
Nuclear Binding Protein 2/Nesfatin-1 Affects Trophoblast Cell Fusion during Placental Development via the EGFR-PLCG1-CAMK4 Pathway
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
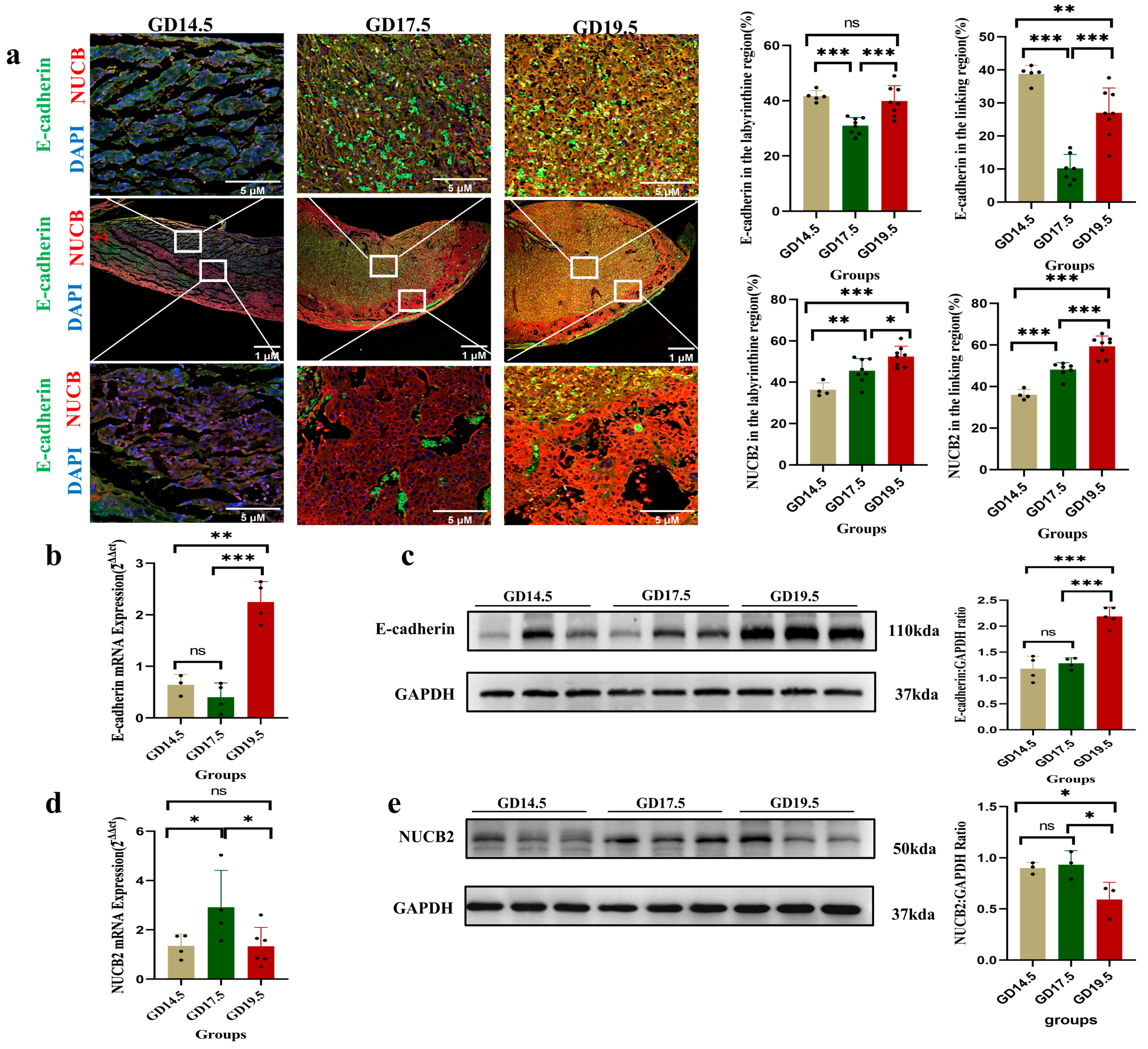
2.1. NUCB2 Expression at Different Stages of Gestation in the Rat Placenta
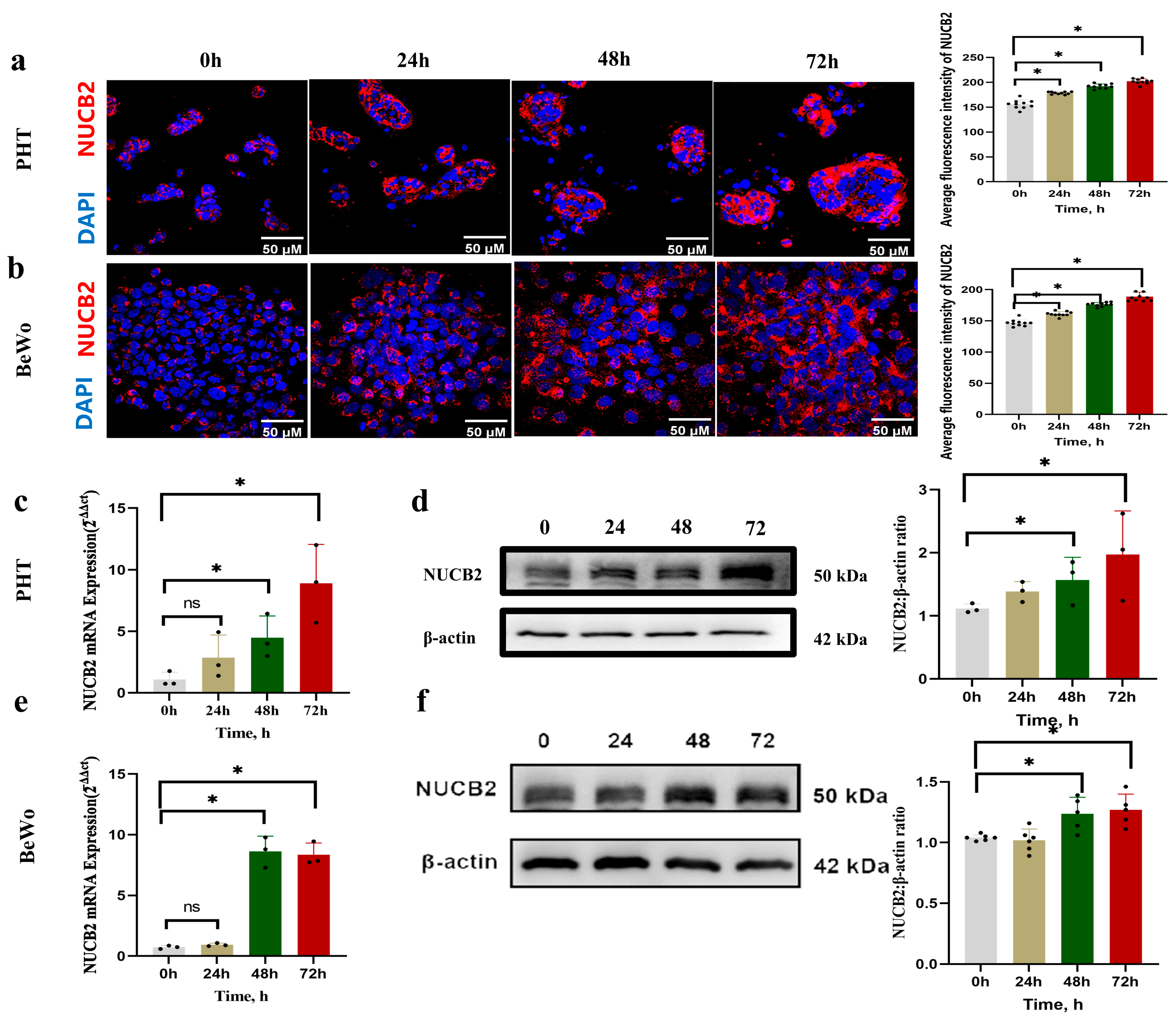
2.2. NUCB2 Expression during Spontaneous Syncytialization of Primary Human Trophoblasts and Forskolin-Induced BeWo Syncytialization
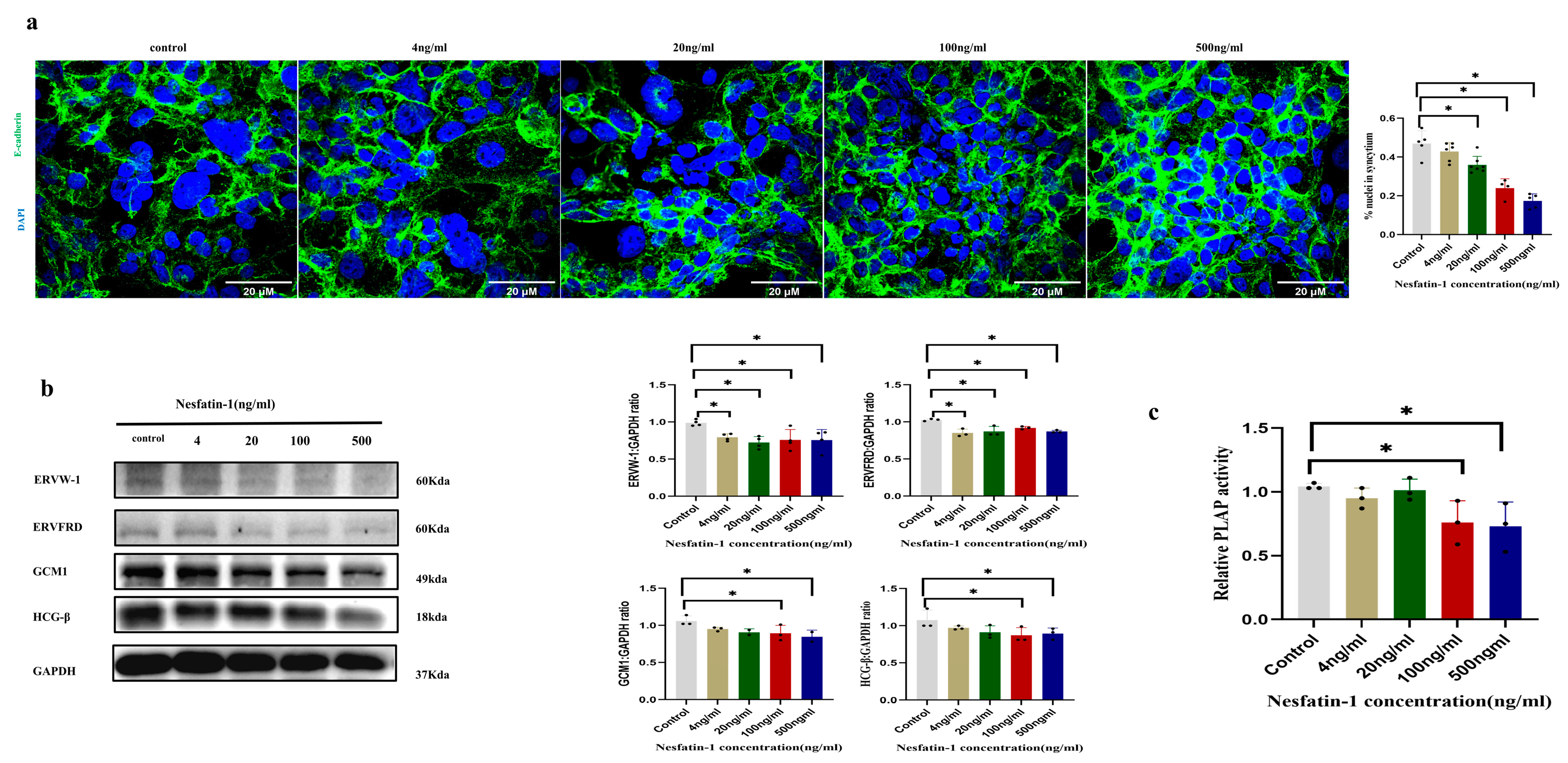
2.3. Effect of Exogenous Nesfatin-1 on Forskolin-Induced Fusion of BeWo Syncytized Trophoblast Cells
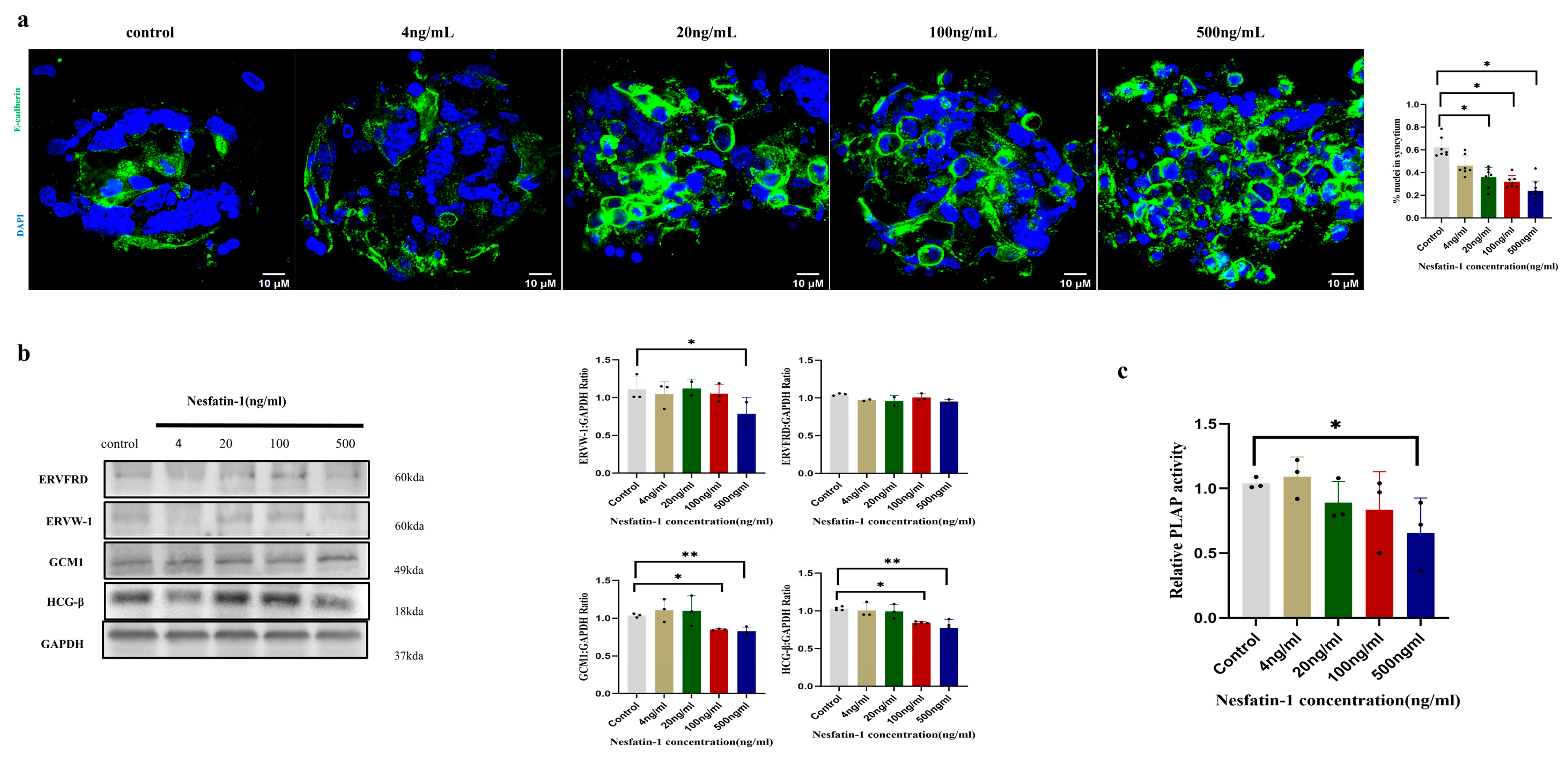
2.4. Effect of Exogenous Nesfatin-1 on Spontaneous Syncytial Processes in Primary Human Trophoblast Cells
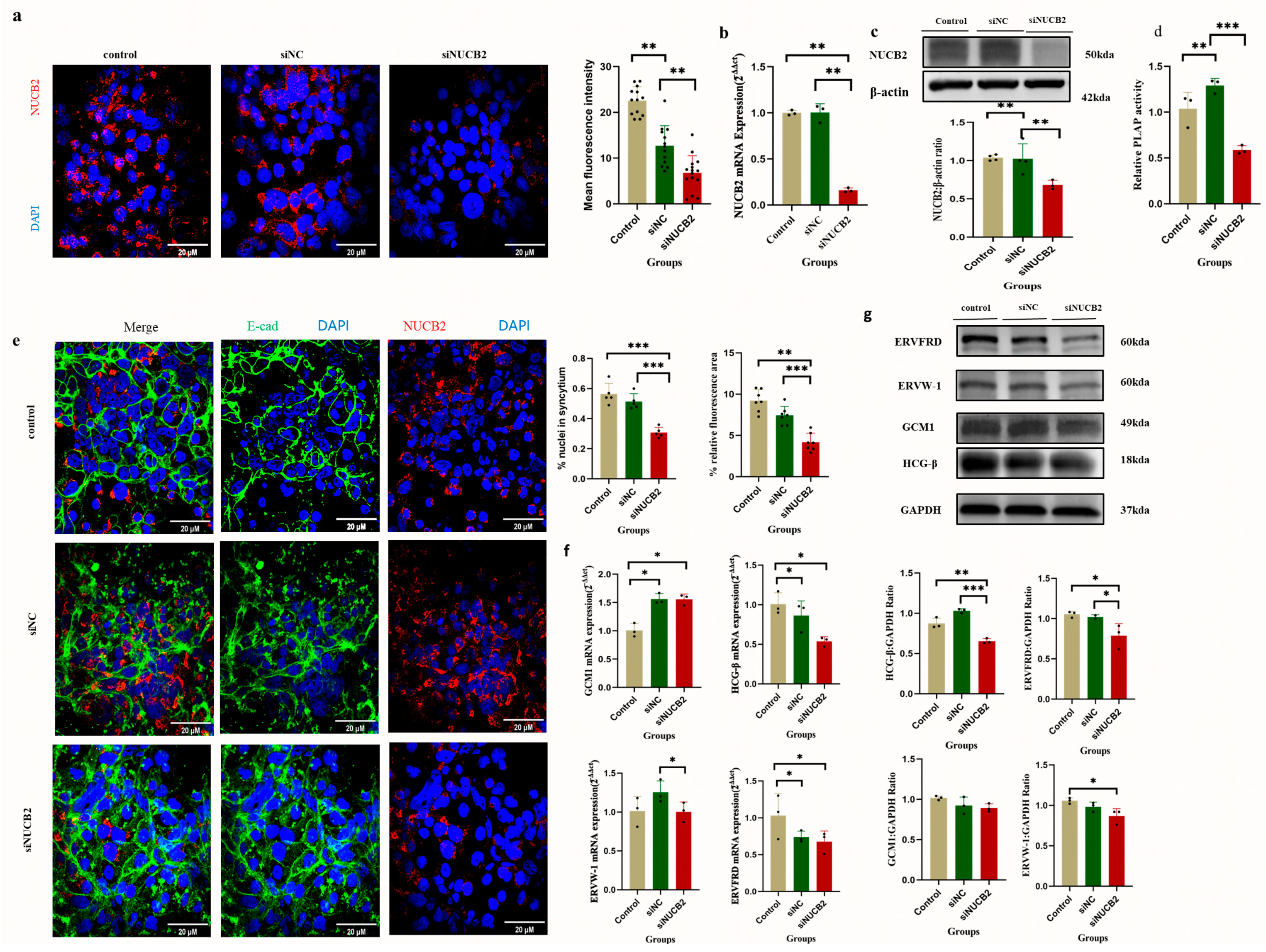
2.5. Effect of Knockdown of NUCB2 on the Extent of Trophoblast Fusion in BeWo Cells
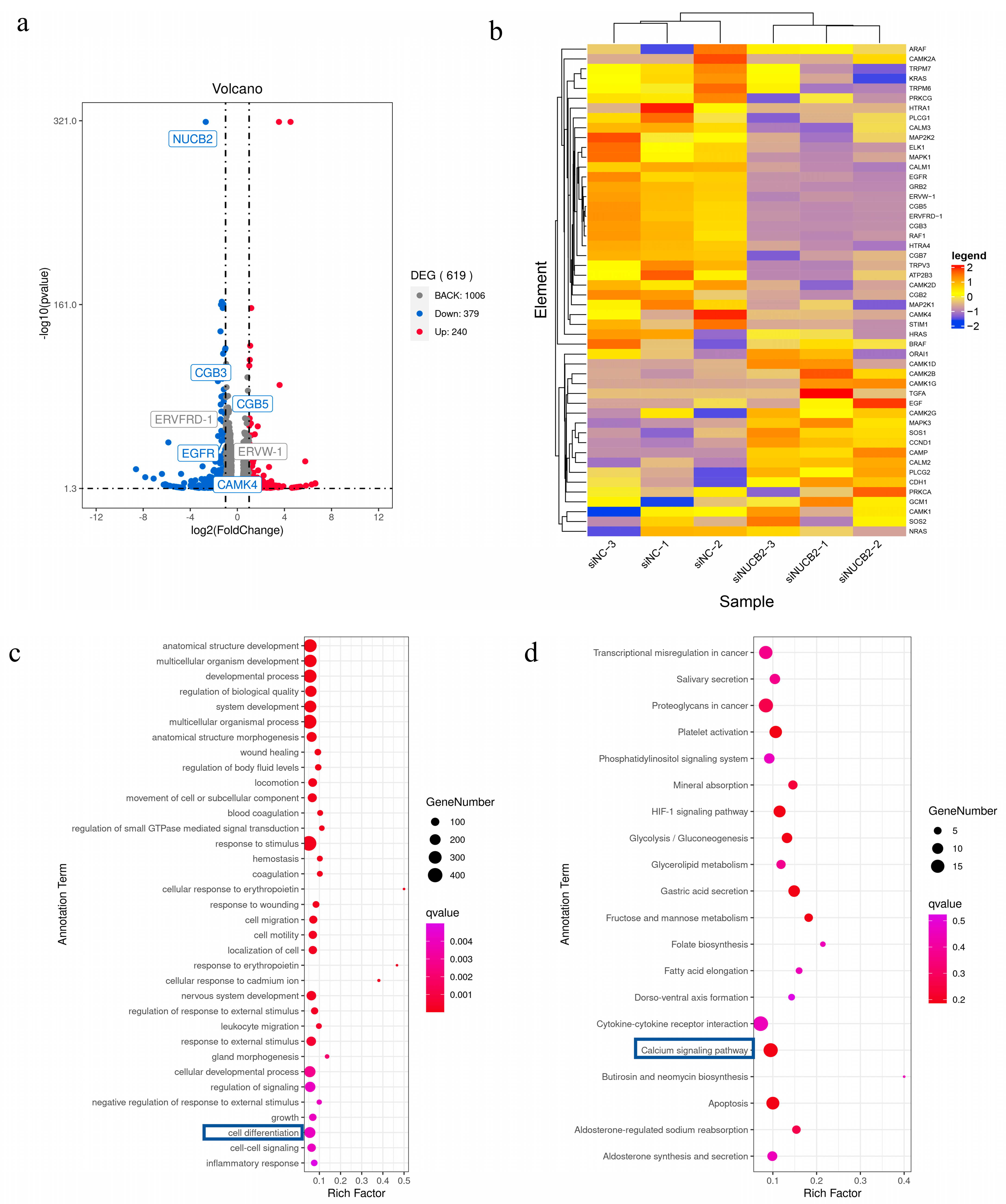
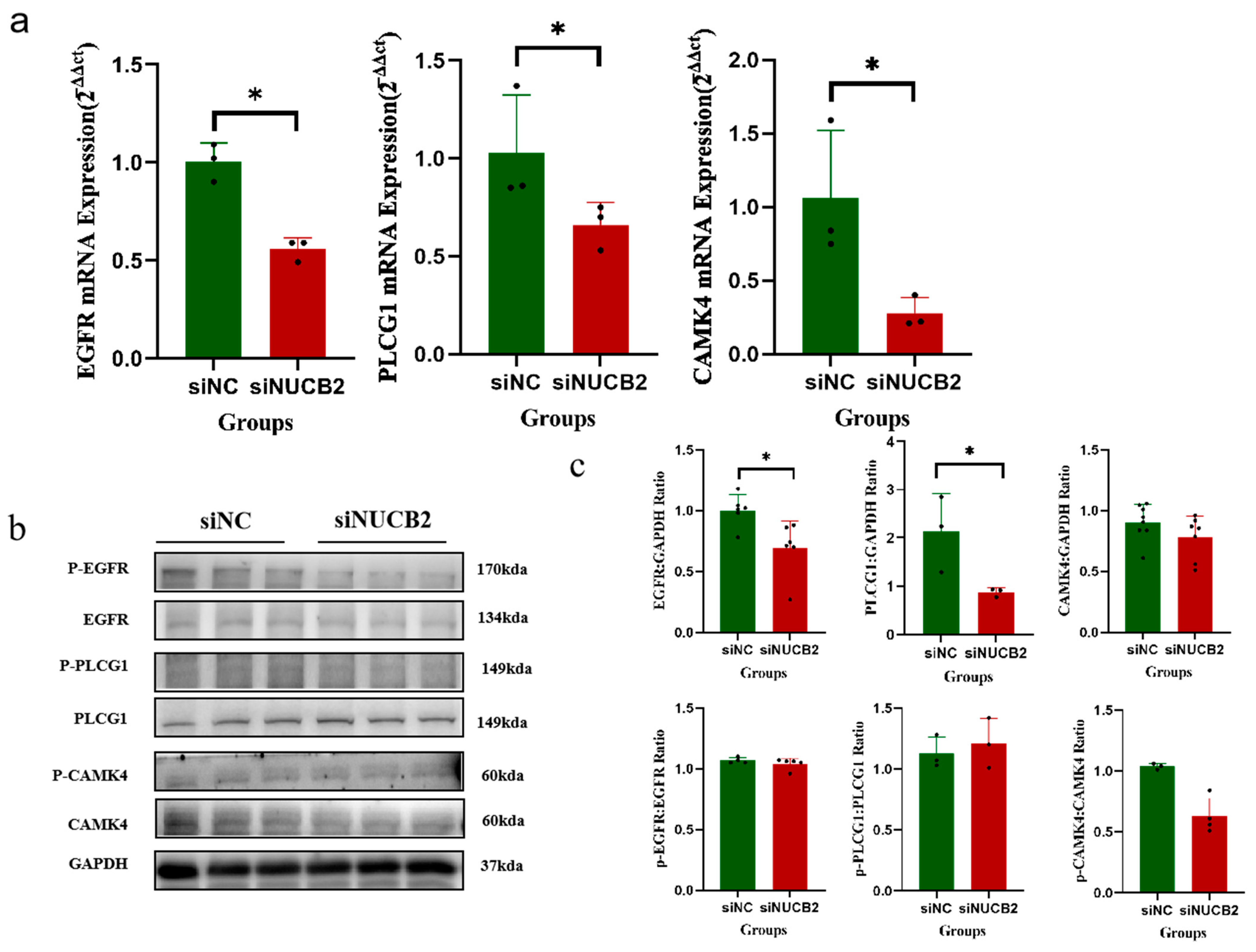
2.6. NUCB2/Nesfatin-1 May Inhibit Trophoblast Differentiation and Fusion through the EGFR-PLCG1-CAMK4 Pathway
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.2. Isolation of Primary Human Trophoblast Cells
4.3. BeWo Cell Culture and Fusion Assay
4.4. Lentiviral Vector Infection for NUCB2 Knockdown
4.5. Placental Alkaline Phosphatase (PLAP) Activity Assay
4.6. Total RNA Extraction and Real-Time PCR
4.7. Western Blotting Analysis
4.8. Immunofluorescence Staining and Confocal Microscopy
4.9. RNA Sequencing
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Zheng, R.; Wang, R.; Lu, X.; Zhu, C.; Lin, H.Y.; Wang, H.; Yu, X.; Fu, J. Involvement of nephrin in human placental trophoblast syncytialization. Reproduction 2015, 149, 339–346. [Google Scholar] [CrossRef]
- Benaitreau, D.; Dos Santos, E.; Leneveu, M.C.; De Mazancourt, P.; Pecquery, R.; Dieudonné, M.N. Adiponectin promotes syncytialisation of BeWo cell line and primary trophoblast cells. Reprod. Biol. Endocrinol. 2010, 8, 128. [Google Scholar] [CrossRef]
- Morosin, S.K.; Delforce, S.J.; Corbisier de Meaultsart, C.; Lumbers, E.R.; Pringle, K.G. FURIN and placental syncytialisation: A cautionary tale. Cell Death Dis. 2021, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, M.; Wang, Y.; Lim, R.; Palmer, K.; Nie, G. HtrA4 is up-regulated during trophoblast syncytialization and BeWo cells fail to syncytialize without HtrA4. Sci. Rep. 2021, 11, 14363. [Google Scholar] [CrossRef] [PubMed]
- Morosin, S.K.; Delforce, S.J.; Lumbers, E.R.; Pringle, K.G. The (pro)renin receptor (ATP6AP2) does not play a role in syncytialisation of term human primary trophoblast cells. Placenta 2020, 97, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Weedon-Fekjær, M.S.; Taskén, K. Review: Spatiotemporal dynamics of hCG/cAMP signaling and regulation of placental function. Placenta 2012, 33, S87–S91. [Google Scholar] [CrossRef]
- Zani, A.; Zhang, L.; McMichael, T.M.; Kenney, A.D.; Chemudupati, M.; Kwiek, J.J.; Liu, S.L.; Yount, J.S. Interferon-induced transmembrane proteins inhibit cell fusion mediated by trophoblast syncytins. J. Biol. Chem. 2019, 294, 19844–19851. [Google Scholar] [CrossRef]
- Drewlo, S.; Johnson, E.; Kilburn, B.A.; Kadam, L.; Armistead, B.; Kohan-Ghadr, H.R. Irisin induces trophoblast differentiation via AMPK activation in the human placenta. J. Cell. Physiol. 2020, 235, 7146–7158. [Google Scholar] [CrossRef]
- Barnikol-Watanabe, S.; Gross, N.A.; Götz, H.; Henkel, T.; Karabinos, A.; Kratzin, H.; Barnikol, H.U.; Hilschmann, N. Human protein NEFA, a novel DNA binding/EF-hand/leucine zipper protein. Molecular cloning and sequence analysis of the cDNA, isolation and characterization of the protein. Biol. Chem. Hoppe Seyler 1994, 375, 497–512. [Google Scholar] [CrossRef]
- Kanai, Y.; Tanuma, S. Purification of a novel B cell growth and differentiation factor associated with lupus syndrome. Immunol. Lett. 1992, 32, 43–48. [Google Scholar] [CrossRef]
- Foo, K.S.; Brauner, H.; Ostenson, C.G.; Broberger, C. Nucleobindin-2/nesfatin in the endocrine pancreas: Distribution and relationship to glycaemic state. J. Endocrinol. 2010, 204, 255–263. [Google Scholar] [CrossRef]
- Gonzalez, R.; Tiwari, A.; Unniappan, S. Pancreatic beta cells colocalize insulin and pronesfatin immunoreactivity in rodents. Biochem. Biophys. Res. Commun. 2009, 381, 643–648. [Google Scholar] [CrossRef]
- Stengel, A.; Goebel, M.; Yakubov, I.; Wang, L.; Witcher, D.; Coskun, T.; Taché, Y.; Sachs, G.; Lambrecht, N.W. Identification and characterization of nesfatin-1 immunoreactivity in endocrine cell types of the rat gastric oxyntic mucosa. Endocrinology 2009, 150, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Legg-St Pierre, C.B.; Mackova, M.; Miskiewicz, E.I.; Hemmings, D.G.; Unniappan, S.; MacPhee, D.J. Insulinotropic nucleobindin-2/nesfatin-1 is dynamically expressed in the haemochorial mouse and human placenta. Reprod. Fertil. Dev. 2018, 30, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Y.; Wang, Y.; Li, J.; Liu, R.; Liu, H. Decreased levels of serum nesfatin-1 in patients with preeclampsia. Biomarkers 2014, 19, 402–406. [Google Scholar] [CrossRef]
- Skheel Aljebory, H.D.; Abdalkareem, Z.M.; Jasim, A.M. Correlation between serum nesfatin-1 level and severity of preeclampsia—A case control study. J. Pak. Med. Assoc. 2021, 71 (Suppl. 8), S49–S52. [Google Scholar]
- Serin, S.; Bakacak, M.; Ercan, Ö.; Köstü, B.; Avci, F.; Arıkan, D.; Kıran, G. The evaluation of Nesfatin-1 levels in patients with and without intrauterine growth restriction. J. Matern. Fetal Neonatal Med. 2016, 29, 1409–1413. [Google Scholar] [CrossRef]
- Li, T.; Wei, S.; Fan, C.; Tang, D.; Luo, D. Nesfatin-1 Promotes Proliferation, Migration and Invasion of HTR-8/SVneo Trophoblast Cells and Inhibits Oxidative Stress via Activation of PI3K/AKT/mTOR and AKT/GSK3β Pathway. Reprod. Sci. 2021, 28, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Li, Y.; Sun, H.; Lu, X.; Sun, B.F.; Wang, R.; Cui, L.; Zhu, C.; Lin, H.Y.; Wang, H. Deep RNA sequencing analysis of syncytialization-related genes during BeWo cell fusion. Reproduction 2016, 153, 35–48. [Google Scholar] [CrossRef]
- Tagaya, Y.; Miura, A.; Okada, S.; Ohshima, K.; Mori, M. Nucleobindin-2 is a positive modulator of EGF-dependent signals leading to enhancement of cell growth and suppression of adipocyte differentiation. Endocrinology 2012, 153, 3308–3319. [Google Scholar] [CrossRef]
- Morrish, D.W.; Dakour, J.; Li, H. Functional regulation of human trophoblast differentiation. J. Reprod. Immunol. 1998, 39, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, K.; Kurachi, H.; Adachi, H.; Morishige, K.I.; Adachi, K.; Imai, T.; Miyake, A. Involvement of epidermal growth factor (EGF)/EGF receptor autocrine and paracrine mechanism in human trophoblast cells: Functional differentiation in vitro. J. Endocrinol. 1994, 143, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.S.; Gupta, S.K. Relevance of the NR4A sub-family of nuclear orphan receptors in trophoblastic BeWo cell differentiation. Cell. Mol. Biol. Lett. 2017, 22, 15. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Zhang, Y.; Li, W.; Liu, Y.; Duan, C.; Ma, J.; Wang, Y.; Zhuang, R.; Ding, Y. Identification of CaMK4 as a sex-difference-related gene in knee osteoarthritis. Ann. Transl. Med. 2023, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Coutifaris, C.; Kao, L.C.; Sehdev, H.M.; Chin, U.; Babalola, G.O.; Blaschuk, O.W.; Strauss, J.F., 3rd. E-cadherin expression during the differentiation of human trophoblasts. Development 1991, 113, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, N.; Ikezaki, M.; Nishitsuji, K.; Yamamoto, M.; Matsuzaki, I.; Kato, N.; Takaoka, N.; Taniguchi, M.; Murata, S.I.; Ino, K.; et al. Extracellularly Released Calreticulin Induced by Endoplasmic Reticulum Stress Impairs Syncytialization of Cytotrophoblast Model BeWo Cells. Cells 2021, 10, 1305. [Google Scholar] [CrossRef] [PubMed]
- Metz, J. On the developing rat placenta. I. Differentiation and junctional alterations of labyrinthine layers II and III. Anat. Embryol. 1980, 159, 289–305. [Google Scholar] [CrossRef]
- Garcés, M.F.; Poveda, N.E.; Sanchez, E.; Sánchez, Á.Y.; Bravo, S.B.; Vázquez, M.J.; Diéguez, C.; Nogueiras, R.; Caminos, J.E. Regulation of NucB2NUCB2/Nesfatin-1 throughout rat pregnancy. Physiol. Behav. 2014, 133, 216–222. [Google Scholar] [CrossRef]
- Garcia-Galiano, D.; Tena-Sempere, M. Emerging roles of NUCB2NUCB2/nesfatin-1 in the metabolic control of reproduction. Curr. Pharm. Des. 2013, 19, 6966–6972. [Google Scholar] [CrossRef]
- Huppertz, B.; Bartz, C.; Kokozidou, M. Trophoblast fusion: Fusogenic proteins, syncytins and ADAMs, and other prerequisites for syncytial fusion. Micron 2006, 37, 509–517. [Google Scholar] [CrossRef]
- Getsios, S.; Chen, G.T.; MacCalman, C.D. Regulation of beta-catenin mRNA and protein levels in human villous cytotrophoblasts undergoing aggregation and fusion in vitro: Correlation with E-cadherin expression. J. Reprod. Fertil. 2000, 119, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Dore, R.; Levata, L.; Lehnert, H.; Schulz, C. Nesfatin-1: Functions and physiology of a novel regulatory peptide. J. Endocrinol. 2017, 232, R45–R65. [Google Scholar] [CrossRef] [PubMed]
- Leitner, K.; Szlauer, R.; Ellinger, I.; Ellinger, A.; Zimmer, K.P.; Fuchs, R. Placental alkaline phosphatase expression at the apical and basal plasma membrane in term villous trophoblasts. J. Histochem. Cytochem. 2001, 49, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Orendi, K.; Gauster, M.; Moser, G.; Meiri, H.; Huppertz, B. The choriocarcinoma cell line BeWo: Syncytial fusion and expression of syncytium-specific proteins. Reproduction 2010, 140, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Malassiné, A.; Blaise, S.; Handschuh, K.; Lalucque, H.; Dupressoir, A.; Evain-Brion, D.; Heidmann, T. Expression of the fusogenic HERV-FRD Env glycoprotein (syncytin 2) in human placenta is restricted to villous cytotrophoblastic cells. Placenta 2007, 28, 185–191. [Google Scholar] [CrossRef]
- Holder, B.S.; Tower, C.L.; Forbes, K.; Mulla, M.J.; Aplin, J.D.; Abrahams, V.M. Immune cell activation by trophoblast-derived microvesicles is mediated by syncytin 1. Immunology 2012, 136, 184–191. [Google Scholar] [CrossRef]
- Lokossou, A.G.; Toudic, C.; Nguyen, P.T.; Elisseeff, X.; Vargas, A.; Rassart, É.; Lafond, J.; Leduc, L.; Bourgault, S.; Gilbert, C.; et al. Endogenous retrovirus-encoded Syncytin-2 contributes to exosome-mediated immunosuppression of T cells. Biol. Reprod. 2020, 102, 185–198. [Google Scholar] [CrossRef]
- Mincheva-Nilsson, L.; Baranov, V. Placenta-derived exosomes and syncytiotrophoblast microparticles and their role in human reproduction: Immune modulation for pregnancy success. Am. J. Reprod. Immunol. 2014, 72, 440–457. [Google Scholar] [CrossRef]
- Schust, D.J.; Bonney, E.A.; Sugimoto, J.; Ezashi, T.; Roberts, R.M.; Choi, S.; Zhou, J. The Immunology of Syncytialized Trophoblast. Int. J. Mol. Sci. 2021, 22, 1767. [Google Scholar] [CrossRef] [PubMed]
- Jeyarajah, M.J.; Jaju Bhattad, G.; Kelly, R.D.; Baines, K.J.; Jaremek, A.; Yang, F.P.; Okae, H.; Arima, T.; Dumeaux, V.; Renaud, S.J. The multifaceted role of GCM1 during trophoblast differentiation in the human placenta. Proc. Natl. Acad. Sci. USA 2022, 119, e2203071119. [Google Scholar] [CrossRef] [PubMed]
- Boutsikou, T.; Briana, D.D.; Boutsikou, M.; Kafalidis, G.; Piatopoulou, D.; Baka, S.; Hassiakos, D.; Gourgiotis, D.; Malamitsi-Puchner, A. Cord blood nesfatin-1 in large for gestational age pregnancies. Cytokine 2013, 61, 591–594. [Google Scholar] [CrossRef]
- Ruebner, M.; Strissel, P.L.; Langbein, M.; Fahlbusch, F.; Wachter, D.L.; Faschingbauer, F.; Beckmann, M.W.; Strick, R. Impaired cell fusion and differentiation in placentae from patients with intrauterine growth restriction correlate with reduced levels of HERV envelope genes. J. Mol. Med. 2010, 88, 1143–1156. [Google Scholar] [CrossRef]
- Newhouse, S.M.; Davidge, S.T.; Winkler-Lowen, B.; Demianczuk, N.; Guilbert, L.J. In vitro differentiation of villous trophoblasts from pregnancies complicated by intrauterine growth restriction with and without pre-eclampsia. Placenta 2007, 28, 999–1003. [Google Scholar] [CrossRef]
- Ticiani, E.; Villegas, J.A.; Murga-Zamalloa, C.; Veiga-Lopez, A. Binding sites in the epidermal growth factor receptor are responsible for bisphenol S effects on trophoblast cell invasion. Chemosphere 2023, 318, 137960. [Google Scholar] [CrossRef] [PubMed]
- Whigham, C.A.; Hastie, R.; Hannan, N.J.; Brownfoot, F.; Pritchard, N.; Cannon, P.; Nguyen, T.V.; Kandel, M.; Masci, J.; Tong, S.; et al. Placental growth factor is negatively regulated by epidermal growth factor receptor (EGFR) signaling. Placenta 2021, 114, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Suh, P.G.; Lee, Y.J.; Shin, K.J.; Cocco, L.; Chae, Y.C. PLCγ1: Potential arbitrator of cancer progression. Adv. Biol. Regul. 2018, 67, 179–189. [Google Scholar] [CrossRef]
- Li, H.Q.; Fan, J.J.; Li, X.H.; Bao, D. MiR-507 inhibits the growth and invasion of trophoblasts by targeting CAMK4. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5856–5862. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, L.; Shao, Y.; Du, R.; Li, L.; Shi, X.; Bai, Y. Calcium/calmodulin dependent protein kinase IV in trophoblast cells under insulin resistance: Functional and metabolomic analyses. Mol. Med. 2023, 29, 82. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Schust, D.J. Isolation, purification and in vitro differentiation of cytotrophoblast cells from human term placenta. Reprod. Biol. Endocrinol. 2015, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Kliman, H.J.; Nestler, J.E.; Sermasi, E.; Sanger, J.M.; Strauss, J.F., 3rd. Purification, characterization, and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 1986, 118, 1567–1582. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, Q.; Zhu, Y.; Zhang, Y.; Hu, Z.; Wei, Y.; Chen, Z.; Jiang, X.; Cai, X.; Yu, H. Nuclear Binding Protein 2/Nesfatin-1 Affects Trophoblast Cell Fusion during Placental Development via the EGFR-PLCG1-CAMK4 Pathway. Int. J. Mol. Sci. 2024, 25, 1925. https://doi.org/10.3390/ijms25031925
Dang Q, Zhu Y, Zhang Y, Hu Z, Wei Y, Chen Z, Jiang X, Cai X, Yu H. Nuclear Binding Protein 2/Nesfatin-1 Affects Trophoblast Cell Fusion during Placental Development via the EGFR-PLCG1-CAMK4 Pathway. International Journal of Molecular Sciences. 2024; 25(3):1925. https://doi.org/10.3390/ijms25031925
Chicago/Turabian StyleDang, Qinyu, Yandi Zhu, Yadi Zhang, Zhuo Hu, Yuchen Wei, Zhaoyang Chen, Xinyin Jiang, Xiaxia Cai, and Huanling Yu. 2024. "Nuclear Binding Protein 2/Nesfatin-1 Affects Trophoblast Cell Fusion during Placental Development via the EGFR-PLCG1-CAMK4 Pathway" International Journal of Molecular Sciences 25, no. 3: 1925. https://doi.org/10.3390/ijms25031925