
The Terminal Extensions of Dbp7 Influence Growth and 60S Ribosomal Subunit Biogenesis in Saccharomyces cerevisiae
 , ,
, ,  and
and 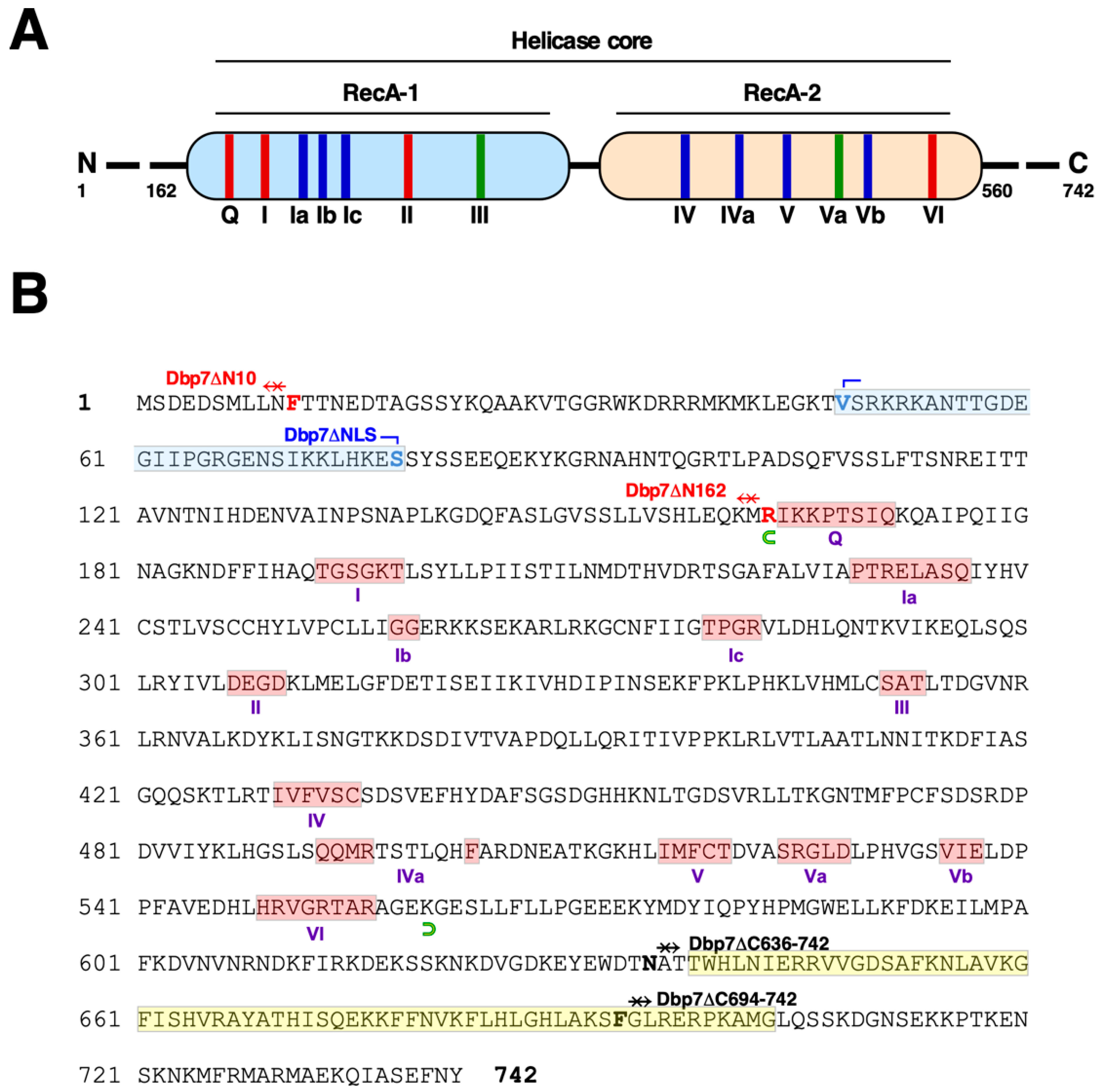{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Truncation of the N- and C-Terminal Extensions of Yeast Dbp7
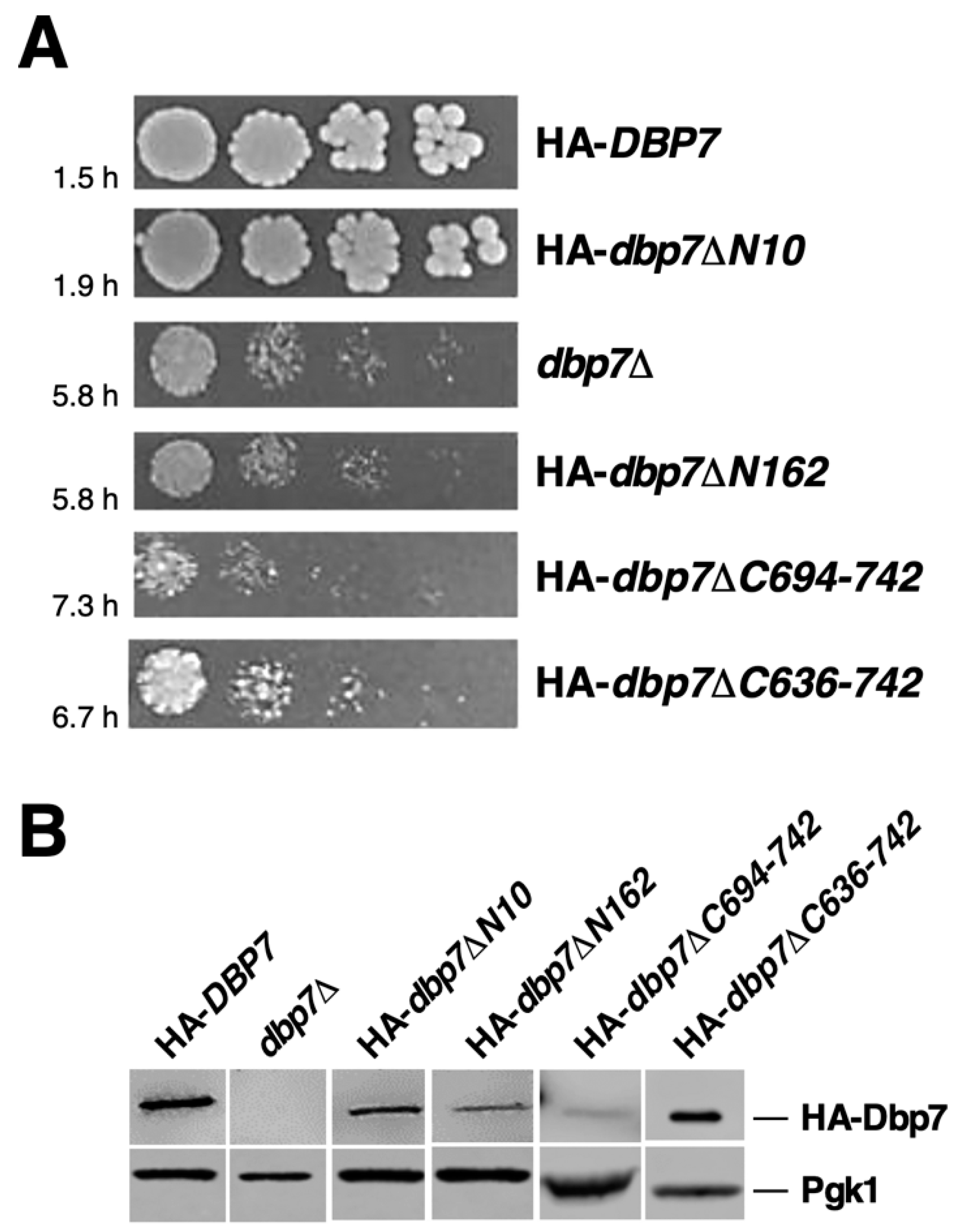
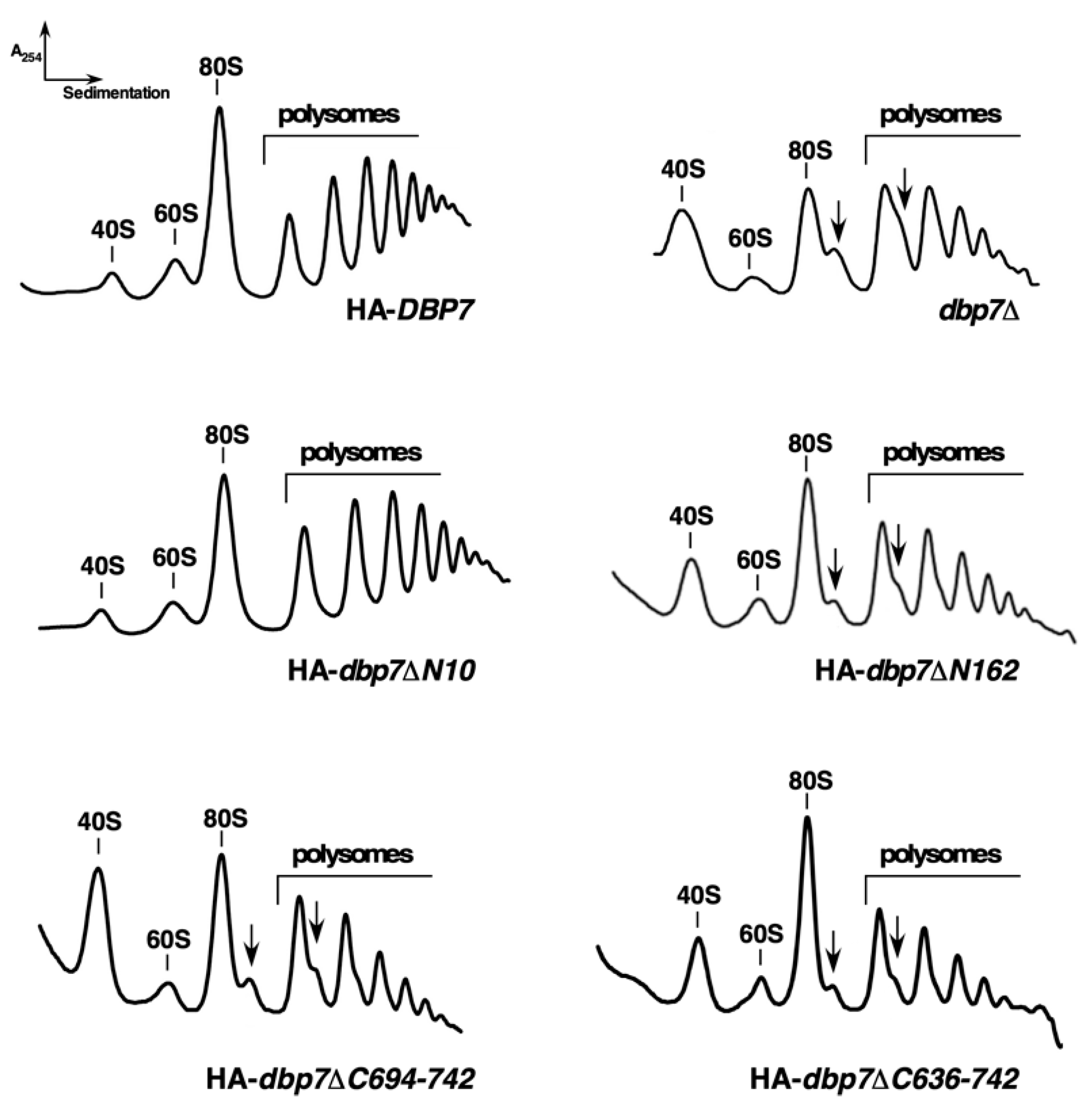
2.2. The N-Terminal Extension of Dbp7 Is Required for Efficient Cell Growth and for Production of 60S r-Subunits
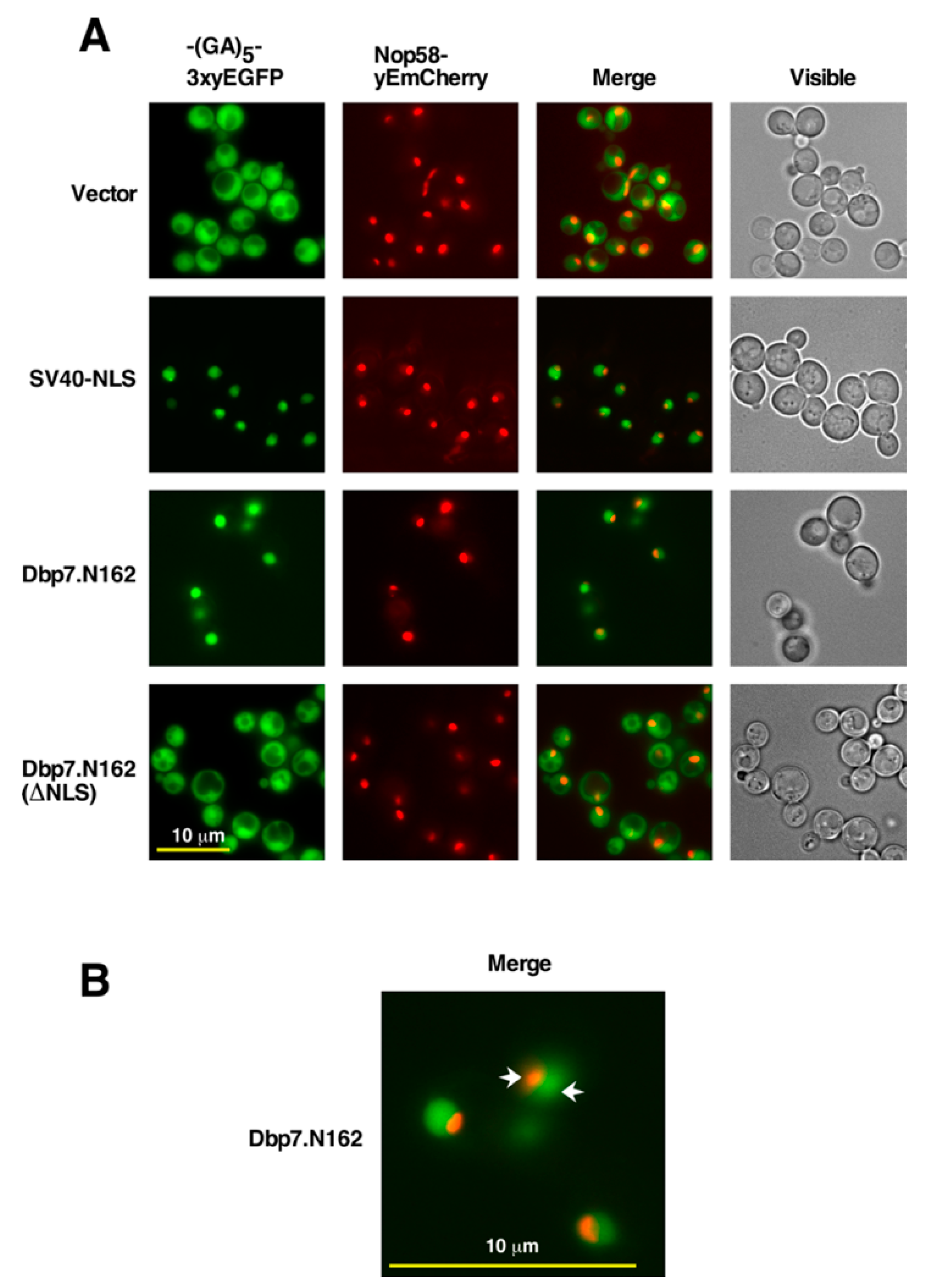
2.3. The N-Terminal Region of Dbp7 Contains a Functional Nuclear Localization Signal
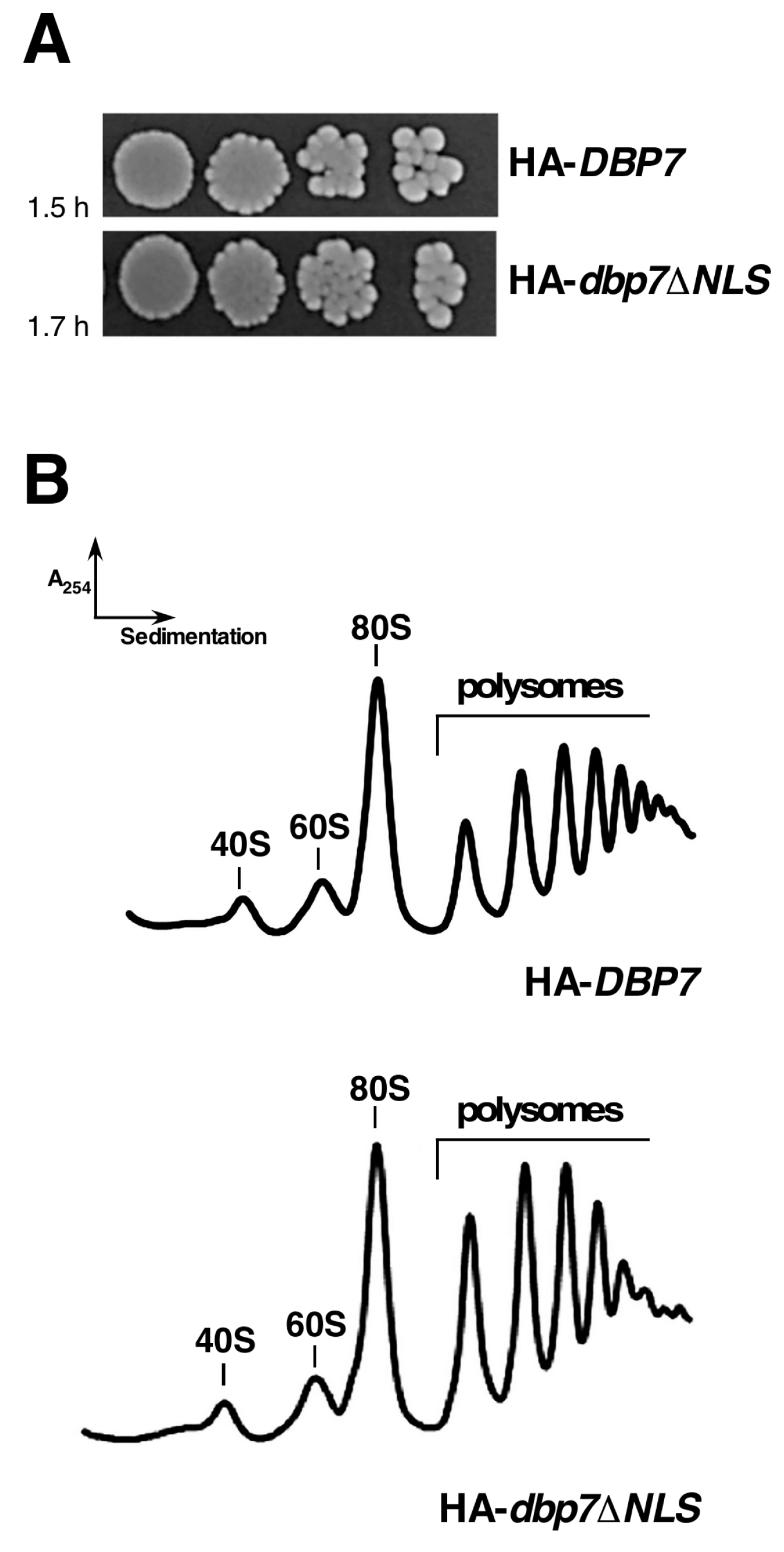
2.4. The NLS (V48 to S78 Sequence) of Dbp7 Is Required for Optimal Growth but Its Deletion Does Not Affect Polysome Profiles
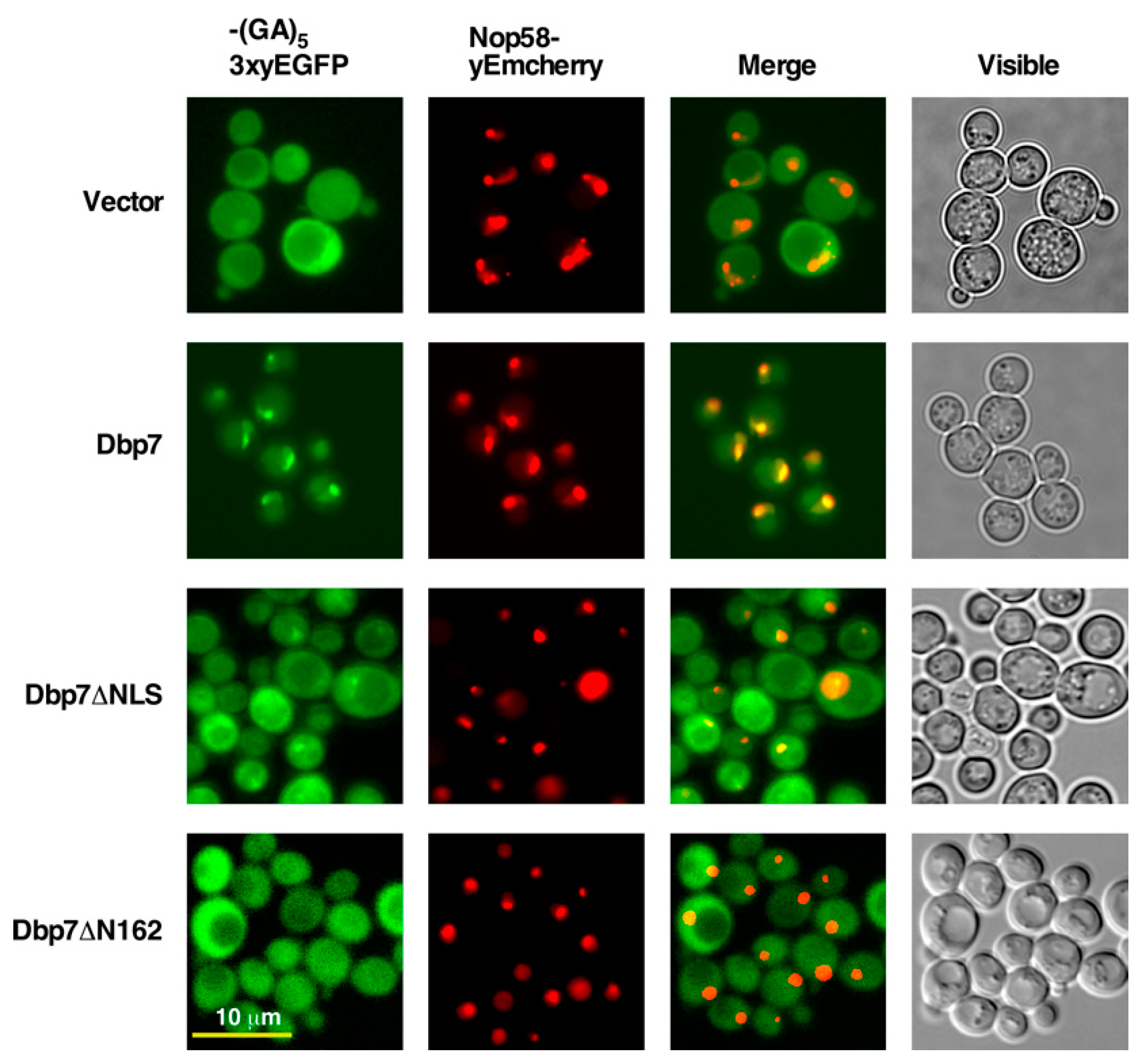
2.5. The N-Terminal Region of Dbp7 Is Necessary for Efficient Nuclear Import
2.6. The C-Terminal Extension of Dbp7 Is also Required for Efficient Cell Growth and for Production of 60S r-Subunits
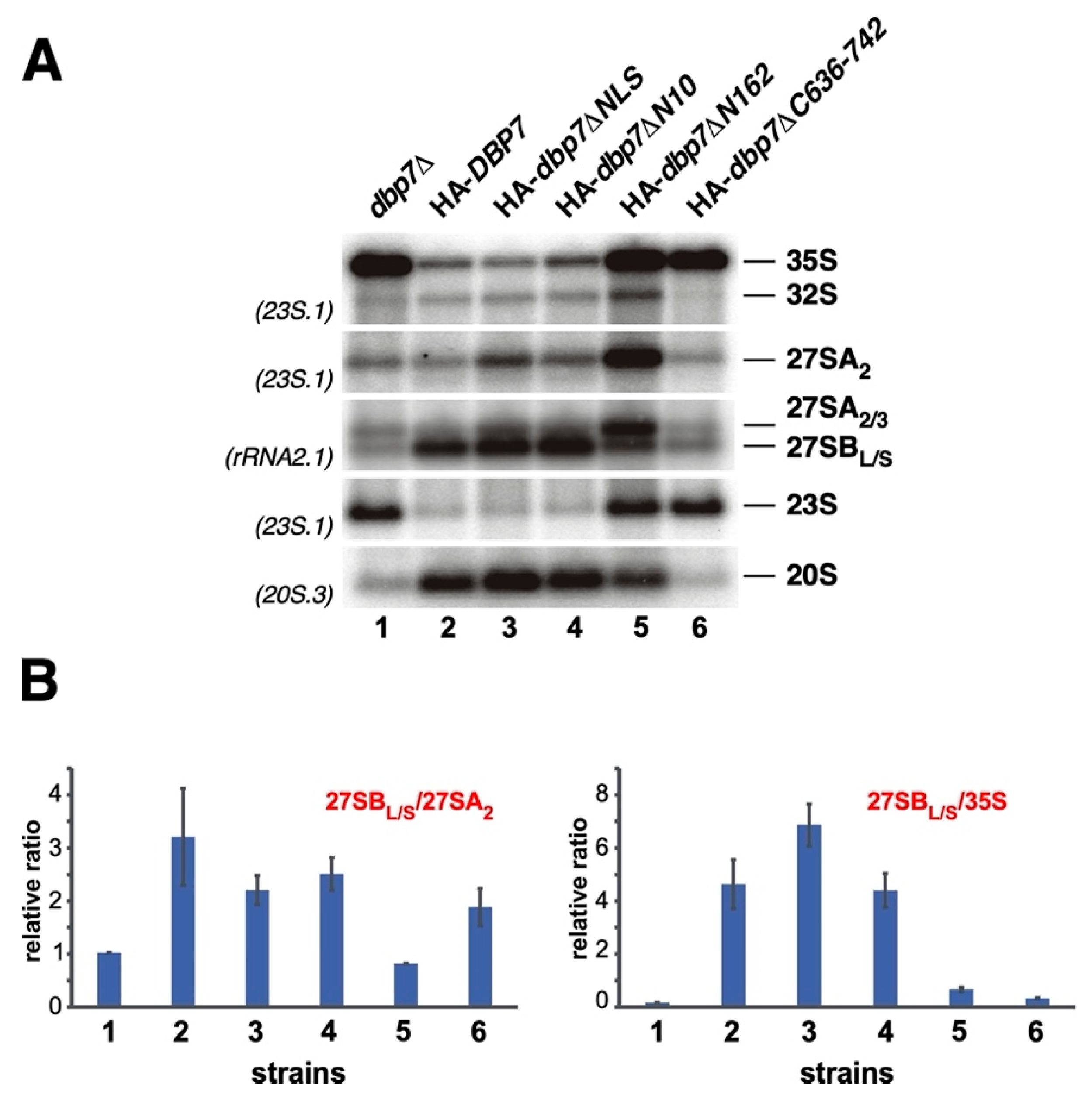
2.7. Importance of Terminal Extensions of Dbp7 in Pre-rRNA Processing
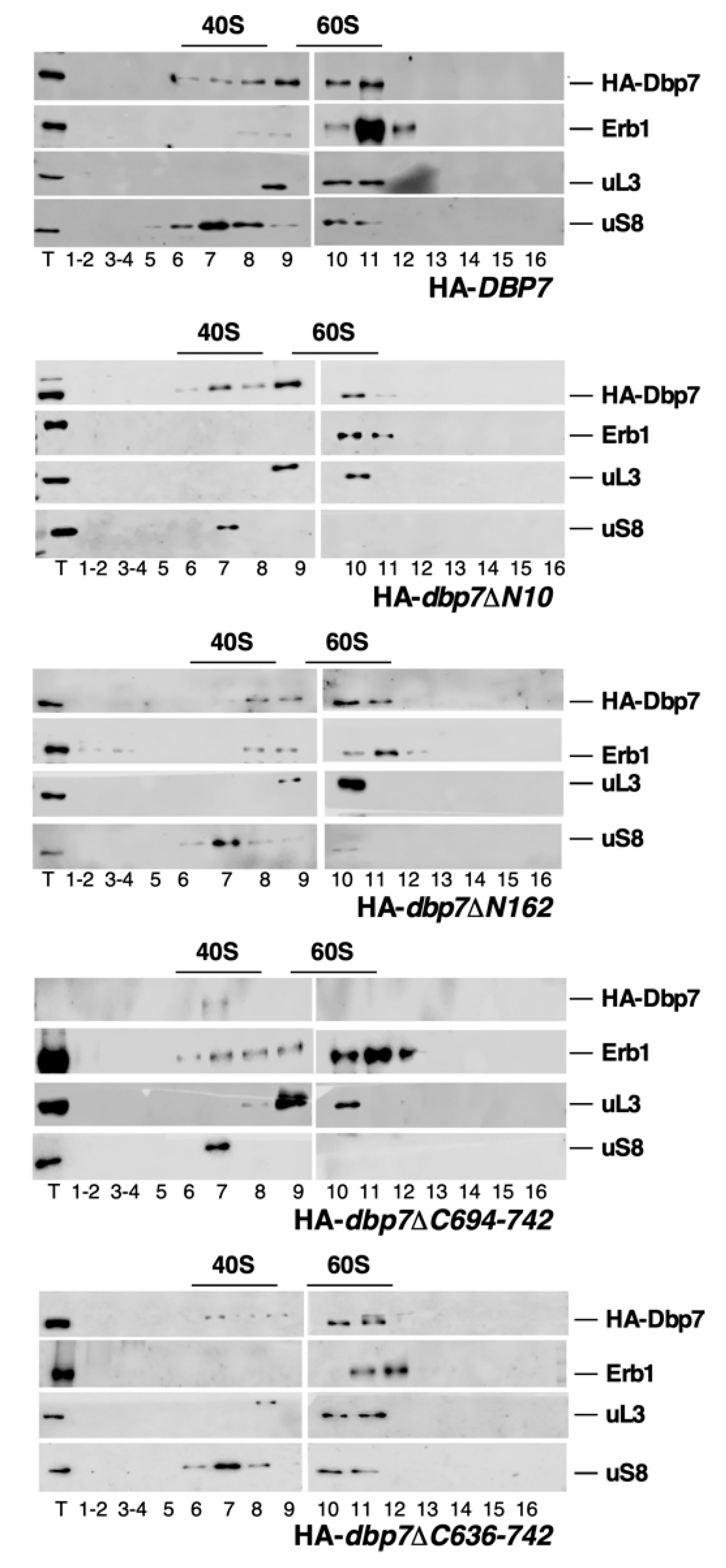
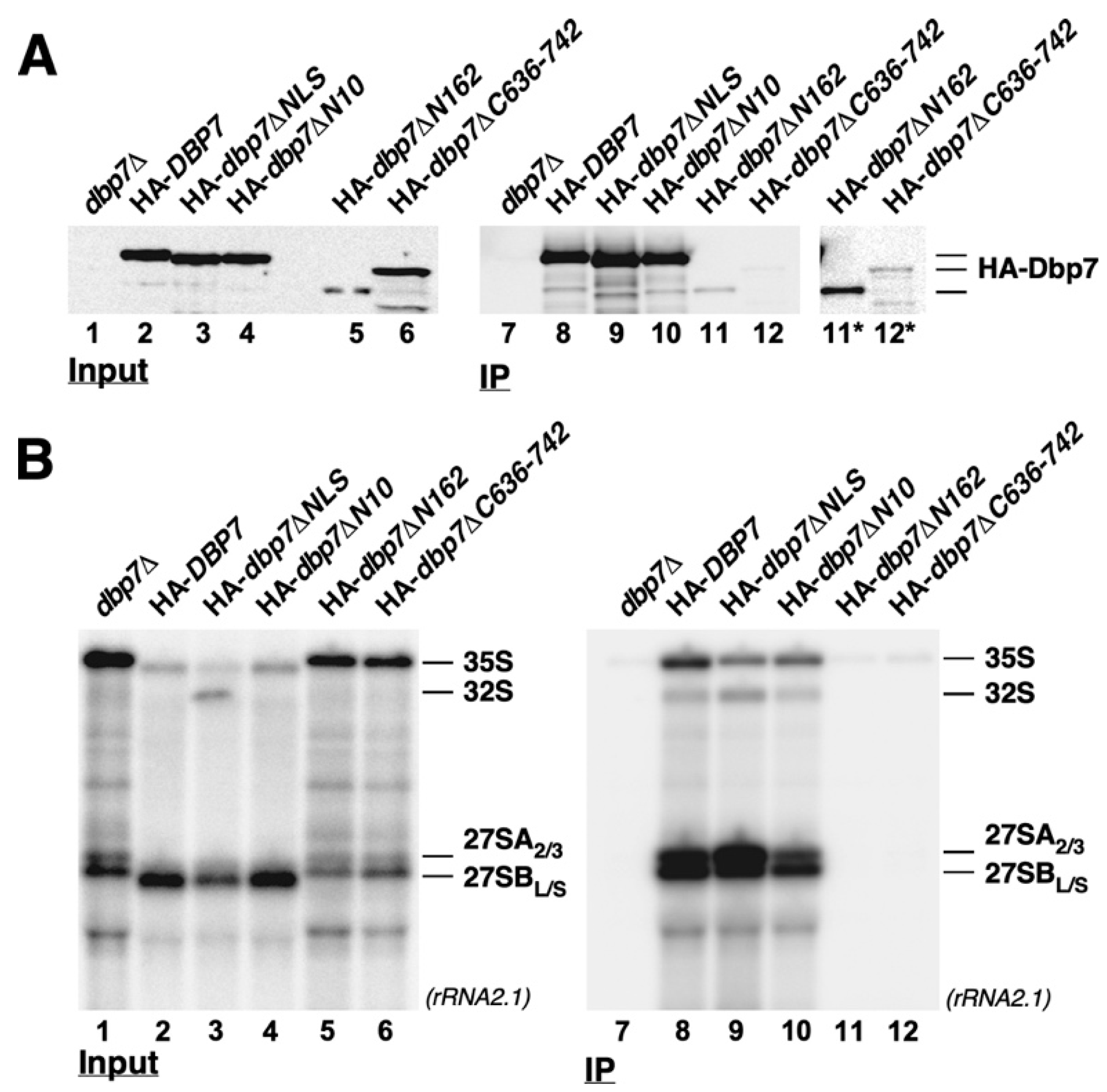
2.8. Influence of the N- or C-Terminal Extensions of Dbp7 on Its Association with Pre-Ribosomal Particles
3. Discussion
4. Materials and Methods
4.1. Strains and Plasmids
4.2. Media and Culturing
4.3. Nucleic Acid Manipulations
4.4. Protein Extraction and Western Blotting Analyses
4.5. RNA Extraction and Northern Blotting Analyses
4.6. Microscopy
4.7. Sucrose Gradient Centrifugation
4.8. Reproducibility
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, D.N.; Doudna Cate, J.H. The structure and function of the eukaryotic ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef]
- Kressler, D.; Hurt, E.; Bassler, J. Driving ribosome assembly. Biochim. Biophys. Acta 2010, 1803, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Woolford, J.L., Jr.; Baserga, S.J. Ribosome biogenesis in the yeast Saccharomyces cerevisiae. Genetics 2013, 195, 643–681. [Google Scholar] [CrossRef]
- Rodríguez-Galán, O.; García-Gómez, J.J.; de la Cruz, J. Yeast and human RNA helicases involved in ribosome biogenesis: Current status and perspectives. Biochim. Biophys. Acta-Gene Regul. Mech. 2013, 1829, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Straub, A.U.; Doebele, C.; Bohnsack, M.T. DExD/H-box RNA helicases in ribosome biogenesis. RNA Biol. 2012, 10, 4–18. [Google Scholar] [CrossRef]
- Mitterer, V.; Pertschy, B. RNA folding and functions of RNA helicases in ribosome biogenesis. RNA Biol. 2022, 19, 781–810. [Google Scholar] [CrossRef] [PubMed]
- Singleton, M.R.; Dillingham, M.S.; Wigley, D.B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 2007, 76, 23–50. [Google Scholar] [CrossRef]
- Jankowsky, E. RNA helicases at work: Binding and rearranging. Trends Biochem. Sci. 2011, 36, 19–29. [Google Scholar] [CrossRef]
- Fairman-Williams, M.E.; Guenther, U.P.; Jankowsky, E. SF1 and SF2 helicases: Family matters. Curr. Opin. Struct. Biol. 2010, 2010, 313–324. [Google Scholar] [CrossRef]
- Pyle, A.M. Translocation and unwinding mechanisms of RNA and DNA helicases. Annu. Rev. Biophys. 2008, 37, 317–336. [Google Scholar] [CrossRef]
- Bourgeois, C.F.; Mortreux, F.; Auboeuf, D. The multiple functions of RNA helicases as drivers and regulators of gene expression. Nat. Rev. Mol. Cell. Biol. 2016, 17, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Jarmoskaite, I.; Russell, R. DEAD-box proteins as RNA helicases and chaperones. Wiley Interdiscip. Rev. RNA 2011, 2, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Jarmoskaite, I.; Russell, R. RNA helicase proteins as chaperones and remodelers. Annu. Rev. Biochem. 2014, 83, 697–725. [Google Scholar] [CrossRef] [PubMed]
- Leitão, A.L.; Costa, M.C.; Enguita, F.J. Unzippers, resolvers and sensors: A structural and functional biochemistry tale of RNA helicases. Int. J. Mol. Sci. 2015, 16, 2269–2293. [Google Scholar] [CrossRef] [PubMed]
- Jankowsky, E.; Gross, C.H.; Shuman, S.; Pyle, A.M. Active disruption of an RNA-protein interaction by a DExH/D RNA helicase. Science 2000, 291, 121–124. [Google Scholar] [CrossRef]
- Linder, P.; Jankowsky, E. From unwinding to clamping-the DEAD box RNA helicase family. Nat. Rev. Mol. Cell. Biol. 2011, 12, 505–516. [Google Scholar] [CrossRef]
- Putnam, A.A.; Jankowsky, E. DEAD-box helicases as integrators of RNA, nucleotide and protein binding. Biochim. Biophys. Acta 2013, 1829, 884–893. [Google Scholar] [CrossRef]
- Schwer, B. A new twist on RNA helicases: DExH/D box proteins as RNPases. Nat. Struct. Biol. 2001, 8, 113–116. [Google Scholar] [CrossRef]
- Linder, P. Dead-box proteins: A family affair--active and passive players in RNP-remodeling. Nucleic Acids Res. 2006, 34, 4168–4180. [Google Scholar] [CrossRef]
- de la Cruz, J.; Kressler, D.; Linder, P. Unwinding RNA in Saccharomyces cerevisiae: DEAD-box proteins and related families. Trends Biochem. Sci. 1999, 24, 192–198. [Google Scholar] [CrossRef]
- Jankowsky, A.; Guenther, U.P.; Jankowsky, E. The RNA helicase database. Nucleic Acids Res. 2011, 39, D338–D341. [Google Scholar] [CrossRef] [PubMed]
- Rocak, S.; Linder, P. DEAD-box proteins: The driving forces behind RNA metabolism. Nat. Rev. Mol.Cell. Biol. 2004, 5, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Koonin, E.V. Helicases: Amino acid sequence comparisons and structure-function relationships. Curr. Opin. Struct. Biol. 1993, 3, 419–429. [Google Scholar] [CrossRef]
- Tanner, N.K.; Linder, P. DExD/H box RNA helicases: From generic motors to specific dissociation functions. Mol. Cell 2001, 8, 251–262. [Google Scholar] [CrossRef]
- Srikant, S. Evolutionary history of ATP-binding cassette proteins. FEBS Lett. 2020, 594, 3882–3897. [Google Scholar] [CrossRef]
- Sloan, K.E.; Bohnsack, M.T. Unravelling the Mechanisms of RNA Helicase Regulation. Trends Biochem. Sci. 2018, 43, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Banroques, J.; Cordin, O.; Doere, M.; Linder, P.; Tanner, N.K. Analyses of the functional regions of DEAD-box RNA “helicases” with deletion and chimera constructs tested in vivo and In vitro. J. Mol. Biol. 2011, 413, 451–472. [Google Scholar] [CrossRef]
- Chang, L.C.; Lee, F.J. The RNA helicase Dhh1p cooperates with Rbp1p to promote porin mRNA decay via its non-conserved C-terminal domain. Nucleic Acids Res. 2012, 40, 1331–1344. [Google Scholar] [CrossRef]
- Wang, Y.; Guthrie, C. PRP16, a DEAH-box RNA helicase, is recruited to the spliceosome primarily via its nonconserved N-terminal domain. RNA 1998, 4, 1216–1229. [Google Scholar] [CrossRef]
- van Nues, R.W.; Beggs, J.D. Functional contacts with a range of splicing proteins suggest a central role for Brr2p in the dynamic control of the order of events in spliceosomes of Saccharomyces cerevisiae. Genetics 2001, 157, 1451–1467. [Google Scholar] [CrossRef]
- Mohr, G.; Del Campo, M.; Mohr, S.; Yang, Q.; Jia, H.; Jankowsky, E.; Lambowitz, A.M. Function of the C-terminal domain of the DEAD-box protein Mss116p analyzed in vivo and In vitro. J. Mol. Biol. 2008, 375, 1344–1364. [Google Scholar] [CrossRef] [PubMed]
- Mohr, G.; Del Campo, M.; Turner, K.G.; Gilman, B.; Wolf, R.Z.; Lambowitz, A.M. High-throughput genetic identification of functionally important regions of the yeast DEAD-box protein Mss116p. J. Mol. Biol. 2011, 413, 952–972. [Google Scholar] [CrossRef] [PubMed]
- Kudlinzki, D.; Schmitt, A.; Christian, H.; Ficner, R. Structural analysis of the C-terminal domain of the spliceosomal helicase Prp22. Biol. Chem. 2012, 393, 1131–1140. [Google Scholar] [CrossRef]
- Roychowdhury, A.; Joret, C.; Bourgeois, G.; Heurgué-Hamard, V.; Lafontaine, D.L.J.; Graille, M. The DEAH-box RNA helicase Dhr1 contains a remarkable carboxyl terminal domain essential for small ribosomal subunit biogenesis. Nucleic Acids Res. 2019, 47, 7548–7563. [Google Scholar] [CrossRef] [PubMed]
- Walbott, H.; Mouffok, S.; Capeyrou, R.; Lebaron, S.; Humbert, O.; van Tilbeurgh, H.; Henry, Y.; Leulliot, N. Prp43p contains a processive helicase structural architecture with a specific regulatory domain. EMBO J. 2010, 29, 2194–2204. [Google Scholar] [CrossRef]
- Jackson, R.N.; Klauer, A.A.; Hintze, B.J.; Robinson, H.; van Hoof, A.; Johnson, S.J. The crystal structure of Mtr4 reveals a novel arch domain required for rRNA processing. EMBO J. 2010, 29, 2205–2216. [Google Scholar] [CrossRef]
- Daugeron, M.-C.; Linder, P. Dbp7p, a putative ATP-dependent RNA helicase from Saccharomyces cerevisiae, is required for 60S ribosomal subunit assembly. RNA 1998, 4, 566–581. [Google Scholar] [CrossRef]
- Aquino, G.R.R.; Hackert, P.; Krogh, N.; Pan, K.T.; Jaafar, M.; Henras, A.K.; Nielsen, H.; Urlaub, H.; Bohnsack, K.E.; Bohnsack, M.T. The RNA helicase Dbp7 promotes domain V/VI compaction and stabilization of inter-domain interactions during early 60S assembly. Nat. Commun. 2021, 12, 6152. [Google Scholar] [CrossRef]
- Jaafar, M.; Contreras, J.; Dominique, C.; Martín-Villanueva, S.; Capeyrou, R.; Vitali, P.; Rodríguez-Galán, O.; Velasco, C.; Humbert, O.; Watkins, N.J.; et al. Association of snR190 snoRNA chaperone with early pre-60S particles is regulated by the RNA helicase Dbp7 in yeast. Nat. Commun. 2021, 12, 6153. [Google Scholar] [CrossRef]
- Bernstein, K.A.; Granneman, S.; Lee, A.V.; Manickam, S.; Baserga, S.J. Comprehensive mutational analysis of yeast DEXD/H box RNA helicases involved in large ribosomal subunit biogenesis. Mol. Cell. Biol. 2006, 26, 1195–1208. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Kinoshita, K. PrDOS: Prediction of disordered protein regions from amino acid sequence. Nucleic Acids Res. 2007, 35, W460–W464. [Google Scholar] [CrossRef]
- Linding, R.; Jensen, L.J.; Diella, F.; Bork, P.; Gibson, T.J.; Russell, R.B. Protein disorder prediction: Implications for structural proteomics. Structure 2003, 11, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Helser, T.L.; Baan, R.A.; Dahlberg, A.E. Characterization of a 40S ribosomal subunit complex in polyribosomes of Saccharomyces cerevisiae treated with cycloheximide. Mol. Cell. Biol. 1981, 1, 51–57. [Google Scholar]
- Rotenberg, M.; Moritz, M.; Woolford, J.L., Jr. Depletion of Saccharomyces cerevisiae ribosomal protein L16 causes a decrease in 60S ribosomal subunits and formation of half-mer polysomes. Genes Dev. 1988, 2, 160–172. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef]
- Kumar, M.; Gouw, M.; Michael, S.; Samano-Sánchez, H.; Pancsa, R.; Glavina, J.; Diakogianni, A.; Valverde, J.A.; Bukirova, D.; Calyseva, J.; et al. ELM-the eukaryotic linear motif resource in 2020. Nucleic Acids Res. 2020, 48, D296–D306. [Google Scholar] [CrossRef]
- Lin, J.R.; Hu, J. SeqNLS: Nuclear localization signal prediction based on frequent pattern mining and linear motif scoring. PLoS One 2013, 8, e76864. [Google Scholar] [CrossRef]
- Tang, L.; Sahasranaman, A.; Jakovljevic, J.; Schleifman, E.; Woolford, J.L., Jr. Interactions among Ytm1, Erb1, and Nop7 required for assembly of the Nop7-subcomplex in yeast preribosomes. Mol. Biol. Cell 2008, 19, 2844–2856. [Google Scholar] [CrossRef] [PubMed]
- Wegrecki, M.; Rodríguez-Galán, O.; de la Cruz, J.; Bravo, J. The structure of Erb1-Ytm1 complex reveals the functional importance of a high-affinity binding between two beta-propellers during the assembly of large ribosomal subunits in eukaryotes. Nucleic Acids Res. 2015, 43, 11017–11030. [Google Scholar] [CrossRef] [PubMed]
- Cordin, O.; Banroques, J.; Tanner, N.K.; Linder, P. The DEAD-box protein family of RNA helicases. Gene 2006, 367, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Granneman, S.; Bernstein, K.A.; Bleichert, F.; Baserga, S.J. Comprehensive mutational analysis of yeast DEXD/H box RNA helicases required for small ribosomal subunit synthesis. Mol. Cell. Biol. 2006, 26, 1183–1194. [Google Scholar] [CrossRef]
- Tanner, N.K.; Cordin, O.; Banroques, J.; Doere, M.; Linder, P. The Q motif: A newly identified motif in DEAD box helicases may regulate ATP binding and hydrolysis. Mol. Cell 2003, 11, 127–138. [Google Scholar] [CrossRef]
- Rocak, S.; Emery, B.; Tanner, N.K.; Linder, P. Characterization of the ATPase and unwinding activities of the yeast DEAD-box protein Has1p and the analysis of the roles of the conserved motifs. Nucleic Acids Res. 2005, 33, 999–1009. [Google Scholar] [CrossRef]
- Schmid, S.R.; Linder, P. Translation initiation factor 4A from Saccharomyces cerevisiae: Analysis of residues conserved in the D-E-A-D family of RNA helicases. Mol. Cell. Biol. 1991, 11, 3463–3471. [Google Scholar]
- Hotz, H.-R.; Schwer, B. Mutational analysis of the yeast DEAH-box splicing factor Prp16. Genetics 1998, 149, 807–815. [Google Scholar] [CrossRef]
- Schwer, B.; Meszaros, T. RNA helicase dynamics in pre-mRNA splicing. EMBO J. 2000, 19, 6582–6591. [Google Scholar] [CrossRef]
- Kossen, K.; Karginov, F.V.; Uhlenbeck, O.C. The carboxy-terminal domain of the DExDH protein YxiN is sufficient to confer specificity for 23S rRNA. J. Mol. Biol. 2002, 324, 625–636. [Google Scholar] [CrossRef]
- Schneider, S.; Schwer, B. Functional domains of the yeast splicing factor Prp22p. J. Biol. Chem. 2001, 276, 21184–21191. [Google Scholar] [CrossRef]
- Pillet, B.; García-Gómez, J.J.; Pausch, P.; Falquet, L.; Bange, G.; de la Cruz, J.; Kressler, D. The dedicated chaperone Acl4 escorts ribosomal protein Rpl4 to its nuclear pre-60S assembly site. PLoS Genet. 2015, 11, e1005565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordonné, R. Functional characterization of nuclear localization signals in yeast Sm proteins. Mol. Cell. Biol. 2000, 20, 7943–7954. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Ku, W.C.; Chen, Y.C.; Chang, Y.L.; Chu, C.Y. Dual mechanisms regulate the nucleocytoplasmic localization of human DDX6. Sci. Rep. 2017, 7, 42853. [Google Scholar] [CrossRef] [PubMed]
- Edwalds-Gilbert, G.; Kim, D.H.; Silverman, E.; Lin, R.J. Definition of a spliceosome interaction domain in yeast Prp2 ATPase. RNA 2004, 10, 210–220. [Google Scholar] [CrossRef]
- Martin, A.; Schneider, S.; Schwer, B. Prp43 is an essential RNA-dependent ATPase required for release of lariat-intron from the spliceosome. J. Biol. Chem. 2002, 277, 17743–17750. [Google Scholar] [CrossRef]
- Mouffok, S.; Capeyrou, R.; Belhabich-Baumas, K.; Joret, C.; Henras, A.K.; Humbert, O.; Henry, Y. The G-patch activators Pfa1 and PINX1 exhibit different modes of interaction with the Prp43 RNA helicase. RNA Biol. 2021, 18, 510–522. [Google Scholar] [CrossRef]
- Absmeier, E.; Wollenhaupt, J.; Mozaffari-Jovin, S.; Becke, C.; Lee, C.T.; Preussner, M.; Heyd, F.; Urlaub, H.; Luhrmann, R.; Santos, K.F.; et al. The large N-terminal region of the Brr2 RNA helicase guides productive spliceosome activation. Genes Dev. 2015, 29, 2576–2587. [Google Scholar] [CrossRef]
- Silverman, E.J.; Maeda, A.; Wei, J.; Smith, P.; Beggs, J.D.; Lin, R.J. Interaction between a G-patch protein and a spliceosomal DEXD/H-box ATPase that is critical for splicing. Mol. Cell. Biol. 2004, 24, 10101–10110. [Google Scholar] [CrossRef]
- de la Cruz, J.; Lacombe, T.; Deloche, O.; Linder, P.; Kressler, D. The putative RNA helicase Dbp6p functionally interacts with Rpl3p, Nop8p and the novel trans-acting factor Rsa3p during biogenesis of 60S ribosomal subunits in Saccharomyces cerevisiae. Genetics 2004, 166, 1687–1699. [Google Scholar] [CrossRef]
- Rosado, I.V.; Dez, C.; Lebaron, S.; Caizergues-Ferrer, M.; Henry, Y.; de la Cruz, J. Characterization of Saccharomyces cerevisiae Npa2p (Urb2p) reveals a low-molecular-mass complex containing Dbp6p, Npa1p (Urb1p), Nop8p, and Rsa3p involved in early steps of 60S ribosomal subunit biogenesis. Mol. Cell. Biol. 2007, 27, 1207–1221. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.J.; Rothstein, R. Elevated recombination rates in transcriptionally active DNA. Cell 1989, 56, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.; Dawson, D.; Stearns, T. Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2000. [Google Scholar]
- Gietz, D.; St. Jean, A.; Woods, R.A.; Schiestl, R.H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res. 1992, 20, 1425. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, R.S.; Boeke, J.D. In vitro mutagenesis and plasmid shuffling: From cloned gene to mutant yeast. Methods Enzymol. 1991, 194, 302–318. [Google Scholar]
- Ho, S.N.; Hunt, H.D.; Horton, R.M.; Pullen, J.K.; Pease, L.R. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene 1989, 77, 51–59. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Joret, C.; Capeyrou, R.; Belhabich-Baumas, K.; Plisson-Chastang, C.; Ghandour, R.; Humbert, O.; Fribourg, S.; Leulliot, N.; Lebaron, S.; Henras, A.K.; et al. The Npa1p complex chaperones the assembly of the earliest eukaryotic large ribosomal subunit precursor. PLoS Genet. 2018, 14, e1007597. [Google Scholar] [CrossRef]
- Kressler, D.; de la Cruz, J.; Rojo, M.; Linder, P. Fal1p is an essential DEAD-box protein involved in 40S-ribosomal-subunit biogenesis in Saccharomyces cerevisiae. Mol. Cell. Biol. 1997, 17, 7283–7294. [Google Scholar] [CrossRef]
- de la Cruz, J.; Kressler, D.; Rojo, M.; Tollervey, D.; Linder, P. Spb4p, an essential putative RNA helicase, is required for a late step in the assembly of 60S ribosomal subunits in Saccharomyces cerevisiae. RNA 1998, 4, 1268–1281. [Google Scholar] [CrossRef]
- Gietz, R.D.; Sugino, A. New yeast-Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking sixbase pair restriction sites. Gene 1988, 74, 527–534. [Google Scholar] [CrossRef]
- Koch, B.; Mitterer, V.; Niederhauser, J.; Stanborough, T.; Murat, G.; Rechberger, G.; Bergler, H.; Kressler, D.; Pertschy, B. Yar1 protects the ribosomal protein Rps3 from aggregation. J. Biol. Chem. 2012, 287, 21806–21815. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras, J.; Ruiz-Blanco, Ó.; Dominique, C.; Humbert, O.; Henry, Y.; Henras, A.K.; de la Cruz, J.; Villalobo, E. The Terminal Extensions of Dbp7 Influence Growth and 60S Ribosomal Subunit Biogenesis in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2023, 24, 3460. https://doi.org/10.3390/ijms24043460
Contreras J, Ruiz-Blanco Ó, Dominique C, Humbert O, Henry Y, Henras AK, de la Cruz J, Villalobo E. The Terminal Extensions of Dbp7 Influence Growth and 60S Ribosomal Subunit Biogenesis in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2023; 24(4):3460. https://doi.org/10.3390/ijms24043460
Chicago/Turabian StyleContreras, Julia, Óscar Ruiz-Blanco, Carine Dominique, Odile Humbert, Yves Henry, Anthony K. Henras, Jesús de la Cruz, and Eduardo Villalobo. 2023. "The Terminal Extensions of Dbp7 Influence Growth and 60S Ribosomal Subunit Biogenesis in Saccharomyces cerevisiae" International Journal of Molecular Sciences 24, no. 4: 3460. https://doi.org/10.3390/ijms24043460