
Comparative Analysis of Tumor-Associated microRNAs and Tetraspanines from Exosomes of Plasma and Ascitic Fluids of Ovarian Cancer Patients
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
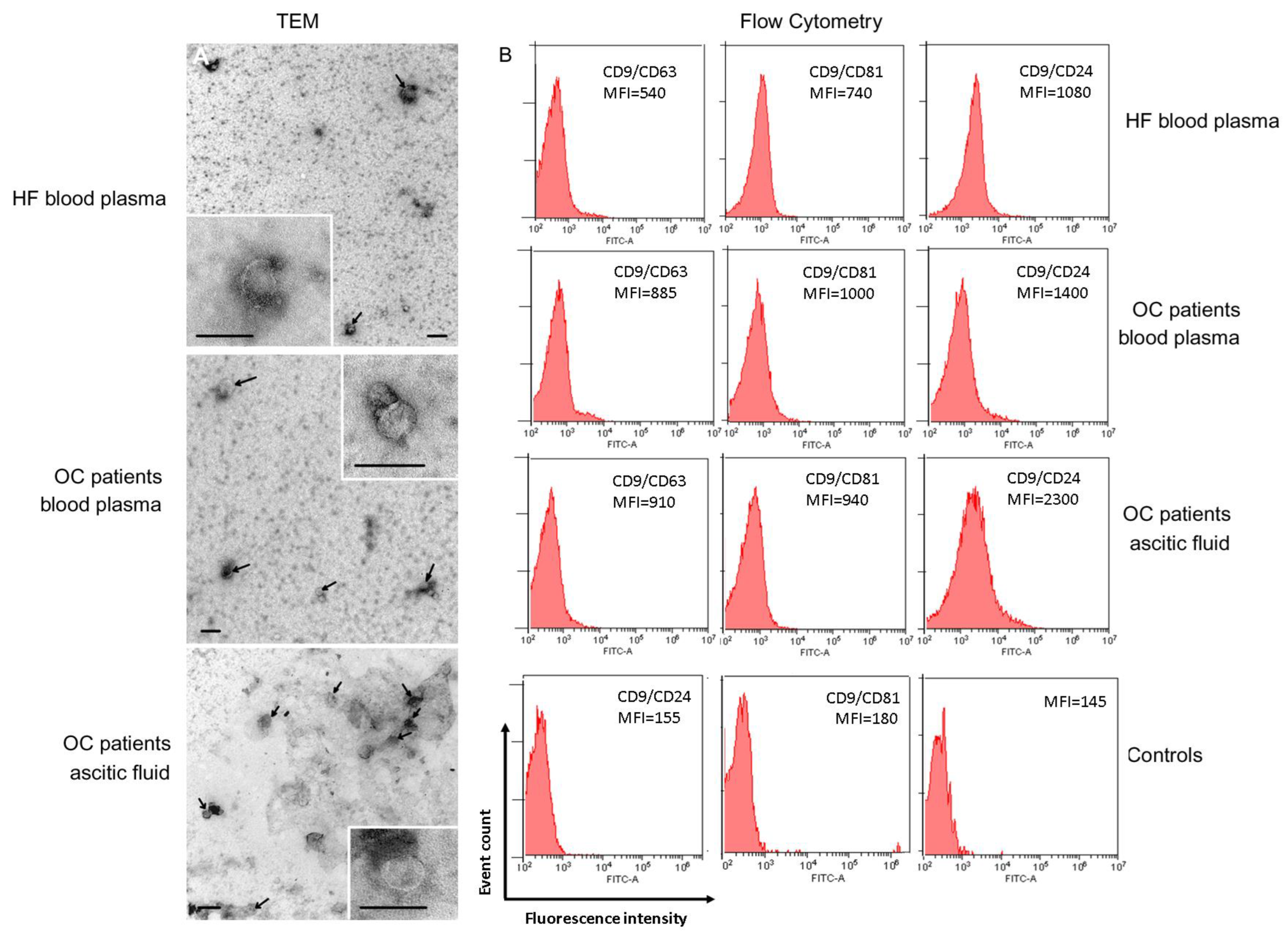
2.1. Characterization of Exosomes
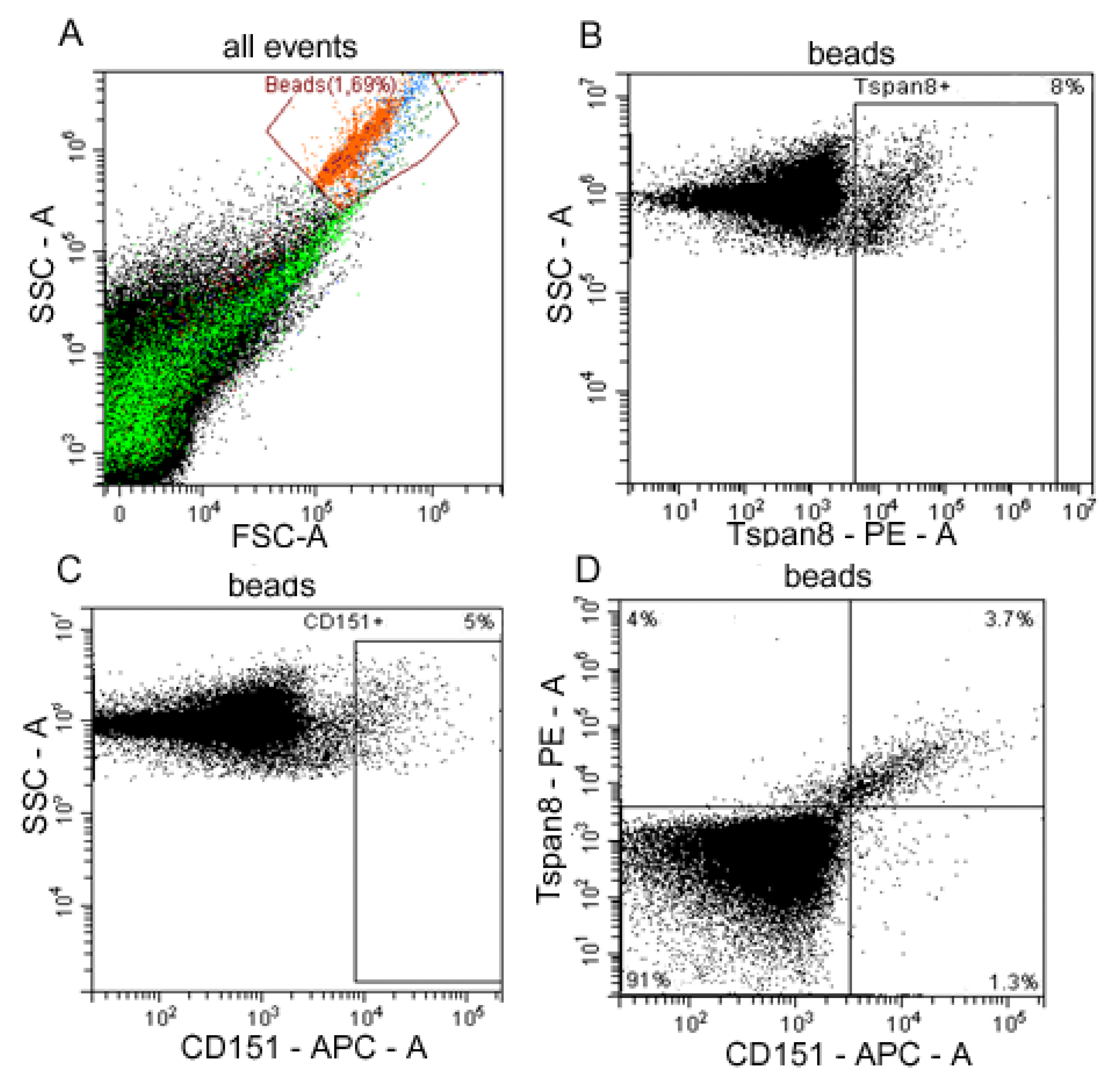
2.2. Distribution of CD151 and Tspan8 on the Surface of Exosomes in HFs and OC Patients
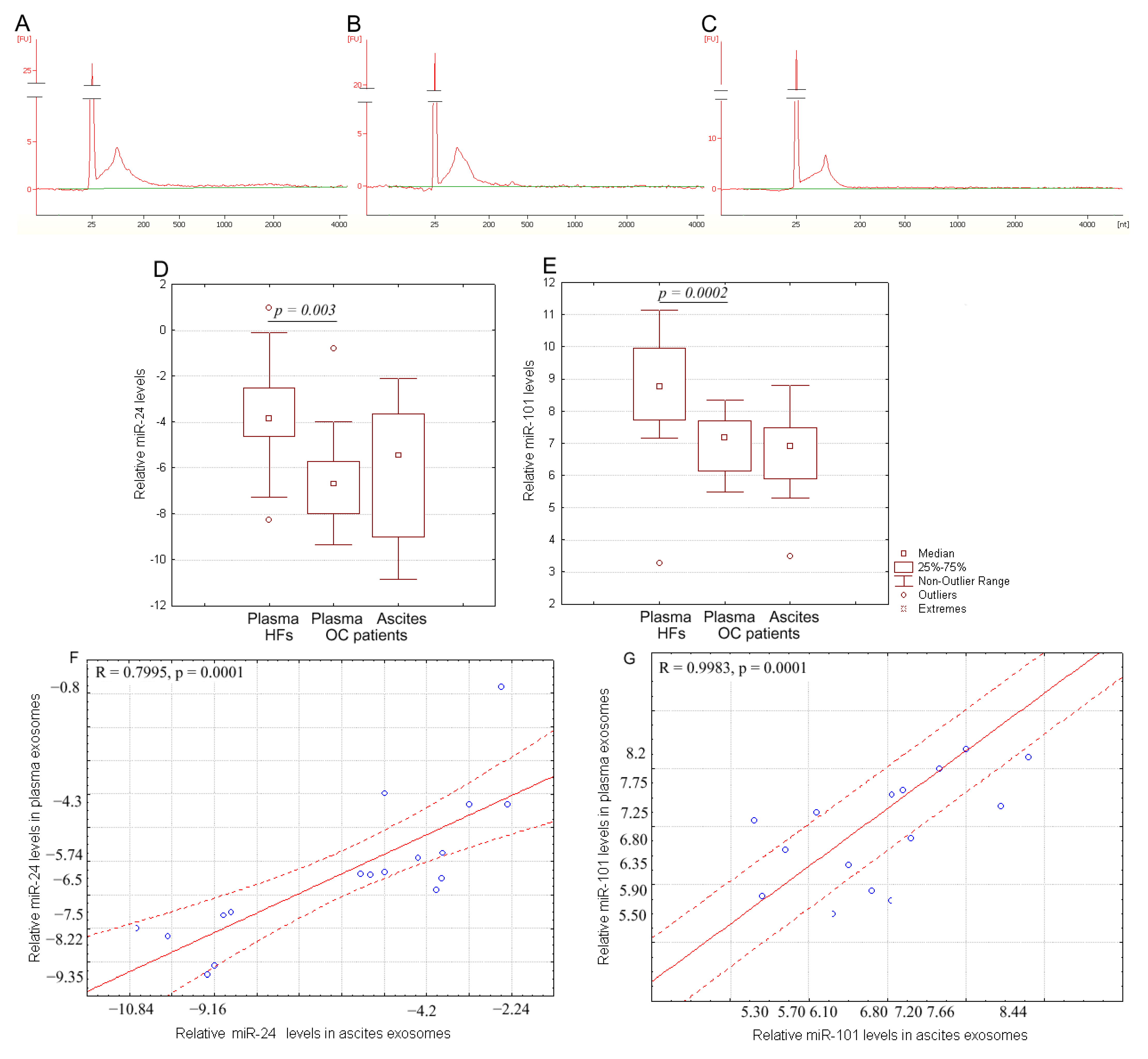
2.3. Expression of miR-24-3p and miR-101 in Exosomes from Plasma and Ascites of OC Patients
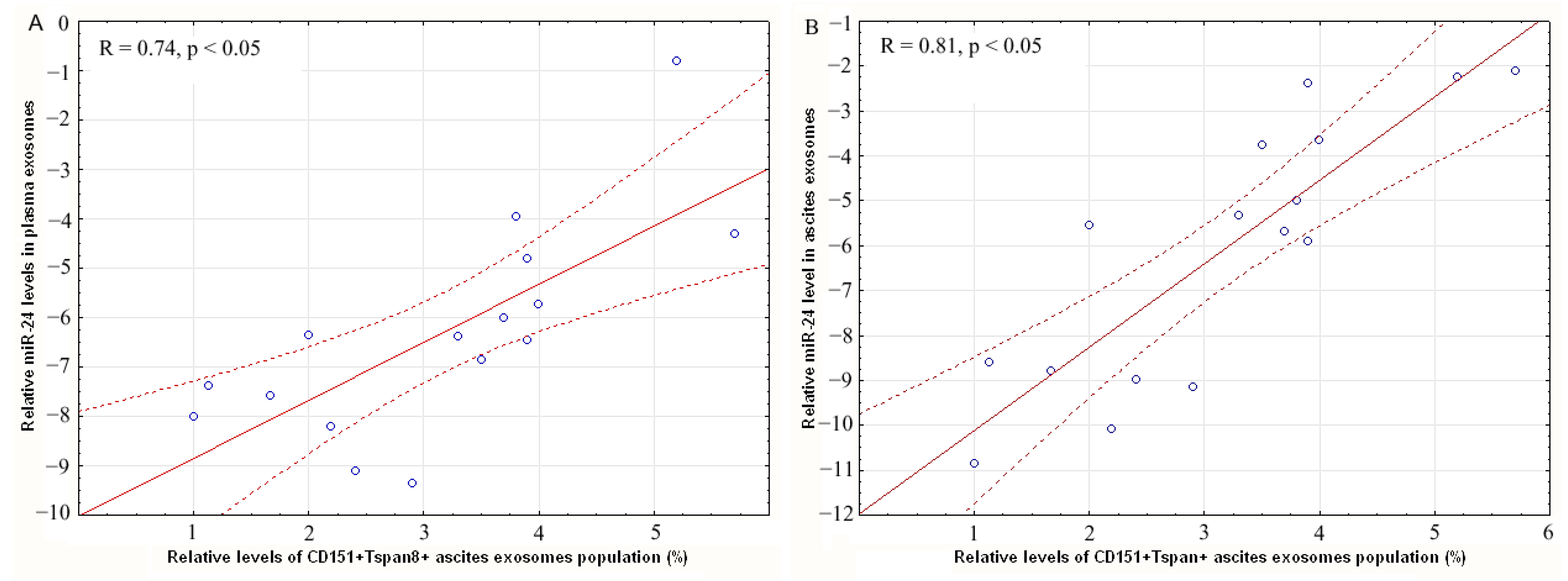
2.4. Levels of miR-24-3p in Plasma and Ascites Exosomes Are Related with Expression of CD151 and Tspan8 on CD9-Positive Exosomes
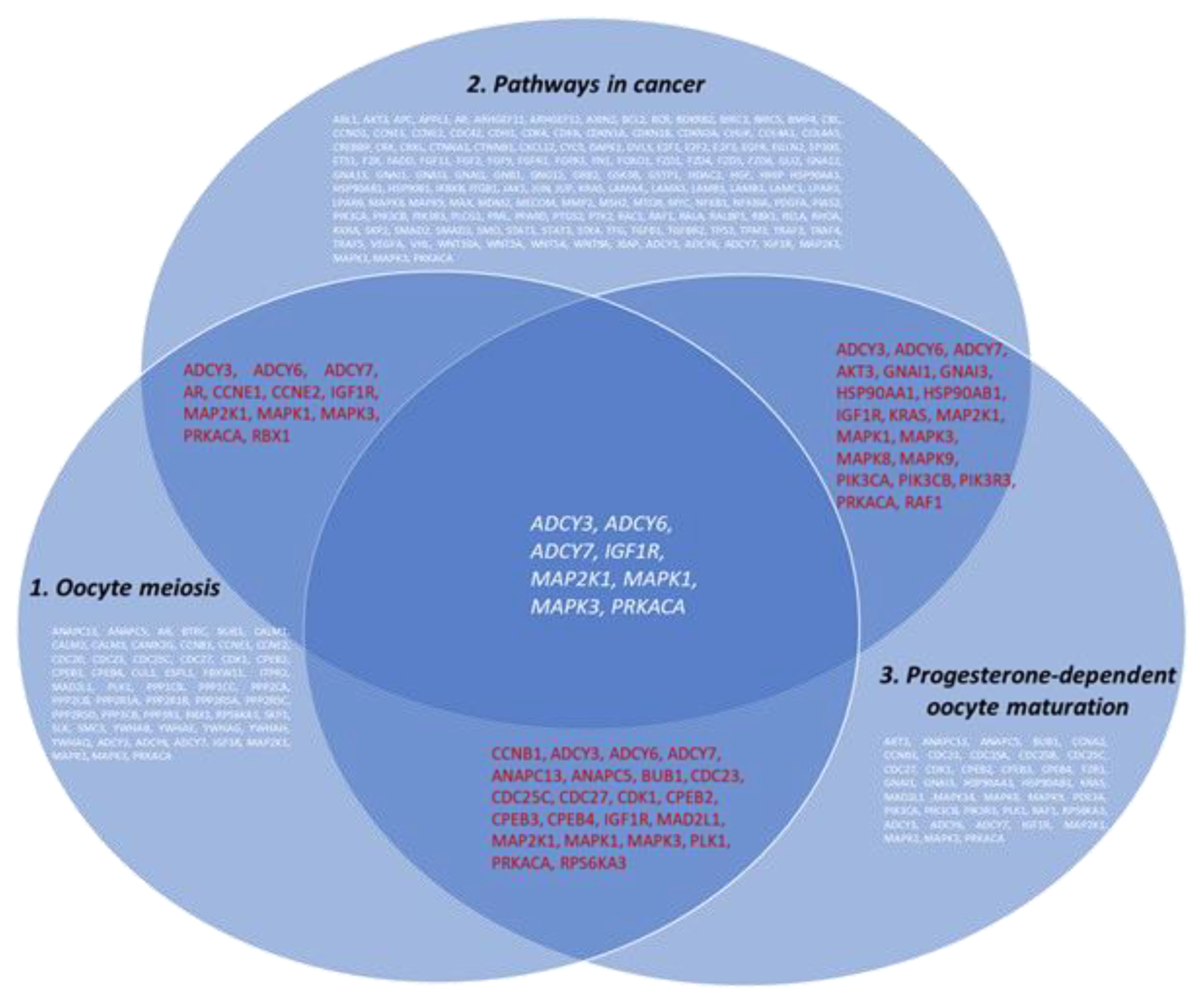
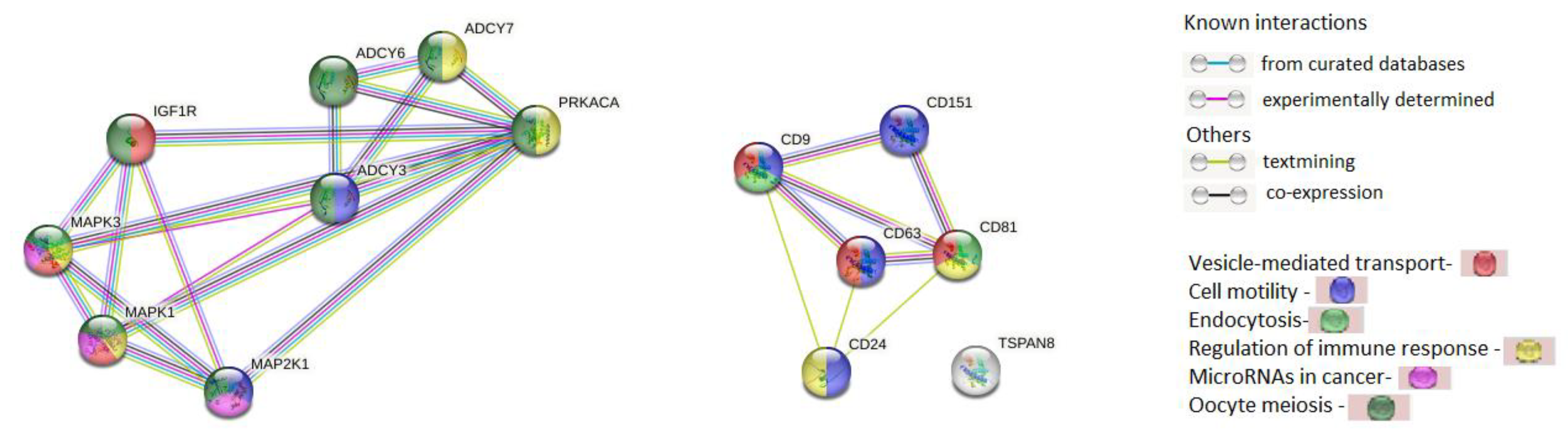
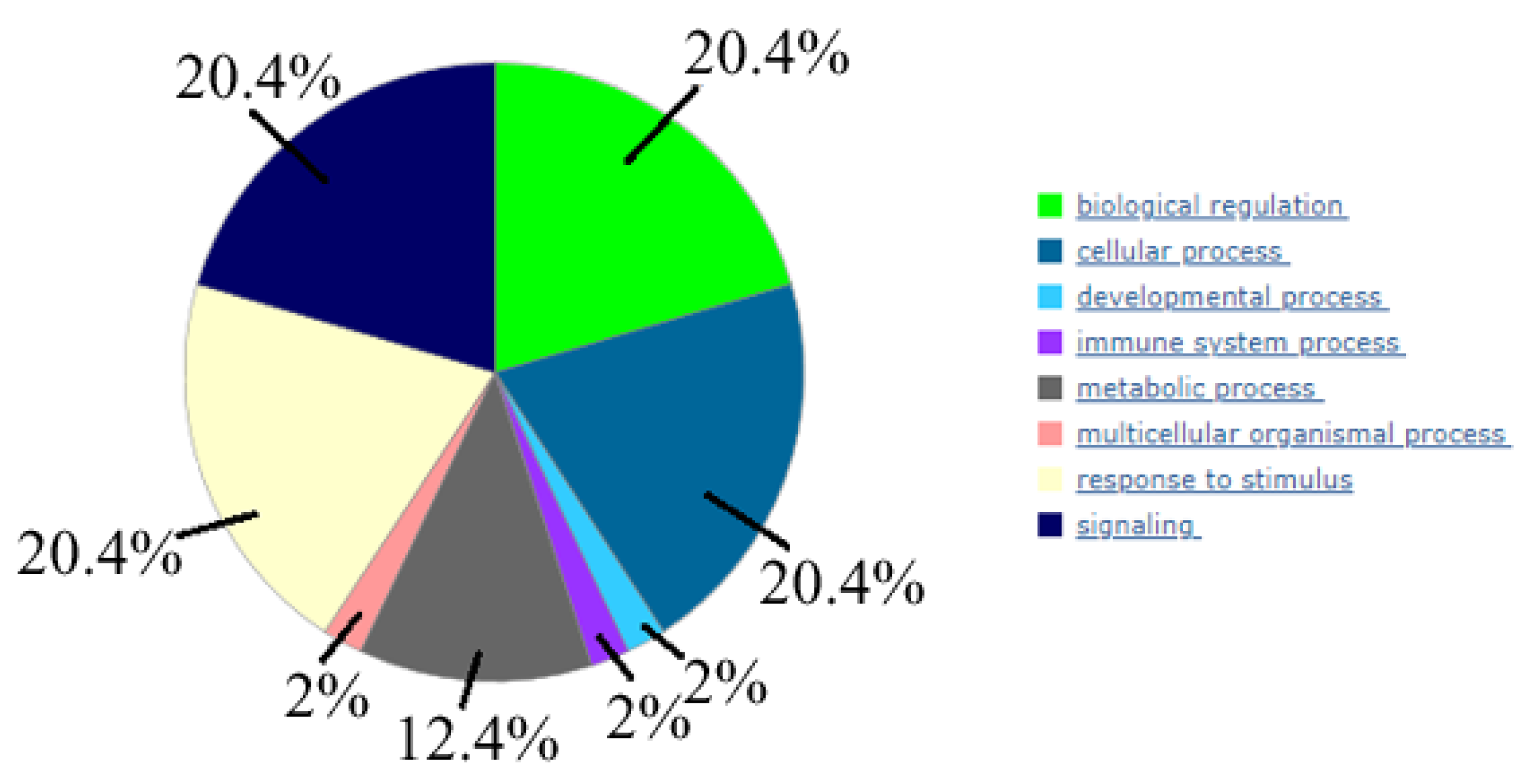
2.5. Bioinformatic Analysis
3. Discussion
4. Materials and Methods
4.1. Female Enrollment
4.2. Isolation and Characterization of Exosomes
4.3. Analysis of Tspan8/CD151 Subpopulations on the Surface of CD9-Positive Exosomes
4.4. Bioinformatic Analysis
4.5. Evaluation of miRNA Concentrations
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ding, H.; Zhang, J.; Zhang, F.; Xu, Y.; Liang, W.; Yu, Y. Nanotechnological approaches for diagnosis and treatment of ovarian cancer: A review of recent trends. Drug Deliv. 2022, 29, 3218–3232. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Guo, N. Exosomes: Potent regulators of tumor malignancy and potential bio-tools in clinical application. Crit. Rev. Oncol. Hematol. 2015, 95, 346–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunusova, N.; Kolegova, E.; Sereda, E.; Kolomiets, L.; Villert, A.; Patysheva, M.; Rekeda, I.; Grigor’eva, A.; Tarabanovskaya, N.; Kondakova, I.; et al. Plasma Exosomes of Patients with Breast and Ovarian Tumors Contain an Inactive 20S Proteasome. Molecules 2021, 26, 6965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ou, X.; Wu, X. Proteomics profiling of plasma exosomes in epithelial ovarian cancer: A potential role in the coagulation cascade, diagnosis and prognosis. Int. J. Oncol. 2019, 54, 1719–1733. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zeringer, E.; Barta, T.; Schageman, J.; Cheng, A.; Vlassov, A.V. Analysis of the RNA Content of the Exosomes Derived from Blood Serum and Urine and its Potential as Biomarkers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130502. [Google Scholar] [CrossRef]
- Tutanov, O.; Shtam, T.; Grigor’eva, A.; Tupikin, A.; Tsentalovich, Y.; Tamkovich, S. Blood Plasma Exosomes Contain Circulating DNA in Their Crown. Diagnostics 2022, 12, 854. [Google Scholar] [CrossRef]
- Yunusova, N.V.; Tugutova, E.A.; Tamkovich, S.N.; Kondakova, I.V. The role of exosomal tetraspanins and proteases in tumor progression. Biochem. Suppl. Ser. B Biomed. Chem. 2018, 12, 191–202. [Google Scholar]
- Yunusova, N.V.; Patysheva, M.R.; Molchanov, S.V.; Zambalova, E.A.; Grigor’eva, A.E.; Kolomiets, L.A.; Ochirov, M.O.; Tamkovich, S.N.; Kondakova, I.V. Metalloproteinases at the surface of small extrcellular vesicles in advanced ovarian cancer: Relationships with ascites volume and peritoneal canceromatosis index. Clin. Chim. Acta 2019, 494, 116–122. [Google Scholar] [CrossRef]
- Kumari, S.; Gayatri Devi, V.; Badana, A.; Dasari, V.R.; Malla, R.R. CD151—A striking marker for cancer therapy. Biomark. Cancer 2015, 7, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Medrano, M.; Communal, L.; Brown, K.R.; Iwanicki, M.; Normand, J.; Paterson, J.; Sircoulomb, F.; Krzyzanowski, P.; Novak, M.; Doodnauth, S.A.; et al. Interrogation of functional cell-surface markers identifies CD151 dependency in high-grade serous ovarian cancer. Cell Rep. 2017, 18, 2343–2358. [Google Scholar] [CrossRef] [Green Version]
- Mosig, R.A.; Lin, L.; Senturk, E.; Shah, H.; Huang, F.; Schlosshauer, P.; Cohen, S.; Fruscio, R.; Marchini, S.; D’Incalci, M.; et al. Application of RNA-seq transcriptome analysis: CD151 is an invasion/migration target in all stages of epithelial ovarian cancer. J. Ovarian Res. 2012, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Llorente, A. Proteomic analysis of microvesicles released by the Human Prostate Cancer Cell Line PC-3. Mol. Cell. Proteom. 2012, 11, M111.012914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemler, M.E. Tetraspanin proteins promote multiple cancer stages. Nat. Rev. Cancer 2014, 14, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Ailane, N.; Greco, C.; Zhu, Y.; Sala-Valdes, M.; Billard, M.; Casal, I.; Bawa, O.; Opolon, P.; Rubinstein, E.; Boucheix, C. Effect of an anti-human co-029/TSPAN8 mouse monoclonal antibody on tumor growth in a nude mouse model. Front. Physiol. 2014, 5, 364. [Google Scholar] [CrossRef] [Green Version]
- Park, C.S.; Kim, T.-K.; Kim, H.G.; Kim, Y.-J.; Jeoung, M.H.; Lee, W.R.; Go, N.K.; Heo, K.; Lee, S. Therapeutic targeting of Tetraspanin8 in epithelial ovarian cancer invasion and metastasis. Oncogene 2016, 35, 4540–4548. [Google Scholar] [CrossRef]
- Joyce, D.P.; Kerin, M.J.; Dwyer, R.M. Exosome-encapsulated micrornas as circulating biomarkers for breast cancer. Int. J. Cancer. 2016, 139, 1443–1448. [Google Scholar] [CrossRef]
- Kalra, H.; Drummen, G.P.C.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Sun, Y.; Hu, L.; Zheng, H.; Ji, P.; Pecot, C.V.; Zhao, Y.; Reynolds, S.; Cheng, H.; Rupaimoole, R.; et al. Integrated analyses identify a master MicroRNA regulatory network for the mesenchymal subtype in serous ovarian cancer. Cancer Cell 2013, 23, 186–199. [Google Scholar] [CrossRef] [Green Version]
- Braga, E.A.; Fridman, M.V.; Kushlinskii, N.E. Molecular mechanisms of ovarian carcinoma metastasis: Key genes and regulatory micrornas. Biochemistry 2017, 82, 529–541. [Google Scholar] [CrossRef]
- Liu, W.; Wang, S.; Zhou, S.; Yang, F.; Jiang, W.; Zhang, Q.; Wang, L. A systems biology approach to identify microRNAs contributing to cisplatin resistance in human ovarian cancer cells. Mol. Biosyst. 2017, 13, 2268–2276. [Google Scholar] [CrossRef]
- Yue, S.; Mu, W.; Zöller, M. Tspan8 and CD151 promote metastasis by distinct mechanisms. Eur. J. Cancer 2013, 49, 2934–2948. [Google Scholar] [CrossRef] [PubMed]
- Hagrass, H.A.; Sharaf, S.; Pasha, H.F.; Tantawy, E.A.; Mohamed, R.H.; Kassem, R. Circulating microRNAs-a new horizon in molecular diagnosis of breast cancer. Genes Cancer 2015, 6, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryzgunova, O.E.; Zaripov, M.M.; Skvortsova, T.E.; Lekchnov, E.A.; Grigor’eva, A.E.; Zaporozhchenko, I.A.; Morozkin, E.S.; Ryabchikova, E.I.; Yurchenko, Y.B.; Voitsitskiy, V.E.; et al. Comparative study of extracellular vesicles from the urine of healthy individuals and prostate cancer patients. PLoS ONE 2016, 11, e0157566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, A.M.; Kerin, M.J.; Miller, N. Identification and validation of miRNAs as endogenous controls for RQ-PCR in blood specimens for breast cancer studies. PLoS ONE 2013, 8, e83718. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Kano, M.; Murakami, K.; Toyozumi, T.; Suito, H.; Takahashi, M.; Sekino, N.; Shiraishi, T.; Kamata, T.; Ryuzaki, T.; et al. Tumor-derived exosomes influence the cell cycle and cell migration of human esophageal cancer cell lines. Cancer Sci. 2020, 111, 4348–4358. [Google Scholar] [CrossRef]
- Pathania, A.S.; Prathipati, P.; Challagundla, K.B. New insights into exosome mediated tumor-immune escape: Clinical Perspectives and Therapeutic Strategies. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188624. [Google Scholar] [CrossRef]
- Paskeh, M.D.A.; Entezari, M.; Mirzaei, S.; Zabolian, A.; Saleki, H.; Naghdi, M.J.; Sabet, S.; Khoshbakht, M.A.; Hashemi, M.; Hushmandi, K.; et al. Emerging role of exosomes in cancer progression and tumor microenvironment remodeling. J. Hematol. Oncol. 2022, 15, 83. [Google Scholar] [CrossRef]
- Sahebi, R.; Langari, H.; Fathinezhad, Z.; Bahari Sani, Z.; Avan, A.; Ghayour Mobarhan, M.; Rezayi, M. Exosomes: New insights into cancer mechanisms. J. Cell Biochem. 2019, 121, 7–16. [Google Scholar] [CrossRef]
- Yi, Y.; Wu, M.; Zeng, H.; Hu, W.; Zhao, C.; Xiong, M.; Lv, W.; Deng, P.; Zhang, Q.; Wu, Y. Tumor-derived exosomal non-coding RNAS: The emerging mechanisms and potential clinical applications in breast cancer. Front. Oncol. 2021, 11, 738945. [Google Scholar] [CrossRef]
- Deb, A.; Gupta, S.; Mazumder, P.B. Exosomes: A New Horizon in modern medicine. Life Sci. 2021, 264, 118623. [Google Scholar] [CrossRef]
- Zheng, H.B.; Zheng, X.G.; Liu, B.P. miRNA-101 inhibits ovarian cancer cells proliferation and invasion by down-regulating expression of SOCS-2. Int. J. Clin. Exp. Med. 2015, 8, 20263–20270. [Google Scholar] [PubMed]
- Liang, H.; Yu, T.; Han, Y.; Jiang, H.; Wang, C.; You, T.; Zhao, X.; Shan, H.; Yang, R.; Yang, L.; et al. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding mir-101-3p to regulate zeb1 expression. Mol. Cancer 2018, 17, 119. [Google Scholar] [PubMed]
- Xu, Y.; Xu, L.; Zheng, J.; Geng, L.; Zhao, S. Mir-101 inhibits ovarian carcinogenesis by repressing the expression of brain-derived neurotrophic factor. FEBS Open Bio 2017, 7, 1258–1266. [Google Scholar] [PubMed]
- Liu, L.; Guo, J.; Yu, L.; Cai, J.; Gui, T.; Tang, H.; Song, L.; Wang, J.; Han, F.; Yang, C.; et al. Mir-101 regulates expression of EZH2 and contributes to progression of and cisplatin resistance in epithelial ovarian cancer. Tumor Biol. 2014, 35, 12619–12626. [Google Scholar] [CrossRef]
- Wang, H.; Chen, C.; Ding, K.; Zhang, W.; Hou, J. Mir-24-3P as a prognostic indicator for multiple cancers: From a meta-analysis view. Biosci. Rep. 2020, 40, BSR20202938. [Google Scholar] [CrossRef]
- Yuan, D.; Cui, X.; Wang, Y.; Zhao, Y.; Li, H.; Hu, S.; Chu, X.; Li, Y.; Li, Q.; Liu, Q.; et al. Enrichment Analysis identifies functional microRNA-disease associations in humans. PLoS ONE 2015, 10, e0136285. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Xu, X.; Chen, K.; Fu, Z.; Wang, S.; Chen, Y.; Zhang, H.; Niu, Y.; Chen, H.; Yu, H.; et al. LncRNA Casc15, Mir-23b cluster and smad3 form a novel positive feedback loop to promote epithelial-mesenchymal transition and metastasis in ovarian cancer. Int. J. Biol. Sci. 2022, 18, 1989–2002. [Google Scholar]
- Sun, H.; Rana, S.; Wang, Z.; Zhao, K.; Schnölzer, M.; Provaznik, J.; Hackert, T.; Lv, Q.; Zöller, M. The pancreatic cancer-initiating cell marker CD44v6 affects transcription, translation, and signaling: Consequences for exosome composition and delivery. J. Oncol. 2019, 2019, 3516973. [Google Scholar] [CrossRef] [Green Version]
- Konoshenko, M.; Sagaradze, G.; Orlova, E.; Shtam, T.; Proskura, K.; Kamyshinsky, R.; Yunusova, N.; Alexandrova, A.; Efimenko, A.; Tamkovich, S. Total blood exosomes in breast cancer: Potential role in crucial steps of tumorigenesis. Int. J. Biol. Sci. 2020, 21, 7341. [Google Scholar]
- Konoshenko, M.Y.; Bryzgunova, O.E.; Lekchnov, E.A.; Amelina, E.V.; Yarmoschuk, S.V.; Pak, S.V.; Laktionov, P.P. The influence of radical prostatectomy on the expression of cell-free miRNA. Diagnostics 2020, 10, 600. [Google Scholar]
- Tamkovich, S.; Tutanov, O.; Efimenko, A.; Grigor’eva, A.; Ryabchikova, E.; Kirushina, N.; Vlassov, V.; Tkachuk, V.; Laktionov, P. Blood circulating exosomes contain distinguishable fractions of free and cell-surface-associated vesicles. Curr. Mol. Med. 2019, 19, 273–285. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subpopulations | HFs (n = 19) | OC Patients (n = 20) | p-Level |
|---|---|---|---|
| Plasma exosomes | |||
| Tspan8+ | 6.9 (5.47–8.96) | 11.02 (7.66–15.6) | 0.046 |
| CD151+ | 1.88 (1.63–2.82) | 3.91 (2.99–5.02) | 0.011 |
| CD151+/Tspan8+ | 1.59 (1.31–2.29) | 3.22 (2.76–4.80) | 0.006 |
| Ascites exosomes | |||
| Tspan8+ | 7.31 (5.40–9.10) | ||
| CD151+ | 3.40 (2.45–4.96) | ||
| CD151+/Tspan8+ | 2.71 (1.77–3.82) |
| OC Patients, n = 20 | |
|---|---|
| Histology | |
| Serous | 20 (100%) |
| FIGO (2013) staging | |
| IIB | 2 (10%) |
| IIIB | 3 (15%) |
| IIIC | 15 (75%) |
| Tumor Grade | |
| High-grade | 20 (100%) |
| Ascites volume | |
| <200 mL | 5 (25%) |
| 200–1000 mL | 5 (25%) |
| >1000 mL | 10 (50%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunusova, N.; Dzhugashvili, E.; Yalovaya, A.; Kolomiets, L.; Shefer, A.; Grigor’eva, A.; Tupikin, A.; Kondakova, I.; Tamkovich, S. Comparative Analysis of Tumor-Associated microRNAs and Tetraspanines from Exosomes of Plasma and Ascitic Fluids of Ovarian Cancer Patients. Int. J. Mol. Sci. 2023, 24, 464. https://doi.org/10.3390/ijms24010464
Yunusova N, Dzhugashvili E, Yalovaya A, Kolomiets L, Shefer A, Grigor’eva A, Tupikin A, Kondakova I, Tamkovich S. Comparative Analysis of Tumor-Associated microRNAs and Tetraspanines from Exosomes of Plasma and Ascitic Fluids of Ovarian Cancer Patients. International Journal of Molecular Sciences. 2023; 24(1):464. https://doi.org/10.3390/ijms24010464
Chicago/Turabian StyleYunusova, Natalia, Ekaterina Dzhugashvili, Alena Yalovaya, Larisa Kolomiets, Aleksei Shefer, Alina Grigor’eva, Alexey Tupikin, Irina Kondakova, and Svetlana Tamkovich. 2023. "Comparative Analysis of Tumor-Associated microRNAs and Tetraspanines from Exosomes of Plasma and Ascitic Fluids of Ovarian Cancer Patients" International Journal of Molecular Sciences 24, no. 1: 464. https://doi.org/10.3390/ijms24010464