
Characterisation of New Foxunavirus Phage Murka with the Potential of Xanthomonas campestris pv. campestris Control

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Their Cultivation
2.1.1. Isolation of Strains
2.1.2. Checking Bacterial Pathogenicity
2.1.3. Genetic Identification of Xcc Strains
2.2. Isolation and Purification of Phage Murka
2.3. Electron Microscopy
2.4. Biological Characterisation of the Phage Murka
2.4.1. Determination of Phage Host Range on Xcc Strains
2.4.2. Phage Adsorption and One-Step Growth Experiments
2.4.3. Phage Stability in Different Conditions
2.5. Phage Genome Sequencing and Annotation
2.6. Genome and Proteome Analysis
2.7. Statistical Analysis
3. Results
3.1. Bacterial Strains
3.2. Isolation of Xcc Phage Murka
3.3. Biological Properties of Xcc Phage Murka
3.3.1. Morphology
3.3.2. Phage Production and Stability
3.4. General Genome and Proteome Features
3.5. Intergenomic Comparisons and Phylogenetic Analysis
3.5.1. Comparisons with Related Phages
3.5.2. VIRIDIC Analysis
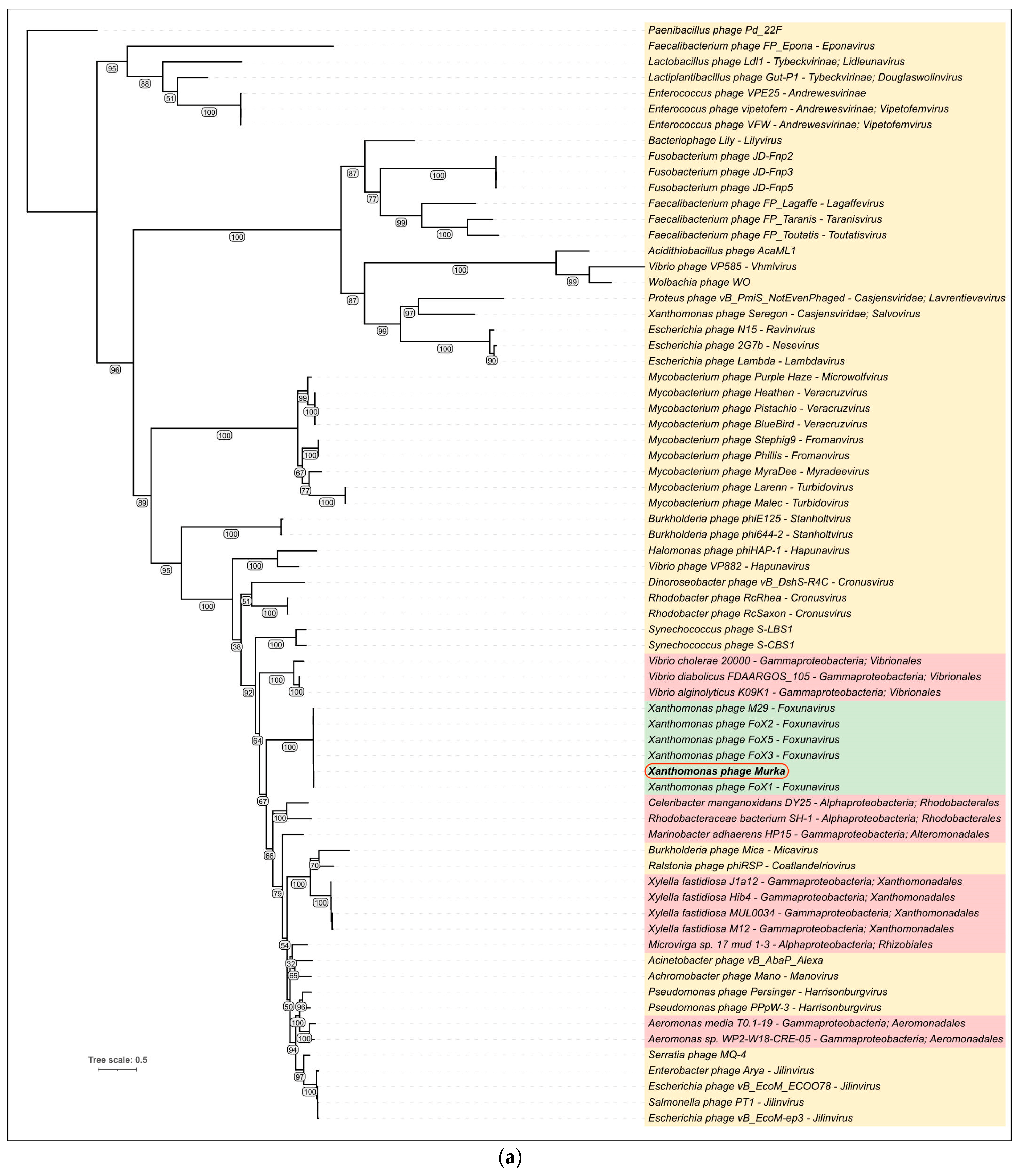
3.5.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Xcc | Xanthomonas campestris pv. campestris |
| cv. | Cultivar |
| CFU | Colony-forming unit |
| PFU | Plaque-forming unit |
| IKI | Solution of molecular iodine (I) and potassium iodide (KI) |
| MOI | Multiplicity of infection |
| OD | Optical density |
| AUGC | Area under growth curve |
References
- Lee, J.; Izzah, N.K.; Jayakodi, M.; Perumal, S.; Joh, H.J.; Lee, H.J.; Lee, S.-C.; Park, J.Y.; Yang, K.-W.; Nou, I.-S.; et al. Genome-Wide SNP Identification and QTL Mapping for Black Rot Resistance in Cabbage. BMC Plant Biol. 2015, 15, 32. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture—Statistical Yearbook 2021; FAO: Roma, Italy, 2021; ISBN 978-92-5-134332-6. [Google Scholar]
- Vicente, J.G.; Holub, E.B. Xanthomonas campestris pv. campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to brassica crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef]
- Cook, A.; Walker, J.; Larson, R. Studies on the disease cycle of black rot of crucifers. Phytopathology 1952, 42, 162. [Google Scholar]
- Williams, P.H. Black rot: A continuing threat to world crucifers. Plant Dis. 1980, 64, 736–742. [Google Scholar] [CrossRef]
- Sharma, B.B.; Kalia, P.; Singh, D.; Sharma, T.R. Introgression of Black Rot Resistance from Brassica carinata to Cauliflower (Brassica oleracea botrytis Group) through Embryo Rescue. Front. Plant Sci. 2017, 8, 1255. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.-H.; Huang, L.; Liu, G.-F.; Leng, M.; Lu, G.-T. PilG and PilH antagonistically control flagellum-dependent and pili-dependent motility in the phytopathogen Xanthomonas campestris pv. campestris. BMC Microbiol. 2020, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Rubel, M.H.; Robin, A.H.K.; Natarajan, S.; Vicente, J.G.; Kim, H.T.; Park, J.I.; Nou, I.S. Whole-genome re-alignment facilitates development of specific molecular markers for races 1 and 4 of Xanthomonas campestris pv. campestris, the cause of black rot disease in Brassica oleracea. Int. J. Mol. Sci. 2017, 18, 2523. [Google Scholar] [CrossRef] [PubMed]
- Tonu, N.N.; Doullah, M.A.-U.; Shimizu, M.; Karim, M.M.; Kawanabe, T.; Fujimoto, R.; Okazaki, K. Comparison of positions of QTLs conferring resistance to Xanthomonas campestris pv. campestris in Brassica oleracea. Am. J. Plant Physiol. 2013, 4, 11–20. [Google Scholar]
- Singh, S.; Dey, S.S.; Bhatia, R.; Batley, J.; Kumar, R. Molecular breeding for resistance to black rot [Xanthomonas campestris pv. campestris (Pammel) Dowson] in Brassicas: Recent advances. Euphytica 2018, 214, 196. [Google Scholar] [CrossRef]
- Cruz, J.; Tenreiro, R.; Cruz, L. Assessment of diversity of Xanthomonas campestris pathovars affecting cruciferous plants in Portugal and disclosure of two novel X. campestris pv. campestris races. J. Plant Pathol. 2017, 99, 403–414. [Google Scholar]
- Fargier, E.; Manceau, C. Pathogenicity assays restrict the species Xanthomonas campestris into three pathovars and reveal nine races within X. campestris pv. campestris. Plant Pathol. 2007, 56, 805–818. [Google Scholar] [CrossRef]
- Singh, D.; Rathaur, P.; Vicente, J. Characterization, genetic diversity and distribution of Xanthomonas campestris pv. campestris races causing black rot disease in cruciferous crops of India. Plant Pathol. 2016, 65, 1411–1418. [Google Scholar] [CrossRef]
- Eichmeier, A.; Peňázová, E.; Pokluda, R.; Vicente, J.G. Detection of Xanthomonas campestris pv. campestris through a real-time PCR assay targeting the Zur gene and comparison with detection targeting the hrpF. Eur. J. Plant Pathol. 2019, 155, 891–902. [Google Scholar] [CrossRef]
- Caracciolo, R.; Sella, L.; De Zotti, M.; Bolzonello, A.; Armellin, M.; Trainotti, L.; Favaron, F.; Tundo, S. Efficacy of Trichoderma longibrachiatum Trichogin GA IV Peptaibol Analogs against the Black Rot Pathogen Xanthomonas campestris pv. campestris and Other Phytopathogenic Bacteria. Microorganisms 2023, 11, 480. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Vikram, A.; Bharat, N. Black rot-A devastating disease of crucifers: A review. Agric. Rev. 2013, 34, 269. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, H.; Wang, J.; Lv, J.; Xie, B.; Luo, S.; Wang, S.; Zhang, B.; Li, Z.; Yue, Z.; et al. Physical, Chemical, and Biological Control of Black Rot of Brassicaceae Vegetables: A Review. Front. Microbiol. 2022, 13, 1023826. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Arora, N.K. Evaluation of rhizospheric Pseudomonas and Bacillus as biocontrol tool for Xanthomonas campestris pv campestris. World J. Microbiol. Biotechnol. 2012, 28, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.; Caproni, A.; Sicurella, M.; Manfredini, S.; Baldisserotto, A.; Marconi, P. Effects of Flavonoids and Phenols from Moringa oleifera Leaf Extracts on Biofilm Processes in Xanthomonas campestris pv. campestris. Plants 2023, 12, 1508. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, A.M.P.; Rodríguez, G.A.A.; Monteiro, F.P.; Faria, A.F.; Silva, J.C.P.; Monteiro, A.C.A.; Carvalho, C.V.; Gomes, L.A.A.; Souza, R.M.; de Souza, J.T.; et al. Bio-based products control black rot (Xanthomonas campestris pv. campestris) and increase the nutraceutical and antioxidant components in kale. Sci. Rep. 2018, 8, 10199. [Google Scholar] [CrossRef]
- Holtappels, D.; Fortuna, K.J.; Moons, L.; Broeckaert, N.; Bäcker, L.E.; Venneman, S.; Rombouts, S.; Lippens, L.; Baeyen, S.; Pollet, S.; et al. The Potential of Bacteriophages to Control Xanthomonas campestris pv. campestris at Different Stages of Disease Development. Microb. Biotechnol. 2022, 15, 1762–1782. [Google Scholar] [CrossRef] [PubMed]
- Orynbayev, A.T.; Dzhalilov, F.S.U.; Ignatov, A.N. Improved Efficacy of Formulated Bacteriophage in Control of Black Rot Caused by Xanthomonas campestris pv. campestris on Cabbage Seedlings. Arch. Phytopathol. Plant Prot. 2020, 53, 379–394. [Google Scholar] [CrossRef]
- Vu, N.T.; Oh, C.-S. Bacteriophage usage for bacterial disease management and diagnosis in plants. Plant Pathol. J. 2020, 36, 204. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the future with bacteriophages in agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E.; Sulakvelidze, A. (Eds.) Bacteriophages: Biology and Applications, 1st ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Harada, L.K.; Silva, E.C.; Campos, W.F.; Del Fiol, F.S.; Vila, M.; Dąbrowska, K.; Krylov, V.N.; Balcão, V.M. Biotechnological Applications of Bacteriophages: State of the Art. Microbiol. Res. 2018, 212–213, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Lelliot, R.A. Methods for the Diagnosis of Bacterial Diseases of Plants; Lellio, R.A., Stead, D.E., Eds.; Wiley-Blackwell: Oxford, UK, 1987; 216p. [Google Scholar]
- Rafi, A.; Hameed, A.; Akhtar, M.A.; Sohail, K.; Shahid, M.; Fahim, M. Identification and Characterization of Xanthomonas oryzae pv. oryzae in North-West of Pakistan. Sarhad J. Agric. 2013, 29, 10. [Google Scholar]
- Hakalova, E.; Kopta, T.; Jurica, M.; Pečenka, J.; Eichmeier, A.; Pokluda, R. Testing of Inoculation Methods and Susceptibility Testing of Perspective Cabbage Breeding Lines (Brassica Oleracea Convar. Capitata) to the Black Rot Disease Caused by Xanthomonas campestris pv. campestris. Acta Univ. Agric. Et Silvic. Mendel. Brun. 2018, 66, 139–148. [Google Scholar]
- Yinur, D.; Zinabu, D.; Mola, F.; Tadesse, B. Isolation and Identification of Enset Wilt Disease Causing Bacteria Using 16S RRNA Gene Sequence Samples Collected from Gurage Zone, Ethiopia. J. Life Sci. Biomed. 2020, 10, 51–58. [Google Scholar]
- Van Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil. Methods Mol. Biol. 2009, 501, 15–21. [Google Scholar]
- Adams, M. Bacteriophages; StatPearls Publishing: Treasure Island, FL, USA, 1959; p. 620. [Google Scholar]
- Adriaenssens, E.M.; Vaerenbergh, J.V.; Vandenheuvel, D.; Dunon, V.; Ceyssens, P.-J.; Proft, M.D.; Kropinski, A.M.; Noben, J.-P.; Maes, M.; Lavigne, R. T4-Related Bacteriophage LIMEstone Isolates for the Control of Soft Rot on Potato Caused by ‘Dickeya solani’. PLoS ONE 2012, 7, e33227. [Google Scholar] [CrossRef]
- Lukianova, A.A.; Shneider, M.M.; Evseev, P.V.; Shpirt, A.M.; Bugaeva, E.N.; Kabanova, A.P.; Obraztsova, E.A.; Miroshnikov, K.K.; Senchenkova, S.N.; Shashkov, A.S.; et al. Morphologically Different Pectobacterium brasiliense Bacteriophages PP99 and PP101: Deacetylation of O-Polysaccharide by the Tail Spike Protein of Phage PP99 Accompanies the Infection. Front. Microbiol. 2019, 10, 3147. [Google Scholar] [CrossRef]
- Esgalhado, M.E.; Roseiro, J.C.; Collaço, M.T.A. Interactive Effects of PH and Temperature on Cell Growth and Polymer Production by Xanthomonas campestris. Process Biochem. 1995, 30, 667–671. [Google Scholar] [CrossRef]
- Guo, M.; Gao, Y.; Xue, Y.; Liu, Y.; Zeng, X.; Cheng, Y.; Ma, J.; Wang, H.; Sun, J.; Wang, Z.; et al. Bacteriophage Cocktails Protect Dairy Cows Against Mastitis Caused By Drug Resistant Escherichia coli Infection. Front. Cell. Infect. Microbiol. 2021, 11, 690377. [Google Scholar] [CrossRef] [PubMed]
- Bouras, G.; Nepal, R.; Houtak, G.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Pharokka: A Fast Scalable Bacteriophage Annotation Tool. Bioinform. Oxf. Engl. 2023, 39, btac776. [Google Scholar] [CrossRef] [PubMed]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying Bacterial Genes and Endosymbiont DNA with Glimmer. Bioinform. Oxf. Engl. 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic Gene Recognition and Translation Initiation Site Identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at Its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, Snoscan and snoGPS Web Servers for the Detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canback, B. ARAGORN, a Program to Detect tRNA Genes and tmRNA Genes in Nucleotide Sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.-H. Clinker & Clustermap.Js: Automatic Generation of Gene Cluster Comparison Figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [PubMed]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- AlQuraishi, M. AlphaFold at CASP13. Bioinform. Oxf. Engl. 2019, 35, 4862–4865. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinform. Oxf. Engl. 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Bull, C.T.; Boer, S.H.D.; Denny, T.P.; Firrao, G.; Saux, M.F.-L.; Saddler, G.S.; Scortichini, M.; Stead, D.E.; Takikawa, Y. Comprehensive list of names of plant pathogenic bacteria, 1980–2007. J. Plant Pathol. 2010, 92, 551–592. [Google Scholar]
- Sharma, A.; Gautam, S.; Wadhawan, S. Xanthomonas . In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 811–817. ISBN 978-0-12-384733-1. [Google Scholar]
- Evseev, P.; Lukianova, A.; Sykilinda, N.; Gorshkova, A.; Bondar, A.; Shneider, M.; Kabilov, M.; Drucker, V.; Miroshnikov, K. Pseudomonas Phage MD8: Genetic Mosaicism and Challenges of Taxonomic Classification of Lambdoid Bacteriophages. Int. J. Mol. Sci. 2021, 22, 10350. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, H.; Jin, L.; Czornyj, E.; Hodes, A.; Hui, W.H.; Nieh, A.W.; Miller, J.F.; Zhou, Z.H. A New Topology of the HK97-like Fold Revealed in Bordetella Bacteriophage by cryoEM at 3.5 Å Resolution. eLife 2013, 2, e01299. [Google Scholar] [CrossRef]
- Evseev, P.; Shneider, M.; Miroshnikov, K. Evolution of Phage Tail Sheath Protein. Viruses 2022, 14, 1148. [Google Scholar] [CrossRef]
- Gonzales, M.F.; Piya, D.K.; Koehler, B.; Zhang, K.; Yu, Z.; Zeng, L.; Gill, J.J. New Insights into the Structure and Assembly of Bacteriophage P1. Viruses 2022, 14, 678. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A. Comparative Molecular Biology of Lambdoid Phages. Annu. Rev. Microbiol. 1994, 48, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Phage Lysis: Three Steps, Three Choices, One Outcome. J. Microbiol. Seoul Korea 2014, 52, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Yao, G.; Vizoso-Pinto, M.G.; Clark, J.; Le, T.; Gonzalez, C.; Gill, J.; Liu, M. Complete Genome Sequence of Burkholderia cenocepacia Phage Mica. Microbiol. Resour. Announc. 2021, 10, e01407-20. [Google Scholar] [CrossRef] [PubMed]
- Bartz, M.; Yao, G.; Le, T.; Liu, M.; Burrowes, B.; Gonzalez, C.; Young, R. Complete Genome Sequence of Achromobacter xylosoxidans Myophage Mano. Microbiol. Resour. Announc. 2021, 10, e01390-20. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Wang, S.; Yan, G.; Sun, C.; Feng, X.; Gu, J.; Han, W.; Lei, L. Genome Sequencing and Analysis of an Escherichia coli Phage vB_EcoM-Ep3 with a Novel Lysin, Lysep3. Virus Genes 2015, 50, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, G.; Liu, X.; Wang, J. Genomic Analysis and Biological Properties of the Novel Serratia liquefaciens Phage vB_SlqM_MQ-4. Arch. Virol. 2023, 168, 38. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.-Y.; Blanco, S.D.; Peng, A.-L.; Fu, J.-Y.; Chen, B.-W.; Luo, M.-C.; Xie, X.-Y.; Lin, Y.-H. Seed Treatment with Calcium Carbonate Containing Bacillus amyloliquefaciens PMB05 Powder Is an Efficient Way to Control Black Rot Disease of Cabbage. Agriculture 2023, 13, 926. [Google Scholar] [CrossRef]
- Hakalová, E.; Čechová, J.; Tekielska, D.A.; Eichmeier, A.; Pothier, J.F. Combined Effect of Thyme and Clove Phenolic Compounds on Xanthomonas campestris pv. campestris and Biocontrol of Black Rot Disease on Cabbage Seeds. Front. Microbiol. 2022, 13, 1007988. [Google Scholar] [CrossRef]
- Schmidtke, C.; Abendroth, U.; Brock, J.; Serrania, J.; Becker, A.; Bonas, U. Small RNA sX13: A Multifaceted Regulator of Virulence in the Plant Pathogen Xanthomonas. PLoS Pathog. 2013, 9, e1003626. [Google Scholar] [CrossRef]
- Villalpando-Aguilar, J.L.; Matos-Pech, G.; López-Rosas, I.; Castelán-Sánchez, H.G.; Alatorre-Cobos, F. Phage Therapy for Crops: Concepts, Experimental and Bioinformatics Approaches to Direct Its Application. Int. J. Mol. Sci. 2023, 24, 325. [Google Scholar] [CrossRef]
- Korniienko, N.; Kharina, A.; Budzanivska, I.; Burketová, L.; Kalachova, T. Phages of phytopathogenic bacteria: High potential, but challenging application. Plant Protect. Sci. 2022, 58, 81–91. [Google Scholar] [CrossRef]
- Jain, L.; Kumar, V.; Jain, S.K.; Kaushal, P.; Ghosh, P.K. Isolation of Bacteriophages Infecting Xanthomonas oryzae pv. oryzae and Genomic Characterization of Novel Phage vB_XooS_NR08 for Biocontrol of Bacterial Leaf Blight of Rice. Front. Microbiol. 2023, 14, 1084025. [Google Scholar] [CrossRef]
- Teklemariam, A.D.; Al Hindi, R.; Qadri, I.; Alharbi, M.G.; Hashem, A.M.; Alrefaei, A.A.; Basamad, N.A.; Haque, S.; Alamri, T.; Harakeh, S. Phage Cocktails—An Emerging Approach for the Control of Bacterial Infection with Major Emphasis on Foodborne Pathogens. Biotechnol. Genet. Eng. Rev. 2023, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Zhang, G.; Xie, G.; Zhu, J.; Lou, M.; Zhou, X.; Zhang, X.; Sun, G.; Li, B.; Zhu, B. Evolution of Xanthomonas Gene Content: Gene Gain/Loss History and Species Divergence. J. Integr. Agric. 2012, 11, 954–961. [Google Scholar] [CrossRef]
- Nakayinga, R.; Makumi, A.; Tumuhaise, V.; Tinzaara, W. Xanthomonas Bacteriophages: A Review of Their Biology and Biocontrol Applications in Agriculture. BMC Microbiol. 2021, 21, 291. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.D.; Capage, M.A.; Kessel, M.; Benson, S.A. Isolation and Characterization of a Generalized Transducing Phage for Xanthomonas campestris pv. campestris. J. Bacteriol. 1994, 176, 3354–3359. [Google Scholar] [CrossRef] [PubMed]
- da Silva, F.P.; Xavier, A.d.S.; Bruckner, F.P.; de Rezende, R.R.; Vidigal, P.M.P.; Alfenas-Zerbini, P. Biological and molecular characterization of a bacteriophage infecting Xanthomonas campestris pv. campestris, isolated from brassica fields. Arch. Virol. 2019, 164, 1857–1862. [Google Scholar] [CrossRef] [PubMed]
- Papaianni, M.; Paris, D.; Woo, S.L.; Fulgione, A.; Rigano, M.M.; Parrilli, E.; Tutino, M.L.; Marra, R.; Manganiello, G.; Casillo, A.; et al. Plant Dynamic Metabolic Response to Bacteriophage Treatment After Xanthomonas campestris pv. campestris Infection. Front. Microbiol. 2020, 11, 732. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Name of the Strain | Date of Isolation | Place of Origin, Plant | Genbank 16S № | Lysis Zone during Interaction with the Phage Murka |
|---|---|---|---|---|---|
| 1 | BK-55 | 10.2017 | Krasnodar region, Russia, white cabbage | OR626094 | + |
| 2 | CK-71 | 10.2017 | Krasnodar region, Russia, cauliflower | OR626097 | + |
| 3 | Xcc 1/1 | 09.2017 | Moscow region, Dmitrov district, Russia, white cabbage | OR626648 | + |
| 4 | Bes-1 | 09.2016 | Moscow region, Dmitrov district, Russia, white cabbage | OR626092 | + |
| 5 | Cas | 09.2016 | Moscow region, Dmitrov district, Russia, cauliflower | OR626095 | + |
| 6 | Tr1 | 11.2012 | Tiraspol, Moldova, cabbage | OR626099 | + |
| 7 | DK-1 | 10.2012 | Moscow region, Serpukhov district, Russia, white cabbage | OR626096 | + |
| 8 | Ram 3-1 | 10.2012 | Moscow region, Ramensky district, Russia, cabbage | OR625211 | + |
| 9 | XУ 1-2 | 10.2012 | Ukraine, white cabbage | OR644606 | - |
| 10 | Bel-2 | 10.2006 | Belarus, white cabbage | OR626091 | - |
| 11 | Bun-1 | 09.2006 | Moscow region, Dmitrov district, Russia, white cabbage | OR626093 | - |
| 12 | Xn-13 | 1997 | Japan, Ano, Capsélla búrsa-pastóris (shepherd’s purse) | OR626098 | + |
| 13 | 306NZ | - | Netherlands | OR626090 | - |
| 14 | NCPPB 528T | 1957 | UK, cabbage | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evseev, P.V.; Tarakanov, R.I.; Vo, H.T.N.; Suzina, N.E.; Vasilyeva, A.A.; Ignatov, A.N.; Miroshnikov, K.A.; Dzhalilov, F.S.-U. Characterisation of New Foxunavirus Phage Murka with the Potential of Xanthomonas campestris pv. campestris Control. Viruses 2024, 16, 198. https://doi.org/10.3390/v16020198
Evseev PV, Tarakanov RI, Vo HTN, Suzina NE, Vasilyeva AA, Ignatov AN, Miroshnikov KA, Dzhalilov FS-U. Characterisation of New Foxunavirus Phage Murka with the Potential of Xanthomonas campestris pv. campestris Control. Viruses. 2024; 16(2):198. https://doi.org/10.3390/v16020198
Chicago/Turabian StyleEvseev, Peter V., Rashit I. Tarakanov, Ha T. N. Vo, Natalia E. Suzina, Anna A. Vasilyeva, Alexander N. Ignatov, Konstantin A. Miroshnikov, and Fevzi S.-U. Dzhalilov. 2024. "Characterisation of New Foxunavirus Phage Murka with the Potential of Xanthomonas campestris pv. campestris Control" Viruses 16, no. 2: 198. https://doi.org/10.3390/v16020198