


The Role of MicroRNA, Long Non-Coding RNA and Circular RNA in the Pathogenesis of Polycystic Ovary Syndrome: A Literature Review
 ,
,
Abstract
:1. Introduction to Polycystic Ovary Syndrome
2. Methodology
3. Introduction to microRNA
4. The Role of miRNA in the Pathogenesis of Common PCOS Complications
4.1. Infertility
4.2. Insulin Resistance

{kind=link}
{kind=link}
| miRNA(s) | Study Finding | References |
|---|---|---|
| miRNA-93 | Upregulated in PCOS, resulting in the downregulation of GLUT4 receptors in adipose tissue. | [47] |
| miRNA-146 | Role in insulin resistance and inflammatory modulation. | [48] |
| miRNA-133a-3p | Upregulation resulting in the inhibition of the PI3K/AKT signaling pathway that is important in insulin activity. | [48] |
| miR-222 miR-221 | Altered expression with the administration of metformin in T2DM patients, leading to increased glucose metabolism and transport. | [49] |
| miR-26a | Altered expression with the administration of metformin for pancreatic stem cell markers. | [54] |
| miR-6763 miR-33 miR-155-5p miR-6356 miR-197 miR-875-5P miR-1197-3p | Altered expression with dipeptidyl peptidase-4 (DPP-4) inhibitors and glucagon-like peptide 1 agonist receptor agonist (GLP-1 RA), leading to increased glucose metabolism and transport. | [50] |
| miR-143-3p | Negatively regulates glycolysis. | [53] |
| miR-155-5p | Positively regulates glycolysis. | [53] |
4.3. Cardiovascular Complications
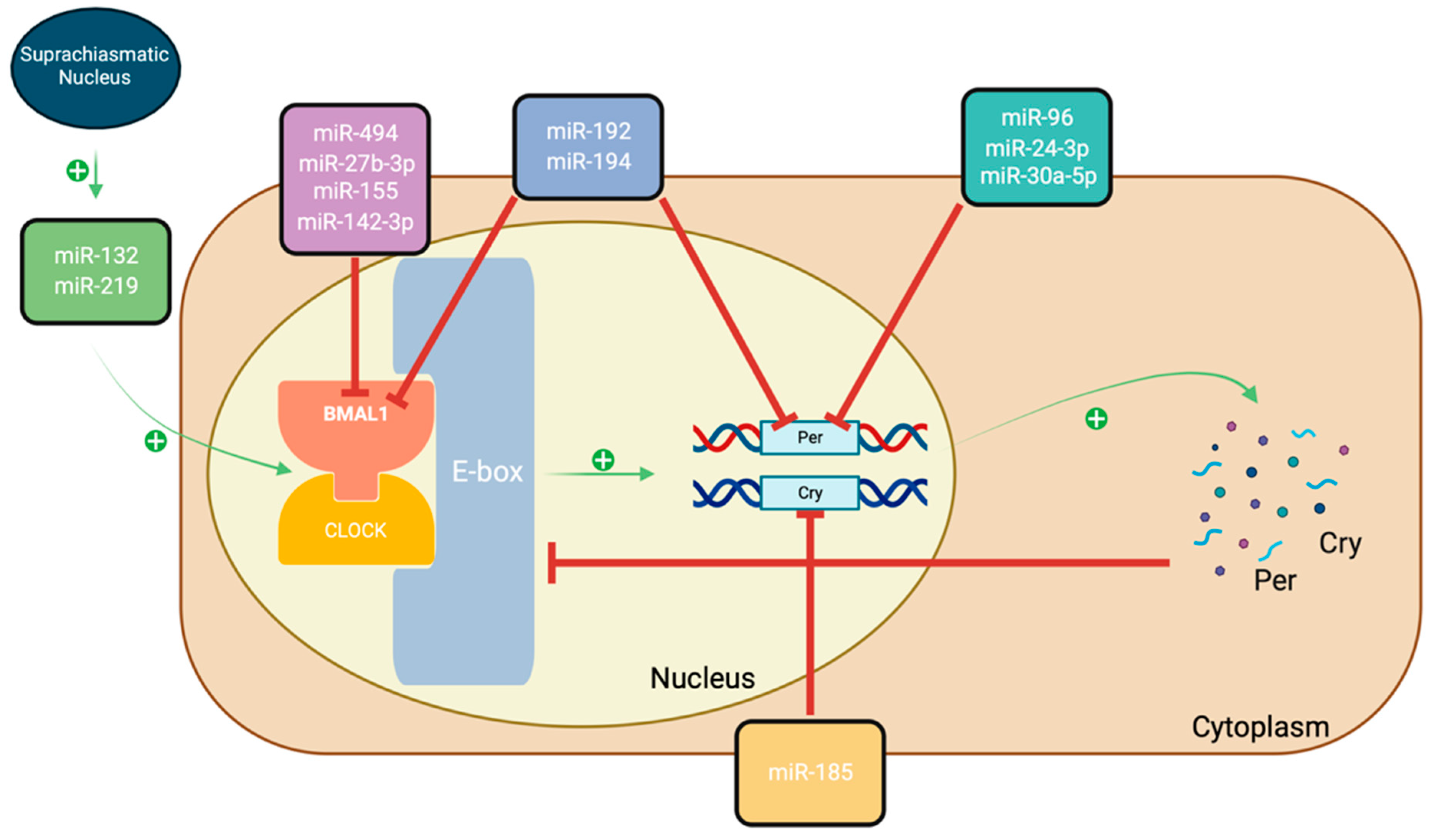
5. The Circadian Rhythm and PCOS
6. Potential Diagnostic and Prognostic Uses of miRNAs in PCOS
6.1. Biomarkers in Serum/Plasma
| miRNA(s) | Sample | Diagnostic/Prognostic Value | References |
|---|---|---|---|
| miR-222 | Serum | Marker of insulin sensitivity and type 2 diabetes | [83] |
| miR-146a | Serum | Negatively correlates with serum testosterone | [83] |
| miR-29a-5p | Serum | Marker of PCOS | [84] |
| miR-320 | Serum | Negatively correlates with insulin resistance | [84] |
| miR-339-5p | Serum | Marker of vascular endothelial health in PCOS | [61] |
| miR-103a-3p miR-21-5p miR-376a-3p | Serum | Positively correlate with androgen levels | [85] |
| miR-155 | Serum | Monitor estroprogestinic treatment in hyperandrogenic PCOS women | [86] |
| miR-93 | Plasma | Marker of PCOS | [31] |
| miR-146a-5p miR-126-3p | Plasma | Related to anovulation and polycystic ovaries | [87] |
| miR-20b-5p | Plasma | Potential marker of insulin resistance | [87] |
| miR-18a-3p | Plasma | Inversely correlates with luteinizing hormone | [87] |
| miR-106a-5p miR-20b-5p miR-18a-3p | Plasma | Correlates with plasma steroid hormones | [87] |
| miR-376a-3p miR-103-3p miR-139-5p miR-28-5p | Serum | Reflect androgenic profile | [88] |
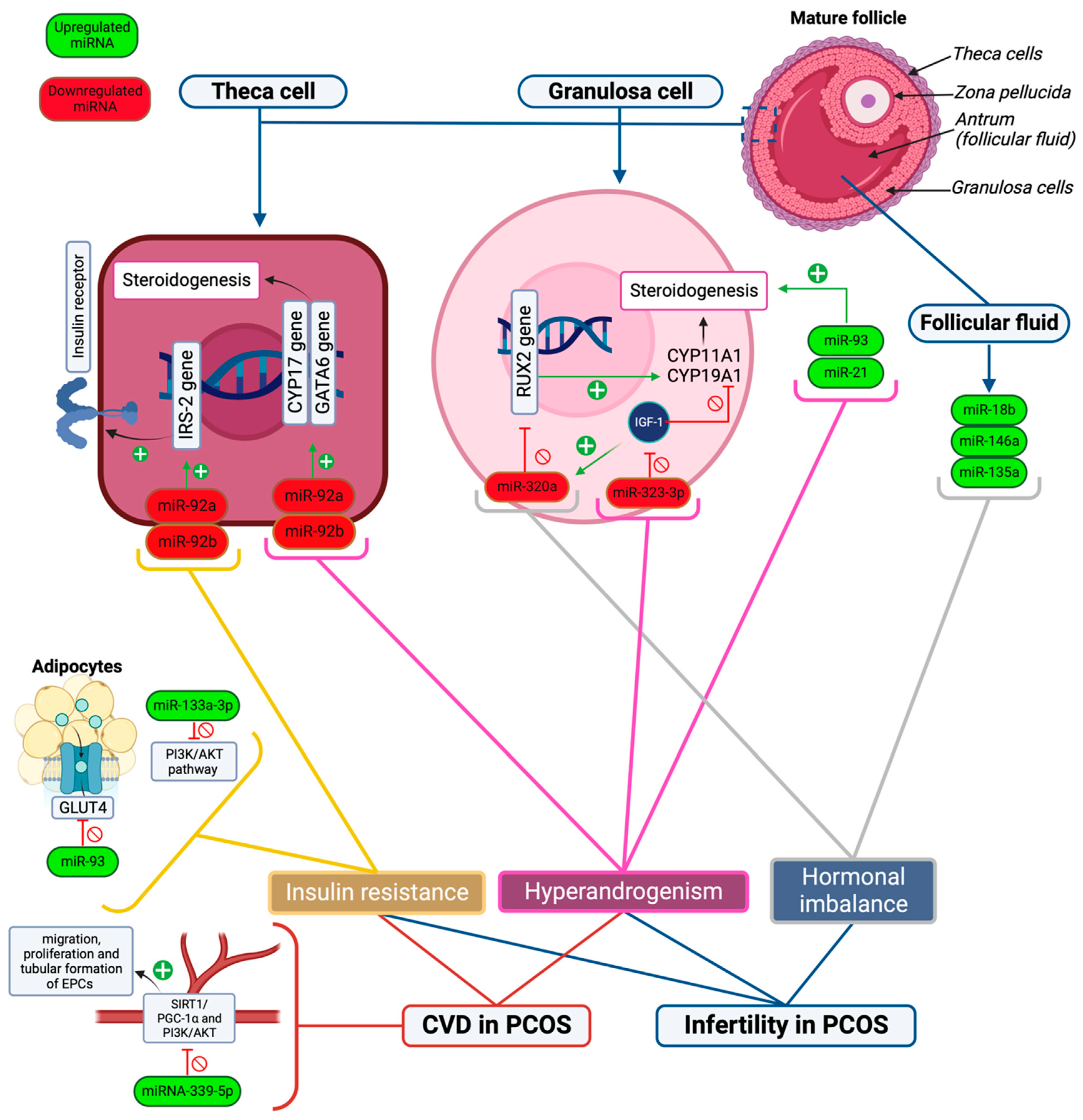
| miR-92b miR-92a | Theca cells | Correlate with hyperinsulinemia/IR and hyperandrogenism profile | [45] |
| miR-423 | Granulosa cells and follicular fluid | Downregulated, marker of PCOS | [89] |
| miR-142 miR-33b | Granulosa cells and follicular fluid | Upregulated, a marker of PCOS | [89] |
| miR-199b-5p | Follicular fluid | Correlates with anti-mullerian hormone | [39] |
| miR-382-5p | Follicular fluid | Correlates with free androgen index and age | [39] |
| miR-93-3p | Follicular fluid | Correlates with C-reactive protein | [39] |
| miR-490-5p miR-212-3p miR-4643 | Follicular fluid and cumulus cells | Identification of PCOS subtype | [34] |
| miR-647 | Follicular fluid and cumulus cells | Identification of PCOS subtype | [34] |
6.2. Biomarkers in Theca Cells
6.3. Biomarkers in Granulosa Cells
6.4. Biomarkers in Follicular Fluid
7. Other RNAs in PCOS
7.1. The Role of Long Non-Coding (lncRNA) RNAs in PCOS
7.1.1. Effects of lncRNAs in PCOS
7.1.2. The Interaction between miRNAs and lncRNAs in PCOS
| lncRNA | Role in PCOS | References |
|---|---|---|
| Effects of lncRNAs in PCOS | ||
| lncRNA CCNL1-3:1 | Overexpression of lncRNA CCNL elevates FOX01 expression, resulting in cell death, decreased glucose transporters, and altered mitochondrial functions. Therefore, lncRNA CCNL leads to two main complications of PCOS, which are follicular atresia and insulin resistance. | [94] |
| lncRNA GDAR | lncRNA GDAR is reduced in PCOS, leading to a greater expression of mRNAs and proteins associated with the apoptosis of granulosa cells. | [91] |
| lncRNA BANCR | LncBANCR is upregulated in PCOS, enhancing cell death and preventing cell growth, indicating a role in the progression of PCOS. | [95] |
| Interactions between lncRNAs and miRNAs in PCOS | ||
| lncRNA ZFAS1 | lncRNA ZFAS1 is increased, and miR-129 is reduced in PCOS patients. The interactions of lncRNA ZFAS1 and miR-129 induce PCOS by increasing apoptosis of granulosa cells via increased HMGB1 gene expression. | [96] |
| lncRNA NEAT1 and lncRNA MALAT1 | In PCOS, lncRNA NEAT1 has elevated levels in serum and granulosa cells; however, lncRNA MALAT1 is repressed. Their involvement in PCOS development relies on adjusting miR-30d-5p and miR-30a-5p, which affects IRS-2, AR, and FST expression. | [98] |
| lncRNA HOTAIR | Upregulated lncRNA HOTAIR competitively binds to miR-130a, leading to higher expression of IGF1 seen in rat models of PCOS. | [99] |
| LncRNA HCP5 | LncRNA HCP5 is upregulated in PCOS, which competitively inhibits miR-27a-3p, leading to increased activity of IGF1. | [100] |
7.2. The Role of Circular RNA (circRNA) in PCOS
8. Future Directions
- Establish a definitive set of miRNAs that are common in the majority of PCOS patients, which can be used as prognostic markers and for disease monitoring purposes.
- Further research to identify the mode of function of miRNAs to understand how they manipulate and are manipulated by insulin, thereby providing potential therapeutic targets not only for PCOS patients but all those suffering from insulin disorders (such as T2DM, pre-diabetes, etc.).
- Previous studies often do not clearly define the PCOS population in terms of the Rotterdam criteria, and therefore, comparisons are often of mixed phenotypes. Consequently, the results can be difficult to interpret.
- Studies to identify potential therapeutic uses of miRNA in PCOS patients.
- Further research is needed to establish reliable, non-invasive methods to diagnose PCOS.
- Studies to identify the potential for MiR-143-3p and MiR-155-5p to act as treatment targets in PCOS patients.
- Further studies are needed to identify the relationship between CVD and PCOS in relation to miRNA, as there is a lack of literature surrounding this area.
- Studies to investigate lncRNAs as circulating biomarkers for PCOS and their potential to monitor treatment effectiveness.
- As various circRNAs have been associated with the progression of PCOS, studies may identify potential diagnostic and therapeutic uses of circRNAs.
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ago2 | Argonaute RISC catalytic component 2 |
| Bax | Bcl-2 associated x protein |
| Bcl-2 | B cell leukemia/lymphoma protein 2 |
| Bcl-9 | B cell leukemia/lymphoma 9 |
| BMAL1 | brain and muscle Arnt-like protein 1 |
| CCNL1 | LncRNA Cyclin L1 |
| ceRNA | Competitive endogenous RNA |
| circRNA | Circular RNA |
| circRNAs | Circular RNAs |
| CLOCK | Circadian locomotor output cycle kaput |
| Cry1, Cry2 | Cryptochrome |
| CYP17 | 17-hydroxylase/C17–20 lyase cytochrome P50 |
| FOX01 | Forkhead box 01 |
| Fox12b | Forkhead transcription factor 12b |
| GATA6 | GATA-binding factor 6 |
| Gdf | Growth differentiation factor |
| hLGCs | Human luteinized granulosa cells |
| HMGB1 | High mobility group box 1 |
| IGF-1 | Insulin-like growth factor 1 |
| KGN cells | Human granulosa-like tumor cell line |
| lncGDAR | Long noncoding granulosa-cells-differentiation-associated RNA |
| MALAT1 | Metastasis-associated lung adenocarcinoma transcript 1 |
| MAPK | Mitogen-activated protein kinase |
| NEAT1 | Nuclear-enriched abundant transcript 1 |
| NFkB | Nuclear factor kappa B |
| Per1, Per2, Per3 | Period |
| PI3K/AKT(PKB) | Phosphoinositide-3-kinase/protein kinase B |
| pri-miRNA | Primary miRNA |
| pre-miRNA | Precursor miRNA |
| PTEN | Phosphatase and tensin homolog |
| SIRT1/PGC-1α | Silent information regulator 1/peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| TRBP | Dicer-transactivation response RNA binding protein |
| ZFAS1 | Zinc finger antisense 1 |
References
- Azziz, R. Polycystic Ovary Syndrome. Obstet. Gynecol. 2018, 132, 321–336. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Deswal, R.; Narwal, V.; Dang, A.; Pundir, C.S. The Prevalence of Polycystic Ovary Syndrome: A Brief Systematic Review. J. Hum. Reprod. Sci. 2020, 13, 261–271. [Google Scholar] [PubMed]
- Witchel, S.F.; Oberfield, S.E.; Pena, A.S. Polycystic Ovary Syndrome: Pathophysiology, Presentation, and Treatment with Emphasis on Adolescent Girls. J. Endocr. Soc. 2019, 3, 1545–1573. [Google Scholar] [CrossRef] [PubMed]
- Randeva, H.S.; Tan, B.K.; Weickert, M.O.; Lois, K.; Nestler, J.E.; Sattar, N.; Lehnert, H. Cardiometabolic aspects of the polycystic ovary syndrome. Endocr. Rev. 2012, 33, 812–841. [Google Scholar] [CrossRef]
- Vink, J.M.; Sadrzadeh, S.; Lambalk, C.B.; Boomsma, D.I. Heritability of polycystic ovary syndrome in a Dutch twin-family study. J. Clin. Endocrinol. Metab. 2006, 91, 2100–2104. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Luque-Ramirez, M.; San Millan, J.L. The molecular-genetic basis of functional hyperandrogenism and the polycystic ovary syndrome. Endocr. Rev. 2005, 26, 251–282. [Google Scholar] [CrossRef]
- Charifson, M.A.; Trumble, B.C. Evolutionary origins of polycystic ovary syndrome: An environmental mismatch disorder. Evol. Med. Public Health 2019, 2019, 50–63. [Google Scholar] [CrossRef]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Fertil. Steril. 2018, 110, 364–379. [Google Scholar] [CrossRef]
- Smet, M.E.; McLennan, A. Rotterdam criteria, the end. Australas. J. Ultrasound Med. 2018, 21, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Hoeger, K.M.; Dokras, A.; Piltonen, T. Update on PCOS: Consequences, Challenges, and Guiding Treatment. J. Clin. Endocrinol. Metab. 2021, 106, e1071–e1083. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.O.; Dumesic, D.A.; Chazenbalk, G.; Azziz, R. Polycystic ovary syndrome: Etiology, pathogenesis and diagnosis. Nat. Rev. Endocrinol. 2011, 7, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The Androgen Excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, K.; Sivasankar, V. MicroRNAs—Biology and clinical applications. J. Oral Maxillofac. Pathol. 2014, 18, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, biogenesis, and their evolving role in animal development and disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef]
- Ilie, I.R.; Georgescu, C.E. Polycystic ovary syndrome-epigenetic mechanisms and aberrant microRNA. Adv. Clin. Chem. 2015, 71, 25–45. [Google Scholar]
- Abdalla, M.; Deshmukh, H.; Atkin, S.L.; Sathyapalan, T. miRNAs as a novel clinical biomarker and therapeutic targets in polycystic ovary syndrome (PCOS): A review. Life Sci. 2020, 259, 118174. [Google Scholar] [CrossRef]
- Flynt, A.S.; Lai, E.C. Biological principles of microRNA-mediated regulation: Shared themes amid diversity. Nat. Rev. Genet. 2008, 9, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Lv, J.; Xu, P.; Gu, L.; Cao, J.; Xu, L.; Xue, L.; Li, Q. Identification of microRNAs and genes associated with hyperandrogenism in the follicular fluid of women with polycystic ovary syndrome. J. Cell. Biochem. 2018, 119, 3913–3921. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.G.; Fulghesu, A.M.; Mikus, M.; Watrowski, R.; D’Alterio, M.N.; Lin, L.T.; Shah, M.; Reyes-Muñoz, E.; Sathyapalan, T.; Angioni, S. The Translational Role of miRNA in Polycystic Ovary Syndrome: From Bench to Bedside—A Systematic Literature Review. Biomedicines 2022, 10, 1816. [Google Scholar] [CrossRef] [PubMed]
- Imbar, T.; Eisenberg, I. Regulatory role of microRNAs in ovarian function. Fertil. Steril. 2014, 101, 1524–1530. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Chang, H.R.; Baek, D. Determinants of Functional MicroRNA Targeting. Mol. Cells 2023, 46, 21–32. [Google Scholar] [CrossRef]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-Based Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Mari-Alexandre, J.; Barcelo-Molina, M.; Belmonte-Lopez, E.; Garcia-Oms, J.; Estelles, A.; Braza-Boils, A.; Gilabert-Estellés, J. Micro-RNA profile and proteins in peritoneal fluid from women with endometriosis: Their relationship with sterility. Fertil. Steril. 2018, 109, 675–684.e2. [Google Scholar] [CrossRef]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids—The mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef]
- Li, P.; Liu, H.; Li, Y.; Wang, Y.; Zhao, L.; Wang, H. miR-339-5p inhibits lung adenocarcinoma invasion and migration by directly targeting BCL6. Oncol. Lett. 2018, 16, 5785–5790. [Google Scholar] [CrossRef]
- Shan, W.; Li, J.; Bai, Y.; Lu, X. miR-339-5p inhibits migration and invasion in ovarian cancer cell lines by targeting NACC1 and BCL6. Tumor Biol. 2016, 37, 5203–5211. [Google Scholar] [CrossRef]
- Sathyapalan, T.; David, R.; Gooderham, N.J.; Atkin, S.L. Increased expression of circulating miRNA-93 in women with polycystic ovary syndrome may represent a novel, non-invasive biomarker for diagnosis. Sci. Rep. 2015, 5, 16890. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xu, P.; Wang, J.; Zhang, C. The role of MiRNA in polycystic ovary syndrome (PCOS). Gene 2019, 706, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Motahari Rad, H.; Mowla, S.J.; Ramazanali, F.; Rezazadeh Valojerdi, M. Characterization of altered microRNAs related to different phenotypes of polycystic ovarian syndrome (PCOS) in serum, follicular fluid, and cumulus cells. Taiwan. J. Obstet. Gynecol. 2022, 61, 768–779. [Google Scholar] [CrossRef]
- Li, M.; Ruan, X.; Mueck, A.O. Management strategy of infertility in polycystic ovary syndrome. Glob. Health J. 2022, 6, 70–74. [Google Scholar] [CrossRef]
- Zhuang, S.; Jing, C.; Yu, L.; Ji, L.; Liu, W.; Hu, X. The relationship between polycystic ovary syndrome and infertility: A bibliometric analysis. Ann. Transl. Med. 2022, 10, 318. [Google Scholar] [CrossRef]
- Sawant, S.; Bhide, P. Fertility Treatment Options for Women with Polycystic Ovary Syndrome. Clin. Med. Insights Reprod. Health 2019, 13, 1179558119890867. [Google Scholar] [CrossRef]
- Gunning, M.N.; Fauser, B. Are women with polycystic ovary syndrome at increased cardiovascular disease risk later in life? Climacteric 2017, 20, 222–227. [Google Scholar] [CrossRef]
- Butler, A.E.; Ramachandran, V.; Hayat, S.; Dargham, S.R.; Cunningham, T.K.; Benurwar, M.; Sathyapalan, T.; Najafi-Shoushtari, S.H.; Atkin, S.L. Expression of microRNA in follicular fluid in women with and without PCOS. Sci. Rep. 2019, 9, 16306. [Google Scholar] [CrossRef]
- Xu, Y.; Qiao, J. Association of Insulin Resistance and Elevated Androgen Levels with Polycystic Ovarian Syndrome (PCOS): A Review of Literature. J. Healthc. Eng. 2022, 2022, 9240569. [Google Scholar] [CrossRef]
- Roth, L.W.; McCallie, B.; Alvero, R.; Schoolcraft, W.B.; Minjarez, D.; Katz-Jaffe, M.G. Altered microRNA and gene expression in the follicular fluid of women with polycystic ovary syndrome. J. Assist. Reprod. Genet. 2014, 31, 355–362. [Google Scholar] [CrossRef]
- Naji, M.; Aleyasin, A.; Nekoonam, S.; Arefian, E.; Mahdian, R.; Amidi, F. Differential Expression of miR-93 and miR-21 in Granulosa Cells and Follicular Fluid of Polycystic Ovary Syndrome Associating with Different Phenotypes. Sci. Rep. 2017, 7, 14671. [Google Scholar] [CrossRef]
- Wang, T.; Liu, Y.; Lv, M.; Xing, Q.; Zhang, Z.; He, X.; Xu, Y.; Wei, Z.; Cao, Y. miR-323-3p regulates the steroidogenesis and cell apoptosis in polycystic ovary syndrome (PCOS) by targeting IGF-1. Gene 2019, 683, 87–100. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wang, H.; Yan, C.Y.; Gao, X.F.; Ling, X.J. Deregulation of RUNX2 by miR-320a deficiency impairs steroidogenesis in cumulus granulosa cells from polycystic ovary syndrome (PCOS) patients. Biochem. Biophys. Res. Commun. 2017, 482, 1469–1476. [Google Scholar] [CrossRef]
- Lin, L.; Du, T.; Huang, J.; Huang, L.L.; Yang, D.Z. Identification of differentially expressed microRNAs in the ovary of polycystic ovary syndrome with hyperandrogenism and insulin resistance. Chin. Med. J. 2015, 128, 169–174. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, J.; Cheng, X.; Nie, X.; He, B. Insulin resistance in polycystic ovary syndrome across various tissues: An updated review of pathogenesis, evaluation, and treatment. J. Ovarian Res. 2023, 16, 9. [Google Scholar] [CrossRef]
- Chen, Y.H.; Heneidi, S.; Lee, J.M.; Layman, L.C.; Stepp, D.W.; Gamboa, G.M.; Chen, B.S.; Chazenbalk, G.; Azziz, R. miRNA-93 inhibits GLUT4 and is overexpressed in adipose tissue of polycystic ovary syndrome patients and women with insulin resistance. Diabetes 2013, 62, 2278–2286. [Google Scholar] [CrossRef]
- Balasubramanyam, M.; Aravind, S.; Gokulakrishnan, K.; Prabu, P.; Sathishkumar, C.; Ranjani, H.; Mohan, V. Impaired miR-146a expression links subclinical inflammation and insulin resistance in Type 2 diabetes. Mol. Cell. Biochem. 2011, 351, 197–205. [Google Scholar] [CrossRef]
- Coleman, C.B.; Lightell, D.J., Jr.; Moss, S.C.; Bates, M.; Parrino, P.E.; Woods, T.C. Elevation of miR-221 and -222 in the internal mammary arteries of diabetic subjects and normalization with metformin. Mol. Cell. Endocrinol. 2013, 374, 125–129. [Google Scholar] [CrossRef]
- Capuani, B.; Pacifici, F.; Della-Morte, D.; Lauro, D. Glucagon Like Peptide 1 and MicroRNA in Metabolic Diseases: Focusing on GLP1 Action on miRNAs. Front. Endocrinol. 2018, 9, 719. [Google Scholar] [CrossRef]
- Radbakhsh, S.; Sathyapalan, T.; Banach, M.; Sahebkar, A. Incretins and microRNAs: Interactions and physiological relevance. Pharmacol. Res. 2020, 153, 104662. [Google Scholar] [CrossRef]
- Angioni, S.; Sanna, S.; Magnini, R.; Melis, G.B.; Fulghesu, A.M. The quantitative insulin sensitivity check index is not able to detect early metabolic alterations in young patients with polycystic ovarian syndrome. Gynecol. Endocrinol. 2011, 27, 468–474. [Google Scholar] [CrossRef]
- Cao, J.; Huo, P.; Cui, K.; Wei, H.; Cao, J.; Wang, J.; Liu, Q.; Lei, X.; Zhang, S. Follicular fluid-derived exosomal miR-143-3p/miR-155-5p regulate follicular dysplasia by modulating glycolysis in granulosa cells in polycystic ovary syndrome. Cell Commun. Signal. 2022, 20, 61. [Google Scholar] [CrossRef]
- Bao, B.; Wang, Z.; Ali, S.; Ahmad, A.; Azmi, A.S.; Sarkar, S.H.; Banerjee, S.; Kong, D.; Li, Y.; Thakur, S.; et al. Metformin inhibits cell proliferation, migration and invasion by attenuating CSC function mediated by deregulating miRNAs in pancreatic cancer cells. Cancer Prev. Res. 2012, 5, 355–364. [Google Scholar] [CrossRef]
- Guan, C.; Zahid, S.; Minhas, A.S.; Ouyang, P.; Vaught, A.; Baker, V.L.; Michos, E.D. Polycystic ovary syndrome: A “risk-enhancing” factor for cardiovascular disease. Fertil. Steril. 2022, 117, 924–935. [Google Scholar] [CrossRef]
- Meun, C.; Gunning, M.N.; Louwers, Y.V.; Peters, H.; Roos-Hesselink, J.; Roeters van Lennep, J.; Rueda Ochoa, O.L.; Appelman, Y.; Lambalk, N.; Boersma, E.; et al. The cardiovascular risk profile of middle-aged women with polycystic ovary syndrome. Clin. Endocrinol. 2020, 92, 150–158. [Google Scholar] [CrossRef]
- Osibogun, O.; Ogunmoroti, O.; Michos, E.D. Polycystic ovary syndrome and cardiometabolic risk: Opportunities for cardiovascular disease prevention. Trends Cardiovasc. Med. 2020, 30, 399–404. [Google Scholar] [CrossRef]
- Gomez, J.M.D.; VanHise, K.; Stachenfeld, N.; Chan, J.L.; Merz, N.B.; Shufelt, C. Subclinical cardiovascular disease and polycystic ovary syndrome. Fertil. Steril. 2022, 117, 912–923. [Google Scholar] [CrossRef]
- Duica, F.; Danila, C.A.; Boboc, A.E.; Antoniadis, P.; Condrat, C.E.; Onciul, S.; Suciu, N.; Creţoiu, S.M.; Varlas, V.N.; Creţoiu, D. Impact of Increased Oxidative Stress on Cardiovascular Diseases in Women with Polycystic Ovary Syndrome. Front. Endocrinol. 2021, 12, 614679. [Google Scholar] [CrossRef]
- Wronska, A. The Role of microRNA in the Development, Diagnosis, and Treatment of Cardiovascular Disease: Recent Developments. J. Pharmacol. Exp. Ther. 2023, 384, 123–132. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, W.; Li, S.; Zhang, J.; Shang, Y.; Gui, J. The role of miRNA-339-5p in the function of vascular endothelial progenitor cells in patients with PCOS. Reprod. Biomed. Online 2022, 44, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.H.; Chiu, W.C.; Hsu, M.I.; Chen, Y.J. Endothelial progenitor cell dysfunction in polycystic ovary syndrome: Implications for the genesis of cardiovascular diseases. Int. J. Fertil. Steril. 2013, 6, 208–213. [Google Scholar] [PubMed]
- Willis, S.K.; Hatch, E.E.; Wise, L.A. Sleep and female reproduction. Curr. Opin. Obstet. Gynecol. 2019, 31, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Huang, Q.Y.; Wang, Z.Y.; Zhuang, X.X.; Lin, S.; Shi, Q.Y. Therapeutic potential of exosomes/miRNAs in polycystic ovary syndrome induced by the alteration of circadian rhythms. Front. Endocrinol. 2022, 13, 918805. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tong, X.; Arthurs, B.; Guha, A.; Rui, L.; Kamath, A.; Inoki, K.; Yin, L. Liver clock protein BMAL1 promotes de novo lipogenesis through insulin-mTORC2-AKT signaling. J. Biol. Chem. 2014, 289, 25925–25935. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Zhao, H.; Lu, Z.; Lei, X.; Zhang, Y. Circadian Rhythms within the Female HPG Axis: From Physiology to Etiology. Endocrinology 2021, 162, bqab117. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Mohanty, S.R.; Minocha, T.; Mishra, N.K.; Yadav, S.K.; Haldar, C. Circadian desynchronization in pregnancy of Golden hamster following long time light exposure: Involvement of Akt/FoxO1 pathway. J. Photochem. Photobiol. B 2022, 234, 112508. [Google Scholar] [CrossRef]
- Soyman, Z.; Durmus, S.; Ates, S.; Simsek, G.; Sozer, V.; Kundaktepe, B.P.; Kurtulus, D.; Gelisgen, R.; Sal, V.; Uzun, H. Circulating Mir-132, Mir-146a, Mir-222, and Mir-320 Expression in Differential Diagnosis of Women with Polycystic Ovary Syndrome. Acta Endocrinol. 2022, 18, 13–19. [Google Scholar] [CrossRef]
- Li, M.; Zhao, H.; Zhao, S.G.; Wei, D.M.; Zhao, Y.R.; Huang, T.; Muhammad, T.; Yan, L.; Gao, F.; Li, L.; et al. The HMGA2-IMP2 Pathway Promotes Granulosa Cell Proliferation in Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2019, 104, 1049–1059. [Google Scholar] [CrossRef]
- Lionett, S.; Kiel, I.A.; Camera, D.M.; Vanky, E.; Parr, E.B.; Lydersen, S.; Hawley, J.A.; Moholdt, T. Circulating and Adipose Tissue miRNAs in Women with Polycystic Ovary Syndrome and Responses to High-Intensity Interval Training. Front. Physiol. 2020, 11, 904. [Google Scholar] [CrossRef]
- Xia, H.; Zhao, Y. miR-155 is high-expressed in polycystic ovarian syndrome and promotes cell proliferation and migration through targeting PDCD4 in KGN cells. Artif. Cells Nanomed. Biotechnol. 2020, 48, 197–205. [Google Scholar] [CrossRef]
- Insenser, M.; Quintero, A.; de Lope, S.; Alvarez-Blasco, F.; Martinez-Garcia, M.A.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Validation of circulating microRNAs miR-142-3p and miR-598-3p in women with polycystic ovary syndrome as potential diagnostic markers. Hum. Reprod. 2023, 38, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.H.; Kojima, S.; Shimomura, K.; Koike, N.; Buhr, E.D.; Furukawa, T.; Ko, C.H.; Glostron, G.; Ayoub, C.; Nohara, K.; et al. Period2 3′-UTR and microRNA-24 regulate circadian rhythms by repressing PERIOD2 protein accumulation. Proc. Natl. Acad. Sci. USA 2017, 114, E8855–E8864. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Huang, J.; Chen, Y.; Yang, Y.; Li, R.; Li, Y.; Chen, X.; Yang, D. Identification of several circulating microRNAs from a genome-wide circulating microRNA expression profile as potential biomarkers for impaired glucose metabolism in polycystic ovarian syndrome. Endocrine 2016, 53, 280–290. [Google Scholar] [CrossRef]
- Zhou, L.; Miller, C.; Miraglia, L.J.; Romero, A.; Mure, L.S.; Panda, S.; Kay, S.A. A genome-wide microRNA screen identifies the microRNA-183/96/182 cluster as a modulator of circadian rhythms. Proc. Natl. Acad. Sci. USA 2021, 118, e2020454118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, S.; Chen, L.; Huang, X.; Wang, M.; Ponikwicka-Tyszko, D.; Ragman, N.A.; Wolcsynski, S.; Yao, B.; Li, X. The molecular mechanism of miR-96-5p in the pathogenesis and treatment of polycystic ovary syndrome. Transl. Res. 2023, 256, 1–13. [Google Scholar] [CrossRef]
- Sorensen, A.E.; Wissing, M.L.; Englund, A.L.; Dalgaard, L.T. MicroRNA Species in Follicular Fluid Associating with Polycystic Ovary Syndrome and Related Intermediary Phenotypes. J. Clin. Endocrinol. Metab. 2016, 101, 1579–1589. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, C.; Lin, Z.; Pan, H.; Li, K.; Ma, H. MiRNAs expression profiling of rat ovaries displaying PCOS with insulin resistance. Arch. Gynecol. Obstet. 2020, 302, 1205–1213. [Google Scholar] [CrossRef]
- Tao, S.C.; Guo, S.C. Extracellular Vesicles: Potential Participants in Circadian Rhythm Synchronization. Int. J. Biol. Sci. 2018, 14, 1610–1620. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, B.; Yang, L.; Bai, Y.G.; Song, J.B.; Ge, Y.L.; Ma, H.Z.; Cheng, J.H.; Ma, J.; Xie, M.J. BMAL1 Disrupted Intrinsic Diurnal Oscillation in Rat Cerebrovascular Contractility of Simulated Microgravity Rats by Altering Circadian Regulation of miR-103/Ca(V)1.2 Signal Pathway. Int. J. Mol. Sci. 2019, 20, 3947. [Google Scholar] [CrossRef]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, A.E.; Wissing, M.L.; Salo, S.; Englund, A.L.; Dalgaard, L.T. MicroRNAs Related to Polycystic Ovary Syndrome (PCOS). Genes 2014, 5, 684–708. [Google Scholar] [CrossRef] [PubMed]
- Long, W.; Zhao, C.; Ji, C.; Ding, H.; Cui, Y.; Guo, X.; Shen, R.; Liu, J. Characterization of serum microRNAs profile of PCOS and identification of novel non-invasive biomarkers. Cell Physiol. Biochem. 2014, 33, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Deswal, R.; Dang, A.S. Dissecting the role of micro-RNAs as a diagnostic marker for polycystic ovary syndrome: A systematic review and meta-analysis. Fertil. Steril. 2020, 113, 661–669.e2. [Google Scholar] [CrossRef]
- De Nardo Maffazioli, G.; Baracat, E.C.; Soares, J.M.; Carvalho, K.C.; Maciel, G.A.R. Evaluation of circulating microRNA profiles in Brazilian women with polycystic ovary syndrome: A preliminary study. PLoS ONE 2022, 17, e0275031. [Google Scholar] [CrossRef] [PubMed]
- Arancio, W.; Calogero Amato, M.; Magliozzo, M.; Pizzolanti, G.; Vesco, R.; Giordano, C. Serum miRNAs in women affected by hyperandrogenic polycystic ovary syndrome: The potential role of miR-155 as a biomarker for monitoring the estroprogestinic treatment. Gynecol. Endocrinol. 2018, 34, 704–708. [Google Scholar] [CrossRef]
- Jiang, X.; Li, J.; Zhang, B.; Hu, J.; Ma, J.; Cui, L.; Chen, Z.J. Differential expression profile of plasma exosomal microRNAs in women with polycystic ovary syndrome. Fertil. Steril. 2021, 115, 782–792. [Google Scholar] [CrossRef]
- Udesen, P.B.; Sorensen, A.E.; Svendsen, R.; Frisk, N.L.S.; Hess, A.L.; Aziz, M.; Wissing, M.L.M.; Englund, A.L.M.; Dalgaard, L.T. Circulating miRNAs in Women with Polycystic Ovary Syndrome: A Longitudinal Cohort Study. Cells 2023, 12, 983. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiang, Y.; Song, Y.; Wan, L.; Yu, G.; Tan, L. Dysregulated miR-142, -33b and -423 in granulosa cells target TGFBR1 and SMAD7: A possible role in polycystic ovary syndrome. Mol. Hum. Reprod. 2019, 25, 638–646. [Google Scholar] [CrossRef]
- Tamaddon, M.; Azimzadeh, M.; Tavangar, S.M. microRNAs and long non-coding RNAs as biomarkers for polycystic ovary syndrome. J. Cell. Mol. Med. 2022, 26, 654–670. [Google Scholar] [CrossRef]
- Butler, A.E.; Hayat, S.; Dargham, S.R.; Malek, J.A.; Abdulla, S.A.; Mohamoud, Y.A.; Suhre, K.; Sathyapalan, T.; Atkin, S.L. Alterations in long noncoding RNAs in women with and without polycystic ovarian syndrome. Clin. Endocrinol. 2019, 91, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, A.; Yu, B.; Li, S. Interplay between miRNAs and lncRNAs: Mode of action and biological roles in plant development and stress adaptation. Comput. Struct. Biotechnol. J. 2021, 19, 2567–2574. [Google Scholar] [CrossRef] [PubMed]
- Ratti, M.; Lampis, A.; Ghidini, M.; Salati, M.; Mirchev, M.B.; Valeri, N.; Hahne, J.C. MicroRNAs (miRNAs) and Long Non-Coding RNAs (lncRNAs) as New Tools for Cancer Therapy: First Steps from Bench to Bedside. Target. Oncol. 2020, 15, 261–278. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, J.; Geng, X.; Chu, W.; Li, S.; Chen, Z.J.; Du, Y. Long non-coding RNA lnc-CCNL1-3:1 promotes granulosa cell apoptosis and suppresses glucose uptake in women with polycystic ovary syndrome. Mol. Ther. Nucleic Acids 2021, 23, 614–628. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, J.; Wang, L.; Deng, A. LncRNA BANCR participates in polycystic ovary syndrome by promoting cell apoptosis. Mol. Med. Rep. 2019, 19, 1581–1586. [Google Scholar] [CrossRef]
- Zhu, H.L.; Chen, Y.Q.; Zhang, Z.F. Downregulation of lncRNA ZFAS1 and upregulation of microRNA-129 repress endocrine disturbance, increase proliferation and inhibit apoptosis of ovarian granulosa cells in polycystic ovarian syndrome by downregulating HMGB1. Genomics 2020, 112, 3597–3608. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhang, Y.; Lu, Y.; Han, J.; Guo, T.; Cui, P.; Brännström, M.; Shao, L.R.; Billig, H. Regulatory mechanisms of HMGB1 and its receptors in polycystic ovary syndrome-driven gravid uterine inflammation. FEBS J. 2023, 290, 1874–1906. [Google Scholar] [CrossRef]
- ElMonier, A.A.; El-Boghdady, N.A.; Fahim, S.A.; Sabry, D.; Elsetohy, K.A.; Shaheen, A.A. LncRNA NEAT1 and MALAT1 are involved in polycystic ovary syndrome pathogenesis by functioning as competing endogenous RNAs to control the expression of PCOS-related target genes. Noncoding RNA Res. 2023, 8, 263–271. [Google Scholar] [CrossRef]
- Jiang, B.; Xue, M.; Xu, D.; Song, J.; Zhu, S. Down-regulated lncRNA HOTAIR alleviates polycystic ovaries syndrome in rats by reducing expression of insulin-like growth factor 1 via microRNA-130a. J. Cell. Mol. Med. 2020, 24, 451–464. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; An, Y.; Liu, B.; Lu, M. LncRNA HCP5 promotes cell proliferation and inhibits apoptosis via miR-27a-3p/IGF-1 axis in human granulosa-like tumor cell line KGN. Mol. Cell. Endocrinol. 2020, 503, 110697. [Google Scholar] [CrossRef]
- Lu, X.; Gao, H.; Zhu, B.; Lin, G. Circular RNA circ_RANBP9 exacerbates polycystic ovary syndrome via microRNA-136-5p/XIAP axis. Bioengineered 2021, 12, 6748–6758. [Google Scholar] [CrossRef]
- Liu, J.; Ding, J.; Qu, B.; Liu, J.; Song, X.; Suo, Q.; Zhou, A.; Yang, J. CircPSMC3 alleviates the symptoms of PCOS by sponging miR-296-3p and regulating PTEN expression. J. Cell. Mol. Med. 2020, 24, 11001–11011. [Google Scholar] [CrossRef]
- Zhang, D.; Yi, S.; Cai, B.; Wang, Z.; Chen, M.; Zheng, Z.; Zhou, C. Involvement of ferroptosis in the granulosa cells proliferation of PCOS through the circRHBG/miR-515/SLC7A11 axis. Ann. Transl. Med. 2021, 9, 1348. [Google Scholar] [CrossRef]
- Chen, Y.; Miao, J.; Lou, G. Knockdown of circ-FURIN suppresses the proliferation and induces apoptosis of granular cells in polycystic ovary syndrome via miR-195-5p/BCL2 axis. J. Ovarian Res. 2021, 14, 156. [Google Scholar] [CrossRef]
- Deng, L.; Chen, Q.; Xie, J.; Wei, W.; Hui, H. circPUM1 promotes polycystic ovary syndrome progression by sponging to miR-760. Gene 2020, 754, 144903. [Google Scholar] [CrossRef]
- Wu, G.; Xia, J.; Yang, Z.; Chen, Y.; Jiang, W.; Yin, T.; Yang, J. CircASPH promotes KGN cells proliferation through miR-375/MAP2K6 axis in Polycystic Ovary Syndrome. J. Cell. Mol. Med. 2022, 26, 1817–1825. [Google Scholar] [CrossRef]
- Xu, L.; Xiong, F.; Bai, Y.; Xiao, J.; Zhang, Y.; Chen, J.; Li, Q. Circ_0043532 regulates miR-182/SGK3 axis to promote granulosa cell progression in polycystic ovary syndrome. Reprod. Biol. Endocrinol. 2021, 19, 167. [Google Scholar] [CrossRef]
- Duan, J.; Cai, H.; Huang, Y.; Shi, L. SNAI2-Induced CircMTO1 Promotes Cell Proliferation and Inhibits Apoptosis Through the miR-320b/MCL1 Axis in Human Granulosa-Like Tumor Cells. Front. Genet. 2021, 12, 689916. [Google Scholar] [CrossRef]
- Xu, J.; Qu, Q.; Liu, B.; Shen, L. The circular RNA circ_0030018/miR-136/migration and invasion enhancer 1 (MIEN1) axis promotes the progression of polycystic ovary syndrome. Bioengineered 2022, 13, 5999–6011. [Google Scholar] [CrossRef]


| Phenotype | Androgen Excess | Ovulatory Dysfunction | Polycystic Ovarian Morphology on Ultrasound |
|---|---|---|---|
| A [Classic PCOS] | X | X | X |
| B [Essential NIH criteria] | X | X | |
| C [Ovulatory PCOS] | X | X | |
| D [Non-hyperandrogenic PCOS] | X | X |
| miRNA(s) | Sample | Finding in PCOS | References |
|---|---|---|---|
| miR-18b | Follicular fluid | Upregulated, promotes the release of progesterone while inhibiting the release of estradiol and testosterone, thus negatively impacting fertility. | [41] |
| miR-146a | Follicular fluid | Upregulated, reduces progesterone, estradiol, and testosterone release, causing irregularities in the menstrual cycle. | [41] |
| miR-135a | Follicular cells | Upregulated, reduces progesterone and testosterone release | [41] |
| miR-93 miR-21 | Granulosa cells | Upregulated in hyperandrogenic patients and has a role in follicular dysfunction. | [42] |
| miR-27a-3p | Granulosa cells | Impacts relative estrogen deficiency, which is believed to induce granulosa cell apoptosis. | [43] |
| miR-320a | Cumulus granulosa cells | Downregulated, has an effect on the IGF-1 regulatory mechanism and estrogen synthesis by targeting the RUNX2 gene. | [44] |
| miR-92a miR-92b | Theca cells | Downregulated, play a role in hyperandrogenism in PCOS by regulating CYP17, GATA6, and IRS-2 gene expression. | [45] |
| miR-323-3p | Cumulus granulosa cells | Downregulated, promotes apoptosis, and upregulates steroidogenesis in cumulus cells. | [43] |
| CircRNA | Role in PCOS | References |
|---|---|---|
| circRANBP9 | circRANBP9 is upregulated in PCOS patients. The loss of circRANBP9 reduces granulosa cell growth and promotes apoptosis via the regulation of miR-136-5p. | [101] |
| circPMSC3 | circPMSC3 is downregulated in PCOS. Overexpression of circPMSC3 modulates miR296-3p that targets PTEN, which could have therapeutic use in PCOS. | [102] |
| circRHBG | circRHBG is upregulated in PCOS; it prevents cell proliferation and inhibits ferroptosis. | [103] |
| circFURIN | circFURIN is upregulated in PCOS; reduction of circFURIN impairs cell proliferation and promotes apoptosis. | [104] |
| circPUM1 | circPUM1 functions as a sponge for miR-760, causing PCOS progression through apoptosis. | [105] |
| circASPH | circASPH is upregulated in PCOS; it restricts cell proliferation while promoting cell death and has an oncogenic role in the pathogenesis of PCOS. | [106] |
| circ_0043532 | circ_0043532 is upregulated; silencing circ_0043532 leads to the suppression of cell proliferation and stimulated cell apoptosis, indicating it as a therapeutic target in PCOS. | [107] |
| circMTO1 | Upregulated by SNAI2 and expressing miR-320b/MCL1 downstream, it aids the progression of human granulosa-like tumor cells. | [108] |
| circ_0030018 | Upregulated in PCOS patients and expresses the miR-136/MEIN1 axis, causing elevated AKT in the cytoplasm. | [109] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasser, J.S.; Altahoo, N.; Almosawi, S.; Alhermi, A.; Butler, A.E. The Role of MicroRNA, Long Non-Coding RNA and Circular RNA in the Pathogenesis of Polycystic Ovary Syndrome: A Literature Review. Int. J. Mol. Sci. 2024, 25, 903. https://doi.org/10.3390/ijms25020903
Nasser JS, Altahoo N, Almosawi S, Alhermi A, Butler AE. The Role of MicroRNA, Long Non-Coding RNA and Circular RNA in the Pathogenesis of Polycystic Ovary Syndrome: A Literature Review. International Journal of Molecular Sciences. 2024; 25(2):903. https://doi.org/10.3390/ijms25020903
Chicago/Turabian StyleNasser, Jenan Sh., Noor Altahoo, Sayed Almosawi, Abrar Alhermi, and Alexandra E. Butler. 2024. "The Role of MicroRNA, Long Non-Coding RNA and Circular RNA in the Pathogenesis of Polycystic Ovary Syndrome: A Literature Review" International Journal of Molecular Sciences 25, no. 2: 903. https://doi.org/10.3390/ijms25020903