
Advances in the Prophylaxis of Respiratory Infections by the Nasal and the Oromucosal Route: Relevance to the Fight with the SARS-CoV-2 Pandemic

 , ,
, ,  ,
, 
Abstract
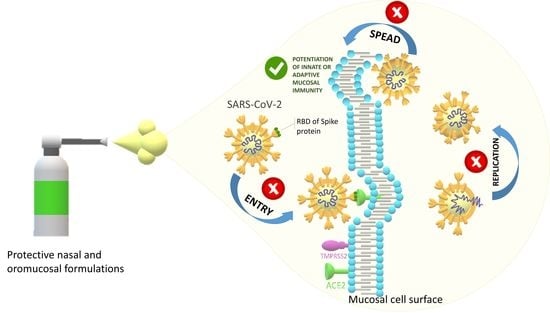
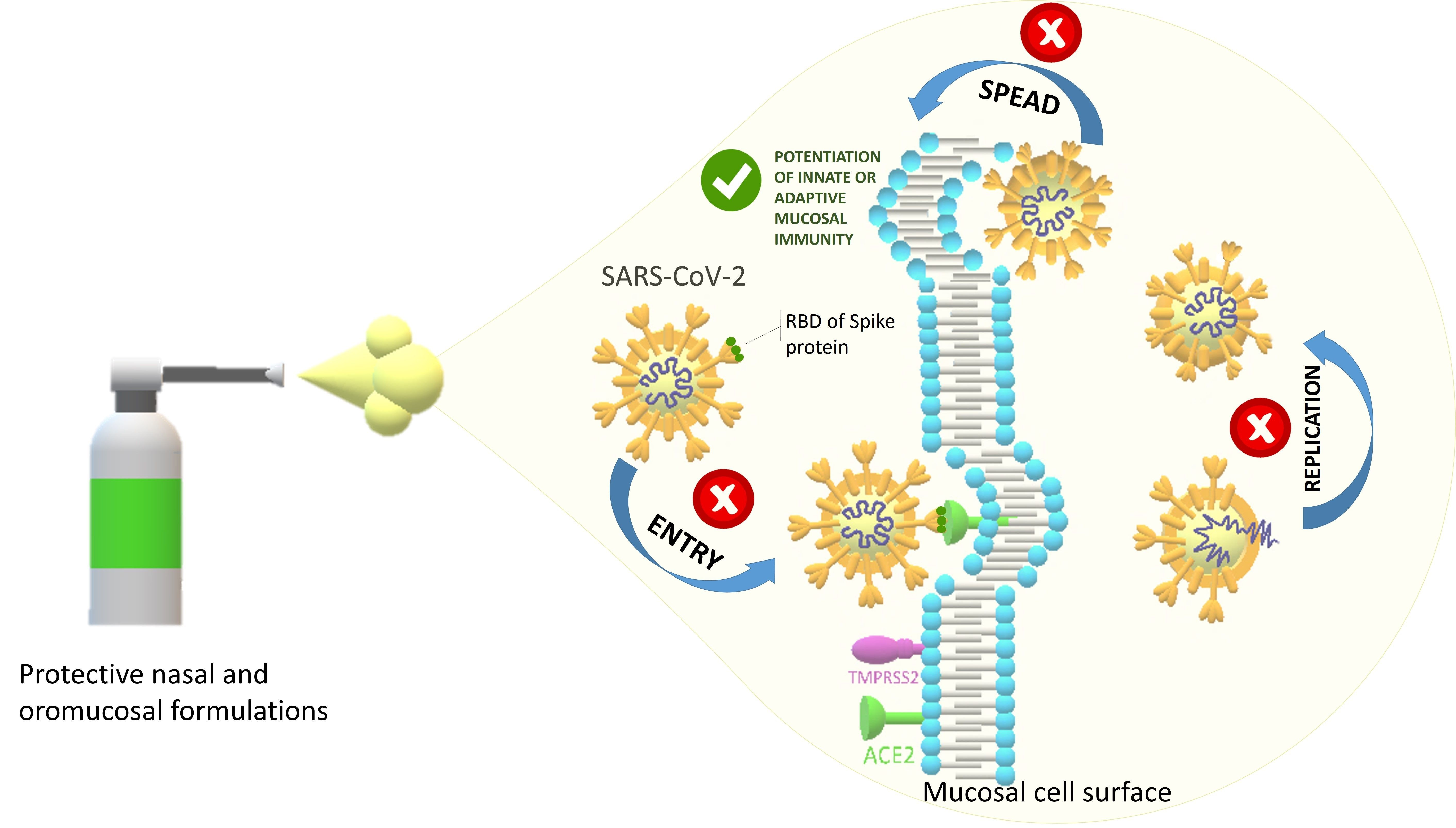
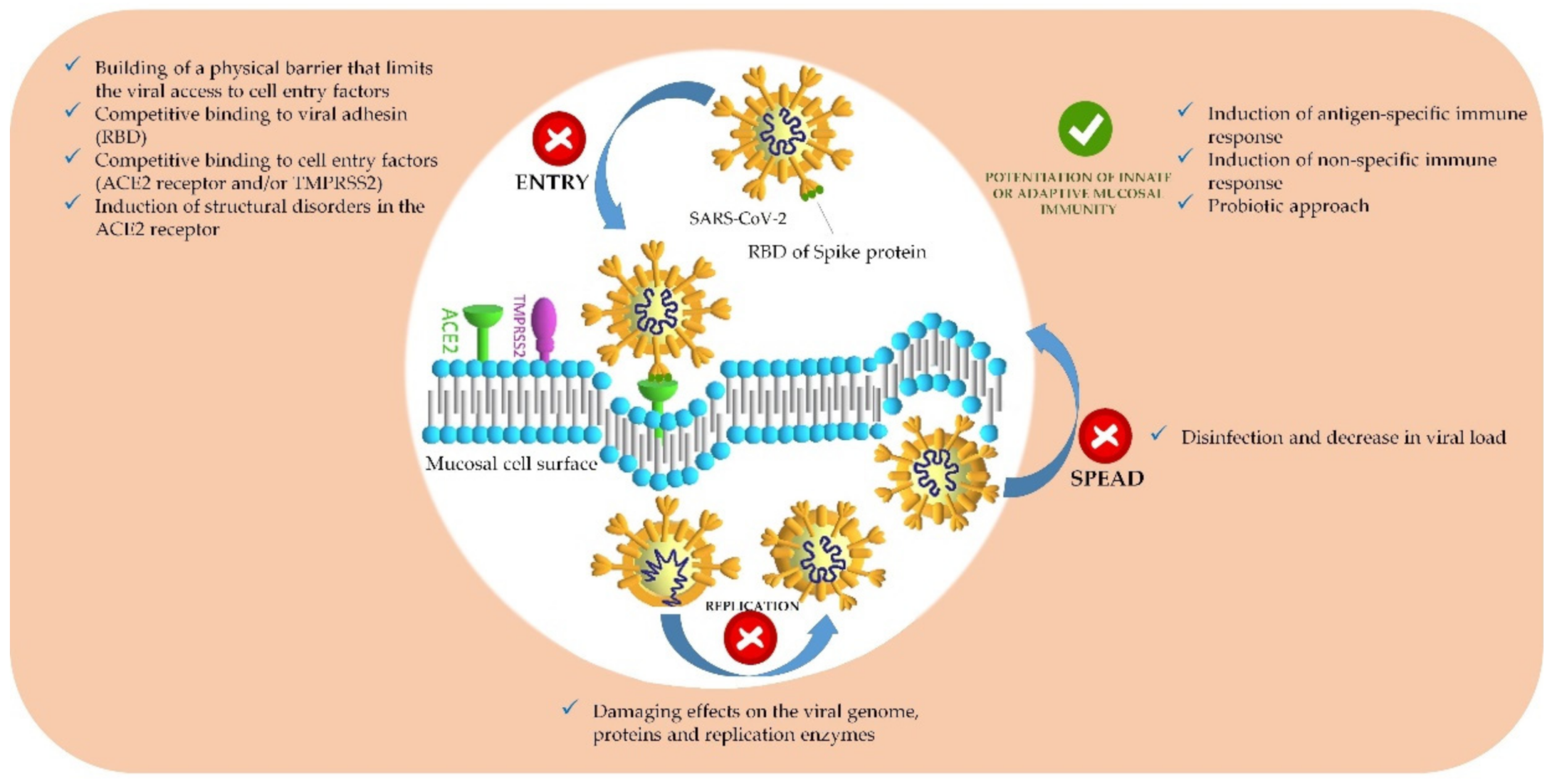
:
1. Introduction
2. An Insight of the Early Phases of Respiratory Tract Infections (RTIs)
3. The Upper Respiratory Tract (URT)
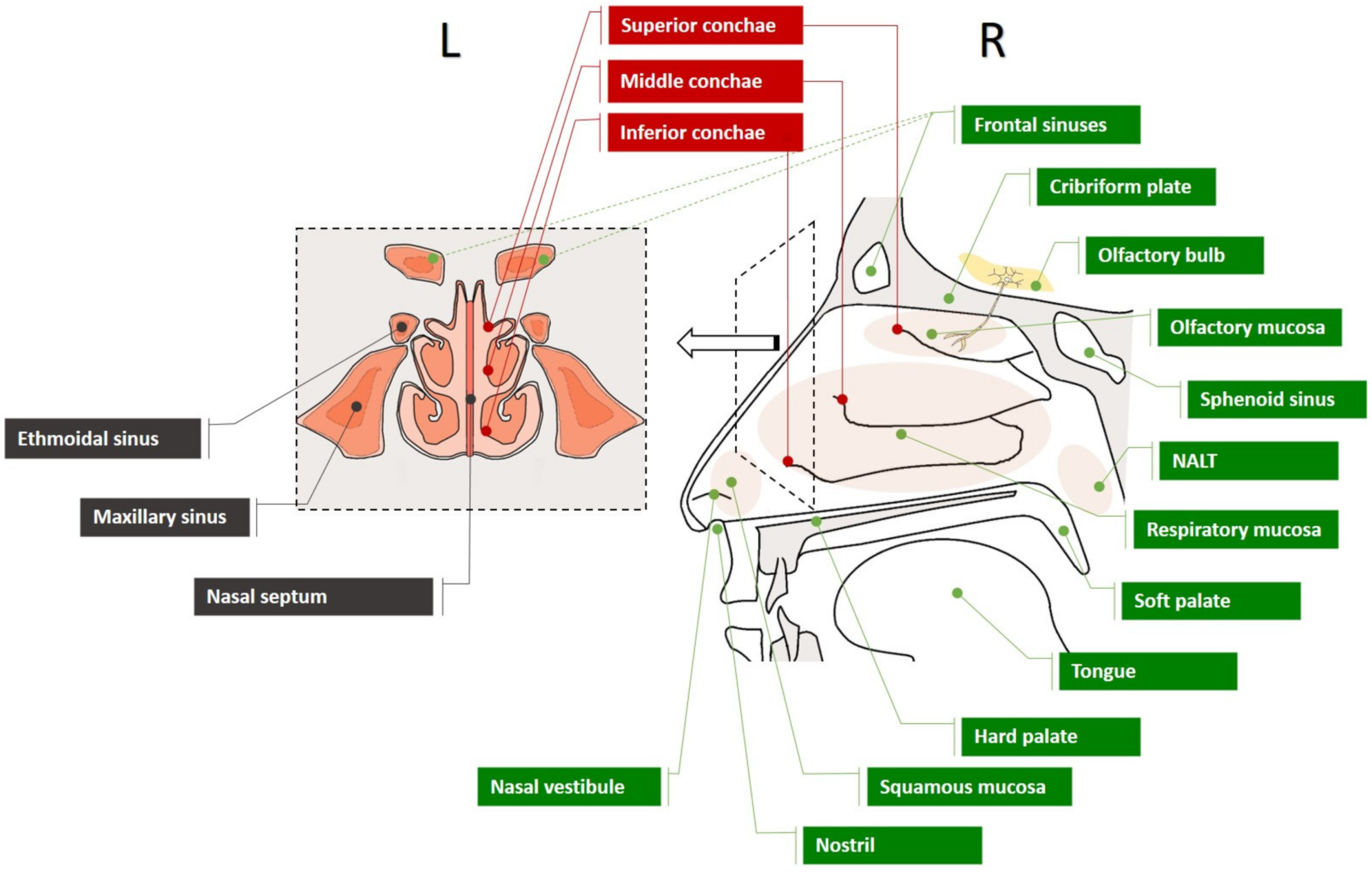
3.1. The Nasal Cavity
3.2. The Nasal Mucosa
3.3. The Mucus
3.4. The Role of the Oral Cavity as a Portal of Entry for Respiratory Pathogens
4. Prevention and Prophylaxis of RTI’s
4.1. Immunoprophylaxis
4.2. Chemoprophylaxis
5. The Nasal and Oromucosal Drug Delivery
5.1. Mucoadhesion—Principles and Utilization in Mucosal Dosage Forms
5.2. Nasal Dosage Forms
5.3. Oromucosal Dosage Forms
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stilianakis, N.I.; Drossinos, Y. Dynamics of infectious disease transmission by inhalable respiratory droplets. J. R. Soc. Interface 2010, 7, 1355–1366. [Google Scholar] [CrossRef] [Green Version]
- Dasaraju, P.V.; Liu, C. Infections of the respiratory system. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 0-9631172-1-1. [Google Scholar]
- Cegolon, L.; Javanbakht, M.; Mastrangelo, G. Nasal disinfection for the prevention and control of COVID-19: A scoping review on potential chemo-preventive agents. Int. J. Hyg. Environ. Health 2020, 230, 113605. [Google Scholar] [CrossRef] [PubMed]
- Joffe, S. Evaluating SARS-CoV-2 Vaccines After Emergency Use Authorization or Licensing of Initial Candidate Vaccines. JAMA 2021, 325, 221–222. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, I.; Liu, M.A.; Peden, K.; Zhou, T.; Kang, H.-N. Development of mRNA Vaccines: Scientific and Regulatory Issues. Vaccines 2021, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- Amanpour, S. The Rapid Development and Early Success of COVID 19 Vaccines Have Raised Hopes for Accelerating the Cancer Treatment Mechanism. Arch. Razi Inst. 2021, 76, 1–6. [Google Scholar]
- Baron, S.; Fons, M.; Albrecht, T. Viral pathogenesis. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 0-9631172-1-1. [Google Scholar]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, T.; Fratini, M.; Frey, F.; Yserentant, K.; Liu, Y.; Weber, E.; Galior, K.; Ohmes, J.; Braun, F.; Herten, D.-P.; et al. Forces during cellular uptake of viruses and nanoparticles at the ventral side. Nat. Commun. 2020, 11, 32. [Google Scholar] [CrossRef]
- Lai, S.K.; Wang, Y.-Y.; Wirtz, D.; Hanes, J. Micro- and macrorheology of mucus. Adv. Drug Deliv. Rev. 2009, 61, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Cole, P. Physiology of the nose and paranasal sinuses. In Diseases of the Sinuses; Gershwin, M.E., Incaudo, G.A., Eds.; Humana Press: Totowa, NJ, USA, 1996; pp. 33–51. ISBN 978-1-4612-0225-7. [Google Scholar]
- Tratnjek, L.; Kreft, M.; Kristan, K.; Kreft, M.E. Ciliary beat frequency of in vitro human nasal epithelium measured with the simple high-speed microscopy is applicable for safety studies of nasal drug formulations. Toxicol. Vitr. 2020, 66, 104865. [Google Scholar] [CrossRef]
- Shaikh, R.; Raj Singh, T.R.; Garland, M.J.; Woolfson, A.D.; Donnelly, R.F. Mucoadhesive drug delivery systems. J. Pharm. Bioallied Sci. 2011, 3, 89–100. [Google Scholar]
- Mallakpour, S.; Azadi, E.; Hussain, C.M. Protection, disinfection, and immunization for healthcare during the COVID-19 pandemic: Role of natural and synthetic macromolecules. Sci. Total Environ. 2021, 776, 145989. [Google Scholar] [CrossRef]
- Moakes, R.J.A.; Davies, S.P.; Stamataki, Z.; Grover, L.M. Formulation of a composite nasal spray enabling enhanced surface coverage and prophylaxis of SARS-COV-2. Adv. Mater. 2021, 33, 2008304. [Google Scholar] [CrossRef]
- Sandri, G.; Saporito, F.; Ferrari, F.; Bonferoni, M.C.; Rossi, S.; Caramella, C. In vitro evaluation of a protective nasal spray: Measurements of mucoadhesion and reconstructive barrier properties towards a tracheobronchial reconstruct. J. Drug Deliv. Sci. Technol. 2015, 30, 368–374. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Esper, F.; Buchheit, K.; Arters, K.; Adkins, I.; Ghannoum, M.A.; Salata, R.A. Randomized, double-blind, placebo-controlled clinical trial to assess the safety and effectiveness of a novel dual-action oral topical formulation against upper respiratory infections. BMC Infect. Dis. 2017, 17, 74. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, N.G.; Megremis, S.; Kitsioulis, N.A.; Vangelatou, O.; West, P.; Xepapadaki, P. Promising approaches for the treatment and prevention of viral respiratory illnesses. J. Allergy Clin. Immunol. 2017, 140, 921–932. [Google Scholar] [CrossRef] [Green Version]
- Mellis, C. Acute upper respiratory tract infections in childhood. In Practical Paediatrics, 6th ed.; Roberton, D.M., South, M., Eds.; Churchill Livingstone/Elsevier: London, UK, 2006; pp. 475–481. ISBN 9780443102806. [Google Scholar]
- Myint, S. Microbiology and epidemiology of upper respiratory tract infections. In Respiratory Infections in Allergy and Asthma, 1st ed.; Johnston, S.L., Papadopoulos, N.G., Eds.; CRC Press: Boca Raton, FL, USA, 2003; ISBN 978-0824741266. [Google Scholar]
- Cappelletty, D. Microbiology of bacterial respiratory infections. Pediatr. Infect. Dis. J. 1998, 17, S55–S61. [Google Scholar] [CrossRef]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef]
- Brachman, P.S. Epidemiology. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 0-9631172-1-1. [Google Scholar]
- Fenner, F. Epidemiology and evolution. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 0-9631172-1-1. [Google Scholar]
- Peterson, J.W. Bacterial pathogenesis. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 0-9631172-1-1. [Google Scholar]
- Kuek, L.E.; Lee, R.J. First contact: The role of respiratory cilia in host-pathogen interactions in the airways. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, L603–L619. [Google Scholar] [CrossRef]
- Johnson, J.A. Pathogenesis of bacterial infections of the respiratory tract. Br. J. Biomed. Sci. 1995, 52, 157–161. [Google Scholar]
- Wanner, A. Clinical aspects of mucociliary transport. Am. Rev. Respir. Dis. 1977, 116, 73–125. [Google Scholar]
- Cole, P.; Wilson, R. Host-microbial interrelationships in respiratory infection. Chest 1989, 95, 217S–221S. [Google Scholar] [CrossRef]
- Wilson, R.; Pitt, T.; Taylor, G.; Watson, D.; MacDermot, J.; Sykes, D.; Roberts, D.; Cole, P. Pyocyanin and 1-hydroxyphenazine produced by Pseudomonas aeruginosa inhibit the beating of human respiratory cilia in vitro. J. Clin. Investig. 1987, 79, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Denny, F.W. Effect of a toxin produced by Haemophilus influenzae on ciliated respiratory epithelium. J. Infect. Dis. 1974, 129, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Chilvers, M.A.; McKean, M.; Rutman, A.; Myint, B.S.; Silverman, M.; O’Callaghan, C. The effects of coronavirus on human nasal ciliated respiratory epithelium. Eur. Respir. J. 2001, 18, 965–970. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Tong, J.; Meng, F.; Hoeltig, D.; Liu, G.; Yin, X.; Herrler, G. Ciliostasis of airway epithelial cells facilitates influenza A virus infection. Vet. Res. 2018, 49, 65. [Google Scholar] [CrossRef] [Green Version]
- Griggs, T.F.; Bochkov, Y.A.; Basnet, S.; Pasic, T.R.; Brockman-Schneider, R.A.; Palmenberg, A.C.; Gern, J.E. Rhinovirus C targets ciliated airway epithelial cells. Respir. Res. 2017, 18, 84. [Google Scholar] [CrossRef]
- Look, D.C.; Walter, M.J.; Williamson, M.R.; Pang, L.; You, Y.; Sreshta, J.N.; Johnson, J.E.; Zander, D.S.; Brody, S.L. Effects of paramyxoviral infection on airway epithelial cell Foxj1 expression, ciliogenesis, and mucociliary function. Am. J. Pathol. 2001, 159, 2055–2069. [Google Scholar] [CrossRef] [Green Version]
- Mata, M.; Sarrion, I.; Armengot, M.; Carda, C.; Martinez, I.; Melero, J.A.; Cortijo, J. Respiratory syncytial virus inhibits ciliagenesis in differentiated normal human bronchial epithelial cells: Effectiveness of N-acetylcysteine. PLoS ONE 2012, 7, e48037. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Kulkarni, H.; Radhakrishnan, P.; Rutman, A.; Bankart, M.J.; Williams, G.; Hirst, R.A.; Easton, A.J.; Andrew, P.W.; O’Callaghan, C. Ciliary dyskinesia is an early feature of respiratory syncytial virus infection. Eur. Respir. J. 2014, 43, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.N.; Sharon, N.; Ofek, I.; Schwartzman, J.D. Adhesion and colonization. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 409–421. ISBN 9780123971692. [Google Scholar]
- Sanyal, S. How SARS-CoV-2 (COVID-19) spreads within infected hosts—What we know so far. Emerg. Top. Life Sci. 2020, 4, 371–378. [Google Scholar]
- Hallstrom, K.N.; McCormick, B.A. Pathogenicity islands: Origins, structure, and roles in bacterial pathogenesis. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 303–314. ISBN 9780123971692. [Google Scholar]
- Nash, A.A.; Dalziel, R.G.; Fitzgerald, J.R. Attachment to and entry of microorganisms into the body. In Mims’ Pathogenesis of Infectious Disease, 6th ed.; Nash, A.A., Dalziel, R.G., Fitzgerald, J.R., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 9–49. ISBN 9780123971883. [Google Scholar]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS corona-virus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.R. HLA class I antigen serves as a receptor for human coronavirus OC43. Immunol. Investig. 1993, 22, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkman, R.; Jebbink, M.F.; Koekkoek, S.M.; Deijs, M.; Jónsdóttir, H.R.; Molenkamp, R.; Ieven, M.; Goossens, H.; Thiel, V.; van der Hoek, L. Isolation and characterization of current human coronavirus strains in primary human epithelial cell cultures reveal differences in target cell tropism. J. Virol. 2013, 87, 6081–6090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essaidi-Laziosi, M.; Brito, F.; Benaoudia, S.; Royston, L.; Cagno, V.; Fernandes-Rocha, M.; Piuz, I.; Zdobnov, E.; Huang, S.; Constant, S.; et al. Propagation of respiratory viruses in human airway epithelia reveals persistent virus-specific signatures. J. Allergy Clin. Immunol. 2018, 141, 2074–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, C.I.; Barclay, W.S.; Zambon, M.C.; Pickles, R.J. Infection of Human Airway Epithelium by Human and Avian Strains of Influenza A Virus. J. Virol. 2006, 80, 8060–8068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittet, L.A.; Hall-Stoodley, L.; Rutkowski, M.R.; Harmsen, A.G. Influenza virus infection decreases tracheal mucociliary velocity and clearance of Streptococcus pneumoniae. Am. J. Respir. Cell Mol. Biol. 2010, 42, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Bella, J.; Kolatkar, P.R.; Marlor, C.W.; Greve, J.M.; Rossmann, M.G. The Structure of the Two Amino-Terminal Domains of Human ICAM-1 Suggests How it Functions as a Rhinovirus Receptor and as an LFA-1 Integrin Ligand. Proc. Natl. Acad. Sci. USA 1998, 95, 4140–4145. [Google Scholar] [CrossRef] [Green Version]
- Sakakura, Y.; Sasaki, Y.; Togo, Y.; Wagner, H.N., Jr.; Hornick, R.B.; Schwartz, A.R.; Proctor, D.F. Mucociliary function during experimentally induced rhinovirus infection in man. Ann. Otol. Rhinol. Laryngol. 1973, 82, 203–211. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other line-age B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Nakayama, T.; Jiang, S.; Goltsev, Y.; Schürch, C.; Zhu, B.; McIlwain, D.; Chu, P.; Chen, H.; Tzankov, A.; et al. SARS-CoV-2 entry factors are expressed in nasal, ocular, and oral tissues: Implications for COVID-19 prophylaxes/therapeutics. J. Allergy Clin. Immunol. 2021, 147, AB2. [Google Scholar] [CrossRef]
- Mete, A.; Akbudak, İ.H. Functional anatomy and physiology of airway. In Tracheal Intubation; Erbay, R.H., Ed.; IntechOpen: London, UK, 2018; ISBN 978-1-78923-489-3. [Google Scholar]
- Triana, B.E.G.; Ali, A.H.; León, I.B.G. Mouth breathing and its relationship to some oral and medical conditions: Physio-pathological mechanisms involved. Rev. Habanera Cienc. Med. 2016, 15, 200–212. [Google Scholar]
- Gizurarson, S. Anatomical and histological factors affecting intranasal drug and vaccine delivery. Curr. Drug Deliv. 2012, 9, 566–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkema, J.R.; Carey, S.A.; Wagner, J.G. The nose revisited: A brief review of the comparative structure, function, and toxicologic pathology of the nasal epithelium. Toxicol. Pathol. 2006, 34, 252–269. [Google Scholar] [CrossRef]
- Ozturk, A.B.; Damadoglu, E.; Karakaya, G.; Kalyoncu, A.F. Does nasal hair (vibrissae) density affect the risk of developing asthma in patients with seasonal rhinitis? Int. Arch. Allergy Immunol. 2011, 156, 75–80. [Google Scholar] [CrossRef]
- Beule, A.G. Physiology and pathophysiology of respiratory mucosa of the nose and the paranasal sinuses. GMS Curr. Top. Otorhinolaryngol. Head Neck Surg. 2010, 9, 1–24. [Google Scholar]
- Anand, S.; Mayya, Y.S. Size distribution of virus laden droplets from expiratory ejecta of infected subjects. Sci. Rep. 2020, 10, 21174. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, G.; Huang, Y.-W. Modeling the load of SARS-CoV-2 virus in human expelled particles during coughing and speaking. PLoS ONE 2020, 15, e0241539. [Google Scholar]
- Bustamante-Marin, X.M.; Ostrowski, L.E. Cilia and Mucociliary Clearance. Cold Spring Harb. Perspect. Biol. 2017, 9, a028241. [Google Scholar] [CrossRef]
- Mitchison, T.J.; Mitchison, H.M. How cilia beat. Nature 2010, 463, 308–309. [Google Scholar] [CrossRef]
- Debertin, A.S.; Tschernig, T.; Tönjes, H.; Kleemann, W.J.; Tröger, H.D.; Pabst, R. Nasal-associated lymphoid tissue (NALT): Frequency and localization in young children. Clin. Exp. Immunol. 2003, 134, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y. Evidence of M cells as portals of entry for antigens in the nasopharyngeal lymphoid tissue of humans. Virchows Arch. 2000, 436, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Lobaina Mato, Y. Nasal route for vaccine and drug delivery: Features and current opportunities. Int. J. Pharm. 2019, 572, 118813. [Google Scholar] [CrossRef]
- Moini, J.; Piran, P. Cranial nerves. In Functional and Clinical Neuroanatomy, 1st ed.; Moini, J., Piran, P., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 319–344. ISBN 9780128174258. [Google Scholar]
- Alberty, J.; Stoll, W.; Rudack, C. The effect of endogenous nitric oxide on mechanical ciliostimulation of human nasal mucosa. Clin. Exp. Allergy 2006, 36, 1254–1259. [Google Scholar] [CrossRef]
- Swart, S.J.; van der Baan, S.; Steenbergen, J.J.E.; Nauta, J.J.P.; van Kamp, G.J.; Biewenga, J. Immunoglobulin concentrations in nasal secretions differ between patients with an IgE-mediated rhinopathy and a non-IgE-mediated rhinopathy. J. Allergy Clin. Immunol. 1991, 88, 612–619. [Google Scholar] [CrossRef]
- Meredith, S.D.; Raphael, G.D.; Baraniuk, J.N.; Banks, S.M.; Kaliner, M.A. The pathophysiology of rhinitis. III. The control of IgG secretion. J. Allergy Clin. Immunol. 1989, 84, 920–930. [Google Scholar] [CrossRef]
- Huang, N.; Pérez, P.; Kato, T.; Mikami, Y.; Okuda, K.; Gilmore, R.C.; Conde, C.D.; Gasmi, B.; Stein, S.; Beach, M.; et al. SARS-CoV-2 infection of the oral cavity and saliva. Nat. Med. 2021, 27, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Altaie, A.M.; Hamdy, R.; Venkatachalam, T.; Hamoudi, R.; Soliman, S.S.M. Estimating the viral loads of SARS-CoV-2 in the oral cavity when complicated with periapical lesions. BMC Oral Health 2021, 21, 567. [Google Scholar] [CrossRef]
- Xiang, Z.; Koo, H.; Chen, Q.; Zhou, X.; Liu, Y.; Simon-Soro, A. Potential implications of SARS-CoV-2 oral infection in the host microbiota. J. Oral Microbiol. 2020, 13, 1853451. [Google Scholar] [CrossRef]
- Walker, D.H. Interactions of pathogens with the host. In Pathobiology of Human Disease; MacManus, L.M., Mitchel, R.N., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 214–216. ISBN 978-0-12-386457-4. [Google Scholar]
- Gaeckle, N.T.; Pragman, A.A.; Pendleton, K.M.; Baldomero, A.K.; Criner, G.J. The Oral-Lung Axis: The Impact of Oral Health on Lung Health. Respir. Care 2020, 65, 1211–1220. [Google Scholar] [CrossRef]
- De Steenhuijsen Piters, W.A.A.; Sanders, E.A.M.; Bogaert, D. The role of the local microbial ecosystem in respiratory health and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, R.A.; Hoit, J.D. Nasal and oral inspiration during natural speech breathing. J. Speech Lang. Hear. Res. 2014, 57, 734–742. [Google Scholar] [CrossRef] [Green Version]
- Abreu, R.R.; Rocha, R.L.; Lamounier, J.A.; Guerra, A.F.M. Etiology, clinical manifestations and concurrent findings in mouth-breathing children. J. Pediatr. 2008, 84, 529–535. [Google Scholar] [CrossRef]
- Bradley, D. Physiotherapy in rehabilitation of breathing pattern disorders. In Recognizing and Treating Breathing Disorders, 2nd ed.; Chaitow, L., Bradley, D., Gilbert, C., Eds.; Churchill Livingstone/Elsevier: London, UK, 2014; pp. 185–196. ISBN 978-0-7020-4980-4. [Google Scholar]
- Lundberg, J.O.; Weitzberg, E. Nasal nitric oxide in man. Thorax 1999, 54, 947–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, S.; Vinayachandran, D. Bioaerosols from mouth-breathing: Under-recognized transmissible mode in COVID-19? Can. Commun. Dis. Rep. 2021, 47, 276–278. [Google Scholar] [CrossRef]
- Martel, J.; Ko, Y.-F.; Young, J.D.; Ojcius, D.M. Could nasal nitric oxide help to mitigate the severity of COVID-19? Microbes Infect. 2020, 22, 168–171. [Google Scholar] [CrossRef]
- Lotz, C.; Muellenbach, R.M.; Meybohm, P.; Mutlak, H.; Lepper, P.M.; Rolfes, C.-B.; Peivandi, A.; Stumpner, J.; Kredel, M.; Kranke, P.; et al. Effects of inhaled nitric oxide in COVID-19-induced ARDS—Is it worthwhile? Acta Anaesthesiol. Scand. 2021, 65, 629–632. [Google Scholar] [CrossRef]
- Wu, R.-Q.; Zhang, D.-F.; Tu, E.; Chen, Q.-M.; Chen, W. The mucosal immune system in the oral cavity-an orchestra of T cell diversity. Int. J. Oral Sci. 2014, 6, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar]
- La Rosa, G.R.M.; Libra, M.; de Pasquale, R.; Ferlito, S.; Pedullà, E. Association of Viral Infections with Oral Cavity Lesions: Role of SARS-CoV-2 Infection. Front. Med. 2021, 7, 571214. [Google Scholar] [CrossRef]
- Xu, H.; Zhong, L.; Deng, J.; Peng, J.; Dan, H.; Zeng, X.; Li, T.; Chen, Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int. J. Oral Sci. 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.M. The Importance of Reducing the Viral Load in the Oral Cavity. Int. J. Oral Dent. Health 2021, 7, 124. [Google Scholar]
- Baghizadeh Fini, M. Oral saliva and COVID-19. Oral Oncol. 2020, 108, 104821. [Google Scholar] [CrossRef]
- Zhang, H.; Kang, Z.; Gong, H.; Xu, D.; Wang, J.; Li, Z.; Li, Z.; Cui, X.; Xiao, J.; Zhan, J.; et al. Digestive system is a potential route of COVID-19: An analysis of single-cell coexpression pattern of key proteins in viral entry process. Gut 2020, 69, 1010–1018. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, Q.; Liu, H. Coronavirus disease 2019 and the gut-lung axis. Int. J. Infect. Dis. 2021, 113, 300–307. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, G.L.V.; Oliveira, C.N.S.; Pinzan, C.F.; de Salis, L.V.V.; de Barros Cardoso, C.R. Microbiota Modulation of the Gut-Lung Axis in COVID-19. Front. Immunol. 2021, 12, 635471. [Google Scholar] [CrossRef]
- Guo, M.; Tao, W.; Flavell, R.A.; Zhu, S. Potential intestinal infection and faecal-oral transmission of SARS-CoV-2. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 269–283. [Google Scholar] [CrossRef]
- Dibner, J.J. Direct COVID-19 infection of enterocytes: The role of hypochlorhydria. Am. J. Infect. Control 2021, 49, 385–386. [Google Scholar] [CrossRef]
- Price, E. Could the severity of COVID-19 be increased by low gastric acidity? Crit. Care 2020, 24, 456. [Google Scholar] [CrossRef]
- Zhou, J.; Li, C.; Zhao, G.; Chu, H.; Wang, D.; Yan, H.H.-N.; Poon, V.K.-M.; Wen, L.; Wong, B.H.-Y.; Zhao, X.; et al. Human intestinal tract serves as an alternative infection route for Middle East respiratory syndrome coronavirus. Sci. Adv. 2017, 3, eaao4966. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Zhang, C.; Dong, J.; Zhao, L.; Li, Y.; Sun, J. Oral Microbiome and SARS-CoV-2: Beware of Lung Co-infection. Front. Microbiol. 2020, 11, 1840. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Transmission of SARS-CoV-2: Implications for Infection Prevention Precautions. Scientific Brief. Available online: https://www.who.int/news-room/commentaries/detail/transmission-of-sars-cov-2-implications-for-infection-prevention-precautions (accessed on 15 August 2020).
- WHO. Controlling the Spread of Infectious Diseases. Advancing the Right to Health: The Vital Role of Law; World Health Organization: Geneva, Switzerland, 2017; pp. 151–164. ISBN 9789241511384.
- Keeling, M.J.; Rohani, P. Controlling infectious diseases. In Modeling Infectious Diseases in Humans and Animals; Keeling, M.J., Rohani, P., Eds.; Princeton University Press: Princeton, NJ, USA, 2008; pp. 291–336. ISBN 9781400841035. [Google Scholar]
- Jain, M.; Kim, S.T.; Xu, C.; Li, H.; Rose, G. Efficacy and Use of Cloth Masks: A Scoping Review. Cureus 2020, 12, e10423. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Seale, H.; MacIntyre, C.R.; Zhang, H.; Zhang, Z.; Zhang, Y.; Wang, X.; Li, X.; Pang, X.; Wang, Q. Mask-wearing and respiratory infection in healthcare workers in Beijing, China. Braz. J. Infect. Dis. 2011, 15, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.; Huang, A.; Li, Z.; Tufekci, Z.; Zdimal, V.; van der Westhuizen, H.-M.; von Delft, A.; Price, A.; Fridman, L.; Tang, L.-H.; et al. An evidence review of face masks against COVID-19. Proc. Natl. Acad. Sci. USA 2021, 118, e2014564118. [Google Scholar] [CrossRef]
- Davies, A.; Thompson, K.-A.; Giri, K.; Kafatos, G.; Walker, J.; Bennet, A. Testing the efficacy of homemade masks: Would they protect in an influenza pandemic? Disaster Med. Public Health Prep. 2013, 7, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, C.J.; Hufert, F.; Siewert, S.; Reisinger, E. Protection from COVID-19: The efficacy of face masks. Dtsch. Arztebl. Int. 2021, 118, 59–65. [Google Scholar] [CrossRef]
- Neupane, B.B.; Mainali, S.; Sharma, A.; Giri, B. Optical microscopic study of surface morphology and filtering efficiency of face masks. PeerJ 2019, 26, e7142. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M. A study on the microbial filtration efficiency of surgical face masks—With special reference to the non-woven fabric mask. Bull. Tokyo Med. Dent. Univ. 1978, 25, 7–15. [Google Scholar]
- Tagle, M.G.; Cilia-López, V.G. The Massive Misuse of Face Mask as a Risk to COVID-19 Pandemic in Latin American: The Case of Mexico. PREPRINT (Version 1). 2021. Available online: https://www.researchsquare.com/article/rs-323037/v1 (accessed on 13 October 2021).
- Neilson, S. The surgical mask is a bad fit for risk reduction. Can. Med. Assoc. J. 2016, 188, 606–607. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Ma, N.; Witt, C.; Rapp, S.; Wild, P.S.; Andreae, M.O.; Pöschl, U.; Su, H. Face masks effectively limit the probability of SARS-CoV-2 transmission. Science 2021, 372, 1439–1443. [Google Scholar] [CrossRef]
- Chou, R.; Dana, T.; Jungbauer, R.; Weeks, C.; McDonagh, M.S. Masks for Prevention of Respiratory Virus Infections, Including SARS-CoV-2, in Health Care and Community Settings. Ann. Intern. Med. 2020, 173, 542–555. [Google Scholar] [CrossRef] [PubMed]
- Chou, R.; Dana, T.; Jungbauer, R.; Weeks, C. Update Alert 3: Masks for Prevention of Respiratory Virus Infections, Including SARS-CoV-2, in Health Care and Community Settings. Ann. Intern. Med. 2020, 173, 169. [Google Scholar] [CrossRef] [PubMed]
- Chou, R.; Dana, T.; Jungbauer, R.; Weeks, C. Update Alert 4: Masks for Prevention of Respiratory Virus Infections, Including SARS-CoV-2, in Health Care and Community Settings. Ann. Intern. Med. 2021, 174, W24. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.M.; Willmore, L.J. Posttraumatic epilepsy. In Handbook of Clinical Neurology; Stefan, H., Theodore, W.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 108, pp. 585–599. ISBN 978-0-444-52899-5. [Google Scholar]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Abbas, A.T.; El-Kafrawy, S.A.; Sohrab, S.S.; Azhar, E.I.A. IgY antibodies for the immunoprophylaxis and therapy of respiratory infections. Hum. Vaccin. Immunoth. 2019, 15, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Peghin, M.; Danziger-Isakov, L. Prevention and treatment of respiratory virus infection. In Infectious Diseases in Solid-Organ Transplant Recipients, 1st ed.; Manuel, O., Ison, M.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 107–129. ISBN 978-3-030-15394-6. [Google Scholar]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2021, 1–15. [Google Scholar] [CrossRef]
- Sparrow, E.; Friede, M.; Sheikh, M.; Torvaldsen, S. Therapeutic antibodies for infectious diseases. Bull. World Health Organ. 2017, 95, 235–237. [Google Scholar] [CrossRef]
- Berry, C.M. Antibody immunoprophylaxis and immunotherapy for influenza virus infection: Utilization of monoclonal or polyclonal antibodies? Hum. Vaccin. Immunoth. 2018, 14, 796–799. [Google Scholar] [CrossRef]
- Rahman, S.; van Nguyen, S.; Icatlo, F.C., Jr.; Umeda, K.; Kodama, Y. Oral passive IgY-based immunotherapeutics: A novel solution for prevention and treatment of alimentary tract diseases. Hum. Vaccin. Immunoth. 2013, 9, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Calvert, R.A.; Sutton, B.J.; Doré, K.A. IgY: A key isotype in antibody evolution. Biol. Rev. Camb. Philos. Soc. 2017, 92, 2144–2156. [Google Scholar] [CrossRef]
- Ikemori, Y.; Peralta, R.C.; Kuroki, M.; Yokoyama, H.; Kodama, Y. Research note: Avidity of chicken yolk antibodies to enterotoxigenic Escherichia coli fimbriae. Poult. Sci. 1993, 72, 2361–2365. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Thömmes, P.; Weiser, T.; Hübscher, U. Efficient production of chicken egg yolk antibodies against a conserved mammalian protein. FASEB J. 1990, 4, 2528–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, K.; Christophersen, L.; Bjarnsholt, T.; Jensen, P.Ø.; Moser, C.; Høiby, N. Anti-Pseudomonas aeruginosa IgY antibodies augment bacterial clearance in a murine pneumonia model. J. Cyst. Fibros. 2016, 15, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Sudjarwo, S.A.; Eraiko, K.; Sudjarwo, G.W. Koerniasari. The potency of chicken egg yolk immunoglobulin (IgY) specific as immunotherapy to Mycobacterium tuberculosis infection. J. Adv. Pharm. Technol. Res. 2017, 8, 91–96. [Google Scholar] [PubMed]
- Ferella, A.; Bellido, D.; Chacana, P.; Wigdorovitz, A.; dus Santos, M.J.; Mozgovoj, M.V. Chicken egg yolk antibodies against bovine respiratory syncytial virus neutralize the virus in vitro. Procedia Vaccinol. 2012, 6, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.Y.; Huang, H.; Wang, X.-M.; Liu, Y.-G.; Wang, Z.-G.; Cui, S.-J.; Gao, H.-L.; Li, Z.; Li, J.-P.; Kong, X.-G. Preparation and evaluation of anti-SARS coronavirus IgY from yolks of immunized SPF chickens. J. Virol. Methods 2006, 133, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Constantin, C.; Neagu, M.; Diana Supeanu, T.; Chiurciu, V.; Spandidos, D.A. IgY—Turning the page toward passive immunization in COVID-19 infection (Review). Exp. Ther. Med. 2020, 20, 151–158. [Google Scholar] [CrossRef]
- Davey, P.; Wilcox, M.H.; Irving, W.; Thwaites, G. Chemoprophylaxis and immunization. In Antimicrobial Chemotherapy, 7th ed.; Davey, P., Wilcox, M.H., Irving, W., Thwaites, G., Eds.; Oxford University Press: Oxford, UK, 2015; ISBN 9780199689774. [Google Scholar]
- Bradley, J.S. Chemoprophylaxis. In Principles and Practice of Pediatric Infectious Diseases, 4th ed.; Long, S.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 68–76. ISBN 9781437727029. [Google Scholar]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 1–12. [Google Scholar] [CrossRef]
- Batten, J. Chemoprophylaxis of respiratory infections. Postgrad. Med. J. 1976, 52, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Oxford, J.S. Chemoprophylaxis and Virus Infections of the Respiratory Tract, 1st ed.; Oxford, J.S., Ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781351070614. [Google Scholar]
- Minozzi, S.; Pifferi, S.; Brazzi, L.; Pecoraro, V.; Montrucchio, G.; D’Amico, R. Topical antibiotic prophylaxis to reduce respiratory tract infections and mortality in adults receiving mechanical ventilation. Cochrane Database Syst. Rev. 2021, 1, CD000022. [Google Scholar]
- Grammatikos, A.; Albur, M.; Gompels, M.; Barnaby, C.L.; Allan, S.; Johnston, S. Antibiotic prophylaxis for the prevention of respiratory tract infections in antibody deficient patients: A retrospective cohort study. Clin. Infect. Pract. 2020, 7–8, 100048. [Google Scholar] [CrossRef]
- Liberati, A.; D’Amico, R.; Pifferi, S.; Torri, V.; Brazzi, L.; Parmelli, E. Antibiotic prophylaxis to reduce respiratory tract infections and mortality in adults receiving intensive care. Cochrane Database Syst. Rev. 2009, 2009, CD000022. [Google Scholar]
- Guitor, A.K.; Wright, G.D. Antimicrobial Resistance and Respiratory Infections. Chest 2018, 154, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Alshehhi, H.S.; Ali, A.A.; Jawhar, D.S.; Aly, E.M.; Swamy, S.; Fattah, M.A.; Drweesh, K.A.; Alsaadi, A. Assessment of implementation of antibiotic stewardship program in surgical prophylaxis at a secondary care hospital in Ras Al Khaimah, United Arab Emirates. Sci. Rep. 2021, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Tappenden, P.; Jackson, R.; Cooper, K.; Rees, A.; Simpson, E.; Read, R.; Nicholson, K. Amantadine, oseltamivir and zanamivir for the prophylaxis of influenza (including a review of existing guidance no. 67): A systematic review and economic evaluation. Health Technol. Assess. 2009, 13, 1–246. [Google Scholar] [CrossRef] [Green Version]
- Prevention of SARS-CoV-2 Infection. Available online: https://www.covid19treatmentguidelines.nih.gov/overview/prevention-of-sars-cov-2/ (accessed on 18 December 2021).
- Shoumann, W.M.; Hegazy, A.A.; Nafae, R.M.; Ragab, M.I.; Samra, S.R.; Ibrahim, D.A.; Al-Mahrouky, T.H.; Sileem, A.E. Use of Ivermectin as a Potential Chemoprophylaxis for COVID-19 in Egypt: A Randomised Clinical Trial. J. Clin. Diagn. Res. 2021, 15, OC27–OC32. [Google Scholar] [CrossRef]
- Omrani, M.; Keshavarz, M.; Nejad Ebrahimi, S.N.; Mehrabi, M.; McGaw, L.J.; Ali Abdalla, M.; Mehrbod, P. Potential Natural Products Against Respiratory Viruses: A Perspective to Develop Anti-COVID-19 Medicines. Front. Pharmacol. 2021, 11, 586993. [Google Scholar] [CrossRef]
- Langeder, J.; Grienke, U.; Chen, Y.; Kirchmair, J.; Schmidtke, M.; Rollinger, J.M. Natural products against acute respiratory infections: Strategies and lessons learned. J. Ethnopharmacol. 2020, 248, 112298. [Google Scholar] [CrossRef]
- Hensel, A.; Bauer, R.; Heinrich, M.; Spiegler, V.; Kayser, O.; Hempel, G.; Kraft, K. Challenges at the Time of COVID-19: Opportunities and Innovations in Antivirals from Nature. Planta Med. 2020, 86, 659–664. [Google Scholar] [CrossRef]
- Chen, X.; Han, W.; Wang, G.; Zhao, X. Application prospect of polysaccharides in the development of anti-novel corona-virus drugs and vaccines. Int. J. Biol. Macromol. 2020, 164, 331–343. [Google Scholar] [CrossRef]
- Alitongbieke, G.; Li, X.-M.; Wu, Q.-C.; Lin, Z.-C.; Huang, J.-F.; Xue, Y.; Liu, J.-N.; Lin, J.-M.; Pan, T.; Chen, Y.-X.; et al. Effect of β-chitosan on the Binding Interaction between SARS-CoV-2 S-RBD and ACE2. PREPRINT. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.07.31.229781v3 (accessed on 24 October 2021).
- Milewska, A.; Chi, Y.; Szczepanski, A.; Barreto-Duran, E.; Liu, K.; Liu, D.; Guo, X.; Ge, Y.; Li, J.; Cui, L.; et al. HTCC as a Highly Effective Polymeric Inhibitor of SARS-CoV-2 and MERS-CoV. Preprint. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.03.29.014183v1 (accessed on 24 October 2021).
- Jang, Y.; Shin, H.; Lee, M.K.; Kwon, O.S.; Shin, J.S.; Kim, Y.-I.; Kim, C.W.; Lee, H.-R.; Kim, M. Antiviral activity of lamb-da-carrageenan against influenza viruses and severe acute respiratory syndrome coronavirus 2. Sci. Rep. 2021, 11, 821. [Google Scholar] [CrossRef] [PubMed]
- Morokutti-Kurz, M.; Graf, C.; Prieschl-Grassauer, E. Amylmetacresol/2,4-dichlorobenzyl alcohol, hexylresorcinol, or carrageenan lozenges as active treatments for sore throat. Int. J. Gen. Med. 2017, 10, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.-L.; Li, Y.; Ni, L.-Q.; Li, Y.-X.; Cui, Y.-S.; Jiang, S.-L.; Xie, E.-Y.; Du, J.; Deng, F.; Dong, C.-X. Structural characterization and antiviral activity of two fucoidans from the brown algae Sargassum henslowianum. Carbohydr. Polym. 2020, 229, 115487. [Google Scholar] [CrossRef]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of Influenza A Virus Infection by Fucoidan Targeting Viral Neuraminidase and Cellular EGFR Pathway. Sci. Rep. 2017, 7, 40760. [Google Scholar] [CrossRef]
- Chang, R.; Ng, T.B.; Sun, W.-Z. Lactoferrin as potential preventative and adjunct treatment for COVID-19. Int. J. Antimicrob. Agents 2020, 56, 106118. [Google Scholar] [CrossRef]
- Salaris, C.; Scarpa, M.; Elli, M.; Bertolini, A.; Guglielmetti, S.; Pregliasco, F.; Blandizzi, C.; Brun, P.; Castagliuolo, I. Protective Effects of Lactoferrin against SARS-CoV-2 Infection In Vitro. Nutrients 2021, 13, 328. [Google Scholar] [CrossRef]
- Chauhan, P.; Kumar, A. Development of a microbial coating for cellulosic surface using aloe vera and silane. Carbohydr. Polym. Technol. Appl. 2020, 1, 100015. [Google Scholar] [CrossRef]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Review of Evidence Available on Hesperidin-Rich Products as Potential Tools against COVID-19 and Hydrodynamic Cavitation-Based Extraction as a Method of Increasing Their Production. Processes 2020, 8, 549. [Google Scholar] [CrossRef]
- Cheng, L.; Zheng, W.; Li, M.; Huang, J.; Bao, S.; Xu, Q.; Ma, Z. Citrus Fruits Are Rich in Flavonoids for Immunoregulation and Potential Targeting ACE2. PREPRINT (Version 1). 2020. Available online: https://www.preprints.org/manuscript/202002.0313/v1 (accessed on 26 October 2021).
- Omar, S.; Bouziane, I.; Bouslama, Z.; Djemel, A. In-Silico Identification of Potent Inhibitors of COVID-19 Main Protease (Mpro) and Angiotensin Converting Enzyme 2 (ACE2) from Natural Products: Quercetin, Hispidulin, and Cirsimaritin Ex-hibited Better Potential Inhibition than Hydroxy-Chloroquine against COVID-19 Main Protease Active Site and ACE2. 2020. Available online: https://chemrxiv.org/engage/chemrxiv/article-details/60c74a53469df45440f43d21 (accessed on 26 October 2021).
- Joshi, R.S.; Jagdale, S.S.; Bansode, S.B.; Shankar, S.S.; Tellis, M.B.; Pandya, V.K.; Chugh, A.; Giri, A.P.; Kulkarni, M.J. Discovery of potential multi-target-directed ligands by targeting host-specific SARS-CoV-2 structurally conserved main protease. J. Biomol. Struct. Dyn. 2021, 39, 3099–3114. [Google Scholar] [CrossRef] [Green Version]
- Wahedi, H.M.; Ahmad, S.; Abbasi, S.W. Stilbene-based natura lompounds as promising drug candidates against COVID-19. J. Biomol. Struct. Dyn. 2020, 39, 3225–3234. [Google Scholar] [PubMed]
- Chen, H.; Du, Q. Potential Natural Compounds for Preventing SARS-CoV-2 (2019-nCoV) Infection. 2020. Available online: https://www.preprints.org/manuscript/202001.0358/v3 (accessed on 26 October 2021).
- Maurya, V.K.; Kumar, S.; Prasad, A.K.; Bhatt, M.L.B.; Saxena, S.K. Structure-based drug designing for potential antiviral activity of selected natural products from Ayurveda against SARS-CoV-2 spike glycoprotein and its cellular receptor. Virus Dis. 2020, 31, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Sa-Ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensut-thivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis paniculata Extract and Its Major Component Andrographolide in Human Lung Epithelial Cells and Cyto-toxicity Evaluation in Major Organ Cell Representatives. J. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Zhu, Z.; Yu, P.; Zhang, X.; Dong, W.; Wang, X.; Chen, Y.; Liu, X. Inhibitory effect of iota-carrageenan on porcine reproductive and respiratory syndrome virus in vitro. Antivir. Ther. 2019, 24, 261–270. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, P.; Yu, G.-L.; Li, C.-X.; Hao, C.; Qi, X.; Zhang, L.-J.; Guan, H.-S. Preparation and anti-influenza A virus activity of κ-carrageenan oligosaccharide and its sulphated derivatives. Food Chem. 2012, 133, 880–888. [Google Scholar] [CrossRef]
- Ter Ellen, B.M.; Kumar, N.D.; Bouma, E.M.; Troost, B.; van de Pol, D.P.I.; van der Ende-Metselaar, H.; Nawijn, M. Resveratrol and Pterostilbene Potentlyinhibit SARS-CoV-2 Infection in Vitro. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.09.24.285940v1 (accessed on 27 October 2021).
- Pasquereau, S.; Nehme, Z.; Haidar Ahmad, S.; Daouad, F.; van Assche, J.; Wallet, C.; Schwartz, C.; Rohr, O.; Morot-Bizot, S.; Herbein, G. Resveratrol Inhibits HCoV-229E and SARS-CoV-2 Coronavirus Replication In Vitro. Viruses 2021, 13, 354. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H.W. Glycyrrhizin, an active component of liquo-rice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Mishra, K.P.; Ganju, L. Broad-spectrum antiviral properties of andrographolide. Arch. Virol. 2017, 162, 611–623. [Google Scholar] [CrossRef]
- Muralidharan, A.; Russell, M.S.; Larocque, L.; Gravel, C.; Sauvé, S.; Chen, Z.; Li, C.; Chen, W.; Cyr, T.; Rosu-Myles, M.; et al. Chitosan alters inactivated respiratory syncytial virus vaccine elicited immune responses without affecting lung histopathology in mice. Vaccine 2019, 37, 4031–4039. [Google Scholar] [CrossRef]
- Aucoin, M.; Cooley, K.; Saunders, P.R.; Carè, J.; Anheyer, D.; Medina, D.N.; Cardozo, V.; Remy, D.; Hannan, N.; Garber, A. The effect of Echinacea spp. on the prevention or treatment of COVID-19 and other respiratory tract infections in hu-mans: A rapid review. Adv. Integr. Med. 2020, 7, 203–217. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, J.; Wang, W.; Liu, X.; Liu, H.; Li, X.; Wu, X. Astragalus polysaccharides enhance the immune response to avian infectious bronchitis virus vaccination in chickens. Microb. Pathog. 2017, 111, 81–85. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, X.; Liu, H.; Wang, W.; Liu, X.; Li, X.; Wu, X. Astragalus polysaccharides inhibit avian infectious bronchitis virus infection by regulating viral replication. Microb. Pathog. 2018, 114, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Garg, P.; Singh, P.; Verma, K. Remdesivir break replication of virus & Rhodiola rosea acts same by enhancing immunity. Int. J. Sci. Eng. Res. 2020, 11, 817–825. [Google Scholar]
- El-Sekaily, A.; Helal, M.; Saad, A. Enhancement of immune tolerance of COVID19 patients might be achieved with alginate supplemented therapy. Int. J. Cancer Biomed. Res. 2020, 4, 21–26. [Google Scholar]
- Abdullahi, A.Y.; Kallon, S.; Yu, X.; Zhang, Y.; Li, G. Vaccination with Astragalus and Ginseng Polysaccharides Improves Immune Response of Chickens against H5N1 Avian Influenza Virus. Biomed. Res. Int. 2016, 2016, 1510264. [Google Scholar] [CrossRef] [Green Version]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan solution enhances both humoral and cell-mediated immune responses to subcutaneous vaccination. Vaccine 2007, 25, 2085–2094. [Google Scholar] [CrossRef] [Green Version]
- Ghendon, Y.; Markushin, S.; Krivtsov, G.; Akopova, I. Chitosan as an adjuvant for parenterally administered inactivated influenza vaccines. Arch. Virol. 2008, 153, 831–837. [Google Scholar] [CrossRef]
- Bolhassani, A.; Talebi, S.; Anvar, A. Endogenous and Exogenous Natural Adjuvants for Vaccine Development. Mini Rev. Med. Chem. 2017, 17, 1442–1456. [Google Scholar] [CrossRef]
- Zhang, Y.-Q.; Tsai, Y.-C.; Monie, A.; Hung, C.-F.; Wu, T.-C. Carrageenan as an adjuvant to enhance peptide-based vaccine potency. Vaccine 2010, 28, 5212–5219. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Dong, B.R.; Wu, T. Probiotics for preventing acute upper respiratory tract infections. Cochrane Database Syst. Rev. 2015, CD006895. [Google Scholar] [CrossRef]
- Ahanchian, H.; Jafari, S.A. Probiotics and prebiotics for prevention of viral respiratory tract infections. In Probiotics, Prebiotics, and Synbiotics, 1st ed.; Watson, R., Preedy, V., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 575–583. ISBN 9780128023716. [Google Scholar]
- Bourdillon, A.T.; Edwards, H.A. Review of probiotic use in otolaryngology. Am. J. Otolaryngol. 2021, 42, 102883. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, A.; Asadi, A.; Ghanavati, R.; Afifirad, R.; Darb Emamie, A.; Kakanj, M.; Talebi, M. The effect of probiotics on respiratory tract infection with special emphasis on COVID-19: Systemic review 2010-20. Int. J. Infect. Dis. 2021, 105, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Gohil, K.; Samson, R.; Dastager, S.; Dharne, M. Probiotics in the prophylaxis of COVID-19: Something is better than nothing. 3 Biotech. 2021, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Vouloumanou, E.K.; Makris, G.C.; Karageorgopoulos, D.E.; Falagas, M.E. Probiotics for the prevention of respiratory tract infections: A systematic review. Int. J. Antimicrob. Agents 2009, 34, 197.e1–197.e10. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Xu, K.; Du, T.; Zhu, P.; Liang, Z.; Liao, S.; Zhang, J.; Raizada, M.K.; Grant, M.B.; Li, Q. Expression of Human ACE2 in Lactobacillus and Beneficial Effects in Diabetic Retinopathy in Mice. Mol. Ther. Methods Clin. Dev. 2019, 14, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Khanzada, H.; Salam, A.; Qadir, M.B.; Phan, D.-N.; Hassan, T.; Munir, M.U.; Pasha, K.; Hassan, N.; Khan, M.Q.; Kim, I.S. Fabrication of Promising Antimicrobial Aloe Vera/PVA Electrospun Nanofibers for Protective Clothing. Materials 2020, 13, 3884. [Google Scholar] [CrossRef]
- Buluş, E.; Buluş, G.S.; Yakuphanoğlu, F. Production of polylactic acid activated charcoal nanofiber membranes for COVID-19 pandemic by electrospinning technique and determination of filtration efficiency. J. Mater. Electron. Devices 2020, 4, 21–26. [Google Scholar]
- Leung, W.W.F.; Sun, Q. Electrostatic charged nanofiber filter for filtering airborne novel coronavirus (COVID-19) and nano-aerosols. Sep. Purif. Technol. 2020, 250, 116886. [Google Scholar] [CrossRef]
- Zhang, Q.; Honko, A.; Zhou, J.; Gong, H.; Downs, S.N.; Vasquez, J.H.; Fang, R.H.; Gao, W.; Griffiths, A.; Zhang, L. Cellular nanosponges inhibit SARS-CoV-2 infectivity. Nano Lett. 2020, 20, 5570–5574. [Google Scholar] [CrossRef]
- Surnar, B.; Kamran, M.Z.; Shah, A.S.; Dhar, S. Clinically approved antiviral drug in an orally administrable nanoparticle for COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 1371–1380. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, K.J.; Webber, M.J.; Anderson, D.G. Materials for non-viral intracellular delivery of messenger RNA therapeutics. J. Control. Release 2016, 240, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Illum, L. Nasal drug delivery—Possibilities, problems and solutions. J. Control. Release 2003, 87, 187–198. [Google Scholar] [CrossRef]
- Maaz, A.; Blagbrough, I.S.; de Bank, P.A. In Vitro Evaluation of Nasal Aerosol Depositions: An Insight for Direct Nose to Brain Drug Delivery. Pharmaceutics 2021, 13, 1079. [Google Scholar] [CrossRef] [PubMed]
- Madav, Y.; Wairkar, S. Strategies for enhanced direct nose-to-brain drug delivery. In Direct Nose-to-Brain Drug Delivery, 1st ed.; Pardeshi, C.V., Souto, E.B., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 169–184. ISBN 9780128225226. [Google Scholar]
- Hua, S. Advances in Nanoparticulate Drug Delivery Approaches for Sublingual and Buccal Administration. Front. Pharmacol. 2019, 10, 1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, J.D. Buccal drug delivery. Expert Opin. Drug Deliv. 2005, 2, 507–517. [Google Scholar] [CrossRef]
- Czerkinsky, C.; Holmgren, J. Topical immunization strategies. Mucosal. Immunol. 2010, 3, 545–555. [Google Scholar] [CrossRef]
- Pabst, R. Mucosal vaccination by the intranasal route. Nose-associated lymphoid tissue (NALT)—Structure, function and species differences. Vaccine 2015, 33, 4406–4413. [Google Scholar] [CrossRef]
- Date, Y.; Ebisawa, M.; Fukuda, S.; Shima, H.; Obata, Y.; Takahashi, D.; Kato, T.; Hanazato, M.; Nakato, G.; Williams, I.R.; et al. NALT M cells are important for immune induction for the common mucosal immune system. Int. Immunol. 2017, 29, 471–478. [Google Scholar] [CrossRef]
- Shakya, A.K.; Chowdhury, M.Y.E.; Tao, W.; Gill, H.S. Mucosal vaccine delivery: Current state and a pediatric perspective. J. Control. Release 2016, 240, 394–413. [Google Scholar] [CrossRef] [Green Version]
- Patil, M. Mucoadhesion as a strategy to enhance the direct nose-to-brain drug delivery. In Direct Nose-to-Brain Drug Delivery, 1st ed.; Pardeshi, C.V., Souto, E.B., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 115–156. ISBN 9780128225226. [Google Scholar]
- Sudhakar, Y.; Kuotsu, K.; Bandyopadhyay, A.K. Buccal bioadhesive drug delivery—A promising option for orally less efficient drugs. J. Control. Release 2006, 114, 15–40. [Google Scholar] [CrossRef]
- Roy, S.; Pal, K.; Anis, A.; Pramanik, K.; Prabhakar, B.R. Polymers in Mucoadhesive Drug-Delivery Systems: A Brief Note. Des. Monomers Polym. 2009, 12, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Hanif, M.; Zaman, M.; Qureshi, S. Thiomers: A Blessing to Evaluating Era of Pharmaceuticals. Int. J. Polym. Sci. 2015, 2015, 146329. [Google Scholar] [CrossRef] [Green Version]
- Bernkop-Schnürch, A. Thiomers: A new generation of mucoadhesive polymers. Adv. Drug Deliv. Rev. 2005, 57, 1569–1582. [Google Scholar] [CrossRef]
- Brannigan, R.P.; Khutoryanskiy, V.V. Progress and Current Trends in the Synthesis of Novel Polymers with Enhanced Mucoadhesive Properties. Macromol. Biosci. 2019, 19, 1900194. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Aspinall, S.; Kaldybekov, D.B.; Buang, F.; Williams, A.C.; Khutoryanskiy, V.V. Synthesis and Evaluation of Methacrylated Poly(2-ethyl-2-oxazoline) as a Mucoadhesive Polymer for Nasal Drug Delivery. ACS Appl. Polym. Mater. 2021, 3, 5882–5892. [Google Scholar] [CrossRef]
- Gennari, C.G.M.; Sperandeo, P.; Polissi, A.; Minghetti, P.; Cilurzo, F. Lysozyme Mucoadhesive Tablets Obtained by Freeze-Drying. J. Pharm. Sci. 2019, 108, 3667–3674. [Google Scholar] [CrossRef] [PubMed]
- Dreijer-van der Glas, S.; Hafner, A. Nose. In Practical Pharmaceutics: An International Guideline for the Preparation, Care and Use of Medicinal Products, 1st ed.; Bouwman-Boer, Y., Fenton-May, V., le Brun, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 139–152. ISBN 978-3-319-15813-6. [Google Scholar]
- Al-Ghananeem, A.M.; Sandefer, E.P.; Doll, W.J.; Page, R.C.; Chang, Y.; Digenis, G.A. Gamma scintigraphy for testing bioequivalence: A case study on two cromolyn sodium nasal spray preparations. Int. J. Pharm. 2008, 357, 70–76. [Google Scholar] [CrossRef]
- Newman, S.P.; Pitcairn, G.R.; Dalby, R.N. Drug delivery to the nasal cavity: In vitro and in vivo assessment. Crit. Rev. Ther. Drug Carr. Syst. 2004, 21, 21–66. [Google Scholar] [CrossRef]
- Djupesland, P.G. Nasal drug delivery devices: Characteristics and performance in a clinical perspective-a review. Drug Deliv. Transl. Res. 2013, 3, 42–62. [Google Scholar] [CrossRef] [Green Version]
- Rygg, A.; Hindle, M.; Longest, P.W. Linking Suspension Nasal Spray Drug Deposition Patterns to Pharmacokinetic Profiles: A Proof-of-Concept Study Using Computational Fluid Dynamics. J. Pharm. Sci. 2016, 105, 1995–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiozzo Fasiolo, L.; Manniello, M.D.; Tratta, E.; Buttini, F.; Rossi, A.; Sonvico, F.; Bortolotti, F.; Russo, P.; Colombo, G. Opportunity and challenges of nasal powders: Drug formulation and delivery. Eur. J. Pharm. Sci. 2018, 113, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, M.H.; Conway, B.R.; Smith, A.M. Development of mucoadhesive sprayable gellan gum fluid gels. Int. J. Pharm. 2015, 488, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherafudeen, S.P.; Vasantha, P.V. Development and evaluation of in situ nasal gel formulations of loratadine. Res. Pharm. Sci. 2015, 10, 466–476. [Google Scholar] [PubMed]
- Cassano, R.; Servidio, C.; Trombino, S. Biomaterials for Drugs Nose–Brain Transport: A New Therapeutic Approach for Neurological Diseases. Materials 2021, 14, 1802. [Google Scholar] [CrossRef]
- Kouchak, M. In situ gelling systems for drug delivery. Jundishapur J. Nat. Pharm. Prod. 2014, 9, e20126. [Google Scholar] [CrossRef] [Green Version]
- Saindane, N.S.; Pagar, K.P.; Vavia, P.R. Nanosuspension based in situ gelling nasal spray of carvedilol: Development, in vitro and in vivo characterization. AAPS PharmSciTech 2013, 14, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, H.; Vaghela, D.; Patel, B.P. Brain targeted intranasal in-situ gelling spray of paroxetine: Formulation, characterization and in-vivo evaluation. J. Drug Deliv. Sci. Technol. 2021, 62, 102317. [Google Scholar] [CrossRef]
- Bianculli, R.H.; Mase, J.D.; Schulz, M.D. Antiviral Polymers: Past Approaches and Future Possibilities. Macromolecules 2020, 53, 9158–9186. [Google Scholar] [CrossRef]
- Tratnjek, L.; Sibinovska, N.; Kristan, K.; Kreft, M.E. In Vitro Ciliotoxicity and Cytotoxicity Testing of Repeated Chronic Exposure to Topical Nasal Formulations for Safety Studies. Pharmaceutics 2021, 13, 1750. [Google Scholar] [CrossRef]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Sjölinder, H.; Jonsson, A.B. Olfactory nerve—A novel invasion route of Neisseria meningitidis to reach the meninges. PLoS ONE 2010, 5, e14034. [Google Scholar] [CrossRef] [PubMed]
- Herbert, R.P.; Harris, J.; Chong, K.P.; Chapman, J.; West, A.K.; Chuah, M.I. Cytokines and olfactory bulb microglia in response to bacterial challenge in the compromised primary olfactory pathway. J. Neuroinflamm. 2012, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-W.; Chen, Y.-T.; Ho, H.-H.; Hsieh, P.-S.; Kuo, Y.-W.; Lin, J.-H.; Liu, C.-R.; Huang, Y.-F.; Chen, C.-W.; Hsu, C.-H.; et al. Lozenges with probiotic strains enhance oral immune response and health. Oral Dis. 2021, 00, 1–10. [Google Scholar] [CrossRef]
- Pattanshetty, S.; Narayana, A.; Radhakrishnan, R. Povidone-iodine gargle as a prophylactic intervention to interrupt the transmission of SARS-CoV-2. Oral Dis. 2021, 27, 752–753. [Google Scholar] [CrossRef]
- Shewale, J.G.; Gelhaus, H.C.; Ratcliff, J.L.; Hernandez-Kapila, Y.L. In vitro antiviral activity of stabilized chlorine dioxide containing oral care products. Oral Dis. 2021, 00, 1–8. [Google Scholar] [CrossRef]
- Schütz, D.; Conzelmann, C.; Fois, G.; Groß, R.; Weil, T.; Wettstein, L.; Stenger, S.; Zelikin, A.; Hoffmann, T.K.; Frick, M.; et al. Carrageenan-containing over-the-counter nasal and oral sprays inhibit SARS-CoV-2 infection of airway epithelial cultures. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L750–L756. [Google Scholar] [CrossRef]
- Mattes, R.D. Physiologic responses to sensory stimulation by food: Nutritional implications. J. Am. Diet. Assoc. 1997, 97, 406–413. [Google Scholar] [CrossRef]
- Campos, J.C.; Cunha, D.; Ferreira, D.C.; Reis, S.; Costa, P.J. Oromucosal precursors of in loco hydrogels for wound-dressing and drug delivery in oral mucositis: Retain, resist, and release. Mater. Sci. Eng. C. Mater. Biol. Appl. 2021, 118, 111413. [Google Scholar] [CrossRef]
- Dreijer-van der Glas, S. Oropharynx. In Practical Pharmaceutics: An International Guideline for the Preparation, Care and Use of Medicinal Products, 1st ed.; Bouwman-Boer, Y., Fenton-May, V., le Brun, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 131–138. ISBN 978-3-319-15813-6. [Google Scholar]
- Hassan, A.O.; Kafai, N.M.; Dmitriev, I.P.; Fox, J.M.; Smith, B.K.; Harvey, I.B.; Chen, R.E.; Winkler, E.S.; Wessel, A.W.; Case, J.B.; et al. A Single-Dose Intranasal ChAd Vaccine Protects Upper and Lower Respiratory Tracts against SARS-CoV-2. Cell 2020, 183, 169–184. [Google Scholar] [CrossRef]
- An, X.; Martinez-Paniagua, M.; Rezvan, A.; Sefat, S.R.; Fathi, M.; Singh, S.; Biswas, S.; Pourpak, M.; Yee, C.; Liu, X.; et al. Single-dose intranasal vaccination elicits systemic and mucosal immunity against SARS-CoV-2. iScience 2021, 24, 103037. [Google Scholar] [CrossRef]
- Bricker, T.L.; Darling, T.L.; Hassan, A.O.; Harastani, H.H.; Soung, A.; Jiang, X.; Dai, Y.-N.; Zhao, H.; Adams, L.J.; Holtzman, M.J.; et al. A single intranasal or intramuscular immunization with chimpanzee adenovirus-vectored SARS-CoV-2 vaccine protects against pneumonia in hamsters. Cell Rep. 2021, 36, 109400. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.O.; Feldmann, F.; Zhao, H.; Curiel, D.T.; Okumura, A.; Tang-Huau, T.-L.; Case, J.B.; Meade-White, K.; Calli-son, J.; Chen, R.E.; et al. A single in-tranasal dose of chimpanzee adenovirus-vectored vaccine protects against SARS-CoV-2 infection in rhesus macaques. Cell Rep. Med. 2021, 2, 100230. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Purushotham, J.N.; Schulz, J.E.; Holbrook, M.G.; Bushmaker, T.; Carmody, A.; Port, J.R.; Yinda, C.K.; Okumura, A.; Saturday, G.; et al. Intranasal ChAdOx1 nCoV-19/AZD1222 Vaccination Reduces Shed-Ding of SARS-CoV-2 D614G in Rhesus Macaques. 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.01.09.426058v1 (accessed on 26 November 2021).
- A Study of Intranasal ChAdOx1 nCOV-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04816019?term=chadox1+ncov-19+intranasal&draw=2&rank=1 (accessed on 26 November 2021).
- King, R.G.; Silva-Sanchez, A.; Peel, J.N.; Botta, D.; Meza-Perez, S.; Allie, R.; Schultz, M.D.; Liu, M.; Bradley, J.E.; Qiu, S.; et al. Single-dose Intranasal Administration of AdCOVID Elicits Systemic and Mucosal Immunity Against SARS-CoV-2 in Mice. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.10.10.331348v1 (accessed on 26 November 2021).
- Nasal Spray Vaccine for Covid-19. Available online: https://www.clinicaltrialsarena.com/comment/nasal-spray-vaccine-covid-19/ (accessed on 26 November 2021).
- Safety and Immunogenicity of an Intranasal RSV Vaccine Expressing SARS-CoV-2 Spike Protein (COVID-19 Vaccine) in Adults. Available online: https://clinicaltrials.gov/ct2/show/NCT04798001?term=meissa+vaccines&draw=2&rank=2 (accessed on 27 November 2021).
- Chavda, V.P.; Vora, L.K.; Pandya, A.K.; Patravale, V.B. Intranasal vaccines for SARS-CoV-2: From challenges to potential in COVID-19 management. Drug Discov. Today 2021, 26, 2619–2636. [Google Scholar] [CrossRef] [PubMed]
- Intravacc Announces Positive Pre-clinical Data for Its SARS-CoV-2 Nose Spray Vaccine. Available online: https://www.intravacc.nl/news/intravacc-announces-positive-pre-clinical-data-intranasal-sars-cov-2-candidate-vaccine/ (accessed on 27 November 2021).
- Ku, Z.; Xie, X.; Hinton, P.R.; Liu, X.; Ye, X.; Muruato, A.E.; Ng, D.C.; Biswas, S.; Zou, J.; Liu, Y.; et al. Nasal delivery of an IgM offers broad protection from SARS-CoV-2 variants. Nature 2021, 595, 718–723. [Google Scholar] [CrossRef]
- Ku, Z.; Xie, X.; Davidson, E.; Ye, X.; Su, H.; Menachery, V.D.; Li, Y.; Yuan, Z.; Zhang, X.; Muruato, A.E.; et al. Molecular determinants and mechanism for antibody cocktail preventing SARS-CoV-2 escape. Nat. Commun. 2021, 12, 469. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Xiang, J.; Cui, Z.; Liu, J.; Liu, C. Intranasal Administration of SARS-CoV-2 Neutralizing Human Antibody Prevents Infection in mice. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.12.08.416677v1 (accessed on 30 November 2021).
- Eureka Therapeutics Announces Successful Preclinical Results of InvisiMask™ Human Antibody Nasal Spray against SARS-CoV-2 Infection. Available online: https://www.eurekatherapeutics.com/media/press-releases/121420/ (accessed on 30 November 2021).
- Lepelletier, D.; Maillard, J.Y.; Pozzetto, B.; Simon, A. Povidone Iodine: Properties, Mechanisms of Action, and Role in Infection Control and Staphylococcus aureus Decolonization. Antimicrob. Agents Chemother. 2020, 64, e00682-20. [Google Scholar] [CrossRef]
- Carrouel, F.; Gonçalves, L.S.; Conte, M.P.; Campus, G.; Fisher, J.; Fraticelli, L.; Gadea-Deschamps, E.; Ottolenghi, L.; Bourgeois, D. Antiviral Activity of Reagents in Mouth Rinses against SARS-CoV-2. J. Dent. Res. 2021, 100, 124–132. [Google Scholar] [CrossRef]
- Anderson, D.E.; Sivalingam, V.; Kang, A.E.Z.; Ananthanarayanan, A.; Arumugam, H.; Jenkins, T.M.; Hadjiat, Y.; Eggers, M. Povidone-iodine demonstrates rapid in vitro virucidal activity against SARS-CoV-2, the virus causing COVID-19 disease. Infect. Dis. Ther. 2020, 9, 669–675. [Google Scholar] [CrossRef]
- Hassandarvish, P.; Tiong, V.; Sazaly, A.B.; Mohamed, N.A.; Arumugam, H.; Ananthanarayanan, A.; Qasuri, M.; Hadjiat, Y. Povidone iodine gargle and mouthwash. Br. Dent. J. 2020, 228, 900. [Google Scholar] [CrossRef]
- Challacombe, S.J.; Kirk-Bayley, J.; Sunkaraneni, V.S.; Combes, J. Povidone iodine. Br. Dent. J. 2020, 228, 656–657. [Google Scholar] [CrossRef] [PubMed]
- Bidra, A.S.; Pelletier, J.S.; Westover, J.B.; Frank, S.; Brown, S.M.; Tessema, B. Rapid In-Vitro Inactivation of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Using Povidone-Iodine Oral Antiseptic Rinse. J. Prosthodont. 2020, 29, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Martínez Lamas, L.; Diz Dios, P.; Pérez Rodríguez, M.T.; del Campo Pérez, V.; Cabrera Alvargonzalez, J.J.; López Domínguez, A.M.; Fernandez Feijoo, J.; Diniz Freitas, M.; Limeres Posse, J. Is povidone iodine mouthwash effective against SARS-CoV-2? First in vivo tests. Oral Dis. 2020, 00, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Effect of Local Treatment (Carrageenan Nasal Spray and PVP-I Mouthwash) in Reducing Viral Load in Patients with COVID-19 (LT-COVID19). Available online: https://clinicaltrials.gov/ct2/show/NCT05049213?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=5 (accessed on 1 December 2021).
- Arefin, M.K. Povidone Iodine (PVP-I) Oro-Nasal Spray: An Effective Shield for COVID-19 Protection for Health Care Worker (HCW), for all. Indian J. Otolaryngol. Head Neck Surg. 2021, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.J.; Rumi, S.K.N.F.; Banu, S.S.; Uddin, A.K.M.N.; Islam, M.S.; Arefin, M.K. Virucidal effect of povidone iodine on COVID-19 in the nasopharynx: A structured summary of a study protocol for an open-label randomized clinical trial. Trials 2021, 22, 2. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Chlorine Dioxide (CLO2) As a Non-Toxic Antimicrobial Agent for Virus, Bacteria and Yeast (Candida Albicans). Int. J. Vaccines Vaccin. 2016, 2, 52. [Google Scholar] [CrossRef] [Green Version]
- Giarratana, N.; Rajan, B.; Kamala, K.; Mendenhall, M.; Reiner, G. A sprayable Acid-Oxidizing solution containing hypochlorous acid (AOS2020) efficiently and safely inactivates SARS-Cov-2: A new potential solution for upper respiratory tract hygiene. Eur. Arch. Otorhinolaryngol. 2021, 278, 3099–3103. [Google Scholar] [CrossRef]
- Use of Hypochlorous Acid as Prophylaxis in Health Personnel at High Risk of Infection by SARS-CoV 2 (COVID19). Available online: https://clinicaltrials.gov/ct2/show/NCT04684550?term=HCLO&cond=SARS (accessed on 4 December 2021).
- Shmuel, K.; Dalia, M.; Tair, L.; Yaakov, N. Low pH Hypromellose (Taffix) nasal powder spray could reduce SARS-CoV-2 infection rate post mass-gathering event at a highly endemic community: An observational prospective open label user survey. Expert Rev. Anti-Infect. Ther. 2021, 19, 1325–1330. [Google Scholar] [CrossRef]
- Bentley, K.; Stanton, R.J. Hydroxypropyl Methylcellulose-Based Nasal Sprays Effectively Inhibit In Vitro SARS-CoV-2 Infection and Spread. Viruses 2021, 13, 2345. [Google Scholar] [CrossRef]
- Paull, J.R.A.; Heery, G.P.; Bobardt, M.D.; Castellarnau, A.; Luscombe, C.A.; Fairley, J.K.; Gallay, P.A. Virucidal and antiviral activity of astodrimer sodium against SARS-CoV-2 in vitro. Antiviral Res. 2021, 191, 105089. [Google Scholar] [CrossRef]
- Paull, J.R.A.; Luscombe, C.A.; Castellarnau, A.; Heery, G.P.; Bobardt, M.D.; Gallay, P.A. Protective Effects of Astodrimer Sodium 1% Nasal Spray Formulation against SARS-CoV-2 Nasal Challenge in K18-hACE2 Mice. Viruses 2021, 13, 1656. [Google Scholar] [CrossRef] [PubMed]
- Pyrć, K.; Milewska, A.; Duran, E.B.; Botwina, P.; Dabrowska, A.; Jedrysik, M.; Benedyk, M.; Lopes, R.; Arenas-Pinto, A.; Badr, M.; et al. SARS-CoV-2 inhibition using a mucoadhesive, amphiphilic chitosan that may serve as an anti-viral nasal spray. Sci Rep. 2021, 11, 20012. [Google Scholar] [CrossRef]
- Effectiveness of a Novel Respirator with Chitosan Nanoparticles. Available online: https://clinicaltrials.gov/ct2/show/NCT04490200 (accessed on 5 December 2021).
- Koenighofer, M.; Lion, T.; Bodenteich, A.; Prieschl-Grassauer, E.; Grassauer, A.; Unger, H.; Mueller, C.A.; Fazekas, T. Carrageenan nasal spray in virus confirmed common cold: Individual patient data analysis of two randomized controlled trials. Multidiscip. Respir. Med. 2014, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Eccles, R.; Winther, B.; Johnston, S.L.; Robinson, P.; Trampisch, M.; Koelsch, S. Efficacy and safety of iota-carrageenan nasal spray versus placebo in early treatment of the common cold in adults: The ICICC trial. Respir. Res. 2015, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prophylaxis Covid-19 in Healthcare Agents by Intensive Treatment with Ivermectin and Iota-carrageenan (Ivercar-Tuc). Available online: https://clinicaltrials.gov/ct2/show/NCT04701710?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=3 (accessed on 7 December 2021).
- Carrageenan Nasal Spray for COVID-19 Prophylaxis (ICE-COVID). Available online: https://clinicaltrials.gov/ct2/show/NCT04590365?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=4 (accessed on 7 December 2021).
- Leibbrandt, A.; Meier, C.; König-Schuster, M.; Weinmüllner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; et al. Iota-carrageenan is a potent inhibitor of influenza A virus infection. PLoS ONE 2010, 5, e14320. [Google Scholar] [CrossRef] [PubMed]
- Usefulness of Topic Ivermectin and Carrageenan to Prevent Contagion of Covid 19 (IVERCAR). Available online: https://clinicaltrials.gov/ct2/show/results/NCT04425850?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=2 (accessed on 13 December 2021).
- Efficacy of a Nasal Spray Containing Iota-Carrageenan in the Prophylaxis of COVID-19 Disease in Health Personnel Dedicated to Patients Care With COVID-19 Disease (CARR-COV-02). Available online: https://clinicaltrials.gov/ct2/show/NCT04521322?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=1 (accessed on 13 December 2021).
- Prophylactic Treatment with Carragelose Nasal Spary to Prevent SARS-CoV-2, COVID-19, Infections in Health Care Workers. Available online: https://clinicaltrials.gov/ct2/show/NCT04681001?term=carrageenan&cond=SARS-CoV2+Infection&draw=2&rank=6 (accessed on 13 December 2021).
- Bansal, S.; Jonsson, C.B.; Taylor, S.L.; Figueroa, J.M.; Dugour, A.V.; Palacios, C.; Vega, J.C. Iota-carrageenan and xylitol inhibit SARS-CoV-2 in Vero cell culture. PLoS ONE 2021, 16, e0259943. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; del Vecchio, C.; Franchin, E.; et al. Lactoferrin as Antiviral Treatment in COVID-19 Management: Preliminary Evidence. Int. J. Environ. Res. Public Health 2021, 18, 10985. [Google Scholar] [CrossRef]
- Campione, E.; Cosio, T.; Rosa, L.; Lanna, C.; di Girolamo, S.; Gaziano, R.; Valenti, P.; Bianchi, L. Lactoferrin as Protective Natural Barrier of Respiratory and Intestinal Mucosa against Coronavirus Infection and Inflammation. Int. J. Mol. Sci. 2020, 21, 4903. [Google Scholar] [CrossRef]
- Winchester, S.; John, S.; Jabbar, K.; John, I. Clinical efficacy of nitric oxide nasal spray (NONS) for the treatment of mild COVID-19 infection. J. Infect. 2021, 83, 237–279. [Google Scholar] [CrossRef]
- Garren, M.R.; Ashcraft, M.; Qian, Y.; Douglass, M.; Brisbois, E.J.; Handa, H. Nitric oxide and viral infection: Recent developments in antiviral therapies and platforms. Appl. Mater. Today 2021, 22, 100887. [Google Scholar] [CrossRef]
- Meng, Z.; Wang, T.; Chen, L.; Chen, X.; Li, L.; Qin, X.; Li, H.; Luo, J. The Effect of Recombinant Human Interferon Alpha Nasal Drops to Prevent COVID-19 Pneumonia for Medical Staff in an Epidemic Area. Curr. Top. Med. Chem. 2021, 21, 920–927. [Google Scholar] [CrossRef]
- Lee, A.C.; Jeong, Y.; Lee, S.; Jang, H.; Zheng, A.; Kwon, S.; Repine, J.E. Nasopharyngeal Type-I Interferon for Immediately Available Prophylaxis Against Emerging Respiratory Viral Infections. Front. Immunol. 2021, 12, 660298. [Google Scholar] [CrossRef]
- Alibek, K.; Tskhay, A. Ahead of a vaccine: A safe method of protection against COVID-19 exists. Res. Ideas Outcomes 2020, 6, e61709. [Google Scholar] [CrossRef]
- Cannon, M.L.; Westover, J.B.; Bleher, R.; Sanchez-Gonzalez, M.A.; Ferrer, G. In Vitro Analysis of the Anti-viral Potential of Nasal Spray Constituents against SARS-CoV-2. 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.12.02.408575v3 (accessed on 13 December 2021).
- Kontiokari, T.; Uhari, M.; Koskela, M. Antiadhesive effects of xylitol on otopathogenic bacteria. J. Antimicrob. Chemother. 1998, 41, 563–565. [Google Scholar] [CrossRef] [Green Version]
- A Study to Evaluate the Efficacy of Xlear vs. Placebo for Acute COVID-19 Infection. Available online: https://clinicaltrials.gov/ct2/show/NCT04858620 (accessed on 13 December 2021).
- Gudmundsdottir, Á.; Scheving, R.; Lindberg, F.; Stefansson, B. Inactivation of SARS-CoV-2 and HCoV-229E In Vitro by ColdZyme® a medical device mouth spray against the common cold. J. Med. Virol. 2021, 93, 1792–1795. [Google Scholar] [CrossRef]
- Davison, G.; Perkins, E.; Jones, A.W.; Swart, G.M.; Jenkins, A.R.; Robinson, H.; Dargan, K. Coldzyme® Mouth Spray reduces duration of upper respiratory tract infection symptoms in endurance athletes under free living conditions. Eur. J. Sport Sci. 2021, 21, 771–780. [Google Scholar] [CrossRef]
- Posch, W.; Vosper, J.; Zaderer, V.; Noureen, A.; Constant, S.; Bellmann-Weiler, R.; Lass-Flörl, C.; Wilflingseder, D. ColdZyme Maintains Integrity in SARS-CoV-2-Infected Airway Epithelia. mBio 2021, 12, e00904-21. [Google Scholar] [CrossRef]
- Nagai, M.; Moriyama, M.; Ichinohe, T. Oral Bacteria Combined with an Intranasal Vaccine Protect from Influenza A Virus and SARS-CoV-2 Infection. mBio 2021, 12, e0159821. [Google Scholar] [CrossRef]
- Paolacci, S.; Ceccarini, M.R.; Codini, M.; Manara, E.; Tezzele, S.; Percio, M.; Capodicasa, N.; Kroni, D.; Dundar, M.; Ergoren, M.C.; et al. Pilot study for the evaluation of safety profile of a potential inhibitor of SARS-CoV-2 endocytosis. Acta Biomed. 2020, 91, e2020009. [Google Scholar]
- Efficacy and Safety of Longan Extract Spray (P80 Spray) in Volunteers with Coronavirus Disease 2019 (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/study/NCT04887233 (accessed on 15 December 2021).
- Gugleva, V.; Ivanova, N.; Sotirova, Y.; Andonova, V. Dermal Drug Delivery of Phytochemicals with Phenolic Structure via Lipid-Based Nanotechnologies. Pharmaceuticals 2021, 14, 837. [Google Scholar] [CrossRef]
- Bioflavonoid Spray Efficacy Against Respiratory Viruses. Available online: https://www.oralhealthgroup.com/features/bioflavonoid-spray-efficacy-against-respiratory-viruses/ (accessed on 15 December 2021).
- Godinho, P.I.C.; Soengas, R.G.; Silva, V.L.M. Therapeutic Potential of Glycosyl Flavonoids as Anti-Coronaviral Agents. Pharmaceuticals 2021, 14, 546. [Google Scholar] [CrossRef]
- Proud, P.C.; Tsitoura, D.; Watson, R.J.; Chua, B.Y.; Aram, M.J.; Bewley, K.R.; Cavell, B.E.; Cobb, R.; Dowall, S.; Fotheringham, S.A.; et al. Prophylactic intranasal administration of a TLR2/6 agonist reduces upper respiratory tract viral shedding in a SARS-CoV-2 challenge ferret model. EBioMedicine 2021, 63, 103153. [Google Scholar] [CrossRef]
- Deliyannis, G.; Wong, C.Y.; McQuilten, H.A.; Bachem, A.; Clarke, M.; Jia, X.; Horrocks, K.; Zeng, W.; Girkin, J.; Scott, N.E.; et al. TLR2-mediated activation of innate responses in the upper airways confers antiviral protection of the lungs. JCI Insight 2021, 6, e140267. [Google Scholar] [CrossRef]
- Pilicheva, B.; Boyuklieva, R. Can the Nasal Cavity Help Tackle COVID-19? Pharmaceutics 2021, 13, 1612. [Google Scholar] [CrossRef]
- Zaidi, A.K.; Dehgani-Mobaraki, P. Retracted article: The mechanisms of action of Ivermectin against SARS-CoV-2: An evidence-based clinical review article. J. Antibiot. 2021, 75, 1–13. [Google Scholar] [CrossRef]
- Jokicevic, K.; Kiekens, S.; Byl, E.; de Boeck, I.; Cauwenberghs, E.; Lebeer, S.; Kiekens, F. Probiotic nasal spray development by spray drying. Eur. J. Pharm. Biopharm. 2021, 159, 211–220. [Google Scholar] [CrossRef]
- Almanza-Reyes, H.; Moreno, S.; Plascencia-López, I.; Alvarado-Vera, M.; Patrón-Romero, L.; Borrego, B.; Reyes-Escamilla, A.; Valencia-Manzo, D.; Brun, A.; Pestryakov, A.; et al. Evaluation of silver nanoparticles for the prevention of SARS-CoV-2 infection in health workers: In vitro and in vivo. PLoS ONE 2021, 16, e0256401. [Google Scholar]
- Jeremiah, S.S.; Miyakawa, K.; Morita, T.; Yamaoka, Y.; Ryo, A. Potent antiviral effect of silver nanoparticles on SARS-CoV-2. Biochem. Biophys. Res. Commun. 2020, 533, 195–200. [Google Scholar] [CrossRef]
- Colunga Biancatelli, R.M.L.; Berrill, M.; Catravas, J.D.; Marik, P.E. Quercetin and Vitamin C: An Experimental, Synergistic Therapy for the Prevention and Treatment of SARS-CoV-2 Related Disease (COVID-19). Front. Immunol. 2020, 11, 1451. [Google Scholar] [CrossRef]
- Williamson, G.; Kerimi, A. Testing of natural products in clinical trials targeting the SARS-CoV-2 (Covid-19) viral spike protein-angiotensin converting enzyme-2 (ACE2) interaction. Biochem. Pharmacol. 2020, 178, 114123. [Google Scholar] [CrossRef]
- Dejani, N.N.; Elshabrawy, H.A.; Bezerra Filho, C.d.S.M.; de Sousa, D.P. Anticoronavirus and Immunomodulatory Phenolic Compounds: Opportunities and Pharmacotherapeutic Perspectives. Biomolecules 2021, 11, 1254. [Google Scholar] [CrossRef]
- Rossi, G.A.; Sacco, O.; Capizzi, A.; Mastromarino, P. Can Resveratrol-Inhaled Formulations Be Considered Potential Adjunct Treatments for COVID-19? Front. Immunol. 2021, 12, 670955. [Google Scholar] [CrossRef]
- Da Silva, J.K.R.; Figueiredo, P.L.B.; Byler, K.G.; Setzer, W.N. Essential oils as antiviral agents. Potential of essential oils to treat SARS-CoV-2 infection: An in-silico investigation. Int. J. Mol. Sci. 2020, 21, 3426. [Google Scholar] [CrossRef]
- Asif, M.; Saleem, M.; Saadullah, M.; Yaseen, H.S.; Al Zarzour, R. COVID-19 and therapy with essential oils having antiviral, anti-inflammatory, and immunomodulatory properties. Inflammopharmacology 2020, 28, 1153–1161. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuy, B.T.P.; My, T.T.A.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thi Phuong Loan, H.; Triet, N.T.; Anh, T.T.V.; Quy, P.T.; Tat, P.V.; et al. Investigation into SARS-CoV-2 Resistance of Compounds in Garlic Essential Oil. ACS Omega 2020, 5, 8312–8320. [Google Scholar] [CrossRef]
- Baker, N.; Williams, A.J.; Tropsha, A.; Ekins, S. Repurposing Quaternary Ammonium Compounds as Potential Treatments for COVID-19. Pharm. Res. 2020, 37, 104. [Google Scholar] [CrossRef]
- Abdulrab, S.; Alkadasi, B.; Al-Maweri, S.; Halboub, E.; Alhadainy, H.; Geerts, G. Statins-based prophylactic mouthwash and nasal spray may protect against coronavirus disease 2019. New Microbes New Infect. 2020, 37, 100751. [Google Scholar] [CrossRef]
- Vofo, G.; Brodie, R.; Gross, M. Nasal lavage containing Angiotensin-Converting Enzyme-2 agonist can prevent and reduce viral load in COVID-19. Med. Hypotheses 2020, 144, 110207. [Google Scholar] [CrossRef]
- Yoon, J.G.; Yoon, J.; Song, J.Y.; Yoon, S.Y.; Lim, C.S.; Seong, H.; Noh, J.Y.; Cheong, H.J.; Kim, W.J. Clinical Significance of a High SARS-CoV-2 Viral Load in the Saliva. J. Korean Med. Sci. 2020, 35, e195. [Google Scholar] [CrossRef]
- Aydemir, D.; Ulusu, N.N. Correspondence: Angiotensin-converting enzyme 2 coated nanoparticles containing respiratory masks, chewing gums and nasal filters may be used for protection against COVID-19 infection. Travel Med. Infect. Dis. 2020, 37, 101697. [Google Scholar] [CrossRef] [PubMed]
- Herrera, D.; Serrano, J.; Roldán, S.; Sanz, M. Is the oral cavity relevant in SARS-CoV-2 pandemic? Clin. Oral Investig. 2020, 24, 2925–2930. [Google Scholar] [CrossRef] [PubMed]
- Bidra, A.S.; Pelletier, J.S.; Westover, J.B.; Frank, S.; Brown, S.M.; Tessema, B. Comparison of in vitro inactivation of SARS CoV-2 with hydrogen peroxide and povidone-iodine oral antiseptic rinses. J. Prosthodont. 2020, 29, 599–603. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Active Agent | Form of Application | Type of Studies Conducted/in Progress | Efficacy | Mechanism of Action | Available Marketed Product(s) | References |
|---|---|---|---|---|---|---|
| I. ANTIGEN-SPECIFIC PROPHYLAXIS | ||||||
| Adenovirus-vectored vaccine encoding the spike (S)-protein/RBD domain of SARS-CoV-2 | Intranasal |
| SARS-CoV-2 | Induction of S-specific and receptor binding domain (RBD)-specific serum and secretory antibodies (IgG and IgA, respectively), and lung-resident T cells | n/a a | [236,237,238,239,240,241,242] |
| Genetically modified live-attenuated vaccines | Intranasal |
| SARS-CoV-2 | Induction of S-specific and receptor binding domain (RBD)-specific secretory antibodies (IgA) | n/a | [243,244,245] |
| S-protein embedded bacterial outer membrane vesicles | Intranasal |
| SARS-CoV-2 | Induction of SARS-CoV-2-specific neutralizing antibodies | n/a | [246] |
| Monoclonal IgM antibodies | Intranasal |
| SARS-CoV-2 | Antigen-specific binding to SARS-CoV-2 RBD domain | n/a | [247,248] |
| Monoclonal IgG antibodies | Nasal spray |
| SARS-CoV-2 | Antigen-specific binding to SARS-CoV-2 RBD domain | InvisiMask™ (Eureka Therapeutics, USA) | [249,250] |
| II. BROAD-SPECTRUM ANTISEPTICS | ||||||
| Povidone-iodine (PVP-I) | Oral rinse |
| Broad-spectrum antimicrobial activity, including against SARS-CoV, MERS, and SARS-CoV-2 | Oxidation-determined damaging on microbial nucleic acids, proteins, and cell membranes | Betadine® 1% (Avrio Health L.P. Purdue Pharma Inc., USA), Halodine® 1.7% (Halodine LLC, USA) | [230,251,252,253,254,255,256,257,258,259,260] |
| Gargling solution | Betadine® gargle 0.5% (Avrio Health L.P. Pur-due Pharma Inc., USA) | |||||
| Oral and nasal spray |
| n/a | ||||
| Nasal irrigation |
| n/a | ||||
| Chlorine dioxide (stabilized) | Oral rinse |
| Broad-spectrum antimicrobial activity, including against HCoV-229E, Influenza A, SARS-CoV, and SARS-CoV-2 | Oxidation-determined damaging on microbial nucleic acids, proteins, and cell membranes | ClōSYS® oral care products (Rowpar Pharmaceuticals, Inc., USA) | [231,261] |
| Hypochlorous acid (HCLO) | Nasal and oral spray |
| Broad-spectrum antibacterial activity, including against SARS-CoV-2 | Oxidation-determined damaging on viral genome | n/a Tehclo™ technology platform (APR Nanotechnologies SA, Switzerland) for delivery of HCLO—Acid-Oxidizing Solution (AOS2020) is used | [262,263] |
| III. POLYMERIC ANTIMICROBIAL AGENTS | ||||||
| Hydroxypropyl methyl cellulose | Nasal spray, Nasal powder spray |
| Nonspecific mucosal protection against respiratory-transmitted pathogens | Formation of a physical barrier on mucosa to facilitate the process of pathogen entrapment and clearance; additionally, Taffix™ (Nasus Pharma Ltd., Israel) nasal spray creates an acidic microenviroment, unfavorable for most respiratory pathogens | Taffix™ (Nasus Pharma Ltd., Israel) nasal spray, Nasaleze nasal spray | [264,265] |
| Astodrimer sodium | Nasal spray |
| Broad-spectrum antiviral activity, including against SARS-CoV-2 | Blocking of viral adhesion and entry by binding to viral adhesins and cell entry factors, such as heparan sulfate proteoglycans | Viraleze™ (Starpharma Pty Ltd., Australia) | [266,267] |
| Chitosan and Chitosan derivatives | Nasal spray |
| Broad -spectrum antiviral activity, including against MERS and SARS-CoV-2 | Binding to and blocking of the SARS-CoV-2 S-protein and RBD and subsequent inhibition of the receptor–adhesin connection and viral entry | n/a | [146,147,268,269] |
| Semifacial respirator with chitosan nanoparticles |
| n/a | ||||
| Iota-, Lambda-, and Kapa Carrageenan | Oral spray |
| Broad-spectrum antiviral activity, including against SARS-CoV-2 | Formation of a negatively charged protective barrier on mucus membranes, viral binding and subsequent viral aggregation, and decreased viral attachment and entry | Coldamaris® (Sigmapharm Arzneimittel GmbH, Austria) Throat Spray, GripVis, Viruseptin, Iovir® (Cube Pharma & Nutrition, Greece) Throat spray | [15,132,149,150,270,271,272,273,274,275,276,277,278] |
| Lozenges |
| Coldamaris® (Sigmapharm Arzneimittel GmbH, Austria) lozenges, Betadine® (Avrio Health L.P. Purdue Pharma Inc., USA) lozenges, Lontax Gola, Viruseptin® (Beampoint AB Joint-stock company, Sweden) lozenges | ||||
| Nasal spray |
| Flo™ Travel nasal spray (ENT Technologies Pty Ltd., Australia), Coldamaris pro. ® (Sigmapharm Arzneimittel GmbH, Austria) nasal spray, GripVis, Betadine® (Avrio Health L.P. Pur-due Pharma Inc., USA) Cold Defence nasal spray, Viruseptin® (Beampoint AB Joint-stock company, Sweden), Agovirax® (GryNumber Health, Ltd., Lithuania), Iovir® (Cube Pharma & Nutrition, Greece) nasal spray, Nasitrol nasal spray | ||||
| IV. NON-POLYMERIC NATURAL ANTIMICROBIAL AGENTS | ||||||
| Lactoferrin | Intranasal liposomal suspension |
| Broad-spectrum antimicrobial activity, including against SARS-CoV-2 | Binding with endogenous cell entry factors, such as heparan sulfate proteoglycans, and SARS-CoV-2 S-protein | n/a | [279,280] |
| Nitric oxide (NO) | Nasal spray |
| Component of the innate immunity | Mediator of immune-inflammatory cascade defense mechanisms | Nitric oxide-releasing systems (NORS™)—Enovid™/VirX™ (SaNOtize Research and Development Corp., Canada) | [281,282] |
| Type I Interferon | Nasal drops |
| Component of the first-line nonspecific (innate) immunity against viruses | Prevents a delay in INF-I natural response | n/a | [283,284,285] |
| Xylitol | Nasal spray |
| Antimicrobial activity, including against RSV and SARS-CoV-2 | Anti-adhesive effects, prebiotic activity | Xlear® (SaNOtize Research and Development Corp., Canada) sinus care products | [278,286,287,288] |
| Trypsin (from cod) + Glycerol | Oral spray |
| Antiviral activity, including against HCoV-229E and SARS-CoV-2 | Formation of a physical barrier on the mucosa that entraps viral particles (Glycerol); proteolytic activity of Trypsin on trypsin-susceptible sites along the SARS-CoV-2 S-protein | ColdZyme® (SaNOtize Research and Development Corp., Canada) mouth spray, Viruprotect mouth spray | [289,290,291] |
| Lysozyme (as pretreatment agent prior to nasal vaccination) | Intranasal |
| Influenza, SARS-CoV-2 | Disruption of nasal bacteria and subsequent release of pathogen-associated molecular patterns, which may act as adjuvants to enhance the virus-specific antibody response to vaccination | n/a | [292] |
| Hydroxytyrosol + α-cyclodextrin | Nasal spray |
| Broad-spectrum antiviral activity | Depletion of sphingolipids from the lipid rafts where the ACE2 receptor, specific for SARS-CoV-2, localizes (α-cyclodextrin) | Endovir Stop | [293] |
| Longan extract (rich in polyphenols) | Nasal spray |
| Potential antiviral, including anti-SARS-CoV-2 activity | Potentially owed to the polyphenols’ unspecific antiviral and anti-inflammatory properties | n/a | [294] |
| Oral spray | P80 Throat spray | |||||
| Flavonoid complex (patent combination Flavabac) | Oral spray |
| Broad-spectrum antimicrobial activity, including against SARS-CoV-2 | Antiviral activity of flavonoids is owed to their anti-oxidant properties and ability to interact with key enzymes, receptors, and membranes | Cold & Flu Guard oral spray | [295,296,297] |
| V. IMMUNE-MODULATORS/ THERAPEUTICS WITH IMMUNE-MODULATORY ACTIVITY | ||||||
| INNA-051 (synthetic PEGylated TLR2/6 agonist) | Nasal spray |
| SARS-CoV-2 | Reduction of the time required for nasal epithelial cells to initiate the innate immune responses following virus exposure | n/a | [298,299,300] |
| Ivermectin | Oral (buccal) drops |
| Antiviral activity, including against SARS-CoV-2 | Interaction with SARS-CoV-2 S-protein and host cells entry factors, and subsequent inhibition of viral cell entry; blocking of the nuclear transport of the SARS-CoV-2 viral proteins by action on Importin superfamily; blocking of viral replication and assembly; immune-modulatory properties | n/a | [272,275,301] |
| VI. OTHER | ||||||
| Probiotic strain Lacticaseibacillus casei AMBR2 | Nasal spray |
| Staphylococcus aureus, Moraxella catarrhalis, and Haemophilus influenzae | Secretion of antimicrobial substances such as lactic acid, bacteriocins, and H2O2 | n/a LiviaOne Probiotics Nasal Spray b | [183,184,302] |
| Ethyl lauroyl arginate hydrochloride (ELAH) | Nasal spray |
| Broad-spectrum antimicrobial activity, including against SARS-CoV-2 | Formation of positively charged physical barrier on mucosa and inhibition of viral adhesion | Covixyl-V | [300] |
| Silver nanoparticles | Nasal and oral rinse |
| Broad-spectrum antimicrobial activity, including against SARS-CoV-2 | Interference with the structural proteins of the virus and inhibiting their ability to bind with cell receptors, or bind to genetic material of viruses and inhibiting their replication | ViruStat-RDS nasal spray (+Xylitol), Silvacol nasal spray | [303,304] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, N.; Sotirova, Y.; Gavrailov, G.; Nikolova, K.; Andonova, V. Advances in the Prophylaxis of Respiratory Infections by the Nasal and the Oromucosal Route: Relevance to the Fight with the SARS-CoV-2 Pandemic. Pharmaceutics 2022, 14, 530. https://doi.org/10.3390/pharmaceutics14030530
Ivanova N, Sotirova Y, Gavrailov G, Nikolova K, Andonova V. Advances in the Prophylaxis of Respiratory Infections by the Nasal and the Oromucosal Route: Relevance to the Fight with the SARS-CoV-2 Pandemic. Pharmaceutics. 2022; 14(3):530. https://doi.org/10.3390/pharmaceutics14030530
Chicago/Turabian StyleIvanova, Nadezhda, Yoana Sotirova, Georgi Gavrailov, Krastena Nikolova, and Velichka Andonova. 2022. "Advances in the Prophylaxis of Respiratory Infections by the Nasal and the Oromucosal Route: Relevance to the Fight with the SARS-CoV-2 Pandemic" Pharmaceutics 14, no. 3: 530. https://doi.org/10.3390/pharmaceutics14030530