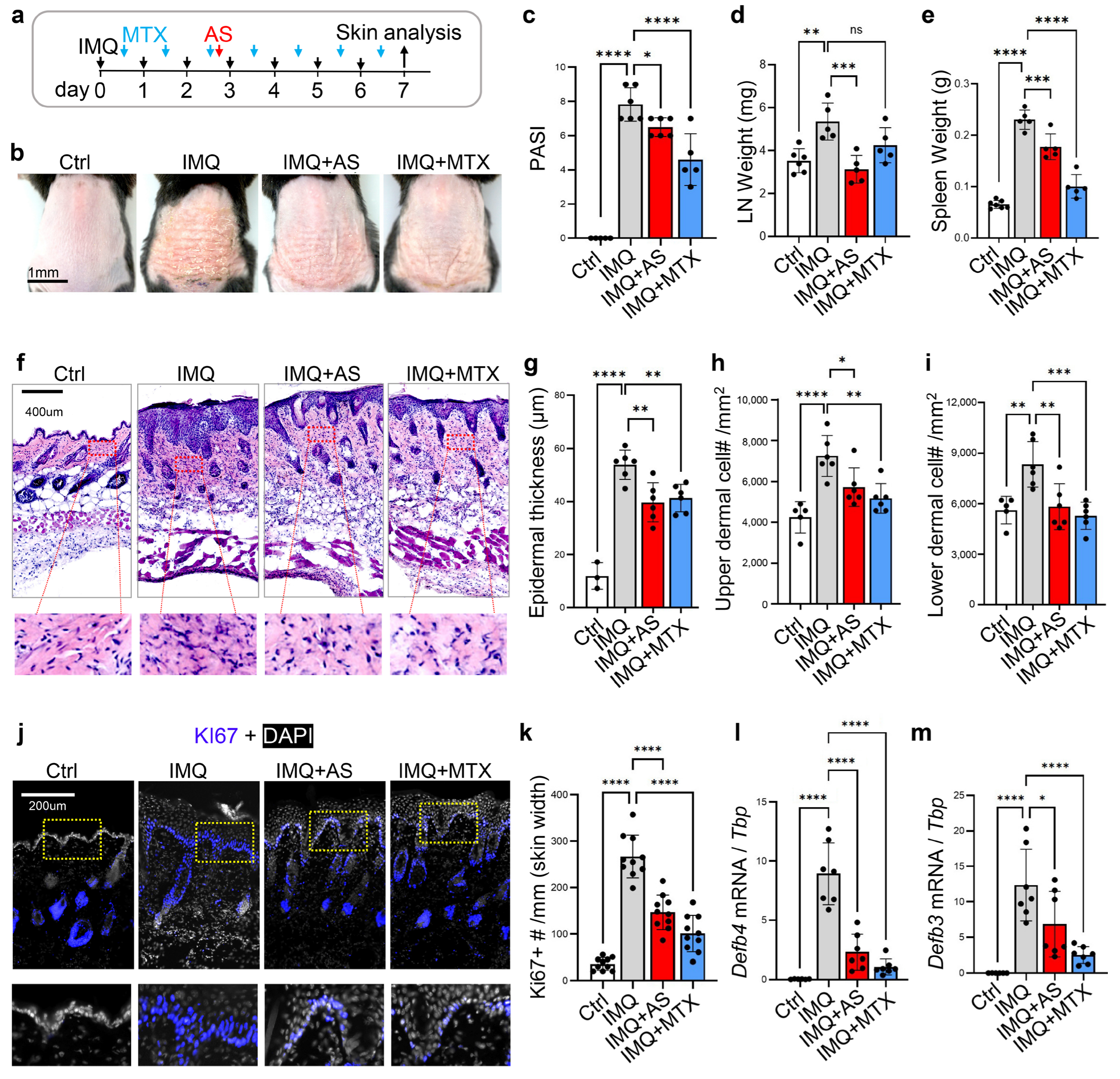
3.1. Administration of Aluminum Salt Alleviates the Development of Psoriasis-like Skin Inflammation in Mice
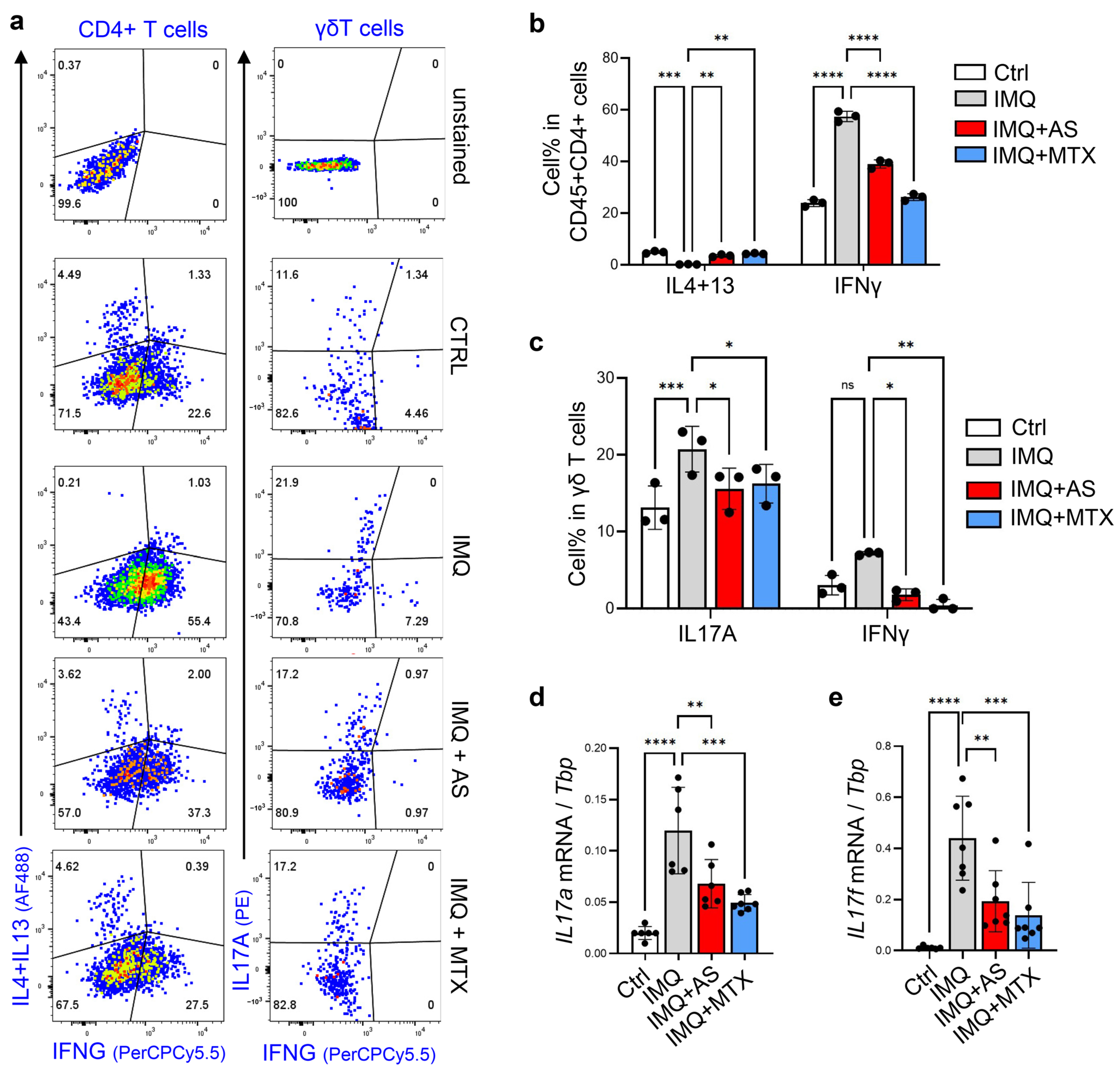
3.2. Administration of Aluminum Salt Inhibited the Development of Th1 and Th17 Immune Responses in the IMQ-Induced Psoriasis Model
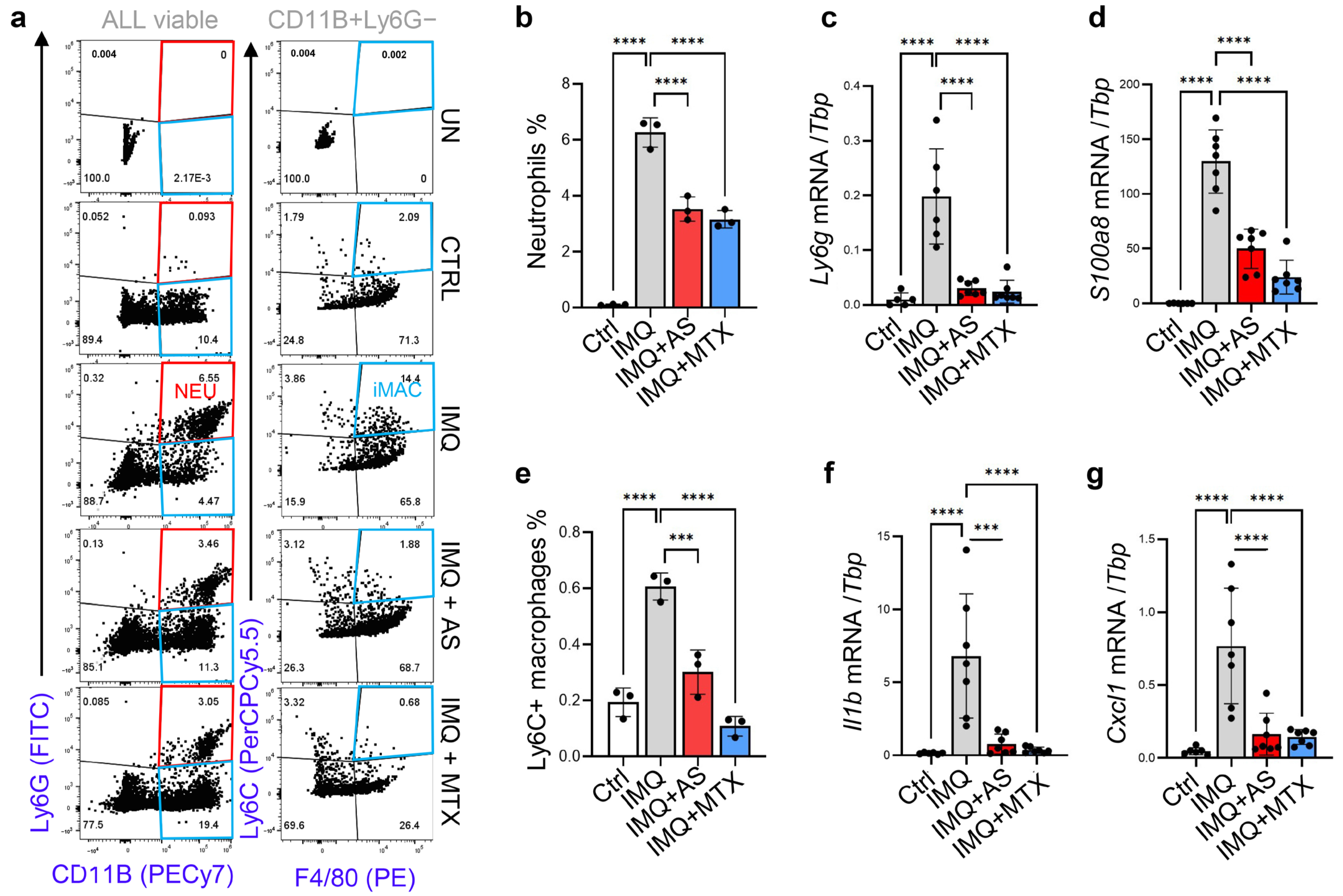
3.3. Administration of Aluminum Salt Reduced the Recruitment and Activation of Myeloid Cells
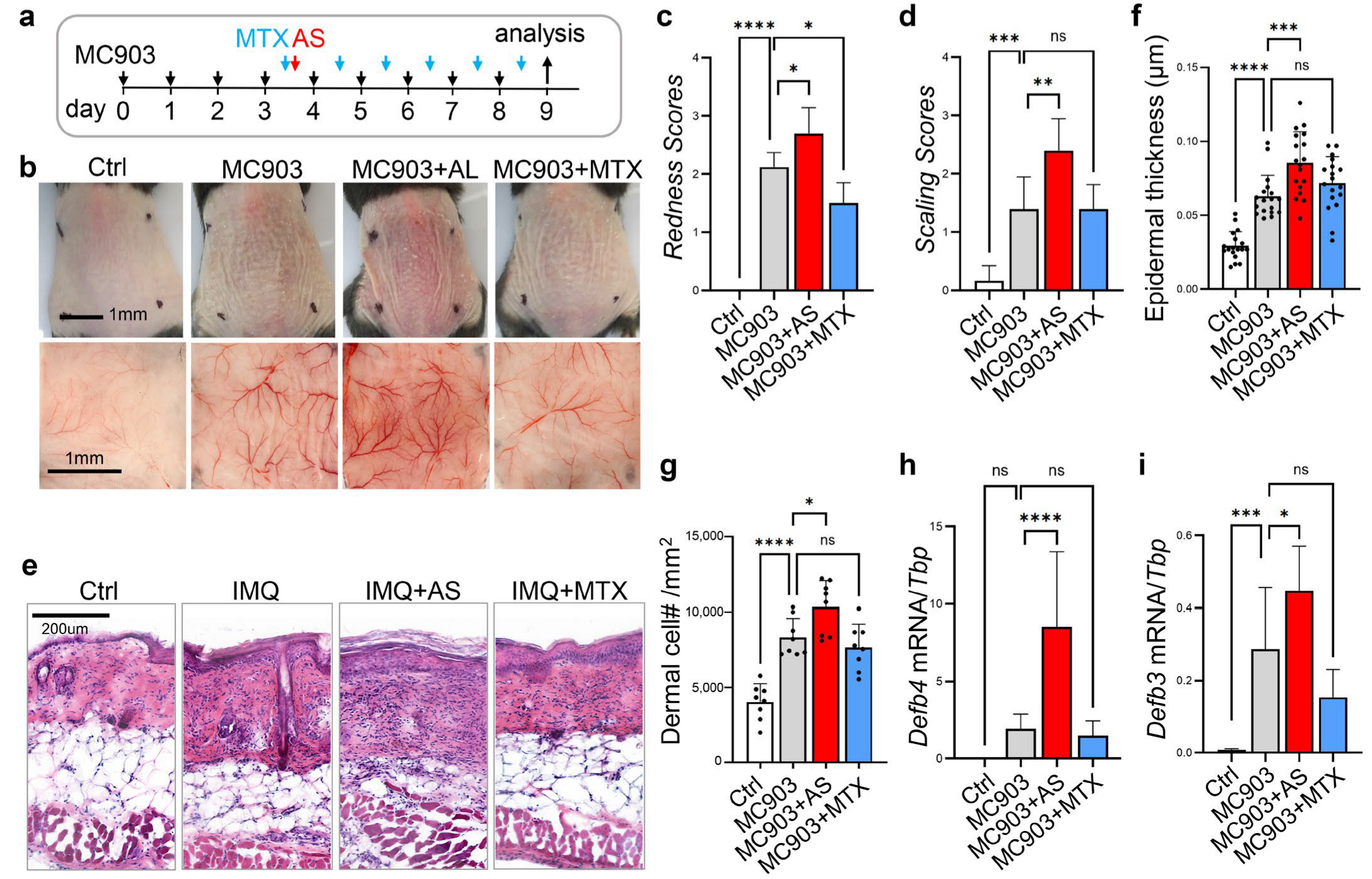
3.4. Administration of Aluminum Salt Promoted the Development of MC903-Induced Atopic Dermatitis-like Skin Inflammation
3.5. Effect of Aluminum Salt Injection in Modulating the MC903-Mediated Activation of T Cells or Myeloid Cells
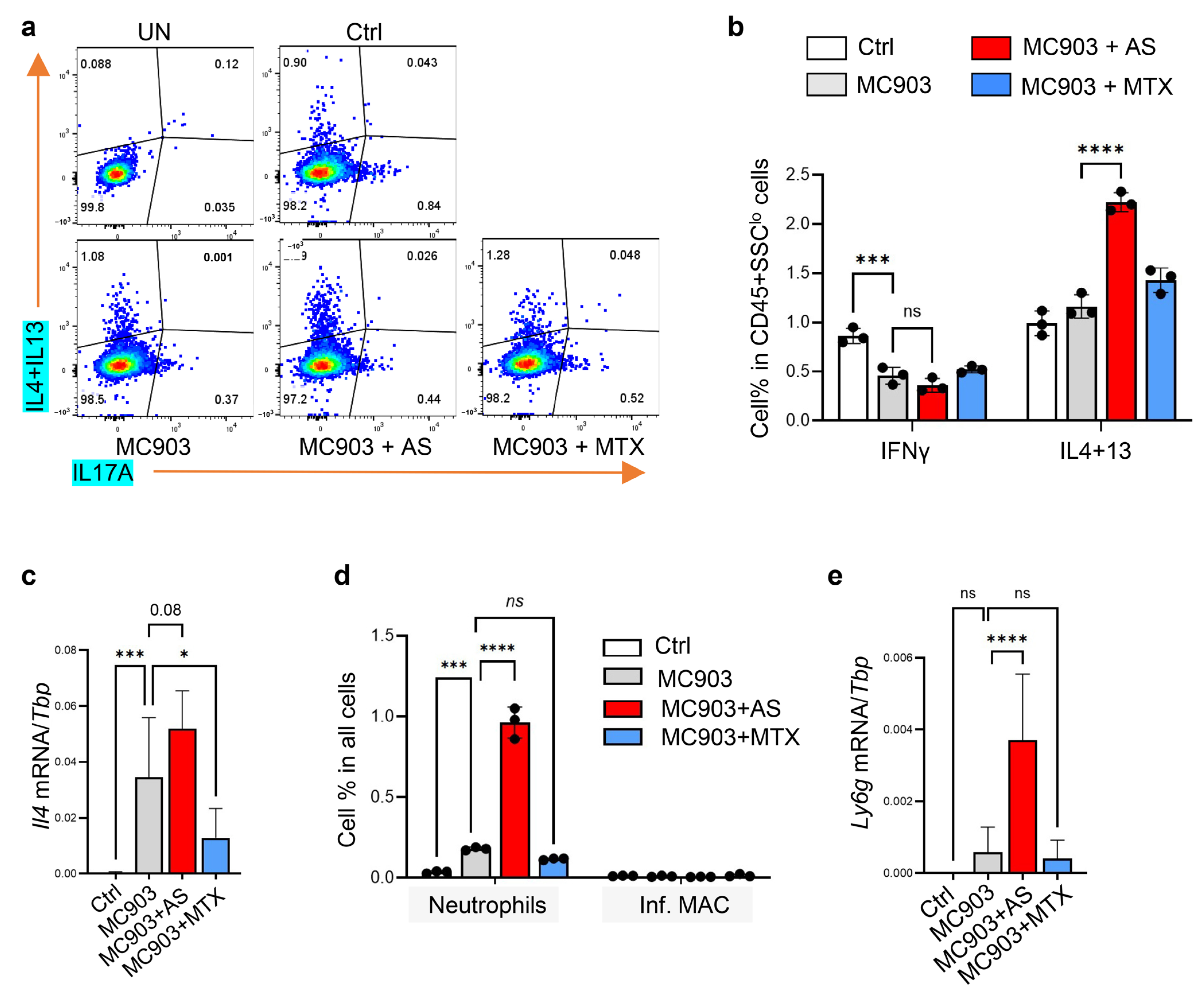
3.5.1. Administration of Aluminum Salt Promoted the Development of Type 2 Immune Response in the MC903-Induced Dermatitis Model
3.5.2. Administration of Aluminum Salt Altered Myeloid Cell Activation in the MC903-Induced Dermatitis Model
3.6. The In Vitro Effect of Aluminum Salt in Modulating T Cell Differentiation and Myeloid Cell Activation
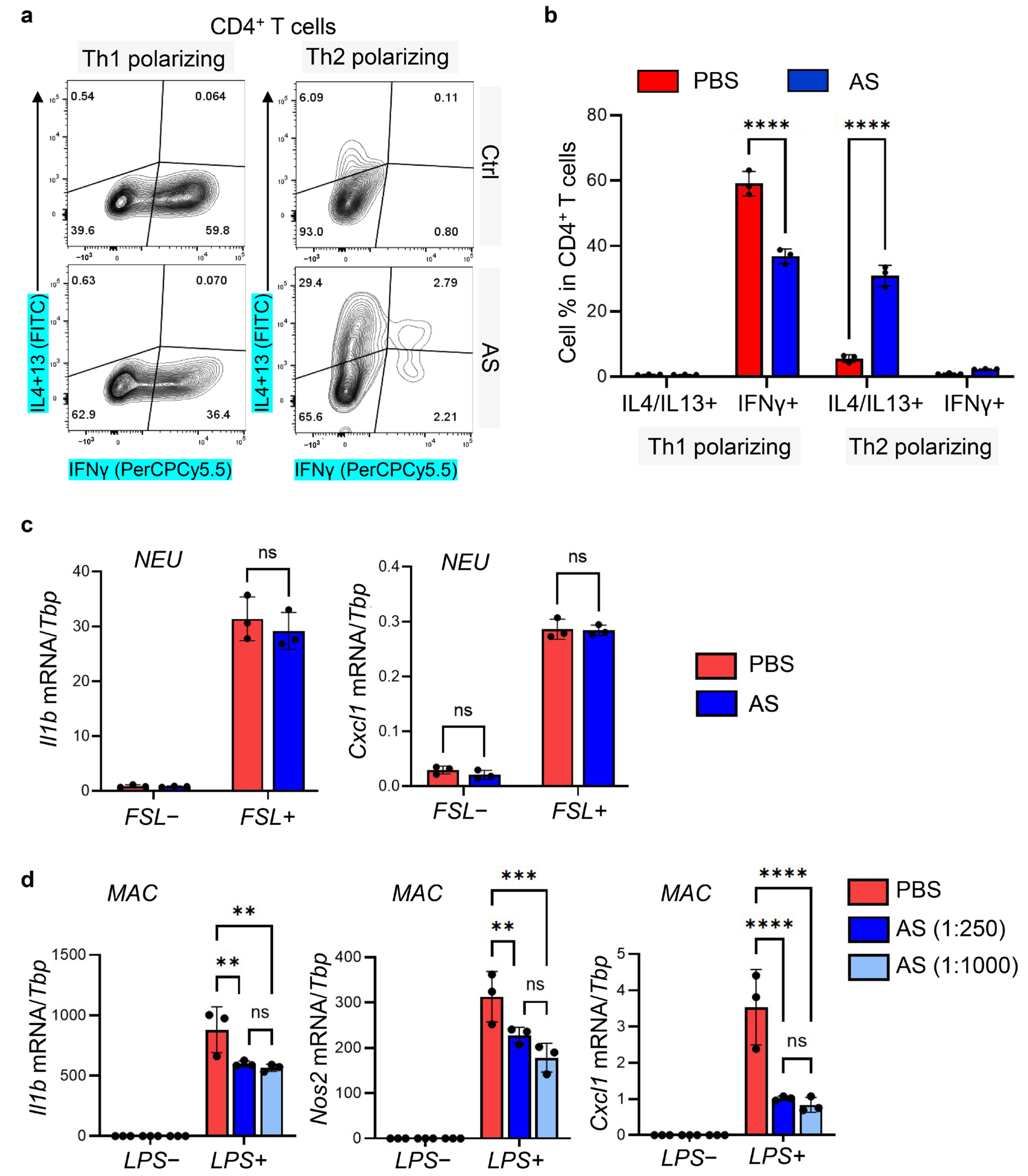
3.6.1. Aluminum Salt Inhibited Th1 but Promoted Th2 Polarization during In Vitro T Cell Differentiation
3.6.2. Aluminum Salt Inhibited Macrophage but Not Neutrophil Activation In Vitro
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, L.; Liu, W.; Zhang, L.J. The Role of Toll-Like Receptors in Skin Host Defense, Psoriasis, and Atopic Dermatitis. J. Immunol. Res. 2019, 2019, 1824624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J. Recent progress at the psoriasis and atopic dermatitis research front: An experimental dermatology perspective. Exp. Derm. 2021, 30, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Inui, N.; Chida, K.; Suda, T.; Nakamura, H. TH1/TH2 and TC1/TC2 profiles in peripheral blood and bronchoalveolar lavage fluid cells in pulmonary sarcoidosis. J. Allergy Clin. Immunol. 2001, 102, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.; Liu, X.; Fang, L.; Chen, X.; Guo, T.; Zhang, J. The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease. Cell. Mol. Immunol. 2010, 7, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Eyerich, S.; Onken, A.T.; Weidinger, S.; Franke, A.; Nasorri, A.; Pennino, D.; Grosber, M.; Pfab, F.; Schmidt-Weber, C.B.; Mempel, M.; et al. Mutual antagonism of T cells causing psoriasis and atopic eczema. N. Engl. J. Med. 2011, 365, 231–238. [Google Scholar] [CrossRef]
- Guo, Z.; Yang, Y.; Liao, Y.; Shi, Y.; Zhang, L.-j. Emerging Roles of Adipose Tissue in the Pathogenesis of Psoriasis and Atopic Dermatitis in Obesity. JID Innov. 2022, 2, 100064. [Google Scholar] [CrossRef]
- Sabat, R.; Wolk, K.; Loyal, L.; Döcke, W.-D.; Ghoreschi, K. T cell pathology in skin inflammation. In Seminar Immunopathology; Springer: Berlin/Heidelberg, Germany, 2019; Volume 41, pp. 359–377. [Google Scholar]
- Ho, A.W.; Kupper, T.S. T cells and the skin: From protective immunity to inflammatory skin disorders. Nat. Rev. Immunol. 2019, 19, 490–502. [Google Scholar] [CrossRef]
- Volk, V.K.; Bunney, W.E. Diphtheria Immunization with fluid toxoid and alum-precipitated toxoid. Am. J. Public Health Nations Health 1942, 32, 690–699. [Google Scholar] [CrossRef]
- Jones, F.G.; Moss, J.M. Studies on Tetanus Toxoid: I. The Antitoxic Titer of Human Subjects Following Immunization with Tetanus Toxoid and Tetanus Alum Precipitated Toxoid. J. Immunol. 1936, 30, 115–125. [Google Scholar] [CrossRef]
- Sokolovska, A.; Hem, S.L.; HogenEsch, H. Activation of dendritic cells and induction of CD4(+) T cell differentiation by aluminum-containing adjuvants. Vaccine 2007, 25, 4575–4585. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Serre, K.; Toellner, K.-M.; Khan, M.; Alexander, J.; Brombacher, F.; MacLennan, I.C.M. Pinpointing IL-4-independent acquisition and IL-4-influenced maintenance of Th2 activity by CD4 T cells. Eur. J. Immunol. 2004, 34, 686–694. [Google Scholar] [CrossRef]
- Comoy, E.E.; Capron, A.; Thyphronitis, G. In vivo induction of type 1 and 2 immune responses against protein antigens. Int. Immunol. 1997, 9, 523–531. [Google Scholar] [CrossRef]
- Brewer, J.M.; Conacher, M.; Satoskar, A.; Bluethmann, H.; Alexander, J. In interleukin-4-deficient mice, alum not only generates T helper 1 responses equivalent to freund’s complete adjuvant, but continues to induce T helper 2 cytokine production. Eur. J. Immunol. 1996, 26, 2062–2066. [Google Scholar] [CrossRef]
- Nie, M.; Liu, Y.; Huang, X.; Zhang, Z.; Zhao, Q. Microtiter Plate-Based Differential Scanning Fluorimetry: A High-Throughput Method for Efficient Formulation Development. J. Pharm. Sci. 2022, 111, 2397–2403. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, X.; Yuan, L.; Wang, S.; Zhang, Y.; Xiong, H.; Chen, R.; Ma, J.; Qi, R.; Nie, M.; et al. A recombinant spike protein subunit vaccine confers protective immunity against SARS-CoV-2 infection and transmission in hamsters. Sci. Transl. Med. 2021, 13, eabg1143. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Yang, M.; Li, Y.; Zheng, Z.; Zhang, X.; Lin, Q.; Wang, Y.; Li, S.; Xia, N.; et al. Comparable quality attributes of hepatitis E vaccine antigen with and without adjuvant adsorption-dissolution treatment. Hum. Vaccines Immunother. 2015, 11, 1129–1139. [Google Scholar] [CrossRef]
- Zong, J.; Cheng, J.; Fu, Y.; Song, J.; Pan, W.; Yang, L.; Zhang, T.; Zhou, M. Serum Metabolomic Profiling Reveals the Amelioration Effect of Methotrexate on Imiquimod-Induced Psoriasis in Mouse. Front. Pharm. 2020, 11, 558629. [Google Scholar] [CrossRef]
- Neu, S.D.; Strzepa, A.; Martin, S.; Sorci-Thomas, M.G.; Pritchard, K.A., Jr.; Dittel, B.N. Myeloperoxidase Inhibition Ameliorates Plaque Psoriasis in Mice. Antioxidants 2021, 10, 1338. [Google Scholar] [CrossRef]
- Langley, R.G.; Ellis, C.N. Evaluating psoriasis with Psoriasis Area and Severity Index, Psoriasis Global Assessment, and Lattice System Physician’s Global Assessment. J. Am. Acad. Derm. 2004, 51, 563–569. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shun, Z.; Silverberg, A.; Chang, C.K.; Ouyang, P. Dunnett’s many-to-one test and least square means. J. Biopharm. Stat. 2003, 13, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic. Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Ishizaki, M.; Akimoto, T.; Muromoto, R.; Yokoyama, M.; Ohshiro, Y.; Sekine, Y.; Maeda, H.; Shimoda, K.; Oritani, K.; Matsuda, T. Involvement of tyrosine kinase-2 in both the IL-12/Th1 and IL-23/Th17 axes in vivo. J. Immunol. 2011, 187, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Baylor, N.W.; Egan, W.; Richman, P. Aluminum salts in vaccines--US perspective. Vaccine 2002, 20, S18–S23. [Google Scholar] [CrossRef]
- Wang, R.; Sun, C.; Ma, J.; Yu, C.; Kong, D.; Chen, M.; Liu, X.; Zhao, D.; Gao, S.; Kou, S.; et al. A Bivalent COVID-19 Vaccine Based on Alpha and Beta Variants Elicits Potent and Broad Immune Responses in Mice against SARS-CoV-2 Variants. Vaccines 2022, 10, 702. [Google Scholar] [CrossRef]
- Mao, Q.; Dong, C.; Li, X.; Gao, Q.; Guo, Z.; Yao, X.; Wang, Y.; Gao, F.; Li, F.; Xu, M.; et al. Comparative analysis of the immunogenicity and protective effects of inactivated EV71 vaccines in mice. PLoS ONE 2012, 7, e46043. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human beta-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Investig. Derm. 2007, 127, 594–604. [Google Scholar] [CrossRef]
- Qi, C.; Wang, Y.; Li, P. Gamma Delta T Cells and Their Pathogenic Role in Psoriasis. Front. Immunol. 2021, 12, 627139. [Google Scholar] [CrossRef]
- Deng, Y.; Chang, C.; Lu, Q. The Inflammatory Response in Psoriasis: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 377–389. [Google Scholar] [CrossRef]
- Rahman, K.; Vengrenyuk, Y.; Loke, P.; Fisher, E.A. Inflammatory Ly6Chi monocytes and their conversion to M2 macrophages drive atherosclerosis regression. J. Clin. Investig. 2017, 127, 2904–2915. [Google Scholar] [CrossRef]
- Cai, Y.; Xue, F.; Quan, C.; Qu, M.; Liu, N.; Zhang, Y.; Fleming, C.; Hu, X.; Zhang, H.-G.; Weuchselbaum, R.; et al. A Critical Role of the IL-1beta-IL-1R Signaling Pathway in Skin Inflammation and Psoriasis Pathogenesis. J. Investig. Derm. 2019, 139, 146–156. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Leyva-Castillo, J.M.; Hener, P.; Michea, P.; Karasuyama, H.; Chan, S.; Soulmelis, V.; Li, M. Skin thymic stromal lymphopoietin initiates Th2 responses through an orchestrated immune cascade. Nat. Commun. 2013, 4, 2847. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Corrigendum: Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS-) vs. Alternatively Activated Macrophages. Front. Immunol. 2020, 11, 234. [Google Scholar] [CrossRef]
- Leventhal, J.S.; Berger, E.M.; Brauer, J.A.; Cohen, D.E. Hypersensitivity reactions to vaccine constituents: A case series and review of the literature. Dermatitis 2012, 23, 102–109. [Google Scholar] [CrossRef]
- Engler, R.J.; Kenner, J.; Leung, D.Y. Smallpox vaccination: Risk considerations for patients with atopic dermatitis. J. Allergy Clin. Immunol. 2022, 110, 357–365. [Google Scholar] [CrossRef]
- Ayasse, M.; Ahmed, A.; McCullum, C.; Espinosa, M.L.; Paller, A.S.; Silverberg, J.I. Vaccines do not cause atopic dermatitis: A systematic review and meta-analysis. Vaccine 2021, 39, 1805–1811. [Google Scholar] [CrossRef]
- Enriquez, R.; Addington, W.; Davis, F.; Freels, S.; Park, C.L.; Hershow, R.S.; Persky, V. The relationship between vaccine refusal and self-report of atopic disease in children. J. Allergy Clin. Immunol. 2005, 115, 737–744. [Google Scholar] [CrossRef]
- Koppen, S.; de Groot, R.; Neijens, H.J.; Nagelkerke, N.; van Eden, W.; Rümke, H.C. No epidemiological evidence for infant vaccinations to cause allergic disease. Vaccine 2004, 22, 3375–3385. [Google Scholar] [CrossRef]
- Linneberg, A.; Jacobsen, R.K.; Jespersen, L.; Abildstrøm, S.Z. Association of subcutaneous allergen-specific immunotherapy with incidence of autoimmune disease, ischemic heart disease, and mortality. J. Allergy Clin. Immunol. 2012, 129, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Mistry, B.D.; DeKoven, J.G. Widespread cutaneous eruption after aluminum-containing vaccination: A case report and review of current literature. Pediatr. Derm. 2021, 38, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Aquino, M.R.; Bingerman, T.A.; Nanda, A.; Maples, K.M. Delayed allergic skin reactions to vaccines. In Allergy and Asthma Proceedings; OceanSide Publications: East Providence, RI, USA, 2022; Volume 43, pp. 20–29. [Google Scholar]
- Hoffmann, S.S.; Thiesson, E.M.; Johansen, J.D.; Hviid, A. Risk factors for granulomas in children following immunization with aluminium-adsorbed vaccines: A Danish population-based cohort study. Contact Dermat. 2022, 87, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.C.; Bartenstein, D.W.; Tajmir, S.H.; Song, J.S.; Hawryluk, E.B. Delayed-type hypersensitivity to vaccine aluminum adjuvant causing subcutaneous leg mass and urticaria in a child. Pediatr. Derm. 2018, 35, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Thierry-Carstensen, B.; Stellfeld, M. Itching nodules and hypersensitivity to aluminium after the use of adsorbed vaccines from SSI. Vaccine 2004, 22, 1845. [Google Scholar] [CrossRef]
- Shi, Y.; HogenEsch, H.; Regnier, F.E.; Hem, S.L. Detoxification of endotoxin by aluminum hydroxide adjuvant. Vaccine 2001, 19, 1747–1752. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Panel A: Immune Cell Analysis | Panel B: T Cell Analysis |
|---|---|
| APC anti-CD11C | AF488 anti-IL4 * |
| PerCP-Cy5.5 anti-LY6C | APC anti-TCRγ/δ |
| PECy7 anti-CD11B | PerCP-Cy5.5 anti-IFNγ * |
| APC/Cyanine7 anti-CD3 | AF700 anti-CD4 |
| AF488 anti-LY6G | AF488 anti-IL13 * |
| PE an-ti-F4/80 | PECy7 anti-CD45 |
| AF700 anti-MHCII | APC/Cyanine7 anti-CD3 |
| Gene | Strand | Primer Sequence |
|---|---|---|
| Tbp | Forward | CCTTGTACCCTTCACCAATGAC |
| Reverse | ACAGCCAAGATTCACGGTAGA | |
| Defb4 | Forward | TGGTGCTGCTGTCTCCACTTGC |
| Reverse | AGGGCACGGACCCCAGCATA | |
| Defb3 | Forward | GGTGCTGCTGTCTCCACCTGC |
| Reverse | TGCACCGATTCCAGCATCTGCC | |
| Il17a | Forward | ACGCGCAAACATGAGTCCAGGG |
| Reverse | TGAGGGATGATCGCTGCTGCCT | |
| Il17f | Forward | ACGTGAATTCCAGAACCGCT |
| Reverse | TGATGCAGCCTGAGTGTCTG | |
| Ly6g | Forward | GACTTCCTGCAACACAACTACC |
| Reverse | ACAGCATTACCAGTGATCTCAGT | |
| S100a8 | Forward | AAATCACCATGCCCTCTACAAG |
| Reverse | CCCACTTTTATCACCATCGCAA | |
| Il1b | Forward | GAAATGCCACCTTTTGACAGTG |
| Reverse | TGGATGCTCTCATCAGGACAG | |
| Cxcl1 | Forward | CACCCGCTCGCTTCTCTG |
| Reverse | TCTTGAGGTGAATCCCAGCC | |
| Il4 | Forward | GAGCCATATCCACGGATGCGAC |
| Reverse | ATGCGAAGCACCTTGGAAGCCC | |
| Nos2 | Forward | GTTCTCAGCCCAACAATACAAGA |
| Reverse | GTGGACGGGTCGATGTCAC | |
| Col1a1 | Forward | GCTCCTCTTAGGGGCCACT |
| Reverse | ATTGGGGACCCTTAGGCCAT | |
| Il22 | Forward | CCTACATGCAGGAGGTGGTG |
| Reverse | AAACAGCAGGTCCAGTTCCC | |
| Il23 | Forward | ATGCTGGATTGCAGAGCAGTA |
| Reverse | ACGGGGCACATTATTTTTAGTCT | |
| Saa3 | Forward | TGCCATCATTCTTTGCATCTTGA |
| Reverse | CCGTGAACTTCTGAACAGCCT | |
| Defb14 | Forward | TGGTGCCTGCTCCAGGGGAC |
| Reverse | CAGCACACCGGCCACCTCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Sun, L.; Nie, M.; Li, J.; Huang, X.; Heng, S.; Zhang, W.; Xia, T.; Guo, Z.; Zhao, Q.; et al. Modulation of Skin Inflammatory Responses by Aluminum Adjuvant. Pharmaceutics 2023, 15, 576. https://doi.org/10.3390/pharmaceutics15020576
Liao Y, Sun L, Nie M, Li J, Huang X, Heng S, Zhang W, Xia T, Guo Z, Zhao Q, et al. Modulation of Skin Inflammatory Responses by Aluminum Adjuvant. Pharmaceutics. 2023; 15(2):576. https://doi.org/10.3390/pharmaceutics15020576
Chicago/Turabian StyleLiao, Yanhang, Lixiang Sun, Meifeng Nie, Jiacheng Li, Xiaofen Huang, Shujun Heng, Wenlu Zhang, Tian Xia, Zhuolin Guo, Qinjian Zhao, and et al. 2023. "Modulation of Skin Inflammatory Responses by Aluminum Adjuvant" Pharmaceutics 15, no. 2: 576. https://doi.org/10.3390/pharmaceutics15020576