Comparative Analysis of Complete Chloroplast Genome Sequences and Insertion-Deletion (Indel) Polymorphisms to Distinguish Five Vaccinium Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Comparison of Cp Genomes and Identification of InDel Loci
2.2. Development and Validation of the Candidate DNA Barcodes
2.3. Phylogenetic Analysis
3. Results
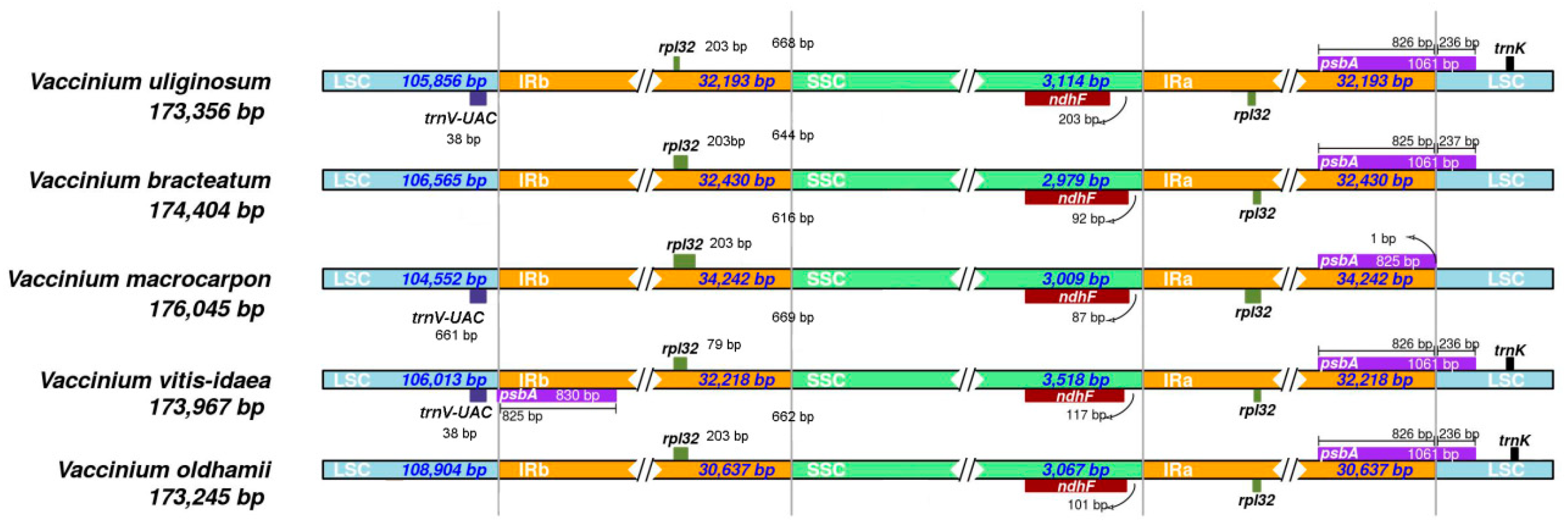
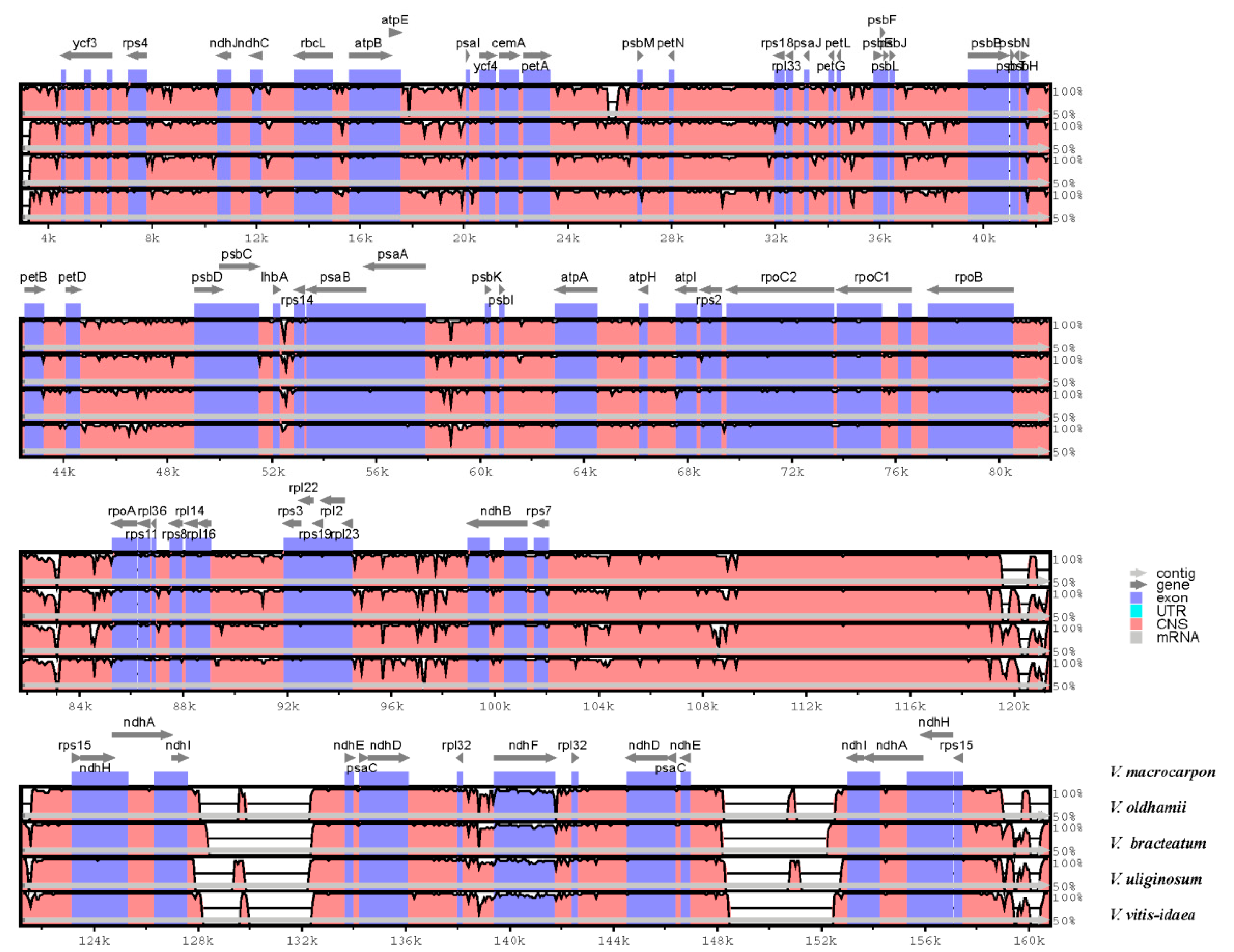
3.1. Comparative Analyses of the Chloroplast Genome of Five Vaccinium Species
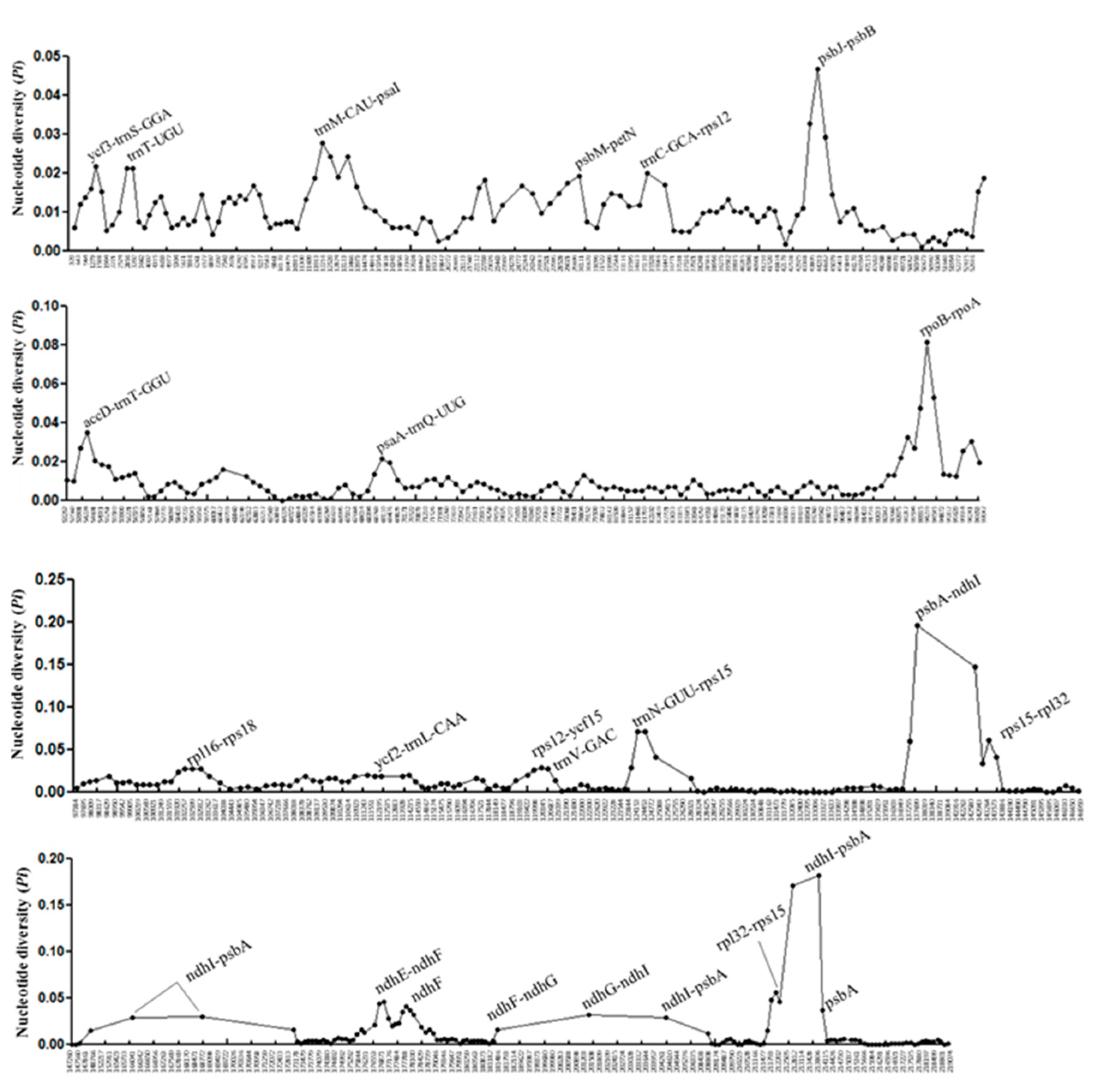
3.2. Divergence Hotspots of Five Vaccinium Cp Genomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luby, J.J.; Ballington, J.R.; Draper, A.D.; Pliszka, K.; Austin, M.E. Blueberries and cranberries (Vaccinium). Genet. Resour. Temp. Fruit Nut Crops 1991, 290, 393–458. [Google Scholar] [CrossRef]
- Ballington, J.R. Collection, utilization, and preservation of genetic resources in Vaccinium. HortScience 2001, 36, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Abreu, O.A.; Barreto, G.; Prieto, S. Vaccinium (Ericaceae): Ethnobotany and pharmacological potentials. Emir. J. Food Agric. 2014, 26, 577. [Google Scholar] [CrossRef]
- Strik, B.C. A review of optimal systems for organic production of blueberry and blackberry for fresh and processed markets in the northwestern United States. Sci. Horticult. 2016, 208, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Primetta, A.K.; Jaakola, L.; Ayaz, F.A.; Inceer, H.; Riihinen, K.R. Anthocyanin fingerprinting for authenticity studies of bilberry (Vaccinium myrtillus L.). Food Control 2013, 30, 662–667. [Google Scholar] [CrossRef]
- Zhu, H.; Senalik, D.; McCown, B.H.; Zeldin, E.L.; Speers, J.; Hyman, J.; Zalapa, J.E. Mining and validation of pyrosequenced simple sequence repeats (SSRs) from American cranberry (Vaccinium macrocarpon Ait.). Theor. Appl. Genet. 2012, 124, 87–96. [Google Scholar] [CrossRef]
- Jothi, G.A.; Sircom, J. Native pollinators alone provide full pollination on small-scale commercial cranberry (Ericaceae) farms. Can. Entomol. 2019, 151, 745–756. [Google Scholar] [CrossRef]
- Jurikova, T.; Skrovankova, S.; Mlcek, J.; Balla, S.; Snopek, L. Bioactive compounds, antioxidant activity, and biological effects of European cranberry (Vaccinium oxycoccos). Molecules 2019, 24, 24. [Google Scholar] [CrossRef] [Green Version]
- Gustavsson, B.A. Genetic variation in horticulturally important traits of fifteen wild lingonberry Vaccinium vitis-idaea L. populations. Euphytica 2001, 120, 173–182. [Google Scholar] [CrossRef]
- Hokkanen, J.; Mattila, S.; Jaakola, L.; Pirttilä, A.M.; Tolonen, A. Identification of phenolic compounds from lingonberry (Vaccinium vitis-idaea L.), bilberry (Vaccinium myrtillus L.) and hybrid bilberry (Vaccinium x intermedium Ruthe L.) leaves. J. Agric. Food Chem. 2009, 57, 9437–9447. [Google Scholar] [CrossRef]
- Lee, T.B. Coloured Flora of Korea; Hyangmunsa: Seoul, Korea, 2003; Volume 2, pp. 27–29. (In Korean) [Google Scholar]
- Kim, C.Y.; Kim, Y.H.; Han, S.H.; Ko, H.C. Current Situations and Prospects on the Cultivation Program of Tropical and Subtropical Crops in Korea. Korean J. Plant Res. 2019, 32, 45–52. [Google Scholar]
- Sherman, B.; Henry, R.J. The Nagoya Protocol and historical collections of plants. Nat. Plants 2020, 6, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.L.; Kang, J.H.; Kim, H.M.; Jeong, S.H.; Jang, D.S.; Jang, Y.P.; Choung, S.Y. Polyphenol-enriched Vaccinium uliginosum L. fractions reduce retinal damage induced by blue light in A2E-laden ARPE19 cell cultures and mice. Nutr. Res. 2016, 36, 1402–1414. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.T.; Jeong, S.Y.; Velmurugan, P.; Park, J.H.; Jeong, D.Y. Probiotic-mediated blueberry (Vaccinium corymbosum L.) fruit fermentation to yield functionalized products for augmented antibacterial and antioxidant activity. J. Biosci. Bioeng. 2017, 124, 542–550. [Google Scholar] [CrossRef]
- Oh, D.R.; Yoo, J.S.; Kim, Y.; Kang, H.; Lee, H.; Lm, S.J.; Hong, J. Vaccinium bracteatum leaf extract reverses chronic restraint stress-induced depression-like behavior in mice: Regulation of hypothalamic-pituitary-adrenal axis, serotonin turnover systems, and ERK/Akt phosphorylation. Front. Pharmacol. 2018, 9, 604. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biolog. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Ravi, V.; Khurana, J.P.; Tyagi, A.K.; Khurana, P. An update on chloroplast genomes. Plant Syst. Evol. 2008, 271, 101–122. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Mileshina, D.; Wallet, C.; Niazi, A.K.; Weber-Lotfi, F.; Dietrich, A. The plant mitochondrial genome: Dynamics and maintenance. Biochimie 2014, 100, 107–120. [Google Scholar] [CrossRef]
- Bi, Y.; Zhang, M.F.; Xue, J.; Dong, R.; Du, Y.P.; Zhang, X.H. Chloroplast genomic resources for phylogeny and DNA barcoding: A case study on Fritillaria. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhou, J.; Cui, Y.; Wang, Y.; Duan, B.; Yao, H. Identification of Ligularia herbs using the complete chloroplast genome as a super-barcode. Front. Pharmacol. 2018, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35 (Suppl. 2), W71–W74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Shin, J.; Oh, D.R.; Kim, D.W.; Lee, H.S.; Choi, C. Complete chloroplast genome sequences of Vaccinium bracteatum Thunb. V. vitis-idaea L. and V. uliginosum L. (Ericaceae). Mitochondrial DNA B 2020, 5, 1843–1844. [Google Scholar]
- Knudsen, B.; Knudsen, T.; Flensborg, M.; Sandmann, H.; Heltzen, M.; Andersen, A.; Mønsted, S. Bio-Qiagen, C.L.C.; CLC Sequence Viewer: Aarhus, Denmark, 2011; Volume 6. [Google Scholar]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2007, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Calie, P.J. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Nock, C.J.; Waters, D.L.; Edwards, M.A.; Bowen, S.G.; Rice, N.; Cordeiro, G.M.; Henry, R.J. Chloroplast genome sequences from total DNA for plant identification. Plant Biotechnol. J. 2011, 9, 328–333. [Google Scholar] [CrossRef]
- Kron, K.A.; Powell, E.A.; Luteyn, J.L. Phylogenetic relationships within the blueberry tribe (Vaccinieae, Ericaceae) based on sequence data from matK and nuclear ribosomal ITS regions, with comments on the placement of Satyria. Am. J. Bot. 2002, 89, 327–336. [Google Scholar] [CrossRef]
- Sultana, N.; Pascual-Díaz, J.P.; Gers, A.; Ilga, K.; Serçe, S.; Vitales, D.; Garcia, S. Contribution to the knowledge of genome size evolution in edible blueberries (genus Vaccinium). J. Berry Res. 2019, 10, 243–257. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Scientific Name (n) | Common Name | Collection Site | Specimen Code |
|---|---|---|---|---|
| 1 | Vaccinium bracteatum Thunb. (n = 3) | Sea blueberry | 33°31′03.5″ N 126°43′00.1″ E | NIBRGR0000424873 |
| 2 | 34°22′17.0″ N 126°17′11.5″ E | NIBRGR0000424891 | ||
| 3 | 34°20′55.1″ N 126°41′30.5″ E | NIBRGR0000067902 | ||
| 4 | Vaccinium vitis-iaea L. (n = 3) | Cowberry | 38°10′14.5″ N 128°27′39.3″ E | JINR000002321 |
| 5 | 38°10′14.5″ N 128°27′39.3″ E | JINR000002322 | ||
| 6 | 38°10′14.5″ N 128°27′39.3″ E | JINR000002323 | ||
| 7 | Vaccinium uliginosum L. (n = 3) | Bog bilberry, Moonberry | 33°19′37.6″ N 126°34′09.3″ E | NIBRGR0000424897 |
| 8 | 38°10′14.5″ N 128°27′39.3″ E | JINR000002324 | ||
| 9 | 38°10′14.5″ N 128°27′39.3″ E | JINR000002325 | ||
| 10 | Vaccinium macrocarpon Ait. (n = 3) | Cranberry | 34°40′54.3″ N 126°54′38.1″ E | JINR000002326 |
| 11 | 34°40′54.3″ N 126°54′38.1″ E | JINR000002327 | ||
| 12 | 34°40′54.3″ N 126°54′38.1″ E | JINR000002328 | ||
| 13 | Vaccinium oldhamii Miq. (n = 7) | Oldham blueberry | 36°20′45.2″N 128°01′54.5″ E | NIBRGR0000424923 |
| 14 | 35°28′18.7″N 126°39′10.6″ E | NIBRGR0000424929 | ||
| 15 | 33°24′47.3″ N 126°24′41.9″ E | NIBRGR0000424965 | ||
| 16 | 36°33′33.7″ N 126°20′20.6″ E | NIBRGR0000424969 | ||
| 17 | 34°45′16.8″ N 127°58′43.8″ E | NIBRGR0000424971 | ||
| 18 | 34°22′36.6″ N 126°17′26.6″ E | NIBRGR0000424895 | ||
| 19 | 34°48′49.7′ N 126°20′10.2″ E | NIBRGR0000374901 |
| Name of Taxon | V. bracteatum | V. uliginosum | V. vitis-idaea | V. macrocarpon | V. oldhamii |
|---|---|---|---|---|---|
| Accession number | LC521967 | LC521968 | LC521969 | NC_019616 | NC_042713 |
| Genome length | 174,404 | 173,356 | 173,967 | 176,045 | 173,245 |
| LSC length | 106,565 | 105,856 | 106,013 | 104,552 | 108,904 |
| SSC length | 2979 | 3114 | 3518 | 3009 | 3067 |
| IR length | 32,430 | 32,193 | 32,218 | 34,242 | 30,637 |
| Total gene number | 117 | 125 | 125 | 147 | 130 |
| No. of protein coding genes | 79 | 79 | 79 | 75 | 85 |
| No. of tRNA genes | 30 | 38 | 38 | 36 | 37 |
| No. of rRNA genes | 8 | 8 | 8 | 8 | 8 |
| GC content in genome (%) | 36.8 | 36.8 | 36.7 | 37.1 | 37.2 |
| Gene Category | Gene Group | Gene Names | Vb | Vu | Vv | Vm | Vo |
|---|---|---|---|---|---|---|---|
| Self-replication | RNA polymerase | rpoA | - | - | - | ● a | ● |
| rpoB | ● | ● | ● | ● | ● | ||
| rpoC1 * | ● | ● | ● | ● | ● | ||
| rpoC2 | ● | ● | ● | ● | ● | ||
| rRNA genes | rrn16a | ● | ● | ● | ● | ● | |
| rrn23a | ● | ● | ● | ● | ● | ||
| rrn4.5a | ● | ● | ● | ● | ● | ||
| rrn5a | ● | ● | ● | ● | ● | ||
| tRNA genes | trnA-UGCa,* | ● | ● | ● | ● | ● | |
| trnC-GCA | ● | ● | ● | ● | ● | ||
| trnD-GUC | - | ● | ● | ● | ● | ||
| trnE-UUC | ● | ● | ● | ● | ● | ||
| trnfM-CAU | - | ● | ● | ● a | ● a | ||
| trnG-GCC | ● | ● | ● | - | - | ||
| trnG-UCC | ● * | ● * | ● * | ● | ●* | ||
| trnH-GUGa | ● | ● | ● | ● | ● | ||
| trnI-CAU | ● | ● | ● | ● | ● | ||
| trnI-GAU * | ● | ● a | ● a | ● a | ● a | ||
| trnK-UUU | - | ● * | ● * | - | ● * | ||
| trnL-CAA | ● | ● | ● | ● | ● | ||
| trnL-UAGa | ● | ● | ● | ● | ● | ||
| trnM-CAU | ● | ● | ● | ● | ● | ||
| trnN-GUUa | ● | ● | ● | ● | ● | ||
| trnP-GGG | - | - | - | ● | - | ||
| trnQ-UUG | ● | ● | ● | ● | ● | ||
| trnR-ACGa | ● | ● | ● | ● | ● | ||
| trnR-UCU | - | ● | ● | ● | ● | ||
| trnS-GCU | ● | ● | ● | ● | ● | ||
| trnS-GGA | ● | ● | ● | ● | ● | ||
| trnS-UGA | ● | ● | ● | ● | ● | ||
| trnT-UGU | ● | ● | ● | ● | ● | ||
| trnT-GGU | ● | ● | ● | ● | ● | ||
| trnV-GAC | ● | ● | ● | ● | ● | ||
| trnV-UAC | - | ● * | ●* | ● * | ●* | ||
| trnW-CCA | ● | ● | ● | ● | ● | ||
| trnY-GUA | - | ● | ● | ● | ● | ||
| Small subunit of ribosome | rps2 | ● | ● | ● | ● | ● | |
| rps3 | ● * | ● * | ● * | ● a | ● | ||
| rps4 | ● | ● | ● | ● | ● | ||
| rps7 | - | - | - | ● | ● | ||
| rps8 | ● | ● | ● | ● | ● | ||
| rps11 | ● | ● | ● | ● | ● | ||
| rps12 | - | - | - | ●a | ●a | ||
| rps14 | ● | ● | ● | ● | ● | ||
| rps15a | ● | ● | ● | ● | ● | ||
| rps16a | ● * | ● * | ● * | - | ● * | ||
| rps18 | ● | ● | - | ● a | ● | ||
| rps19 | ● | ● | ● | ● | ● | ||
| Large subunit of ribosome | rpl2 | - | ● | ● | ● | ● | |
| rpl14 | ● | ● | ● | ● | ● | ||
| rpl16 | - | - | - | ● | ● | ||
| rpl20 | ● | ● | ● | - | ● | ||
| rpl22 | ● | ● | ● | ● a | ● | ||
| rpl23 | ● | ● | ● | ● | ● | ||
| rpl32a | ● | ● | ● | ● | ● | ||
| rpl33 | ● | ● | ● | ● | ● | ||
| rpl36 | ● | ● | ● | ● | ● | ||
| Photosynthesis | Photosystem I | psaA | ● | ● | ● | ● | ● |
| psaB | ● | ● | ● | ● | ● | ||
| psaCa | ● | ● | ● | ● | ● | ||
| psaI | ● | ● | ● | ● | ● | ||
| psaJ | ● | ● | ● | ● | ● | ||
| ycf3 ** | ● | ● | ● | ● | ● | ||
| Photosystem II | lhbAa | - | - | - | ● | - | |
| psbA | - | - | ● a | ● * | - | ||
| psbB | ● | ● | ● | ● | ● | ||
| psbC | ● | ● | ● | ● | ● | ||
| psbD | ● | ● | ● | ● | ● | ||
| psbE | ● | ● | ● | ● | ● | ||
| psbF | ● | ● | ● | ● | ● | ||
| psbH | ● | ● | ● | ● | ● | ||
| psbI | ● | ● | ● | ● | ● | ||
| psbJ | ● | ● | ● | ● | ● | ||
| psbK | ● | ● | ● | ● | ● | ||
| psbL | ● | ● | ● | ● | ● | ||
| psbM | ● | ● | ● | ● | ● | ||
| psbN | ● | ● | ● | ● | ● | ||
| psbT | ● | ● | ● | ● | ● | ||
| psbZ | ● | ● | ● | - | ● | ||
| ATP synthase | atpA | ● | ● | ● | ● | ● | |
| atpB | ● | ● | ● | ● | ● | ||
| atpE | ● | ● | ● | ● | ● | ||
| atpF * | ● | ● | ● | ● | ● | ||
| atpH | ● | ● | ● | ● | ● | ||
| atpI | ● | ● | ● | ● | ● | ||
| NADH dehydrogenase | ndhAa,* | ● | ● | ● | ● | ● | |
| ndhB * | ● | ● | ● | ● | ● | ||
| ndhC | ● | ● | ● | ● | ● | ||
| ndhDa | ● | ● | ● | ● | ● | ||
| ndhEa | ● | ● | ● | ● | ● | ||
| ndhF | ● | ● | ● | ● | ● | ||
| ndhGa | ● | ● | ● | ● | ● | ||
| ndhHa | ● | ● | ● | ● | ● | ||
| ndhIa | ● | ● | ● | ● | ● | ||
| ndhK | ● | ● | ● | ● | ● | ||
| ndhJ | ● | ● | ● | ● | ● | ||
| Cytochrome b/f complex | petA | ● | ● | ● | ● | ● | |
| petB | - | - | - | ● | ● * | ||
| petD | - | - | - | ● * | ● * | ||
| petG | ● | ● | ● | ● | ● | ||
| petL | ● | ● | ● | ● | ● | ||
| petN | ● | ● | ● | ● | ● | ||
| rbcL | ● | ● | ● | ● | ● | ||
| Rubisco | accD | ● | ● | ● | ● | ● | |
| Other genes | Subunit of acetyl-CoA-carboxylase | ccsA | ● | ● | ● | ● | ● |
| C-type cytochrome synthesis gene | infA | - | - | - | ● | - | |
| Translational initiation factor | cemA | ● | ● | ● | ● | ● | |
| Envelop membrane protein | matK | ● | ● | ● | ● | ● | |
| Gene of unknown function | Open reading frame | ycf4 | ● | ● | ● | ● | ● |
| Putative pseudogenes | ycf2 | - | - | - | ● | - | |
| ycf15b | - | - | - | ● | - | ||
| ycf68 | - | - | - | ● a | - |
| No. | Locus | Forward Primer (Sequence 5′ to 3′) | Reverse Primer (Sequence 5′ to 3′) | Product Range (bp) | AS | No. of InDels | Mean Pairwise Distance |
|---|---|---|---|---|---|---|---|
| 1 | trnT-UGU | CAGTAATCTTTGCAAAAGGAAAAAC | TTCGTCGTAACTTACACCTTTATGA | 617–653 | 40 | 4 | 0.01683 |
| 2 | psbM-petN | ATGAGAGCTTCTTCGAATAATTTTG | CATTTTCTCTTTCACTCGTAGTATGG | 650–665 | 60 | 2 | 0.01467 |
| 3 | trnC-GCA-rps12 | AATTCGATTGAATAAAATGGAGGA | GGAAATTGCCAACGTCAA | 611–630 | 80 | 3 | 0.02496 |
| 4 | accD-trnT-GGU | GGATCTAAATTAGGCCTCGTGTG | ATGATAGAGTCGACTTGACAATGC | 630–639 | 100 | 8 | 0.02504 |
| 5 | psaA-trnQ-UUG | ATCCCCCGGTATCTTATCTACATT | TTGCTGAATATCAAGTCAAACAGAA | 682–718 | 60 | 6 | 0.01364 |
| 6 | rpoB-rpoA (1) | AAAAAGCCAATTACAAGCCAAATA | ATCCAACGGAAATGACATTCTTAT | 603–706 | 20 | 5 | 0.02594 |
| 7 | rpoB-rpoA (2) | GCACTGAGATCTGCCACTTTATT | GTCATCGACGAGATTTTTGTAGC | 552–659 | 100 | 14 | 0.05182 |
| 8 | rpoB-rpoA (3) | CTTTCTTCGCTTTGATCCTCATA | TCCCCTCTTATGTATGTTTTTGC | 725–770 | 100 | 7 | 0.02646 |
| 9 | ycf2-trnL-CAA | ATTCTTTCGACTCATTTTCCTGAC | CTAGGAGCCAAAACTATGTGATTG | 643–924 | 100 | 11 | 0.01768 |
| 10 | rps12-ycf15 | CTTACACTCGGTCCCAAAGAAC | CTTTTCTCATGGGACAATGCTCT | 698–709 | 100 | 6 | 0.01991 |
| 11 | trnV-GAC | GAGCTCTTAAATGGAAATGGAAAA | GCCATTGTATAACCATTCATCAAC | 625-647 | 100 | 11 | 0.03506 |
| 12 | ndhE-ndhF | AATTCTATGAGGCACTGTTTCGAT | GAAGATTTTTCGTTGCTCTTGG | 587–643 | 100 | 7 | 0.04406 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Shin, J.; Oh, D.-R.; Kim, A.-Y.; Choi, C. Comparative Analysis of Complete Chloroplast Genome Sequences and Insertion-Deletion (Indel) Polymorphisms to Distinguish Five Vaccinium Species. Forests 2020, 11, 927. https://doi.org/10.3390/f11090927
Kim Y, Shin J, Oh D-R, Kim A-Y, Choi C. Comparative Analysis of Complete Chloroplast Genome Sequences and Insertion-Deletion (Indel) Polymorphisms to Distinguish Five Vaccinium Species. Forests. 2020; 11(9):927. https://doi.org/10.3390/f11090927
Chicago/Turabian StyleKim, Yonguk, Jawon Shin, Dool-Ri Oh, Ah-Young Kim, and Chulyung Choi. 2020. "Comparative Analysis of Complete Chloroplast Genome Sequences and Insertion-Deletion (Indel) Polymorphisms to Distinguish Five Vaccinium Species" Forests 11, no. 9: 927. https://doi.org/10.3390/f11090927