
ATP-Binding Cassette Protein ABCC10 Deficiency Prevents Diet-Induced Obesity but Not Atherosclerosis in Mice
 , , and
, , and 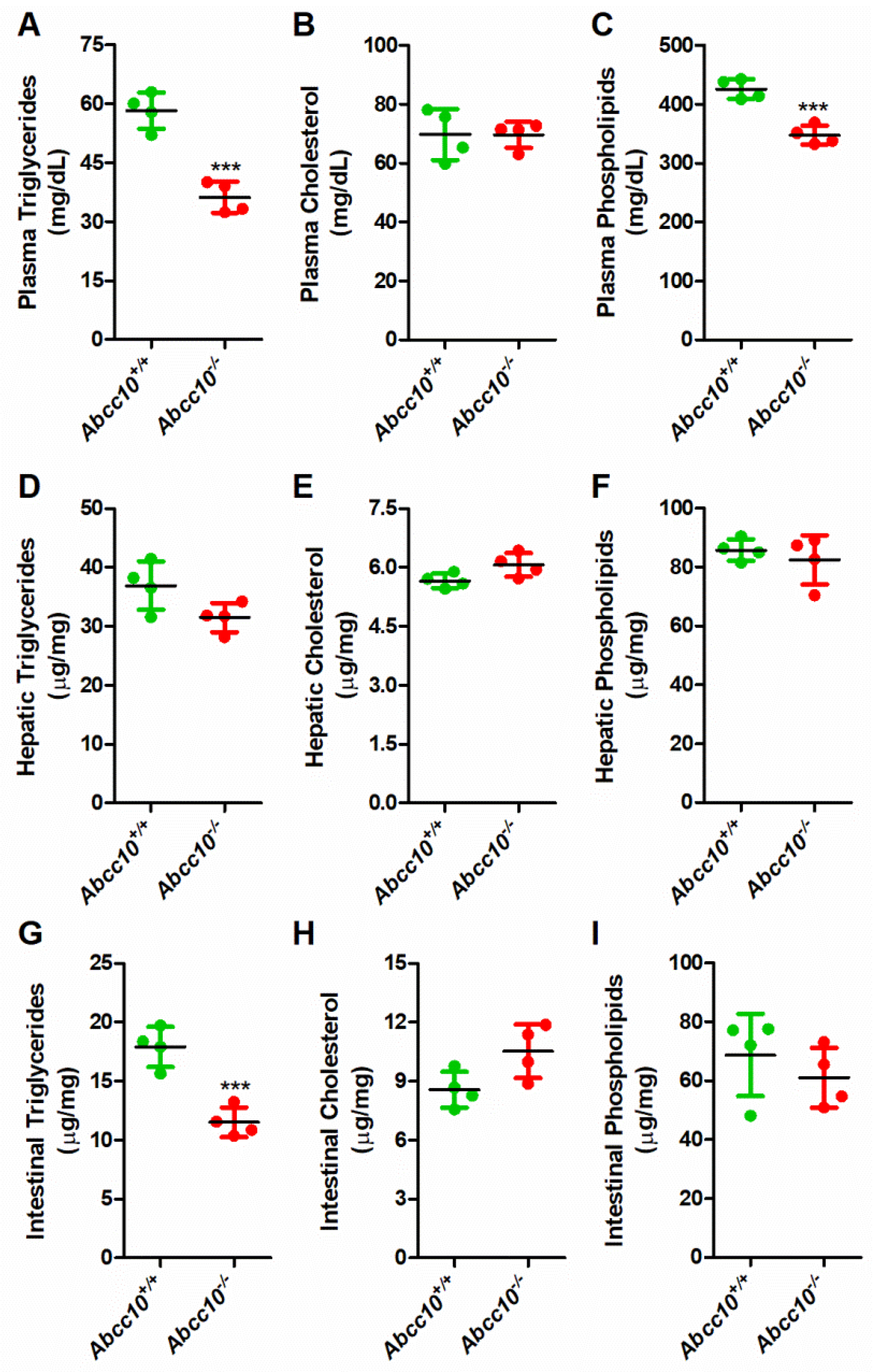{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Deletion of ABCC10 in Chow-Fed Mice Decreases Plasma and Tissue Triglycerides
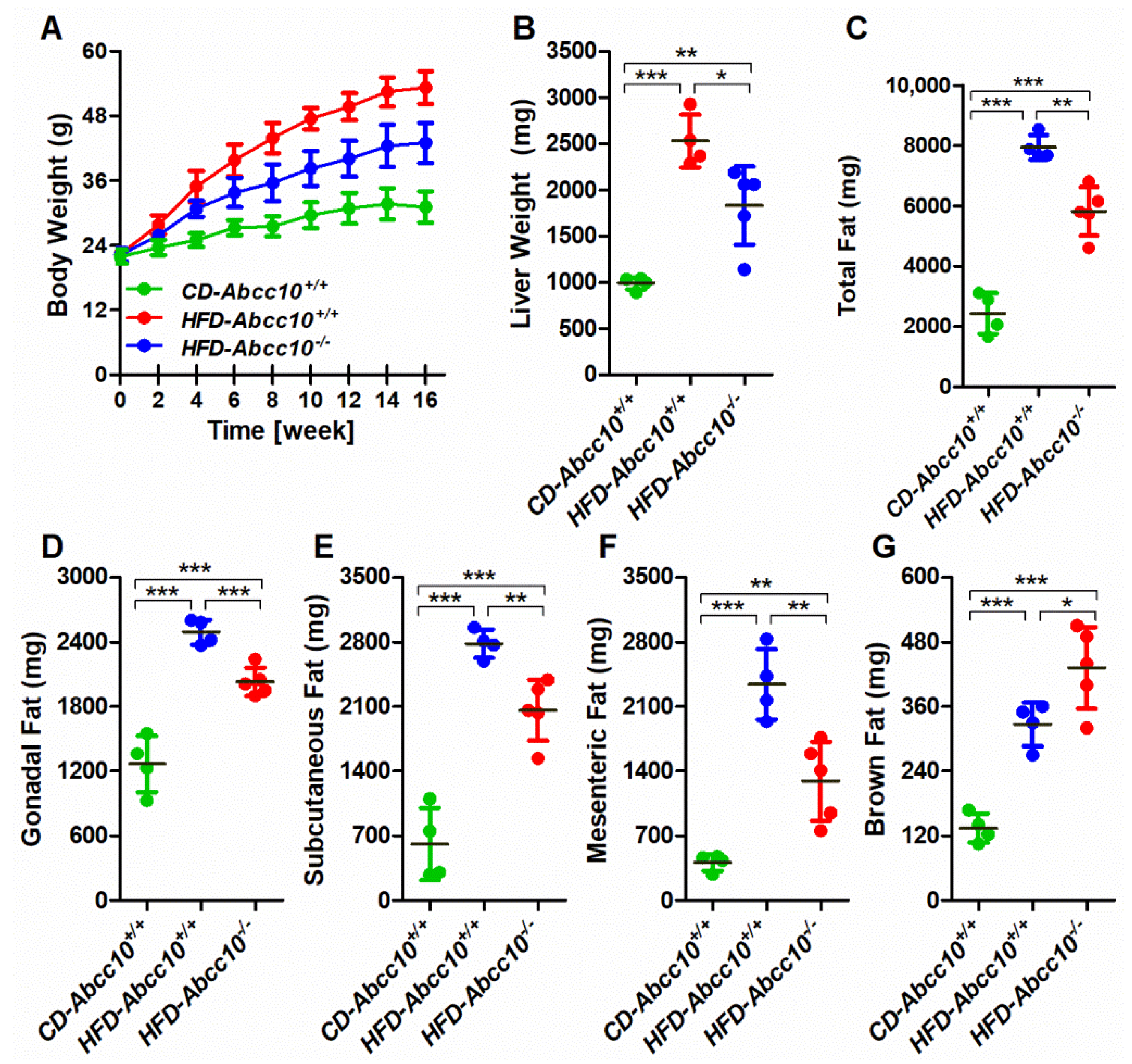
2.2. Deletion of ABCC10 Prevents Diet-Induced Obesity in Mice
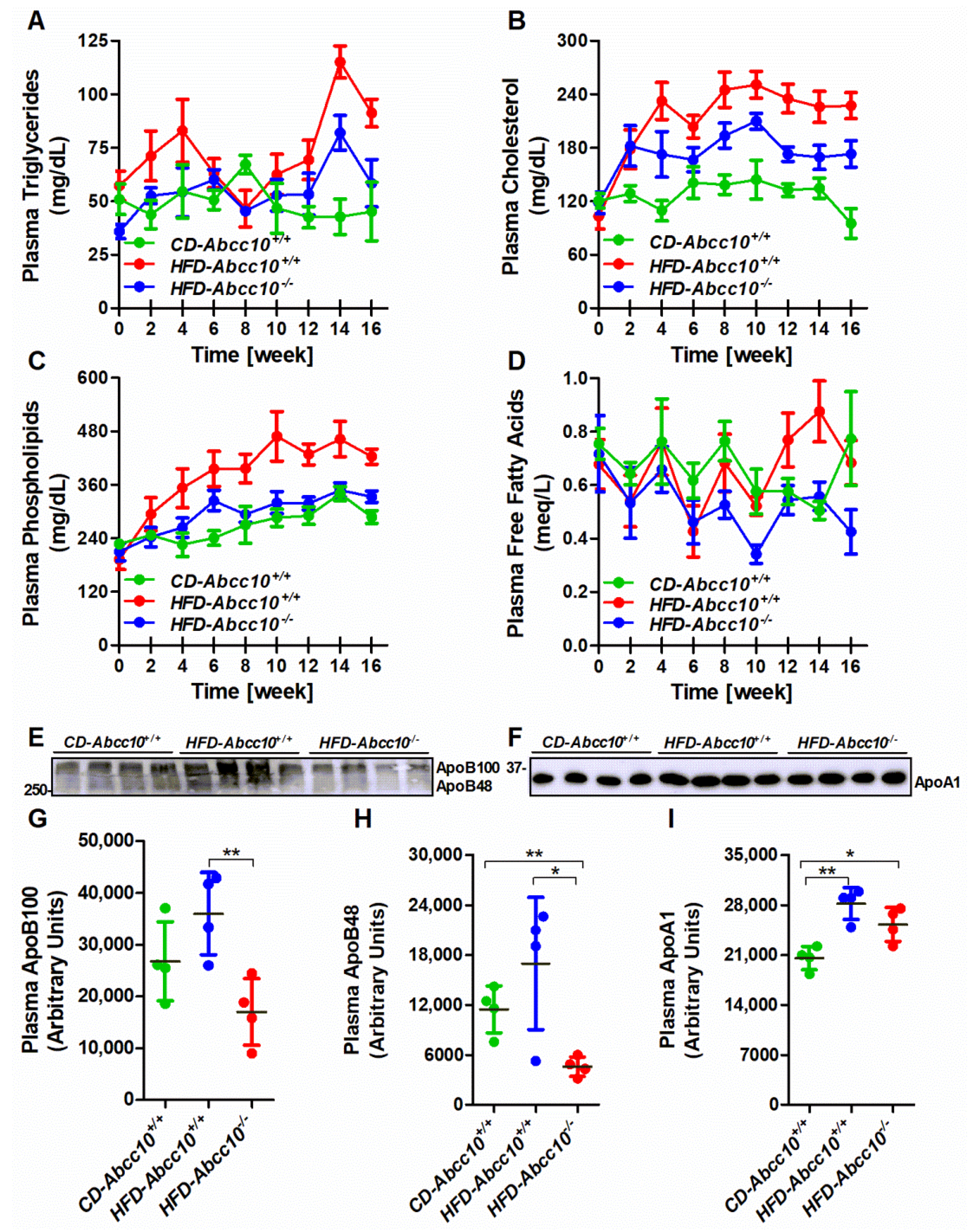
2.3. Plasma Lipid and ApoB Lipoprotein Levels Are Lower in High-Fat-Diet-Fed ABCC10-Deficient Mice
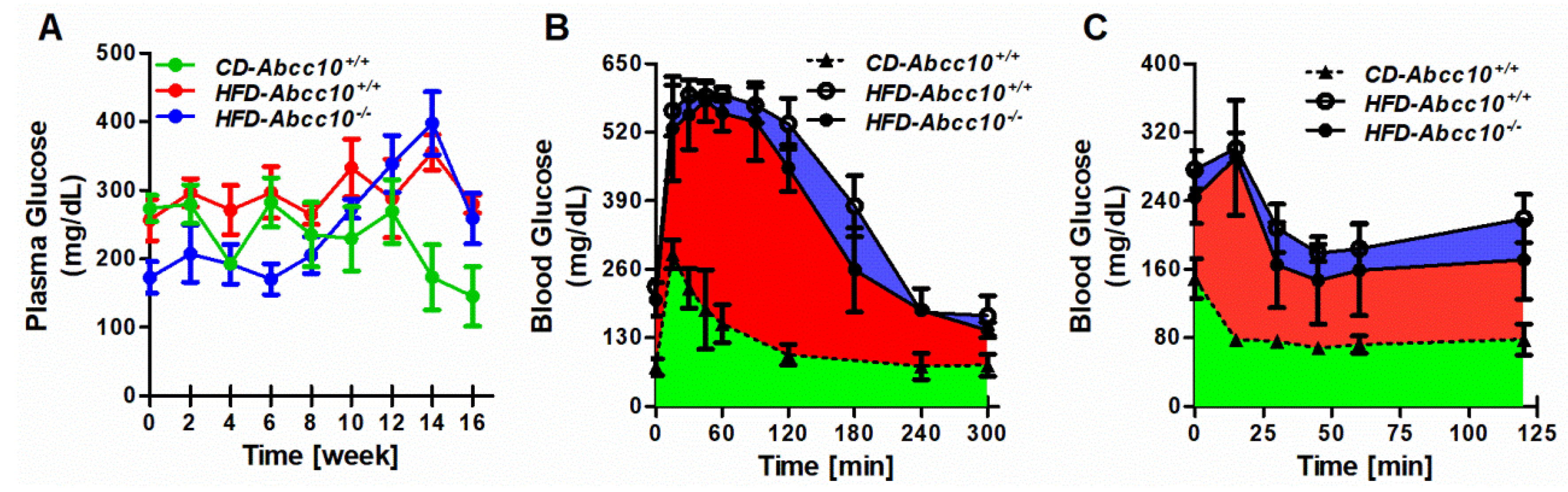
2.4. Plasma Glucose Levels Are Lower during Early Stages of High-Fat-Diet Feeding in ABCC10-Deficient Mice
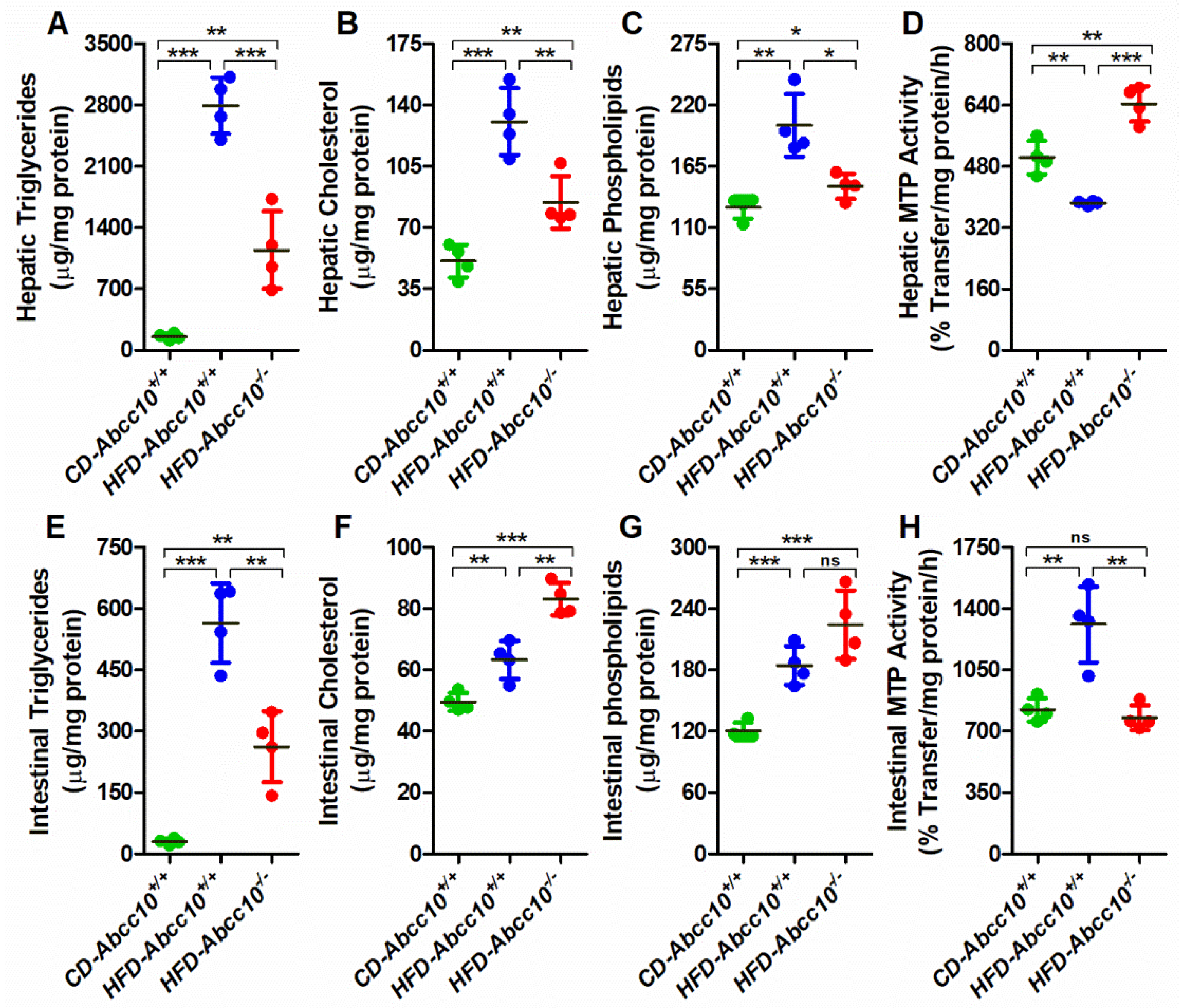
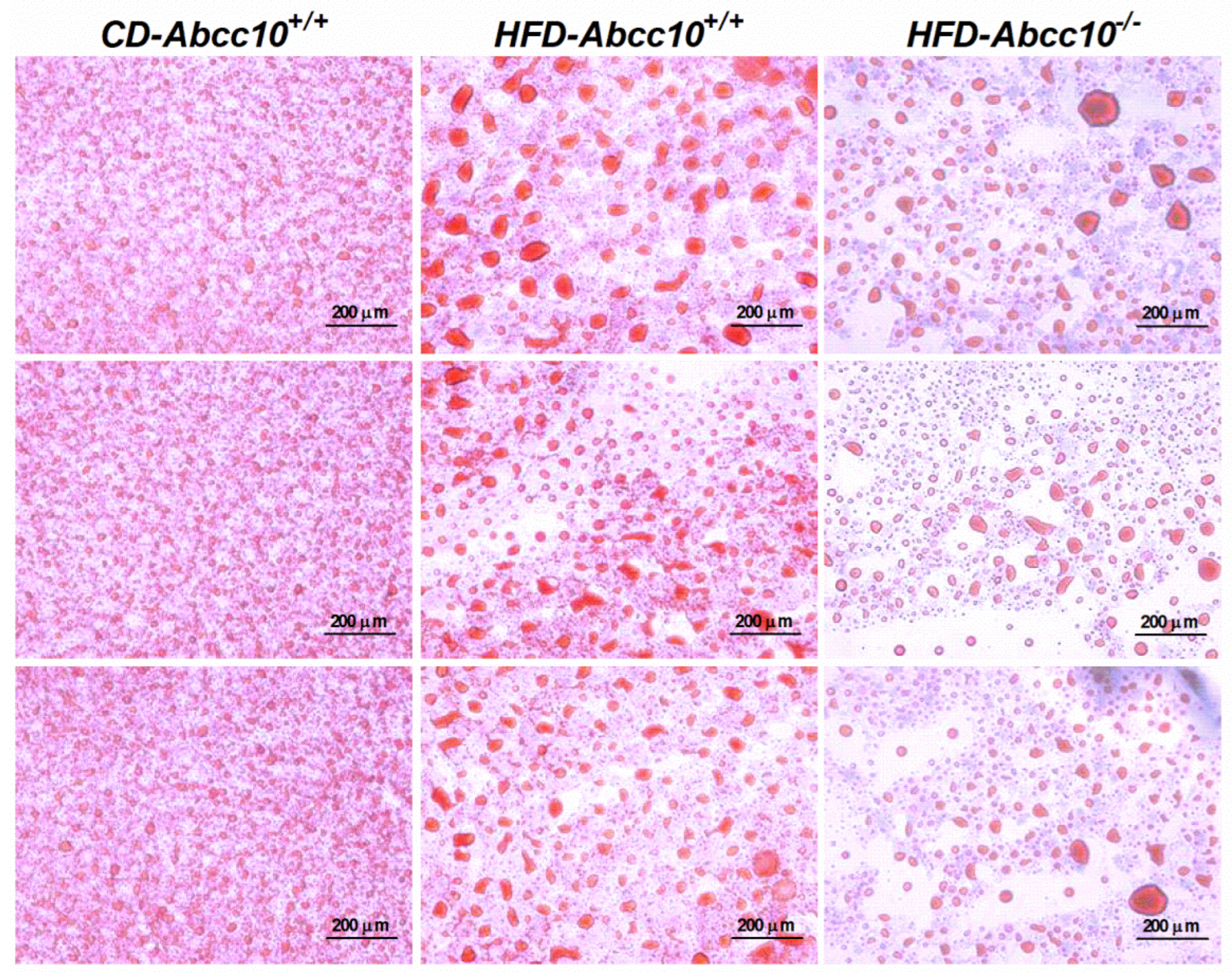
2.5. ABCC10 Deficiency Prevents Accumulation of Triglycerides in the Tissues
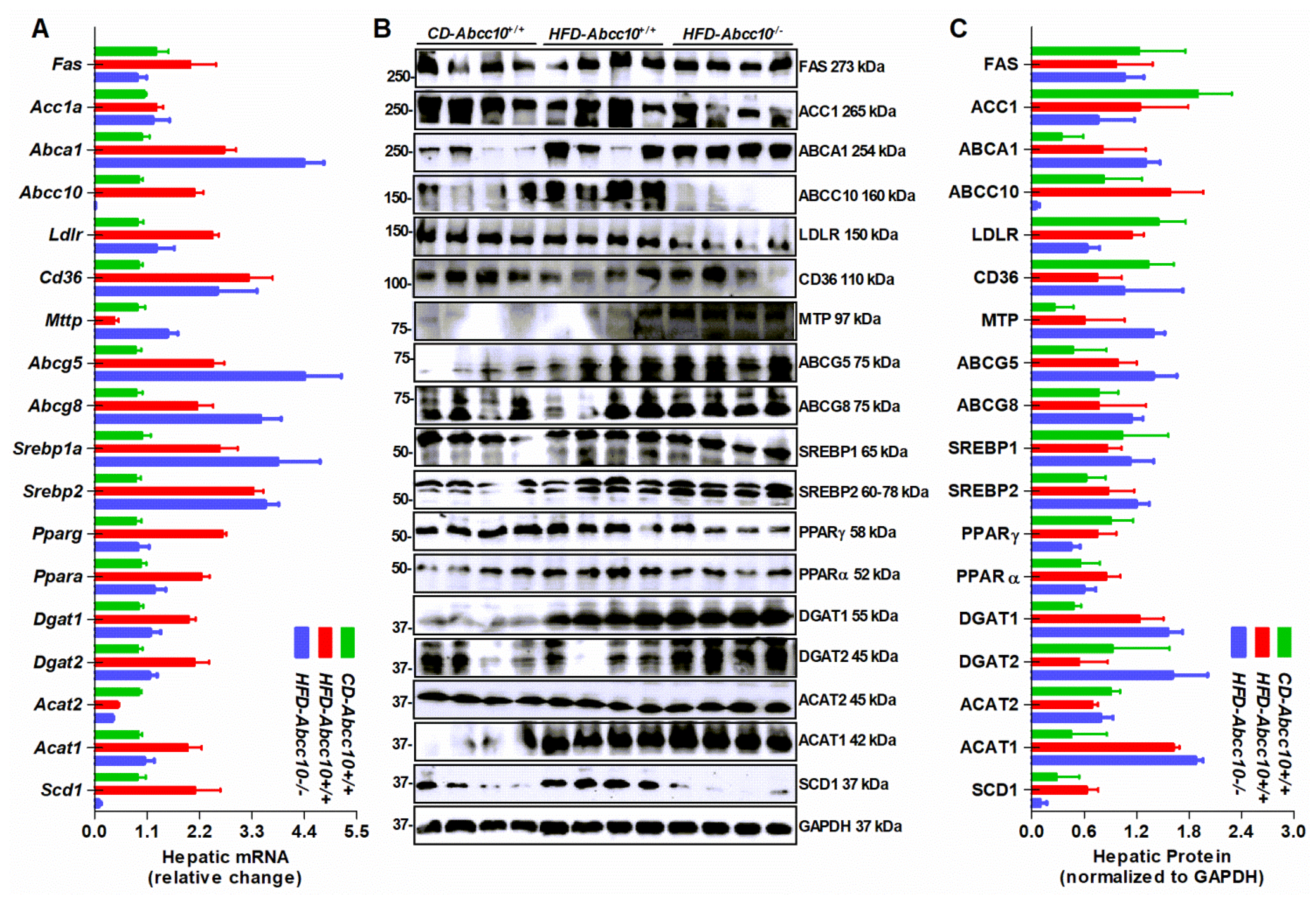
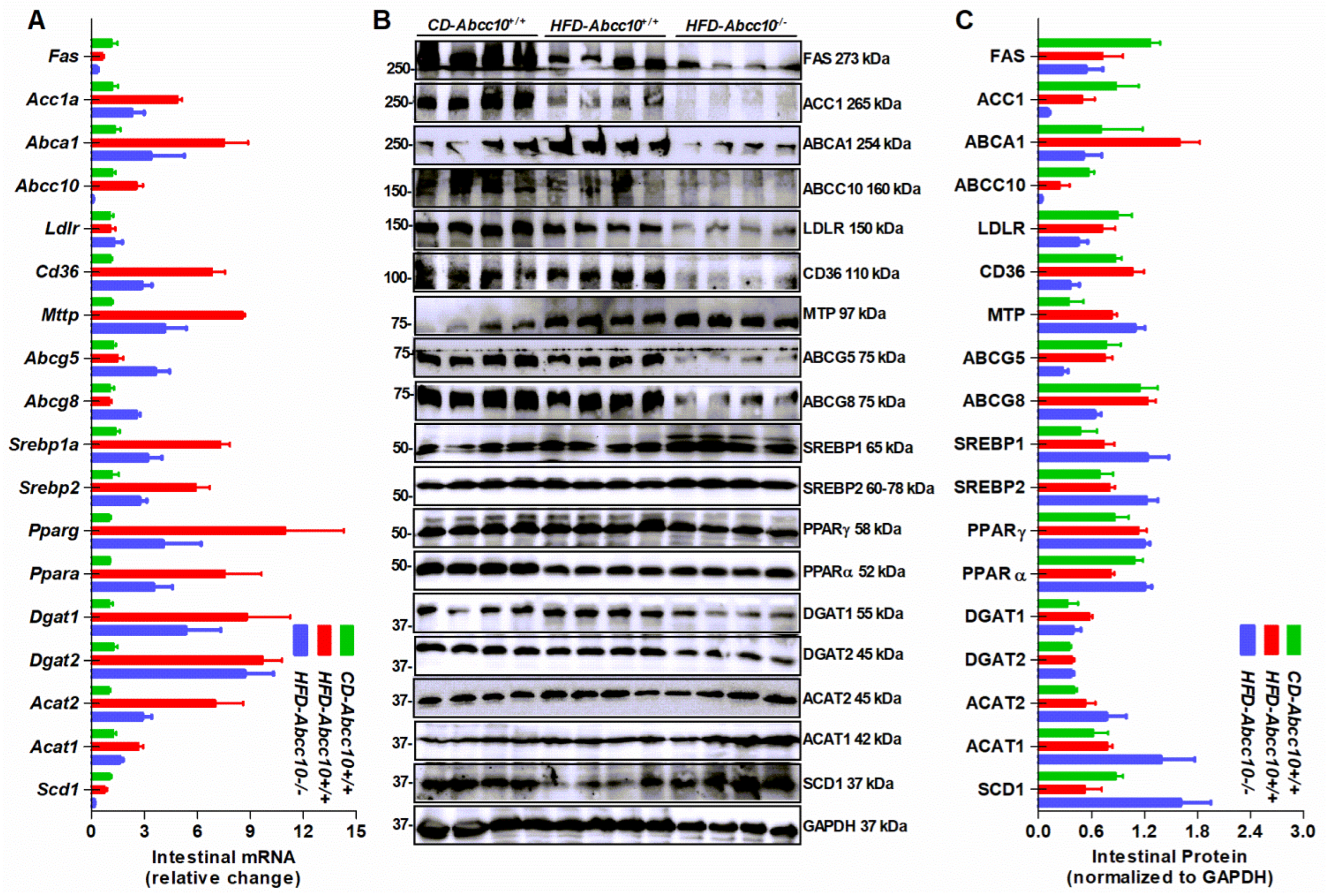
2.6. Deletion of ABCC10 in Mice Alters Expression of Several Lipid Metabolism Genes in the Liver and the Intestine
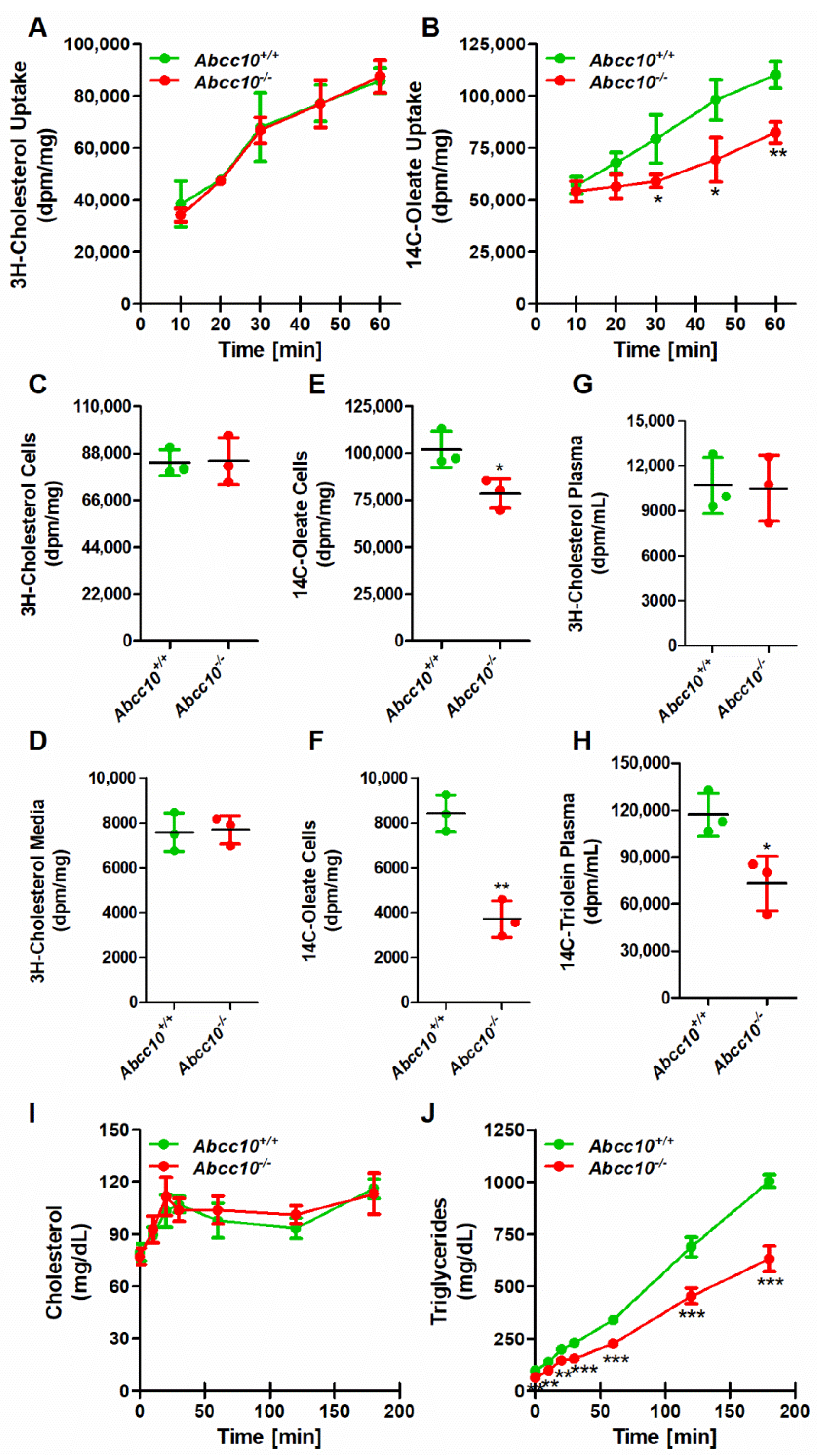
2.7. Deletion of ABCC10 in Mice Affects Absorption of Triglycerides in the Intestine
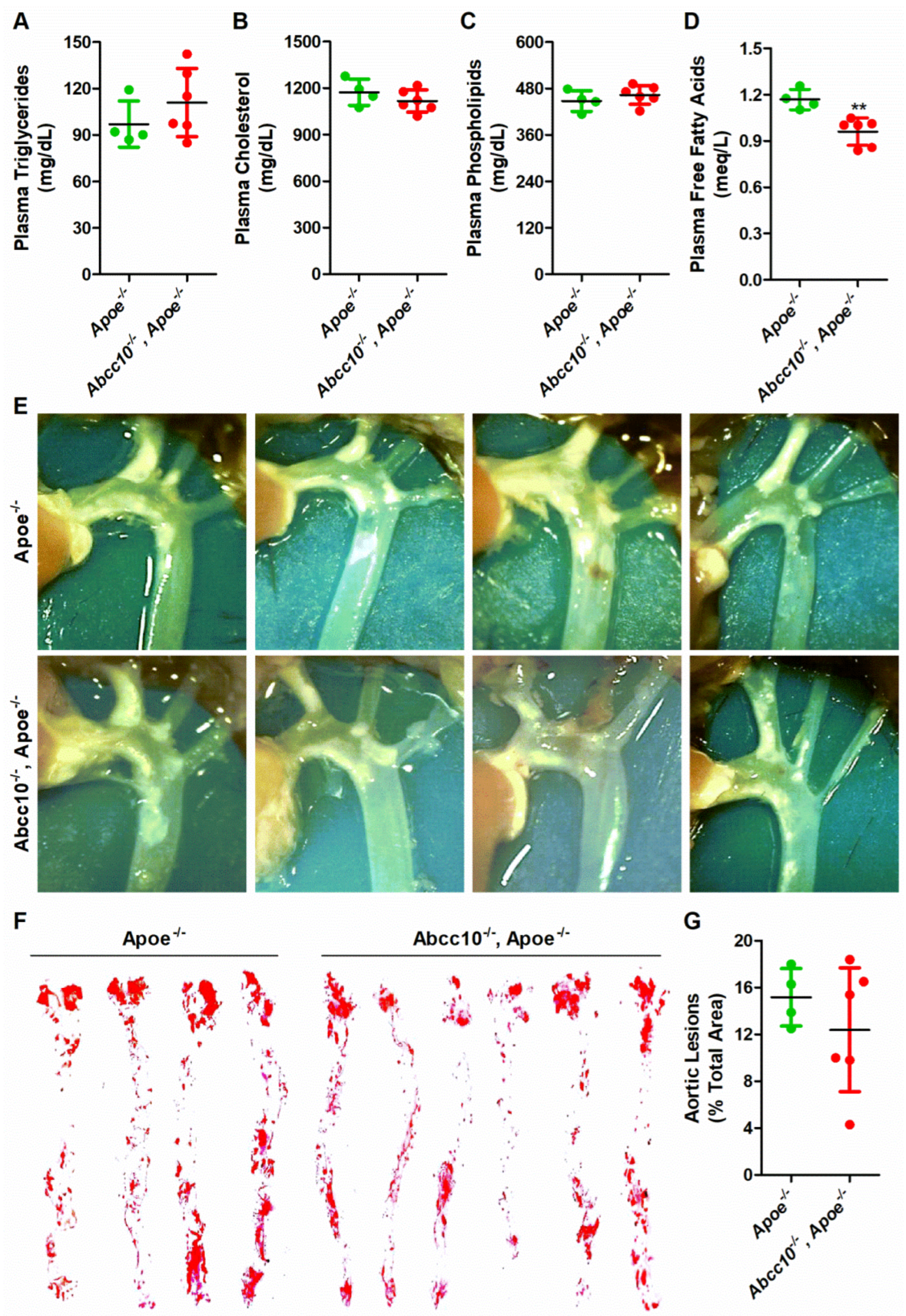
2.8. ABCC10 Deficiency Does Not Affect Atherosclerosis in Western-Type-Diet-Fed ApoE Knockout Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Diets
4.3. Lipid Measurements and Oil Red O Staining
4.4. Glucose Analysis, Glucose and Insulin Tolerance Tests
4.5. Uptake and Secretion of Lipids by Primary Enterocytes
4.6. Short-Term Lipid Absorption Studies
4.7. Hepatic Lipid Mobilization and Metabolic Studies
4.8. Determination of MTP Activity
4.9. Western Blot Analysis
4.10. mRNA Quantification
4.11. Mouse Atherosclerotic Lesion Measurement
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, M.M.; Shi, J.; Dreizen, P. Microsomal triglyceride transfer protein and its role in apoB-lipoprotein assembly. J. Lipid Res. 2003, 44, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Hussain, M.M. Intestinal lipid absorption. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1183–E1194. [Google Scholar] [CrossRef] [Green Version]
- Packard, C.J.; Boren, J.; Taskinen, M.R. Causes and Consequences of Hypertriglyceridemia. Front. Endocrinol. 2020, 11, 252. [Google Scholar] [CrossRef]
- Nissen, S.E.; Nicholls, S.J.; Sipahi, I.; Libby, P.; Raichlen, J.S.; Ballantyne, C.M.; Davignon, J.; Erbel, R.; Fruchart, J.C.; Tardif, J.C.; et al. Effect of very high-intensity statin therapy on regression of coronary atherosclerosis: The ASTEROID trial. JAMA 2006, 295, 1556–1565. [Google Scholar] [CrossRef] [Green Version]
- Boekholdt, S.M.; Hovingh, G.K.; Mora, S.; Arsenault, B.J.; Amarenco, P.; Pedersen, T.R.; LaRosa, J.C.; Waters, D.D.; DeMicco, D.A.; Simes, R.J.; et al. Very low levels of atherogenic lipoproteins and the risk for cardiovascular events: A meta-analysis of statin trials. J. Am. Coll. Cardiol. 2014, 64, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Taskinen, M.R. Diabetic dyslipidaemia: From basic research to clinical practice. Diabetologia 2003, 46, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Adiels, M.; Olofsson, S.O.; Taskinen, M.R.; Boren, J. Diabetic dyslipidaemia. Curr. Opin. Lipidol. 2006, 17, 238–246. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Zhang, Y.L.; Hernandez-Ono, A. Metabolic syndrome: Focus on dyslipidemia. Obesity 2006, 14 (Suppl. S1), 41S–49S. [Google Scholar] [CrossRef]
- Chan, D.C.; Watts, G.F.; Redgrave, T.G.; Mori, T.A.; Barrett, P.H. Apolipoprotein B-100 kinetics in visceral obesity: Associations with plasma apolipoprotein C-III concentration. Metabolism 2002, 51, 1041–1046. [Google Scholar] [CrossRef]
- Veilleux, A.; Grenier, E.; Marceau, P.; Carpentier, A.C.; Richard, D.; Levy, E. Intestinal lipid handling: Evidence and implication of insulin signaling abnormalities in human obese subjects. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 644–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dassa, E.; Bouige, P. The ABC of ABCS: A phylogenetic and functional classification of ABC systems in living organisms. Res. Microbiol. 2001, 152, 211–229. [Google Scholar] [CrossRef]
- Kimura, Y.; Morita, S.Y.; Matsuo, M.; Ueda, K. Mechanism of multidrug recognition by MDR1/ABCB1. Cancer Sci. 2007, 98, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, H.; Kimura, Y.; Kioka, N.; Matsuo, M.; Ueda, K. ATPase activity of human ABCG1 is stimulated by cholesterol and sphingomyelin. J. Lipid Res. 2013, 54, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Sharom, F.J. ABC multidrug transporters: Structure, function and role in chemoresistance. Pharmacogenomics 2008, 9, 105–127. [Google Scholar] [CrossRef]
- Schmitz, G.; Langmann, T. Structure, function and regulation of the ABC1 gene product. Curr. Opin. Lipidol. 2001, 12, 129–140. [Google Scholar] [CrossRef]
- Francis, G.A.; Knopp, R.H.; Oram, J.F. Defective removal of cellular cholesterol and phospholipids by apolipoprotein A-I in Tangier Disease. J. Clin. Investig. 1995, 96, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Cuchel, M.; Tarugi, P.; Hegele, R.A.; Davidson, N.O.; Rader, D.J.; Klein, R.L.; Hussain, M.M. Microsomal triglycerdie transfer protein transfers and determines plasma concentrations of ceramide and sphingomyelin but not glycosylceramide. J. Biol. Chem. 2015, 290, 25863–25875. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Suarez, M.D.; Yadav, P.K.; Walsh, M.T.; Li, Y.; Wu, Y.; Huang, Z.; James, A.W.; Escobar, V.; Mokbe, A.; et al. ATP-binding cassette protein ABCA7 deficiency impairs sphingomyelin synthesis, cognitive discrimination, and synaptic plasticity in the entorhinal cortex. J. Biol. Chem. 2022, 2022, 102411. [Google Scholar] [CrossRef]
- Raggers, R.J.; van Helvoort, A.; Evers, R.; van Meer, G. The human multidrug resistance protein MRP1 translocates sphingolipid analogs across the plasma membrane. J. Cell Sci. 1999, 112 Pt 3, 415–422. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Kohyama-Koganeya, A.; Hirabayashi, Y. New insights on glucosylated lipids: Metabolism and functions. Biochim. Biophys. Acta 2013, 1831, 1475–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budani, M.; Auray-Blais, C.; Lingwood, C. ATP-binding cassette transporters mediate differential biosynthesis of glycosphingolipid species. J. Lipid Res. 2021, 62, 100128. [Google Scholar] [CrossRef] [PubMed]
- Kathawala, R.J.; Wang, Y.J.; Ashby, C.R., Jr.; Chen, Z.S. Recent advances regarding the role of ABC subfamily C member 10 (ABCC10) in the efflux of antitumor drugs. Chin. J. Cancer 2014, 33, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarling, E.J.; de Aguiar Vallim, T.Q.; Edwards, P.A. Role of ABC transporters in lipid transport and human disease. Trends Endocrinol. Metab. 2013, 24, 342–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerterp, M.; Bochem, A.E.; Yvan-Charvet, L.; Murphy, A.J.; Wang, N.; Tall, A.R. ATP-binding cassette transporters, atherosclerosis, and inflammation. Circ. Res. 2014, 114, 157–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahnke, J.; Langer, O.; Krohn, M. Alzheimer’s and ABC transporters—New opportunities for diagnostics and treatment. Neurobiol. Dis. 2014, 72 Pt A, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Kotlyarov, S.; Kotlyarova, A. Analysis of ABC Transporter Gene Expression in Atherosclerosis. Cardiogenetics 2021, 11, 204–218. [Google Scholar] [CrossRef]
- Surwit, R.S.; Kuhn, C.M.; Cochrane, C.; McCubbin, J.A.; Feinglos, M.N. Diet-induced type II diabetes in C57BL/6J mice. Diabetes 1988, 37, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [Green Version]
- Bakillah, A.; Hussain, M.M. Mice subjected to aP2-Cre mediated ablation of microsomal triglyceride transfer protein are resistant to high fat diet induced obesity. Nutr. Metab. 2016, 13, 1. [Google Scholar] [CrossRef]
- Iqbal, J.; Dai, K.; Seimon, T.; Jungreis, R.; Oyadomari, M.; Kuriakose, G.; Ron, D.; Tabas, I.; Hussain, M.M. IRE1beta inhibits chylomicron production by selectively degrading MTP mRNA. Cell Metab. 2008, 7, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Newberry, E.P.; Young, S.G.; Robine, S.; Hamilton, R.L.; Wong, J.S.; Luo, J.; Kennedy, S.; Davidson, N.O. Compensatory increase in hepatic lipogenesis in mice with conditional intestine-specific Mttp deficiency. J. Biol. Chem. 2006, 281, 4075–4086. [Google Scholar] [CrossRef] [Green Version]
- Kendrick, J.S.; Chan, L.; Higgins, J.A. Superior role of apolipoprotein B48 over apolipoprotein B100 in chylomicron assembly and fat absorption: An investigation of apobec-1 knock-out and wild-type mice. Biochem. J. 2001, 356, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Fukuda, S.; Aoyama, H.; Taniuchi, N.; Ishihara, T.; Ohashi, N.; Sato, H.; Wakimoto, K.; Shiotani, M.; Oku, A. MGAT2 deficiency ameliorates high-fat diet-induced obesity and insulin resistance by inhibiting intestinal fat absorption in mice. Lipids Health Dis. 2012, 11, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, C.L.; Cheong, M.L.; Grueter, C.; Zhou, P.; Moriwaki, J.; Wong, J.S.; Hubbard, B.; Marmor, S.; Farese, R.V., Jr. Deficiency of the intestinal enzyme acyl CoA:monoacylglycerol acyltransferase-2 protects mice from metabolic disorders induced by high-fat feeding. Nat. Med. 2009, 15, 442–446. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.K.; Huckestein, B.R.; Edmunds, L.R.; Petersen, M.C.; Nasiri, A.; Butrico, G.M.; Abulizi, A.; Harmon, D.B.; Lu, C.; Mantell, B.S.; et al. Reduced intestinal lipid absorption and body weight-independent improvements in insulin sensitivity in high-fat diet-fed Park2 knockout mice. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E105–E116. [Google Scholar] [CrossRef] [Green Version]
- Buhman, K.K.; Smith, S.J.; Stone, S.J.; Repa, J.J.; Wong, J.S.; Knapp, F.F., Jr.; Burri, B.J.; Hamilton, R.L.; Abumrad, N.A.; Farese, R.V., Jr. DGAT1 is not essential for intestinal triacylglycerol absorption or chylomicron synthesis. J. Biol. Chem. 2002, 277, 25474–25479. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.J.; Cases, S.; Jensen, D.R.; Chen, H.C.; Sande, E.; Tow, B.; Sanan, D.A.; Raber, J.; Eckel, R.H.; Farese, R.V., Jr. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nat. Genet. 2000, 25, 87–90. [Google Scholar] [CrossRef]
- Chen, H.C.; Ladha, Z.; Smith, S.J.; Farese, R.V., Jr. Analysis of energy expenditure at different ambient temperatures in mice lacking DGAT1. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E213–E218. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Qarni, A.A.; Bakillah, A. Diet-induced differential effects on plasma lipids secretion by the inositol-requiring transmembrane kinase/endoribonuclease 1alpha. Front. Biosci. 2021, 26, 11–21. [Google Scholar] [CrossRef]
- Iqbal, J.; Parks, J.S.; Hussain, M.M. Lipid absorption defects in intestine-specific microsomal triglyceride transfer protein and ATP-binding cassette transporter A1-deficient mice. J. Biol. Chem. 2013, 288, 30432–30444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, R.H. Hyperlipidemia as a risk factor for cardiovascular disease. Prim. Care 2013, 40, 195–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopper-Borge, E.A.; Churchill, T.; Paulose, C.; Nicolas, E.; Jacobs, J.D.; Ngo, O.; Kuang, Y.; Grinberg, A.; Westphal, H.; Chen, Z.S.; et al. Contribution of Abcc10 (Mrp7) to in vivo paclitaxel resistance as assessed in Abcc10(-/-) mice. Cancer Res. 2011, 71, 3649–3657. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Rudel, L.L.; Hussain, M.M. Microsomal triglyceride transfer protein enhances cellular cholesteryl esterification by relieving product inhibition. J. Biol. Chem. 2008, 283, 19967–19980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, J.E.; Samuel, V.T.; Morton, G.J.; Obici, S.; Croniger, C.M.; Shulman, G.I.; Wasserman, D.H.; McGuinness, O.P. Standard operating procedures for describing and performing metabolic tests of glucose homeostasis in mice. Dis. Model. Mech. 2010, 3, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Dinger, K.; Mohr, J.; Vohlen, C.; Hirani, D.; Hucklenbruch-Rother, E.; Ensenauer, R.; Dotsch, J.; Alejandre Alcazar, M.A. Intraperitoneal Glucose Tolerance Test, Measurement of Lung Function, and Fixation of the Lung to Study the Impact of Obesity and Impaired Metabolism on Pulmonary Outcomes. J. Vis. Exp. 2018, 133, e56685. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, 131, e56672. [Google Scholar] [CrossRef]
- Iqbal, J.; Hussain, M.M. Evidence for multiple complementary pathways for efficient cholesterol absorption in mice. J. Lipid Res. 2005, 46, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Anwar, K.; Hussain, M.M. Multiple, independently regulated pathways of cholesterol transport across the intestinal epithelial cells. J. Biol. Chem. 2003, 278, 31610–31620. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Queiroz, J.; Li, Y.; Jiang, X.C.; Ron, D.; Hussain, M.M. Increased intestinal lipid absorption caused by Ire1beta deficiency contributes to hyperlipidemia and atherosclerosis in apolipoprotein E-deficient mice. Circ. Res. 2012, 110, 1575–1584. [Google Scholar] [CrossRef]
- Athar, H.; Iqbal, J.; Jiang, X.C.; Hussain, M.M. A simple, rapid, and sensitive fluorescence assay for microsomal triglyceride transfer protein. J. Lipid Res. 2004, 45, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Huan, C.; Chakraborty, M.; Zhang, H.; Lu, D.; Kuo, M.S.; Cao, G.; Jiang, X.C. Macrophage sphingomyelin synthase 2 deficiency decreases atherosclerosis in mice. Circ. Res. 2009, 105, 295–303. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otaibi, A.A.; Mubarak, S.A.; Qarni, A.A.; Hawwari, A.; Bakillah, A.; Iqbal, J. ATP-Binding Cassette Protein ABCC10 Deficiency Prevents Diet-Induced Obesity but Not Atherosclerosis in Mice. Int. J. Mol. Sci. 2022, 23, 13813. https://doi.org/10.3390/ijms232213813
Otaibi AA, Mubarak SA, Qarni AA, Hawwari A, Bakillah A, Iqbal J. ATP-Binding Cassette Protein ABCC10 Deficiency Prevents Diet-Induced Obesity but Not Atherosclerosis in Mice. International Journal of Molecular Sciences. 2022; 23(22):13813. https://doi.org/10.3390/ijms232213813
Chicago/Turabian StyleOtaibi, Abeer Al, Sindiyan Alshaikh Mubarak, Ali Al Qarni, Abbas Hawwari, Ahmed Bakillah, and Jahangir Iqbal. 2022. "ATP-Binding Cassette Protein ABCC10 Deficiency Prevents Diet-Induced Obesity but Not Atherosclerosis in Mice" International Journal of Molecular Sciences 23, no. 22: 13813. https://doi.org/10.3390/ijms232213813