
AR and PI3K/AKT in Prostate Cancer: A Tale of Two Interconnected Pathways

1
Department of Molecular Medicine, Sapienza University of Rome, Viale Regina Elena 324, 00161 Rome, Italy
2
Department of Urology, Sapienza University of Rome, Viale del Policlinico, 00161 Rome, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(3), 2046; https://doi.org/10.3390/ijms24032046
Submission received: 30 December 2022
/
Revised: 12 January 2023
/
Accepted: 18 January 2023
/
Published: 20 January 2023
(This article belongs to the Special Issue Prostate Cancer: Pathology, Pathobiology and Therapy)
Abstract
:Prostate cancer (PCa) is the most common cancer in men. The androgen receptor (AR) has a pivotal role in the pathogenesis and progression of PCa. Many therapies targeting AR signaling have been developed over the years. AR signaling inhibitors (ARSIs), including androgen synthesis inhibitors and AR antagonists, have proven to be effective in castration-sensitive PCa (CSPC) and improve survival, but men with castration-resistant PCa (CRPC) continue to have a poor prognosis. Despite a good initial response, drug resistance develops in almost all patients with metastatic CRPC, and ARSIs are no longer effective. Several mechanisms confer resistance to ARSI and include AR mutations but also hyperactivation of other pathways, such as PI3K/AKT/mTOR. This pathway controls key cellular processes, including proliferation and tumor progression, and it is the most frequently deregulated pathway in human cancers. A significant interaction between AR and the PI3K/AKT/mTOR signaling pathway has been shown in PCa. This review centers on the current scene of different AR and PI3K signaling pathway inhibitors, either as monotherapy or in combination treatments in PCa, and the treatment outcomes involved in both preclinical and clinical trials. A PubMed-based literature search was conducted up to November 2022. The most relevant and recent articles were selected to provide essential information and current evidence on the crosstalk between AR and the PI3K signaling pathways. The ClinicalTrials.gov registry was used to report information about clinical studies and their results using the Advanced research tool, filtering for disease and target.
1. Introduction
Prostate cancer (PCa) is the most common cancer in men and the second-leading cause of cancer [1].
A more widespread prevention through screening with the prostate-specific antigen (PSA) has shown an increase in PCa incidence rate [2]. Most cases are diagnosed in the early stages (78% in localized stages and 12% with regional involvement), but 5% of cases are identified as already metastatic [3].
The time of diagnosis influences the prognosis. PCa can be a serious disease, but the majority of diagnosed patients do not die from it unless it becomes more aggressive and castration-resistant. Patients with localized PCa have a survival rate of 90% versus only 67% in metastatic PCa [4,5].
Treatment for localized PCa includes surgery (radical prostatectomy (RP)), radiotherapy (RT), or active surveillance, depending on the stage of the disease and other factors [3]. Despite the high rates of progression-free survival (PFS), about 20 to 30% of patients with PCa can experience cancer recurrence [6,7], characterized by rising serum PSA [4]. Clinical relapse occurs, on average, after 5 years (1–8 years) [8,9]. These patients with relapsed disease are either treated with salvage radiation therapy (SRT) or androgen deprivation therapy (ADT) [10]. ADT involves biochemical inhibition of male sex hormones, blocking the production of androgens in the testes via the hypothalamus–pituitary–gonadal (HPG) axis with luteinizing hormone-releasing hormone (LHRH) agonists (e.g., leuprolide) or antagonists (e.g., degarelix) [11,12,13].
Despite a good initial response, androgen-independent and castration-resistant prostate cancer (CRPC) can occur and the outcome is poor [14,15,16]. Both patients with CRPC and with metastatic disease at diagnosis can have improvements in overall survival (OS) with many treatments that include chemotherapy, such as docetaxel and, recently, cabazitaxel; radium-223, used for the treatment of bone; and denosumab, an antagonist of receptor activator of nuclear factor kappa-B ligand (RANKL). There is also increased interest in new androgen synthesis inhibitors, such as abiraterone acetate, or second-generation competitive AR antagonists, enzalutamide, as well as apalutamide and darolutamide [17,18,19,20,21,22,23].
Furthermore, clinical studies on agents that target the immune checkpoint, such as cytotoxic T-lymphocyte-associated protein 4 (CTLA4), programmed cell death protein 1 (PD1), or programmed death-ligand 1 (PD-L1), have been evaluated in the clinic [24]. To date, the Food and Drug Administration (FDA) has approved Sipuleucel-T immunotherapy, which is the only dendritic cell vaccine for asymptomatic or minimally symptomatic CRPC without visceral metastases [25]. In recent years, genetic DNA repair alterations were also found in some PCa patients and, in these cases, poly ADP ribose polymerase (PARP) inhibitors, including olaparib and rucaparib, both approved by the FDA, or talazoparib and niraparib have shown efficacy [26].
Hence, in this complex scenario focused on identifying both disease biomarkers and novel and effective treatments that will benefit patients with PCa, this review centers on the current scene of different Phosphatidylinositol-3-kinase (PI3K) signaling pathway inhibitors, either as monotherapy or in combination treatments in PCa, and the treatment outcomes involved in both preclinical and clinical trials.
2. Androgen Receptor Signaling Pathway
2.1. Androgens and Androgen Receptor
AR signaling has a pivotal role in prostate development and homeostasis as well as in prostate tumorigenesis [27,28], due to the fact that inhibition of AR signaling remains the mainstay therapeutic target in PCa [28].
Briefly, androgens, such as testosterone, are synthesized primarily by the Leydig cells in the testes and are tightly regulated by the hypothalamic–pituitary–gonadal (HPG) axis.
Once produced, testosterone mostly circulates bound to serum sex hormone-binding globulin (SHBG) and albumin [29]. Only the free form enters prostate cells. Intracellularly, testosterone is converted into a more potent 5α-reduced metabolite of testosterone, 5α-dihydrotestosterone (DHT), essential for the growth and survival of these cells. Circulating testosterone and DHT represent the main native agonists for AR.
AR is a member of the steroid and nuclear hormone receptor superfamily and is encoded by the AR gene mapped on the long arm of the X-chromosome (locus: Xq11-q12) [30,31]. The AR consists of four domains: an amino-terminal transcriptional domain (NTD), DNA-binding domain (DBD), a hinge region, and a carboxy-terminal ligand-binding domain (LBD) [32].
In the cytoplasm, AR is bound with heat-shock proteins (HSPs)-90, HSP-70, HSP-56, and other chaperone proteins to protect the receptor against degradation [32,33].
The binding of the ligand promotes the dissociation of AR from these complexes and AR can shuttle to the nucleus, where it dimerizes and binds to the androgen response elements (AREs) [33,34]. AREs also include genes that are involved in PSA expression, androgen biosynthesis, DNA synthesis and repair, cell cycle regulation, and proliferation [35,36].
Furthermore, AR transactivation is modulated by coregulators. At least 300 proteins have been identified that can coactivate or co-repress AR-dependent transcription [37].
In addition to the DNA-binding-dependent actions of the AR, commonly referred to as genomic or canonical, the complex androgen-AR can also trigger a signal through a non-DNA-binding-dependent mechanism, referred to as nongenomic. The non-canonical androgen signaling pathways can be classified as non-genomic pathways (triggered by androgens) and outlaw pathways (androgen-independent, but AR-dependent) [38].
Non-genomic androgen signaling pathways include androgen-induced activation of G-protein-coupled receptors (GPCRs), the protein kinase A (PKA) pathway, and via AR variants [39].
2.2. Therapies Targeted to AR Signaling
Considering that androgens and AR signals are essential regulators in the growth of the normal prostate and play a key role in PCa pathogenesis, they represent a target for many agents, used both as monotherapy or in combination, with the intent to treat and improve the outcome of distinct clinical phenotypes of prostate disease: localized, CSPC, mCSPC, CRPC, and mCRPC. Therapies that aim to inhibit AR signaling, collectively called AR signaling inhibitors (ARSIs), have undergone a significant advance since the 1940s. At that time, Huggins and Hodges demonstrated the therapeutic effect of gonadal testosterone deprivation via orchiectomy [40]. Nowadays, the available drugs target the gonadotropin-releasing hormone (GnRH) to prevent luteinizing hormone (LH) releasing, cytochrome P450 17α-hydroxylase/17,20-lyase (CYP17A1) to restrain androgen synthesis, or directly AR, to inhibit AR transcriptional activity [16,17,18,19,20].
2.2.1. Androgen Deprivation Therapy (ADT)
ADT via surgical or chemical castration has become crucial in the treatment of PCa and metastatic disease with the aim to suppress serum testosterone to castration levels and to block the activation of the AR [14,15]. Gonadotropin-releasing hormone (GnRH) agonists, such as histrelin, goserelin, leuprolide, and triptorelin, approved in the 1980s by the FDA, continue to be the mainstay of ADT. Their action is based on the activation of GnRH receptors, leading to an increase in follicle-stimulating hormone (FSH), LH, and, consequentially, testosterone production. Despite this, the continuous stimulation, which is the opposite of the physiological pulsatile action of GnRH receptors, leads to their downregulation and subsequent desensitization of the pituitary gland to the effects of GnRH [41]. Hence, testosterone production decreases to castration levels over 3 to 4 weeks [42]. These agents may be associated with an increased likelihood of adverse events compared to orchiectomy, including cardiovascular events, fractures, and peripheral arterial disease [43,44]. GnRH antagonists induce testosterone suppression by directly inhibiting GnRH receptors. Injectable degarelix [45] and relugolix [46], oral GnRH antagonists, were approved by the FDA in 2009 and 2020, respectively, for advanced PCa. It seems that the risk of cardiovascular events in men taking GnRH antagonists is lower compared to those taking GnRH agonists [47]. The androgen synthesis inhibitor targets CYP17, a member of the cytochrome P450 family. CYP17 catalyzes and converts precursor steroids to testosterone, DHT, and dehydroepiandrosterone (DHEA). Abiraterone acetate is a selective inhibitor of 17α-hydroxylase/C17,20-lyase (CYP17) and blocks the production of androgen in the adrenal glands, testes, placenta, as well as in PCa cells [48]. Because of interaction with numerous targets, steroid supplementation is necessary to overcome cortisol insufficiency and limit some adverse effects (AEs) [20]. Some studies have demonstrated that a dose of 800 mg once daily is effective in suppressing testosterone to below detectable levels in humans and provided preliminary insight into its safety profile [49]. This drug was FDA-approved in 2011 for patients with mCRPC who had received prior chemotherapy associated with prednisone, and in 2012, for patients with mCRPC and, together with ADT, for metastatic high-risk CSPC [49,50,51]. Furthermore, in the early 1980s, it was shown that castration levels of serum testosterone can be induced also by high doses of the imidazole antifungal agent ketoconazole (Nizoral), by blocking cytochrome P450 enzymes (including CYP17A1) [52].
2.2.2. Androgen Receptor Antagonists
Androgen receptor antagonists (ARAs) bind to the LBD of the AR and prevent the binding of testosterone and DHT, so that AR translocation into the nucleus is inhibited and, consequently, the interactions with AREs. The non-steroidal antiandrogens flutamide (Eulexin), nilutamide (Anandron) and bicalutamide (Casodex) represent that the first generation of ARAs do not completely block AR activity [53] and show a short-lived benefit [16]. Enzalutamide (MDV3100), a second-generation ARA, has higher affinity for the AR and minimal no-agonist activity [54]. After the encouraging results from several clinical trials [21,55], the FDA approved enzalutamide in the pre- and post-chemotherapy mCRPC (2012), CRPC (2018) as well as in mCSPC (2019). Apalutamide (ARN 509) has a similar mechanism of action as enzalutamide [56] but a greater efficacy [23]; the FDA approved apalutamide for the treatment of patients with non-metastatic (nm) CRPC (2018) and for patients with mCSPC (2019). Apalutamide significantly improves OS and radiographic progression-free survival (rPFS) in patients with mCSPC receiving ongoing ADT, as analyzed in TITAN [57]. Darolutamide (ODM-201) has a particular molecular structure that is distinct from other AR antagonists [58,59]. Darolutamide and its metabolite have a tighter binding to the AR than enzalutamide and apalutamide with lower toxicity and higher OS. Moreover, Darolutamide can also antagonize some mutations within AR, which confer resistance for enzalutamide and apalutamide. It is the last AR inhibitor approved by the FDA for treatment of nmCRPC (2019) and for mCSPC (2022) [55,56].
2.3. Alterations in AR Signaling
Despite these therapies suppressing most PCas, some high-risk prostate cancers gradually progress to CRPC and mCRPC. The AR signaling pathway plays a central role also in the progression of PCa during hormonal therapy [60,61,62]. Several mechanisms can restore AR signaling, including intracrine androgen synthesis, AR overexpression and amplification, point mutations, acquisition of constitutively active AR splice variants, deregulated AR coactivators/corepressors that sensitize AR in response to ligand binding, and, finally, ligand-independent signaling [3]. These alterations are not exclusive and can even coexist in the same patient [62].
2.3.1. Amplification of the AR Gene and Overexpression of the AR Protein
AR gene amplification leads to an overexpression of the AR protein and is the most common genetic mutation among patients with CRPC [63]. Results from studies using fluorescence in situ hybridization (FISH) have shown that AR amplification is the most frequent genetic alteration detected in CRPC, observed in more than 50% of cases, while it is only rarely detected in untreated primary tumors, suggesting that AR amplification could be an adaptive response to ADT. AR gene amplification was also analyzed at the mRNA level. By using reverse-transcription polymerase chain reaction (RT-PCR), it has been observed that the expression of AR mRNA in CRPC with AR amplification was two-fold higher than CRPC without AR amplification. Elevated AR protein levels were also linked to CRPC [64].
Furthermore, it has been shown that AR gene amplification is frequently detected in the circulating tumor cells (CTCs) of patients with CRPC [65]. As well, AR amplification is more common in patients resistant to both enzalutamide and abiraterone (more enzalutamide than abiraterone) [66], suggesting that AR amplification could be a mechanism responsible for ADT resistance [34]. Finally, ADT may induce upregulation of AR transcription by disrupting the AR negative-feedback loop, then resulting in elevated AR-FL and AR-Vs protein levels [67,68]. Moreover, in CRPC, decreased androgen levels may relieve AR suppression of genes mediating DNA synthesis, thereby contributing to tumor cell proliferation [67].
2.3.2. Point Mutations in Androgen Receptor
Point mutations in the AR gene are rarely detectable in early-stage PCa but are detected in 15–20% of CRPC patients and up to 40% of CRPC patients treated with AR antagonists, most frequently in the LBD, followed by the NTD [66,69]. The acquisition of AR mutations results in broadening ligand specificity by weak adrenal androgens and other steroid hormones, including DHEA, progesterone, estrogen, and glucocorticoids, as well as in reversing antagonists into agonists [70]. T878A was the first identified point mutation resulting in the loss of specificity for the agonist. Progesterone, estrogens, flutamide, bicalutamide, and enzalutamide can activate AR carrying the T878A point mutation [71]. Moreover, T878A also causes resistance to second-generation AR agonists [66]. Resistance to enzalutamide is also induced by the point mutation F876L, in the LBD domain [64,65,66]. Finally, H875Y, T878A, and T878S mutations detected in CRPC patients induce agonist effects by enzalutamide and apalutamide [70]. Mutations, such as T878A or L702H, have been identified in the plasma of 13% of CRPC patients progressing during abiraterone treatment. Other AR point mutations, T877A, L702H, and H875Y, have been detected in cell-free DNA from patients with CRPC and have also been associated with resistance to abiraterone and enzalutamide [72].
2.3.3. AR Splice Variants
Androgen receptor splice variants (AR-Vs) are the main causes of abnormalities in AR regulation [73,74]. AR-Vs are derived by a splicing mechanism from full length and are truncated receptors with a modified LBD domain, but normal NTD and DBD domains, required for endogenous AR functions. The effect of the previously reported modification is that AR may still mediate signaling in the absence of the ligand, but PC cells become resistant to many ARSIs, including abiraterone and enzalutamide. Generally, these AR-Vs have been associated with poor prognosis in advanced PCa [75]. AR-V7 is the most prevalent variant among more than 20 AR-Vs identified in CRPC, followed by the exon-skipping AR-v567 [74,76]. AR-V7 has been detected in 75% of mCRPC but only rarely in early-stage disease (<1%) [77]. This variant has also been associated with an increased risk of recurrent disease after prostatectomy in CSCR and with low survival in CRPC. Furthermore, the identification of AR-V7 mRNA levels in whole blood of mCRPC patients receiving abiraterone has been correlated with poor outcome [78]. This and other studies support that AR-V7 detection in patients with CRPC could be correlated with resistance to enzalutamide and abiraterone [79]. Some studies are aimed at better understanding if AR-V7 protein expression is simply associated with enhanced AR-FL expression as a marker of resistance to ARSI or whether a critical level of AR-V7 is required for such ARSI refractory lethal cancer growth [80,81,82]. Some studies have focused on understanding the functional differences, as compared to canonical AR-FL. AR-V7 exhibits fast nuclear import kinetics via a pathway distinct from the nuclear localization signal-dependent importin-α/β pathway used by AR-FL. The dimerization box domain, known to mediate AR dimerization and transactivation, is required for AR-V7 nuclear import but not for AR-FL. In the nucleus, AR-V7 is transcriptionally active and exhibits unusually high intranuclear mobility and transient chromatin interactions, unlike the stable chromatin association of liganded AR-FL. The high intranuclear mobility of AR-V7 together with its high transcriptional output suggest a hit-and-run mode of transcription [83,84]. These mechanisms regulating AR-V7 activity could offer the opportunity to develop selective therapeutic interventions [85].
AR-V567es is one of the two most frequently observed AR variants. It has exons 5–7 spliced out and only contains a small portion of the LBD. Results from studies using transgenic mice have shown that ARV567es induces carcinogenesis and accelerates tumor progression [86]. Studies are ongoing to explore if AR-V567es confers relatively greater taxane sensitivity than AR-V7 [87,88].
3. PI3K/AKT/mTOR Signaling Pathway
The PI3K/AKT/mammalian target of rapamycin (mTOR) pathway is considered to be a pivotal intracellular signaling pathway and its hyperactivity is correlated with tumor progression in a wide assortment of cancers, counting breast, gastric, ovarian, colorectal, prostate, glioblastoma, and endometrial cancers [89]. PI3K kinase activation constitutes a central mechanism between upstream growth signals and downstream signal transduction mechanisms involved in numerous cellular processes, such as protein synthesis, metabolism, inflammation, cell survival, motility, and tumor progression. PI3K is a large family of lipid enzymes capable of phosphorylating the 3′-OH group of phosphatidylinositol present on the plasma membrane. It was discovered more than 25 years ago and initially associated with the transforming ability of viral oncoproteins. Three classes of PI3Ks (class I, class II, and class III) have been identified in mammals. Kinases belonging to class IA consist of a catalytic subunit and a regulatory subunit. The catalytic subunits p110α, p110β, or p110δ are encoded by the PIK3CA, PIK3CB, and PIK3CD genes, respectively. In contrast, the regulatory subunits consist of p85α (in the isoforms p85α, p55α, p50α), p85β, and p55γ, which are encoded by the PIK3R1, PIK3R2, and PIK3R3 genes. Class IB consists of only one catalytic subunit, P110γ, and two regulatory subunits, p84 and p101 [90]. Class II includes three different monomeric isoforms and remains the most enigmatic of all PI3Ks, although recent studies have provided new clues about its role in signal transduction [91]. Finally, the only member of class III is known as Vps34 (Vacuolar protein signal 34), expressed in all eukaryotic organisms. Vps34 was first discovered in yeast and is implicated in the integration of cellular responses and changes in nutritional status [92]. A variety of signals stimulate PI3K activity, mainly through receptor tyrosine kinases (RTKs), but also through GPCRs and oncogenes, such as Ras, that directly bind p110. After stimulation, the catalytic subunit of PI3K phosphorylates phosphatidylinositol-4,5-bisphosphate (PIP2) to phosphatidylinositol-3,4,5-triphosphate (PIP3), which acts as a second messenger to recruit a series of proteins containing homology domain with the pleckstrin (PH) of the cell membrane. Uncontrolled signaling of PI3K is very common in cancer, also due to the different roles played by its catalytic subunits p110α and p110β. Mutations in the PI3KA gene encoding p110α have been shown in cancer cells from the colon, lung, prostate, liver, and brain [93]. This gene, in addition to being involved in the processes of cell cycle regulation and growth, acquires a very important role in endothelial cells, promoting angiogenesis and, thus, the formation of a vascular network essential for the delivery of nutrients and oxygen, which can ultimately ensure a pathway of metastasis from the primary lesion. In oncogenesis, the p110α isoform is required for tumors driven by activated receptor tyrosine kinases and oncogenes. The p110β is mainly required for GPCR downstream signaling but it was also found to be essential for the development of high-grade prostatic intraepithelial neoplasia (HG-PIN) [94]. In an animal model of Phosphatase and Tensin homolog (PTEN)-deficient PCa, ablation of p110β, but not that of p110α, impeded tumorigenesis, with a concomitant diminution in AKT phosphorylation [95]. Consistently, data from the latest studies suggested that while blockade of p110α had negligible effects in the development of PTEN-null CRPC, genetic or pharmacological disruption of p110β dramatically slowed the initiation and progression of CRPC [94,95,96].
A key molecule in the regulation of PI3K/AKT is PTEN. The function of PTEN as an oncosuppressor is carried out through its phosphatase activity; it dephosphorylates PIP3 to PIP2, negatively regulating the activation of the PI3K/AKT pathway. This phosphatase can act on both lipids and proteins, and acts by inhibiting cell proliferation and inducing apoptosis. Mutations to PTEN inhibit its oncosuppressive activity. Two major mutations affect the phosphatase domain: one results in the loss of phosphatase activity on both lipids and proteins while the other impairs phosphatase activity on protein substrates. In addition to regulating the PI3K/AKT signaling pathway, PTEN has many other critical roles in tumors, including genomic instability, tumor cell renewal, cell senescence, cell migration, and metastasis. Finally, PTEN plays a significant role in regulating the tumor microenvironment [97]. Mutations in the PTEN gene have been observed in breast, prostate, endometrial, ovarian, colon, melanoma, glioblastoma, and lymphoma cancer [98]. Studies in animal models have also shown that the loss of a single copy of the PTEN gene is sufficient to disrupt cell signaling and initiate uncontrolled cell growth [99]. PI3K activation leads to phosphorylation and then activation of AKT, or protein kinase B (PKB), a serine/threonine kinase of the AGC family of kinases. It exists in three structurally similar isoforms: AKT1, AKT2, and AKT. The three isoforms are composed of characteristic domains. The Pleckstrin Homology (PH) domain has a remarkably conserved tertiary structure, although the amino acid sequence may differ; this domain is responsible for binding to PIP3. The LIN domain, of 39 amino acids is the hinge region connecting the PH domain with the catalytic domain, which is poorly conserved among AKT isoforms (17–46% identical) and has no significant homology with any other human protein. The kinase domain extends from amino acids 148–411 and terminates in a hydrophobic regulatory motif (CTD), ATP-binding portion of the enzyme; the ATP-binding site of 25 residues has 96–100% homology across the three isoforms. The C-terminal hydrophobic domain appears to be conserved in the AGC family of kinases. These hydrophobic residues play a critical role in the complete activation of AKT for substrate phosphorylation. Within it is another key residue for enzyme activation, Ser473. While AKT1 is ubiquitously expressed at high levels, except for the kidney, liver, and spleen, AKT2 expression is high in insulin-sensitive tissues, such as brown fat, skeletal muscle, and liver. AKT3 expression is ubiquitous, although low levels of expression have been found in liver and skeletal muscle. These different isoforms appear to be implicated in specific functions. For example, amplification and overexpression of AKT2 correlate with increased cell motility and invasion, whereas increased AKT3 activity appears to contribute to the aggressiveness of steroid-hormone-insensitive tumors [100]. All three isoforms are activated through phosphorylation: the first occurs on a threonine residue while the second occurs on a serine residue in the hydrophobic motif. Once activated, AKT recognizes and phosphorylates serine or threonine residues of numerous substrates, such as tuberosis sclerosis complex 2 (TSC 2), glycogen synthase kinase 3 (GSK 3), forkhead box transcription factors (FOXO), p21WAF1/CIP1, p27KIP1, caspase-9, Bcl-2 associated death promoter (BAD), and inducible Nitric Oxide Synthase (iNOS), which regulate numerous processes that coordinate cell life and death, metabolism, and angiogenesis. Hyperactivation of AKT has been shown in numerous cancers, such as multiple myeloma, lung cancer, glioblastoma, breast cancer, prostate cancer, etc. [101]. The best-studied downstream substrate of AKT is mTOR kinase. AKT can directly phosphorylate and activate mTOR and can cause indirect activation of mTOR by phosphorylating and inactivating tuberous sclerosis 2, also called tuberin (TSC2), which normally inhibits mTOR. The consequence of mTOR activation is increased protein translation [102]. Finally, it has recently been shown that AKT activity can be negatively regulated by the PH domain of leucine repeat sequence-rich phosphatase (PHLPP), which specifically dephosphorylates the hydrophobic motif of AKT (Ser473 in Akt1) [103]. mTOR (mammalian target of rapamycin) is a serine/threonine kinase that regulates cell growth, proliferation, motility and survival, transcription, and protein synthesis. mTOR plays an important role in regulating the body’s energy balance and weight; it is activated by amino acids, glucose, insulin, and other hormones involved in regulating metabolism. Recent studies have shown that mTOR is not only a substrate of AKT but also a crucial activator of AKT. In fact, mTOR forms a complex (TORC2) with the protein Rapamycin-insensitive companion of mTOR (RICTOR) and then directly phosphorylates the Ser473 of AKT [104]. Activation of TORC2 could then explain the sequestration of newly formed mTOR molecules within cells during long-term rapamycin treatments. In fact, this drug is particularly effective in inducing apoptosis and suppressing the proliferation of AKT-overexpressing cells because, over time, it interferes with the reassembly of the complex by joining it [105].
4. Crosstalk between AR and PI3K Signaling
The PI3K/AKT/mTOR signaling pathway has been shown to be deregulated in a wide range of cancers. Genetic alterations have been identified in all components of this signaling pathway. In PCa, the PI3K/AKT/mTOR pathway is deregulated in 42% of localized and 100% of advanced disease cases, indicating that alterations in these signals might be an essential prerequisite for the development of CRPC [106]. The existence of negative feedback regulation within AR and PI3K/AKT signaling networks has been demonstrated [107] (Figure 1). Thus, gene mutations and amplifications, and changes in mRNA expression in components of the PI3K pathway, are strictly correlated with the prognosis of PCa patients. For example, reduced expression of PTEN is associated with higher Gleason, biochemical recurrence after prostatectomy, and shorter time to metastatic progression [108]. In addition, high levels of phospho-4EBP1 and eI4E are associated with increased mortality in patients with PCa, indicating that effectors further down the pathway are also predictive of disease progression [109].
Results from studies in knockout (KO) and transgenic mouse models have also clearly shown the role of PI3K/AKT/mTOR in the development of PCa. Specifically, overexpression of AKT or biallelic loss of the oncosuppressor PTEN in prostate epithelial cells leads to hyperactivation of the pathway and is sufficient for PCa development in vivo [110]. PTEN deletion has also been shown to inhibit the progression of PCa in mouse models with conditional KO of mTOR [111]. Others have also shown in vivo that the progression of PCa is reduced when PTEN and RICTOR, a subunit of mTORC2 complex, are lost [112]. This demonstrates that the progression of PCa can be sufficiently induced by the hyperactivation of PI3K/AKT/mTOR. Loss of the oncosuppressor PTEN and subsequent uncontrolled activation of the PI3K signaling pathway has been found in 40% of primary tumors and 70% of metastatic forms [113]. Moreover, AKT was shown to phosphorylate AR at Ser-210 and Ser-791, but the effect on AR activity and protein stability is debated, as studies have demonstrated either repression or activation of AR function [114]. From a functional point of view, AKT repressed AR transactivation in a reporter assay in AR-insensitive DU145 mPCa cells when the AR was exogenously expressed [115]. However, in the androgen-sensitive PCa LNCaP cell line, AKT phosphorylation activated the PSA reporter and promoted cell survival [115]. These differential effects suggest that AKT-mediated AR phosphorylation on AR function could be attributed to cell context. The AKT pathway is considerably sensitive to feedback regulation. Furthermore, inhibition of the PI3K pathway also stimulates the upstream of HER2/3, thereby activating the androgen receptor axis in murine and human tumors with PTEN deletion [116].
On the other hand, inactivation of AR would lead to over-regulation of the PI3K/AKT pathway, which has been correlated with altered control of cell growth and survival, increased metastatic competence, angiogenesis, and resistance to chemotherapy.
Indeed, it has been demonstrated that AR inhibition activates AKT signaling by reducing the expression of the AKT phosphatase PHLPP [117]. AR blockade reduces FKBP5 levels, then impairing PHLPP function and leading to upregulation of pAKT [118].
Thus, these two pathways are regulated by a reciprocal feedback mechanism in that the inhibition of one inactivates the other, allowing for cancer cell survival and progression.
Combination Therapy
Because inhibiting either AR or AKT often activates the other, a combination therapy might be advantageous. Over 40 compounds targeting key components in the PI3K-induced signaling pathway have been investigated to date. AZD5363, an inhibitor of all isoforms of Akt, has been reported to inhibit proliferation and induce apoptosis in prostate cancer cell lines expressing AR and has antitumor activity in vivo in androgen-sensitive LNCaP xenograft models resistant to castration [119]. However, resistance occurs already after about 30 days of treatment. This is proposed to be since AZD5363 induces an increase in the binding affinity of AR to AREs and an increase in the transcriptional activity of AR and the expression of AR-dependent genes, such as PSA and NKX3.1. These effects were overcome by the combination of AZD5363 and the earlier antiandrogen Bicalutamide, resulting not only in a synergistic inhibition of cell proliferation and induction of apoptosis in vitro, but also in a prolongation of tumor growth inhibition and PSA stabilization [120]. Moreover, clinical data from the latest ongoing clinical trials support the hypothesis that combinatorial therapies may have a good response in treating advanced PCa (Table 1). The phase II ProCAID clinical trial suggested that addition of capivasertib (pan-AKT inhibitor) to docetaxel improved OS benefit in mCRPC patients. Median OS was 25.3 months for capivasertib plus docetaxel versus 20.3 months for placebo plus docetaxel (hazard ratio (HR) 0.70, 95% confidence interval (CI) 0.47–1.05; nominal p = 0.09) [121]. Another pan-AKT inhibitor, Ipatasertib, has been used in the recent randomized, double-blind, phase III trial combined with abiraterone (IPATential150). This combination led to prolonged radiographic progression-free survival and antitumor activity over a placebo with abiraterone among patients with mCRPC with PTEN loss (median 18.5 vs. 16.5 months, HR = 0.77; p = 0.0335) [122]. The phase I/II study investigating AZD8186, a potent and selective inhibitor of PI3K, supported combination treatment with abiraterone acetate [123]. Moreover, a phase II trial of everolimus (mTOR inhibitor) plus bicalutamide showed encouraging efficacy in men with bicalutamide-naïve CRPC [124]. It must be mentioned, however, that one of the limitations of the use of PI3K/AKT inhibitors is undoubtedly the occurrence of AEs, usually hyperglycemia, rash, and diarrhea. For this reason, numerous studies are focusing on understanding the mechanisms and management of toxicity. In addition, new phase I studies are aimed at optimizing the dosing schedule to improve drug-related toxicity. Noteworthily, most of the clinical trials to date are directed towards patients with advanced or mCRPC, which are very different from the earlier, localized, high-risk disease. Hence, the effects of the combined targeting of AR and PTEN/AKT pathways in the setting of localized prostate cancer need to be investigated.
5. Conclusions
The high incidence of prostate cancer in the global male population has resulted in many efforts being channeled into finding the best therapeutic strategies. In particular, the prognosis of mCRPC remains very poor to date since patients develop resistance to treatments. Indeed, there is currently no single therapeutic choice for these patients, but recommended therapies involve the use of agents targeting different signaling pathways, often in combination or as neoadjuvant therapies. Many studies have confirmed the role of the PI3K/AKT/mTOR signaling pathway in the development of treatment resistance and tumor progression. Indeed, activation of PI3K increases proliferation and prevents apoptosis of prostate cancer cells, while inactivation promotes cell cycle arrest in the G phase. On the other hand, there is evidence that AR inhibition promotes upregulation of the PI3K pathway, and vice versa, via a negative feedback mechanism.
Therefore, studies aiming to understand the key mechanisms that induce resistance within this continuous crosstalk between the two signaling pathways are needed to improve the outcome of patients with prostate cancer.
Author Contributions
Conceptualization, E.T. and I.S.; writing—original draft preparation, I.S. and E.T.; writing—review and editing, E.T., I.S., S.G. and A.S.; supervision, I.S. and E.T.; funding acquisition, I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
BioRender software (Biorender.com) was used to create Figure 1.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ADT | androgen deprivation therapy |
| AGC | protein kinase A, G and C families |
| AKT | protein kinase B |
| AR-FL | androgen receptor full length |
| AR-V | androgen receptor splice variants |
| AR | androgen receptor |
| ARA | androgen receptor antagonist |
| ARE | androgen response element |
| ARSI | androgen receptor signaling inhibitor |
| ATP | adenosine triphosphate |
| BAD | Bcl-2-associated death promoter |
| BAT | bipolar androgen therapy |
| CRPC | castration-resistant prostate cancer |
| CSPC | castration-sensitive prostate cancer |
| CTC | circulating tumor cells |
| CTD | hydrophobic regulatory motif domain |
| CTLA4 | cytotoxic T-lymphocyte-associated protein 4 |
| CYP17A1 | cytochrome P450 17α-hydroxylase/17,20-lyase |
| DBD | DNA-binding domain |
| DHEA | dehydroepiandrosterone |
| DHT | 5α-dihydrotestosterone |
| DNA | deoxyribonucleic acid |
| FDA | Food and Drug Administration |
| FISH | fluorescence in situ hybridization |
| FKBP5 | FK506 binding protein 5 |
| FOXO | forkhead box transcription factors |
| FSH | follicle-stimulating hormone |
| GnRH | gonadotropin-releasing hormone |
| GPCR | G-protein-coupled receptor |
| GSK 3 | glycogen synthase kinase 3 |
| HER | human epidermal growth factor receptor |
| HPG | hypothalamus–pituitary–gonadal axis |
| HSP | heat-shock protein |
| iNOS | inducible nitric oxide synthase |
| KO | knockout |
| LBD | ligand-binding domain |
| LH | luteinizing hormone |
| LHRH | luteinizing hormone-releasing hormone |
| LNCaP | androgen-sensitive human prostate adenocarcinoma cells |
| mCRPC | metastatic castration-resistant prostate cancer |
| mCSPC | metastatic castration-sensitive prostate cancer |
| mRNA | messenger ribonucleic acid |
| mTOR | mammalian target of rapamycin |
| mTORC1/2 | mTOR complex ½ |
| nmCRPC | non metastatic castration-resistant prostate cancer |
| NOS | nitric oxide synthase |
| NTD | amino-terminal transcriptional domain |
| OS | overall survival |
| p70S6K | p70S6 kinase |
| PARP | poly ADP-ribose polymerase |
| PCa | prostate cancer |
| PCR | polymerase chain reaction |
| PD-L1 | programmed death-ligand 1 |
| PD1 | programmed cell death protein 1 |
| PDK1 | phosphoinositide-dependent kinase 1 |
| PFS | progression-free survival |
| PH | pleckstrin homology domain |
| PHLPP | PH domain of leucine repeat sequence-rich phosphatase |
| PI(3,4)P2 | phosphatidylinositol-3,4-biphosphate |
| PI(4,5)P2 | phosphatidylinositol-4,5-biphosphate |
| PI3K | phosphatidylinositol-3-kinase |
| PIP3 | phosphatidylinositol-3,4,5-triphosphate |
| PKA | protein kinase A |
| PSA | prostate-specific antigen |
| PTEN | phosphatase and tensin homolog |
| RANKL | receptor activator of nuclear factor kappa-B ligand |
| RICTOR | rapamycin-insensitive companion of mTOR |
| RNA | ribonucleic acid |
| RP | radical prostatectomy |
| rPFS | radiographic progression-free survival |
| RT-PCR | real time polymerase chain reaction |
| RT | radiotherapy |
| RTK | receptor tyrosine kinase |
| SHBG | sex hormone-binding globulin |
| TSC 2 | tuberosis sclerosis complex 2 |
| Vps34 | vacuolar protein signal 34 |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.M.; Cacciamani, G.E.; Gill, K.; Zhang, J.; Liu, L.; Abreu, A.; Gill, I.S. Trends in Incidence of Metastatic Prostate Cancer in the US. JAMA Netw. Open 2022, 5, e222246. [Google Scholar] [CrossRef] [PubMed]
- Rebello, R.J.; Oing, C.; Knudsen, K.E.; Loeb, S.; Johnson, D.C.; Reiter, R.E.; Gillessen, S.; Van der Kwast, T.; Bristow, R.G. Prostate cancer. Nat. Rev. Dis. Prim. 2021, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Fedewa, S.A.; Ma, J.; Siegel, R.; Lin, C.C.; Brawley, O.W.; Ward, E.M. Prostate Cancer Incidence and PSA Testing Patterns in Relation to USPSTF Screening Recommendations. JAMA J. Am. Med. Assoc. 2015, 314, 2054–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campá, J.; Mar-Barrutia, G.; Extramiana, J.; Arróspide, A.; Mar, J. Advanced prostate cancer survival in Spain according to the Gleason score, age and stage. Actas Urológicas Españolas 2016, 40, 499–506. [Google Scholar] [CrossRef]
- Steele, C.B.; Li, J.; Huang, B.; Weir, H.K. Prostate cancer survival in the United States by race and stage (2001–2009): Findings from the CONCORD-2 study. Cancer 2017, 123, 5160–5177. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [Green Version]
- Cooperberg, M.R.; Broering, J.M.; Carroll, P.R. Time Trends and Local Variation in Primary Treatment of Localized Prostate Cancer. J. Clin. Oncol. 2010, 28, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Heidenreich, A.; Bastian, P.J.; Bellmunt, J.; Bolla, M.; Joniau, S.; van der Kwast, T.; Mason, M.; Matveev, V.; Wiegel, T.; Zattoni, F.; et al. EAU guidelines on prostate cancer. Part II: Treatment of advanced, relapsing, and castration-resistant prostate cancer. Eur. Urol. 2014, 65, 467–479. [Google Scholar] [CrossRef]
- Nayak, A.L.; Flaman, A.S.; Mallick, R.; Lavallée, L.T.; Fergusson, D.A.; Cagiannos, I.; Morash, C.; Breau, R.H. Do androgen-directed therapies improve outcomes in prostate cancer patients undergoing radical prostatectomy? A systematic review and meta-analysis. Can. Urol. Assoc. J. 2021, 15, 269. [Google Scholar] [CrossRef] [PubMed]
- Crona, D.J.; Whang, Y.E. Androgen Receptor-Dependent and -Independent Mechanisms Involved in Prostate Cancer Therapy Resistance. Cancers 2017, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Sanda, M.G.; Cadeddu, J.A.; Kirkby, E.; Chen, R.C.; Crispino, T.; Fontanarosa, J.; Freedland, S.; Greene, K.; Klotz, L.H.; Makarov, D.V.; et al. Clinically Localized Prostate Cancer: AUA/ASTRO/SUO Guideline. Part I: Risk Stratification, Shared Decision Making, and Care Options. J. Urol. 2018, 199, 683–690. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Boorjian, S.A.; Inman, B.; Bagniewski, S.; Bergstralh, E.J.; Blute, M.L. Timing of Androgen Deprivation Therapy and its Impact on Survival After Radical Prostatectomy: A Matched Cohort Study. J. Urol. 2008, 179, 1830–1837. [Google Scholar] [CrossRef]
- Ferraldeschi, R.; Welti, J.; Luo, J.; Attard, G.; de Bono, J.S. Targeting the androgen receptor pathway in castration-resistant prostate cancer: Progresses and prospects. Oncogene 2014, 34, 1745–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, L.; Schweizer, M.T. Targeting persistent androgen receptor signaling in castration-resistant prostate cancer. Med. Oncol. 2016, 33, 44. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.H.; Muñoz, D.M.; Rodríguez, A.R.; Ruiz, J.S.; Rodríguez, D.R.; Couñago, F. Changing the History of Prostate Cancer with New Targeted Therapies. Biomedicines 2021, 9, 392. [Google Scholar] [CrossRef]
- Attard, G.; Reid, A.H.; A’Hern, R.; Parker, C.; Oommen, N.B.; Folkerd, E.; Messiou, C.; Molife, L.R.; Maier, G.; Thompson, E.; et al. Selective Inhibition of CYP17 With Abiraterone Acetate Is Highly Active in the Treatment of Castration-Resistant Prostate Cancer. J. Clin. Oncol. 2009, 27, 3742–3748. [Google Scholar] [CrossRef] [Green Version]
- Crawford, E.D.; Heidenreich, A.; Lawrentschuk, N.; Tombal, B.; Pompeo, A.C.L.; Mendoza-Valdes, A.; Miller, K.; Debruyne, F.M.J.; Klotz, L. Androgen-targeted therapy in men with prostate cancer: Evolving practice and future considerations. Prostate Cancer Prostatic Dis. 2019, 22, 24–38. [Google Scholar] [CrossRef] [Green Version]
- de Bono, J.S.; Logothetis, C.J.; Molina, A.; Fizazi, K.; North, S.; Chu, L.; Chi, K.N.; Jones, R.J.; Goodman, O.B., Jr.; Saad, F.; et al. Abiraterone and Increased Survival in Metastatic Prostate Cancer. N. Engl. J. Med. 2011, 364, 1995–2005. [Google Scholar] [CrossRef]
- Beer, T.M.; Armstrong, A.J.; Rathkopf, D.E.; Loriot, Y.; Sternberg, C.N.; Higano, C.S.; Iversen, P.; Bhattacharya, S.; Carles, J.; Chowdhury, S.; et al. Enzalutamide in Metastatic Prostate Cancer before Chemotherapy. N. Engl. J. Med. 2014, 371, 424–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, C.N.; Fizazi, K.; Saad, F.; Shore, N.D.; De Giorgi, U.; Penson, D.F.; Ferreira, U.; Efstathiou, E.; Madziarska, K.; Kolinsky, M.P.; et al. Enzalutamide and Survival in Nonmetastatic, Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 382, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, W.; Xiao, Y.-T.; Huang, H.; Di Gu, D.; Ren, S. Targeting signaling pathways in prostate cancer: Mechanisms and clinical trials. Signal Transduct. Target. Ther. 2022, 7, 198. [Google Scholar] [CrossRef]
- Silvestri, I.; Tortorella, E.; Giantulli, S.; Scarpa, S.; Sciarra, A. Immunotherapy in Prostate Cancer: Recent Advances and Future Directions. EMJ Urol. 2019, 7, 51–61. [Google Scholar]
- Caram, M.E.V.; Ross, R.; Lin, P.; Mukherjee, B. Factors Associated with Use of Sipuleucel-T to Treat Patients with Advanced Prostate Cancer. JAMA Netw. Open 2019, 2, e192589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, V.; Paunu, K.; Ahlskog, J.K.; Varnai, R.; Sipeky, C.; Sundvall, M. PARP Inhibitors in Prostate Cancer—The Preclinical Rationale and Current Clinical Development. Genes 2019, 10, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messner, E.A.; Steele, T.M.; Tsamouri, M.M.; Hejazi, N.; Gao, A.C.; Mudryj, M.; Ghosh, P.M. The Androgen Receptor in Prostate Cancer: Effect of Structure, Ligands and Spliced Variants on Therapy. Biomedicines 2020, 8, 422. [Google Scholar] [CrossRef]
- Jacob, A.; Raj, R.; Allison, D.B.; Myint, Z.W. Androgen Receptor Signaling in Prostate Cancer and Therapeutic Strategies. Cancers 2021, 13, 5417. [Google Scholar] [CrossRef]
- Sharifi, N.; Auchus, R.J. Steroid biosynthesis and prostate cancer. Steroids 2012, 77, 719–726. [Google Scholar] [CrossRef]
- van Laar, J.; Vries, J.-D.; Voorhorst-Ogink, M.; Brinkmann, A. The human androgen receptor is a 110 kDa protein. Mol. Cell. Endocrinol. 1989, 63, 39–44. [Google Scholar] [CrossRef]
- Shaffer, P.L.; Jivan, A.; Dollins, D.E.; Claessens, F.; Gewirth, D.T. Structural basis of androgen receptor binding to selective androgen response elements. Proc. Natl. Acad. Sci. USA 2004, 101, 4758–4763. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.E.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, R.A.; Grossmann, M. Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Androg. Recept. Biol. Clin. Biochem. Rev. 2016, 37, 3–15. [Google Scholar]
- Dai, C.; Heemers, H.; Sharifi, N. Androgen signaling in prostate cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a030452. [Google Scholar] [CrossRef] [Green Version]
- Heemers, H.V.; Tindall, D.J. Androgen Receptor (AR) Coregulators: A Diversity of Functions Converging on and Regulating the AR Transcriptional Complex. Endocr. Rev. 2007, 28, 778–808. [Google Scholar] [CrossRef] [Green Version]
- Dehm, S.M.; Tindall, D.J. Androgen Receptor Structural and Functional Elements: Role and Regulation in Prostate Cancer. Mol. Endocrinol. 2007, 21, 2855–2863. [Google Scholar] [CrossRef] [Green Version]
- DePriest, A.D.; Fiandalo, M.; Schlanger, S.; Heemers, F.; Mohler, J.L.; Liu, S.; Heemers, H.V. Regulators of Androgen Action Resource: A one-stop shop for the comprehensive study of androgen receptor action. Database 2016, 2016, bav125. [Google Scholar] [CrossRef] [Green Version]
- Pang, T.P.; Clarke, M.V.; Ghasem-Zadeh, A.; Lee, N.K.; Davey, R.A.; MacLean, H.E. A physiological role for androgen actions in the absence of androgen receptor DNA binding activity. Mol. Cell. Endocrinol. 2012, 348, 189–197. [Google Scholar] [CrossRef]
- Dahiya, V.; Bagchi, G. Non-canonical androgen signaling pathways and implications in prostate cancer. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2022, 1869, 119357. [Google Scholar] [CrossRef]
- Huggins, C.; Hodges, C.V. Milestone in Urology Studies on Prostatic Cancer I. The Effect of Castration, of Estrogen and of Androgen Injection on Serum Phosphatases in Metastatic Carcinoma of the Prostate. J. Urol. 2002, 168, 9–12. [Google Scholar] [CrossRef]
- Conn, P.M.; Crowley, W.F., Jr. Gonadotropin-releasing hormone and its analogues. N. Engl. J. Med. 1991, 324, 93–103. [Google Scholar] [PubMed]
- Limonta, P.; Marelli, M.M.; Moretti, R.M. LHRH analogues as anticancer agents: Pituitary and extrapituitary sites of action. Expert Opin. Investig. Drugs 2001, 10, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Choueiri, T.K.; Hamnvik, O.P.R.; Preston, M.A.; De Velasco, G.; Jiang, W.; Loeb, S.; Nguyen, P.L.; Trinh, Q.D. Comparison of gonadotropin-releasing hormone agonists and orchiectomy: Effects of androgen-deprivation therapy. JAMA Oncol. 2016, 2, 500–507. [Google Scholar] [CrossRef] [Green Version]
- Keating, N.L.; O’Malley, A.J.; Smith, M.R. Diabetes and Cardiovascular Disease During Androgen Deprivation Therapy for Prostate Cancer. J. Clin. Oncol. 2006, 24, 4448–4456. [Google Scholar] [CrossRef] [PubMed]
- Tosco, L.; Laenen, A.; Gevaert, T.; Salmon, I.; Decaestecker, C.; Davicioni, E.; Buerki, C.; Claessens, F.; Swinnen, J.; Goffin, K.; et al. Neoadjuvant degarelix with or without apalutamide followed by radical prostatectomy for intermediate and high-risk prostate cancer: ARNEO, a randomized, double blind, placebo-controlled trial. BMC Cancer 2018, 18, 354. [Google Scholar] [CrossRef]
- Shore, N.D.; Saad, F.; Cookson, M.S.; George, D.J.; Saltzstein, D.R.; Tutrone, R.; Akaza, H.; Bossi, A.; van Veenhuyzen, D.F.; Selby, B.; et al. Oral Relugolix for Androgen-Deprivation Therapy in Advanced Prostate Cancer. N. Engl. J. Med. 2020, 382, 2187–2196. [Google Scholar] [CrossRef]
- Albertsen, P.C.; Klotz, L.; Tombal, B.; Grady, J.; Olesen, T.K.; Nilsson, J. Cardiovascular Morbidity Associated with Gonadotropin Releasing Hormone Agonists and an Antagonist. Eur. Urol. 2014, 65, 565–573. [Google Scholar] [CrossRef]
- Odonnell, A.G.; Judson, I.; Dowsett, M.; Raynaud, F.; Dearnaley, D.P.; Mason, M.G.; Harland, S.J.; Robbins, A.S.; Halbert, G.; Nutley, B.; et al. Hormonal impact of the 17α-hydroxylase/C17,20-lyase inhibitor abiraterone acetate (CB7630) in patients with prostate cancer. Br. J. Cancer 2004, 90, 2317–2325. [Google Scholar] [CrossRef]
- Rehman, Y.; Rosenberg, J.E. Abiraterone acetate: Oral androgen biosynthesis inhibitor for treatment of castration-resistant prostate cancer. Drug Des. Dev. Ther. 2012, 6, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Hoyle, A.P.; Ali, A.; James, N.D.; Cook, A.; Parker, C.C.; De Bono, J.S.; Attard, G.; Chowdhury, S.; Cross, W.R.; Dearnaley, D.P.; et al. Abiraterone in “High-” and “Low-risk” Metastatic Hormone-sensitive Prostate Cancer (Figure presented). Eur. Urol. 2019, 76, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Barata, P.C.; Sartor, A.O. Metastatic castration-sensitive prostate cancer: Abiraterone, docetaxel, or…. Cancer 2019, 125, 1777–1788. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Liaw, B.; Oh, W. The role of ketoconazole in current prostate cancer care. Nat. Rev. Urol. 2018, 15, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Kolvenbag, G.; Furr, B.; Blackledge, G. Receptor affinity and potency of non-steroidal antiandrogens: Translation of preclinical findings into clinical activity. Prostate Cancer Prostatic Dis. 1998, 1, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scher, H.I.; Fizazi, K.; Saad, F.; Taplin, M.E.; Sternberg, C.N.; Miller, K.; De Wit, R.; Mulders, P.; Chi, K.N.; Shore, N.D.; et al. Increased Survival with Enzalutamide in Prostate Cancer after Chemotherapy. N. Engl. J. Med. 2012, 367, 1187–1197. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, A.J.; Szmulewitz, R.Z.; Petrylak, D.P.; Holzbeierlein, J.; Villers, A.; Azad, A.; Alcaraz, A.; Alekseev, B.; Iguchi, T.; Shore, N.D.; et al. ARCHES: A Randomized, Phase III Study of Androgen Deprivation Therapy with Enzalutamide or Placebo in Men with Metastatic Hormone-Sensitive Prostate Cancer. J. Clin. Oncol. 2019, 37, 2974–2986. [Google Scholar] [CrossRef]
- Clegg, N.J.; Wongvipat, J.; Joseph, J.D.; Tran, C.; Ouk, S.; Dilhas, A.; Chen, Y.; Grillot, K.; Bischoff, E.D.; Cai, L.; et al. ARN-509: A Novel Antiandrogen for Prostate Cancer Treatment. Cancer Res. 2012, 72, 1494–1503. [Google Scholar] [CrossRef] [Green Version]
- Chi, K.N.; Chowdhury, S.; Bjartell, A.; Chung, B.H.; de Santana Gomes, A.J.P.; Given, R.; Juárez, A.; Merseburger, A.S.; Özgüroğlu, M.; Uemura, H.; et al. Apalutamide in Patients with Metastatic Castration-Sensitive Prostate Cancer: Final Survival Analysis of the Randomized, Double-Blind, Phase III TITAN Study. J. Clin. Oncol. 2021, 39, 2294–2303. [Google Scholar] [CrossRef]
- Shore, N.D. Darolutamide (ODM-201) for the treatment of prostate cancer. Expert Opin. Pharmacother. 2017, 18, 945–952. [Google Scholar] [CrossRef]
- Fizazi, K.; Smith, M.R.; Tombal, B. Clinical Development of Darolutamide: A Novel Androgen Receptor Antagonist for the Treatment of Prostate Cancer. Clin. Genitourin. Cancer 2018, 16, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.; Nocera, L.; Ruvolo, C.C.; Würnschimmel, C.; Tian, Z.; Shariat, S.F.; Saad, F.; Tilki, D.; Graefen, M.; Kluth, L.A.; et al. Overall survival and adverse events after treatment with darolutamide vs. apalutamide vs. enzalutamide for high-risk non-metastatic castration-resistant prostate cancer: A systematic review and network meta-analysis. Prostate Cancer Prostatic Dis. 2022, 25, 139–148. [Google Scholar] [CrossRef]
- Zhou, Y.; Bolton, E.C.; O Jones, J. Androgens and androgen receptor signaling in prostate tumorigenesis. J. Mol. Endocrinol. 2015, 54, R15–R29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Jiang, X.; Liang, X.; Jiang, G. Molecular and cellular mechanisms of castration resistant prostate cancer (Review). Oncol. Lett. 2018, 15, 6063–6076. [Google Scholar] [CrossRef] [PubMed]
- Visakorpi, T.; Hyytinen, E.; Koivisto, P.; Tanner, M.; Keinänen, R.; Palmberg, C.; Palotie, A.; Tammela, T.; Isola, J.; Kallioniemi, O.P. In vivo amplification of the androgen receptor gene and progression of human prostate cancer. Nat. Genet. 1995, 9, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.D.; Welsbie, D.S.; Tran, C.; Baek, S.H.; Chen, R.; Vessella, R.; Rosenfeld, M.G.; Sawyers, C.L. Molecular determinants of resistance to antiandrogen therapy. Nat. Med. 2004, 10, 33–39. [Google Scholar] [CrossRef]
- Attard, G.; Swennenhuis, J.F.; Olmos, D.; Reid, A.H.; Vickers, E.; A’Hern, R.; Levink, R.; Coumans, F.; Moreira, J.; Riisnaes, R.; et al. Characterization of ERG, AR and PTEN gene status in circulating tumor cells from patients with castration-resistant prostate cancer. Cancer Res. 2009, 69, 2912–2918. [Google Scholar] [CrossRef] [Green Version]
- Azad, A.A.; Volik, S.V.; Wyatt, A.W.; Haegert, A.; Le Bihan, S.; Bell, R.H.; Anderson, S.A.; McConeghy, B.; Shukin, R.; Bazov, J.; et al. Androgen Receptor Gene Aberrations in Circulating Cell-Free DNA: Biomarkers of Therapeutic Resistance in Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2015, 21, 2315–2324. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; He, H.H.; Chen, S.; Coleman, I.; Wang, H.; Fang, Z.; Chen, S.; Nelson, P.S.; Liu, X.S.; Brown, M.; et al. Androgen Receptor Gene Expression in Prostate Cancer Is Directly Suppressed by the Androgen Receptor through Recruitment of Lysine-Specific Demethylase 1. Cancer Cell 2011, 20, 457–471. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Chen, S.; Sowalsky, A.G.; Voznesensky, O.S.; Mostaghel, E.A.; Nelson, P.S.; Cai, C.; Balk, S.P. Rapid Induction of Androgen Receptor Splice Variants by Androgen Deprivation in Prostate Cancer. Clin. Cancer Res. 2014, 20, 1590–1600. [Google Scholar] [CrossRef] [Green Version]
- Grasso, C.S.; Wu, Y.-M.; Robinson, D.R.; Cao, X.; Dhanasekaran, S.M.; Khan, A.P.; Quist, M.J.; Jing, X.; Lonigro, R.J.; Brenner, J.C.; et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature 2012, 487, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Jernberg, E.; Bergh, A.; Wikström, P. Clinical relevance of androgen receptor alterations in prostate cancer. Endocr. Connect. 2017, 6, R146–R161. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Kinoshita, H.; Segawa, T.; Nakamura, E.; Inoue, T.; Shimizu, Y.; Kamoto, T.; Ogawa, O. Antiandrogen Bicalutamide Promotes Tumor Growth in a Novel Androgen-Dependent Prostate Cancer Xenograft Model Derived from a Bicalutamide-Treated Patient. Cancer Res. 2005, 65, 9611–9616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledet, E.M.; Lilly, M.B.; Sonpavde, G.; Lin, E.; Nussenzveig, R.H.; Barata, P.C.; Yandell, M.; Nagy, R.J.; Kiedrowski, L.; Agarwal, N.; et al. Comprehensive Analysis of AR Alterations in Circulating Tumor DNA from Patients with Advanced Prostate Cancer. Oncologist 2020, 25, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Paschalis, A.; Sharp, A.; Welti, J.C.; Neeb, A.; Raj, G.V.; Luo, J.; Plymate, S.R.; de Bono, J.S. Alternative splicing in prostate cancer. Nat. Rev. Clin. Oncol. 2018, 15, 663–675. [Google Scholar] [CrossRef]
- Kiliccioglu, I.; Bilen, C.Y.; Sozen, S.; Konac, E. Upregulation of potential regulatory signaling molecules correlate with androgen receptor splice variants AR-V7 and AR-V567es in prostate cancer metastasis. Gene 2021, 772, 145377. [Google Scholar] [CrossRef]
- Sciarra, A.; Maggi, M.; Salciccia, S.; Nicolai, A.; Tortorella, E.; Giantulli, S.; Magliocca, F.M.; Silvestri, I.; Taglieri, L.; Cattarino, S.; et al. Tissue Expression of Androgen Receptor Splice Variant 7 at Radical Prostatectomy Predicts Risk of Progression in Untreated Nonmetastatic Prostate Cancer. Oncology 2021, 99, 251–255. [Google Scholar] [CrossRef]
- Wadosky, K.M.; Koochekpour, S. Androgen receptor splice variants and prostate cancer: From bench to bedside. Oncotarget 2017, 8, 18550–18576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonarakis, E.S.; Lu, C.; Wang, H.; Luber, B.; Nakazawa, M.; Roeser, J.C.; Chen, Y.; Mohammad, T.A.; Chen, Y.; Fedor, H.L.; et al. AR-V7 and Resistance to Enzalutamide and Abiraterone in Prostate Cancer. N. Engl. J. Med. 2014, 371, 1028–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todenhöfer, T.; Azad, A.; Stewart, C.; Gao, J.; Eigl, B.; Gleave, M.; Joshua, A.; Black, P.C.; Chi, K.N. AR-V7 Transcripts in Whole Blood RNA of Patients with Metastatic Castration Resistant Prostate Cancer Correlate with Response to Abiraterone Acetate. J. Urol. 2017, 197, 135–142. [Google Scholar] [CrossRef]
- Li, Y.; Chan, S.C.; Brand, L.J.; Hwang, T.H.; Silverstein, K.A.; Dehm, S.M. Androgen Receptor Splice Variants Mediate Enzalutamide Resistance in Castration-Resistant Prostate Cancer Cell Lines. Cancer Res. 2013, 73, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Dalrymple, S.L.; Coleman, I.; Zheng, S.L.; Xu, J.; Hooper, J.E.; Antonarakis, E.S.; De Marzo, A.M.; Meeker, A.K.; Nelson, P.S.; et al. Role of androgen receptor splice variant-7 (AR-V7) in prostate cancer resistance to 2nd-generation androgen receptor signaling inhibitors. Oncogene 2020, 39, 6935–6949. [Google Scholar] [CrossRef]
- Liu, L.L.; Xie, N.; Sun, S.; Plymate, S.; Mostaghel, E.; Dong, X. Mechanisms of the androgen receptor splicing in prostate cancer cells. Oncogene 2014, 33, 3140–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.; Bai, S.; Qi, Y.; Zhan, Y.; Ungerleider, N.; Zhang, D.Y.; Neklesa, T.; Corey, E.; Dehm, S.M.; Zhang, K.; et al. Increased transcription and high translation efficiency lead to accumulation of androgen receptor splice variant after androgen deprivation therapy. Cancer Lett. 2021, 504, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhan, Y.; Qi, Y.; Cao, B.; Bai, S.; Xu, W.; Gambhir, S.S.; Lee, P.; Sartor, O.; Flemington, E.K.; et al. Androgen Receptor Splice Variants Dimerize to Transactivate Target Genes. Cancer Res. 2015, 75, 3663–3671. [Google Scholar] [CrossRef] [Green Version]
- Roggero, C.M.; Jin, L.; Cao, S.; Sonavane, R.; Kopplin, N.G.; Ta, H.Q.; Ekoue, D.N.; Witwer, M.; Ma, S.; Liu, H.; et al. A detailed characterization of stepwise activation of the androgen receptor variant 7 in prostate cancer cells. Oncogene 2021, 40, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Au, C.C.; Bin Jamalruddin, M.A.; Abou-Ghali, N.E.; Mukhtar, E.; Portella, L.; Berger, A.; Worroll, D.; Vatsa, P.; Rickman, D.S.; et al. AR-V7 exhibits non-canonical mechanisms of nuclear import and chromatin engagement in castrate-resistant prostate cancer. eLife 2022, 11, e73396. [Google Scholar] [CrossRef]
- Liu, G.; Sprenger, C.; Sun, S.; Epilepsia, K.S.; Haugk, K.; Zhang, X.; Coleman, I.; Nelson, P.S.; Plymate, S. AR Variant ARv567es Induces Carcinogenesis in a Novel Transgenic Mouse Model of Prostate Cancer. Neoplasia 2013, 15, 1009–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonarakis, E.S.; Lu, C.; Luber, B.; Wang, H.; Chen, Y.; Zhu, Y.; Silberstein, J.L.; Taylor, M.N.; Maughan, B.L.; Denmeade, S.R.; et al. Clinical Significance of Androgen Receptor Splice Variant-7 mRNA Detection in Circulating Tumor Cells of Men With Metastatic Castration-Resistant Prostate Cancer Treated With First- and Second-Line Abiraterone and Enzalutamide. J. Clin. Oncol. 2017, 35, 2149–2156. [Google Scholar] [CrossRef]
- Tagawa, S.T.; Antonarakis, E.S.; Gjyrezi, A.; Galletti, G.; Kim, S.; Worroll, D.; Stewart, J.; Zaher, A.; Szatrowski, T.P.; Ballman, K.V.; et al. Expression of AR-V7 and ARv567es in Circulating Tumor Cells Correlates with Outcomes to Taxane Therapy in Men with Metastatic Prostate Cancer Treated in TAXYNERGY. Clin. Cancer Res. 2019, 25, 1880–1888. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [Green Version]
- Geering, B.; Cutillas, P.R.; Nock, G.; Gharbi, S.I.; Vanhaesebroeck, B. Class IA phosphoinositide 3-kinases are obligate p85-p110 heterodimers. Proc. Natl. Acad. Sci. USA 2007, 104, 7809–7814. [Google Scholar] [CrossRef] [Green Version]
- Gewinner, C.; Wang, Z.C.; Richardson, A.; Teruya-Feldstein, J.; Etemadmoghadam, D.; Bowtell, D.; Barretina, J.; Lin, W.M.; Rameh, L.; Salmena, L.; et al. Evidence that Inositol Polyphosphate 4-Phosphatase Type II Is a Tumor Suppressor that Inhibits PI3K Signaling. Cancer Cell 2009, 16, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backer, J.M. The regulation and function of Class III PI3Ks: Novel roles for Vps34. Biochem. J. 2008, 410, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.A.; Bogomolniy, F.; Yee, C.J.; Lash, A.; Barakat, R.R.; Borgen, P.I.; Boyd, J. Frequent Mutation of the PIK3CA Gene in Ovarian and Breast Cancers. Clin. Cancer Res. 2005, 11, 2875–2878. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wang, Y.; Ribeiro, C.F.; Manokaran, C.; Chang, H.; Von, T.; Rodrigues, S.; Cizmecioglu, O.; Jia, S.; Korpal, M.; et al. Blocking PI3K p110β Attenuates Development of PTEN-Deficient Castration-Resistant Prostate Cancer. Mol. Cancer Res. 2022, 20, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Liu, Z.; Zhang, S.; Liu, P.; Zhang, L.; Lee, S.H.; Zhang, J.; Signoretti, S.; Loda, M.; Roberts, T.M.; et al. Essential roles of PI(3)K-p110β in cell growth, metabolism and tumorigenesis. Nature 2008, 454, 776–779. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.J.; Liu, Z.; Wang, L.; Shin, E.; Loda, M.F.; Roberts, T.M. The oncogenic properties of mutant p110α and p110β phosphatidylinositol 3-kinases in human mammary epithelial cells. Proc. Natl. Acad. Sci. USA 2005, 102, 18443–18448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conciatori, F.; Bazzichetto, C.; Falcone, I.; Ciuffreda, L.; Ferretti, G.; Vari, S.; Ferraresi, V.; Cognetti, F.; Milella, M. PTEN Function at the Interface between Cancer and Tumor Microenvironment: Implications for Response to Immunotherapy. Int. J. Mol. Sci. 2020, 21, 5337. [Google Scholar] [CrossRef]
- Bunney, T.D.; Katan, M. Phosphoinositide signalling in cancer: Beyond PI3K and PTEN. Nat. Rev. Cancer 2010, 10, 342–352. [Google Scholar] [CrossRef]
- Knowles, M.A.; Platt, F.M.; Ross, R.L.; Hurst, C.D. Phosphatidylinositol 3-kinase (PI3K) pathway activation in bladder cancer. Cancer Metastasis Rev. 2009, 28, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Hinz, N.; Jücker, M. Distinct functions of AKT isoforms in breast cancer: A comprehensive review. Cell Commun. Signal. 2019, 17, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.M.; Sharma, A.; Cheung, M.; Zimmerman, M.; Cheng, J.Q.; Bosenberg, M.W.; Kester, M.; Sandirasegarane, L.; Robertson, G.P. Deregulated Akt3 Activity Promotes Development of Malignant Melanoma. Cancer Res. 2004, 64, 7002–7010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Guan, K.-L. mTOR as a central hub of nutrient signalling and cell growth. Nat. Cell Biol. 2019, 21, 63–71. [Google Scholar] [CrossRef]
- Yan, X.; Li, W.; Yang, L.; Dong, W.; Chen, W.; Mao, Y.; Xu, P.; Li, D.-J.; Yuan, H.; Li, Y.-H. MiR-135a Protects Vascular Endothelial Cells Against Ventilator-Induced Lung Injury by Inhibiting PHLPP2 to Activate PI3K/Akt Pathway. Cell. Physiol. Biochem. 2018, 48, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.-H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. Rictor, a Novel Binding Partner of mTOR, Defines a Rapamycin-Insensitive and Raptor-Independent Pathway that Regulates the Cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellen, P.; Saqcena, M.; Salloum, D.; Feng, J.; Preda, A.; Xu, L.; Rodrik-Outmezguine, V.; Foster, D.A. High-dose rapamycin induces apoptosis in human cancer cells by dissociating mTOR complex 1 and suppressing phosphorylation of 4E-BP1. Cell Cycle 2011, 10, 3948–3956. [Google Scholar] [CrossRef] [Green Version]
- Pungsrinont, T.; Kallenbach, J.; Baniahmad, A. Role of pi3k-akt-mtor pathway as a pro-survival signaling and resistance-mediating mechanism to therapy of prostate cancer. Int. J. Mol. Sci. 2021, 22, 11088. [Google Scholar] [CrossRef]
- Chandarlapaty, S. Negative Feedback and Adaptive Resistance to the Targeted Therapy of Cancer. Cancer Discov. 2012, 2, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotan, T.L.; Carvalho, F.L.; Peskoe, S.B.; Hicks, J.L.; Good, J.; Fedor, H.L.; Humphreys, E.; Han, M.; Platz, E.A.; Squire, J.A.; et al. PTEN loss is associated with upgrading of prostate cancer from biopsy to radical prostatectomy. Mod. Pathol. 2015, 28, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Edlind, M.P.; Hsieh, A.C. PI3K-AKT-mTOR signaling in prostate cancer progression and androgen deprivation therapy resistance. Asian J. Androl. 2014, 16, 378–386. [Google Scholar]
- Wang, S.; Gao, J.; Lei, Q.-Y.; Rozengurt, N.; Pritchard, C.; Jiao, J.; Thomas, G.V.; Li, G.; Roy-Burman, P.; Nelson, P.S.; et al. Prostate-specific deletion of the murine Pten tumor suppressor gene leads to metastatic prostate cancer. Cancer Cell 2003, 4, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Nardella, C.; Carracedo, A.; Alimonti, A.; Hobbs, R.M.; Clohessy, J.G.; Chen, Z.; Egia, A.; Fornari, A.; Fiorentino, M.; Loda, M.; et al. Differential Requirement of mTOR in Postmitotic Tissues and Tumorigenesis. Sci. Signal. 2009, 2, ra2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guertin, D.A.; Stevens, D.M.; Saitoh, M.; Kinkel, S.; Crosby, K.; Sheen, J.-H.; Mullholland, D.J.; Magnuson, M.A.; Wu, H.; Sabatini, D.M. mTOR Complex 2 Is Required for the Development of Prostate Cancer Induced by Pten Loss in Mice. Cancer Cell 2009, 15, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Sheikh, S.S.; Romanska, H.M.; Abel, P.; Domin, J.; Lalani, E.-N. Predictive Value of PTEN and AR Coexpression of Sustained Responsiveness to Hormonal Therapy in Prostate Cancer—A Pilot Study. Neoplasia 2008, 10, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Koryakina, Y.; Ta, H.Q.; Gioeli, D. Androgen receptor phosphorylation: Biological context and functional consequences. Endocr.-Relat. Cancer 2014, 21, T131–T145. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-K.; Yeh, S.; Kang, H.-Y.; Chang, C. Akt suppresses androgen-induced apoptosis by phosphorylating and inhibiting androgen receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 7200–7205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, B.S.; Chapinski, C.; Wongvipat, J.; Hieronymus, H.; Chen, Y.; Chandarlapaty, S.; Arora, V.K.; Le, C.; Koutcher, J.; Scher, H.; et al. Reciprocal Feedback Regulation of PI3K and Androgen Receptor Signaling in PTEN-Deficient Prostate Cancer. Cancer Cell 2011, 19, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, A.C.; Trotman, L.C. Turning off AKT: PHLPP as a drug target. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 537–558. [Google Scholar] [CrossRef] [Green Version]
- Serra, V.; Scaltriti, M.; Prudkin, L.; Eichhorn, P.; Ibrahim, Y.H.; Chandarlapaty, S.; Markman, B.; Rodriguez, O.; Guzman, M.; Gili, M.; et al. PI3K inhibition results in enhanced HER signaling and acquired ERK dependency in HER2-overexpressing breast cancer. Oncogene 2011, 30, 2547–2557. [Google Scholar] [CrossRef]
- Toren, P.; Kim, S.; Cordonnier, T.; Crafter, C.; Davies, B.R.; Fazli, L.; Gleave, M.E.; Zoubeidi, A. Combination AZD5363 with Enzalutamide Significantly Delays Enzalutamide-resistant Prostate Cancer in Preclinical Models. Eur. Urol. 2015, 67, 986–990. [Google Scholar] [CrossRef]
- Thomas, C.; Lamoureux, F.; Crafter, C.; Davies, B.R.; Beraldi, E.; Fazli, L.; Kim, S.; Thaper, D.; Gleave, M.E.; Zoubeidi, A. Synergistic Targeting of PI3K/AKT Pathway and Androgen Receptor Axis Significantly Delays Castration-Resistant Prostate Cancer Progression In Vivo. Mol. Cancer Ther. 2013, 12, 2342–2355. [Google Scholar] [CrossRef] [Green Version]
- Crabb, S.J.; Griffiths, G.; Dunkley, D.; Downs, N.; Ellis, M.; Radford, M.; Light, M.; Northey, J.; Whitehead, A.; Wilding, S.; et al. Overall Survival Update for Patients with Metastatic Castration-resistant Prostate Cancer Treated with Capivasertib and Docetaxel in the Phase 2 ProCAID Clinical Trial. Eur. Urol. 2022, 82, 512–515. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, C.; Bracarda, S.; Sternberg, C.N.; Chi, K.N.; Olmos, D.; Sandhu, S.; Massard, C.; Matsubara, N.; Alekseev, B.; Parnis, F.; et al. Ipatasertib plus abiraterone and prednisolone in metastatic castration-resistant prostate cancer (IPATential150): A multicentre, randomised, double-blind, phase 3 trial. Lancet 2021, 398, 131–142. [Google Scholar] [CrossRef]
- De Bono, J.; Hansen, A.; Choudhury, A.; Cook, N.; Heath, E.; Higano, C.; Linch, M.; Martin-Liberal, J.; Rathkopf, D.; Wisinski, K.; et al. AZD8186, a potent and selective inhibitor of PI3Kβ/δ, as monotherapy and in combination with abiraterone acetate plus prednisone (AAP), in patients (pts) with metastatic castrate-resistant prostate cancer (mCRPC). Ann. Oncol. 2018, 29, viii291–viii292. [Google Scholar] [CrossRef]
- Chow, H.; Ghosh, P.M.; deVere White, R.; Evans, C.P.; Dall’Era, M.A.; Yap, S.A.; Li, Y.; Beckett, L.A.; Lara, P.N., Jr.; Pan, C.X. A Phase II clinical trial of everolimus plus bicalutamide for castration-resistant prostate cancer. Cancer 2016, 122, 1897–1904. [Google Scholar] [CrossRef] [PubMed]
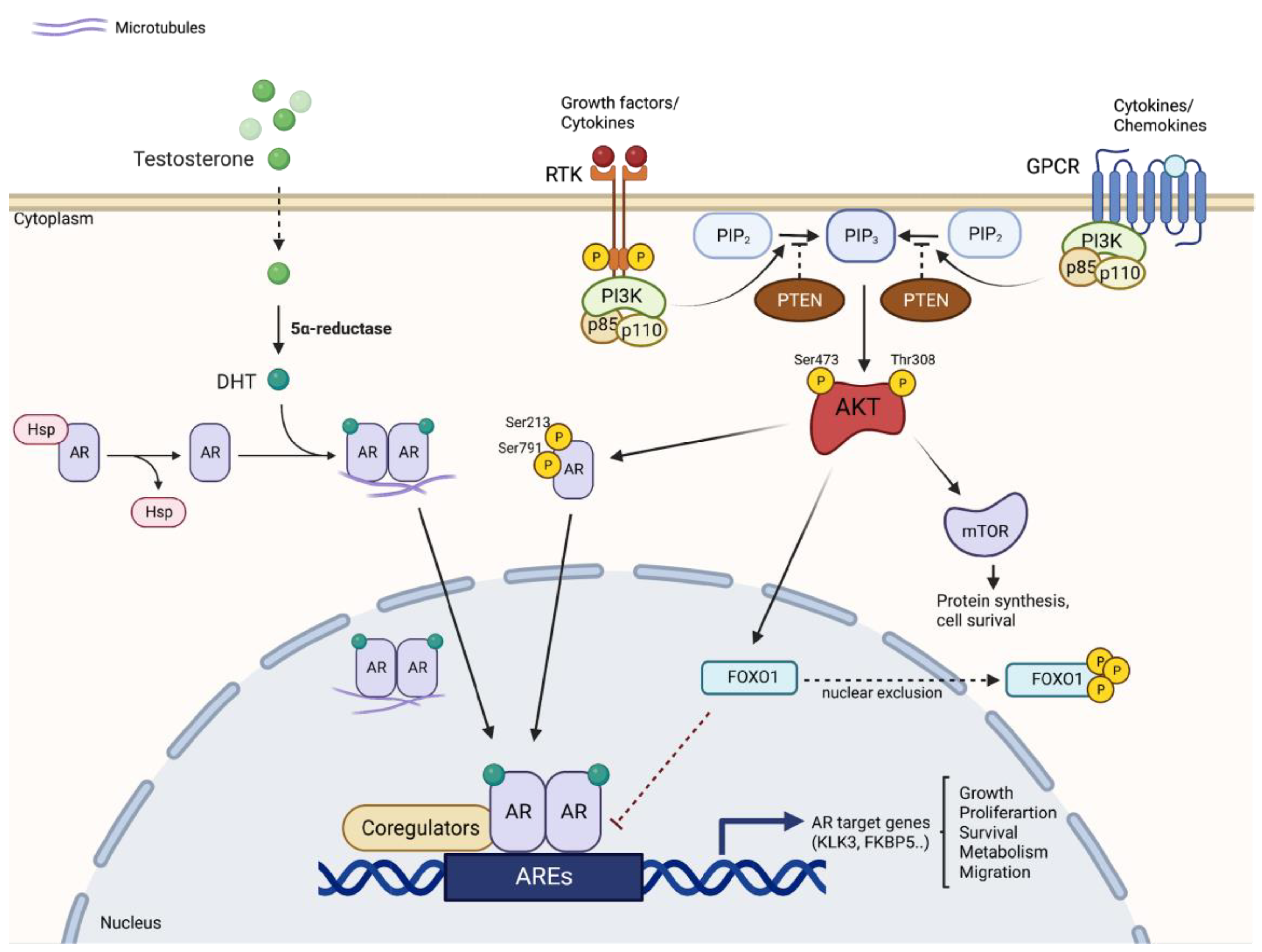
Figure 1.
Crosstalk between PI3K/AKT/mTOR and AR signaling pathways. The two signaling pathways are closely connected and regulated according to a reciprocal feedback mechanism. AR inhibition reduces FKBP5 levels, inhibiting PHLPP-mediated suppression of AKT, thereby activating AKT. Activation of AKT then generates upregulation of AR by several mechanisms, including direct phosphorylation of AR and nuclear exclusion of FOXO1. AKT: protein kinase B; AR: androgen receptor; ARE: androgen response element; FKBP5: FK506 binding protein 5; FOXO1: forkhead box transcription factor 1; GPCR: G-protein coupled receptor; mTOR: mammalian target of rapamycin; PIP3: phosphatidylinositol-3,4,5-triphosphate; PHLPP: PH domain of leucine repeat sequence-rich phosphatase; PSA: prostate-specific antigen; PTEN: phosphatase and tensin homolog; RTK: receptor tyrosine kinase.
Figure 1.
Crosstalk between PI3K/AKT/mTOR and AR signaling pathways. The two signaling pathways are closely connected and regulated according to a reciprocal feedback mechanism. AR inhibition reduces FKBP5 levels, inhibiting PHLPP-mediated suppression of AKT, thereby activating AKT. Activation of AKT then generates upregulation of AR by several mechanisms, including direct phosphorylation of AR and nuclear exclusion of FOXO1. AKT: protein kinase B; AR: androgen receptor; ARE: androgen response element; FKBP5: FK506 binding protein 5; FOXO1: forkhead box transcription factor 1; GPCR: G-protein coupled receptor; mTOR: mammalian target of rapamycin; PIP3: phosphatidylinositol-3,4,5-triphosphate; PHLPP: PH domain of leucine repeat sequence-rich phosphatase; PSA: prostate-specific antigen; PTEN: phosphatase and tensin homolog; RTK: receptor tyrosine kinase.


{kind=link}
Table 1.
Clinical studies on PI3K/AKT/mTOR pathway inhibitors in PCa.
| Target | Agent | Phase | Regimen | Study Population | Status | Clinicaltrial.Gov ID |
|---|---|---|---|---|---|---|
| AKT | MK2206 | II | + Bicalutamide | High-Risk of Progression PCa with rising PSA | Active, not recruiting | NCT01251861 |
| I | + hydroxychloroquine | Stage IV PCa | Active, not recruiting | NCT01480154 | ||
| AZD5363 (Capivasertib) | I | + Enzalutamide | Advanced Solid Tumors Harboring Mutations in AKT1, AKT2, or AKT3 | Active, not recruiting | NCT03310541 | |
| III | + Docetaxel | mCRPC | Recruiting | NCT05348577 | ||
| II | + Abiraterone acetate | High Risk Localized PCa With PTEN Loss | Not yet recruiting | NCT05593497 | ||
| I/II | + Docetaxel/Prednisolone | mCRPC | Completed | NCT02121639 | ||
| Afuresertib | I/II | + LAE001/prednisone | mCRPC | Recruiting | NCT04060394 | |
| PI3K | AZD8186 | I | + Docetaxel | mPCa with PTEN or PIK3CB mutations | Active, not recruiting | NCT03218826 |
| I | + Abiraterone acetate | Advanced CRPC | Completed | NCT01884285 | ||
| GDC-0068 (Ipatasertib) | I/II | + Atezolizumab | PTEN-loss CRPC | Recruiting | NCT03673787 | |
| III | + Abiraterone acetate and prednisone/prednisolone | mCRPC | Active, not recruiting | NCT03072238 | ||
| II | + Abiraterone acetate and prednisone/prednisolone | CRPC previously treated with Docetaxel | Completed | NCT01485861 | ||
| I | + Rucaparib | Advanced PCa | Completed | NCT03840200 | ||
| GSK2636771 | I | + Enzalutamide | PTEN(-) mCRPC | Completed | NCT02215096 | |
| BKM120 (Buparlisib) | Ib | + Abiraterone acetate | CRPC | Completed | NCT01634061 | |
| LY3023414 | II | + Enzalutamide | mCRPC | Completed | NCT02407054 | |
| CYH33 | I | + Olaparib | PCa with DDR and/or PIK3CA mutations | Recruiting | NCT04586335 | |
| mTOR | RAD001 (Everolimus) | III | Monotherapy | CRPC patients with PI3K-AKT-mTOR signaling pathway deficiency | Not yet recruiting | NCT03580239 |
| II | Monotherapy | mCRPC | Completed | NCT00636090 | ||
| I | + Apalutamide | mCRPC after treatment with Abiraterone acetate | Completed | NCT02106507 | ||
| I | + standard radiation therapy | PCa with rising PSA following RP | Completed | NCT01548807 | ||
| II | + Carboplatin/Prednisone | mPCa that progressed after docetaxel | Completed | NCT01051570 | ||
| I/II | + Docetaxel/Prednisone | mCRPC | Completed | NCT00459186 | ||
| II | Monotherapy | HRPC | Completed | NCT00629525 | ||
| I/II | + Docetaxel/Bevacizumab | Advanced PCa | Completed | NCT00574769 | ||
| CCI-779 (Temsirolimus) | I/II | + Cixutumumab | mPCa | Completed | NCT01026623 | |
| II | + conventional surgery | newly diagnosed PCa at high risk of relapse | Completed | NCT00071968 | ||
| CC-115 | I | Monotherapy | PCa | Completed | NCT01353625 | |
| MLN0128 (Sepanisertib) | II | Monotherapy | CRPC | Completed | NCT02091531 | |
| AZD2014 | I | Monotherapy prior to RP | High Risk PCa | Completed | NCT02064608 |
(+) indicates co-treatments; ADT: androgen deprivation therapy; CRPC: castration-resistant prostate cancer; HRPC: hormone-resistant prostate cancer; mPCa: metastatic prostate cancer; mCRPC: metastatic castration-resistant prostate cancer; PSA: prostate-specific antigen; RP: radical prostatectomy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tortorella, E.; Giantulli, S.; Sciarra, A.; Silvestri, I. AR and PI3K/AKT in Prostate Cancer: A Tale of Two Interconnected Pathways. Int. J. Mol. Sci. 2023, 24, 2046. https://doi.org/10.3390/ijms24032046
AMA Style
Tortorella E, Giantulli S, Sciarra A, Silvestri I. AR and PI3K/AKT in Prostate Cancer: A Tale of Two Interconnected Pathways. International Journal of Molecular Sciences. 2023; 24(3):2046. https://doi.org/10.3390/ijms24032046
Chicago/Turabian StyleTortorella, Elisabetta, Sabrina Giantulli, Alessandro Sciarra, and Ida Silvestri. 2023. "AR and PI3K/AKT in Prostate Cancer: A Tale of Two Interconnected Pathways" International Journal of Molecular Sciences 24, no. 3: 2046. https://doi.org/10.3390/ijms24032046
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.