
Living Algal Community Voucher and Taxonomy in Dickinson County Iowa: A Case for Collection-Based Research

1
Department of Biological and Environmental Sciences, Georgia College & State University, Milledgeville, GA 31061, USA
2
Institute of Environment and Department of Biological Sciences, Florida International University, Miami, FL 33199, USA
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(1), 21; https://doi.org/10.3390/d16010021
Submission received: 26 November 2023
/
Revised: 16 December 2023
/
Accepted: 22 December 2023
/
Published: 27 December 2023
(This article belongs to the Section Freshwater Biodiversity)
Abstract
:Integrative taxonomy is necessary for advancing the field of phycology and understanding regional algal diversity. Collection-based studies for the purposes of providing descriptive voucher flora of living algal communities inform future long-term monitoring and genetic barcoding studies. In this study, littoral composite samples were taken from eight sites (Center Lake, Emerson Bay, Freda Haffner Kettlehole State Preserve, Millers Bay, East Okoboji Lake, West Okoboji Lake, Big Spirit Lake, Upper Gar Lake) in Dickinson County, Iowa, for the purposes of fine level taxonomy. This region has a history of eutrophication, and algal blooms have caused Iowa water resource issues in the past. Micrography for these voucher floras was conducted for taxa representing at least 10% or more of the total estimated algal biomass of evaluated slides. Our voucher floras comprise micrographs of living cells across nine different algal groups and 54 unique taxonomic entities. Unique taxa from these sites include Glaucocystis nostochinearum Itzigsohn, Draparnaldia acuta (C.Agardh) Kützing, and Chaetophoropsis elegans (Roth) B. Wen Liu, Qian Xiong, X.Dong Liu, Z. Yu Hu & G. Xiang Liu. We also document species belonging to toxin-producing Cyanobacteria genera Mycrocystis and Dolichospermum.
1. Introduction
Documenting algal biodiversity for water quality studies has long been important for ecologists and conservationists. Understanding the communities of aquatic biota aids in policies aimed at providing potable water to surrounding communities. However, over the years, there have been taxonomic challenges and disagreements on how species are interpreted and documented [1,2,3]. To overcome these obstacles, there has been a technological trend of using molecular work or genomic information to document algal species. Although this technique has proven to capture cryptic species [4,5], information about the species’ living environment, cell physiologies, and aspects of natural history is lost. Another limitation to currently using genomic information to document freshwater algal diversity is the lack of published freshwater genome sequences compared to the number of algal species [6]. An emerging solution is providing voucher flora or micrographs to standardize species identifications [7,8,9,10,11,12]. These vouchers provide consistency in taxonomic identification for long-term studies and help avoid misrepresentation, which is possible when only lists or tables of species names are provided. Many of these published vouchers contain micrographs of non-living cells and are often of one algal group (diatoms) as their silica cell wall preserves them well. In fact, few publications aimed at assessing aquatic communities document diatoms and other algal groups [13]. Collecting freshwater algal taxa from other groups for the purposes of providing descriptions and voucher flora is not without its own impediments: funding and support. In “The Silent Extinction of Species and Taxonomists—An Appeal to Science Policymakers and Legislators,” the authors describe this issue and the paradox it creates: there is a lack of support and funding for taxonomic baseline research and collection-based approaches when it is needed most [14]. This issue has been ongoing for decades [15].
In this collection-based study, we use composite samples from eight sites in Dickinson County, Iowa (Center Lake, Emerson Bay, Freda Haffner Kettlehole State Preserve, Millers Bay, East Okoboji Lake, West Okoboji Lake, Big Spirit Lake, Upper Gar Lake) for the purposes of fine level taxonomy. With voucher flora, we document algal communities, describe algal species, and record information about these species’ living environments. Our voucher flora provides micrographs of living cells across different algal groups. We do not make assumptions about these sites as a whole, nor do we claim to document all algal species in these communities. We chose these sites for a variety of reasons: (1) Iowa waters have a history of eutrophication and documented aquatic community changes [16,17,18,19,20], (2) algal blooms have caused issues with surface water resources in Iowa in the past [21,22,23], (3) with the exception of paleo research and Ambient Lake Monitoring program through Iowa State University, species-specific ecology and distribution records from Iowa and the region remain sparse [24,25,26,27], and (4) students from Iowa Lakeside Laboratory visit these sites with their summer courses on diatom and algal ecology and systematics most years providing an avenue for training taxonomists for future long-term studies. The goal of this study is to provide descriptive voucher flora to inform future long-term studies and provide complementary information to molecular studies documenting diversity at these sites.
2. Materials and Methods
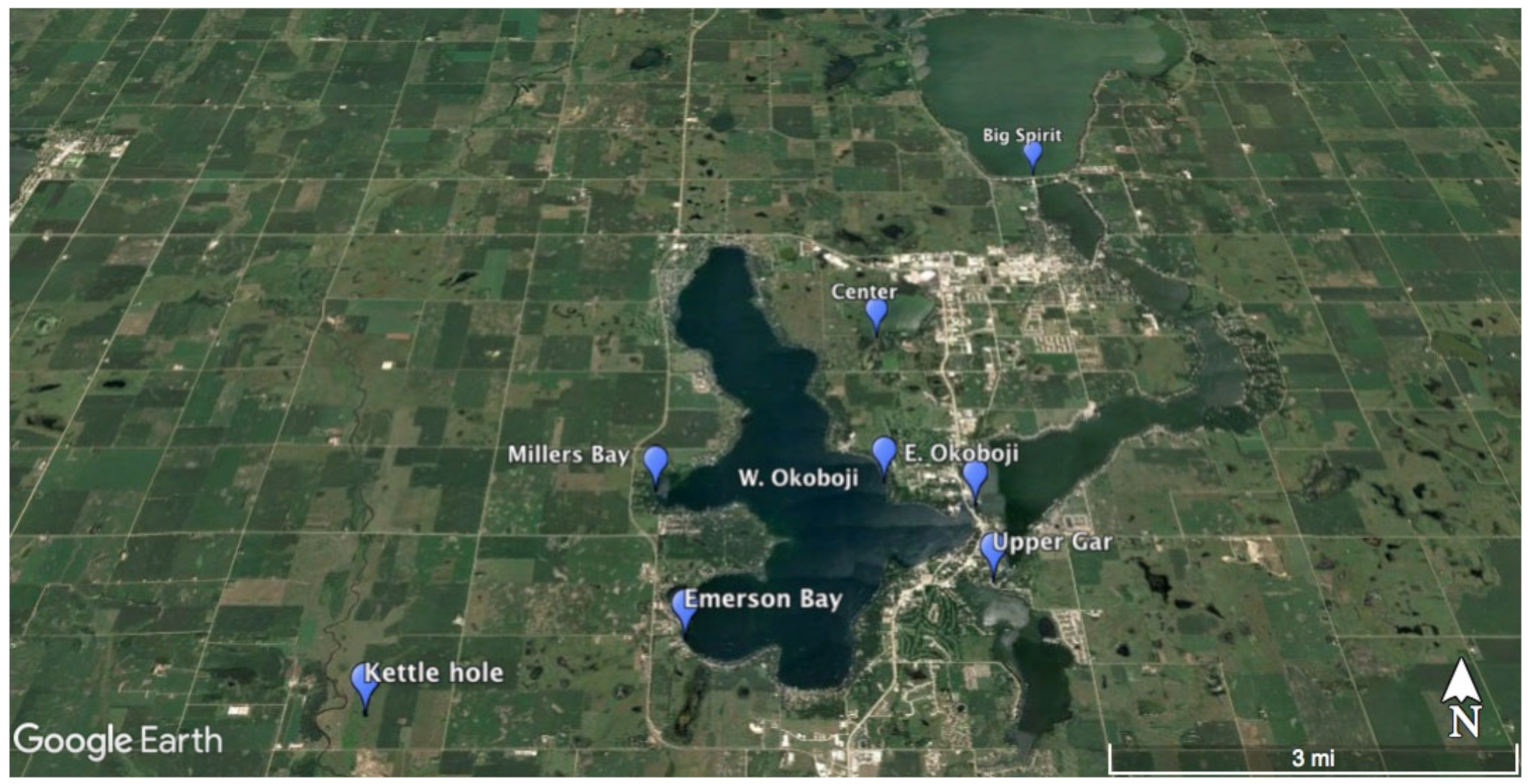
Composite littoral zone samples, following APHA standard methods and EPA periphyton protocols [28,29,30], were taken from all eight sites (Center Lake, Emerson Bay, Freda Haffner Kettlehole State Preserve, Millers Bay, East Okoboji Lake, West Okoboji Lake, Big Spirit Lake, Upper Gar Lake) during the months of June and July 2017 (Figure 1). All GPS coordinates for our sampling sites are located in Table 1. According to the Iowa Department of Natural Resources, the descriptions of these sites are as follows [31,32].
2.1. Sampling Site Descriptions
Center Lake—This lake has a surface area of 257 acres, a mean depth of 3.7 m, and a maximum depth of 5.5 m. Center Lake is found in the Des Moines Lobe ecoregion, and its surrounding land is predominantly grassland, followed by urban development, agriculture, and forest cover.
West Okoboji Lake—West Okoboji Lake has a surface area of 3847 acres, a mean depth of 11.6 m, and a maximum depth of 40.8 m. This lake’s surrounding land use is predominantly grassland, followed by agriculture, forest, and urban areas. This lake is located 274.3–457.2 m above sea levels and is also within the Des Moines Lobe ecoregion. Millers Bay and Emerson Bay are located within West Okoboji Lake: Millers Bay is on the southwestern side of the lake, and Emerson Bay is east of Millers Bay. Emerson Bay is open to the public for boat ramp access.
Freda Haffner Kettlehole State Preserve—sampling was conducted in a glacial kettle hole found in the Freda Haffner State Preserve. The land around this site consists of mainly native prairies, of which this State Preserve protects 110 acres.
East Okoboji Lake—This site is located at elevations of 274.3–457.2 m above sea levels within the Des Moines Lobe ecoregion. The surface area of East Okoboji Lake is 1835 acres, with a mean depth of 3 m and a maximum depth of 6.7 m. This lake’s surrounding land use consists mostly of agriculture, followed by grasslands, forest cover, and urban areas.
Big Spirit Lake—comprises the largest surface area of all eight sites (5684 acres). This lake has a mean depth of 5.18 m and a maximum depth of 7.32 m. Big Spirit Lake and its surrounding watershed are located in the Des Moines Lobe ecoregion, which is characterized by bluestem prairie, mollisols, and plains. The elevation of this ecoregion ranges from 274.3–457.2 m above sea levels. Surrounding land use of this site consists predominantly of agriculture, followed by grasslands, forest, and urban development.
Upper Gar Lake—This lake and its surrounding watershed are located in the Des Moines Lobe ecoregion. Upper Gar Lake comprises a surface area of 36.1 acres, a maximum depth of 1.5 m, and a mean depth of approximately 1 m. The surrounding land use of this lake consists mostly of agriculture, followed by grassland, forest, and urban development.
2.2. Taxonomic Evaluations
Live algal units were evaluated from samples within two days of sampling. Preservation changes the shape and color of chloroplasts. Therefore, after observation, the representative sample was preserved for further analysis and deposited in the Georgia College Natural History Museum Algae Collection.
Identification of algal units and micrography were conducted at 100X–1000X magnification using a Leica DM2000 microscope and Leica DFC295 camera (Leica Microsystems, Wetzlar, Germany). For each collected sample, a biomass estimation of physiologically active algae was made by scanning at least two entire slides for algal taxa that represented at least 10% or more of the total estimated algal biomass (relative abundance of the taxa documented per 100 units times cell biovolume). Species richness was documented for each site. An opportunistic approach was taken for algal taxa micrography, as it was not conducted for all taxa present at sites, but rather only for those that represented at least 10% or more observed biomass and which clearly displayed key identification characteristics like reproductive structures, pyrenoids, etc. Species identification was conducted using morphometrics for all documented populations by measuring 15 cells where possible. The nomenclatural status for all species was verified by using AlgaeBase [26].
While conducting taxonomic evaluations, handwritten archives from 1915 by Lewis Hanford Tiffany were found at Iowa Lakeside Laboratory. These archives document algal species collected from sites in our study. Critical evaluation of the document is valuable for knowing if any species have persisted for over 100 years at the same location because G. W. Prescott reported no studies of Iowa algae prior to 1920 [33]. The 1915 archives provide specific dates and locations for reference collections sampled by Lewis Hanford Tiffany (Ohio State University) during his visit to Iowa’s Lakeside Laboratory by the invitation of the then director, Dr. R. B. Wylie. Because collection methods and frequency are unknown, we do not compare our samples to the past written archives, but we do note if any species have persisted for over 100 years at these locations (Supplemental Materials Table S1 and Figure S1). Our review of Tiffany’s 1915 records was conducted through a current taxonomic perspective (i.e., all nomenclatural changes were considered).
3. Results
Overall, the taxa we collected represented nine different algal groups: Charophyta, (Zygnematophyceae), Cyanobacteria, Bacillariophyta, Ochrophyta, Chlorophyta, Dinophyceae, Euglenophyceae, and Glaucophyta and 54 unique taxonomic entities were documented in this study (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9; Supplemental Materials Table S2). Only 49% of the taxa were identified to species and variety level (reproductive structures were rarely encountered). Overall, 35% of these species were green algae, 26% were diatoms, 15% were cyanobacteria, 9% were euglenoids, and 7% and less were desmids, yellow-green algae, dinoflagellates, and glaucophytes. The Center Lake sample had the highest species richness (n = 13), followed by the kettle hole (n = 11). The kettle hole and Center Lake samples had the most desmid species (n = 3). However, the kettlehole sample had the highest number of Euglenoid algal species recorded (n = 5) and was the only site where Euglenoid genera were found to be dominant in the collected sample. Cyanobacteria were found in all site samples except East Okoboji Lake and Upper Gar Lake. Diatoms were present at all sites. Species richness for diatoms was highest in the Center Lake sample (n = 4), followed by East Okoboji Lake, West Okoboji Lake, and Big Spirit Lake (n = 3). Filamentous green algae were present at all sites but were not found in the kettle hole sample. Big Spirit Lake and West Okoboji Lake samples had the highest species richness for green algae (n = 4), followed by those for Center Lake and Emerson Bay (n = 3). One dinoflagellate and one Glaucophyte species were documented at the kettle hole. Big Spirit Lake and Emerson Bay comprised the only yellow-green algae found in these samples. The kettle hole sample was the most diverse in species richness across algal groups, with representatives of desmids, dinoflagellates, diatoms, glaucophytes, and Euglenoids. However, we did not find green algae in the kettle hole sample. Fifty percent of sites were dominated by green algae species, 25% by diatoms, and 12.5% by Euglenoid species.
Two species were found in our samples and 1915 records in the same location: Melosira varians in Center Lake and Cladophora glomerata in Millers Bay. Five taxa from our study were recorded in the 1915 archives, but in different lakes: Chaetophoropsis elegans (Roth) B. Wen Liu, Qian Xiong, X. Dong Liu, Z. Yu Hu & G. Xiang Liu 2019, Cladophora glomerata (L.) Kützing, Gloeotrichia echinulata P. G. Richter, Hydrodictyon reticulatum (Linnaeus) Bory 1824, Microcystis aeruginosa (Kützing) Kützing. In the 1915 records, filamentous green algae were documented in all locations, and cyanobacteria were recorded in all locations except East Okoboji Lake and the kettle hole.
3.1. Common Taxa
Cladophora glomerata (Linnaeus) Kützing 1843
Figure 3A–D; Figure 5D–F; Figure 6C; Figure 7D;
References: [40] (p. 138), Pl. 20, Figures 8 and 9; Pl. 21, Figures 1 and 2.
Documented from: Millers Bay, Emerson Bay, East and West Okoboji Lakes.
Cell sizes in our population vary, with cylindrical cells in main and side branches gradually decreasing in diameter and length-to-width ratio. Filaments profusely branched and attached by rhizoids, dark green, branching pseudodichotomous with insertion of side branches generally oblique to horizontal; cells cylindrical; chloroplast filling the volume of the cell parietal net-like with numerous pyrenoids; main axis up to 300 µm in diameter; cells of side branches 21.9–35.9 µm wide, 1.5 to 10 times longer than wide (41.4–100 µm in length).
Closterium parvulum Nägeli 1849
Figure 4I
References: [42] (p. 38), Pl. 6, Figure H.
Documented from: Freda Haffner Kettlehole State Preserve, Dickinson County.
Cell sizes in the current population fit the available literature. Cells strongly curved, outer margin with arc of 102–158°, inner margin sometimes straight, attenuating to acutely rounded apices; girdle bands absent; chloroplasts with 5–6 longitudinal ridges and 3–6 axial pyrenoids; wall smooth, colorless; terminal vacuoles containing 2–8 ovoid gypsum crystals; cells 14.4–19.4 µm wide, 137.4–146.2 µm long.
Closterium cf. dianae var. minus Hieronymus 1895
Figure 4J
References: [35] (p. 623), Pl. 153, Figure L.
Documented from: Freda Haffner Kettlehole State Preserve, Dickinson County.
Cells strongly curved, inner margin slightly tumid, attenuating to acutely rounded apices; girdle bands absent; chloroplasts with longitudinal ridges; wall smooth, colorless; cells 15 µm wide, 145 µm long.
Cocconeis pediculus Ehrenberg 1838
Figure 6C
References: [43] (p. 240), Pl. 15, Figures 3 and 4.
Documented from: East Okoboji Lake
Cell sizes in the current population vary, with cells ranging in sizes from 12–22 µm in breadth and 18.5–31.7 µm in length.
Cymbella mexicana (Ehrenberg) Cleve 1894
Figure 3J; Figure 7E
References: [37], vol. 2., Pl. 12, Figures 1 and 2
Documented from: Emerson Bay and West Okoboji Lake
Cell sizes in the current population vary, with cells ranging in sizes from 35.8–37.8 µm in breadth and 111.9–174.9 µm in length with striae 8 in 10 µm.
Dolichospermum circinale (Rabenhorst ex Bornet & Flahault) Wacklin, Hoffmann & Komárek 2009
Figure 2A
References: [34] (p. 703), Figure 867;
Documented from: Center Lake
Trichomes are coiled, usually solitary, embedded in diffluent colorless mucilage. Cells spherical, slightly bent, 10.4–12.4 µm wide, 7–11 µm long, filled with aerotopes. Heterocyst spherical, 10–11 µm in diameter. Akinete was not observed. In the collection from 1923, this taxon was reported as common in West Okoboji Lake [47].
Gloeotrichia echinulata P.G.Richter 1894
Figure 3L; Figure 5A–C
References: [40] (pp. 557–558) Pl. 134, Figures 1 and 2
Documented from: Emerson Bay and Millers Bay
Colonies are spherical, free-floating, yellow to brown, soft, and bullate. Colonies visible macroscopically. Trichomes are slightly bent, usually ending in a distinct hair radiating from a common center. Cells 2.1–5.2 µm wide, 2.9–23.23 µm in length, barrel-shaped, becoming quadrate or cylindrical in the apical area, sometimes thinning to a hair. Heterocyst basal, more or less spherical, 6–12 µm in diameter. Akinetes solitary, adjacent to heterocyst, cylindrical, straight or bent, with a thick, smooth wall, 10–18 µm wide, 40–250 µm long. In the collection from 1923, this taxon was reported as abundant in West Okoboji Lake and rare in Little Spirit and Spirit Lakes [47].
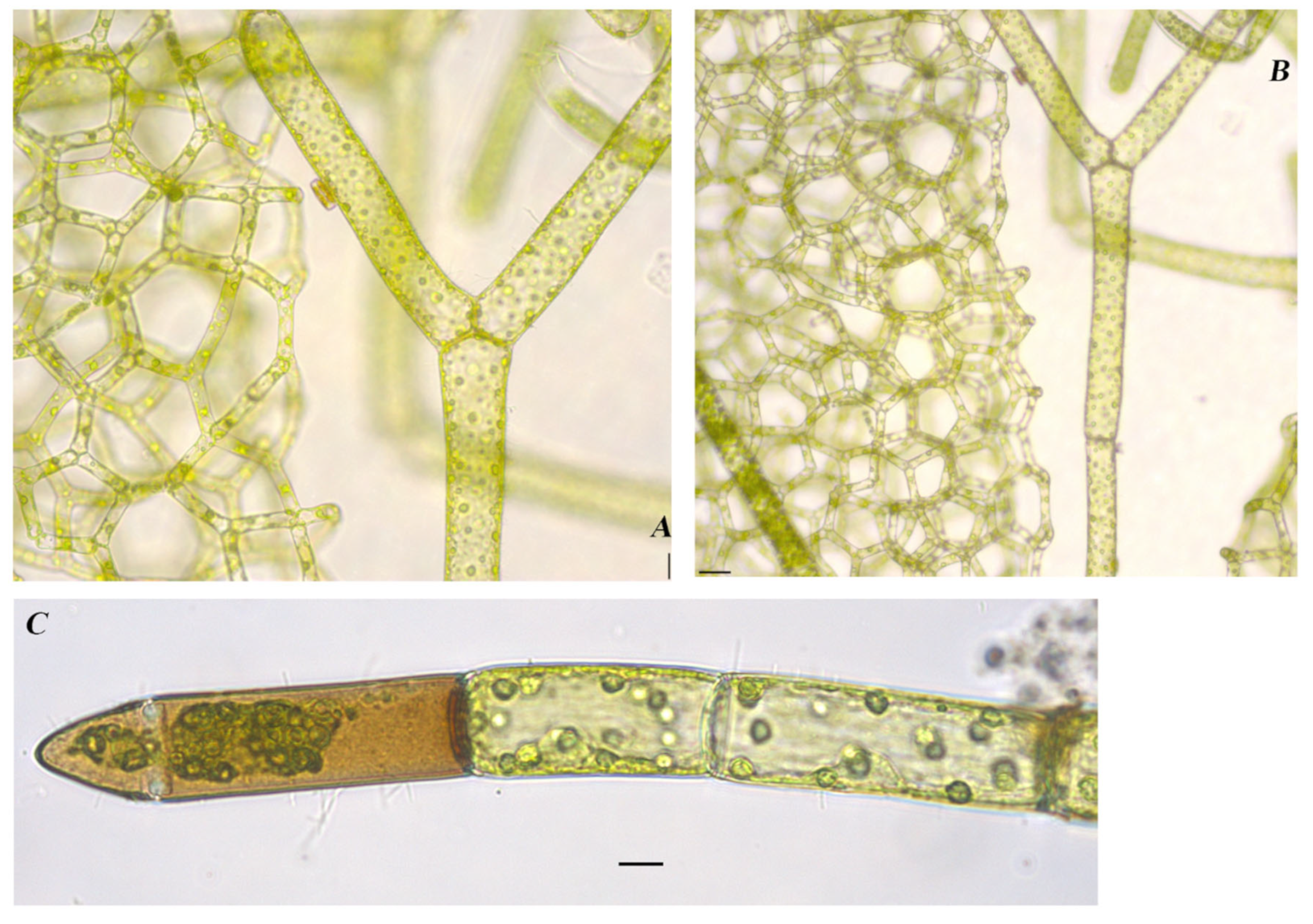
Hydrodictyon reticulatum (Linnaeus) Bory 1824
Figure 9A,B
References: [40] (p. 219), Pl. 47, Figure 1
Documented from: Upper Gar Lake
Coenobia contains cylindrical cells, which form colonies of a mesh or a net-like appearance. Cell arrangement forms pentagonal-shaped spaces with three to five cells connected by their edges throughout. Cell sizes in the population vary greatly, with small cells ranging in size from 11.3–12.5 µm in diameter and 47.5–62.5 µm in length to large cells ranging from 32.5–37.5 in diameter and 230–315 µm in length. Chloroplast parietal with many pyrenoids.
Klebsormidium cf. mucosum (J.B. Petersen) Lokhorst 1985
Figure 8J–N
References: [35] (p. 556), Pl. 138E.
Documented from: Big Spirit Lake
Cell sizes in the population vary, with cells ranging in size from 21.9–35.9 µm in diameter and 41.4–61.7 µm in length. Ours are slightly larger than what is in reference.
Microcystis aeruginosa (Kützing) Kützing 1846
Figure 3H,I
References: [35] (p. 69), Pls. 4C, 11K.
Documented from: Emerson Bay
Colonies are microscopic, mucilaginous, irregular, and clathrate with distinct holes or composed of subcolonies with irregularly and densely packed cells. Mucilage colorless. Cells spherical to slightly elongate, with numerous aerotopes 4.3–4.6 µm in diameter and 4.6–10.3 µm in length. In the collection from 1923, this taxon was reported as rare in East Okoboji Lake, Welch Lake, and Pleasant Lake [47].
Phacus acuminatus A. Stokes 1885
Figure 4B
References: [35] (p. 206), Pl. 50S.
Documented from: Freda Haffner Kettlehole State Preserve
Cells ovate to oval in outline with greatest width below middle, thin, shallow dorsal furrow extending half to ¾ cell length, short cauda at posterior end, shallow incision at anterior end; numerous parietal disc-shaped chloroplasts; 1–2 ring-like paramylon bodies; visible pellicle strips longitudinally striated, flagellum approximately length of cell. Cell size 22.8 µm wide and 24.5 µm long. An eye spot is evident at the base of the flagellum.
Spirogyra cf. borgeana Transeau 1916
Figure 7B
Documented from: West Okoboji Lake
Cell sizes in the population range in sizes of 27–27.3 µm in diameter and 70.8–72 µm in length. Size fits reports from North American populations, with cell length about two times the diameter. Chloroplast in the form of a spiral band spiraling around five times. Several pyrenoids in a linear row. Gametangia and zygospores were not observed.
Woronichinia naegeliana (Unger) Elenkin 1933
Figure 2M,N
References: [35] (p. 77), Pls. 6B, 13 C, D.
Documented from: Center Lake
Spherical or irregular microscopic colony, with densely arranged cells at the periphery of the colony with numerous aerotopes, up to 96- 165 µm in diameter and 155.2–177.7 µm in length. Cell obovoid/ellipsoid of 5.2–7.25 µm in diameter and 4.7–7.9 µm in length. This taxon was reported by Smith [47] as Coelosphaerium Naegelianum Unger and was reported as rare in Center Lake, Upper Gar Lake, and several sampling locations of East and West Okoboji Lakes.
3.2. Unique Ttaxa
Chaetophoropsis elegans (Roth) B.Wen Liu, Qian Xiong, X.Dong Liu, Z.Yu Hu & G.Xiang Liu 2019
Taxonomic synonym: Chaetophora elegans (Roth) C. Agardh 1812
Figure 8D,E
Documented from: Big Spirit Lake
Thallus globose, with irregular light or dark green mucilaginous masses. Attached by rhizoids. Colonies are continuous and intertwined, most often with uniseriate, dichotomous branching, which radiates from a common center. The uppermost branches of the thallus are more numerous and densely arranged than those branching near the center, ending infrequently with pointed cells. Cells 10–11.6 µm in diameter near the center of thallus, 21.6–37.12 µm long in the main axis. Chloroplast is a parietal band with 1-more pyrenoids.
Draparnaldia acuta (C.Agardh) Kützing 1845
Figure 6A,B
References: [40] (pp. 119–120), Pl. 15, Figure 5
Documented from: East Okoboji Lake
Filaments branched under a right angle from the main axis, attached by rhizoids, and enclosed within a soft mucilaginous envelope. Primary branches bearing oppositely, alternatively or in whorls tufts of richly divided and smaller-celled secondary branches lateral branching under sharp angles. Small branching terminal cells end in a fine hair-like structure. Cells of primary branches 37.8–47 µm in diameter and 59.4–84.4 µm in length, square-shaped without constrictions. Chloroplast is a parietal band in the cells of the main axis with several pyrenoids; secondary branches alternatively divided and with a distinct main axis, 25.9–37 µm in diameter and 27.3–38.8 µm in length. This taxon was attached, and the filaments were enclosed in a soft mucilaginous envelope.
Glaucocystis nostochinearum Itzigsohn 1866
Figure 4A,C
Documented from: Freda Haffner Kettlehole State Preserve
Cell sizes vary from 29.64–35.74 µm in length and 17.8–20.7 µm in width. Cells were observed solitary and in 8 cell colonies. The documented colony in Figure 4 measures an old mother cell with visible nodules, which was 73.71 in length and 54.72 in width. Important taxon from soft water habitats [40] showing round, potentially coccoid cyanobacteria, endosymbionts with bright blue coloring due to phycocyanin and allophycocyanin [49].
4. Discussion
Authors’ description-based voucher flora is important for informing long-term monitoring and future technologies, especially with current issues of harmful algal blooms in surface water resources. We observed cyanobacteria genera at four sampling locations: Millers Bay, Emerson Bay, West Okoboji Lake, and Center Lake. One species known to potentially produce toxins, Microcystis aeruginosa, was found in Emerson Bay. In Center Lake, we observed Dolichospermum, which is a genus also associated with toxic algal blooms [50]. Records from the summer of 1915 of the same locations sampled in our study document potentially toxin-producing cyanobacteria in all sites except for the kettle hole and Emerson Bay. Although uncertainty exists when referring to historical archives, such as unclear sampling methodology and proximity of location, they provide rare historical evidence of gamma diversity that contributes to a complete understanding of the aquatic ecology of the region. Due to these challenges and the taxonomic nature of our study, the abundance or changes of cyanobacteria populations at these sites is unclear and would require more in-depth sampling. Because a significant increase in cyanobacteria might not cause systems to collapse but could yield lower gamma biodiversity, which could go unnoticed [51], we recommend continued monitoring of these sites.
In the 1915 archives, many green algal species were documented to occur at the kettle hole. However, in our kettle hole sample, no green algae were documented. This could potentially be due to the higher precipitation during our study year [52] or sampling effort, given the nature of this study. Representatives of Zygnemataceae (phylum Charophyta), filamentous or coccoid algal family with potential for sexual reproduction by conjugation, which produces large amounts of biomass, require high light availability [53], and prefer standing water with minimal mechanical disturbances as they are holdfast-challenged [54]. It is interesting that the sample from the kettle hole (recognized as a protected site) would comprise more euglenoid taxa than any of the other samples, given its formation occurred from glacier melting and there is no boat traffic as observed at other sites. Studies have shown that species of class Euglenophyceae are found in sewage water and have been used as biological indicators for pollution and acidic peat bogs [55]. With high evaporation, the kettle hole preserve site gets shallower, and macrophyte vegetation might shade representatives of Zygnemataceae leading to the proliferation of euglenoid representatives. The kettle hole and Center Lake had the highest species richness for desmids. Desmids are usually found in mostly acidic soft water wetlands, Sphagnum bogs, or shallow pools with poor aeration [56]. Another algal group known to occur in Sphagnum bogs is Glaucophytes [57]. We document a Glaucocystis species in its living environment at the kettle hole. This taxon is especially unique given it is a primitive genus that has been indicated in the primary endosymbiosis event, making it the closest relative to green plants and red algae [58].
The value of historical documents is rarely critically evaluated by ecologists. However, the value that historical data provide cannot be overlooked [59,60,61]. When examining biodiversity, there is value in understanding what biota survive and persist over time. A diatom species, Melosira varians, was found at the same site (Center Lake) 100 years later. This is not surprising since this species is common in North America and is found in pristine or poor water quality environments [45,62]. Another alga occurring at the same site 100 years later is Cladophora glomerata from Millers Bay. C. glomerata was found at two of our study sites and six of the 1915 sites. C. glomerata can be found in oligotrophic environments, but blooms are usually associated with eutrophic conditions from human activity and a pH ranging from 7 to 10 [63,64]. The presence of C. glomerata at most of the sites in 1915 could suggest nutrient enrichment of these waters, but without species abundance data, it remains unknown. C. glomerata is considered sensitive to heavy metals and should be absent in conditions of heavy metal contamination [65]. Interestingly, a unique taxon from Big Spirit Lake, Chaetophoropsis elegans has the ability to remove pollutants like heavy metals from water via biosorption [66,67]. Chaetophorales have also been shown to develop hairs in P-limited conditions [68]. In our East Okoboji Lake sample, we observed another unique taxon belonging to the genus Draparnaldia, which has the ability to form hairs in P-limited conditions.
Benthic and littoral algal community analyses of the northwest region of Iowa have not been completed recently, and little is known about the history of the algal species that are present in Iowa’s great lakes. With the exception of extensive paleo research from cores [69] and monitoring and recording of phytoplankton to genus level [27], there are no current natural history studies from the area that use cell physiology and community composition. Taxonomic data with micrographs of seven hydrologically related aquatic habitats and a nature preserve provide valuable insight into community composition and diversity in this region. Our paper is one of the few documenting and describing these algal communities. A continued long-term composite sampling of these sites is crucial as higher biodiversity may be found in niche partitioning [70,71] and, still to be uncovered, molecular diversity, as shown for a Zygnematophyceae community [72]. Although it is also difficult to draw conclusions about these sites as a whole from one “snapshot” sampling event, focusing on fine-level taxonomy is a valuable tradeoff.
5. Conclusions
In the field of phycology, there remains a gap between technological advances and publicly available reference information to use these advanced tools accurately. As the authors point out, there is a lack of support for collection-based taxonomic approaches that would provide this necessary reference information [14]. Literature on algal taxonomy is often difficult to access, out of print, or outdated, relying on hand drawings for species descriptions. Therefore, providing descriptive voucher flora with micrographs is essential when documenting and monitoring biodiversity and for the advancement of the field of phycology.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16010021/s1, Figure S1. Image of L.H. Tiffany’s handwritten algal records; Table S1. Algal taxa from nine sites in Dickinson County, Iowa, transcribed from archives of L.H. Tiffany’s handwritten documents from 1915. Taxa with an asterisk (*) were found in current samples from the same site. Taxa denoted by a cross (†) were found in current samples from a different site; Table S2. Algal taxa were documented in a 2017 study at eight sites in Dickinson County, Iowa.
Author Contributions
Conceptualization, K.M.J. and K.M.M.; methodology, K.M.J.; software, K.M.J.; validation, K.M.J. and K.M.M.; formal analysis, K.M.J.; investigation, K.M.J.; resources, K.M.M.; data curation, K.M.M., writing—original draft preparation, K.M.J.; writing—review and editing, K.M.J. and K.M.M.; visualization, K.M.J.; supervision, K.M.M.; project administration, K.M.M.; funding acquisition, K.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no other funding but from GCSU.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Material.
Acknowledgments
Thank you to the students of the 2017 Ecology and Systematics of Algae course at Iowa Lakeside Lab for their help in collecting the material. We are thankful to Mary Skopec and the Friends of Lakeside at Iowa Lakeside Laboratory for awarding a visiting research scholarship to Katherine M. Johnson. This manuscript benefitted from discussions and review by Mindy Morales and Rosalina Christova. This work was supported by the Department of Biological and Environmental Sciences and the Graduate School at Georgia College & State University. Finally, we want to extend our gratitude to the 2 anonymous reviewers and the editorial team for their constructive comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Godfray, H.C.J. Challenges for taxonomy. Nature 2002, 417, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, M.; Ács, É.; Almeida, S.F.; Blanco, S.; Dreßler, M.; Ector, L.; Karjalainen, S.M.; Liess, A.; Mertens, A.; Van der Wal, J.; et al. Quality assurance of diatom counts in Europe: Towards harmonized datasets. Hydrobiologia 2016, 772, 1–14. [Google Scholar] [CrossRef]
- Conix, S.; Cuypers, V.; Zachos, F.E.; De Block, A. Taxonomic disagreement about ranks in gray-area taxa: A vignette study. BioScience 2023, 73, 728–737. [Google Scholar] [CrossRef]
- De Luca, D.; Piredda, R.; Sarno, D.; Kooistra, W. Resolving cryptic species complexes in marine protists: Phylogenetic haplotype networks meet global DNA metabarcoding datasets. ISME J. 2021, 15, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Zieritz, A.; Lee, P.S.; Eng, W.W.H.; Lim, S.Y.; Sing, K.W.; Chan, W.N.; Loo, J.S.; Mahadzir, F.N.; Ng, T.H.; Yeo, D.C.; et al. DNA metabarcoding unravels unknown diversity and distribution patterns of tropical freshwater invertebrates. Freshw. Biol. 2022, 67, 1411–1427. [Google Scholar] [CrossRef]
- Lai, J.; Liang, Q.; Zhang, X.; Liu, Y.; Wang, M.; Yang, W.; Sun, T.; Li, Y.; Jin, H.; Liu, Y.; et al. FWAlgaeDB, an integrated genome database of freshwater algae. Front. Environ. Sci. 2023, 11, 1178097. [Google Scholar] [CrossRef]
- Noble, P.J.; Seitz, C.; Lee, S.S.; Manoylov, K.M.; Chandra, S. Characterization of algal community composition and structure from the nearshore environment, Lake Tahoe (United States). Front. Ecol. Evol. 2023, 10, 1053499. [Google Scholar] [CrossRef]
- Alers-García, J.; Lee, S.S.; Spaulding, S.A. Resources and practices to improve diatom data quality. Limnol. Oceanogr. Bull. 2021, 30, 48–53. [Google Scholar] [CrossRef]
- Johnson, K.M.; Gaiser, E.; Manoylov, K.M. Diatom Voucher Flora and Comparison of Collection and Taxonomic Methods for Biodiversity Hotspot Upper Three Runs Creek. Water 2023, 15, 2578. [Google Scholar] [CrossRef]
- Brown, S.; Manoylov, K.M. Assessing Diatom Community Dynamics in a Recovering Agricultural Stream in Middle Georgia, USA. Phycology 2023, 3, 294–304. [Google Scholar] [CrossRef]
- Bishop, I.W.; Esposito, R.M.; Tyree, M.; Spaulding, S.A. A diatom voucher flora from selected southeast rivers (USA). Phytotaxa 2017, 332, 101–140. [Google Scholar] [CrossRef]
- Spaulding, S.A.; Potapova, M.; Bishop, I.W.; Lee, S.S.; Gasperak, T.S.; Jovanoska, E.; Furey, P.C.; Edlund, M.B. Diatoms.org: Supporting taxonomists, connecting communities. Diatom Res. 2021, 36, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Barinova, S.; Smith, T. Flora of Algae and Cyanobacteria of Continental Waters of Israel in the XXI Century: Taxonomy, Autecology and Water Quality Indicators. Diversity 2022, 14, 328. [Google Scholar] [CrossRef]
- Löbl, I.; Klausnitzer, B.; Hartmann, M.; Krell, F.T. The silent extinction of species and taxonomists—An appeal to science policymakers and legislators. Diversity 2023, 15, 1053. [Google Scholar] [CrossRef]
- Wheeler, Q.D.; Raven, P.H.; Wilson, E.O. Taxonomy: Impediment or expedient? Science 2004, 303, 285. [Google Scholar] [CrossRef]
- Heathcote, A.J.; Downing, J.A. Impacts of eutrophication on carbon burial in freshwater lakes in an intensively agricultural landscape. Ecosystems 2012, 15, 60–70. [Google Scholar] [CrossRef]
- Volker, R.; Smith, S.G. Changes in the aquatic vascular flora of Lake East Okoboji in historic times. Proc. Iowa Acad. Sci. 1965, 72, 65–72. [Google Scholar]
- Egertson, C.J.; Kopaska, J.A.; Downing, J.A. A century of change in macrophyte abundance and composition in response to agricultural eutrophication. Hydrobiologia 2004, 524, 145–156. [Google Scholar] [CrossRef]
- Braun, L.E. Deciduous Forests of Eastern North America; Hafner Publishing Company: New York, NY, USA, 1964. [Google Scholar]
- Smith, D.D. Iowa prairie: Original extent and loss, preservation and recovery attempts. J. Iowa Acad. Sci. JIAS 1998, 105, 94–108. [Google Scholar]
- Rose, E.T. Toxic algae in Iowa lakes. Proc. Iowa Acad. Sci. 1953, 60, 738–746. [Google Scholar]
- Greene, S.B.D.; LeFevre, G.H.; Markfort, C.D. Improving the spatial and temporal monitoring of cyanotoxins in Iowa lakes using a multiscale and multi-modal monitoring approach. Sci. Total Environ. 2021, 760, 143327. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and algae blooms: Review of health and environmental data from the harmful algal bloom-related illness surveillance system (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [PubMed]
- Heathcote, A.J.; Ramstack Hobbs, J.M.; Anderson, N.J.; Frings, P.; Engstrom, D.R.; Downing, J.A. Diatom floristic change and lake paleoproduction as evidence of recent eutrophication in shallow lakes of the Midwestern USA. J. Paleolimnol. 2014, 53, 17–34. [Google Scholar] [CrossRef]
- Wehr, J.D.; Sheath, R.G.; Kociolek, J.P. Freshwater Algae of North America, 2nd Edition: Ecology and Classification; Academic Press: Cambridge, MA, USA, 2015; p. 1066. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. In World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2023; Available online: http://www.algaebase.org (accessed on 22 November 2023).
- Iowa DNR Ambient Lake Monitoring. Available online: https://www.iowadnr.gov/Environmental-Protection/Water-Quality/Water-Monitoring/Ambient-Lake-Monitoring (accessed on 22 November 2023).
- American Public Health Association; American Water Works Association; Water Pollution Control Federation; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater Centennial Edition; American Public Health Association: Washington, DC, USA, 2005; pp. 10–32. [Google Scholar]
- Stevenson, R.J.; Bahls, L.L. Periphyton protocols. In Rapid Bioassessment Protocols for Use in Wadeable Streams and Rivers; Barbour, M.T., Gerritsen, J., Snyder, B.D., Stribling, J.B., Eds.; EPA 841-B-99-002; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999; pp. 6.1–6.22. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish; US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999; Volume 339.
- Iowa DNR Lakes Ponds & Reservoirs. Available online: https://www.iowadnr.gov/Fishing/Where-to-Fish/Lakes-Ponds-Reservoirs (accessed on 22 November 2023).
- Iowa DNR Iowa’s Ecoregions. Available online: https://programs.iowadnr.gov/bionet/Docs/Ecoregions/ (accessed on 22 November 2023).
- Prescott, G.W. Iowa Algae; The University of Iowa Press: Iowa City, IA, USA, 1931. [Google Scholar]
- Komárek, J. Süsswasserflora von Mitteleuropa. Cyanoprokaryota: 3rd Part: Heterocystous Genera; Springer Spektrum: Heidelberg, Germany, 2013; Volume 19, pp. 1–1130. [Google Scholar]
- John, D.M.; Whitton, B.A.; Brook, A.J. The Freshwater Algae Flora of the British Isles; Press Syndicate of the University of Cambridge: Cambridge, UK, 2002. [Google Scholar]
- Potapova, M. Diatoma vulgaris. In Diatoms of North America. 2009. Available online: https://diatoms.org/species/diatoma_vulgaris (accessed on 22 November 2023).
- Patrick, R.; Reimer, C.W. The Diatoms of the United States, Exclusive of Alaska and Hawaii: Entomoneidaceae, Cymbellaceae, Gomphonemaceae, Epithemiaceae; Monographs of the Academy of Natural Sciences of Philadelphia: Philadelphia, PA, USA, 1975; Volume 2. [Google Scholar]
- Potapova, M. Melosira varians. In Diatoms of North America. 2009. Available online: https://diatoms.org/species/melosira_varians (accessed on 22 November 2023).
- Prescott, G.W. Algae of the Western Great Lakes Area. With an Illustrated Key to the Genera of Desmids and Freshwater Diatoms, 2nd ed.; Brown Company Publishers: Dubuque, IA, USA, 1962; pp. 1–977. [Google Scholar]
- Prescott, G.W. Algae of the Western Great Lakes Area Exclusive of Desmids and Diatoms; Bulletin of the Cranbrook Institute of Science: Bloomfield Hills, MI, USA, 1951; Volume 31, pp. 1–946. [Google Scholar]
- Brook, A.J.; Williamson, D.B.; John, D.M. Phylum Chlorophyta. Suborder Desmidiineae. In The Freshwater Algal Flora of the British Isles. An Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; John, D.M., Whitton, B.A., Brook, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 636–741. [Google Scholar]
- John, D.M.; Williamson, D.B. A Practical Guide to the Desmids of the West of Ireland; Martin Ryan Institute, National University of Ireland: Galway, Ireland, 2009; p. 38. [Google Scholar]
- Patrick, R.; Reimer, C.W. The Diatoms of the United States, Exclusive of Alaska and Hawaii: Fragilariaceae, Eunotiaceae, Achnanthaceae, Naviculacae; Monographs of the Academy of Natural Sciences of Philadelphia: Philadelphia, PA, USA, 1966. [Google Scholar]
- Transeau, E.N. Notes on the Zygnemales. Ohio J. Sci. 1916, 16, 17–31. [Google Scholar]
- Woods, P. Stephanodiscus Reimeri. In Diatoms of North America. 2011. Available online: https://diatoms.org/species/stephanodiscus_reimeri (accessed on 22 November 2023).
- Komárek, J. Studies on the Cyanophytes of Cuba 4-6. Folia Geobot. 1989, 24, 57–97. [Google Scholar] [CrossRef]
- Smith, G.M. The plankton algae of the Okoboji Region. Trans. Am. Microsc. Soc. 1926, 45, 156–233. [Google Scholar] [CrossRef]
- Takahashi, T.; Nishida, T.; Tuji, A.; Saito, C.; Matsuzaki, R.; Sato, M.; Toyooka, K.; Yasuda, H.; Nozaki, H. Delineation of six species of the primitive algal genus Glaucocystis based on in situ ultrastructural characteristics. Sci. Rep. 2016, 6, 29209. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.E.; Graham, J.M.; Wilcox, L.W. Algae, 2nd ed.; Benjamin Cummings: San Francisco, CA, USA, 2009; p. 616. [Google Scholar]
- Li, X.; Dreher, T.W.; Li, R. An overview of diversity, occurrence, genetics and toxin production of bloom-forming Dolichospermum (Anabaena) species. Harmful Algae 2016, 54, 54–68. [Google Scholar] [CrossRef]
- Dayton, P.K.; Tegner, M.J.; Edwards, P.B.; Riser, K.L. Sliding baselines, ghosts, and reduced expectations in kelp forest communities. Ecol. Appl. 1998, 8, 309–322. [Google Scholar] [CrossRef]
- Iowa State University Iowa Environmental Mesonet. Available online: https://mesonet.agron.iastate.edu/ASOS/reports/mon_prec.php?year=2017 (accessed on 22 November 2023).
- Graham, J.M.; Lembi, C.A.; Adrian, H.L.; Spencer, D.E. Physiological responses to temperature and irradiance in Spirogyra (Zygnematales, Charophyceae). J. Phycol. 1995, 31, 531–540. [Google Scholar] [CrossRef]
- Tiffany, L.H. The filamentous algae of Northwestern Iowa with special reference to the Oedogoniaceae. Trans. Am. Microsc. Soc. 1926, 45, 69–132. [Google Scholar] [CrossRef]
- Munawar, M. Ecological studies of Euglenineae in certain polluted and unpolluted environments. Hydrobiologia 1972, 39, 307–320. [Google Scholar] [CrossRef]
- Prescott, G.W. Desmids. Bot. Rev. 1948, 14, 644–676. [Google Scholar] [CrossRef]
- Griffiths, B.M. On Glaucocystis Nostochinearum, Itzigsohn. Ann. Bot. 1915, 3, 423–432. [Google Scholar] [CrossRef]
- Moreira, D.; Le Guyader, H.; Philippe, H. The origin of red algae and the evolution of chloroplasts. Nature 2000, 405, 69–72. [Google Scholar] [CrossRef] [PubMed]
- McClenachan, L.; Ferretti, F.; Baum, J.K. From archives to conservation: Why historical data are needed to set baselines for marine animals and ecosystems. Conserv. Lett. 2012, 5, 349–359. [Google Scholar] [CrossRef]
- Zu Ermgassen, P.S.; Spalding, M.D.; Blake, B.; Coen, L.D.; Dumbauld, B.; Geiger, S.; Grabowski, J.H.; Grizzle, R.; Luckenbach, M.; McGraw, K.; et al. Historical ecology with real numbers: Past and present extent and biomass of an imperilled estuarine habitat. Procedings R. Soc. B Biol. Sci. 2012, 279, 3393–3400. [Google Scholar] [CrossRef]
- Thurstan, R.H.; Campbell, A.B.; Pandolfi, J.M. Nineteenth century narratives reveal historic catch rates for Australian snapper (Pagrus auratus). Fish Fish. 2016, 17, 210–225. [Google Scholar] [CrossRef]
- Cardinale, B.J. Biodiversity improves water quality through niche partitioning. Nature 2011, 472, 86. [Google Scholar] [CrossRef]
- Whitton, B.A. Biology of Cladophora in freshwaters. Water Res. 1970, 4, 457–476. [Google Scholar] [CrossRef]
- Higgins, S.N.; Malkin, S.Y.; Todd Howell, E.; Guildford, S.J.; Campbell, L.; Hiriart-Baer, V.; Hecky, R.E. An ecological review of Cladophora glomerata (Chlorophyta) in the Laurentian Great Lakes. J. Phycol. 2008, 44, 839–854. [Google Scholar] [CrossRef] [PubMed]
- Whitton, B.A.; Kelly, M.G. Use of algae and other plants for monitoring rivers. Aust. J. Ecol. 1995, 20, 45–56. [Google Scholar] [CrossRef]
- Andrade, A.D.; Rollemberg, M.C.E.; Nóbrega, J.A. Proton and metal binding capacity of the green freshwater alga Chaetophora elegans. Process Biochem. 2005, 40, 1931–1936. [Google Scholar] [CrossRef]
- El Jamal, M.M.; Ncibi, M.C. Biosorption of methylene blue by Chaetophora elegans algae: Kinetics, equilibrium and thermodynamic studies. Acta Chim. Slov. 2012, 59, 24–31. [Google Scholar] [PubMed]
- Gibson, M.T.; Whitton, B.A. Hairs, phosphatase activity and environmental chemistry in Stigeoclonium, Chaetophora and Draparnaldia (Chaetophorales). Br. Phycol. J. 1987, 22, 11–22. [Google Scholar] [CrossRef]
- Heathcote, A.J. Anthropogenic Eutrophication and Ecosystem Functioning in Freshwater Lakes. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2013; pp. 1–158. [Google Scholar]
- Hutchinson, G.E. The Ecological Theater and the Evolutionary Play; Yale University Press: New Haven, CT, USA, 1965. [Google Scholar]
- Tilman, D. Resource competition between planktonic algae: An experimental and theoretical approach. Ecology 1977, 58, 338–348. [Google Scholar] [CrossRef]
- Pichrtová, M.; Holzinger, A.; Kulichová, J.; Ryšánek, D.; Šoljaková, T.; Trumhová, K.; Nemcova, Y. Molecular and morphological diversity of Zygnema and Zygnemopsis (Zygnematophyceae, Streptophyta) from Svalbard (High Arctic). Eur. J. Phycol. 2018, 53, 492–508. [Google Scholar] [CrossRef]
Figure 1.
Dickinson County, IA algal sampling sites (Center Lake, Emerson Bay, Freda Haffner Kettlehole State Preserve, Millers Bay, East Okoboji Lake, West Okoboji Lake, Big Spirit Lake, Upper Gar Lake) for summer 2017.
Figure 1.
Dickinson County, IA algal sampling sites (Center Lake, Emerson Bay, Freda Haffner Kettlehole State Preserve, Millers Bay, East Okoboji Lake, West Okoboji Lake, Big Spirit Lake, Upper Gar Lake) for summer 2017.

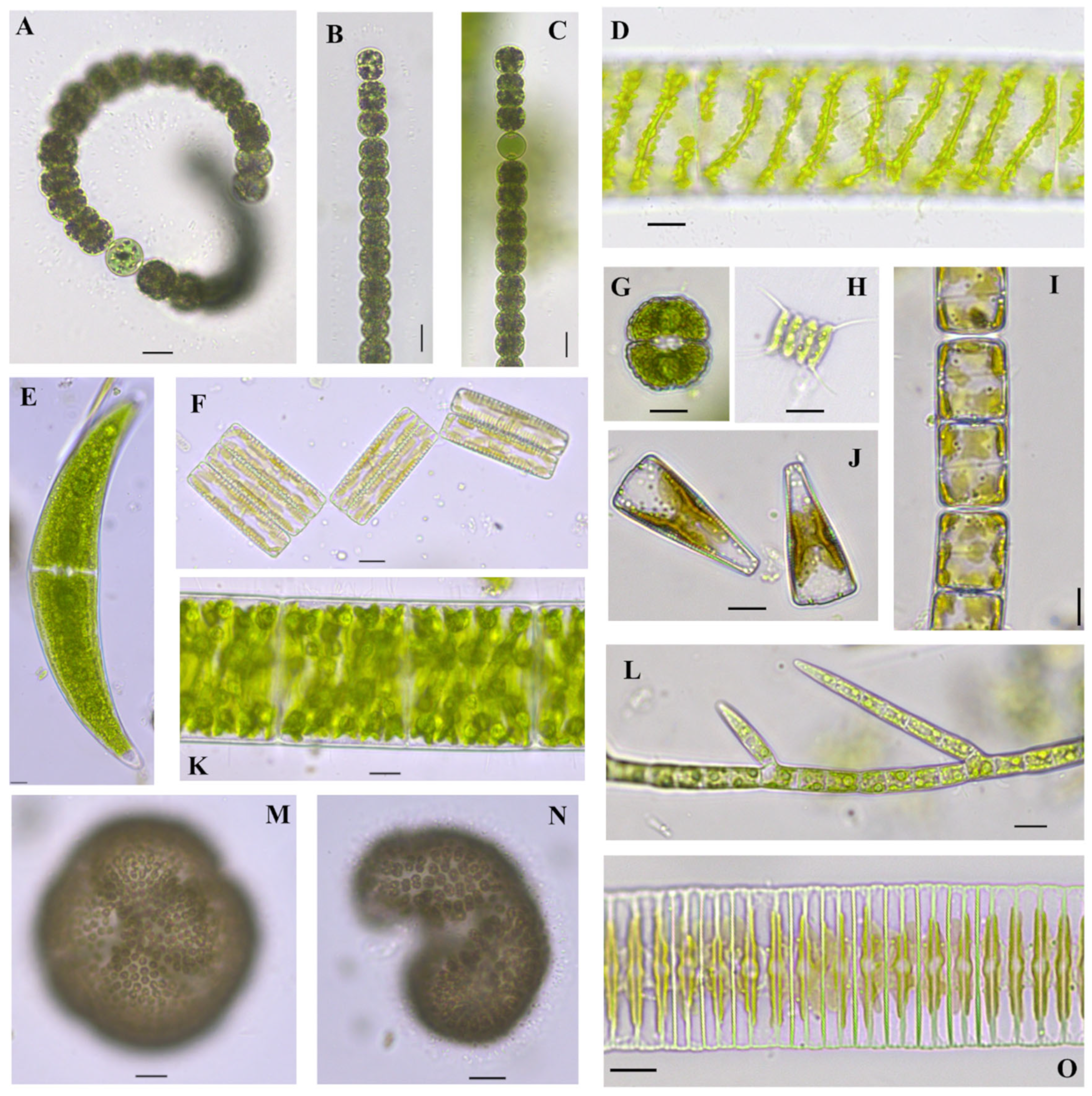
Figure 2.
Algae found in Center Lake, Dickinson County, Iowa. (A). Dolichospermum circinale (Rabenhorst ex Bornet & Flahault) Wacklin, Hoffmann & Komárek 2009 [34], (B,C). Dolichospermum viguieri (Denis and Frémy) Wacklin, L.Hoffmann and Komárek 2009 [34], (D). Spirogyra sp. 1, (E). Closterium ehrenbergii Meneghini ex Ralfs 1848 [35], (F). Diatoma vulgaris Bory 1824 [36] girdle view chain, (G). Cosmarium subcrenatum Hantzsch 1868 [26], (H). Scenedesmus quadricauda (Turpin) Brébisson 1835 [37], (I). Melosira varians C.Agardh 1827 [38] chain, (J). Gomphonema sp. girdle view, (K). Spirogyra sp. 2, (L). Stigeoclonium lubricum (Dillwyn) Kützing 1845 [39], (M,N). Woronichinia naegeliana (Unger) Elenkin 1933 [35] scale bar 20 µm, (O). Fragilaria sp. chain. Scale bars equal 10 µm if not otherwise indicated.
Figure 2.
Algae found in Center Lake, Dickinson County, Iowa. (A). Dolichospermum circinale (Rabenhorst ex Bornet & Flahault) Wacklin, Hoffmann & Komárek 2009 [34], (B,C). Dolichospermum viguieri (Denis and Frémy) Wacklin, L.Hoffmann and Komárek 2009 [34], (D). Spirogyra sp. 1, (E). Closterium ehrenbergii Meneghini ex Ralfs 1848 [35], (F). Diatoma vulgaris Bory 1824 [36] girdle view chain, (G). Cosmarium subcrenatum Hantzsch 1868 [26], (H). Scenedesmus quadricauda (Turpin) Brébisson 1835 [37], (I). Melosira varians C.Agardh 1827 [38] chain, (J). Gomphonema sp. girdle view, (K). Spirogyra sp. 2, (L). Stigeoclonium lubricum (Dillwyn) Kützing 1845 [39], (M,N). Woronichinia naegeliana (Unger) Elenkin 1933 [35] scale bar 20 µm, (O). Fragilaria sp. chain. Scale bars equal 10 µm if not otherwise indicated.

Figure 3.
Algae found in Emerson Bay, Dickinson County, Iowa. (A,D). Cladophora sp. scale bar 40 µm, (B,C). Cladophora sp. scale bar 20 µm, (E). Spirogyra sp. 3, (F). Spirogyra sp. 4, (G). Spirogyra sp. 5, (H,I). Microcystis aeruginosa (Kützing) Kützing 1846 [35] scale bar 20 µm, (J). Cymbella mexicana (Ehrenberg) Cleve 1894 [37], (K). Tribonema sp., (L). Gloeotrichia echinulata P.G. Richter 1894 [40], (M). Quadrigula chodatii (Tanner-Füllemann) G.M.Smith 1920 [39]. Scale bars equal 10 µm if not otherwise indicated.
Figure 3.
Algae found in Emerson Bay, Dickinson County, Iowa. (A,D). Cladophora sp. scale bar 40 µm, (B,C). Cladophora sp. scale bar 20 µm, (E). Spirogyra sp. 3, (F). Spirogyra sp. 4, (G). Spirogyra sp. 5, (H,I). Microcystis aeruginosa (Kützing) Kützing 1846 [35] scale bar 20 µm, (J). Cymbella mexicana (Ehrenberg) Cleve 1894 [37], (K). Tribonema sp., (L). Gloeotrichia echinulata P.G. Richter 1894 [40], (M). Quadrigula chodatii (Tanner-Füllemann) G.M.Smith 1920 [39]. Scale bars equal 10 µm if not otherwise indicated.

Figure 4.
Algae found in the Freda Haffner Kettlehole State Preserve, Dickinson County, Iowa. (A,C). Glaucocystis nostochinearum Itzigsohn 1866 [40], (B). Phacus acuminatus A.Stokes 1885 [35], (D). Xanthidium uncinatum (Brébisson ex Ralfs) Stastny, Skaloud and Neustupa 2013 [41], (E–G). Pleurotaenium cf. ehrenbergii (Ralphs) De Bary 1858 [26,41], (H). Monomorphina sp., (I). Closterium parvulum Nägeli 1849 [42], (J). Closterium cf. dianae var. minus Hieronymus 1895 [35], (K). Closterium gracile Brébisson ex Ralfs 1848 [35], (L). Phacus orbicularis Hübner 1886 [25], (M,N). Trachelomonas armata (Ehrenberg) F.Stein 1878 [35], (N). flagellate cell leaving broken lorica, (O). Eunotia bilunaris (Ehrenberg) Schaarschmidt 1880 [26], epiphytic, (P). Cystodinium cf. cornifax (A.J.Schilling) G.A.Klebs 1912 [35]. Scale bars equal 10 µm, except in F scale bar equal to 20 µm.
Figure 4.
Algae found in the Freda Haffner Kettlehole State Preserve, Dickinson County, Iowa. (A,C). Glaucocystis nostochinearum Itzigsohn 1866 [40], (B). Phacus acuminatus A.Stokes 1885 [35], (D). Xanthidium uncinatum (Brébisson ex Ralfs) Stastny, Skaloud and Neustupa 2013 [41], (E–G). Pleurotaenium cf. ehrenbergii (Ralphs) De Bary 1858 [26,41], (H). Monomorphina sp., (I). Closterium parvulum Nägeli 1849 [42], (J). Closterium cf. dianae var. minus Hieronymus 1895 [35], (K). Closterium gracile Brébisson ex Ralfs 1848 [35], (L). Phacus orbicularis Hübner 1886 [25], (M,N). Trachelomonas armata (Ehrenberg) F.Stein 1878 [35], (N). flagellate cell leaving broken lorica, (O). Eunotia bilunaris (Ehrenberg) Schaarschmidt 1880 [26], epiphytic, (P). Cystodinium cf. cornifax (A.J.Schilling) G.A.Klebs 1912 [35]. Scale bars equal 10 µm, except in F scale bar equal to 20 µm.

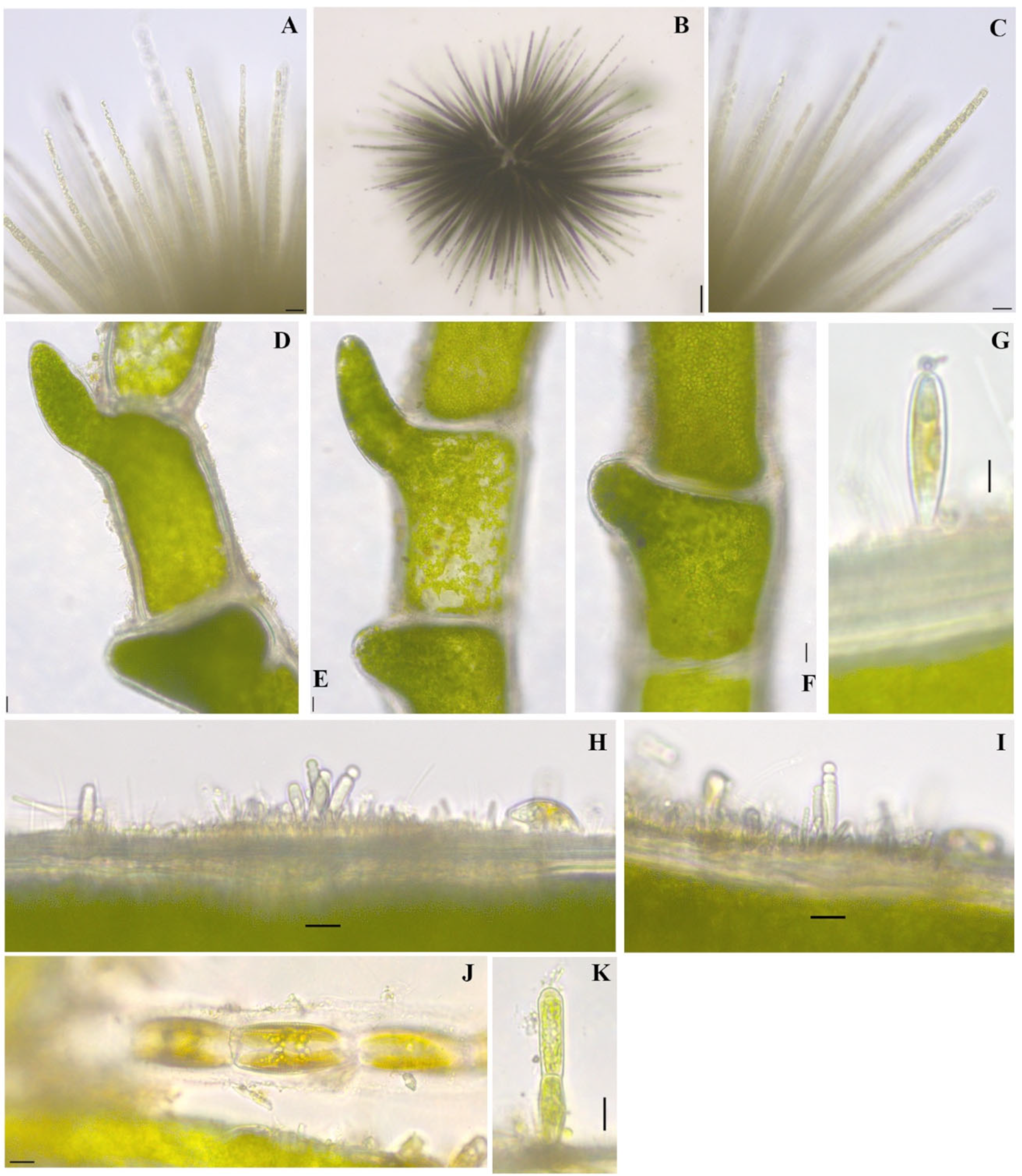
Figure 5.
Algae found in Millers Bay, Dickinson County, Iowa. (A–C). Gloeotrichia echinulata P.G.Richter 1894 [40], (B). entire colony, scale bar 40 µm, (A,C). heterodiametric trichome tips not thinning to a hair (D–F). Cladophora glomerata (Linnaeus) Kützing 1843 [40], short branches, scale bar 20 µm, (G). Gomphonema sp., (H,I). Chamaesiphon incrustans Grunow 1865 [25], (J). Encyonema sp. in tube, (K). Oedogonium sp., initial 2-cell filament, epiphytic on Cladophora sp. Scale bars equal 10 µm if not otherwise indicated.
Figure 5.
Algae found in Millers Bay, Dickinson County, Iowa. (A–C). Gloeotrichia echinulata P.G.Richter 1894 [40], (B). entire colony, scale bar 40 µm, (A,C). heterodiametric trichome tips not thinning to a hair (D–F). Cladophora glomerata (Linnaeus) Kützing 1843 [40], short branches, scale bar 20 µm, (G). Gomphonema sp., (H,I). Chamaesiphon incrustans Grunow 1865 [25], (J). Encyonema sp. in tube, (K). Oedogonium sp., initial 2-cell filament, epiphytic on Cladophora sp. Scale bars equal 10 µm if not otherwise indicated.

Figure 6.
Algae found in East Okoboji Lake, Dickinson County, Iowa. (A,B). Draparnaldia acuta (C.Agardh) Kützing 1845 [40] scale bars 40 µm, (C). Cladophora glomerata (Linnaeus) Kützing 1843 [40] with epiphytic Cocconeis pediculus Ehrenberg 1838 [43] and Gomphonema sp. scale bar 20 µm.

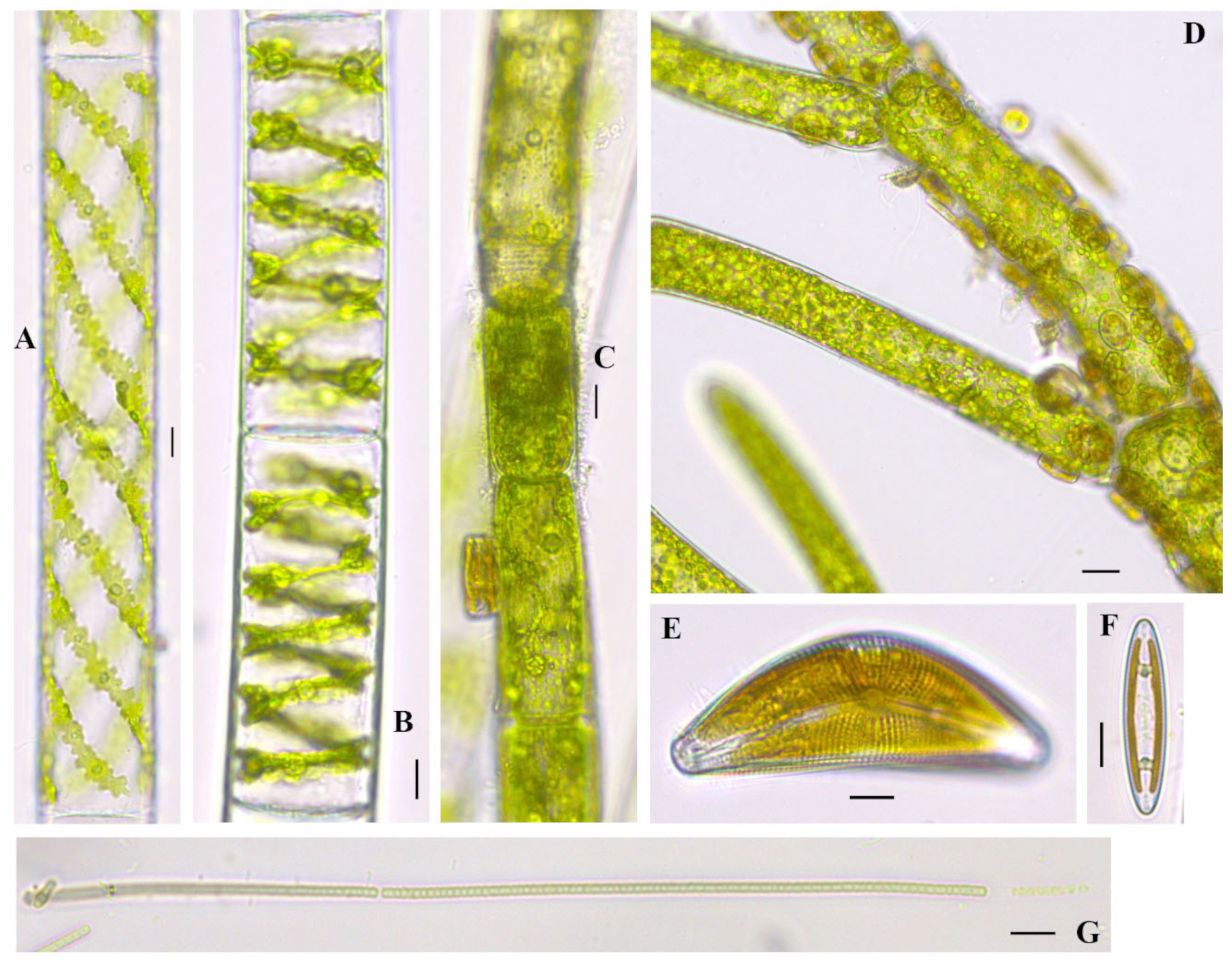
Figure 7.
Algae found in West Okoboji Lake, Dickinson County, Iowa. (A). Spirogyra sp., (B). Spirogyra cf. borgeana Transeau 1916 [35,44], (C). Oedogonium sp. 2, (D). Cladophora sp. with epiphytic Cocconeis sp. scale bar 20 µm, (E). Cymbella mexicana (Ehrenberg) Cleve 1894 [37], (F). Navicula sp., (G). Leptolyngbya sp. Scale bars equal 10 µm if not otherwise indicated.
Figure 7.
Algae found in West Okoboji Lake, Dickinson County, Iowa. (A). Spirogyra sp., (B). Spirogyra cf. borgeana Transeau 1916 [35,44], (C). Oedogonium sp. 2, (D). Cladophora sp. with epiphytic Cocconeis sp. scale bar 20 µm, (E). Cymbella mexicana (Ehrenberg) Cleve 1894 [37], (F). Navicula sp., (G). Leptolyngbya sp. Scale bars equal 10 µm if not otherwise indicated.

Figure 8.
Algae found in Big Spirit Lake, Dickinson County, Iowa. (A–C). Stephanodiscus reimeri Theriot and Stoermer 1992 [45], (D,E). Chaetophoropsis elegans (Roth) B.Wen Liu, Qian Xiong, X.Dong Liu, Z.Yu Hu & G.Xiang Liu 2019 [26,40], scale bar 20 µm, E. scale bar 40 µm, (F,G). Aulacoseira sp. 1, (H). Micractinium sp., (I). Coelomoron cf. pusillum (Van Goor) Komárek 1988 [46], (J–M). Klebsormidium cf. mucosum (J.B.Petersen) Lokhorst 1985 [35] scale bar 20 µm, (N). Characiopsis sp. epiphytic on K. cf. mucosum, (O). Aulacoseira sp. 2, (P,Q). Diatoma vulgaris, chain, (R). Monactinus simplex (Meyen) Corda 1839 [26,39]. Scale bars equal 10 µm if not otherwise indicated.
Figure 8.
Algae found in Big Spirit Lake, Dickinson County, Iowa. (A–C). Stephanodiscus reimeri Theriot and Stoermer 1992 [45], (D,E). Chaetophoropsis elegans (Roth) B.Wen Liu, Qian Xiong, X.Dong Liu, Z.Yu Hu & G.Xiang Liu 2019 [26,40], scale bar 20 µm, E. scale bar 40 µm, (F,G). Aulacoseira sp. 1, (H). Micractinium sp., (I). Coelomoron cf. pusillum (Van Goor) Komárek 1988 [46], (J–M). Klebsormidium cf. mucosum (J.B.Petersen) Lokhorst 1985 [35] scale bar 20 µm, (N). Characiopsis sp. epiphytic on K. cf. mucosum, (O). Aulacoseira sp. 2, (P,Q). Diatoma vulgaris, chain, (R). Monactinus simplex (Meyen) Corda 1839 [26,39]. Scale bars equal 10 µm if not otherwise indicated.

Figure 9.
Algae found in Upper Gar Lake, Dickinson County, Iowa. (A,B). Hydrodictyon reticulatum (Linnaeus) Bory 1824 [40]. (A). scale bar 20 µm, (B). scale bar 40 µm, (C). Oedogonium sp., scale bar 10 µm.
Figure 9.
Algae found in Upper Gar Lake, Dickinson County, Iowa. (A,B). Hydrodictyon reticulatum (Linnaeus) Bory 1824 [40]. (A). scale bar 20 µm, (B). scale bar 40 µm, (C). Oedogonium sp., scale bar 10 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
GPS coordinates of algal sampling locations for summer 2017 sites.
| Site | Latitude | Longitude |
|---|---|---|
| Center Lake | 43.406908° | −95.140512° |
| Emerson Bay | 43.356369° | −95.174774° |
| Freda Haffner Kettlehole State Preserve | 43.346369° | −95.221853° |
| Millers Bay | 43.378333° | −95.181111° |
| East Okoboji Lake | 43.376111° | −95.126111° |
| West Okoboji Lake | 43.379920° | −95.141378° |
| Big Spirit Lake | 43.445810° | −95.102304° |
| Upper Gar Lake | 43.364444° | −95.125000° |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Johnson, K.M.; Manoylov, K.M. Living Algal Community Voucher and Taxonomy in Dickinson County Iowa: A Case for Collection-Based Research. Diversity 2024, 16, 21. https://doi.org/10.3390/d16010021
AMA Style
Johnson KM, Manoylov KM. Living Algal Community Voucher and Taxonomy in Dickinson County Iowa: A Case for Collection-Based Research. Diversity. 2024; 16(1):21. https://doi.org/10.3390/d16010021
Chicago/Turabian StyleJohnson, Katherine M., and Kalina M. Manoylov. 2024. "Living Algal Community Voucher and Taxonomy in Dickinson County Iowa: A Case for Collection-Based Research" Diversity 16, no. 1: 21. https://doi.org/10.3390/d16010021
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.