


Altered Extracellular Vesicle miRNA Profile in Prodromal Alzheimer’s Disease
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
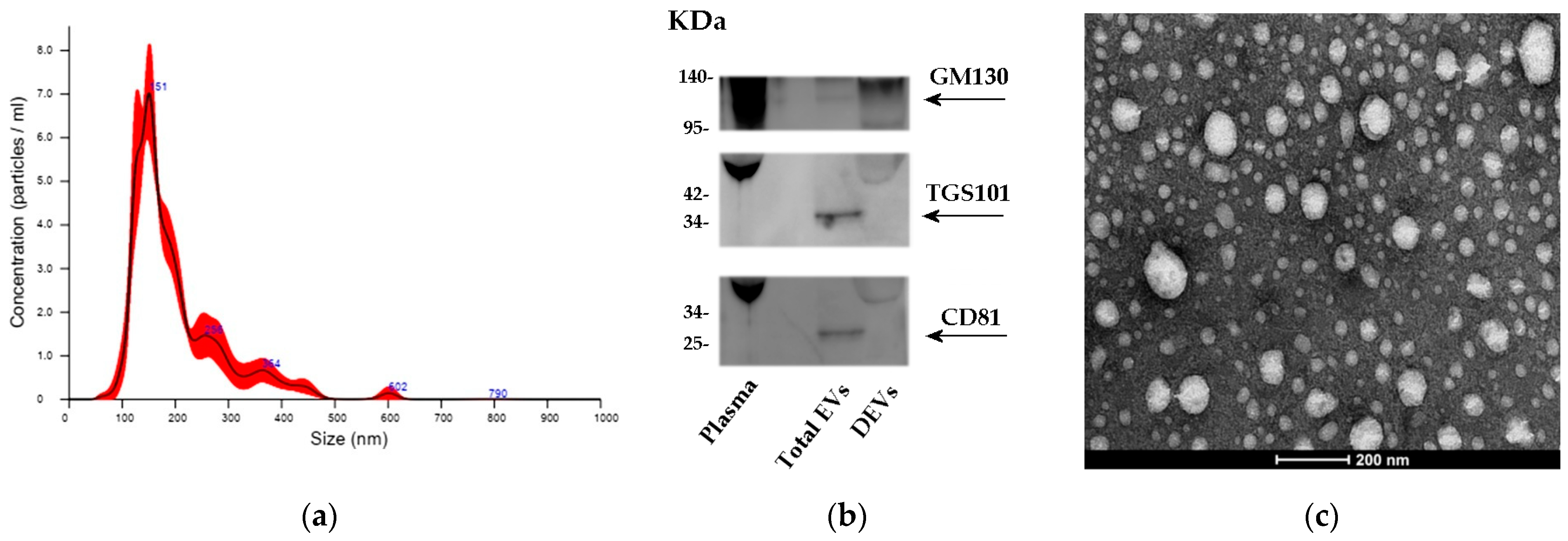
2.1. Characterization of Total EVs from Plasma
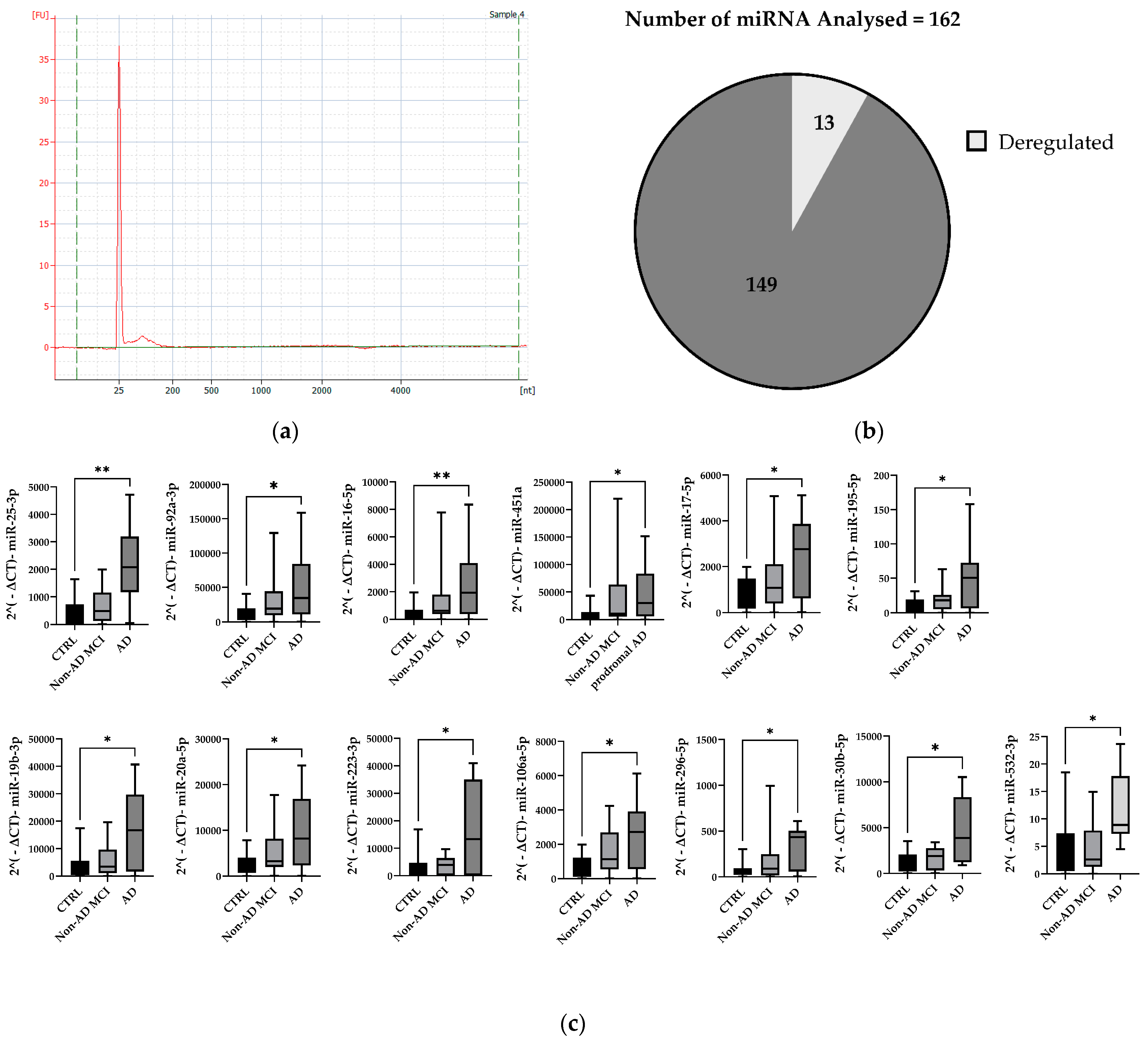
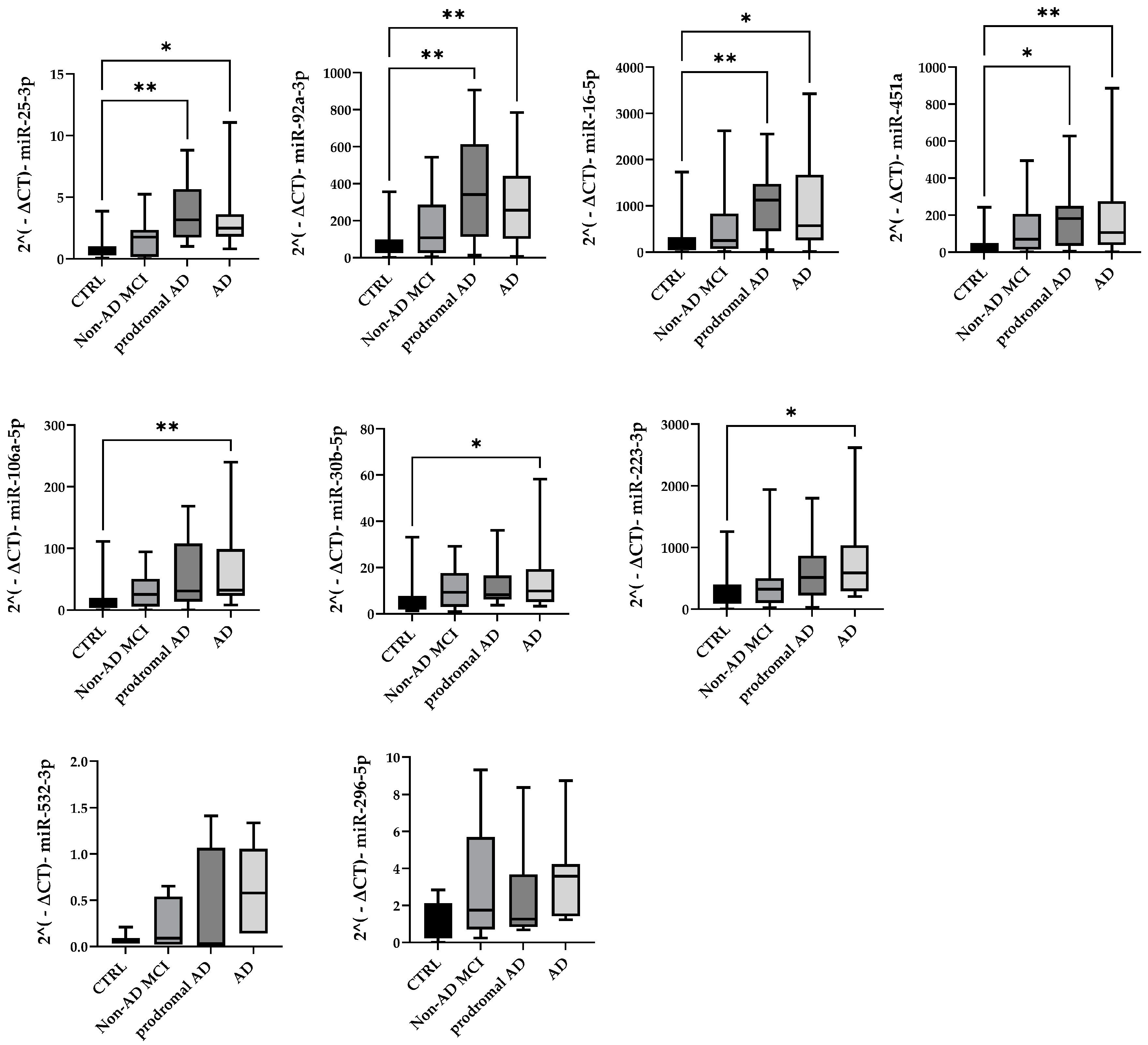
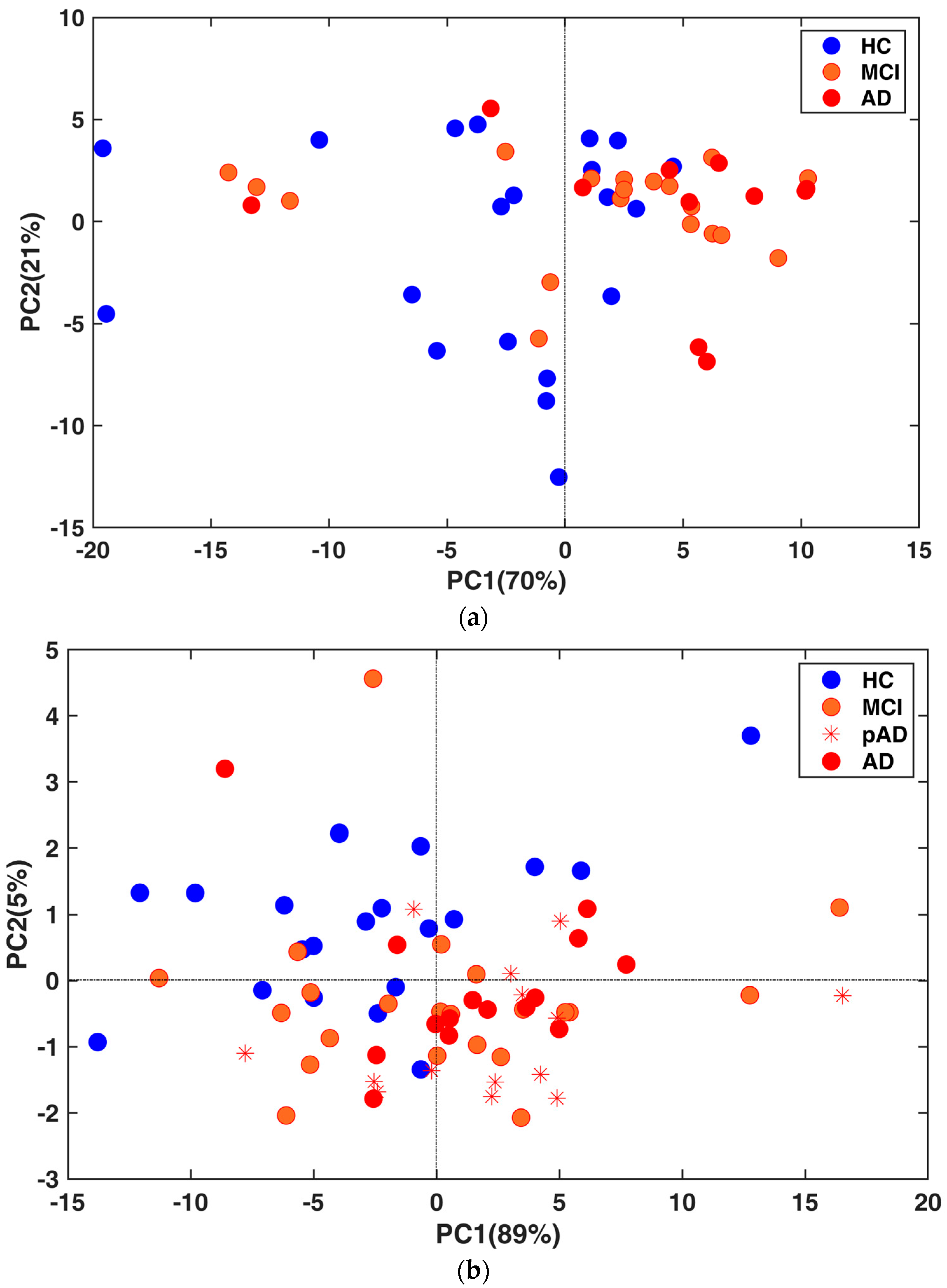
2.2. MiRNA Expression Profile in Total Plasma EVs
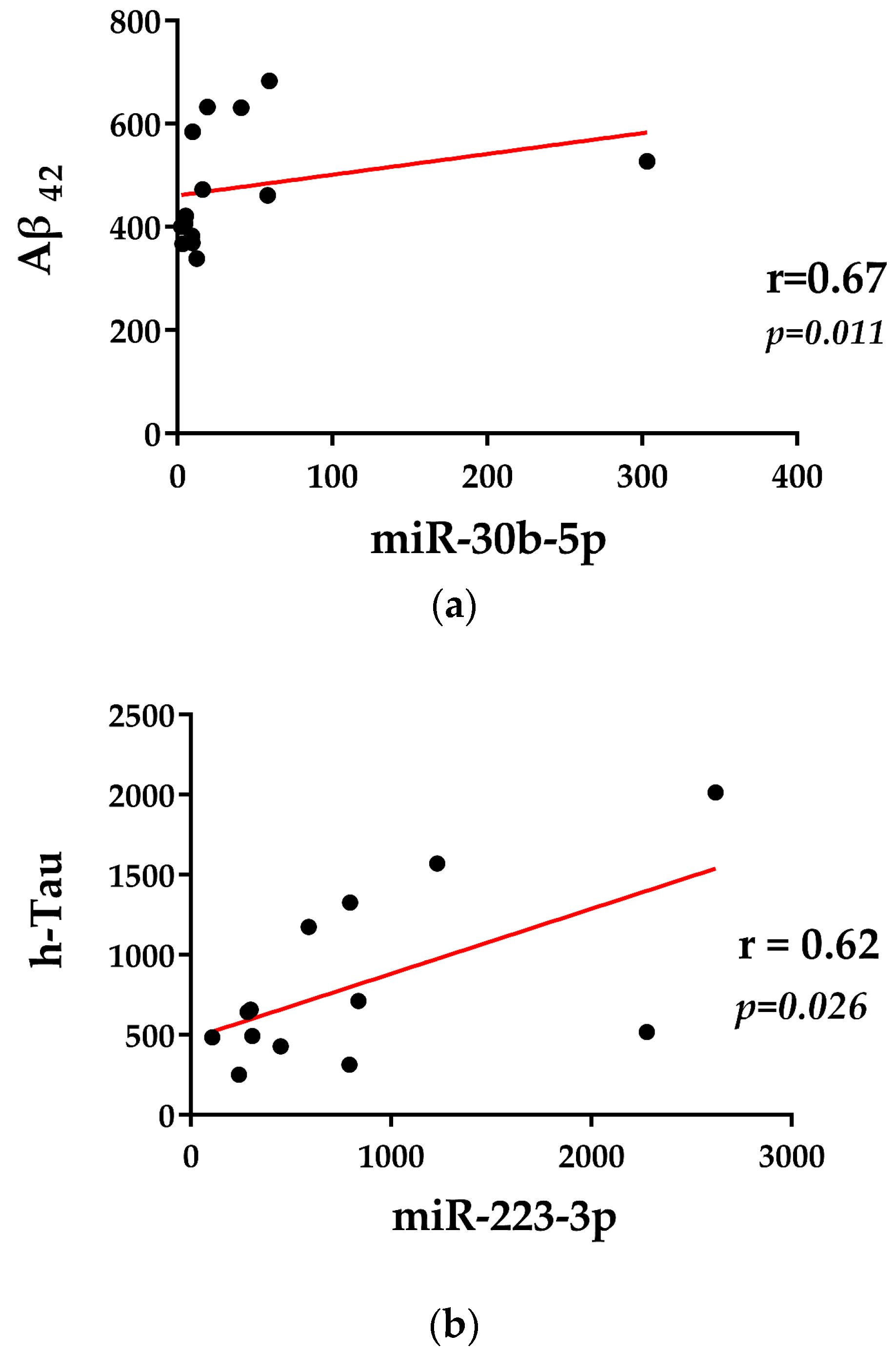
2.3. Correlations with Demographics and CSF Biomarkers
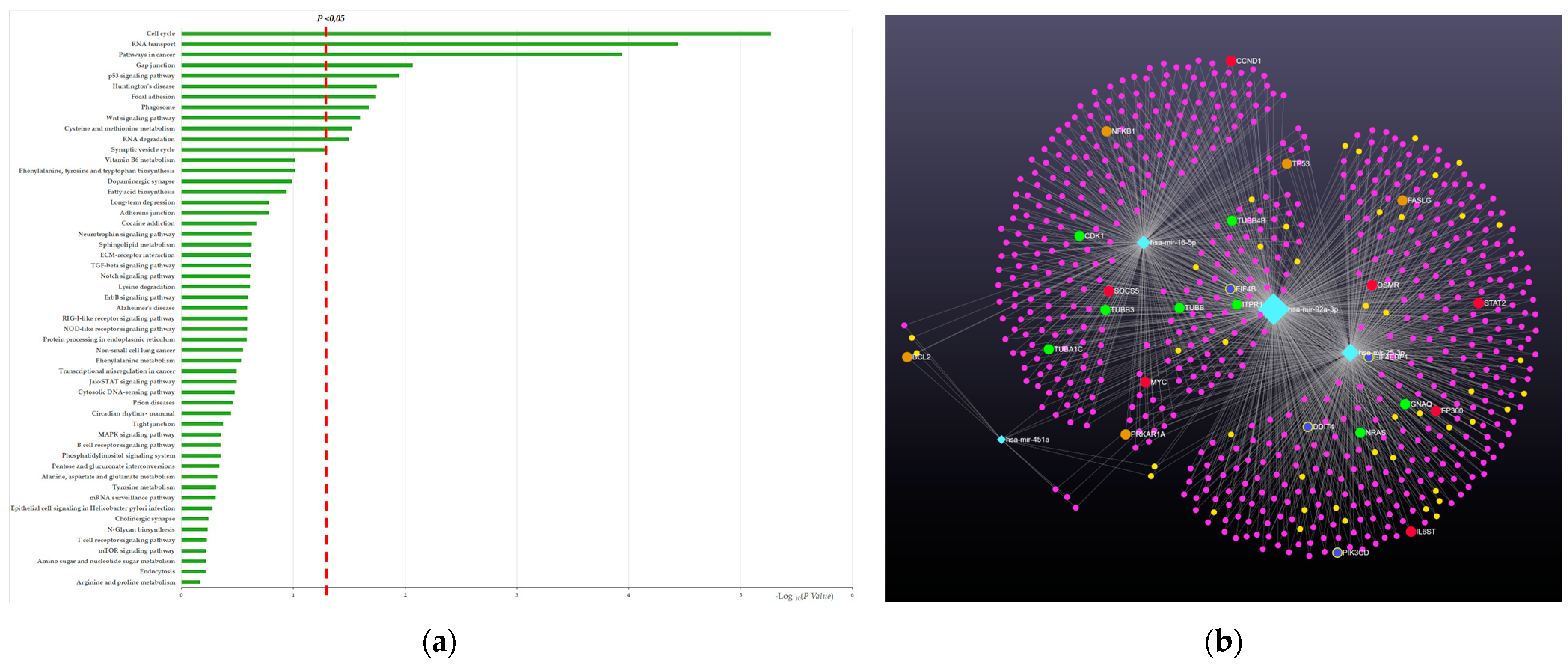
2.4. miRNA Target and Pathway Prediction
3. Discussion
4. Materials and Methods
4.1. Population and Sample Collection
4.2. CSF Processing and Biomarker Analysis
4.3. Isolation of Total Vesicles and Characterization
4.4. RNA Extraction and Quality Control
4.5. Expression Analysis of microRNA Using TaqMan® OpenArray® MicroRNA Panels
4.6. Validation Analysis with Taqman MicroRNA Assays
4.7. Target Prediction and Pathway Enrichment Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liss, J.L.; Assunção, S.S.M.; Cummings, J.; Atri, A.; Geldmacher, D.S.; Candela, S.F.; Devanand, D.P.; Fillit, H.M.; Susman, J.; Mintzer, J.; et al. Practical recommendations for timely, accurate diagnosis of symptomatic Alzheimer’s disease (MCI and dementia) in primary care: A review and synthesis. J. Intern. Med. 2021, 290, 310–334. [Google Scholar] [CrossRef]
- Blennow, K. A Review of Fluid Biomarkers for Alzheimer’s Disease: Moving from CSF to Blood. Neurol. Ther. 2017, 6, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Aharon, A.; Spector, P.; Ahmad, R.S.; Horrany, N.; Sabbach, A.; Brenner, B.; Aharon-Peretz, J. Extracellular Vesicles of Alzheimer’s Disease Patients as a Biomarker for Disease Progression. Mol. Neurobiol. 2020, 57, 4156–4169. [Google Scholar] [CrossRef]
- Aulston, B.; Liu, Q.; Mante, M.; Florio, J.; Rissman, R.A.; Yuan, S.H. Extracellular Vesicles Isolated from Familial Alzheimer’s Disease Neuronal Cultures Induce Aberrant Tau Phosphorylation in the Wild-Type Mouse Brain. J. Alzheimer’s Dis. JAD 2019, 72, 575–585. [Google Scholar] [CrossRef]
- Duggan, M.R.; Lu, A.; Foster, T.C.; Wimmer, M.; Parikh, V. Exosomes in Age-Related Cognitive Decline: Mechanistic Insights and Improving Outcomes. Front. Aging Neurosci. 2022, 14, 834775. [Google Scholar] [CrossRef]
- Xiao, T.; Zhang, W.; Jiao, B.; Pan, C.Z.; Liu, X.; Shen, L. The Role of Exosomes in the Pathogenesis of Alzheimer’ Disease. Transl. Neurodegener. 2017, 6, 3. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A Compendium of RNA, Proteins, Lipids and Metabolites in Extracellular Vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- De Toro, J.; Herschlik, L.; Waldner, C.; Mongini, C. Emerging Roles of Exosomes in Normal and Pathological Conditions: New Insights for Diagnosis and Therapeutic Applications. Front. Immunol. 2015, 6, 203. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Abdelmohsen, K.; Mustapic, M.; Kapogiannis, D.; Gorospe, M. RNA in Extracellular Vesicles. Wiley Interdiscip. Rev. RNA 2017, 8, e1413. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Riva, P.; Ratti, A.; Venturin, M. The Long Non-Coding RNAs in Neurodegenerative Diseases: Novel Mechanisms of Pathogenesis. Curr. Alzheimer Res. 2016, 13, 1219–1231. [Google Scholar] [CrossRef]
- Galimberti, D.; Villa, C.; Fenoglio, C.; Serpente, M.; Ghezzi, L.; Cioffi, S.M.G.; Arighi, A.; Fumagalli, G.; Scarpini, E. Circulating miRNAs as Potential Biomarkers in Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 42, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Reddy, P.H. Are Circulating microRNAs Peripheral Biomarkers for Alzheimer’s Disease? Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 1617–1627. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Campdelacreu, J.; Vilas, D.; Ispierto, L.; Reñé, R.; Álvarez, R.; Armengol, M.P.; Borràs, F.E.; Beyer, K. Exploratory Study on microRNA Profiles from Plasma-Derived Extracellular Vesicles in Alzheimer’s Disease and Dementia with Lewy Bodies. Transl. Neurodegener. 2019, 8, 1–17. [Google Scholar] [CrossRef]
- Sproviero, D.; Gagliardi, S.; Zucca, S.; Arigoni, M.; Giannini, M.; Garofalo, M.; Olivero, M.; Dell’Orco, M.; Pansarasa, O.; Bernuzzi, S.; et al. Different miRNA Profiles in Plasma Derived Small and Large Extracellular Vesicles from Patients with Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 2737. [Google Scholar] [CrossRef]
- Martinez, B.; Peplow, P.V. MicroRNAs as Diagnostic and Therapeutic Tools for Alzheimer’s Disease: Advances and Limitations. Neural Regen. Res. 2019, 14, 242–255. [Google Scholar] [CrossRef]
- Serpente, M.; Fenoglio, C.; D’Anca, M.; Arcaro, M.; Sorrentino, F.; Visconte, C.; Arighi, A.; Fumagalli, G.G.; Porretti, L.; Cattaneo, A.; et al. MiRNA Profiling in Plasma Neural-Derived Small Extracellular Vesicles from Patients with Alzheimer’s Disease. Cells 2020, 9, 1443. [Google Scholar] [CrossRef]
- Hulse, A.M.; Cai, J.J. Genetic Variants Contribute to Gene Expression Variability in Humans. Genetics 2013, 193, 95–108. [Google Scholar] [CrossRef]
- Lukiw, W.J. Variability in Micro RNA (miRNA) Abundance, Speciation and Complexity amongst Different Human Populations and Potential Relevance to Alzheimer’s Disease (AD). Front. Cell Neurosci. 2013, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- Visconte, C.; Golia, M.T.; Fenoglio, C.; Serpente, M.; Gabrielli, M.; Arcaro, M.; Sorrentino, F.; Busnelli, M.; Arighi, A.; Fumagalli, G.; et al. Plasma Microglial-Derived Extracellular Vesicles Are Increased in Frail Patients with Mild Cognitive Impairment and Exert a Neurotoxic Effect. Geroscience 2023, 45, 1557–1571. [Google Scholar] [CrossRef] [PubMed]
- Fitz, N.F.; Wang, J.; Kamboh, M.I.; Koldamova, R.; Lefterov, I. Small Nucleolar RNAs in Plasma Extracellular Vesicles and Their Discriminatory Power as Diagnostic Biomarkers of Alzheimer’s Disease. Neurobiol. Dis. 2021, 159, 105481. [Google Scholar] [CrossRef] [PubMed]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 Cluster: A Comprehensive Update on Its Genomics, Genetics, Functions and Increasingly Important and Numerous Roles in Health and Disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Khuu, C.; Utheim, T.P.; Sehic, A. The Three Paralogous MicroRNA Clusters in Development and Disease, miR-17-92, miR-106a-363, and miR-106b-25. Scientifica 2016, 2016, 1379643. [Google Scholar] [CrossRef]
- Mendell, J.T. miRiad Roles for the miR-17-92 Cluster in Development and Disease. Cell 2008, 133, 217–222. [Google Scholar] [CrossRef]
- McKeever, P.M.; Schneider, R.; Taghdiri, F.; Weichert, A.; Multani, N.; Brown, R.A.; Boxer, A.L.; Karydas, A.; Miller, B.; Robertson, J.; et al. MicroRNA Expression Levels Are Altered in the Cerebrospinal Fluid of Patients with Young-Onset Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 8826–8841. [Google Scholar] [CrossRef]
- Riancho, J.; Vázquez-Higuera, J.L.; Pozueta, A.; Lage, C.; Kazimierczak, M.; Bravo, M.; Calero, M.; Gonalezález, A.; Rodríguez, E.; Lleó, A.; et al. MicroRNA Profile in Patients with Alzheimer’s Disease: Analysis of miR-9-5p and miR-598 in Raw and Exosome Enriched Cerebrospinal Fluid Samples. J. Alzheimer’s Dis. JAD 2017, 57, 483–491. [Google Scholar] [CrossRef]
- Harrison, E.B.; Hochfelder, C.G.; Lamberty, B.G.; Meays, B.M.; Morsey, B.M.; Kelso, M.L.; Fox, H.S.; Yelamanchili, S.V. Traumatic Brain Injury Increases Levels of miR-21 in Extracellular Vesicles: Implications for Neuroinflammation. FEBS Open Bio 2016, 6, 835–846. [Google Scholar] [CrossRef]
- Liu, C.G.; Song, J.; Zhang, Y.Q.; Wang, P.C. MicroRNA-193b Is a Regulator of Amyloid Precursor Protein in the Blood and Cerebrospinal Fluid Derived Exosomal microRNA-193b Is a Biomarker of Alzheimer’s Disease. Mol. Med. Rep. 2014, 10, 2395–2400. [Google Scholar] [CrossRef]
- Soares Martins, T.; Trindade, D.; Vaz, M.; Campelo, I.; Almeida, M.; Trigo, G.; da Cruz Silva, O.A.B.; Gabriela Henriques, A. Diagnostic and Therapeutic Potential of Exosomes in Alzheimer’s Disease. J. Neurochem. 2020, 156, 162–181. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Doecke, J.D.; Sharples, R.A.; Villemagne, V.L.; Fowler, C.J.; Rembach, A.; Martins, R.N.; Rowe, C.C.; Macaulay, S.L.; Masters, C.L.; et al. Prognostic Serum miRNA Biomarkers Associated with Alzheimer’s Disease Shows Concordance with Neuropsychological and Neuroimaging Assessment. Mol. Psychiatry 2015, 20, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes Provide a Protective and Enriched Source of miRNA for Biomarker Profiling Compared to Intracellular and Cell-Free Blood. J. Extracell. Vesicles 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Vella, L.J.; Barnham, K.J.; McLean, C.; Masters, C.L.; Hill, A.F. Small RNA Fingerprinting of Alzheimer’s Disease Frontal Cortex Extracellular Vesiclesand Their Comparison with Peripheral Extracellular Vesicles. J. Extracell. Vesicles 2020, 9, 1766822. [Google Scholar] [CrossRef] [PubMed]
- Lugli, G.; Cohen, A.M.; Bennett, D.A.; Shah, R.C.; Fields, C.J.; Hernandez, A.G.; Smalheiser, N.R. Plasma Exosomal miRNAs in Persons with and without Alzheimer Disease: Altered Expression and Prospects for Biomarkers. PLoS ONE 2015, 10, e0139233. [Google Scholar] [CrossRef]
- Nie, C.; Sun, Y.; Zhen, H.; Guo, M.; Ye, J.; Liu, Z.; Yang, Y.; Zhang, X. Differential Expression of Plasma Exo-miRNA in Neurodegenerative Diseases by Next-Generation Sequencing. Front. Neurosci. 2020, 14, 438. [Google Scholar] [CrossRef]
- Li, F.; Xie, X.Y.; Sui, X.F.; Wang, P.; Chen, Z.; Zhang, J.B. Profile of Pathogenic Proteins and MicroRNAs in Plasma-Derived Extracellular Vesicles in Alzheimer’s Disease: A Pilot Study. Neuroscience 2020, 432, 240–246. [Google Scholar] [CrossRef]
- Bleazard, T.; Lamb, J.A.; Griffiths-Jones, S. Bias in microRNA Functional Enrichment Analysis. Bioinformatics 2015, 31, 1592–1598. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian mRNAs Are Conserved Targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Tanzer, A.; Stadler, P.F. Molecular Evolution of a microRNA Cluster. J. Mol. Biol. 2004, 339, 327–335. [Google Scholar] [CrossRef]
- Ventura, A.; Young, A.G.; Winslow, M.M.; Lintault, L.; Meissner, A.; Erkeland, S.J.; Newman, J.; Bronson, R.T.; Crowley, D.; Stone, J.R.; et al. Targeted Deletion Reveals Essential and Overlapping Functions of the miR-17 through 92 Family of miRNA Clusters. Cell 2008, 132, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Boström, A.-K.; Impens, F.; Fredlund, E.; Van Peer, G.; De Antonellis, P.; von Stedingk, K.; Ghesquière, B.; Schulte, S.; Dews, M.; et al. The miR-17-92 microRNA Cluster Regulates Multiple Components of the TGF-β Pathway in Neuroblastoma. Mol. Cell 2010, 40, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zhang, Y.; Jiang, Y.; Tan, M.; Liu, C. Circular RNA Rbms1 Inhibited the Development of Myocardial Ischemia Reperfusion Injury by Regulating miR-92a/BCL2L11 Signaling Pathway. Bioengineered 2022, 13, 3082–3092. [Google Scholar] [CrossRef] [PubMed]
- Xiu, M.; Luan, H.; Gu, X.; Liu, C.; Xu, D. MicroRNA-17-5p Protects against Propofol Anesthesia-Induced Neurotoxicity and Autophagy Impairment via Targeting BCL2L11. Comput. Math. Methods Med. 2022, 2022, 6018037. [Google Scholar] [CrossRef]
- Petrocca, F.; Vecchione, A.; Croce, C.M. Emerging Role of miR-106b-25/miR-17-92 Clusters in the Control of Transforming Growth Factor Beta Signaling. Cancer Res. 2008, 68, 8191–8194. [Google Scholar] [CrossRef]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Bergmans, B.; Papadopoulou, A.S.; Delacourte, A.; De Strooper, B. MicroRNA Regulation of Alzheimer’s Amyloid Precursor Protein Expression. Neurobiol. Dis. 2009, 33, 422–428. [Google Scholar] [CrossRef]
- Rani, A.; O’Shea, A.; Ianov, L.; Cohen, R.A.; Woods, A.J.; Foster, T.C. miRNA in Circulating Microvesicles as Biomarkers for Age-Related Cognitive Decline. Front. Aging Neurosci. 2017, 9, 323. [Google Scholar] [CrossRef]
- Grillari, J.; Hackl, M.; Grillari-Voglauer, R. miR-17-92 Cluster: Ups and Downs in Cancer and Aging. Biogerontology 2010, 11, 501–506. [Google Scholar] [CrossRef]
- Hackl, M.; Brunner, S.; Fortschegger, K.; Schreiner, C.; Micutkova, L.; Mück, C.; Laschober, G.T.; Lepperdinger, G.; Sampson, N.; Berger, P.; et al. miR-17, miR-19b, miR-20a, and miR-106a Are down-Regulated in Human Aging. Aging Cell 2010, 9, 291–296. [Google Scholar] [CrossRef]
- Feng, H.; Hu, P.; Chen, Y.; Sun, H.; Cai, J.; He, X.; Cao, Q.; Yin, M.; Zhang, Y.; Li, Q.; et al. Decreased miR-451a in Cerebrospinal Fluid, a Marker for Both Cognitive Impairment and Depressive Symptoms in Alzheimer’s Disease. Theranostics 2023, 13, 3021–3040. [Google Scholar] [CrossRef]
- Xie, B.; Zhou, H.; Zhang, R.; Song, M.; Yu, L.; Wang, L.; Liu, Z.; Zhang, Q.; Cui, D.; Wang, X.; et al. Serum miR-206 and miR-132 as Potential Circulating Biomarkers for Mild Cognitive Impairment. J. Alzheimers Dis. 2015, 45, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Fa, W.; Wang, N.; Peng, Y.; Liu, C.; Zhu, M.; Tian, N.; Wang, Y.; Han, X.; Qiu, C.; et al. Exosomal miR-532-5p Induced by Long-Term Exercise Rescues Blood-Brain Barrier Function in 5XFAD Mice via Downregulation of EPHA4. Aging Cell 2023, 22, e13748. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Huang, M.; Guo, L.; Zhu, L.; Hou, J.; Zhang, L.; Pero, A.; Ng, S.; El Gaamouch, F.; Elder, G.; et al. MicroRNA-195 Rescues ApoE4-Induced Cognitive Deficits and Lysosomal Defects in Alzheimer’s Disease Pathogenesis. Mol. Psychiatry 2021, 26, 4687–4701. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xu, Y.; Xu, W.; Zhou, Q.; Chen, Q.; Yang, M.; Feng, F.; Liu, Y.; Zhu, X.; Yu, M.; et al. Serum Exosomal miR-223 Serves as a Potential Diagnostic and Prognostic Biomarker for Dementia. Neuroscience 2018, 379, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Hu, M.; Zhang, J.; Teng, Z.-Q.; Chen, C. A Novel Mechanism of Synaptic and Cognitive Impairments Mediated via microRNA-30b in Alzheimer’s Disease. EBioMedicine 2019, 39, 409–421. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, S.H.; Park, Y.; Park, J.; Lee, J.H.; Kim, B.C.; Song, W.K. miR-16-5p Is Upregulated by Amyloid β Deposition in Alzheimer’s Disease Models and Induces Neuronal Cell Apoptosis through Direct Targeting and Suppression of BCL-2. Exp. Gerontol. 2020, 136, 110954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, C.-F.; Wang, A.-H.; Lin, Q.-F. MiR-16 Regulates Cell Death in Alzheimer’s Disease by Targeting Amyloid Precursor Protein. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4020–4027. [Google Scholar]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.-M.; Kellermann, O. miR-16 Targets the Serotonin Transporter: A New Facet for Adaptive Responses to Antidepressants. Science 2010, 329, 1537–1541. [Google Scholar] [CrossRef]
- Zhu, H.-C.; Wang, L.-M.; Wang, M.; Song, B.; Tan, S.; Teng, J.-F.; Duan, D.-X. MicroRNA-195 Downregulates Alzheimer’s Disease Amyloid-β Production by Targeting BACE1. Brain Res. Bull. 2012, 88, 596–601. [Google Scholar] [CrossRef]
- Kanduri, C. Kcnq1ot1: A Chromatin Regulatory RNA. Semin. Cell Dev. Biol. 2011, 22, 343–350. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, G.; He, M.; Huang, H.; Cao, L.; Yin, A.; Wang, P.; Wang, L. SZRD1 Is a Novel Protein That Functions as a Potential Tumor Suppressor in Cervical Cancer. J. Cancer 2017, 8, 2132–2141. [Google Scholar] [CrossRef]
- Laugesen, A.; Højfeldt, J.W.; Helin, K. Role of the Polycomb Repressive Complex 2 (PRC2) in Transcriptional Regulation and Cancer. Cold Spring Harb. Perspect. Med. 2016, 6, a026575. [Google Scholar] [CrossRef]
- Blackledge, N.P.; Rose, N.R.; Klose, R.J. Targeting Polycomb Systems to Regulate Gene Expression: Modifications to a Complex Story. Nat. Rev. Mol. Cell Biol. 2015, 16, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Kouznetsova, V.L.; Tchekanov, A.; Li, X.; Yan, X.; Tsigelny, I.F. Polycomb Repressive 2 Complex—Molecular Mechanisms of Function. Protein Sci. 2019, 28, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- von Schimmelmann, M.; Feinberg, P.A.; Sullivan, J.M.; Ku, S.M.; Badimon, A.; Duff, M.K.; Wang, Z.; Lachmann, A.; Dewell, S.; Ma’ayan, A.; et al. Polycomb Repressive Complex 2 (PRC2) Silences Genes Responsible for Neurodegeneration. Nat. Neurosci. 2016, 19, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.; Herdy, J.R.; Traxler, L.; Schafer, S.T.; Schlachetzki, J.C.M.; Böhnke, L.; Reid, D.A.; Lee, H.; Zangwill, D.; Fernandes, D.P.; et al. Age-Dependent Instability of Mature Neuronal Fate in Induced Neurons from Alzheimer’s Patients. Cell Stem Cell 2021, 28, 1533–1548. [Google Scholar] [CrossRef]
- Pollard, H.B.; Arispe, N.; Rojas, E. Ion Channel Hypothesis for Alzheimer Amyloid Peptide Neurotoxicity. Cell. Mol. Neurobiol. 1995, 15, 513–526. [Google Scholar] [CrossRef]
- Koh, S.-H.; Noh, M.Y.; Kim, S.H. Amyloid-Beta-Induced Neurotoxicity Is Reduced by Inhibition of Glycogen Synthase Kinase-3. Brain Res. 2008, 1188, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Youn, K.; Yoon, J.-H.; Lee, B.; Kim, D.H.; Jun, M. The Role of Fucoxanthin as a Potent Nrf2 Activator via Akt/GSK-3β/Fyn Axis against Amyloid-β Peptide-Induced Oxidative Damage. Antioxidants 2023, 12, 629. [Google Scholar] [CrossRef]
- Arcaro, M.; Fenoglio, C.; Serpente, M.; Arighi, A.; Fumagalli, G.G.; Sacchi, L.; Floro, S.; D’Anca, M.; Sorrentino, F.; Visconte, C.; et al. A Novel Automated Chemiluminescence Method for Detecting Cerebrospinal Fluid Amyloid-Beta 1-42 and 1-40, Total Tau and Phosphorylated-Tau: Implications for Improving Diagnostic Performance in Alzheimer’s Disease. Biomedicines 2022, 10, 2667. [Google Scholar] [CrossRef]
- Dubois, B.; Feldman, H.H.; Jacova, C.; Hampel, H.; Molinuevo, J.L.; Blennow, K.; DeKosky, S.T.; Gauthier, S.; Selkoe, D.; Bateman, R.; et al. Advancing Research Diagnostic Criteria for Alzheimer’s Disease: The IWG-2 Criteria. Lancet Neurol. 2014, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | AD | Non-AD MCI | CTRL |
|---|---|---|---|
| N | 11 | 19 | 20 |
| Gender (M:F) | 4:7 | 9:10 | 8:12 |
| Mean aβ42 ± SEM (pg/mL) | 551 ± 1.79 | 869.63 ± 2.48 | - |
| Mean h-tau ± SEM (pg/mL) | 842.64 ± 58 | 434.74 ± 3.7 | - |
| Mean p-tau ± SEM (pg/mL) | 95.18 ± 1.18 | 72.53 ± 1.5 | - |
| Mean Age (years ± SD) | 75 ± 1 | 75 ± 6 | 79 ± 6 |
| MMSE (mean ± SD) | 18.30 ± 5.58 | 26.37 ± 2.61 | 28.15 ± 1.35 |
| Variable | AD | Prodromal AD | Non-AD MCI | CTRL |
|---|---|---|---|---|
| N | 15 | 14 | 22 | 21 |
| Gender (M:F) | 7:8 | 4:10 | 12:10 | 6:15 |
| Mean aβ42 ± SEM (pg/mL) | 479.47 ± 1.31 | 518.79 ± 1.67 | 916.14 ± 2.33 | - |
| Mean h-tau ± SEM (pg/mL) | 829.40 ± 4.89 | 661.36 ± 4.17 | 410.24 ± 3.00 | - |
| Mean p-tau ± SEM (pg/mL) | 94.13 ± 1.01 | 112.57 ± 1.55 | 64.38 ± 1.30 | - |
| Mean Age (years ± SD) | 71 ± 8 | 71 ± 9 | 74 ± 6 | 80 ± 4 |
| MMSE (mean ± SD) | 18.36 ± 6.98 | 26.77 ± 1.96 | 26.68 ± 2.12 | 28.15 ± 1.27 |
| miRNA | Log FC | p-Value |
|---|---|---|
| hsa-miR-16-5p | 3.058795 | 0.006041 |
| hsa-miR-92a-3p | 2.339818 | 0.007746 |
| hsa-miR-106a-5p | 2.251733 | 0.006168 |
| hsa-miR-451a | 2.875201 | 0.012556 |
| hsa-miR-19b-3p | 2.642532 | 0.005323 |
| hsa-miR-17-5p | 2.078854 | 0.009754 |
| hsa-miR-223-3p | 4.468833 | 0.015466 |
| hsa-miR-296-5p | 1.851545 | 0.014716 |
| hsa-miR-20a-5p | 2.042569 | 0.014164 |
| hsa-miR-195-5p | 2.541182 | 0.016437 |
| hsa-miR-532-3p | 2.709 | 0.018207 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visconte, C.; Fenoglio, C.; Serpente, M.; Muti, P.; Sacconi, A.; Rigoni, M.; Arighi, A.; Borracci, V.; Arcaro, M.; Arosio, B.; et al. Altered Extracellular Vesicle miRNA Profile in Prodromal Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 14749. https://doi.org/10.3390/ijms241914749
Visconte C, Fenoglio C, Serpente M, Muti P, Sacconi A, Rigoni M, Arighi A, Borracci V, Arcaro M, Arosio B, et al. Altered Extracellular Vesicle miRNA Profile in Prodromal Alzheimer’s Disease. International Journal of Molecular Sciences. 2023; 24(19):14749. https://doi.org/10.3390/ijms241914749
Chicago/Turabian StyleVisconte, Caterina, Chiara Fenoglio, Maria Serpente, Paola Muti, Andrea Sacconi, Marta Rigoni, Andrea Arighi, Vittoria Borracci, Marina Arcaro, Beatrice Arosio, and et al. 2023. "Altered Extracellular Vesicle miRNA Profile in Prodromal Alzheimer’s Disease" International Journal of Molecular Sciences 24, no. 19: 14749. https://doi.org/10.3390/ijms241914749