
Virus Infection and mRNA Nuclear Export

1
University of Chinese Academy of Sciences, Beijing 100049, China
2
Shanxi Provincial Key Laboratory of Protein Structure Determination, Shanxi Academy of Advanced Research and Innovation, Taiyuan 030012, China
3
Key Laboratory of Molecular Virology and Immunology, Chinese Academy of Sciences, Shanghai 200031, China
4
Shanghai Huashen Institute of Microbes and Infections, Shanghai 200052, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(16), 12593; https://doi.org/10.3390/ijms241612593
Submission received: 21 April 2023
/
Revised: 29 July 2023
/
Accepted: 3 August 2023
/
Published: 9 August 2023
(This article belongs to the Special Issue RNA-Binding Proteins — Structure, Function, Networks and Diseases)
Abstract
:Gene expression in eukaryotes begins with transcription in the nucleus, followed by the synthesis of messenger RNA (mRNA), which is then exported to the cytoplasm for its translation into proteins. Along with transcription and translation, mRNA export through the nuclear pore complex (NPC) is an essential regulatory step in eukaryotic gene expression. Multiple factors regulate mRNA export and hence gene expression. Interestingly, proteins from certain types of viruses interact with these factors in infected cells, and such an interaction interferes with the mRNA export of the host cell in favor of viral RNA export. Thus, these viruses hijack the host mRNA nuclear export mechanism, leading to a reduction in host gene expression and the downregulation of immune/antiviral responses. On the other hand, the viral mRNAs successfully evade the host surveillance system and are efficiently exported from the nucleus to the cytoplasm for translation, which enables the continuation of the virus life cycle. Here, we present this review to summarize the mechanisms by which viruses suppress host mRNA nuclear export during infection, as well as the key strategies that viruses use to facilitate their mRNA nuclear export. These studies have revealed new potential antivirals that may be used to inhibit viral mRNA transport and enhance host mRNA nuclear export, thereby promoting host gene expression and immune responses.
1. Introduction
The presence of a nucleus is the primary distinguishing factor between eukaryotic and prokaryotic organisms. The nucleus is a membrane-bound organelle found in eukaryotic cells that contains genetic material, including DNA and RNA, which directs the cellular activities of the organism. The nucleus is surrounded by the nuclear envelope, which is a double-layered membrane and acts as a physical barrier to protect the genetic material from damage and regulates the transport of molecules in and out of the nucleus. The nuclear pore complex (NPC) is a large protein complex that spans the double-layered membrane of the nuclear envelope in eukaryotic cells. The NPC acts as a gatekeeper, controlling the selective transport of molecules such as proteins, RNA, and other macromolecules between the nucleus and the cytoplasm [1,2,3].
To adapt to diverse stimuli and environmental cues, such as pathogen (viral, bacterial, or fungal) infections, cells must change their genetic expression and regulatory mechanisms. These changes include the activation or repression of specific genes through transcriptional regulation, as well as the processing, splicing, and degradation of mRNAs through post-transcriptional regulation. The resulting changes in gene expression can alter the production of proteins and other molecules that are critical for the cellular response to the stimulus or cue. By modulating gene expression, cells can adapt to changing conditions and mount an effective response to various challenges, including pathogen infections [4,5].
Regulations in innate immune responses to pathogens involve various molecular and cellular mechanisms that help a host defend against infection. These mechanisms include the recognition of pathogens by pattern recognition receptors (PRRs), the activation of immune signaling pathways, the transcriptional and post-transcriptional regulation of immune-related gene expression, and the regulation of nucleo-cytoplasmic trafficking. Nuclear export of mRNA, which encodes proteins for cellular defense, is an essential part of this regulation and allows the host to develop an effective response to pathogenic threats [6,7,8]. Viruses are considered to be one of the most significant types of pathogens due to their ability to cause a range of diseases in humans and other organisms. Compared with other pathogens, viruses are unique in their nature as they cannot reproduce or carry out metabolic processes without a host cell. They infect host cells and hijack their cellular machinery for their survival, with effects on their hosts ranging from trivial to deadly [9]. Viruses possess a variety of strategies to impair the host cell’s response to infection. As the nuclear export of host mRNAs is a critical step for the production of antiviral proteins, certain viruses have developed various strategies to effectively block host mRNAs nuclear export, including targeting transcription factors, mRNA processing pathway, and mRNA export factors to impair their functions in the nuclear export of mRNAs. With different genomic material, viruses can be divided into DNA virus and RNA virus. During infection, viruses replicate in the cells by hijacking cellular mechanisms, such as transcription, mRNA processing, mRNA transportation, and translation [5,9,10].
The cellular nuclear export machinery is used by most DNA viruses and a few RNA viruses whose replication occurs in the nucleus [11]. Studies of the mechanisms by which viruses export viral RNA have shown that two important nuclear export factors for host RNA, named CRM1 (chromosome region maintenance 1, also known as exportin 1 or XPO1) and NXF1 (nuclear export factor 1, also known as tip-associated protein or TAP) [12,13,14], are utilized by several types of viruses to promote the export of viral mRNA while also inhibiting the transport of host mRNA to prevent an appropriate host immune response [15,16].
The mRNA precursors (pre-mRNAs) must be properly processed into mature mRNAs by capping, splicing, and polyadenylation before being exported from the nucleus [17,18]. Following these processing, mRNA export factors start loading on the mRNA and draw the export receptor. The transcriptional export (TREX) complex coordinates the nuclear export of the mRNA via transcription and processing [19,20,21]. This complex consists of a multisubunit THO complex [20,22,23,24] that plays a role in transcription elongation, the export factor UAP56 (also known as HEL), and Aly/REF (also known as THOC4). UAP56 binds to Aly/REF and facilitates the binding of Aly/REF to mRNAs [25,26,27]. Aly/REF interacts with the main mRNA export receptor nuclear RNA export factor 1 (NXF1, also known as TAP)–Ntf2-like export factor 1 (NXT1, also known as TAP-p15), a heterodimer that mediates the nuclear export of mRNA by binding to the phenylalanine-glycine (FG) repeat domain of Nups [14,28,29,30]. Nups containing FG, such as Nup98, are translocated via NPC-mediated mRNP (messenger ribonucleoprotein) complexes [30,31]. Rae1 (also known as Mrnp41) is another mRNA export factor involved in this process. It is a nucleo-cytoplasmic shuttle protein that interacts with mRNA, NXF1, and Nup98 [32,33,34,35,36,37,38,39,40]. Rae1 can recruit NXF1 to Nup98 [34,41,42,43]. CRM1 is another important receptor which is known to mediate the nuclear export certain types of RNA, including miRNA, and mRNA subgroups [41,42,43]. RNA nuclear export via CRM1 requires an adaptor protein that binds to cargo RNA, which interacts with CRM1 via nuclear export signal (NES) [44].
Although many host cell processes are involved in viral replication and assembly, we discuss below the mechanisms by which viruses hijack or usurp the host’s mRNA export pathway to inhibit the host’s mRNA nuclear export, thereby reducing the expression of the host’s immune genes and causing a successful and sustained infection [45,46].
2. Cellular mRNA Nuclear Export
mRNA nuclear export, a key step before functional protein production, is a crucial process that plays an important role in gene expression regulation. In eukaryotic cells, transcription occurs in the nucleus, and the newly synthesized mRNA molecules must be exported to the cytoplasm for translation to occur. The export of mRNA from the nucleus to the cytoplasm involves a complex series of interactions between various transcription factors, such as activators/repressors and pre-mRNA processing factors, with TREX complex, transcription and export 2 (TREX-2) complex, and the NPC being the crucial elements necessary for mRNA transport [31,47,48].
The mechanism of mRNA nuclear export is facilitated by the NPC, which is a large protein complex that spans the double-layered membrane of the nuclear envelope in eukaryotic cells. The NPC acts as a gatekeeper, controlling the selective transport of molecules such as proteins, RNA, and other macromolecules between the nucleus and the cytoplasm. The NPC is a cylindrical channel that spans the nuclear envelope and is made up of numerous copies of around 30 different Nups, which are placed in an octagonal symmetry. The central channel of the NPC is lined with phenylalanine-glycine (FG) repeats, which form a selective barrier that allows only certain molecules to pass through. The NPC is critical for many cellular processes, including gene expression, RNA processing, and protein transport. The NPC also serves as a quality control mechanism for the exported mRNA molecules. The export process is mediated by a group of proteins, including exportins and TREX complex [49,50,51,52].
There are two main pathways for mRNA nuclear export: NXF1–NXT1-dependent and CRM1-dependent pathways. After capping, splicing, and polyadenylation, the pre-mRNA becomes mature mRNA [17,18]. Then, mRNA export factors begin to load on the mRNAs and recruit the TREX complex. In eukaryotic cells, the mRNA nuclear export process is significantly influenced by the TREX complex. The THO complex is responsible for the recognition and binding of mRNA molecules after they are synthesized [23]. In the ATP-bound condition, the UAP56 forms a complex with Aly/REF, leading to an increased affinity of Aly/REF for mRNA [25]. Subsequently, Aly/REF recruitsNXF1 -NXT1 to the mRNA [53]. The NXF1–NXT1 heterodimer is the major mRNA nuclear export receptor for the bulk of cellular poly(A)-RNA. NXF1 is the primary component of this complex, while NXT1 acts as an adaptor protein that mediates the interaction between NXF1 and mRNA. By binding to the phenylalanine-glycine (FG) repeat domains found on NPC proteins (nucleoporins or Nups), such as Nup62 and Nup98, the NXF1–NXT1 heterodimer facilitates the transport of mRNA through the NPC and into the cytoplasm for translation [29,30,31]. Through the interaction with GANP, NXF1 can associate with the TREX-2 complex, which includes GANP, PCID2, DSS1, ENY2, and CETN3. With a stable association with the nuclear pore basket, the TREX-2 complex has been found to play a role in the nuclear export of a subset of cellular mRNAs. GANP could interact with nucleoporin TPR on the nucleo-cytoplasmic side of NPC. Messenger ribonucleoprotein (mRNP) translocation via the NPC is mediated by the binding of NXF1-NXT1 with GANP during mRNA nuclear export [54,55,56].
The CRM1-dependent mRNA nuclear export pathway is another mechanism by which certain mRNAs are exported from the nucleus to the cytoplasm. This pathway is dependent on the activity of CRM1, a member of the karyopherin beta protein family that functions as an export receptor for certain NESs found on cargo proteins, including certain mRNAs, tRNAs, and viral RNAs. In this pathway, CRM1 does not directly bind to mRNA, but requires the assistance of RNA export adaptor proteins that can directly bind to the RNA molecules. These adaptor proteins contain NESs that are recognized by CRM1, which then interacts with RanGTP to facilitate the export of the RNA through the NPC [44,57].
In addition, for the host RNA export, RNA length plays an important role in determining which pathway the RNA takes. McCloskey et al. showed that the heterologous nuclear ribonucleoprotein (hnRNP) C tetramer has the function of measuring RNA length for the classification export of RNA polymerase II transcripts [58]. Dantsuji et al. demonstrated that hnRNP C tetramers play a key role in the identification and measurement of RNA length. The heterotetramer of the heterogeneous nuclear ribonucleoprotein (hnRNP) C1/C2 acts as a molecular ruler to measure the length of the transcripts by selectively binding to the unstructured RNA regions longer than 200 to 300 nucleotides. Thus, the tetramer sorts the transcripts into two RNA categories: mRNA or uridine-rich small nuclear RNA (U snRNA). Further mechanistic studies proved that the hnRNP-C tetramer binds the cap-binding complex (CBC) on mRNA and blocks PHAX (an adaptor protein for splicosomal U snRNA output) recruitment for the classification of RNA polymerase II transcripts, so as to direct these RNAs to the mRNA nuclear export pathway [59]. This discovery of the hnRNP C tetramer for length-specific RNA classification contributes to a deeper understanding of RNA biogenesis and provides potential strategies for future disease treatments.
In conclusion, mRNA nuclear export is a complex and highly regulated process that plays a vital role in gene expression regulation. Understanding the underlying mechanisms of mRNA nuclear export provides important implications for our understanding of cellular function and disease pathology.
3. Viral Infection Inhibits the Nuclear Export of Host mRNA
Viruses utilize the host mRNA nuclear export pathway and prevent the export of host mRNA, which is crucial for the translation of mRNA encoding cellular defense proteins required to generate a proper immune response against the invading pathogen. Thus, the mRNA export pathway is an attractive target for viruses to block the expression of host antiviral genes (Figure 1).
In order to block the expression of host genes, viral proteins can utilize host mRNA nuclear export mechanisms to interfere with the host mRNA nuclear export. Multiple viral proteins can be involved in this process. For instance, non-structural protein 1 (NS1) of influenza A virus (IAV) prevents the interaction between NXF1–NXT1 and the nucleoporins, thereby blocking the export of the host’s mRNA [60]. IAV NS1 also interacts with the mRNA nuclear export factor Rae1 and E1B-AP5 to inhibit host mRNA export [60]. Similarly, the M (matrix) protein of vesicular stomatitis virus (VSV) interacts with Rae1 and NPC (via NUP98) to form M-Rae1-NUP98, blocking host mRNA nuclear export [36,61]. The M proteins of other vesicle viruses, such as the Chandipura virus (CV) and spring viraemia of carp virus (SVCV), also block the export of host mRNA by interacting with Rae1 and NUP98 [62]. In addition, the adenovirus (AdV) protein E1B-55K inhibits the nuclear export of host mRNA by interacting with E1B-AP5. Moreover, E1B-55K is assisted by its E4orf6 protein, which can form [63] part of the E3 ubiquitin ligase with extender proteins B and C, cullin 5, RBX1, and other proteins of the host cell [63]. This ubiquitin ligase targets cellular p53 [64], MRN (MRE11–RAD50–NBS1 complex, involved in DNA double-strand break repair) [65], DNA ligase IV [66], and integrin α3 [67] to alter the function of host cells. Thus, viral proteins impair the export of host nuclear mRNAs through a variety of pathways by interacting with specific mRNA export factors.
In the downregulation of host mRNA nuclear export by viral proteases, viral proteins can also manipulate the host’s nucleo-cytoplasmic transport by degrading nucleoporins. For instance, the 2A protease (2APro) of picornaviruses, such as poliovirus (PV) and human rhinovirus (HRV), causes protein degradation in nucleoporins, including NUP62, NUP98, and NUP153, leading to changes in the NPC structure and inhibiting host mRNA nuclear export [68,69,70,71,72,73,74]. Similarly, HRV’s 3C protease (3CPro) is involved in the proteolytic degradation of NUP153, NUP214, and NUP358, thereby affecting NPC and mRNA export [75]. Unlike these viruses, picornaviruses, such as Theiler’s murine encephalomyelitis virus (TMEV) and encephalomyocarditis virus (EMCV), lack proteolytic activity in 2APro, but they still impact RanGTP’s involvement in CRM1-mediated mRNA export by the viral leader (L) proteins [11,76,77,78]. The interaction between L proteins and RanGTP also affects protein transport, as RanGTP and CRM1 are involved in protein transport.
In addition to targeting host mRNA nuclear export factors and nucleoporins, viruses downregulate host gene expression by destabilizing host mRNA. For example, viruses such as IAV and IBV (influenza B virus) target the 5’ cap of the host’s mRNA for its degradation [79]. The influenza virus’s RNA polymerase (vRdRp: viral RNA-dependent RNA polymerase) has three subunits, PA, PB1, and PB2, and is responsible for this process. PB2’s mRNA cap-binding domain binds to the host mRNA cap, and then PA’s endonuclease cleaves the host RNA at the 10th–13th nucleotide downstream of the cap. Viruses can also stabilize host mRNA to alter its gene expression. For example, Zika virus, an insect-borne RNA-containing flavivirus, produces long non-coding RNA (lncRNA) called sfRNAs, which are 300–400 nucleotides long. The repression of XRN1 (a 5’ to 3’ exoribonuclease) by sfRNA located in the 3‘ UTR leads to a reversible inhibition of enzyme activity, likely due to the slow release of the inhibited enzyme from the RNA substrate. The Zika virus sfRNA interacts with a common set of 21 RNA-binding proteins (such as DEAD-box helicase 6 (DDX6), mRNA uncapped enhancer 3 (EDC3), etc.) that help regulate cellular post-transcriptional processes, including splicing, RNA stability, and translation [80].
3.1. RNA Viruses Inhibit Host mRNA Nuclear Export
The influenza virus is an enveloped negative-strand RNA virus. The viral ribonucleoproteins (vRNPs), assembly by RNA-dependent RNA polymerase (RdRP) and nuclnucleoprotein (NP), is responsible for viral RNA transcription. In contrast to most negative-strand RNA viruses, influenza viruses transcribe their mRNA in the nucleus. Transcription is initiated by the RdRP excising 12–15-nucleotide RNAs from the mRNA of the infected cell in a process called “cap grabbing” [81] and polyadenylation by repeated replication occurs around the 5′ end of the vRNA template’s poly(U) stretch [82]. Two viral transcripts, M and NS, undergo alternative splicing. The nuclear export of various types of mRNAs, including spliced mRNAs (M2 and NS2), intron-containing mRNAs (M1 and NS1), and intron-free mRNAs (HA, NA, NP, PB1, PB2, and PA), is essential for viral replication. Similar to most viral infections, the proliferation of IAV within vertebrate cells gets detected by the innate immune system. After recognition, the innate immune system initiates signal transduction pathways that result in the synthesis of type I interferons (IFNs), a group of antiviral cytokines that stimulate the expression of mRNAs encoding antiviral factors [83], including nucleoporins [68,84]. In order to suppress host anti-viral response, IAVs have developed several strategies, primarily utilizing non-structural protein 1 (NS1), which stays mainly in the nucleus during early stage of infection, to inhibit mRNA processing and export [5]. There are close linkages between the mRNA processing and nuclear export since some proteins interacting with mRNA are involved in both processes, while others are replaced by particular factors [85]. By binding to the spliceosome component or interacting with the splicing factor, such as NS1 binding protein (NS1-BP), NS1 inhibits pre-mRNA splicing [86,87]. NS1 further disrupts the splitting and polyadenylation specific factor (CPSF30) and poly(A) binding protein II (PABII) of the host’s mRNA processing-bound 30 kDa subunit, participating in the binding polyadenylation signal and the extension of the poly(A) chain mRNA, respectively [88,89]. The interaction of NS1 with these proteins inhibits the 3’-terminal processing of host mRNA and helps to inhibit the expression of host genes. However, because viral polymerase complexes synthesize the poly(A) tail on viral mRNAs, NS1-mediated interruption of mRNA processing has no impact on the generation of viral transcripts [82,90]. In addition, by interacting with host mRNA-binding proteins, NS1-BP, and hnRNP K, NS1 facilitates the splicing of viral M1 mRNA fragments [91]. This enables the efficient processing of viral mRNA transcripts before nuclear export occurs. In addition to disrupting the host mRNA processing, IAV further downregulates the expression of host antiviral genes through NS1 interacting with the host mRNA nuclear export mechanism. NS1 directly interacts with the mRNA export factor NXF1–NXT1 (Figure 2A), which forms complexes with Rae1 and E1B-AP5, result in the retention of poly(A) RNA in the nucleus [60,92]. The NS1–NXF1–NXT1 complex is solved by crystallization. In the protein complex, two NS1 molecules bind to two NXF1–NXT1 heterodimers. The two NS1 molecules dimerize through their RNA binding domains. One effector domain binds to the LRR domain of NXF1. The phenylalanine 103 of the other effector domain binds to the NTF2L domain of the neighboring NXF1, which is also the nucleoporin binding domain, leading to the inhibition of NXF1 docking to NPC. Using the reverse genetic method, a mutant influenza virus carrying the NS1 mutations, in which key residues involved in the interaction with NXF1 were mutated, was generated. The NS1 mutant virus, which lost the interaction with NXF1, did not inhibit the host mRNA nuclear export, resulting in the release of an antiviral response. Therefore, the replication of the mutant virus was attenuated [16]. Taken together, these investigations propose that the IAV NS1 employs diverse strategies to impede the intricate and precisely controlled procedures of mRNA processing and export. Notably, NS1-mediated inhibition of mRNA nuclear export results in the retention of mRNAs encoding antiviral genes in the nucleus [93], indicating that the virus potentially disrupts these pathways as a strategy to enhance viral replication and evade host immune responses.
Belonging to the Rhabdoviridae family, Vesicular Stomatitis Virus (VSV) has a single-stranded RNA genome. VSV is known for causing vesicular stomatitis, a viral disease that affects cattle, horses, and pigs, as well as humans. The virus is transmitted through direct contact with infected animals, their saliva, or contaminated objects. Five genes are encoded by the VSV RNA genome, and they are organized in the following sequence: 3’-N-P-M-G-L-5’. The M gene encodes the matrix protein (M), which is necessary for virion assembly and budding. During VSV infection, the cellular mRNA nuclear export can be blocked by the interaction between M and RNA binding protein Rae1, a component of the mRNA export machinery, which forms a complex with nucleoporin Nup98 (Figure 2B) [36,61,94,95]. The interaction between VSV M protein and Rae1 leads to the sequestration of Rae1 in the nucleus, preventing its normal function in mRNA nuclear export. The participation of the VSV M protein in the inhibition of the host mRNA nuclear export has been detected by multiple methods, including poly(A) RNA-FISH, qPCR of nucleo-cytoplasmic fractionation samples, and nuclear export assays. The residue methionine 51 plays a key role in the interaction between M and Rae1. The M51R mutation of M loses its function in the mRNA nuclear export blockage [96]. The M–Rae1–Nup98 complex leading to a blockage of bulk mRNA nuclear export may be due to two reasons: the nuclear export of a subset of mRNAs that encode gene expression regulators that are blocked, or the Rae1-Nup98 complex involved in regulating the NXF1-mediated mRNA nuclear export. As a result, the expression of antiviral factors is downregulated [93]. The inhibition of the host’s mRNA export by VSV enables the virus to transport its limited number of mRNAs into the cytoplasm, preventing competition for the translation machinery with the host mRNAs. Overall, hindering cellular mRNA trafficking by M is one of the most important strategies to enable VSV to regulate host gene expression and promote its own replication [97].
Global public health has been significantly affected by the pandemic caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [98]. The SARS-CoV-2 genome is approximately 30 kb in length and consists of positive-sense single-strand RNA. It contains fourteen open reading frames, which encode a large polyprotein, four structural proteins (spike (S), membrane (M), envelope (E), and nucleocapsid (N)), and nine accessory proteins (ORF3a, ORF3b, ORF6, ORF7a, ORF7b, ORF8, ORF9b, ORF9c, and ORF10) [99]. SARS-CoV-2 hijacks the host cellular machinery to promote genome replication, which occurs in the cytoplasm. To favor replication, SARS-CoV-2 adopts multiple strategies to suppress the host immune and antiviral responses, including inhibiting host mRNA nuclear export. One of the identified mechanisms involves SARS-CoV-2 Nsp1. Nsp1 was shown to interact with the NXF1–NXT1 heterodimer, leading to the retention of cellular mRNAs in the nucleus. An immunoprecipitation assay with a nuclear fraction showed that Nsp1 displaces NXF1 from the NPC. mRNA inhibition by SARS-CoV-2 was reverted by the overexpression of NXF1 in vero-E6 cells, resulting in the reduction of SARS-CoV-2 replication [99]. SARS-CoV-2 may also target the Nup98–Rae1 complex by Orf6 (Figure 2C) to subvert nucleo-cytoplasmic trafficking, including protein import and nuclear export of mRNA transcripts, such as IFN-upregulated genes. The interaction between Orf6 and Nup98-Rae1 is greatly decreased by the M58R mutation [46,100,101]. Several other SARS-CoV-2 proteins may also interact with mRNA nuclear export factors or nucleoporins, indicating their unknown roles in regulating cellular mRNA nuclear export [102]. The underlying mechanisms that SARS-CoV-2 may use to inhibit host mRNA nuclear export need to be further studied.
3.2. DNA Viruses Inhibit Host mRNA Nuclear Export
AdVs, a non-envelope DNA virus, contains around 36 kbp long linear double-stranded DNA genome. They got their name because of the first discovery in adenoid cell cultures. AdVs are widely distributed, and infections in the immunocompetent host are typically mild or asymptomatic and resolve independently. Nonetheless, Advs infection can result in a more severe illness in people with compromised immune systems [103]. The genome of human AdVs is quite dense, with more than 40 genes. The total length of AdV genomes can vary among different serotypes, ranging from approximately 26,000 to 45,000 base pairs [104]. To ensure efficient viral replication, these genes are expressed in an orderly manner that allows the host cell to replicate to the virus without alerting the immune system or inducing cell damage that leads to apoptosis [105]. Early gene 1A (E1A) was the first gene to be expressed, and it promotes the production of other early proteins from E2, E3, and E4 transcription units [106]. Subsequently, the expression profile of Adv genes gradually changes to a major late promoter (MLP) upon infection, leading to the promotion of the expression of late gene products, which is responsible for the assembly of the new viral particle [106]. In general, a variety of mechanisms, including alternative splicing, which leaves specific transcripts partially or entirely unspliced, highly regulate the transcription program of AdV [107]. Additionally, specific viral transcripts are exported to the cytoplasm for expression at the appropriate time [106]. To date, studies on the export mechanism of AdV mRNA have focused on late messengers, and the picture appears to be quite complex. In 1986, the possible function of viral E1B-55k protein on AdV late transcript export was first observed [108]. However, it is believed to play a relatively indirect and upstream role in export, occurring before the initiation of mRNA nuclear export [109]. Instead, E1B-55K is the primary coordinator leading to the viral protein E4orf3, the promyelocytic leukemia (PML) nucleosome, viral transcription, and replicating occurrence [110,111]. During the later stages of infection, a different viral protein known as E4orf6, competes with E4orf3 to trigger the reorientation of E1B-55K towards the nuclear blob, which serves as the storage site for splicing factors, export capacity, and both viral and cellular transcripts [111]. These series of modifications induced by viruses in nuclear structure hinder the maturation and export of cellular mRNAs, while simultaneously facilitating the export of viral transcripts that include introns. [112]. In addition, to efficient nuclear export of viral late mRNA, the ubiquitin-3-ligase complexes formed by E1B-55k, E4orf6, and some cellular components are required [113]. The relevant mechanisms depend on the cellular components necessary to inhibit the degradation of intron-containing transcripts [114]. However, the understanding of the numerous other functions of E1B-55k in regulating AdV gene expression still need further investigation [115].
Kaposi’s sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus 8 (HHV-8), is a nuclear DNA virus belonging to Herpeseviridae. KSHV is a complex virus with a double-stranded DNA genome that encodes for more than 80 genes. The virus has two distinct phases in its life cycle: a latent phase and a lytic phase. During the latent phase, the virus resides within the host cell without causing any harm, while during the lytic phase, the virus replicates and produces infectious particles. KSHV encodes several viral proteins that can modulate the host cell’s immune response [106]. The ORF10 gene, which can produce ORF10 protein, is located in the viral genome downstream of the viral interleukin-6 (vIL-6) gene and is expressed during the late phase of the viral life cycle, when viral particles are being assembled. Several studies with RNA-FISH and immunoprecipitation methods have suggested that ORF10 may play a role in immune evasion by the virus, including binding to the Rae1–Nup98 complex to selectively inhibit cellular mRNA nuclear export (Figure 2D). By solving the crystal structure of the ORF10–Rae1–Nup98 complex, Feng et al. showed that L60 and M413, two highly conserved residues, are critical for both the assembly of the Orf10-Rae1-Nup98 complex and the suppression of mRNA nuclear export. In vitro binding assays showed that although ORF10 occupies the RNA binding surface of Rae1–Nup98, the ORF10–Rae1–Nup98 complex still binds RNA through ORF10. When mutations are introduced on ORF10’s RNA-binding surface, the protein’s ability to inhibit mRNA export is disrupted. The ability of KSHV to selectively inhibit RNA export can limit host gene expression and optimize the facilitation of replication [116,117].
In sum, during virus infection, a significant strategy utilized by viruses is to trap host mRNA in the nucleus. This results in a downregulation of the host’s gene expression and the inhibition of antiviral responses, which allows the virus to evade host immune responses and establish a successful infection. Various viruses use distinct mechanisms to inhibit host mRNA nuclear export (as summarized in Table 1). Gaining a better understanding of the molecular mechanisms involved in this process could provide valuable insights for the development of novel antiviral therapies.
4. The Nuclear Export of Viral mRNA
The nuclear export of viral mRNA refers to the process by which viral mRNA is released from the nucleus. This process is the first step that viruses take to infect cells, and it can be achieved through the action of viral proteins. The viral protein will bind to the viral mRNA and release it from the nucleus so that the viral mRNA can enter the cytoplasm, where it can be transcribed and translated to produce new viral proteins.
Viruses usurp or hijack host mRNA nuclear export mechanisms to transport their own RNA for viral replication and assembly and block host mRNA export, thereby reducing the expression of host genes, including genes involved in immune responses (i.e., interferon (IFN), inducer genes, and NF-kb regulatory genes [123,124,125]) for successful viral infection and reproduction. By promoting its mRNA export, a virus can also deceive the host immune system by expressing genes that encode their own cytokines, molecules that neutralize host cytokines and homologues of TNF (tumor necrosis factor) or MHC (major histocompatibility complex) classes, acting as decoys for NK (natural killer) cells [123,124,125,126,127]. In addition, virus-encoded lncRNA (long non-coding RNA) has also been implicated in antiviral responses against IFN [128,129,130,131,132,133]. For example, the PANRNA (polyadenylated nuclear RNA) of KSHV (Kaposi sarcoma-associated herpes virus) reduces IFNα and IFNγ expression [128,129]. sfRNAs (subgenomic flavivirus RNA) of several viruses, including JEV (Japanese encephalitis virus), Dengue virus, and WNV (West Nile virus), inhibit IFNβ expression and IFN signaling, thereby impairing the host’s antiviral response [128,129,130,131,132,133]. In addition, by enhancing its RNA export and producing molecules that inhibit caspase (a major player in apoptosis), viruses can also block host cell apoptosis strategies to get rid of virus-infected cells [96]. Thus, viruses hijack the host’s mRNA export mechanism to help induce a successful infection and subsequent reproduction [123]. This hijacking is mediated through the interaction of viral proteins with host mRNA nuclear export factors.
4.1. The Virus Targets the Host mRNA Export Receptor CRM1
The nuclear export receptor CRM1 recognizes cargo proteins containing a leucine-rich NES. During virus infection, some viruses hijack the CRM1-dependent mRNA export pathway to export their own viral mRNA molecules out of the nucleus and into the cytoplasm, where they can be translated into viral proteins. Viruses can achieve this process by encoding NES-containing proteins, which are recognized by CRM1. The CRM1-mediated nuclear export pathway is a critical step in the replication cycle of many viruses, including human immunodeficiency virus 1 (HIV-1), prototype foamy virus (HFV), human T-cell leukemia virus type 1 (HTLV-1), and human papillomavirus (HPV) (Figure 3A). Targeting this pathway with specific inhibitors has been shown to inhibit viral replication, and could be a potential therapeutic strategy for viral infections.
HIV-1 and other lentiviruses use the host mRNA export receptor CRM1 to export their RNA into the cytoplasm. The major HIV-1-encoding proteins are Gag, Pol, Env, Tat, Rev, and Nef [134]. The genome replication of HIV-1 occurs in the nucleus. Thus, the HIV-1 mRNA should be exported to the cytoplasm for translation. The Rev protein contains a leucine-rich NES domain, which is recognized by CRM1. In addition, the Rev protein interacts with the RRE (Rev response element, a highly structured RNA), which is often unspliced. The Rev also acts as an arginine-rich NLS (nuclear location signal) [10,135,136,137]. This viral RNA Rev-CRM1 interaction network exports viral RNA into the cytoplasm with the help of RanGTP. Thus, viral RNA-bound Revs hijack host CRM1 to export viral RNA via NPCS [10,135,136,137]. Recent work has suggested that other NES-containing proteins, such as RNA binding protein 14 (RBM14) [138] and Phosphofurin acidic cluster-sorting protein 1 (PACS1) [139], could serve as Rev co-factors and play a crucial role in the nuclear export of viral RNAs mediated by Revs. In addition, several cellular factors have also been identified to regulate the nuclear export of HIV-1 mRNA, including the mRNA export adaptor protein Aly/REF and the RNA helicase DDX3 [140]. Further understanding of the molecular mechanisms underlying HIV-1 mRNA nuclear export could provide new insights into the development of therapies for HIV-1 infection.
Host CRM1 is also used to export viral RNA via the Rex protein of Delta retrovirus HTLV-1, which causes T-cell leukemia. Similar to the Rev protein, Rex binds to the RxRE (Rex response element) on unspliced and spliced viral RNA and interacts with CRM1 to export viral RNA into the cytoplasm with the help of RanGTP [140,141]. Similarly, the retrovirus MMTV (mouse mammary tumor virus) encodes the Rem protein, which interacts with viral RNA and CRM1 to export viral RNA to the cytoplasm [141,142,143]. Interestingly, the host adapter of CRM1, HuR, has also been used for the export of viral RNA by foamy viruses [144]. CRM1 is also involved in exporting IAV RNA by interacting with viral NP (with viral RNA complex) in the presence of RanGTP [120,145,146]. Thus, CRM1 is targeted by viral proteins for its advantages (i.e., the nuclear export of its RNA). Since CRM1 is associated with the nuclear export of mRNA, rRNA, and snRNA, viruses that use CRM1 to help export their own RNA to the cytoplasm can inhibit the export of host RNA and thus downregulate the expression of host genes. Viruses can deceive the host’s immune system by enhancing viral mRNA nuclear export to promote their cytokine gene expression. In addition, viruses can promote the export of viral mRNAs that encode inhibitors of host caspases to block cell apoptosis. Thus, viruses hijack the host’s mRNA nuclear export mechanism to help them trigger a successful infection and its subsequent spread [147].
4.2. The Virus Targets the Host mRNA Export Receptor NXF1
Similar to CRM1, the mRNA export receptor NXF1 is also a target for viral proteins exported by viral RNA (Figure 3B). NXF1 binds to CTEs (constitutive transporter elements). The NXF1-interacting protein NXT1 [148] further enhances this binding. Thus, unlike host mRNA, which requires the adapter protein to interact with NXF1, viral RNA interacts directly with NXF1 and is transported to the cytoplasm, thus having an advantage over the host system. The CTEs, which are located in the stem-loop RNA, are responsible for the nuclear export of the unspliced or incompletely spliced RNA of Mason-Pfizer monkey virus (MPMV) or type D retrovirus [14,149,150]. Similarly, other retroviruses, such as avian sarcoma and the leukemia virus, have used CTEs to export unspliced viral RNA [149,151]. Gamma retroviruses, including murine leukemia virus (MLV) and heterophilic murine leukemia virus-associated virus (XMRV), also use NXF1 for viral RNA export [152]. The nuclear export of MLV unspliced RNA is co-regulated by NXF1 and CRM1 pathway [153]. This export is mediated through NXF1’s interaction with the CAE (cytoplasmic accumulation element) of viral RNA [154]. Certain IAV mRNAs (e.g., viral mRNAs encoding M1 matrix proteins and M2 ion channels) also depend on NXF1 for export. The viral NS1 protein acts as an adapter (interacting with viral RNA and NXF1) for the nuclear export of viral RNA [16]. Using the auxin-induced deoxyribonucleic acid (AID) system, a recent study by Bhat et al. demonstrated that the nuclear export of influenza virus mRNAs is dependent on the TREX-2 complex. By labeling different components in mRNA and TREX-2 complex or targeting specific genetic components, they successfully revealed the interaction between the extranuclear transport process of influenza virus mRNA and the TREX-2 complex. Further RNA sequencing techniques have demonstrated the importance of this mode of transport in influenza virus extrinsic replication and infection [155]. The NP (which binds to viral RNA) of Ebola virus (EBOV, an RNA virus) also interacts with NXF1 to export viral RNA into the cytoplasm [118,156,157]. The infected cell protein 27 (ICP27) (which binds to viral RNA) of the herpes simplex virus 1 (HSV-1) interacts with NXF1 to achieve viral RNA export [118,158]. Similarly, the herpesvirus saimiri (HVS) TIP protein also interacts with NXF1 [156]. NXF1, therefore, is primarily a viral protein targeting viral RNA export. Since NXF1 is involved in the nuclear export of a large amount of mRNA in the host body, as described above, the viral hijacking of NXF1 can severely block the export of the host mRNA and thus block the expression of the host gene. AdV early protein E1B-55K, along with its cellular partner, E1B-55K associated protein 5 (E1B-AP5), interacts with viral mRNAs and facilitates the recruitment of NXF1–NXT1, which in turn stimulates the nuclear export of mRNP through the TREX-2–NPC interaction [157,159]. Hepatitis B virus (HBV), a member of the Hepadnaviridae family, can cause liver disease. The genetic material of HBV is composed of partially double-stranded DNA. The transcripts produced by HBV can be categorized into three distinct groups based on their splicing status and genomic location: unspliced pg (pregenomic) and preC (precore) mRNAs, spliced pg and preC mRNAs, and intronless subgenomic mRNAs [160]. The post-transcriptional regulatory element (PRE), a cis-acting RNA element that can be divided into two sub-elements, PRE 1 and 2 (SEP1 and SEP2), is present in all transcripts of HBV [161]. The zinc finger CCCH domain-containing protein 18 (ZC3H18) has been identified as binding SEP1 and recruiting components of the TREX complex for SEP1 nuclear export through the NXF1–NXT1 pathway [162]. TAR DNA binding protein (TARDBP) has been suggested to play a key role in the export of unspliced pgRNA isoforms [163]. The HBV X protein interacts with UAP56, which is potentially functional in the nuclear export of single-exon HBV mRNAs [164]. With a better understanding of the nuclear export mechanisms utilized by HBV mRNA, it may be possible to create novel targeted antiviral therapies that are required for the management of Hepatitis B.
To summarize, viruses have developed diverse strategies to manipulate the cellular pathways of mRNA nuclear export. The mechanisms utilized by viruses to facilitate their mRNA nuclear export are extremely variable, even within the same family (Table 2). Current and forthcoming research endeavors are expected to provide a comprehensive understanding of the molecular mechanisms of viruses’s mRNA nuclear export, which will provide novel anti-viral therapeutic strategies by targeting virus mRNA nuclear export.
5. Conclusions and Future Perspectives
To prevent the expression of mRNA encoding antiviral factors and to favor the competition with host mRNA in translation mechanisms, various research groups have identified ways by which distinct viruses impede the nuclear export of cellular mRNA. Overall, this leads to the inhibition of host protein translation. In addition, viruses have significantly affected our understanding of the nuclear export of cellular mRNA and the key roles involved.
The significance of these pathways in both proviral and antiviral mechanisms is underscored by these findings. These studies suggest that the virus invests many resources to disrupt nucleo-cytoplasmic transport, and that the host can antagonize these effects. Therefore, the nuclear import/export pathway is an attractive target for the development of novel antiviral therapeutics. An anti-IAV compound screening by targeting the nuclear export of IAV M mRNA has been reported by Esparza et al [172]. To effectively nuclear export of the viral mRNA, viruses utilize multiple and complex strategies to hijack cellular mRNA nuclear export machinary. To date, the mechanisms about these strategies still remain incomplete discovered, sparking considerable interest among researchers. Gaining a deeper understanding on the mRNA nuclear export of various types of viruses will offer new insights into both the virus replication process and our knowledge about cellular mRNA nuclear export mechanisms.
Author Contributions
Writing—original draft preparation: J.G., Y.Z., X.M., G.S., B.L. and K.Z.; J.G. and Y.Z. wrote the paper; J.G., X.M. and G.S. made the figures; J.G. and Y.Z. made the tables; J.G., B.L. and K.Z made significant contributions in the revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Shanghai Pujiang Program 22PJ1414600 (to K.Z.), the Shanghai Pujiang Program 21PJ1414500 (to B.L.), and the Shanghai Municipal Science and Technology Major Project HS2021SHZX001 (to B.L.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Callan, H.G.; Tomlin, S.G. Experimental studies on amphibian oocyte nuclei. I. Investigation of the structure of the nuclear membrane by means of the electron microscope. Proc. R Soc. Lond. B Biol. Sci. 1950, 137, 367–378. [Google Scholar]
- Gall, J.G. Observations on the nuclear membrane with the electron microscope. Exp. Cell Res. 1954, 7, 197–200. [Google Scholar] [CrossRef]
- Lin, D.H.; Hoelz, A. The Structure of the Nuclear Pore Complex (An Update). Annu. Rev. Biochem. 2019, 88, 725–783. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W. Gene Regulation and Cellular Metabolism: An Essential Partnership. Trends Genet. 2021, 37, 389–400. [Google Scholar] [CrossRef]
- Kuss, S.K.; Mata, M.A.; Zhang, L.; Fontoura, B.M. Nuclear imprisonment: Viral strategies to arrest host mRNA nuclear export. Viruses 2013, 5, 1824–1849. [Google Scholar] [CrossRef] [Green Version]
- Baum, A.; García-Sastre, A. Induction of type I interferon by RNA viruses: Cellular receptors and their substrates. Amino Acids 2010, 38, 1283–1299. [Google Scholar] [CrossRef] [Green Version]
- Finlay, B.B.; McFadden, G. Anti-immunology: Evasion of the host immune system by bacterial and viral pathogens. Cell 2006, 124, 767–782. [Google Scholar] [CrossRef] [Green Version]
- Fensterl, V.; Sen, G.C. Interferons and viral infections. Biofactors 2009, 35, 14–20. [Google Scholar] [CrossRef]
- Dimmock, N.J.; Easton, A.J.; Leppard, K.N. Introduction to Modern Virology, 7th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Fontoura, B.M.; Faria, P.A.; Nussenzveig, D.R. Viral interactions with the nuclear transport machinery: Discovering and disrupting pathways. IUBMB Life 2005, 57, 65–72. [Google Scholar] [CrossRef]
- Knipe, D.M.; Howley, P.M. Virus Replication Strategies, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PE, USA, 2007. [Google Scholar]
- Fornerod, M.; Ohno, M.; Yoshida, M.; Mattaj, I.W. CRM1 is an export receptor for leucine-rich nuclear export signals. Cell 1997, 90, 1051–1060. [Google Scholar] [CrossRef] [Green Version]
- Stade, K.; Ford, C.S.; Guthrie, C.; Weis, K. Exportin 1 (Crm1p) is an essential nuclear export factor. Cell 1997, 90, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Grüter, P.; Tabernero, C.; von Kobbe, C.; Schmitt, C.; Saavedra, C.; Bachi, A.; Wilm, M.; Felber, B.K.; Izaurralde, E. TAP, the human homolog of Mex67p, mediates CTE-dependent RNA export from the nucleus. Mol. Cell 1998, 1, 649–659. [Google Scholar] [CrossRef]
- Yang, C.C.; Chang, C.H.; Chen, H.L.; Chou, M.C.; Yang, C.J.; Jhou, R.S.; Huang, E.Y.; Li, H.C.; Suen, C.S.; Hwang, M.J.; et al. CRM1-spike-mediated nuclear export of hepatitis B virus encapsidated viral RNA. Cell Rep. 2022, 38, 110472. [Google Scholar] [CrossRef]
- Zhang, K.; Xie, Y.; Muñoz-Moreno, R.; Wang, J.; Zhang, L.; Esparza, M.; García-Sastre, A.; Fontoura, B.M.A.; Ren, Y. Structural basis for influenza virus NS1 protein block of mRNA nuclear export. Nat. Microbiol. 2019, 4, 1671–1679. [Google Scholar] [CrossRef]
- Shatkin, A.J.; Manley, J.L. The ends of the affair: Capping and polyadenylation. Nat. Struct. Biol. 2000, 7, 838–842. [Google Scholar] [CrossRef]
- Cheng, H.; Dufu, K.; Lee, C.S.; Hsu, J.L.; Dias, A.; Reed, R. Human mRNA export machinery recruited to the 5′ end of mRNA. Cell 2006, 127, 1389–1400. [Google Scholar] [CrossRef] [Green Version]
- Strässer, K.; Masuda, S.; Mason, P.; Pfannstiel, J.; Oppizzi, M.; Rodriguez-Navarro, S.; Rondón, A.G.; Aguilera, A.; Struhl, K.; Reed, R.; et al. TREX is a conserved complex coupling transcription with messenger RNA export. Nature 2002, 417, 304–308. [Google Scholar] [CrossRef]
- Masuda, S.; Das, R.; Cheng, H.; Hurt, E.; Dorman, N.; Reed, R. Recruitment of the human TREX complex to mRNA during splicing. Genes Dev. 2005, 19, 1512–1517. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Luo, M.J.; Straesser, K.; Katahira, J.; Hurt, E.; Reed, R. The protein Aly links pre-messenger-RNA splicing to nuclear export in metazoans. Nature 2000, 407, 401–405. [Google Scholar] [CrossRef]
- Chávez, S.; Beilharz, T.; Rondón, A.G.; Erdjument-Bromage, H.; Tempst, P.; Svejstrup, J.Q.; Lithgow, T.; Aguilera, A. A protein complex containing Tho2, Hpr1, Mft1 and a novel protein, Thp2, connects transcription elongation with mitotic recombination in Saccharomyces cerevisiae. Embo. J. 2000, 19, 5824–5834. [Google Scholar] [CrossRef] [Green Version]
- Rehwinkel, J.; Herold, A.; Gari, K.; Köcher, T.; Rode, M.; Ciccarelli, F.L.; Wilm, M.; Izaurralde, E. Genome-wide analysis of mRNAs regulated by the THO complex in Drosophila melanogaster. Nat. Struct. Mol. Biol. 2004, 11, 558–566. [Google Scholar] [CrossRef]
- Piruat, J.I.; Aguilera, A. A novel yeast gene, THO2, is involved in RNA pol II transcription and provides new evidence for transcriptional elongation-associated recombination. Embo. J. 1998, 17, 4859–4872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.L.; Zhou, Z.; Magni, K.; Christoforides, C.; Rappsilber, J.; Mann, M.; Reed, R. Pre-mRNA splicing and mRNA export linked by direct interactions between UAP56 and Aly. Nature 2001, 413, 644–647. [Google Scholar] [CrossRef]
- Dufu, K.; Livingstone, M.J.; Seebacher, J.; Gygi, S.P.; Wilson, S.A.; Reed, R. ATP is required for interactions between UAP56 and two conserved mRNA export proteins, Aly and CIP29, to assemble the TREX complex. Genes Dev. 2010, 24, 2043–2053. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, I.; Ohno, M. ATP-dependent recruitment of export factor Aly/REF onto intronless mRNAs by RNA helicase UAP56. Mol. Cell Biol. 2008, 28, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Stutz, F.; Bachi, A.; Doerks, T.; Braun, I.C.; Séraphin, B.; Wilm, M.; Bork, P.; Izaurralde, E. REF, an evolutionary conserved family of hnRNP-like proteins, interacts with TAP/Mex67p and participates in mRNA nuclear export. RNA 2000, 6, 638–650. [Google Scholar] [CrossRef]
- Strässer, K.; Hurt, E. Yra1p, a conserved nuclear RNA-binding protein, interacts directly with Mex67p and is required for mRNA export. Embo. J. 2000, 19, 410–420. [Google Scholar]
- Bachi, A.; Braun, I.C.; Rodrigues, J.P.; Panté, N.; Ribbeck, K.; von Kobbe, C.; Kutay, U.; Wilm, M.; Görlich, D.; Carmo-Fonseca, M.; et al. The C-terminal domain of TAP interacts with the nuclear pore complex and promotes export of specific CTE-bearing RNA substrates. RNA 2000, 6, 136–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmody, S.R.; Wente, S.R. mRNA nuclear export at a glance. J. Cell Sci. 2009, 122 Pt 12, 1933–1937. [Google Scholar] [CrossRef] [Green Version]
- Bailer, S.M.; Siniossoglou, S.; Podtelejnikov, A.; Hellwig, A.; Mann, M.; Hurt, E. Nup116p and nup100p are interchangeable through a conserved motif which constitutes a docking site for the mRNA transport factor gle2p. Embo. J. 1998, 17, 1107–1119. [Google Scholar] [CrossRef]
- Bharathi, A.; Ghosh, A.; Whalen, W.A.; Yoon, J.H.; Pu, R.; Dasso, M.; Dhar, R. The human RAE1 gene is a functional homologue of Schizosaccharomyces pombe rae1 gene involved in nuclear export of Poly(A) + RNA. Gene 1997, 198, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Blevins, M.B.; Smith, A.M.; Phillips, E.M.; Powers, M.A. Complex formation among the RNA export proteins Nup98, Rae1/Gle2, and TAP. J. Biol. Chem. 2003, 278, 20979–20988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.A.; Bharathi, A.; Ghosh, A.; Whalen, W.; Fitzgerald, E.; Dhar, R. A mutation in the Schizosaccharomyces pombe rae1 gene causes defects in poly(A)+ RNA export and in the cytoskeleton. J. Biol. Chem. 1995, 270, 7411–7419. [Google Scholar] [CrossRef] [Green Version]
- Faria, P.A.; Chakraborty, P.; Levay, A.; Barber, G.N.; Ezelle, H.J.; Enninga, J.; Arana, C.; van Deursen, J.; Fontoura, B.M. VSV disrupts the Rae1/mrnp41 mRNA nuclear export pathway. Mol. Cell 2005, 17, 93–102. [Google Scholar] [CrossRef]
- Kraemer, D.; Blobel, G. mRNA binding protein mrnp 41 localizes to both nucleus and cytoplasm. Proc. Natl. Acad. Sci. USA 1997, 94, 9119–9124. [Google Scholar] [CrossRef]
- Pritchard, C.E.; Fornerod, M.; Kasper, L.H.; van Deursen, J.M. RAE1 is a shuttling mRNA export factor that binds to a GLEBS-like NUP98 motif at the nuclear pore complex through multiple domains. J. Cell Biol. 1999, 145, 237–254. [Google Scholar] [CrossRef]
- Ren, Y.; Seo, H.S.; Blobel, G.; Hoelz, A. Structural and functional analysis of the interaction between the nucleoporin Nup98 and the mRNA export factor Rae1. Proc. Natl. Acad. Sci. USA 2010, 107, 10406–10411. [Google Scholar] [CrossRef]
- Whalen, W.A.; Bharathi, A.; Danielewicz, D.; Dhar, R. Advancement through mitosis requires rae1 gene function in fission yeast. Yeast 1997, 13, 1167–1179. [Google Scholar] [CrossRef]
- Tsai, N.P.; Lin, Y.L.; Tsui, Y.C.; Wei, L.N. Dual action of epidermal growth factor: Extracellular signal-stimulated nuclear-cytoplasmic export and coordinated translation of selected messenger RNA. J. Cell Biol. 2010, 188, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Bogerd, H.P.; Wang, P.J.; Page, D.C.; Cullen, B.R. Two closely related human nuclear export factors utilize entirely distinct export pathways. Mol. Cell 2001, 8, 397–406. [Google Scholar] [CrossRef]
- Culjkovic, B.; Topisirovic, I.; Skrabanek, L.; Ruiz-Gutierrez, M.; Borden, K.L. eIF4E is a central node of an RNA regulon that governs cellular proliferation. J. Cell Biol. 2006, 175, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Natalizio, B.J.; Wente, S.R. Postage for the messenger: Designating routes for nuclear mRNA export. Trends Cell Biol. 2013, 23, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Shang, L.; Zhang, J.; Liu, Y.; Jin, C.; Zhao, Y.; Lei, X.; Wang, W.; Xiao, X.; Zhang, X.; et al. An antibody-based proximity labeling map reveals mechanisms of SARS-CoV-2 inhibition of antiviral immunity. Cell Chem. Biol. 2022, 29, 5–18.e6. [Google Scholar] [CrossRef] [PubMed]
- Miorin, L.; Kehrer, T.; Sanchez-Aparicio, M.T.; Zhang, K.; Cohen, P.; Patel, R.S.; Cupic, A.; Makio, T.; Mei, M.; Moreno, E.; et al. SARS-CoV-2 Orf6 hijacks Nup98 to block STAT nuclear import and antagonize interferon signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 28344–28354. [Google Scholar] [CrossRef]
- Stewart, M. Nuclear export of mRNA. Trends Biochem. Sci. 2010, 35, 609–617. [Google Scholar] [CrossRef]
- Katahira, J. Nuclear export of messenger RNA. Genes 2015, 6, 163–184. [Google Scholar] [CrossRef]
- Strambio-De-Castillia, C.; Niepel, M.; Rout, M.P. The nuclear pore complex: Bridging nuclear transport and gene regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 490–501. [Google Scholar] [CrossRef]
- Dultz, E.; Wojtynek, M.; Medalia, O.; Onischenko, E. The Nuclear Pore Complex: Birth, Life, and Death of a Cellular Behemoth. Cells 2022, 11, 1456. [Google Scholar] [CrossRef]
- Bley, C.J.; Nie, S.; Mobbs, G.W.; Petrovic, S.; Gres, A.T.; Liu, X.; Mukherjee, S.; Harvey, S.; Huber, F.M.; Lin, D.H.; et al. Architecture of the cytoplasmic face of the nuclear pore. Science 2022, 376, eabm9129. [Google Scholar] [CrossRef]
- Petrovic, S.; Samanta, D.; Perriches, T.; Bley, C.J.; Thierbach, K.; Brown, B.; Nie, S.; Mobbs, G.W.; Stevens, T.A.; Liu, X.; et al. Architecture of the linker-scaffold in the nuclear pore. Science 2022, 376, eabm9798. [Google Scholar] [CrossRef]
- Hautbergue, G.M.; Hung, M.L.; Golovanov, A.P.; Lian, L.Y.; Wilson, S.A. Mutually exclusive interactions drive handover of mRNA from export adaptors to TAP. Proc. Natl. Acad. Sci. USA 2008, 105, 5154–5159. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, V.O.; McMurtrie, P.I.; Mills, A.D.; Takei, Y.; Penrhyn-Lowe, S.; Amagase, Y.; Main, S.; Marr, J.; Stewart, M.; Laskey, R.A. mRNA export from mammalian cell nuclei is dependent on GANP. Curr. Biol. 2010, 20, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, V.O.; Andrews, R.; Ellis, P.; Langford, C.; Gurdon, J.B.; Stewart, M.; Venkitaraman, A.R.; Laskey, R.A. Selective nuclear export of specific classes of mRNA from mammalian nuclei is promoted by GANP. Nucleic Acids Res. 2014, 42, 5059–5071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksenova, V.; Smith, A.; Lee, H.; Bhat, P.; Esnault, C.; Chen, S.; Iben, J.; Kaufhold, R.; Yau, K.C.; Echeverria, C.; et al. Nucleoporin TPR is an integral component of the TREX-2 mRNA export pathway. Nat. Commun. 2020, 11, 4577. [Google Scholar] [CrossRef]
- Culjkovic-Kraljacic, B.; Borden, K.L. Aiding and abetting cancer: mRNA export and the nuclear pore. Trends Cell Biol. 2013, 23, 328–335. [Google Scholar] [CrossRef] [Green Version]
- McCloskey, A.; Taniguchi, I.; Shinmyozu, K.; Ohno, M. hnRNP C tetramer measures RNA length to classify RNA polymerase II transcripts for export. Science 2012, 335, 1643–1646. [Google Scholar] [CrossRef]
- Dantsuji, S.; Ohno, M.; Taniguchi, I. The hnRNP C tetramer binds to CBC on mRNA and impedes PHAX recruitment for the classification of RNA polymerase II transcripts. Nucleic Acids Res. 2023, 51, 1393–1408. [Google Scholar] [CrossRef]
- Satterly, N.; Tsai, P.L.; van Deursen, J.; Nussenzveig, D.R.; Wang, Y.; Faria, P.A.; Levay, A.; Levy, D.E.; Fontoura, B.M. Influenza virus targets the mRNA export machinery and the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2007, 104, 1853–1858. [Google Scholar] [CrossRef]
- von Kobbe, C.; van Deursen, J.M.; Rodrigues, J.P.; Sitterlin, D.; Bachi, A.; Wu, X.; Wilm, M.; Carmo-Fonseca, M.; Izaurralde, E. Vesicular stomatitis virus matrix protein inhibits host cell gene expression by targeting the nucleoporin Nup98. Mol. Cell 2000, 6, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.M.; Her, L.S.; Dahlberg, J.E. Multiple vesiculoviral matrix proteins inhibit both nuclear export and import. Proc. Natl. Acad. Sci. USA 2001, 98, 8590–8595. [Google Scholar] [CrossRef]
- Harada, J.N.; Shevchenko, A.; Shevchenko, A.; Pallas, D.C.; Berk, A.J. Analysis of the adenovirus E1B-55K-anchored proteome reveals its link to ubiquitination machinery. J. Virol. 2002, 76, 9194–9206. [Google Scholar] [CrossRef] [Green Version]
- Querido, E.; Blanchette, P.; Yan, Q.; Kamura, T.; Morrison, M.; Boivin, D.; Kaelin, W.G.; Conaway, R.C.; Conaway, J.W.; Branton, P.E. Degradation of p53 by adenovirus E4orf6 and E1B55K proteins occurs via a novel mechanism involving a Cullin-containing complex. Genes Dev. 2001, 15, 3104–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracker, T.H.; Carson, C.T.; Weitzman, M.D. Adenovirus oncoproteins inactivate the Mre11-Rad50-NBS1 DNA repair complex. Nature 2002, 418, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Rohleder, K.J.; Hanakahi, L.A.; Ketner, G. Adenovirus E4 34k and E1b 55k oncoproteins target host DNA ligase IV for proteasomal degradation. J. Virol. 2007, 81, 7034–7040. [Google Scholar] [CrossRef] [Green Version]
- Dallaire, F.; Blanchette, P.; Groitl, P.; Dobner, T.; Branton, P.E. Identification of integrin alpha3 as a new substrate of the adenovirus E4orf6/E1B 55-kilodalton E3 ubiquitin ligase complex. J. Virol. 2009, 83, 5329–5338. [Google Scholar] [CrossRef] [Green Version]
- Castelló, A.; Izquierdo, J.M.; Welnowska, E.; Carrasco, L. RNA nuclear export is blocked by poliovirus 2A protease and is concomitant with nucleoporin cleavage. J. Cell Sci. 2009, 122 Pt 20, 3799–3809. [Google Scholar] [CrossRef] [Green Version]
- Gustin, K.E.; Sarnow, P. Effects of poliovirus infection on nucleo-cytoplasmic trafficking and nuclear pore complex composition. Embo. J. 2001, 20, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Gustin, K.E.; Sarnow, P. Inhibition of nuclear import and alteration of nuclear pore complex composition by rhinovirus. J. Virol. 2002, 76, 8787–8796. [Google Scholar] [CrossRef] [Green Version]
- Belov, G.A.; Lidsky, P.V.; Mikitas, O.V.; Egger, D.; Lukyanov, K.A.; Bienz, K.; Agol, V.I. Bidirectional increase in permeability of nuclear envelope upon poliovirus infection and accompanying alterations of nuclear pores. J. Virol. 2004, 78, 10166–10177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, N.; Katikaneni, P.; Skern, T.; Gustin, K.E. Differential targeting of nuclear pore complex proteins in poliovirus-infected cells. J. Virol. 2008, 82, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Park, N.; Skern, T.; Gustin, K.E. Specific cleavage of the nuclear pore complex protein Nup62 by a viral protease. J. Biol. Chem. 2010, 285, 28796–28805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watters, K.; Palmenberg, A.C. Differential processing of nuclear pore complex proteins by rhinovirus 2A proteases from different species and serotypes. J. Virol. 2011, 85, 10874–10883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghildyal, R.; Jordan, B.; Li, D.; Dagher, H.; Bardin, P.G.; Gern, J.E.; Jans, D.A. Rhinovirus 3C protease can localize in the nucleus and alter active and passive nucleocytoplasmic transport. J. Virol. 2009, 83, 7349–7352. [Google Scholar] [CrossRef] [Green Version]
- Ricour, C.; Borghese, F.; Sorgeloos, F.; Hato, S.V.; van Kuppeveld, F.J.; Michiels, T. Random mutagenesis defines a domain of Theiler’s virus leader protein that is essential for antagonism of nucleocytoplasmic trafficking and cytokine gene expression. J. Virol. 2009, 83, 11223–11232. [Google Scholar] [CrossRef] [Green Version]
- Ricour, C.; Delhaye, S.; Hato, S.V.; Olenyik, T.D.; Michel, B.; van Kuppeveld, F.J.; Gustin, K.E.; Michiels, T. Inhibition of mRNA export and dimerization of interferon regulatory factor 3 by Theiler’s virus leader protein. J. Gen. Virol. 2009, 90 Pt 1, 177–186. [Google Scholar] [CrossRef]
- Porter, F.W.; Bochkov, Y.A.; Albee, A.J.; Wiese, C.; Palmenberg, A.C. A picornavirus protein interacts with Ran-GTPase and disrupts nucleocytoplasmic transport. Proc. Natl. Acad. Sci. USA 2006, 103, 12417–12422. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.W.; Crépin, T.; Hart, D.J.; Cusack, S. Towards an atomic resolution understanding of the influenza virus replication machinery. Curr. Opin. Struct. Biol. 2010, 20, 104–113. [Google Scholar] [CrossRef]
- Michalski, D.; Ontiveros, J.G.; Russo, J.; Charley, P.A.; Anderson, J.R.; Heck, A.M.; Geiss, B.J.; Wilusz, J. Zika virus noncoding sfRNAs sequester multiple host-derived RNA-binding proteins and modulate mRNA decay and splicing during infection. J. Biol. Chem. 2019, 294, 16282–16296. [Google Scholar] [CrossRef]
- Plotch, S.J.; Bouloy, M.; Ulmanen, I.; Krug, R.M. A unique cap(m7GpppXm)-dependent influenza virion endonuclease cleaves capped RNAs to generate the primers that initiate viral RNA transcription. Cell 1981, 23, 847–858. [Google Scholar] [CrossRef]
- Poon, L.L.; Pritlove, D.C.; Fodor, E.; Brownlee, G.G. Direct evidence that the poly(A) tail of influenza A virus mRNA is synthesized by reiterative copying of a U track in the virion RNA template. J. Virol. 1999, 73, 3473–3476. [Google Scholar] [CrossRef]
- García-Sastre, A. Induction and evasion of type I interferon responses by influenza viruses. Virus Res. 2011, 162, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enninga, J.; Levy, D.E.; Blobel, G.; Fontoura, B.M. Role of nucleoporin induction in releasing an mRNA nuclear export block. Science 2002, 295, 1523–1525. [Google Scholar] [CrossRef] [PubMed]
- Luna, R.; Gaillard, H.; González-Aguilera, C.; Aguilera, A. Biogenesis of mRNPs: Integrating different processes in the eukaryotic nucleus. Chromosoma 2008, 117, 319–331. [Google Scholar] [CrossRef]
- Qiu, Y.; Nemeroff, M.; Krug, R.M. The influenza virus NS1 protein binds to a specific region in human U6 snRNA and inhibits U6-U2 and U6-U4 snRNA interactions during splicing. RNA 1995, 1, 304–316. [Google Scholar]
- Wolff, T.; O’Neill, R.E.; Palese, P. NS1-Binding protein (NS1-BP): A novel human protein that interacts with the influenza A virus nonstructural NS1 protein is relocalized in the nuclei of infected cells. J. Virol. 1998, 72, 7170–7180. [Google Scholar] [CrossRef]
- Nemeroff, M.E.; Barabino, S.M.; Li, Y.; Keller, W.; Krug, R.M. Influenza virus NS1 protein interacts with the cellular 30 kDa subunit of CPSF and inhibits 3′end formation of cellular pre-mRNAs. Mol. Cell 1998, 1, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Krug, R.M. Influenza A virus NS1 protein targets poly(A)-binding protein II of the cellular 3′-end processing machinery. Embo. J. 1999, 18, 2273–2283. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.S.; Schubert, M.; Lazzarini, R.A. Polyadenylation sites for influenza virus mRNA. J. Virol. 1981, 38, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.L.; Chiou, N.T.; Kuss, S.; García-Sastre, A.; Lynch, K.W.; Fontoura, B.M. Cellular RNA binding proteins NS1-BP and hnRNP K regulate influenza A virus RNA splicing. PLoS Pathog. 2013, 9, e1003460. [Google Scholar] [CrossRef]
- Xie, Y.; Ren, Y. Mechanisms of nuclear mRNA export: A structural perspective. Traffic 2019, 20, 829–840. [Google Scholar] [CrossRef]
- Zhang, L.; Das, P.; Schmolke, M.; Manicassamy, B.; Wang, Y.; Deng, X.; Cai, L.; Tu, B.P.; Forst, C.V.; Roth, M.G.; et al. Inhibition of pyrimidine synthesis reverses viral virulence factor-mediated block of mRNA nuclear export. J. Cell Biol. 2012, 196, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Her, L.S.; Lund, E.; Dahlberg, J.E. Inhibition of Ran guanosine triphosphatase-dependent nuclear transport by the matrix protein of vesicular stomatitis virus. Science 1997, 276, 1845–1848. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.M.; Her, L.S.; Varvel, V.; Lund, E.; Dahlberg, J.E. The matrix protein of vesicular stomatitis virus inhibits nucleocytoplasmic transport when it is in the nucleus and associated with nuclear pore complexes. Mol. Cell Biol. 2000, 20, 8590–8601. [Google Scholar] [CrossRef] [Green Version]
- Quan, B.; Seo, H.S.; Blobel, G.; Ren, Y. Vesiculoviral matrix (M) protein occupies nucleic acid binding site at nucleoporin pair (Rae1 • Nup98). Proc. Natl. Acad. Sci. USA 2014, 111, 9127–9132. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Seemann, J.; Mishra, R.K.; Wei, J.H.; Weil, L.; Nussenzveig, D.R.; Heiber, J.; Barber, G.N.; Dasso, M.; Fontoura, B.M. Vesicular stomatitis virus inhibits mitotic progression and triggers cell death. EMBO Rep. 2009, 10, 1154–1160. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and ultrastructural findings of fatal COVID-19 infections in Washington State: A case series. Lancet 2020, 396, 320–332. [Google Scholar] [CrossRef]
- Zhang, K.; Miorin, L.; Makio, T.; Dehghan, I.; Gao, S.; Xie, Y.; Zhong, H.; Esparza, M.; Kehrer, T.; Kumar, A.; et al. Nsp1 protein of SARS-CoV-2 disrupts the mRNA export machinery to inhibit host gene expression. Sci. Adv. 2021, 7, eabe7386. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wen, Y.; Guo, H.; Yang, T.; Yang, H.; Ji, X. Molecular Mechanism of SARS-CoVs Orf6 Targeting the Rae1-Nup98 Complex to Compete With mRNA Nuclear Export. Front Mol. Biosci. 2021, 8, 813248. [Google Scholar] [CrossRef]
- Hall, R.; Guedán, A.; Yap, M.W.; Young, G.R.; Harvey, R.; Stoye, J.P.; Bishop, K.N. SARS-CoV-2 ORF6 disrupts innate immune signalling by inhibiting cellular mRNA export. PLoS Pathog. 2022, 18, e1010349. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Rowe, W.P.; Huebner, R.J.; Gilmore, L.K.; Parrott, R.H.; Ward, T.G. Isolation of a cytopathogenic agent from human adenoids undergoing spontaneous degeneration in tissue culture. Proc. Soc. Exp. Biol. Med. 1953, 84, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Sprengel, J.; Schmitz, B.; Heuss-Neitzel, D.; Zock, C.; Doerfler, W. Nucleotide sequence of human adenovirus type 12 DNA: Comparative functional analysis. J. Virol. 1994, 68, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Berk, A. Adenoviridae: The viruses and their replication. In Fields Virology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 2355–2394. [Google Scholar]
- Fessler, S.P.; Young, C.S. Control of adenovirus early gene expression during the late phase of infection. J. Virol. 1998, 72, 4049–4056. [Google Scholar] [CrossRef] [PubMed]
- Biasiotto, R.; Akusjärvi, G. Regulation of human adenovirus alternative RNA splicing by the adenoviral L4-33K and L4-22K proteins. Int. J. Mol. Sci. 2015, 16, 2893–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilder, S.; Moore, M.; Logan, J.; Shenk, T. The adenovirus E1B-55K transforming polypeptide modulates transport or cytoplasmic stabilization of viral and host cell mRNAs. Mol. Cell Biol. 1986, 6, 470–476. [Google Scholar]
- Leppard, K.N.; Shenk, T. The adenovirus E1B 55 kd protein influences mRNA transport via an intranuclear effect on RNA metabolism. Embo. J. 1989, 8, 2329–2336. [Google Scholar] [CrossRef]
- Leppard, K.N.; Everett, R.D. The adenovirus type 5 E1b 55K and E4 Orf3 proteins associate in infected cells and affect ND10 components. J. Gen. Virol. 1999, 80 Pt 4, 997–1008. [Google Scholar] [CrossRef]
- Ornelles, D.A.; Shenk, T. Localization of the adenovirus early region 1B 55-kilodalton protein during lytic infection: Association with nuclear viral inclusions requires the early region 4 34-kilodalton protein. J. Virol. 1991, 65, 424–429. [Google Scholar] [CrossRef]
- Beltz, G.A.; Flint, S.J. Inhibition of HeLa cell protein synthesis during adenovirus infection. Restriction of cellular messenger RNA sequences to the nucleus. J. Mol. Biol. 1979, 131, 353–373. [Google Scholar] [CrossRef]
- Blanchette, P.; Kindsmüller, K.; Groitl, P.; Dallaire, F.; Speiseder, T.; Branton, P.E.; Dobner, T. Control of mRNA export by adenovirus E4orf6 and E1B55K proteins during productive infection requires E4orf6 ubiquitin ligase activity. J. Virol. 2008, 82, 2642–2651. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.L.; Berk, A.J. Adenovirus ubiquitin-protein ligase stimulates viral late mRNA nuclear export. J. Virol. 2007, 81, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, P.; Ip, W.H.; Dobner, T.; Gonzalez, R.A. The biology of the adenovirus E1B 55K protein. FEBS Lett. 2019, 593, 3504–3517. [Google Scholar] [CrossRef]
- Gong, D.; Kim, Y.H.; Xiao, Y.; Du, Y.; Xie, Y.; Lee, K.K.; Feng, J.; Farhat, N.; Zhao, D.; Shu, S.; et al. A Herpesvirus Protein Selectively Inhibits Cellular mRNA Nuclear Export. Cell Host Microbe 2016, 20, 642–653. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Tian, H.; Wang, Y.; Zhang, Q.; Lin, N.; Liu, S.; Yu, Y.; Deng, H.; Gao, P. Molecular mechanism underlying selective inhibition of mRNA nuclear export by herpesvirus protein ORF10. Proc. Natl.Acad. Sci. USA 2020, 117, 26719–26727. [Google Scholar] [CrossRef]
- Tunnicliffe, R.B.; Tian, X.; Storer, J.; Sandri-Goldin, R.M.; Golovanov, A.P. Overlapping motifs on the herpes viral proteins ICP27 and ORF57 mediate interactions with the mRNA export adaptors ALYREF and UIF. Sci. Rep. 2018, 8, 15005. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Ikliptikawati, D.K.; Kobayashi, A.; Kondo, H.; Lim, K.; Hazawa, M.; Wong, R.W. Overexpression of SARS-CoV-2 protein ORF6 dislocates RAE1 and NUP98 from the nuclear pore complex. Biochem. Biophys. Res. Commun. 2021, 536, 59–66. [Google Scholar] [CrossRef]
- Elton, D.; Simpson-Holley, M.; Archer, K.; Medcalf, L.; Hallam, R.; McCauley, J.; Digard, P. Interaction of the influenza virus nucleoprotein with the cellular CRM1-mediated nuclear export pathway. J. Virol. 2001, 75, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, T.; Torres, R.; Salgado, E.N.; Berciu, C.; Stoddard, D.; Nicastro, D.; Jenni, S.; Harrison, S.C. Functional refolding of the penetration protein on a non-enveloped virus. Nature 2021, 590, 666–670. [Google Scholar] [CrossRef]
- Lau, S.; Weber, F. Nuclear pore protein Nup98 is involved in replication of Rift Valley fever virus and nuclear import of virulence factor NSs. J. Gen. Virol. 2020, 101, 712–716. [Google Scholar] [CrossRef]
- Ploegh, H.L. Viral strategies of immune evasion. Science 1998, 280, 248–253. [Google Scholar] [CrossRef]
- Schulz, K.S.; Mossman, K.L. Viral Evasion Strategies in Type I IFN Signaling-A Summary of Recent Developments. Front Immunol. 2016, 7, 498. [Google Scholar] [CrossRef] [Green Version]
- Beachboard, D.C.; Horner, S.M. Innate immune evasion strategies of DNA and RNA viruses. Curr. Opin. Microbiol. 2016, 32, 113–119. [Google Scholar] [CrossRef]
- Smyth, R.P.; Negroni, M.; Lever, A.M.; Mak, J.; Kenyon, J.C. RNA Structure-A Neglected Puppet Master for the Evolution of Virus and Host Immunity. Front Immunol. 2018, 9, 2097. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Chida, J. Roles of Prion Protein in Virus Infections. DNA Cell Biol. 2018, 37, 808–811. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, T.; Tang, Q.; Li, G.; Wu, P.; Chen, K. Long Non-coding RNAs: Regulators of Viral Infection and the Interferon Antiviral Response. Front Microbiol. 2018, 9, 1621. [Google Scholar] [CrossRef]
- Rossetto, C.C.; Pari, G.S. Kaposi’s sarcoma-associated herpesvirus noncoding polyadenylated nuclear RNA interacts with virus- and host cell-encoded proteins and suppresses expression of genes involved in immune modulation. J. Virol. 2011, 85, 13290–13297. [Google Scholar] [CrossRef] [Green Version]
- Schuessler, A.; Funk, A.; Lazear, H.M.; Cooper, D.A.; Torres, S.; Daffis, S.; Jha, B.K.; Kumagai, Y.; Takeuchi, O.; Hertzog, P.; et al. West Nile virus noncoding subgenomic RNA contributes to viral evasion of the type I interferon-mediated antiviral response. J. Virol. 2012, 86, 5708–5718. [Google Scholar] [CrossRef] [Green Version]
- Bidet, K.; Dadlani, D.; Garcia-Blanco, M.A. Correction: G3BP1, G3BP2 and CAPRIN1 Are Required for Translation of Interferon Stimulated mRNAs and Are Targeted by a Dengue Virus Non-coding RNA. PLoS Pathog. 2017, 13, e1006295. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.Y.; Hsu, T.W.; Chen, Y.L.; Liu, S.F.; Tsai, Y.J.; Lin, Y.T.; Chen, Y.S.; Fan, Y.H. Japanese encephalitis virus non-coding RNA inhibits activation of interferon by blocking nuclear translocation of interferon regulatory factor 3. Vet. Microbiol. 2013, 166, 11–21. [Google Scholar] [CrossRef]
- Kondo, S.; Yoshida, K.; Suzuki, M.; Saito, I.; Kanegae, Y. Adenovirus-encoding virus-associated RNAs suppress HDGF gene expression to support efficient viral replication. PLoS ONE 2014, 9, e108627. [Google Scholar] [CrossRef]
- Kapoor, A.K.; Padival, S. HIV-2 Infection. In StatPearls; StatPearls Publishing Copyright © 2023; StatPearls Publishing LLC.: Tampa, FL, USA; Treasure Island (FL) Ineligible Companies: West Palm Beach, FL, USA, 2023. [Google Scholar]
- Malim, M.H.; Hauber, J.; Le, S.Y.; Maizel, J.V.; Cullen, B.R. The HIV-1 rev trans-activator acts through a structured target sequence to activate nuclear export of unspliced viral mRNA. Dis. Markers 1990, 8, 39. [Google Scholar] [CrossRef]
- Fischer, U.; Meyer, S.; Teufel, M.; Heckel, C.; Lührmann, R.; Rautmann, G. Evidence that HIV-1 Rev directly promotes the nuclear export of unspliced RNA. Embo. J. 1994, 13, 4105–4112. [Google Scholar] [CrossRef]
- Malim, M.H.; Cullen, B.R. HIV-1 structural gene expression requires the binding of multiple Rev monomers to the viral RRE: Implications for HIV-1 latency. Cell 1991, 65, 241–248. [Google Scholar] [CrossRef]
- Budhiraja, S.; Liu, H.; Couturier, J.; Malovannaya, A.; Qin, J.; Lewis, D.E.; Rice, A.P. Mining the human complexome database identifies RBM14 as an XPO1-associated protein involved in HIV-1 Rev function. J. Virol. 2015, 89, 3557–3567. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Hu, P.W.; Budhiraja, S.; Misra, A.; Couturier, J.; Lloyd, R.E.; Lewis, D.E.; Kimata, J.T.; Rice, A.P. PACS1 is an HIV-1 cofactor that functions in Rev-mediated nuclear export of viral RNA. Virology 2020, 540, 88–96. [Google Scholar] [CrossRef]
- Yedavalli, V.S.; Neuveut, C.; Chi, Y.H.; Kleiman, L.; Jeang, K.T. Requirement of DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell 2004, 119, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Mertz, J.A.; Simper, M.S.; Lozano, M.M.; Payne, S.M.; Dudley, J.P. Mouse mammary tumor virus encodes a self-regulatory RNA export protein and is a complex retrovirus. J. Virol. 2005, 79, 14737–14747. [Google Scholar] [CrossRef] [Green Version]
- Indik, S.; Günzburg, W.H.; Salmons, B.; Rouault, F. A novel, mouse mammary tumor virus encoded protein with Rev-like properties. Virology 2005, 337, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mertz, J.A.; Lozano, M.M.; Dudley, J.P. Rev and Rex proteins of human complex retroviruses function with the MMTV Rem-responsive element. Retrovirology 2009, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Bodem, J.; Schied, T.; Gabriel, R.; Rammling, M.; Rethwilm, A. Foamy virus nuclear RNA export is distinct from that of other retroviruses. J. Virol. 2011, 85, 2333–2341. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.; Hughes, M.T.; Kawaoka, Y. Influenza A virus NS2 protein mediates vRNP nuclear export through NES-independent interaction with hCRM1. Embo. J. 2000, 19, 6751–6758. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Chen, J.; Chen, Q.; Wang, H.; Yao, Y.; Chen, J.; Chen, Z. A second CRM1-dependent nuclear export signal in the influenza A virus NS2 protein contributes to the nuclear export of viral ribonucleoproteins. J. Virol. 2013, 87, 767–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, S.; Bhaumik, S.R. Viral regulation of mRNA export with potentials for targeted therapy. Biochim. Biophys. Acta Gene Regul. Mech 2021, 1864, 194655. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Ernst, R.K.; Lund, E.; Grimm, C.; Zapp, M.L.; Rekosh, D.; Hammarskjöld, M.L.; Dahlberg, J.E. The constitutive transport element (CTE) of Mason-Pfizer monkey virus (MPMV) accesses a cellular mRNA export pathway. Embo. J. 1997, 16, 7500–7510. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bor, Y.C.; Misawa, Y.; Xue, Y.; Rekosh, D.; Hammarskjöld, M.L. An intron with a constitutive transport element is retained in a Tap messenger RNA. Nature 2006, 443, 234–237. [Google Scholar] [CrossRef]
- Kang, Y.; Bogerd, H.P.; Cullen, B.R. Analysis of cellular factors that mediate nuclear export of RNAs bearing the Mason-Pfizer monkey virus constitutive transport element. J. Virol. 2000, 74, 5863–5871. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Cullen, B.R. Structural and functional analysis of the avian leukemia virus constitutive transport element. Rna 1999, 5, 1645–1655. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, T.; Davila, J.I.; Malcolm, J.A.; Kocher, J.P.; Tonne, J.M.; Ikeda, Y. Murine leukemia virus uses NXF1 for nuclear export of spliced and unspliced viral transcripts. J. Virol. 2014, 88, 4069–4082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mougel, M.; Akkawi, C.; Chamontin, C.; Feuillard, J.; Pessel-Vivares, L.; Socol, M.; Laine, S. NXF1 and CRM1 nuclear export pathways orchestrate nuclear export, translation and packaging of murine leukaemia retrovirus unspliced RNA. RNA Biol. 2020, 17, 528–538. [Google Scholar] [CrossRef]
- Paca, R.E.; Ogert, R.A.; Hibbert, C.S.; Izaurralde, E.; Beemon, K.L. Rous sarcoma virus DR posttranscriptional elements use a novel RNA export pathway. J. Virol. 2000, 74, 9507–9514. [Google Scholar] [CrossRef] [Green Version]
- Bhat, P.; Aksenova, V.; Gazzara, M.; Rex, E.A.; Aslam, S.; Haddad, C.; Gao, S.; Esparza, M.; Cagatay, T.; Batten, K.; et al. Influenza virus mRNAs encode determinants for nuclear export via the cellular TREX-2 complex. Nat. Commun. 2023, 14, 2304. [Google Scholar] [CrossRef]
- Yoon, D.W.; Lee, H.; Seol, W.; DeMaria, M.; Rosenzweig, M.; Jung, J.U. Tap: A novel cellular protein that interacts with tip of herpesvirus saimiri and induces lymphocyte aggregation. Immunity 1997, 6, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Gabler, S.; Schütt, H.; Groitl, P.; Wolf, H.; Shenk, T.; Dobner, T. E1B 55-kilodalton-associated protein: A cellular protein with RNA-binding activity implicated in nucleocytoplasmic transport of adenovirus and cellular mRNAs. J. Virol. 1998, 72, 7960–7971. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F.P.; Sandri-Goldin, R.M. Head-to-tail intramolecular interaction of herpes simplex virus type 1 regulatory protein ICP27 is important for its interaction with cellular mRNA export receptor TAP/NXF1. mBio 2010, 1, 1110–1128. [Google Scholar] [CrossRef] [Green Version]
- Yatherajam, G.; Huang, W.; Flint, S.J. Export of adenoviral late mRNA from the nucleus requires the Nxf1/Tap export receptor. J. Virol. 2011, 85, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.C.; Nickens, D.G.; Lentz, T.B.; Loeb, D.D.; Zlotnick, A. Encapsidated hepatitis B virus reverse transcriptase is poised on an ordered RNA lattice. Proc. Natl. Acad. Sci. USA 2014, 111, 11329–11334. [Google Scholar] [CrossRef]
- Smith, G.J., 3rd; Donello, J.E.; Lück, R.; Steger, G.; Hope, T.J. The hepatitis B virus post-transcriptional regulatory element contains two conserved RNA stem-loops which are required for function. Nucleic Acids Res. 1998, 26, 4818–4827. [Google Scholar] [CrossRef] [Green Version]
- Chi, B.; Wang, K.; Du, Y.; Gui, B.; Chang, X.; Wang, L.; Fan, J.; Chen, S.; Wu, X.; Li, G.; et al. A Sub-Element in PRE enhances nuclear export of intronless mRNAs by recruiting the TREX complex via ZC3H18. Nucleic Acids Res. 2014, 42, 7305–7318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makokha, G.N.; Abe-Chayama, H.; Chowdhury, S.; Hayes, C.N.; Tsuge, M.; Yoshima, T.; Ishida, Y.; Zhang, Y.; Uchida, T.; Tateno, C.; et al. Regulation of the Hepatitis B virus replication and gene expression by the multi-functional protein TARDBP. Sci. Rep. 2019, 9, 8462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Yu, L.; Zhu, N.; Xie, J. Cellular UAP56 interacts with the HBx protein of the hepatitis B virus and is involved in viral RNA nuclear export in hepatocytes. Exp. Cell Res. 2020, 390, 111929. [Google Scholar] [CrossRef]
- Nakano, K.; Watanabe, T. HTLV-1 Rex Tunes the Cellular Environment Favorable for Viral Replication. Viruses 2016, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Mathew, C.; Ghildyal, R. CRM1 Inhibitors for Antiviral Therapy. Front. Microbiol. 2017, 8, 1171. [Google Scholar] [CrossRef] [Green Version]
- Bartels, H.; Luban, J. Gammaretroviral pol sequences act in cis to direct polysome loading and NXF1/NXT-dependent protein production by gag-encoded RNA. Retrovirology 2014, 11, 73. [Google Scholar]
- Scheifele, L.Z.; Garbitt, R.A.; Rhoads, J.D.; Parent, L.J. Nuclear entry and CRM1-dependent nuclear export of the Rous sarcoma virus Gag polyprotein. Proc. Natl. Acad. Sci. USA 2002, 99, 3944–3949. [Google Scholar] [CrossRef] [PubMed]
- Renault, N.; Tobaly-Tapiero, J.; Paris, J.; Giron, M.L.; Coiffic, A.; Roingeard, P.; Saïb, A. A nuclear export signal within the structural Gag protein is required for prototype foamy virus replication. Retrovirology 2011, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolowski, M.; Furneaux, H.; Schwartz, S. The inhibitory activity of the AU-rich RNA element in the human papillomavirus type 1 late 3’ untranslated region correlates with its affinity for the elav-like HuR protein. J. Virol. 1999, 73, 1080–1091. [Google Scholar] [CrossRef]
- Soussan, P.; Tuveri, R.; Nalpas, B.; Garreau, F.; Zavala, F.; Masson, A.; Pol, S.; Brechot, C.; Kremsdorf, D. The expression of hepatitis B spliced protein (HBSP) encoded by a spliced hepatitis B virus RNA is associated with viral replication and liver fibrosis. J. Hepatol. 2003, 38, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Esparza, M.; Mor, A.; Niederstrasser, H.; White, K.; White, A.; Zhang, K.; Gao, S.; Wang, J.; Liang, J.; Sho, S.; et al. Chemical intervention of influenza virus mRNA nuclear export. PLoS Pathog. 2020, 16, e1008407. [Google Scholar] [CrossRef]
Figure 1.
Viruses utilize multiple strategies to suppress host mRNA nuclear export. After transcription and splicing, the mRNA nuclear export factors start binding to the mRNA. Several factors are involved in the process, including the TREX complex, Aly/REF, and UAP56. The NXF1–NXT1 heterodimer is a major mRNA nuclear export factor that recruits mRNA to dock into the NPC. After several steps of translocation, the mRNA is exported from the nucleus to the cytoplasm for translation into protein. This process is targeted by viral proteins. IAV NS1 binds to CPSF30 and PAB II, resulting in the deficiency of mRNA polyadenylation. In addition, IAV NS1 can block the host mRNA nuclear export by directly binding to NXF1, inhibiting the host mRNA docking to the NPC. Several SARS-CoV-2 proteins could be involved in the inhibition of host mRNA nuclear export. SARS-CoV-2 Nsp1 directly binds to NXF1, preventing NXF1 from docking at the NPC. SARS-CoV-2 Orf6 forms a complex with the Rae1-Nup98 heterodimer, and its C-terminus occupies the mRNA binding groove in the Rae1-Nup98 complex. VSV and KSHV also block the host mRNA nuclear export by targeting the Rae1-Nup98 complex by VSV M, or KSHV ORF10 directly binds to the Rae1-Nup98 complex. AdV infection inhibits NXF1-mediated nuclear export of cellular mRNA. The AdV early protein E1B-55K, which cooperates with the AdV E4orf6 protein, recruits E1B-AP5 to bind NXF1.
Figure 1.
Viruses utilize multiple strategies to suppress host mRNA nuclear export. After transcription and splicing, the mRNA nuclear export factors start binding to the mRNA. Several factors are involved in the process, including the TREX complex, Aly/REF, and UAP56. The NXF1–NXT1 heterodimer is a major mRNA nuclear export factor that recruits mRNA to dock into the NPC. After several steps of translocation, the mRNA is exported from the nucleus to the cytoplasm for translation into protein. This process is targeted by viral proteins. IAV NS1 binds to CPSF30 and PAB II, resulting in the deficiency of mRNA polyadenylation. In addition, IAV NS1 can block the host mRNA nuclear export by directly binding to NXF1, inhibiting the host mRNA docking to the NPC. Several SARS-CoV-2 proteins could be involved in the inhibition of host mRNA nuclear export. SARS-CoV-2 Nsp1 directly binds to NXF1, preventing NXF1 from docking at the NPC. SARS-CoV-2 Orf6 forms a complex with the Rae1-Nup98 heterodimer, and its C-terminus occupies the mRNA binding groove in the Rae1-Nup98 complex. VSV and KSHV also block the host mRNA nuclear export by targeting the Rae1-Nup98 complex by VSV M, or KSHV ORF10 directly binds to the Rae1-Nup98 complex. AdV infection inhibits NXF1-mediated nuclear export of cellular mRNA. The AdV early protein E1B-55K, which cooperates with the AdV E4orf6 protein, recruits E1B-AP5 to bind NXF1.

Figure 2.
Structure of the viral protein and mRNA nuclear export factor complex. (A) Crystal structure of the IAV NS1 and NXF1–NXT1 complex. In the crystal, two NXF1–NXT1 heterodimers are linked by the two NXT1 molecules. Two NS1 molecules dimerize through their RBD domains. An effector domain binds to the LRR domain of NXF1. Another effector domain binds to the NTF2L domain of the neighboring NXF1, which is also the nucleoporin binding domain. (B) Crystal structure of VSV M and Rae1-Nup98 complex. The association of VSV M with Rae1-Nup98 is mainly due to the interaction of two extensions of the M global domain with Rae1. The highly conserved Methionine 51 facilitates interaction with Rae1. (C) Crystal structure of SARS-CoV-2 Orf6 and Rae1-Nup98 complex. This model shows that the SARS-CoV-2 Orf6 C-terminal domain and Rae1-Nup98GLEB form a heterotrimer in which the Orf6 C-terminal domain binds directly to Rae1. The highly conserved methionine 58 of Orf6 plays an essential role in the interaction and is located at the mRNA binding site of Rae1. (D) Crystal structure of the KSHV ORF10 and Rae1-Nup98 complex. KSHV ORF10-Rae1-Nup98GLEB forms a heterotrimer, and the KSHV ORF10 C-terminal domain interacts directly with Rae1. The conserved residues of ORF10 methionine 413 and leucine 60 majorly contribute to the interaction with Rae1.
Figure 2.
Structure of the viral protein and mRNA nuclear export factor complex. (A) Crystal structure of the IAV NS1 and NXF1–NXT1 complex. In the crystal, two NXF1–NXT1 heterodimers are linked by the two NXT1 molecules. Two NS1 molecules dimerize through their RBD domains. An effector domain binds to the LRR domain of NXF1. Another effector domain binds to the NTF2L domain of the neighboring NXF1, which is also the nucleoporin binding domain. (B) Crystal structure of VSV M and Rae1-Nup98 complex. The association of VSV M with Rae1-Nup98 is mainly due to the interaction of two extensions of the M global domain with Rae1. The highly conserved Methionine 51 facilitates interaction with Rae1. (C) Crystal structure of SARS-CoV-2 Orf6 and Rae1-Nup98 complex. This model shows that the SARS-CoV-2 Orf6 C-terminal domain and Rae1-Nup98GLEB form a heterotrimer in which the Orf6 C-terminal domain binds directly to Rae1. The highly conserved methionine 58 of Orf6 plays an essential role in the interaction and is located at the mRNA binding site of Rae1. (D) Crystal structure of the KSHV ORF10 and Rae1-Nup98 complex. KSHV ORF10-Rae1-Nup98GLEB forms a heterotrimer, and the KSHV ORF10 C-terminal domain interacts directly with Rae1. The conserved residues of ORF10 methionine 413 and leucine 60 majorly contribute to the interaction with Rae1.

Figure 3.
Viruses hijack the cellular machinery to facilitate the nuclear export of viral mRNA. (A) Examples of viral mRNA nuclear export dependent on the CRM1 pathway. Nuclear export of HFV mRNA, HPV mRNA, and unspliced human HIV-1 mRNA depends on the CRM1 pathway. The RRE RNA element is recognized by HIV Rev protein, which recruits the CRM1 for mRNA nuclear export. The nuclear export of HPV and HFV depends on the indirect recruitment of CRM1 by the cellular protein HuR. HuR can bind to the negative regulator element (NRE) RNA element on HPV mRNA. The mRNA-protein complex binds to RanGTP and is then translocated via NPC. (B) Examples of viral mRNA nuclear export dependent on NXF1–NXT1 mediating pathway. The nuclear export of HBV mRNA, AdV mRNA, HSV-1 mRNA, IAV mRNA, and MPMV mRNA is mediated by NXF1–NXT1 pathway. The PRE is recognized by cellular proteins, such as the ZC3H18, which is responsible for recruiting members of the TREX complex. The nuclear export of AdV mRNA involved the participation of E1B-55K and E1B-AP5. E1B-AP5 plays the role of interacting with both viral mRNA and NXF1. HSV-1 ICP27 is an export adaptor that links NXF1 and Aly/REF to the viral mRNA for nuclear export. The nuclear export of different types (intronless, unspliced, and spliced) of IAV mRNAs through the NXF1–NXT1 pathway is regulated by various cellular factors, including NS1-BP, UAP56, DDX19, and eIF4A3. The constitutive transport element (CTE) on MPMV mRNA strongly interacts with NXF1 for nuclear export.
Figure 3.
Viruses hijack the cellular machinery to facilitate the nuclear export of viral mRNA. (A) Examples of viral mRNA nuclear export dependent on the CRM1 pathway. Nuclear export of HFV mRNA, HPV mRNA, and unspliced human HIV-1 mRNA depends on the CRM1 pathway. The RRE RNA element is recognized by HIV Rev protein, which recruits the CRM1 for mRNA nuclear export. The nuclear export of HPV and HFV depends on the indirect recruitment of CRM1 by the cellular protein HuR. HuR can bind to the negative regulator element (NRE) RNA element on HPV mRNA. The mRNA-protein complex binds to RanGTP and is then translocated via NPC. (B) Examples of viral mRNA nuclear export dependent on NXF1–NXT1 mediating pathway. The nuclear export of HBV mRNA, AdV mRNA, HSV-1 mRNA, IAV mRNA, and MPMV mRNA is mediated by NXF1–NXT1 pathway. The PRE is recognized by cellular proteins, such as the ZC3H18, which is responsible for recruiting members of the TREX complex. The nuclear export of AdV mRNA involved the participation of E1B-55K and E1B-AP5. E1B-AP5 plays the role of interacting with both viral mRNA and NXF1. HSV-1 ICP27 is an export adaptor that links NXF1 and Aly/REF to the viral mRNA for nuclear export. The nuclear export of different types (intronless, unspliced, and spliced) of IAV mRNAs through the NXF1–NXT1 pathway is regulated by various cellular factors, including NS1-BP, UAP56, DDX19, and eIF4A3. The constitutive transport element (CTE) on MPMV mRNA strongly interacts with NXF1 for nuclear export.


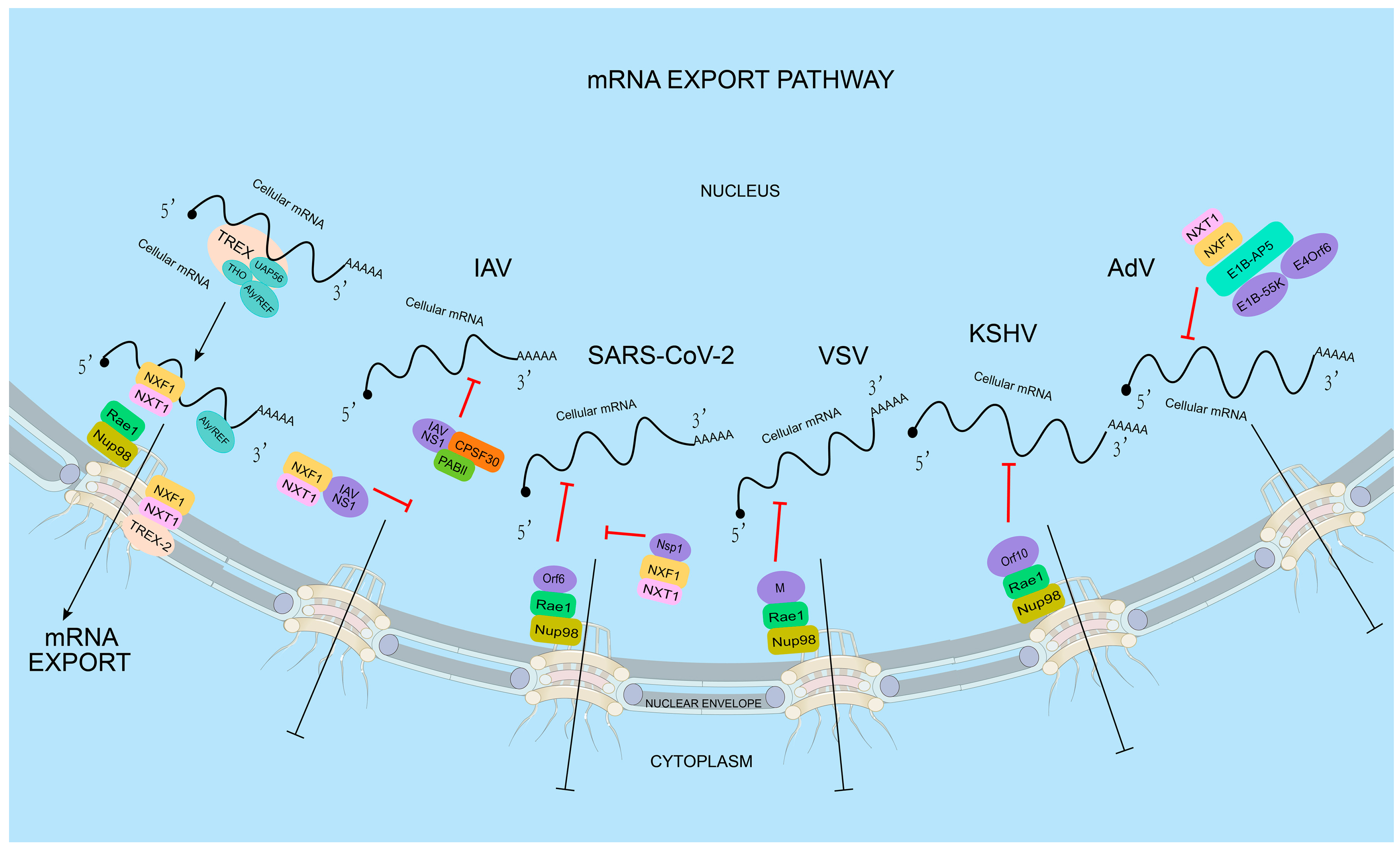{kind=link}
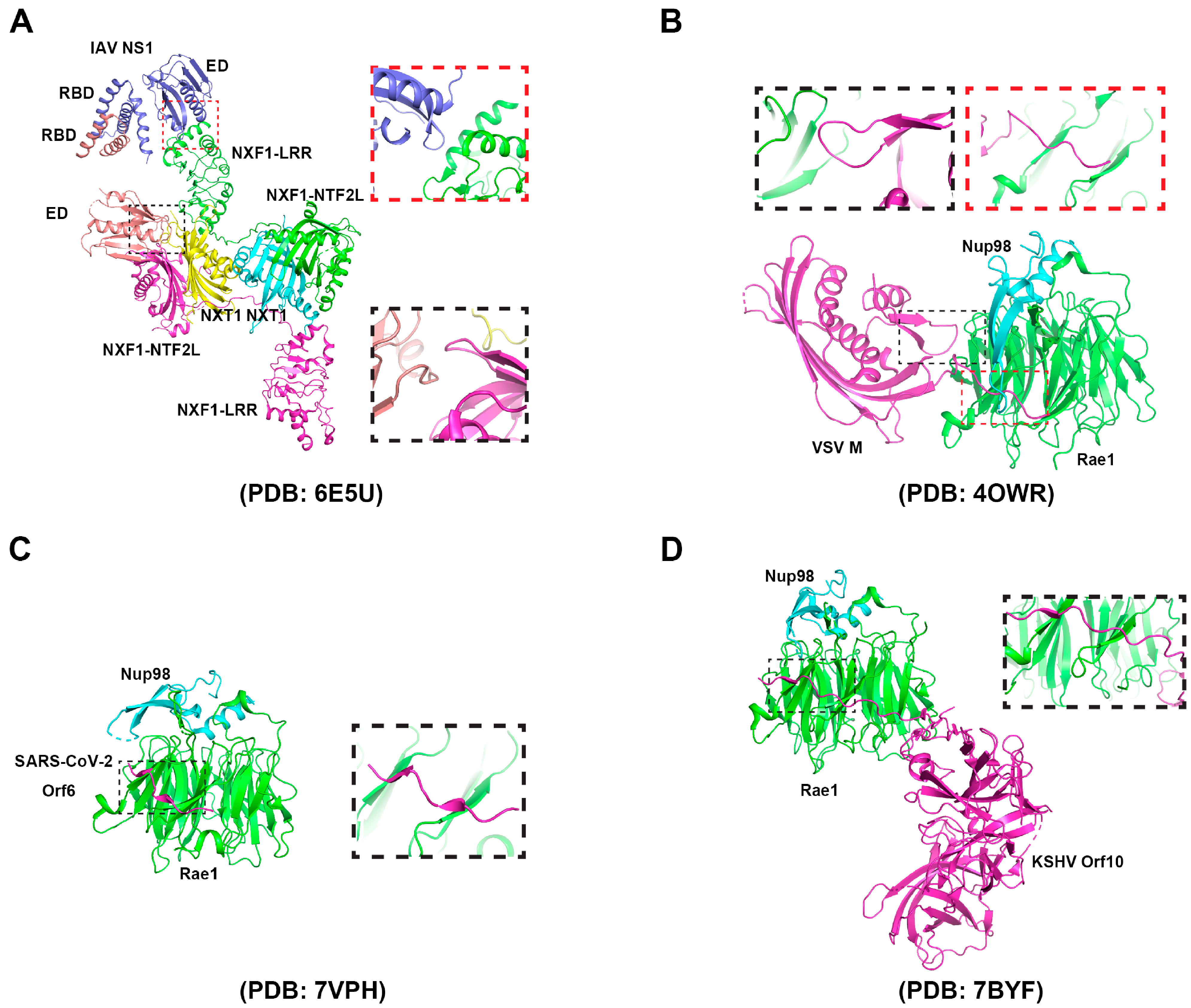{kind=link}
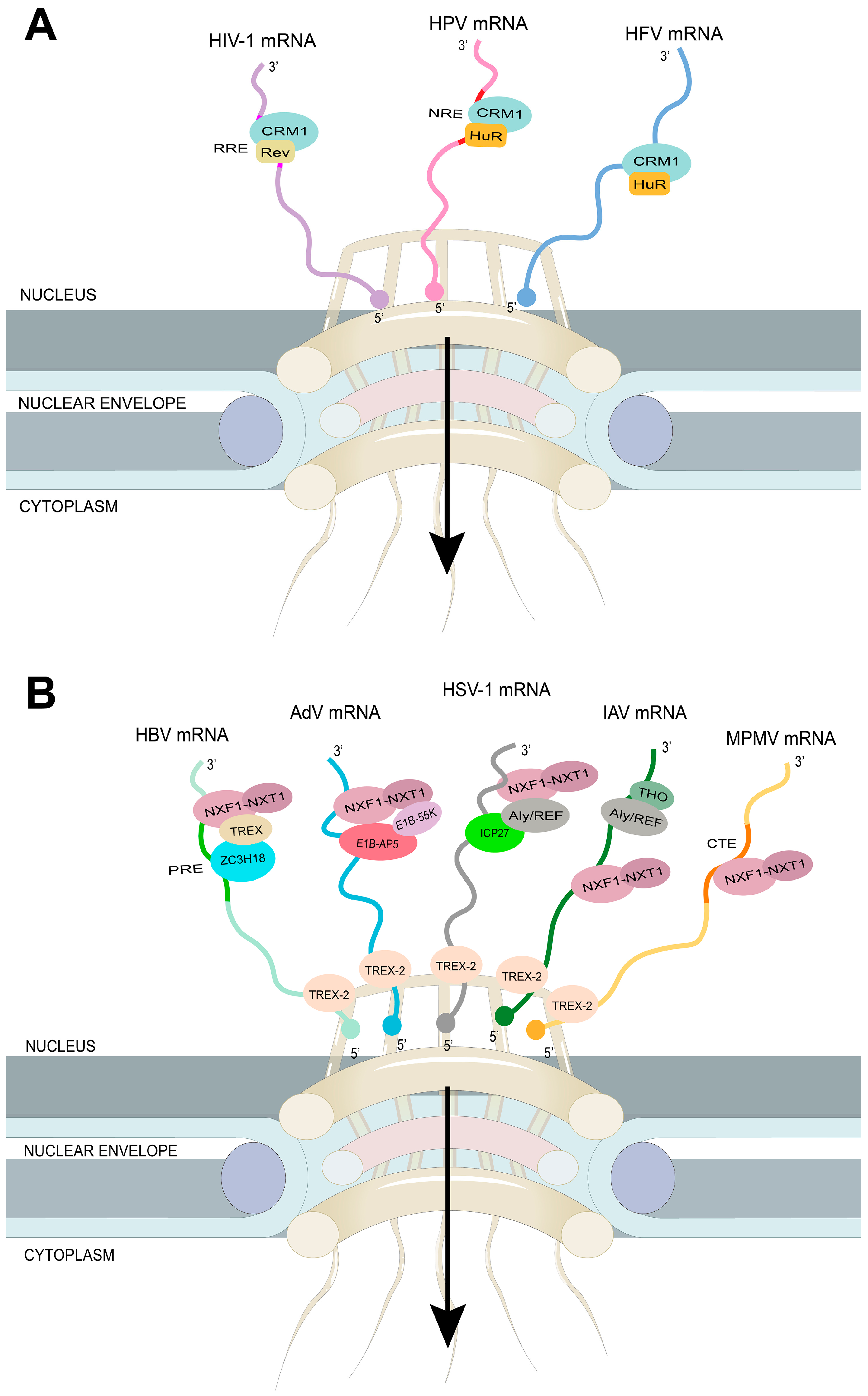{kind=link}
Table 1.
Mechanisms of viruses’ arrest of host mRNA nuclear export.
| Group | Virus (Family) | Viral Protein | Mechanism |
|---|---|---|---|
| RNA-Virus | Influenza A virus (Orthomyxoviridae) | NS1 | Disruption of NXF1 docking to the NPC [60] |
| NS1 | Inhibition of mRNA polyadenylation [60] | ||
| SARS-CoV (Coronaviridae) | NSP1 | Alteration of Nup93 localization [118] | |
| SARS-CoV-2 (Coronaviridae) | NSP1 | Prevention of NXF1 docking at NPC [100] | |
| Orf6 | Interaction with Rae1-Nup98 complex [119] | ||
| Vesicular stomatitis virus (Rhabdoviridae) | M | Hijacking the Rae1-Nup98 complex [61,120] | |
| Poliovirus (Picornaviridae) | 2Apro | Degradation of Nup62, Nup98, and Nup153 [61,120] | |
| Human rhinovirus (Picornaviridae) | 3Cpro | Degradation of Nup153, Nup214, and Nup358 [61,120] | |
| Encephalomyocarditis virus (Picornaviridae) | L | Altering the RanGDP/GTP gradient [76,77,78,79] | |
| Theiler’s Murine encephalomyelitis virus (Picornaviridae) | L | Hyper-phosphorylation of Nup98 [77] | |
| Rhesus rotavirus (Reoviridae) | NSP3 | Hyperadenylation of cellular mRNA [121] | |
| Zika virus (Flaviviridae) | Degradation of UPF1 [80] | ||
| Rift Valley fever virus (Phenuiviridae) | NSs | Unknown [122] | |
| DNA-Virus | Adenoviruses (Adenoviridae) | E1B-55K, E4Orf6 | Interaction with E1B-AP5 [112] |
| Kaposi’s sarcoma-associated herpesvirus (Herpesviridae) | ORF10 | Hijacking the Rae1-Nup98 complex [116] |
Table 2.
Mechanisms of viruses’s mRNA nuclear export.
| Group | Virus (Family) | Pathway | Mechanism |
|---|---|---|---|
| RNA-Virus | Influenza A virus (Orthomyxoviridae) | NXF1–NXT1 | Regulation by viral protein NS1, and cellular proteins NS1-BP, UAP56, DDX19, and eIF4A3 [14,28,29,30]. |
| Human Immunodeficiency Virus 1 (Retroviridae) | CRM1 | The recruitment of CRM1 by HIV-1 Rev [10,135,136,137]. | |
| Mason-pfizer Monkey virus (Retroviridae) | NXF1–NXT1 | The recruitment of cellular export receptor NXF1–NXT1 by constitutive transport elements [148,149,150]. | |
| Human T-cell leukemia virus type 1 (Retroviridae) | CRM1 | The recruitment of CRM1 by HTLV-1 Rex [165,166]. | |
| Murine leukemia virus (Retroviridae) | CRM1 | Unknown (unspliced mRNAs for packaging) [152]. | |
| NXF1–NXT1 | Dependent on UAP56 (spliced mRNA) [152]. | ||
| Dependent on THOC5, THOC7, and SRp20 (unspliced genomic mRNA) [152,167]. | |||
| Rous Sarcoma Virus (Retroviridae) | CRM1 | The contribution of RSV Gag (unspliced mRNAs) [168]. | |
| NXF1–NXT1 | The recruitment of NXF1 and DDX5 by direct repeat sequences flanking the src gene [154]. | ||
| Prototype foamy virus (Retroviridae) | CRM1 | The recruitment [163] of CRM1 by the host adapter HuR [169]. | |
| DNA-Virus | Adenoviruses (Adenoviridae) | NXF1–NXT1 | Regulation by AdV E1B-55K, and cellular protein E1B-AP5 [112]. |
| Herpes simplex virus 1 (Herpesviridae) | NXF1–NXT1 | The interaction of HSV-1 ICP27 with NXF1, Aly/REF, and viral mRNAs [118,158]. | |
| Human papillomavirus (Papoviviridae) | CRM1 | The recruitment of host adapter HuR by viral AU rich element [170]. | |
| Hepatitis B virus (Hepadnaviridae) | NXF1–NXT1 | The recruitment of TREX components by ZC3H18 (intronless mRNA) [162]. | |
| The recruitment of NXF1 by an unknown cellular factor (unspliced PreC mRNA) [171]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, J.; Zhu, Y.; Ma, X.; Shang, G.; Liu, B.; Zhang, K. Virus Infection and mRNA Nuclear Export. Int. J. Mol. Sci. 2023, 24, 12593. https://doi.org/10.3390/ijms241612593
AMA Style
Guo J, Zhu Y, Ma X, Shang G, Liu B, Zhang K. Virus Infection and mRNA Nuclear Export. International Journal of Molecular Sciences. 2023; 24(16):12593. https://doi.org/10.3390/ijms241612593
Chicago/Turabian StyleGuo, Jiayin, Yaru Zhu, Xiaoya Ma, Guijun Shang, Bo Liu, and Ke Zhang. 2023. "Virus Infection and mRNA Nuclear Export" International Journal of Molecular Sciences 24, no. 16: 12593. https://doi.org/10.3390/ijms241612593
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.