
Pioglitazone Modifies Kupffer Cell Function and Protects against Escherichia coli-Induced Bacteremia in Burned Mice
 , , and
, , and
Abstract
:1. Introduction
2. Results
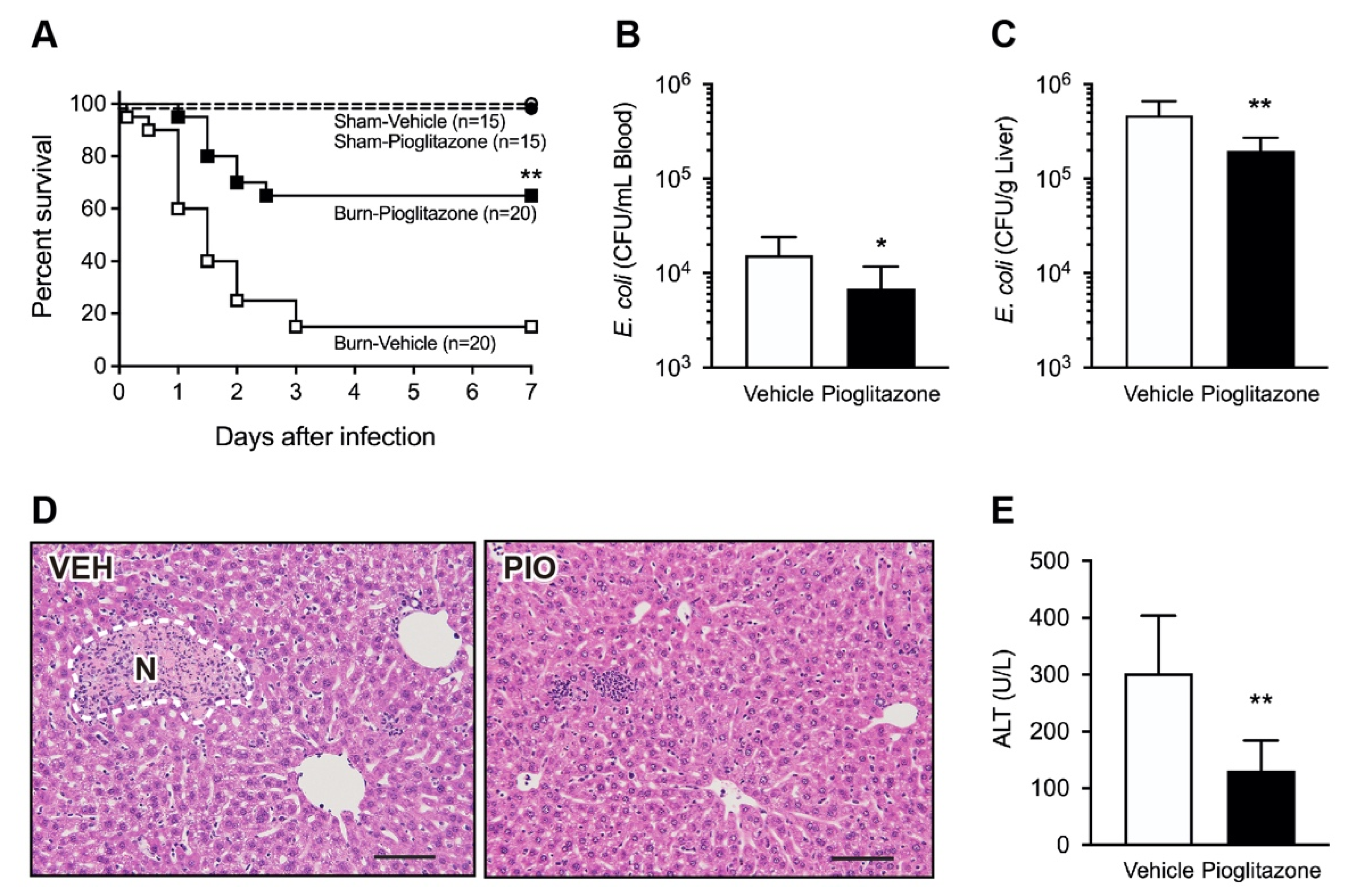
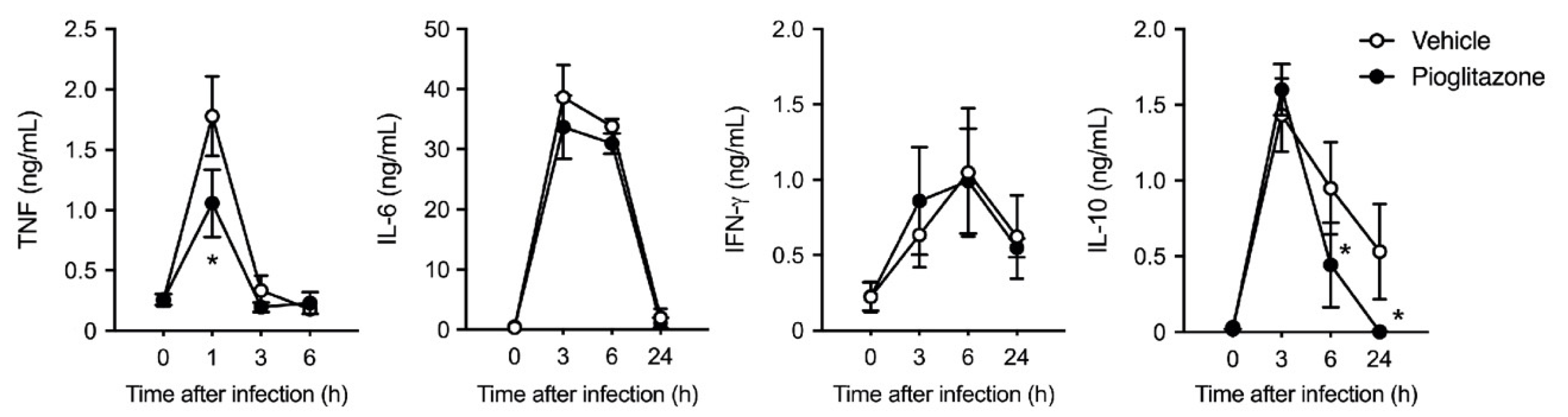
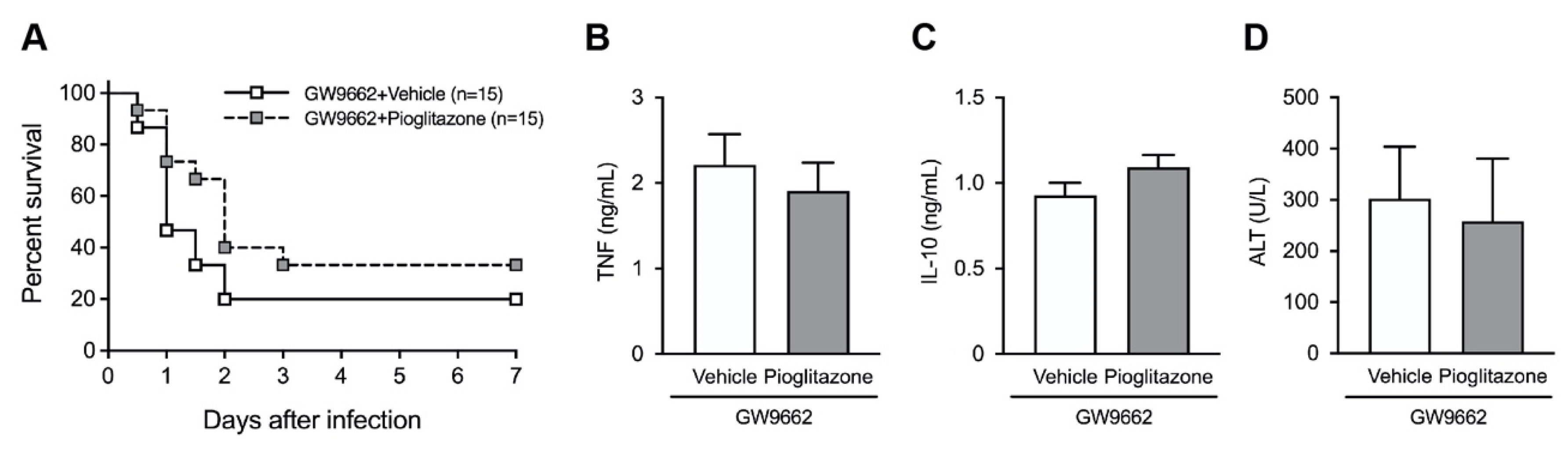
2.1. PPARγ Activation Protects Mice against Postburn Bacterial Infection
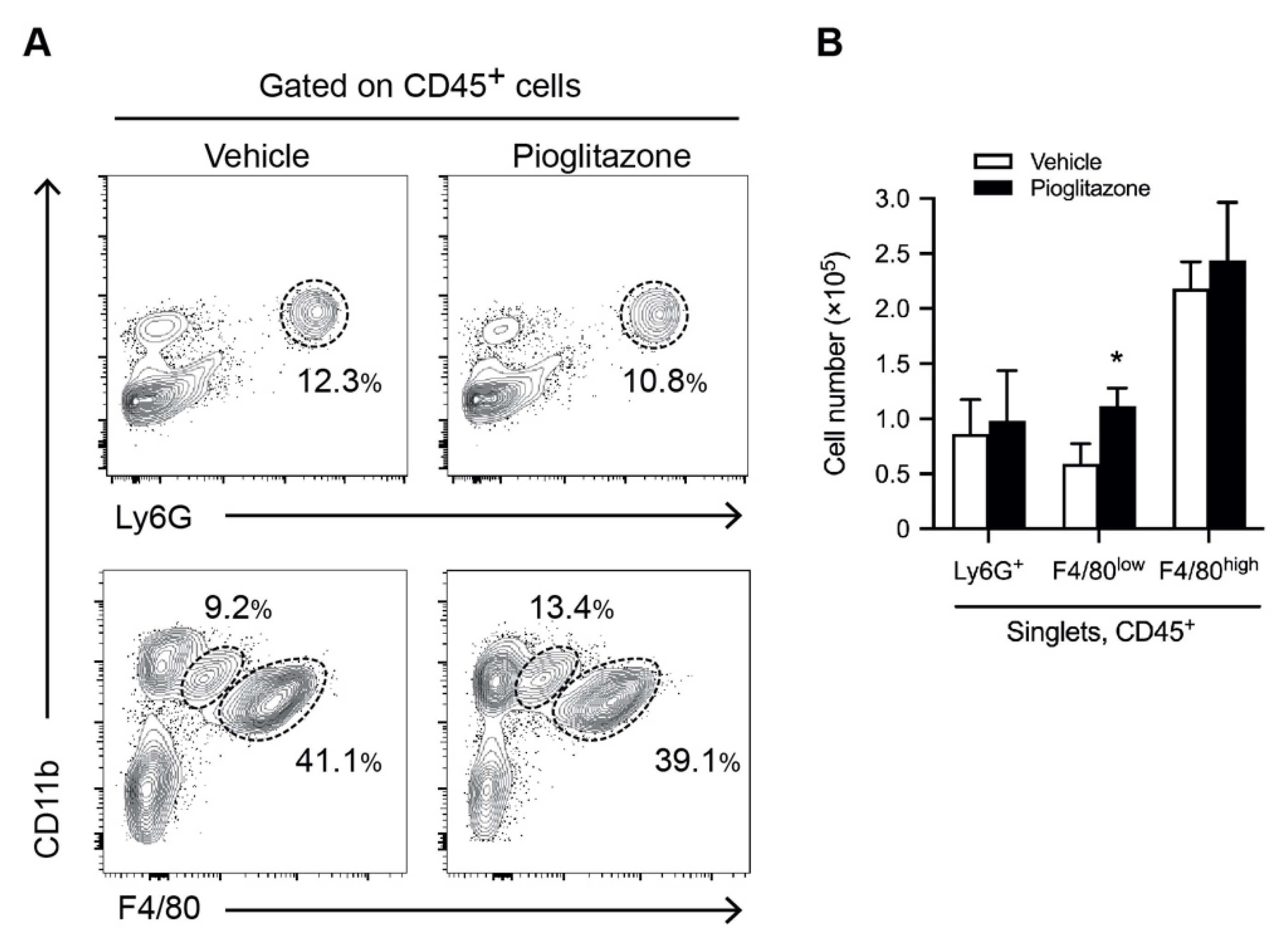
2.2. Pioglitazone Promotes the Recruitment of Inflammatory Monocyte/Macrophages into the Liver
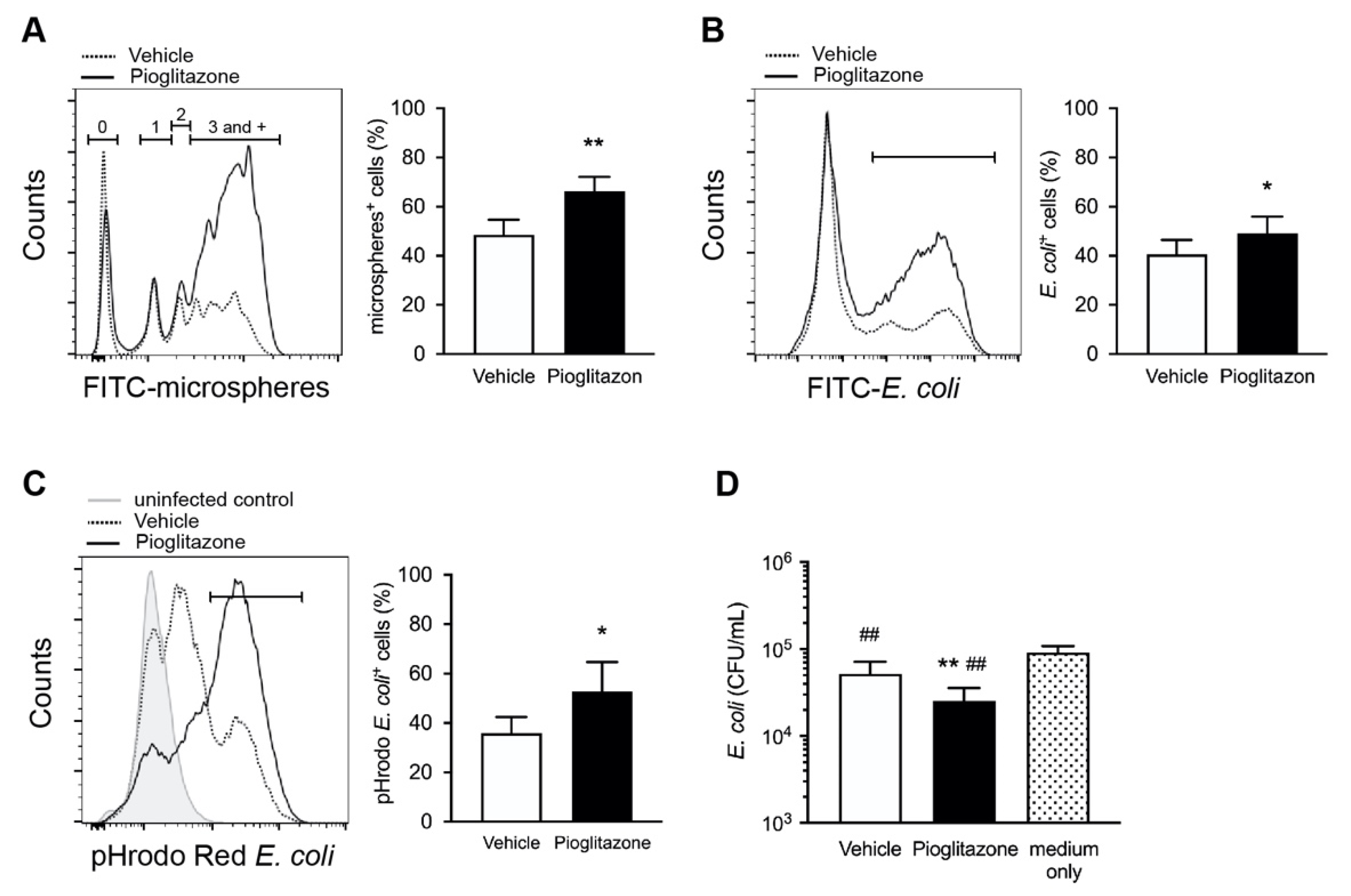
2.3. Treatment with Pioglitazone Enhances Phagocytosis and Bactericidal Activity in Burned Mice
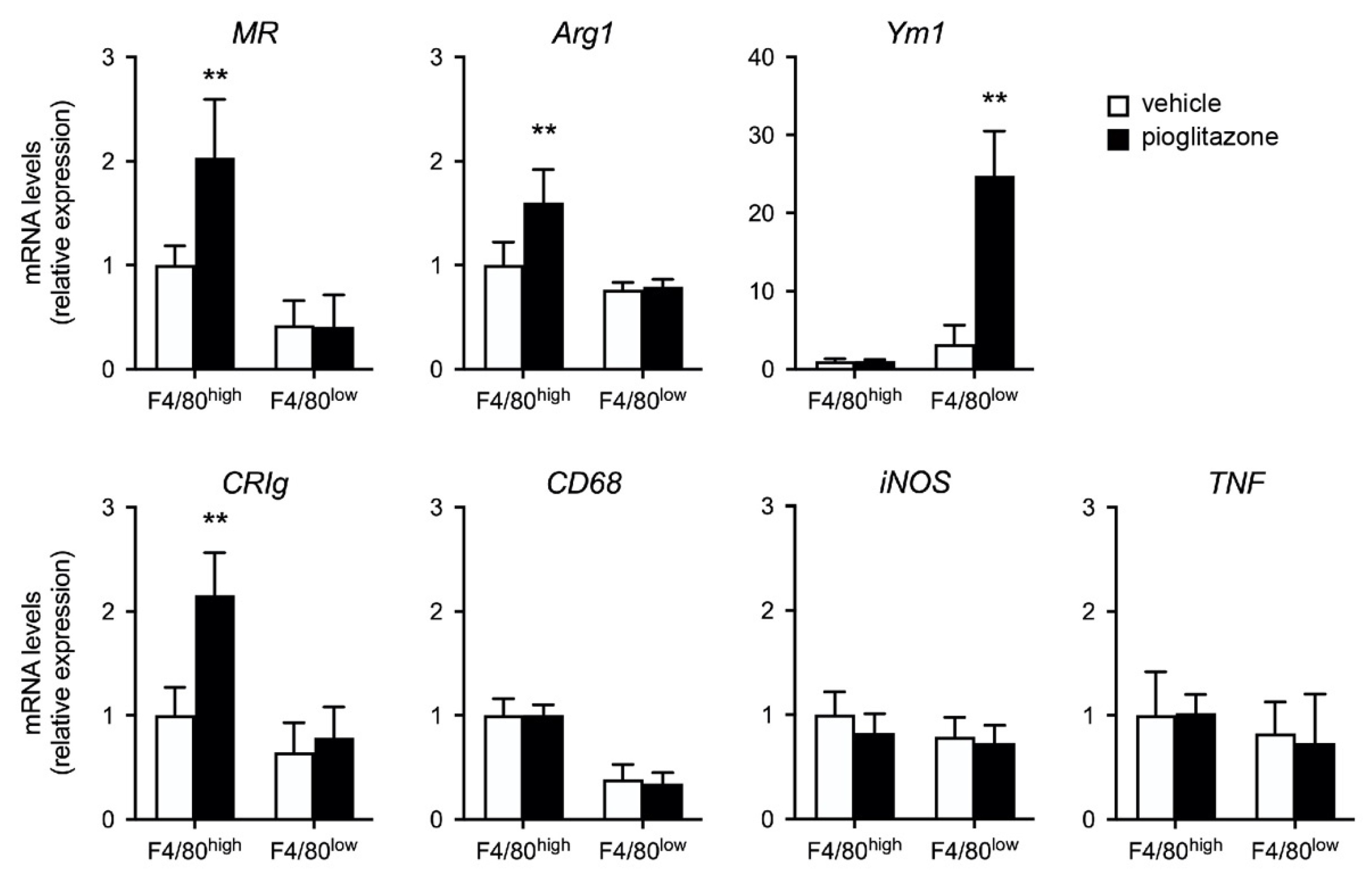
2.4. Pioglitazone Directs Liver F4/80+ Cells toward M2-like Phenotype in Burn-Injured Mice
3. Discussion
4. Materials and Methods
4.1. Animal Model of Burn Injury and Pioglitazone Treatment
4.2. Systemic Bacterial Challenge
4.3. Isolation of Hepatic Immune Cells Including KCs
4.4. Flow Cytometry
4.5. Phagocytosis, Bactericidal Activity, and In Vitro Killing Assays
4.6. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. ALT Level Determination
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lachiewicz, A.M.; Hauck, C.G.; Weber, D.J.; Cairns, B.A.; van Duin, D. Bacterial Infections After Burn Injuries: Impact of Multidrug Resistance. Clin. Infect. Dis. 2017, 65, 2130–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeschke, M.G.; van Baar, M.E.; Choudhry, M.A.; Chung, K.K.; Gibran, N.S.; Logsetty, S. Burn injury. Nat. Rev. Dis. Prim. 2020, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Belba, M.K.; Petrela, E.Y.; Belba, A.G. Epidemiology and outcome analysis of sepsis and organ dysfunction/failure after burns. Burns 2017, 43, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, P.; Frew, Q.; Green, A.; Martin, R.; Dziewulski, P. Cause of death and correlation with autopsy findings in burns patients. Burns 2013, 39, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zou, B.; Liou, Y.C.; Huang, C. The pathogenesis and diagnosis of sepsis post burn injury. Burns Trauma 2021, 9, tkaa047. [Google Scholar] [CrossRef]
- Van der Heide, D.; Weiskirchen, R.; Bansal, R. Therapeutic Targeting of Hepatic Macrophages for the Treatment of Liver Diseases. Front. Immunol. 2019, 10, 2852. [Google Scholar] [CrossRef] [Green Version]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef]
- Dhainaut, J.F.; Marin, N.; Mignon, A.; Vinsonneau, C. Hepatic response to sepsis: Interaction between coagulation and inflammatory processes. Crit. Care Med. 2001, 29, S42–S47. [Google Scholar] [CrossRef]
- Li, P.; He, K.; Li, J.; Liu, Z.; Gong, J. The role of Kupffer cells in hepatic diseases. Mol. Immunol. 2017, 85, 222–229. [Google Scholar] [CrossRef]
- Inatsu, A.; Kinoshita, M.; Nakashima, H.; Shimizu, J.; Saitoh, D.; Tamai, S.; Seki, S. Novel mechanism of C-reactive protein for enhancing mouse liver innate immunity. Hepatology 2009, 49, 2044–2054. [Google Scholar] [CrossRef]
- Kinoshita, M.; Seki, S.; Ono, S.; Shinomiya, N.; Hiraide, H. Paradoxical effect of IL-18 therapy on the severe and mild Escherichia coli infections in burn-injured mice. Ann. Surg. 2004, 240, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, H.; Kinoshita, M.; Ono, S.; Nakashima, M.; Hara, E.; Ohno, H.; Seki, S.; Saitoh, D. Augmented bacterial elimination by Kupffer cells after IL-18 pretreatment via IFN-gamma produced from NK cells in burn-injured mice. Burns 2011, 37, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Xiu, F.; Jeschke, M.G. Perturbed mononuclear phagocyte system in severely burned and septic patients. Shock 2013, 40, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPARgamma and the Innate Immune System Mediate the Resolution of Inflammation. PPAR Res. 2015, 2015, 549691. [Google Scholar] [CrossRef] [Green Version]
- Lehrke, M.; Lazar, M.A. The many faces of PPARgamma. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef] [Green Version]
- Bouhlel, M.A.; Derudas, B.; Rigamonti, E.; Dievart, R.; Brozek, J.; Haulon, S.; Zawadzki, C.; Jude, B.; Torpier, G.; Marx, N.; et al. PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 2007, 6, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Zingarelli, B.; Cook, J.A. Peroxisome proliferator-activated receptor-gamma is a new therapeutic target in sepsis and inflammation. Shock 2005, 23, 393–399. [Google Scholar] [CrossRef]
- Bedi, B.; Yuan, Z.; Joo, M.; Zughaier, S.M.; Goldberg, J.B.; Arbiser, J.L.; Hart, C.M.; Sadikot, R.T. Enhanced Clearance of Pseudomonas aeruginosa by Peroxisome Proliferator-Activated Receptor Gamma. Infect. Immun. 2016, 84, 1975–1985. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, A.E.; Sisti, F.; Sonego, F.; Wang, S.; Filgueiras, L.R.; Brandt, S.; Serezani, A.P.; Du, H.; Cunha, F.Q.; Alves-Filho, J.C.; et al. PPAR-gamma/IL-10 axis inhibits MyD88 expression and ameliorates murine polymicrobial sepsis. J. Immunol. 2014, 192, 2357–2365. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.; Nowell, M.; Chima, R.; Zingarelli, B. Pioglitazone reduces inflammation through inhibition of NF-kappaB in polymicrobial sepsis. Innate Immun. 2014, 20, 519–528. [Google Scholar] [CrossRef]
- Liu, S.Y.; Huang, C.C.; Huang, S.F.; Liao, T.L.; Kuo, N.R.; Yang, Y.Y.; Li, T.H.; Liu, C.W.; Hou, M.C.; Lin, H.C. Pioglitazone Ameliorates Acute Endotoxemia-Induced Acute on Chronic Renal Dysfunction in Cirrhotic Ascitic Rats. Cells 2021, 10, 3044. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Kinoshita, M.; Nakashima, H.; Kotani, A.; Ishikiriyama, T.; Kato, S.; Hiroi, S.; Seki, S. Pioglitazone improves phagocytic activity of liver recruited macrophages in elderly mice possibly by promoting glucose catabolism. Innate Immun. 2019, 25, 356–368. [Google Scholar] [CrossRef]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Lefere, S.; Puengel, T.; Hundertmark, J.; Penners, C.; Frank, A.K.; Guillot, A.; de Muynck, K.; Heymann, F.; Adarbes, V.; Defrene, E.; et al. Differential effects of selective- and pan-PPAR agonists on experimental steatohepatitis and hepatic macrophages. J. Hepatol. 2020, 73, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Linares, I.; Farrokhi, K.; Echeverri, J.; Kaths, J.M.; Kollmann, D.; Hamar, M.; Urbanellis, P.; Ganesh, S.; Adeyi, O.A.; Yip, P.; et al. PPAR-gamma activation is associated with reduced liver ischemia-reperfusion injury and altered tissue-resident macrophages polarization in a mouse model. PLoS ONE 2018, 13, e0195212. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Xu, Q.; Wang, Q.; Wu, H.; Hua, J. Effect of modulation of PPAR-gamma activity on Kupffer cells M1/M2 polarization in the development of non-alcoholic fatty liver disease. Sci. Rep. 2017, 7, 44612. [Google Scholar] [CrossRef] [Green Version]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Red Eagle, A.; Vats, D.; Brombacher, F.; Ferrante, A.W.; et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Jiang, Y.; Xiao, X.; Peng, Y.; Xiao, X.; Yang, M. Protective effect of pioglitazone on sepsis-induced intestinal injury in a rodent model. J. Surg. Res. 2015, 195, 550–558. [Google Scholar] [CrossRef]
- Kimura, T.; Nada, S.; Takegahara, N.; Okuno, T.; Nojima, S.; Kang, S.; Ito, D.; Morimoto, K.; Hosokawa, T.; Hayama, Y.; et al. Polarization of M2 macrophages requires Lamtor1 that integrates cytokine and amino-acid signals. Nat. Commun. 2016, 7, 13130. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Zhang, Y.; Pan, G.; Xiang, L.X.; Luo, D.C.; Shao, J.Z. Occurrences and Functions of Ly6C(hi) and Ly6C(lo) Macrophages in Health and Disease. Front. Immunol. 2022, 13, 901672. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, N.; Asano, K.; Kikuchi, K.; Uchida, Y.; Ikegami, H.; Takagi, R.; Yotsumoto, S.; Shibuya, T.; Makino-Okamura, C.; Fukuyama, H.; et al. Emergence of immunoregulatory Ym1(+)Ly6C(hi) monocytes during recovery phase of tissue injury. Sci. Immunol. 2018, 3, eaat0207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.H.; Jenne, C.N.; Petri, B.; Chrobok, N.L.; Kubes, P. Nucleation of platelets with blood-borne pathogens on Kupffer cells precedes other innate immunity and contributes to bacterial clearance. Nat. Immunol. 2013, 14, 785–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannagan, R.S.; Jaumouille, V.; Grinstein, S. The cell biology of phagocytosis. Annu. Rev. Pathol. 2012, 7, 61–98. [Google Scholar] [CrossRef] [PubMed]
- Helmy, K.Y.; Katschke, K.J., Jr.; Gorgani, N.N.; Kljavin, N.M.; Elliott, J.M.; Diehl, L.; Scales, S.J.; Ghilardi, N.; van Lookeren Campagne, M. CRIg: A macrophage complement receptor required for phagocytosis of circulating pathogens. Cell 2006, 124, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Fu, Y.; Yosri, M.; Chen, Y.; Sun, P.; Xu, J.; Zhang, M.; Sun, D.; Strickland, A.B.; Mackey, Z.B.; et al. CRIg plays an essential role in intravascular clearance of bloodborne parasites by interacting with complement. Proc. Natl. Acad. Sci. USA 2019, 116, 24214–24220. [Google Scholar] [CrossRef]
- Gorgani, N.N.; Thathaisong, U.; Mukaro, V.R.; Poungpair, O.; Tirimacco, A.; Hii, C.S.; Ferrante, A. Regulation of CRIg expression and phagocytosis in human macrophages by arachidonate, dexamethasone, and cytokines. Am. J. Pathol. 2011, 179, 1310–1318. [Google Scholar] [CrossRef]
- Gauglitz, G.G.; Song, J.; Herndon, D.N.; Finnerty, C.C.; Boehning, D.; Barral, J.M.; Jeschke, M.G. Characterization of the inflammatory response during acute and post-acute phases after severe burn. Shock 2008, 30, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Flannagan, R.S.; Cosio, G.; Grinstein, S. Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef]
- Canton, J.; Khezri, R.; Glogauer, M.; Grinstein, S. Contrasting phagosome pH regulation and maturation in human M1 and M2 macrophages. Mol. Biol. Cell 2014, 25, 3330–3341. [Google Scholar] [CrossRef]
- Ganeshan, K.; Chawla, A. Metabolic regulation of immune responses. Annu. Rev. Immunol. 2014, 32, 609–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forwards | Reverse |
|---|---|---|
| MR | 5′-CCCAAGGGCTCTTCTAAAGCA-3′ | 5′-CGCCGGCACCTATCACA-3′ |
| Arg1 | 5′-CTCCAAGCCAAAGTCCTTAGAG-3′ | 5′-AGGAGCTGTCATTAGGGACATC-3′ |
| Ym1 | 5′-TCACTTACACACATGAGCAAGAC-3′ | 5′-CGGTTCTGAGGAGTAGAGACCA-3′ |
| CRIg | 5′-GTCCTGCAGCGGAACAAGATATAA-3′ | 5′-GACTTGACCACTAATGGGACTGGAA-3′ |
| CD68 | 5′-CCACAGTTTCTCCCACCACA-3′ | 5′-AATTTGGGGCTTGGAGCTGAA-3′ |
| iNOS | 5′-GCAGAGATTGGAGGCCTTGTG-3′ | 5′-GGGTTGTTGCTGAACTTCCAGTC-3′ |
| TNF | 5′-CCAGAAAAGACACCATGAGCAC-3′ | 5′-TCACCCCGAAGTTCAGTAGACA-3′ |
| Rps18 | 5′-TTCTGGCCAACGGTCTAGACAAC-3′ | 5′-CCAGTGGTCTTGGTGTGCTGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyazaki, H.; Kinoshita, M.; Nakashima, H.; Nakamura, S.; Saitoh, D. Pioglitazone Modifies Kupffer Cell Function and Protects against Escherichia coli-Induced Bacteremia in Burned Mice. Int. J. Mol. Sci. 2022, 23, 12746. https://doi.org/10.3390/ijms232112746
Miyazaki H, Kinoshita M, Nakashima H, Nakamura S, Saitoh D. Pioglitazone Modifies Kupffer Cell Function and Protects against Escherichia coli-Induced Bacteremia in Burned Mice. International Journal of Molecular Sciences. 2022; 23(21):12746. https://doi.org/10.3390/ijms232112746
Chicago/Turabian StyleMiyazaki, Hiromi, Manabu Kinoshita, Hiroyuki Nakashima, Shingo Nakamura, and Daizoh Saitoh. 2022. "Pioglitazone Modifies Kupffer Cell Function and Protects against Escherichia coli-Induced Bacteremia in Burned Mice" International Journal of Molecular Sciences 23, no. 21: 12746. https://doi.org/10.3390/ijms232112746