
Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer


State Key Laboratory of Esophageal Cancer Prevention and Treatment, Key Laboratory of Advanced Pharmaceutical Technology, Ministry of Education of China, School of Pharmaceutical Sciences, Zhengzhou University, Zhengzhou 450001, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(17), 9574; https://doi.org/10.3390/ijms23179574
Submission received: 24 July 2022
/
Revised: 16 August 2022
/
Accepted: 18 August 2022
/
Published: 24 August 2022
(This article belongs to the Section Molecular Oncology)
Abstract
:Polycomp group (PcG) proteins are members of highly conserved multiprotein complexes, recognized as gene transcriptional repressors during development and shown to play a role in various physiological and pathological processes. PcG proteins consist of two Polycomb repressive complexes (PRCs) with different enzymatic activities: Polycomb repressive complexes 1 (PRC1), a ubiquitin ligase, and Polycomb repressive complexes 2 (PRC2), a histone methyltransferase. Traditionally, PRCs have been described to be associated with transcriptional repression of homeotic genes, as well as gene transcription activating effects. Particularly in cancer, PRCs have been found to misregulate gene expression, not only depending on the function of the whole PRCs, but also through their separate subunits. In this review, we focused especially on the recent findings in the transcriptional regulation of PRCs, the oncogenic and tumor-suppressive roles of PcG proteins, and the research progress of inhibitors targeting PRCs.
1. Introduction
Aberrant regulation of epigenetic pathways is thought to be a frequent event in cancer. Understanding the roles of these epigenetic regulators has facilitated the discovery of new therapeutic approaches for cancer [1]. Polycomb group (PcG) proteins are a group of widely studied epigenetic regulators, originally discovered in Drosophila melanogaster and found to be functionally and compositionally conserved in other animals, including Caenorhabditis elegans, mice, and humans [2,3,4]. The PcG proteins play a critical role in the progression of cancer forming multimeric complexes involved in transcriptional repression, including Polycomb repressive complex 1 (PRC1) and Polycomb repressive complex 2 (PRC2) [5].
Polycomb repressive complexes (PRCs) mainly mediate transcription and gene expression by regulating post-translational modifications of histones, in which PRC1 catalyzes the mono-ubiquitination of histone H2A at Lys119 (H2AK119ub1), whereas PRC2 catalyzes mono-, di-, and trimethylation of histone H3 at Lys27 (H3K27me1, H3K27me2, and H3K27me3) [6,7]. PRC2-catalyzed H3K27me3 is a hallmark of transcriptional silencing. Furthermore, PRC1 and PRC2 were found to spatially converge on the same sites in the genome to form Polycomb chromatin domains, with H2AK119ub1 and H3K27me3 uniquely enriched in these domains [8,9,10]. Gene repression is thought to be mediated by PRC1 and PRC2 cooperatively, although the specific mechanisms have not been fully defined. As shown in Figure 1, Polycomb complexes make up the catalytic core, and they bind accessory proteins to constitute distinct PRC1 and PRC2 complexes. In recent years, it has gradually emerged that PRC1 is not a single complex, and that there are at least eight different complexes [11]. These complexes can be further divided into canonical PRC1 (cPRC1) and non-canonical PRC1 (ncPRC1) depending on whether they contain one of two homologous proteins, YY1-associated factor 2 (YAF2) and RING1 and YY1 binding protein (RYBP), or a Chromobox (CBX) protein, respectively [12]. Moreover, proteomic and biochemical analyses have revealed that both cPRC1 and ncPRC1 possess E3 ubiquitin ligase RING1A/B, a core subunit that catalyzes the ubiquitination of histone H2A [12]. In mammals, PRC2 mainly contains four core subunits, enhancer of zeste homolog 1/2 (EZH1/2), embryonic ectoderm development (EED), suppressor of zeste 12 (SUZ12), and retinoblastoma protein-associated proteins 46/48 (RBAP46/48) [13,14]. In addition, these core proteins associate with different cofactors to create two distinct PRC2 variants, PRC2.1 and PRC2.2 (Figure 1B).
Studies have shown that the expression level of EZH2, the core catalytic subunit of PRC2, is positively correlated with tumor grade in prostate and breast cancers and PRC2 is initially thought to have an oncogenic function [15,16,17]. Subsequently, catalytically hyperactivating mutations of EZH2 in somatic cells were identified in patients with non-Hodgkin lymphoma (NHL), indicating that PRC2 is critically involved in the progression of lymphoma [18,19,20]. However, loss-of-function mutations in the catalytic subunit of PRC2 or loss of other components are found in some cancers (such as leukemia, myeloproliferative neoplasms, and malignant peripheral nerve sheath tumors), suggesting that PRC2 can also have tumor-suppressive functions [21,22,23]. Similar to PRC2, PRC1 also plays dual roles as an oncogenic and tumor suppressor. Its E3 ubiquitin ligase RING1A/B, subunits CBX2/4, and Polycomb group ring finger 1/4 (PCGF1/4) are oncogenic in leukemias, lymphomas, gliomas, and other tumors [24,25,26,27,28]. In contrast, some subunits of PRC1 have also been reported to have tumor-suppressive roles such as CBX6 and PCGF2 in breast cancer [29,30,31]. As discussed above, PRCs have both oncogenic and tumor-suppressive functions that may limit the use of inhibitors and highlight the importance of expounding the specific function of PRCs in the tumor.
In this review, we provide an overview of the diverse PRC1 and PRC2 variants generated by different combinations of PcG proteins. Additionally, we discuss the classical functions of PRCs as well as the recently identified novel functions of PcG proteins, providing some references for therapeutic directions of related inhibitors. Overall, we focus on two major themes: functions of PRCs in mammalian transcription, and the dual roles of core and accessory subunits of PRCs in cancer.
2. Composition of PRC1 and the Role It Plays in Transcription
PRC1 was first purified from Drosophila melanogaster embryos, which is mainly composed of Sex Combs Extra (Sce), Polycomb (Pc), Posterior Sex Combs (Psc), Polyhomeotic (Ph), and Sex Comb on Midleg (Scm) [32]. A similar complex containing the core subunits of D. melanogaster PRC1 and various homologous proteins was later purified in mammalian cells [4]. Proteomic and biochemical analyses over the last decade have revealed the enormous complexity of mammalian PRC1, which is now known to include many canonical and non-canonical complex variants (Figure 1A).
2.1. Composition of PRC1 and Its Ubiquitin Ligase Activity
All PRC1 variants contain the E3 ubiquitin ligase RING1A/B and are designated PRC1.1-PRC1.6 based on the six PCGF proteins they contain [12]. They are generally categorized as canonical PRC1 (cPRC1) or non-canonical PRC1 (ncPRC1, Figure 1A), a nomenclature that reflects the compositional similarity of these complexes to PRC1 in D. melanogaster. The mammalian cPRC1 complexes assemble around either PCGF2 or PCGF 4 (also referred to as MEL-18 or BMI-1) and include five chromodomain proteins (CBX2/4/6/7/8), three Polyhomeotic subunits (PHC1/2/3) and SCM homologs (SCMH1, SCML1/2), which is largely consistent with the composition of the originally discovered Drosophila PRC1 [4,12,33,34,35]. Six PCGF proteins were present in ncPRC1 variants, but only PCGF2 and PCGF4 were found in the cPRC1 [12]. Thus, PRC1.2 and PRC1.4 can be further subdivided into cPRC1.2/4 and ncPRC1.2/4. In addition, the ncPRC1 complexes were also composed of RYBP or its homologous protein YAF2 and various PCGF-specific cofactors [12,33]. Furthermore, a major difference between cPRC1 and ncPRC1 variants is whether their occupancy on chromatin is dependent on PRC2-deposited H3K27me3, and the CBX proteins of cPRC1 can bind to H3K27me3, which is required for their occupancy on chromatin [33]. By contrast, the RYBP or YAF2 subunits of ncPRC1 variants can bind chromatin independently of PRC2 proteins and their activity significantly stimulates the E3 ubiquitin ligase activity of RING1B in vitro [8,33,34,36]. An in-depth discussion of the specific classification of PRC1 is detailed in REF 5.
A key function of the mammalian PRC1 is to catalyze H2AK119ub1, an epigenetic modification closely associated with PRCs-mediated gene silencing [7]. Although all six PCGF proteins have highly similar RING domains, the variants of PRC1 complexes that they form vary considerably in their E3 ubiquitin ligase activity due to differences in cofactors. It is reported that these cofactors can specifically enhance the catalytic activity of PRC1 or promote its recruitment to nucleosomes [5]. Notably, ncPRC1 variants exhibit significantly stronger activity on nucleosome substrates than cPRC1 complexes, in which E3 ligase activity is significantly stimulated by the incorporation of the ncPRC1-specific accessory subunit RYBP [12,36].
2.2. PRC1 in Transcription Repression
Accumulating evidence suggests that PRC1 plays an important role in the repression of gene transcription through chromatin modifications [37,38]. Two mechanisms, H2AK119ub1, and chromatin compaction are currently proposed to account for the mediation of this gene silencing (Figure 2A–C) [7,39]. RING1A/B-catalyzed H2AK119ub1 provides a general mechanism for PRC1-mediated transcriptional repression. Genome-wide studies have shown a strong correlation between H2AK119ub1 deposition and gene inhibition [40,41,42]. Moreover, the occupancy of H2AK119ub1 on chromatin is able to maintain the conformational equilibrium of RNA polymerase II (RNAP) and inhibits its facilitating effect on transcription elongation, which supports the gene transcription repression function of PRC1 [41,43]. Phosphorylation of amino acid residues within the carboxy-terminal domain (CTD) of RNAP is associated with transcription initiation, elongation, and termination, where active transcription sites are typically characterized by phosphorylation of its Ser2 residues, whereas inactive or stable genes bind Ser5-phosphorylated RNAP in promoter-proximal regions [44]. H2AK119ub1 has been reported to inhibit the transcriptional activation of RNAP by balancing the recruitment of Ser5-phosphorylated RNAP and Ser2-phosphorylated RNAP at gene loci [41]. On the other hand, it can block the binding of RNAP at the early stages of elongation by preventing the recruitment of Facilitates Chromatin Transcription (FACT) to the promoter region of transcription [43]. In addition to its ability to counteract RNAP binding and transcription initiation, PRC1 may also play a critical role at promoters, controlling the frequency of transcriptional bursts to drive Polycomb-mediated gene repression by counteracting low-level or inappropriate transcriptional signals emanating from regulatory elements such as enhancers [45]. Furthermore, PRC1 can induce chromatin compaction through disordered regions of highly positively charged amino acids in its Psc subunits while repressing the transcription of genes [46].
PRC1 often cooperates closely with PRC2 to target, establish, and maintain transcriptional repression of PcG-targeted genes (Figure 2A,B) [47,48]. This cooperation was initially thought to be initiated by PRC2-catalyzed H3K27me3, which is then recognized by the CBX proteins of PRC1, driving cPRC1 to pre-occupied loci of PRC2 to exert transcriptional repression (Figure 2A) [6,47,49]. However, because there are no CBX proteins in ncPRC1, this model can only be applied to cPRC1 recruitment. Indeed, in PRC2-deficient mouse embryonic stem cells (ESCs), RING1B can also occupy the majority of gene loci targeted by PcG, suggesting that PRC1 can be recruited to nucleosomes in an H3K27me3-independent manner (Figure 2B) [33,50]. In addition, recent studies have complemented the mechanism of cooperation of ncPRC1 with PRC2, and researchers have shown that ncPRC1-mediated H2AK119ub1 recruits PRC2 [35,51]. Similar to the mechanism by which PRC2 recruits cPRC1, H2AK119ub1 catalyzed by ncPRC1 can be recognized and bound by JARID2, driving PRC2 to catalyze H3K27me3 at PcG-targeted gene loci to exert transcriptional repression [52,53].
PRC1 can also restrict gene transcription in a chromatin compaction manner, independent of histone H2A ubiquitination (Figure 2C) [32,39,54]. Preliminary in vitro analyses with short nucleosome arrays suggest that core components of the D. melanogaster PRC1, especially the Psc subunits, generate a compact chromatin structure by a mechanism involving interactions with nucleosomes without the need for histone tails [32,39]. Subsequent studies on ESCs with loss-of-function mutations in mouse RING1B (I53A) identified chromatin decompaction, and the addition of RING1B restored chromatin compaction in vivo, providing further evidence of a chromatin compaction role for PRC1 [54]. Furthermore, characterization of the organization of PcG-targeted genes in ESCs and neural progenitor cells using 5C and super-resolution microscopy revealed that chromatin compaction at PRC1-repressed loci formed isolated self-interacting domains and that chromatin compaction was only associated with cPRC1 [55]. The Psc subunits of the PRC1 complex play a key role in chromatin compaction in Drosophila, consistent with the predominant role of CBX proteins in mammals. Similar to Psc, CBX2 also has a disordered region of highly positively charged amino acids that is critical for inducing chromatin compaction in vitro [46,56]. Importantly, in a mouse model carrying CBX2 with a mutant nucleosome compaction region, homologous heterogeneous transformations similar to those observed with PcG loss-of-function mutations were observed, suggesting that CBX2-driven nucleosome compaction is a key mechanism by which PRCs maintain transcriptional repression during mouse development [57].
2.3. PRC1 in Transcription Activation
Traditionally, PRC1 is well known for its transcriptional repressive role. However, recent studies have found that PRC1 also functions as a transcriptional activator [58,59,60,61,62,63]. Genome-wide research of murine megakaryoblastic cells identified the PRC1 core component RING1B as sharing a large number of target gene loci with the transcription factor complex Runx1/CBFβ, and interestingly, knockdown of RING1B led not only to upregulation of target genes, but also to downregulation of target genes, suggesting that PRC1 might play a role in both transcriptional repression and transcriptional activation [64]. Subsequent studies of various mammalian cell types have implicated multiple components of PRC1, including PCGF1, CBX8, and RING1B in transcriptional activation of target genes [62,65,66,67]. However, the detailed molecular mechanisms of PRC1-mediated transcriptional activation are not fully understood as yet. Currently, the identified molecular mechanisms of PRC1 involved in transcriptional activation mainly include two aspects: the synergistic effect of PRC1 with other epigenetic regulators and the inhibition of intrinsic ubiquitination activity of PRC1 (Figure 2D,E) [58,59,60].
Studies in mouse neural progenitor cells have demonstrated that the AUTS2 subunit in ncPRC1.5 can recruit P300, a transcriptional coactivator and histone acetyltransferase that promotes acetylation of histone H3 at Lys27 (H3K27ac), facilitating transcriptional activation of genes [60]. In addition, Zhao et al. used a proteomic approach and promoter occupancy analysis to identify several novel PCGF3/5-interacting proteins, including testis expressed 10 (Tex10), which can directly contribute to transcriptional activation through P300 (Figure 2D) [59]. Furthermore, depletion of PCGF3/5 in ESCs significantly reduced the occupancy of Tex10 and P300 on target gene loci, indicating that PCGF3/5 acted as transcriptional activators through the interaction of Tex10 and P300 [59]. The mechanism by which P300 cooperates with ncPRC1.3/5 to activate gene transcription has been further confirmed in more recent studies [68,69].
As mentioned above, the E3 ubiquitin ligase activity of RING1B can be enhanced by several other PRC1 components, thereby promoting gene repression. In contrast, factors that inhibit RING1B-mediated H2AK119ub1 have been identified, which provides new insights into the mechanism of PRC1-mediated transcriptional activation [58,60]. Genome-wide ChIP-sequencing analysis (ChIP-seq) of mouse quiescent lymphoid B cells revealed that Aurora B kinase and cPRC1.4 colocalize at active promoters and they are required for RNAP binding to active promoters [58]. In addition, Aurora B kinase not only inhibits H2AK119ub1 by phosphorylating and inactivating the E2 enzyme UBE2D3 but also promotes H2A deubiquitination by phosphorylating and enhancing the activity of the deubiquitinating enzyme USP16 (Figure 2E, top) [58]. Further research subsequently uncovered an alternative mechanism for the inhibition of H2A mono-ubiquitination (Figure 2E, bottom). Interestingly, ncPRC1.3/5 tends to localize to gene loci lacking H2AK119ub1 compared to other PRC1 complexes, and in vitro reporter assays have identified a role for ncPRC1.3/5 in transcriptional activation [60]. Indeed, the AUTS2 subunit of the ncPRC1.5 complex recruited CK2, which then promoted gene transcriptional activation by binding to and phosphorylating Ser168 of RING1B, leading to a decrease in the E3 enzymatic activity of RING1B [60]. The reduction in PRC1 activity not only inhibits PRC2-mediated gene silencing, it also leads to the upregulation of transcriptional regulators that in turn activate target genes. For example, the reduction of H2AK119ub1 occupancy on chromatin disrupts the conformational equilibrium of RNAP, with an increase in Ser2-phosphorylated RNAP and a decrease in Ser5-phosphorylated RNAP, promoting its transcriptional initiation and elongation effects [41]. Decreased PRC1 activity would also deregulate low-level or inappropriate transcriptional signals from enhancers, thus promoting a transcriptional burst, rather than facilitating gene activation [45].
3. Composition of PRC2 and the Role It Plays in Transcription
PRC2 has generally been considered the relatively smaller of the two PRCs that have fewer PcG auxiliary subunits, but it is now becoming increasingly clear that, like PRC1, it also does not only exist as a single entity. Affinity purification coupled with tandem mass spectrometry (AP-MS) studies using a single PRC2 subunit as bait revealed that PRC2 comprises at least two distinct functional subcomplexes, referred to as PRC2.1 and PRC2.2 (Figure 1B) [70]. Here, we elaborate the composition of these two complexes and the function of these constituent subunits in PRC2-mediated transcriptional regulation.
3.1. Composition of PRC2 and Its Methyltransferase Activity
PRC2 is composed of four core subunits, EZH2 or its homologs EZH1, EED, SUZ12, and RBAP46/48 (also known as RBBP4/7), and is divided into PRC2.1 and PRC2.2, depending on their auxiliary subunits [71,72,73]. PRC2.1 includes a Polycomb-like (PCL) protein (PCL1/2/3), as well as Elongin BC and Polycomb repressive complex 2-associated protein (EPOP) or PRC2-associated LCOR isoform 1/2 (PALI1/2), whereas PRC2.2 includes Adipocyte enhancer binding protein 2 (AEBP2) and Jumanji and AT-rich interaction domain containing 2 (JARID2) [70,74,75,76].
PRC2 promotes chromatin compaction and gene silencing mainly through methylation (mono-, di-, and trimethylation) of H3K27 catalyzed by the enzyme subunits EZH1/2 [73,77]. Methylation of H3K27 is progressive (H3K27me3 is the result of mono-methylation of H3K27me2), and H3K27me3 is a stable mark [78]. In contrast to H3K27me3, the importance of H3K27me2 in maintaining gene repression appears limited [79]. However, H3K27me2 is an important intermediary PRC2 product that not only constitutes a substrate for subsequent H3K27me3 formation but may also prevent H3K27 from being acetylated. Acetylated H3K27 is thought to be antagonistic to PcG-mediated gene silencing and is enriched in the absence of PRC2 [80]. Unlike H3K27me2/3, H3K27me1 is still detectable in cells carrying non-functional PRC2 and its enrichment correlates with actively transcribed genes [81]. Exactly how H3K27me1 is generated is still an issue of debate. H3K27me1 may be catalyzed by PRC2, while its presence in actively transcribed genes also results from the demethylation of H3K27me2/3 by the demethylases lysine demethylase 6B (KDM6B) or (UTX histone demethylase) UTX [82].
Structural and biochemical analyses targeting PRC2 have focused on EZH2-containing complexes, which in most cases exhibit a more pronounced catalytic function for H3K27 methylation [73]. In PRC2, the stability of the core subunits EED, SUZ12, EZH2, and RBAP46/48 is strongly interdependent, and the steady state of the core complex is also necessary for PRC2 to function as a methyltransferase [83,84,85]. After EZH2 trimethylates H3K27, EED binds to H3K27me3 through its WD40 domain, causing the SET (Su(var)3-9, Enhancer-of-Zeste and Trithorax) domain of EZH2 to conform into the active conformation, allosterically activating its catalytic activity [86]. In addition, PRC2 can also spread to neighboring nucleosomes to exert methyltransferase activity by binding H3K27me3 via EED subunits [87,88,89]. SUZ12 associates with EZH2 and EED through its VEFS (VRN2-EMF2-FIS2-SUZ12) domain to promote PRC2 stabilization, and its interaction with PCL proteins play an important role in recruiting PRC2 to chromatin [90,91,92]. RBAP46/48 proteins can associate via its helix 1 with free histone H4, H3-H4 dimers, and tetramers, consistent with being required for PRC2 binding to unmodified nucleosomes, and they are also essential for PRC2 to fully exert its methyltransferase activity [83,93,94]. Importantly, although PRC2.1-specific and PRC2.2-specific subunits are not essential for PRC2 methyltransferase activity, recent studies have found that they also affect PRC2 activity (Figure 3) [95,96,97].
3.2. PRC2 in Transcription Regulation
PRC2-catalyzed H3K27me3 is generally considered a hallmark of gene silencing. In the field of developmental biology, there are two main types of H3K27me3-marked genes: (1) genes with both H3K27me3 and H3K4me3 are denoted “bivalent genes”, which are OFF but can be rapidly activated under subsequent cues [98]; (2) genes that have H3K27me3 and H2AK119ub1, which are OFF and appear to be more difficult to activate at subsequent stages.
Unlike PRC1, the catalytic activity of PRC2 is dependent on the presence and integrity of only four core subunits, and deletion of any one core subunit results in loss of catalytic activity with the remaining auxiliary subunits being nonessential for PRC2 activity but associated with their recruitment to chromatin and regulation of catalytic activity [6,71,72,96,97,99,100]. PRC2 is usually divided into two variants, PRC2.1 and PRC2.2, of which PRC2.1 can be further subdivided into EPOP-containing and PALI-containing PRC2 variants depending on the dissimilarity of its accessory subunits (Figure 1B) [5]. Both EPOP and PALI1/2 proteins individually interact with core PRC2 subunits, but their regulatory effects on PRC2 transcriptional repression function are mutually exclusive, with the presence of PALI1/2 enhancing its methylation enzymatic activity [70,74,97]. EPOP bridges PRC2.1 to Elongin B/C (EloB/C), which interacts with Elongin A (EloA) and promotes RNAP elongation, further inhibiting PRC2-mediated transcriptional repression (Figure 3A, bottom) [75,101]. In addition, both PRC2.1 variants have three PCL proteins: PCL1 (also referred to as PHF1), PCL2 (also referred to as MTF2), and PCL3 (also referred to as PHF19), which promote PRC2.1 recruitment to PcG-targeted gene loci and enhanced catalytic activity (Figure 3A, top) [79,102,103,104,105,106]. Of these, PCL2 promotes de novo recruitment of PRC2.1 by binding to unmethylated CpG islands, whereas PCL1/3 mediates PRC2.1 recruitment to chromatin by recognizing and binding H3K36me3 [106,107,108,109,110]. In particular, PCL1 can increase the residence time of PRC2.1 on chromatin, increasing H3K27me3 deposition at PcG-targeted gene loci [111]. Furthermore, PCL3 can recruit the demethylase NO66 for H3K27me3, helping to promote H3K36me3 removal and H3K27me3 deposition, thereby enhancing PRC2.1-mediated transcriptional repression [102].
Within PRC2.2, a core component of PRC2 associates with AEBP2 and JARID2, which synergistically and drastically increase the catalytic activity of EZH2 [112,113,114]. Recent studies have shown that the catalytic stimulation of EZH2 by JARID2 is partially mediated by the trimethylation of JARID2 at Lys116 (JARID2-K116me3): PRC2 first catalyzes JARID2-K116me3, then the binding of JARID2-K116me3 to the WD40 domain of EED, induces a conformational change of EZH2, leading to the increased catalytic activity of PRC2.2 (Figure 3B, top) [95,115]. However, the stimulation of EZH2 catalytic activity by AEBP2 is not fully understood, possibly because it increases the stability of PRC2.2, which increases EZH2 catalytic activity [116]. Moreover, AEBP2 stimulated PRC2.2 binding to nucleosomes by binding H2AK119ub1, thereby enhancing EZH2-mediated H3K27me3 deposition [52,96]. Interestingly, EZH inhibitory protein (EZHIP), a protein expressed predominantly in the gonads, was recently shown to interact with the allosterically activated PRC2.2 and inhibited its methyltransferase activity (Figure 3B, bottom) [117,118,119,120].
Collectively, the transcriptional repression exerted by PRC2 through catalyzing H3K27me3 requires the stable existing forms of the four core components, while other specific accessory subunits are also very important for its recruitment on chromatin and regulation of methyltransferase activity.
4. Polycomb Repressive Complexes in Cancer
Although gene repression mediated by the cooperation of PRC1 with PRC2 can essentially fully explain the role of PRCs in cancer, the sheer amount of data on these genes has been difficult to fully research, and thus the recent focus has shifted to investigate the roles played by subunits of PRCs in the context of cancer. Multiple components of PRCs play important roles in a variety of cancers, and they not only are implicated in cancer initiation and development but also function as tumor suppressors (Table 1 and Table 2).
4.1. Oncogenic Role of PRC1
PCGF4 (also known as BMI-1), the best-studied PRC1 gene in cancer, was one of the first PcG genes found in mammals and has been defined as a proto-oncogene that cooperates with the c-Myc oncoprotein to promote tumorigenesis [232,233,234,235]. The c-Myc protein can promote the transcription of BMI-1, thereby enhancing the transcriptional repression of genes such as p16 and p19ARF, which are tumor suppressors encoded by the Ink4a/ARF locus (Figure 4A) [131,236]. In addition, BMI-1 is overexpressed in gastric, colorectal, ovarian, breast, and other cancers, promoting cell proliferation and immortality by repressing transcription of the Ink4a/ARF locus [131,132,237,238,239]. However, silencing of this locus is unlikely to be the only mechanism by which it exerts its oncogenic effects, and BMI-1 has been found to promote stem cell expansion and tumorigenesis in an Ink4a/ARF independent manner in some cancers [28,135]. Post-translational modifications (PTMs) of BMI-1 have been reported to enhance its oncogenic effects. O-GlcNAcylation of BMI-1 at Ser255 mediated by O-GlcNAc transferase (OGT) increased the stability of the protein and its oncogenic activity (Figure 4B) [240]. Meanwhile, other homologous proteins of PCGF4 such as PCGF1, PCGF3, and PCGF6 have also been described to play oncogenic roles in various cancers (Figure 4C). Among them, PCGF1 was found to enhance stemness and promote cell proliferation in colorectal cancer (CRC) cells by activating the expression of genes including CD133, CD44, and ALDH1A1 [164]. PCGF3 was shown to promote proliferation and migration of non-small cell lung cancer (NSCLC) through the PI3K/Akt signaling pathway, while mutations in PCGF6 enhanced breast cancer cell migration and metastasis by upregulating the expression of epithelial-mesenchymal transition (EMT)-related genes [166,167].
Moreover, RING1B (also known as RNF2), the core catalytic subunit of PRC1, was found to be overexpressed and promoted oncogene expression [25,121]. It has been reported to have a dual role of gene repression and activation, which drove cell proliferation by activating the expression of CCND2 and promoted cell invasion by inhibiting the expression of LTBP2 (Figure 4D) [24]. Additionally, RING1B has also been shown to promote EMT and metastatic progression of cancer cells by activating the expression of ZEB2 and inhibiting the expression of E-cadherin, further demonstrating its important role in cancer development (Figure 4D) [24,122]. Recently, RING1B has been described to play multiple functions in basal-like and luminal breast cancer, where it elevated enhancer activity and promoted gene transcription [123]. Interestingly, RING1A and RING1B can also play oncogenic roles by denaturing other non-histone substrates, and both of them promote p53 protein degradation in colorectal, hepatocellular, and germ cell tumors [126,127]. In addition, RING1B can negatively regulate autophagy by binding Lys45 (K45) of DCAF3 in mouse embryonic fibroblasts (MEFs), which may also be implicated in exerting its oncogenic effects, but the specific mechanism remains to be further explored [241]. Not just the core subunits, but also numerous accessory subunits of PRC1 have also been shown to primarily play oncogenic roles (for a detailed description, see Table 1).
4.2. Tumor-Suppressive Role of PRC1
Generally, PRC1 is known for its oncogenic role. However, several studies have found that its components also exert tumor-suppressive effects. Most notably, unlike other PCGF homologs, PCGF2 (also known as MEL-18) has tumor-suppressive activity [31,128,129,242,243]. The expression levels of MEL-18 and BMI-1 are negatively correlated in various cancers [30,130]. Overexpression of MEL-18 leads to downregulation of c-Myc protein, a transcriptional activator of BMI-1, resulting in decreased levels of BMI-1 protein, promoting p16 and p19ARF upregulation, ultimately inhibiting cell proliferation and driving cellular senescence (Figure 4A) [236]. Meanwhile, MEL-18 also negatively regulated the ubiquitination activity of RING1B by inhibiting BMI-1 transcription [244]. In addition, previous studies have also shown that MEL-18 loss causes aggressive phenotypes in breast cancer, such as facilitating stem cell activity, angiogenesis, cell cycle progression, and EMT (Figure 4E) [31,128,129,243]. More recently, MEL-18 loss was described as mediating estrogen receptor-α (ER-α) downregulation, resulting in a hormone-independent phenotype of breast cancer associated with poor prognosis, implicating a key role in the hormonal regulation of breast cancer [245]. PHC3, another component of cPRC1, is also known to act as a tumor suppressor, and its frequent loss of heterozygosity (LoH) in osteosarcoma promotes tumorigenesis [140,141].
Importantly, multiple accessory subunits also act as tumor suppressors, such as CBX proteins, RYBP, KDM2B, and BCOR (Table 1). Among them, the CBX4, CBX6, and CBX7 proteins have dual roles as oncogenic and tumor suppressors. CBX proteins mainly regulate cell migration, proliferation, and EMT, but interestingly in different tumors, they play quite opposite roles [29,146,147,148,151,153]. This may be because the auxiliary subunits of PRC1 are dynamically assembled into different PRC1 in a context-dependent manner, and thus play different or even opposite roles in the occurrence and development of cancer, but the specific mechanisms need to be further investigated clearly [246]. Unlike the CBX proteins, RYBP plays a predominantly tumor-suppressive role and is oncogenic in only a few cancers [168,169].
In summary, whether PRC1 plays an oncogenic or tumor-suppressive role is mainly related to its function in cell proliferation, cell cycle progression, metastasis, and invasion (References are shown in Table 1). Given the prominent oncogenic role that PRC1 plays, inhibitors targeting PRC1 for antitumor therapy have also attracted attention, and a subset of inhibitors have been reported [247,248,249,250]. However, the precise roles of the various subunits of PRC1 in cancer remain to be defined, and future work should further delineate the molecular implications of these components in depth and identify appropriate therapeutic approaches to rescue their dysregulation in different cancers.
4.3. Oncogenic Role of PRC2
Currently, it has been reported that the pro-tumor activity of PRC2 is mainly associated with its EZH2, EED, and SUZ12 subunits. The first clinically relevant finding in the PcG protein field was that EZH2 promoted prostate cancer progression and poor prognosis [15]. It was subsequently identified as being downstream of the pRB-E2F pathway, which was essential for tumor cell proliferation [17]. Dysregulation of EZH2, as well as its roles, has been discussed in several solid malignancies including prostate, hepatocellular, colorectal, and breast cancer, as well as in some hematologic malignancies [251]. EZH2 is involved in cancer initiation and progression mainly due to its transcriptional repression activity in the PRC2 complex, and gain-of-function (GOF) mutants of EZH2 (Y647F/N, A677G, and A687V) are frequently generated in several lymphomas, further promoting tumorigenesis (Figure 5A) [19,20,189]. Similar to BMI1, PTMs of EZH2 have also been reported to enhance its oncogenic effects. Acetylation at Lys348 (K348) and O-GlcNAcylation at Ser73 (S73) increased the protein stability and catalytic activity of EZH2, further enhancing its ability to promote cancer cell migration and invasion (Figure 5B) [252,253]. In particular, EZH2 may exert oncogenic effects not only by mediating H3K27me3, but also by methylating non-histone proteins or binding to other proteins in a PRC2-independent manner. Recent studies have found that EZH2 can methylate the transcription factor RORα at Lys38 (K38), promoting the degradation of RORα and reducing RORα-mediated activation of gene transcription. Interestingly, the levels of EZH2 and RORα were inversely correlated in breast cancer, implying that EZH2 might play an oncogenic role by inhibiting RORα-mediated tumor suppression [254]. In contrast, phosphorylated EZH2 methylates the transcription factor STAT3 at Lys180 (K180), which enhances STAT3-mediated transcriptional activation but also promotes cancer stem cell self-renewal and exerts oncogenic effects (Figure 5C) [255]. Moreover, EZH2 is able to directly interact with β-catenin and ERα in a PRC2-independent manner to activate cyclin D1 and c-Myc expression upon estrogen stimulation to induce the proliferation of breast cancer cells (Figure 5D) [256]. Comparably, the β-catenin activation complex can also recruit EZH2 via PCNA associating factor (PAF) to activate the Wnt signaling pathway that drives tumorigenesis (Figure 5D) [257]. However, the specific mechanism by which EZH2 activates gene expression in a PRC2-independent manner remains unclear. Taken together, EZH2, whether acting in a PRC2-dependent or PRC2-independent manner for gene regulation, has been described to be closely associated with tumorigenesis and progression.
Additionally, EED and SUZ12 are upregulated in multiple cancers, including lymphoma, breast cancer, head, and neck squamous cell carcinoma (NHSCC), and colorectal cancer [200,201,206]. The knockdown of SUZ12 significantly inhibited cell proliferation, invasion, and migration in HNSCC cells and inhibited xenograft tumor growth [206]. However, the upregulation of EED and SUZ12 is often accompanied by the upregulation of EZH2, exerting oncogenic effects mainly through PRC2-mediated gene silencing [200,201]. In addition, the oncogenic roles played by other subunits of PRC2, such as PCL1/2/3 and EPOP have also been linked to their regulation of PRC2 activity or promotion of PRC2 recruitment to chromatin (Table 2) [75,215,219,222]. In contrast, RBAP46 bound the transcription factor Sp1 in a PRC2-independent manner, resulting in the downregulation of reversion-inducing cysteine-rich protein with Kazal motifs (RECK), a protein that suppresses tumor metastasis and angiogenesis [213].
4.4. Tumor-Suppressive Role of PRC2
Although PRC2 has oncogenic properties in most tumors, it has been shown that EZH2 and other PRC2 subunits also have tumor-suppressive functions in some types of tumors [23,191,209,230]. In contrast to GOF mutants, loss-of-function (LOF) mutants of EZH2 have also been identified in T-cell acute lymphoblastic leukemia (T-ALL). LOF mutants of EZH2 (G266E, T393M, and C606Y) that result in loss of PRC2 function and drive Notch signaling activation, and increase the in vivo tumorigenic potential of T-ALL cells, suggesting that PRC2 may have a tumor-suppressor function, although the specific mechanisms remain to be further explored (Figure 5A) [192]. Recently, PTMs of EZH2 have also been shown to exert tumor-suppressive effects. Methylation at Lys735 (K735) and Phosphorylation at Thr261 (T261) increased the protein stability and catalytic activity of EZH2, further attenuating its transcriptional repression on tumor suppressors (Figure 5B) [258,259]. Moreover, in malignant peripheral nerve sheath tumors (MPNSTs), there is the frequent deletion of PRC2 subunit genes, which also leads to loss of H3K27me2 and H3K27me3 and is associated with poor prognosis [23,260] On the other hand, by exogenously expressing the deleted PRC2 subunit in MPNST cells with frequent deletion of the PRC2 gene, H3K27me3 levels were increased and cell proliferation was inhibited [23]. Furthermore, treatment with EZH2 inhibitors had no effect on the proliferation of MPNST cells [198]. Similarly, SUZ12 loss in T-ALL, which results in decreased gene silencing function of the PRC2 complex, promotes oncogene upregulation [261]. These results further support that PRC2 has tumor suppressor properties in specific cancers.
Interestingly, deletion of EED in mice resulted in hyperproliferation of myeloid progenitors and lymphoid cells, which accelerated lymphoid tumor formation after exposure of mice to genotoxic drugs, although this did not induce tumorigenesis [262,263]. Likewise, EZH2 or SUZ12 deletion accelerated Myc-driven lymphomagenesis by limiting self-renewal of B cell progenitors [264]. In addition, EZH2 loss significantly promoted the development of myelodysplastic syndrome induced by transcription factor Runx1 mutation, and loss of SUZ12 synergizes with neurofibromin 1 (NF1) mutations to amplify Ras signaling to drive cancer [210,265,266].
In conclusion, in addition to the partial non-canonical function of EZH2, PRC2 mainly exerts oncogenic or tumor-suppressive effects through its gene silencing function, depending on the cancer type. Both enhancement and attenuation of PRC2 catalytic activity can promote tumor development, suggesting that PcG genes are context-dependent tumor suppressors or oncogenes. Further observations of the context-dependent roles of PRC2 have revealed that the effects of loss-of-function and gain-of-function alterations do not simply segregate based on tissue or tumor type [267]. Furthermore, several studies have shown that the role of PRC2 in cancer depends on tumorigenic alterations in other genes [268,269]. For example, in NSCLC with loss-of-function mutations in BRG1 (W764R) or gain-of-function mutations in EGFR (T790M and L858R), inhibition of EZH2 catalytic activity promotes apoptosis and sensitivity to topoisomerase II (TopoII) inhibitors. Conversely, in BRG1 wild-type tumors, inhibition of EZH2 upregulates BRG1 and eventually confers stronger resistance to TopoII inhibitors [268]. Furthermore, in a Kras-driven mouse model of NSCLC, EED loss accelerated or delayed tumor formation depending on p53. In a WT-p53 background, EED loss promotes inflammation, whereas p53 inactivation leads to invasive mucinous adenocarcinoma [269]. Thus, the context-dependent role of PRC2 suggests that its function in specific cancer types is enormously complex and future work will be beneficial to exhaustively characterize its molecular implications in different cancers that will also help in identifying appropriate approaches to reverse their deregulation in different cells and provide suitable therapies for PRC2-dependent cancers. It is also notable that certain tumors are addicted to specific PRC2 subunits, independent of other components, the reason for which also remains ambiguous. Clearly, a better understanding of the cell-type-specific functions of each PRC2 subunit will require future research.
4.5. Development of Inhibitors Targeting Polycomb Repressive Complexes
PcG components have been reported to be associated with the growth and survival of different tumors, considered as targets for cancer therapy, and extensively explored [270,271]. Several PRC1-related inhibitors including PRT4165, PTC-209, IFM-11958, and RB-3 have been reported to date, but no inhibitor has entered clinical trials (Figure 6) [247,248,249,250]. PRT4165 was shown to inhibit the E3 enzymatic activity of PRC1, whereas PTC-209 and IFM-11958 were described as inhibitors of the BMI-1 expression. Furthermore, PTC-209 treatment significantly inhibits proliferation and promotes apoptosis in multiple myeloma (MM) cells, suggesting that BMI-1 may serve as an attractive anti-tumor drug target [248]. However, certain questions remain to be addressed regarding these inhibitors. Although PRT4165 was shown to be able to inhibit PRC1-mediated H2A ubiquitination, the mechanism by which it directly or indirectly inhibits the catalytic activity of PRC1 is not well elucidated. PTC-209 and IFM-11958 have been described to exert antitumor effects by inhibiting BMI-1 expression, but whether this effect is through inhibition of the function of PRC1 or other PRC1-independent functions of BMI-1 is unclear. Therefore, there is a critical need to develop inhibitors that specifically target PRC1. RB-3 was subsequently developed to inhibit the binding of RING1B and BMI-1, while specifically inhibiting the catalytic activity of PRC1 [247]. Importantly, RB-3 treatment drastically reduced the global level of ubiquitination of H2A and induced differentiation of leukemia cell lines, implying that inhibitors of PRC1 might be used to carry out the treatment of leukemia [247]. However, although numerous accessory subunits of PRC1 are oncogenic, PRC1 exerts oncogenic functions not only by repressing tumor suppressors but also by activating oncogenes, so it remains to be explored whether PRC1 components are suitable therapeutic targets [40,67,123,131].
Unlike PRC1, PRC2 exerts oncogenic effects mainly dependent on its transcriptional repression properties [5]. Therefore, the development of an inhibitor that targets PRC2, either by inhibiting its methyltransferase activity or by interfering with the stability of the complexes, could be a promising strategy for the treatment of PRC2-dependent tumors [182,271]. One of the first therapeutic agents reported to target PRC2 was 3-deazaneplanocin A (DZNep), which reduces H3K27me3 levels and causes apoptosis in cancer cells [272,273]. However, subsequent studies showed that instead of specifically inhibiting PRC2, DZNep reduced H3K27me3 levels by altering other histone methylation (H4K20me3) levels [274]. Medicinal chemists then have developed EZH2-specific enzymatic inhibitors including GSK126 and EPZ005687, both of them act by binding competitively to the SET domain of EZH2 and they have shown better selectivity and greater potency compared to DZNep [188,275]. Furthermore, both compounds not only target wild-type EZH2, but are also extremely sensitive to lymphoma cells harboring EZH2-activating mutations [188,275]. To date, multiple inhibitors targeting EZH2 have been reported, of which Tazemetostat was approved by the Food and Drug Administration (FDA) in January 2020 for the treatment of advanced epithelioid sarcoma and follicular lymphoma [276,277]. There are also many potent and more bioavailable EZH2 catalytic inhibitors currently undergoing phase 1/2/3 clinical trials, alone or in combination with other drugs, for the treatment of several solid tumors, mainly lymphoma, prostate cancer, and small cell lung cancer (Table 3). In addition, Tazemetostat is being used in several phase 2 clinical trials for the treatment of diffuse large B-cell lymphoma, hematologic neoplasms, mantle cell lymphoma, and peripheral nerve sheath tumors (NCT05205252, NCT04917042). Constellation pharmaceuticals has also tested multiple EZH2 inhibitors in clinical trials, including CPI-1205 and CPI-1205; CPI-1205 (NCT03480646), as their first-generation EZH2 inhibitor, is currently in phase 2 testing for metastatic castration-resistant prostate cancer, while a second-generation EZH2 inhibitor (CPI-0209) is also being tested in phase 2 for solid tumors (NCT04104776). An EZH2 inhibitor (PF-06821497) developed by Pfizer has entered phase 1 testing in patients with small cell lung cancer, castration-resistant prostate cancer, and follicular lymphoma (NCT03460977). As discussed earlier, the integrity and stability of PRC2 is essential for its methyltransferase activity, which provides additional possibilities for the development of inhibitors targeting PRC2. In addition to EZH2 catalytic inhibitors, an alternative strategy is to inhibit the allosteric activation of EZH2 enzymatic activity by preventing the interaction of EED-H3K27me3 using small molecule inhibitors (Figure 6) [203,278,279]. A potential advantage of this approach is that the specificity of the EED-H3K27me3 interaction may increase the selectivity for inhibition of PRC2, whereas other SET-domain-containing methyltransferases may be targeted by poorly selective EZH2 enzymatic inhibitors. EED226 was one of the first EED inhibitors reported and was found to specifically inhibit H3K27 methylation, either in lymphoma cells harboring WT-EZH2 or EZH2 mutants [278]. Furthermore, gene expression microarray and Chip-PCR experiments revealed that PRC2-regulated gene expression was significantly increased by treatment with EED226 and was similar to that of EZH2 catalytic inhibitor (EI1) treatment, demonstrating that EED226 also has a significant inhibitory effect on the gene-silencing function of PRC2 [278]. Interestingly, although EED226 failed to enter clinical trials, another EED inhibitor, MAK683, developed on the basis of this scaffold, is currently in phase 2 clinical trials for the treatment of lymphoma (Table 3) [280]. Moreover, EEDi-5285 is the most potent EED inhibitor that has been reported, and it achieves complete and long-lasting tumor regression in mice, further suggesting great potential for the treatment of PRC2-dependent cancers by inhibiting the EED-H3K27me3 interaction [279]. Other approaches to inhibiting PRC2 activity in cancer include a staple peptide inhibitor that disrupts the protein-protein interaction (PPI) of EZH2-EED, with significant antiproliferative functions even in cells resistant to EZH2 inhibitors [281]. Subsequent studies reported that the FDA-approved drug astemizole and the natural compound wedelolactone could also inhibit PRC2 activity by blocking PPI of EZH2-EED, but these inhibitors were less potent than known EZH2 inhibitors or EED inhibitors [282,283].
5. Concluding Remarks and Perspective
PRCs have attracted increasing attention because of their important role in a wide range of physiological and pathological processes, especially in cancer treatment (Table 1 and Table 2). The gradually discovered new members of PcG in the complexity of PRCs open new perspectives for research in this field. In addition, the identification and characterization of multiple PRC variants have also provided a new understanding of the specific mechanisms by which PRC1 and PRC2 exert catalytic activity. In this review, we have discussed the currently identified PRC1 variants and PRC2 variants and classified them separately using a similar classification system (Figure 1). This classification is based on the relatively new but robust and cross-validated biochemical experimental results reported so far. Although components of different PRC variants have been reported, important aspects such as their abundance, stoichiometry, assembly of different variants, and an exact number of PRC variants formed in the cell remain to be addressed. Importantly, further exploration of the role of newly identified accessory subunits may have significant implications for complementing the function of PRC variants.
PcG proteins are involved in highly dynamic biological processes by regulating the transcription of genes. Among them, PRC1 is reported to have dual roles of gene silencing and activation, which may be related to its enormous complexity and a large number of subunits. It will be valuable to expend effort to uncover the molecular principles underlying PRCs gene regulation and decipher their mechanisms in Polycomb-related disorders. In particular, PcG proteins are hallmark components of cancer research, and many agents targeting PRCs have been developed and have advanced into clinical trials (Table 3). Considering that PRCs may play opposite functions in different cancers [284], understanding the specific role of each complex variant in a specific cancer type is even more critical and will help to develop effective therapeutic strategies.
Despite our established understanding of PRCs, it is clear that we are only beginning to unravel the possibilities behind these rather complex epigenetic complexes, with a plethora of PcG proteins whose functions in cancer have yet to be discovered and many questions remain to be addressed. Current studies have found both canonical and non-canonical activities and functions of PRCs in tumor initiation and progression, but what are their exact contributions? PMTs of multiple subunits of PRCs were shown to render them functionally altered, but what are the specific mechanisms that trigger the occurrence of these PMTs in cancer? PRCs have been reported to harbor a large number of variants, but is there functional crosstalk between these different variants in specific cancers? If present, what are the specific functional crosstalk mechanisms? The physiological, pathological, and therapeutic possibilities behind such epigenetic complexes remain to be revealed with significant effort. With the remarkable development of our research techniques and methods, the coming years will inevitably be fast-paced and exciting years to explore and define PRCs.
Author Contributions
G.-J.D. wrote and revised the manuscript; J.-L.X. and Y.-R.Q. collected the literature; Z.-Q.Y. and W.Z. revised and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (Grant No. 81870297).
Conflicts of Interest
The authors declare no competing financial interest.
Abbreviations
Enhancer of zeste homolog 1/2, EZH1/2; Embryonic ectoderm development, EED; Suppressor of zeste 12, SUZ12; Retinoblastoma protein-associated proteins 46/48, RBAP46/48; YY1-associated factor 2, YAF2; RING1 and YY1 binding protein, RYBP; Scm polycomb group protein homolog 1/2, SCMH1/2; Scm polycomb group protein like 1, SCML1; BCL6 corepressor, BCOR; Lysine-specific demethylase 2B, KDM2B; S-phase kinase-associated protein 1, SKP1; Ubiquitin carboxyl-terminal hydrolase 7, USP7; BCL6 Corepressor Like 1, BCORL1; Autism susceptibility protein 2, AUTS2; Fibrosin, FBRS; Fibrosin Like 1, FBRSL1; Casein Kinase 2, CK2; Histone deacetylase 1/2, HDAC1/2; Dimerization partner 1/2, DP-1/2; E2F transcription factor 6, E2F6; MYC-associated factor X, MAX; L3MBTL histone methyl-lysine binding protein 2, L3MBTL2; MAX gene-associated protein, MGA; WD repeat domain 5, WDR5; Polycomblike 1/2/3, PCL1/2/3; Elongin BC and PRC2-associated protein, EPOP; PRC2-associated LCOR isoform 1/2, PALI1/2; Jumanji and AT-rich interaction domain containing 2, JARID2; Adipocyte enhancer binding protein 2, AEBP2; Sex Combs Extra, Sce; Polycomb, Pc; Polyhomeotic, Ph; Posterior Sex Combs, Psc; Sex Comb on Midleg, Scm; Polycomp group, PcG; Polycomb repressive complex, PRC; RNA polymerase II, RNAP; Facilitates Chromatin Transcription, FACT; EZH inhibitory protein, EZHIP; Elongin B/C, EloB/C; Elongin A, EloA; O-GlcNAc transferase, OGT; Cyclin D2, CCND2; Zinc finger E-box binding homeobox 2, ZEB2; Latent transforming growth factor beta binding protein 2, LTBP2; Hypoxia inducible factor 1 subunit alpha, HIF1α; Estrogen receptor-α, ER-α; P300/CBP-associated factor, PCAF; SET domain containing 2, SETD2; Cyclin dependent kinase 5, CDK5; Signal transducer and activator of transcription 3, STAT3; Suppressor of cytokine signaling 3, SOCS3.
References
- Helin, K.; Dhanak, D. Chromatin proteins and modifications as drug targets. Nature 2013, 502, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Gerd, J. A group of genes controlling spatial expression of the bithorax complex in Drosophila. Nature 1985, 316, 153–155. [Google Scholar]
- Levine, S.S.; Weiss, A.; Erdjument-Bromage, H.; Shao, Z.; Tempst, P.; Kingston, R.E. The core of the polycomb repressive complex is compositionally and functionally conserved in flies and humans. Mol. Cell. Biol. 2002, 22, 6070–6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piunti, A.; Shilatifard, A. The roles of Polycomb repressive complexes in mammalian development and cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Wang, L.; Wang, H.; Xia, L.; Erdjument-Bromage, H.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science 2002, 298, 1039–1043. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, L.; Erdjument-Bromage, H.; Vidal, M.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of histone H2A ubiquitination in Polycomb silencing. Nature 2004, 431, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef]
- Bracken, A.P.; Dietrich, N.; Pasini, D.; Hansen, K.H.; Helin, K. Genome-wide mapping of Polycomb target genes unravels their roles in cell fate transitions. Genes Dev. 2006, 20, 1123–1136. [Google Scholar] [CrossRef] [Green Version]
- Fursova, N.A.; Blackledge, N.P.; Nakayama, M.; Ito, S.; Koseki, Y.; Farcas, A.M.; King, H.W.; Koseki, H.; Klose, R.J. Synergy between Variant PRC1 Complexes Defines Polycomb-Mediated Gene Repression. Mol. Cell 2019, 74, 1020–1036.e8. [Google Scholar] [CrossRef] [Green Version]
- Geng, Z.; Gao, Z. Mammalian PRC1 Complexes: Compositional Complexity and Diverse Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 8594. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, J.; Bonasio, R.; Strino, F.; Sawai, A.; Parisi, F.; Kluger, Y.; Reinberg, D. PCGF homologs, CBX proteins, and RYBP define functionally distinct PRC1 family complexes. Mol. Cell 2012, 45, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Mierlo, G.; Veenstra, G.J.C.; Vermeulen, M.; Marks, H. The Complexity of PRC2 Subcomplexes. Trends Cell Biol. 2019, 29, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varambally, S.; Dhanasekaran, S.M.; Zhou, M.B.T.; Kumar-Sinha, C.; Sanda, M.G.G.D.; Pienta, K.J.; Sewalt, R.G.; Otte, A.P.; Rubin, M.A.; Chinnaiyan, A.M. The polycomb group protein EZH2 is involved in progression of prostate cancer. Nature 2002, 419, 624–629. [Google Scholar] [CrossRef]
- Kleer, C.G.; Cao, Q.; Varambally, S.; Shen, R.; Ota, I.; Tomlins, S.A.; Ghosh, D.; Sewalt, R.G.; Otte, A.P.; Hayes, D.F.; et al. EZH2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 11606–11611. [Google Scholar] [CrossRef] [Green Version]
- Bracken, A.P.; Pasini, D.; Capra, M.; Prosperini, E.; Colli, E.; Helin, K. EZH2 is downstream of the pRB-E2F pathway, essential for proliferation and amplified in cancer. EMBO J. 2003, 22, 5323–5335. [Google Scholar] [CrossRef] [Green Version]
- Morin, R.D.; Johnson, N.A.; Severson, T.M.; Mungall, A.J.; An, J.; Goya, R.; Paul, J.E.; Boyle, M.; Woolcock, B.W.; Kuchenbauer, F.; et al. Somatic mutations altering EZH2 (Y641) in follicular and diffuse large B-cell lymphomas of germinal-center origin. Nat. Genet. 2010, 42, 181–185. [Google Scholar] [CrossRef]
- Yap, D.B.; Chu, J.; Berg, T.; Schapira, M.; Cheng, S.W.; Moradian, A.; Morin, R.D.; Mungall, A.J.; Meissner, B.; Boyle, M.; et al. Somatic mutations at EZH2 Y641 act dominantly through a mechanism of selectively altered PRC2 catalytic activity, to increase H3K27 trimethylation. Blood 2011, 117, 2451–2459. [Google Scholar] [CrossRef] [Green Version]
- McCabe, M.T.; Graves, A.P.; Ganji, G.; Diaz, E.; Halsey, W.S.; Jiang, Y.; Smitheman, K.N.; Ott, H.M.; Pappalardi, M.B.; Allen, K.E.; et al. Mutation of A677 in histone methyltransferase EZH2 in human B-cell lymphoma promotes hypertrimethylation of histone H3 on lysine 27 (H3K27). Proc. Natl. Acad. Sci. USA 2012, 109, 2989–2994. [Google Scholar] [CrossRef] [Green Version]
- Score, J.; Hidalgo-Curtis, C.; Jones, A.V.; Winkelmann, N.; Skinner, A.; Ward, D.; Zoi, K.; Ernst, T.; Stegelmann, F.; Dohner, K.; et al. Inactivation of polycomb repressive complex 2 components in myeloproliferative and myelodysplastic/myeloproliferative neoplasms. Blood 2012, 119, 1208–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Sanada, M.; Matsui, H.; Yamasaki, N.; Honda, Z.I.; Shih, L.Y.; Mori, H.; Inaba, T.; Ogawa, S.; Honda, H. EED mutants impair polycomb repressive complex 2 in myelodysplastic syndrome and related neoplasms. Leukemia 2012, 26, 2557–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Teckie, S.; Wiesner, T.; Ran, L.; Prieto Granada, C.N.; Lin, M.; Zhu, S.; Cao, Z.; Liang, Y.; Sboner, A.; et al. PRC2 is recurrently inactivated through EED or SUZ12 loss in malignant peripheral nerve sheath tumors. Nat. Genet. 2014, 46, 1227–1232. [Google Scholar] [CrossRef] [Green Version]
- Rai, K.; Akdemir, K.C.; Kwong, L.N.; Fiziev, P.; Wu, C.J.; Keung, E.Z.; Sharma, S.; Samant, N.S.; Williams, M.; Axelrad, J.B.; et al. Dual Roles of RNF2 in Melanoma Progression. Cancer Discov. 2015, 5, 1314–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Boom, V.; Maat, H.; Geugien, M.; Rodriguez Lopez, A.; Sotoca, A.M.; Jaques, J.; Brouwers-Vos, A.Z.; Fusetti, F.; Groen, R.W.; Yuan, H.; et al. Non-canonical PRC1.1 Targets Active Genes Independent of H3K27me3 and Is Essential for Leukemogenesis. Cell Rep. 2016, 14, 332–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, L.J.; Watson, Z.L.; Qamar, L.; Yamamoto, T.M.; Post, M.D.; Berning, A.A.; Spillman, M.A.; Behbakht, K.; Bitler, B.G. CBX2 identified as driver of anoikis escape and dissemination in high grade serous ovarian cancer. Oncogenesis 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qin, G.; Liang, X.; Wang, W.; Wang, Z.; Liao, D.; Zhong, L.; Zhang, R.; Zeng, Y.X.; Wu, Y.; et al. Targeting the CK1alpha/CBX4 axis for metastasis in osteosarcoma. Nat. Commun. 2020, 11, 1141. [Google Scholar] [CrossRef] [Green Version]
- Bruggeman, S.W.; Hulsman, D.; Tanger, E.; Buckle, T.; Blom, M.; Zevenhoven, J.; van Tellingen, O.; van Lohuizen, M. Bmi1 controls tumor development in an Ink4a/Arf-independent manner in a mouse model for glioma. Cancer Cell 2007, 12, 328–341. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Guan, X.; Gong, L.; Zeng, J.; Zhang, H.; Chen, M.Y.; Li, G. CBX6 is negatively regulated by EZH2 and plays a potential tumor suppressor role in breast cancer. Sci. Rep. 2019, 9, 197. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.J.; Zeng, M.S.; Yadav, A.; Song, L.B.; Guo, B.H.; Band, V.; Dimri, G.P. Mel-18 acts as a tumor suppressor by repressing Bmi-1 expression and down-regulating Akt activity in breast cancer cells. Cancer Res. 2007, 67, 5083–5089. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Jang, K.-S.; Shin, D.-H.; Oh, M.-Y.; Kim, H.-J.; Kim, Y.; Kong, G. Mel-18 Negatively Regulates INK4a/ARF-Independent Cell Cycle Progression via Akt Inactivation in Breast Cancer. Cancer Res. 2008, 68, 4201–4209. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Raible, F.; Mollaaghababa, R.; Guyon, J.R.; Wu, C.-T.; Bender, W.; Kingston, R.E. Stabilization of Chromatin Structure by PRC1, a Polycomb Complex. Cell 1999, 98, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Tavares, L.; Dimitrova, E.; Oxley, D.; Webster, J.; Poot, R.; Demmers, J.; Bezstarosti, K.; Taylor, S.; Ura, H.; Koide, H.; et al. RYBP-PRC1 complexes mediate H2A ubiquitylation at polycomb target sites independently of PRC2 and H3K27me3. Cell 2012, 148, 664–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morey, L.; Aloia, L.; Cozzuto, L.; Benitah, S.A.; Di Croce, L. RYBP and Cbx7 define specific biological functions of polycomb complexes in mouse embryonic stem cells. Cell Rep. 2013, 3, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackledge, N.P.; Farcas, A.M.; Kondo, T.; King, H.W.; McGouran, J.F.; Hanssen, L.L.P.; Ito, S.; Cooper, S.; Kondo, K.; Koseki, Y.; et al. Variant PRC1 complex-dependent H2A ubiquitylation drives PRC2 recruitment and polycomb domain formation. Cell 2014, 157, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Rose, N.R.; King, H.W.; Blackledge, N.P.; Fursova, N.A.; Ember, K.J.; Fischer, R.; Kessler, B.M.; Klose, R.J. RYBP stimulates PRC1 to shape chromatin-based communication between Polycomb repressive complexes. Elife 2016, 5, e18591. [Google Scholar] [CrossRef]
- Simon, J.A.; Kingston, R.E. Occupying chromatin: Polycomb mechanisms for getting to genomic targets, stopping transcriptional traffic, and staying put. Mol. Cell 2013, 49, 808–824. [Google Scholar] [CrossRef] [Green Version]
- Schuettengruber, B.; Bourbon, H.M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef] [Green Version]
- Francis, N.J.; Kingston, R.E.; Woodcock, C.L. Chromatin compaction by a polycomb group protein complex. Science 2004, 306, 1574–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scelfo, A.; Fernandez-Perez, D.; Tamburri, S.; Zanotti, M.; Lavarone, E.; Soldi, M.; Bonaldi, T.; Ferrari, K.J.; Pasini, D. Functional Landscape of PCGF Proteins Reveals Both RING1A/B-Dependent-and RING1A/B-Independent-Specific Activities. Mol. Cell 2019, 74, 1037–1052.e7. [Google Scholar] [CrossRef] [Green Version]
- Stock, J.K.; Giadrossi, S.; Casanova, M.; Brookes, E.; Vidal, M.; Koseki, H.; Brockdorff, N.; Fisher, A.G.; Pombo, A. Ring1-mediated ubiquitination of H2A restrains poised RNA polymerase II at bivalent genes in mouse ES cells. Nat. Cell Biol. 2007, 9, 1428–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endoh, M.; Endo, T.A.; Endoh, T.; Isono, K.; Sharif, J.; Ohara, O.; Toyoda, T.; Ito, T.; Eskeland, R.; Bickmore, W.A.; et al. Histone H2A mono-ubiquitination is a crucial step to mediate PRC1-dependent repression of developmental genes to maintain ES cell identity. PLoS Genet. 2012, 8, e1002774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Zhu, P.; Wang, J.; Pascual, G.; Ohgi, K.A.; Lozach, J.; Glass, C.K.; Rosenfeld, M.G. Histone H2A monoubiquitination represses transcription by inhibiting RNA polymerase II transcriptional elongation. Mol. Cell 2008, 29, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phatnani, H.P.; Greenleaf, A.L. Phosphorylation and functions of the RNA polymerase II CTD. Genes Dev. 2006, 20, 2922–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrinic, P.; Szczurek, A.T.; Klose, R.J. PRC1 drives Polycomb-mediated gene repression by controlling transcription initiation and burst frequency. Nat. Struct. Mol. Biol. 2021, 28, 811–824. [Google Scholar] [CrossRef]
- Grau, D.J.; Chapman, B.A.; Garlick, J.D.; Borowsky, M.; Francis, N.J.; Kingston, R.E. Compaction of chromatin by diverse Polycomb group proteins requires localized regions of high charge. Genes Dev. 2011, 25, 2210–2221. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Brown, J.L.; Cao, R.; Zhang, Y.; Kassis, J.A.; Jones, R.S. Hierarchical Recruitment of Polycomb Group Silencing Complexes. Mol. Cell 2004, 14, 637–646. [Google Scholar] [CrossRef]
- Cao, R.; Tsukada, Y.; Zhang, Y. Role of Bmi-1 and Ring1A in H2A ubiquitylation and Hox gene silencing. Mol. Cell 2005, 20, 845–854. [Google Scholar] [CrossRef]
- Min, J.; Zhang, Y.; Xu, R.M. Structural basis for specific binding of Polycomb chromodomain to histone H3 methylated at Lys 27. Genes Dev. 2003, 17, 1823–1828. [Google Scholar] [CrossRef] [Green Version]
- Schoeftner, S.; Sengupta, A.K.; Kubicek, S.; Mechtler, K.; Spahn, L.; Koseki, H.; Jenuwein, T.; Wutz, A. Recruitment of PRC1 function at the initiation of X inactivation independent of PRC2 and silencing. EMBO J. 2006, 25, 3110–3122. [Google Scholar] [CrossRef]
- Sugishita, H.; Kondo, T.; Ito, S.; Nakayama, M.; Yakushiji-Kaminatsui, N.; Kawakami, E.; Koseki, Y.; Ohinata, Y.; Sharif, J.; Harachi, M.; et al. Variant PCGF1-PRC1 links PRC2 recruitment with differentiation-associated transcriptional inactivation at target genes. Nat. Commun. 2021, 12, 5341. [Google Scholar] [CrossRef] [PubMed]
- Kalb, R.; Latwiel, S.; Baymaz, H.I.; Jansen, P.W.; Muller, C.W.; Vermeulen, M.; Muller, J. Histone H2A monoubiquitination promotes histone H3 methylation in Polycomb repression. Nat. Struct. Mol. Biol. 2014, 21, 569–571. [Google Scholar] [CrossRef]
- Cooper, S.; Dienstbier, M.; Hassan, R.; Schermelleh, L.; Sharif, J.; Blackledge, N.P.; De Marco, V.; Elderkin, S.; Koseki, H.; Klose, R.; et al. Targeting polycomb to pericentric heterochromatin in embryonic stem cells reveals a role for H2AK119u1 in PRC2 recruitment. Cell Rep. 2014, 7, 1456–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskeland, R.; Leeb, M.; Grimes, G.R.; Kress, C.; Boyle, S.; Sproul, D.; Gilbert, N.; Fan, Y.; Skoultchi, A.I.; Wutz, A.; et al. Ring1B compacts chromatin structure and represses gene expression independent of histone ubiquitination. Mol. Cell 2010, 38, 452–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, S.; Ji, F.; Sunwoo, H.; Jain, G.; Lee, J.T.; Sadreyev, R.I.; Dekker, J.; Kingston, R.E. Polycomb Repressive Complex 1 Generates Discrete Compacted Domains that Change during Differentiation. Mol. Cell 2017, 65, 432–446e435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plys, A.J.; Davis, C.P.; Kim, J.; Rizki, G.; Keenen, M.M.; Marr, S.K.; Kingston, R.E. Phase separation of Polycomb-repressive complex 1 is governed by a charged disordered region of CBX2. Genes Dev. 2019, 33, 799–813. [Google Scholar] [CrossRef] [Green Version]
- Lau, M.S.; Schwartz, M.G.; Kundu, S.; Savol, A.J.; Wang, P.I.; Marr, S.K.; Grau, D.J.; Schorderet, P.; Sadreyev, R.I.; Tabin, C.J.; et al. Mutation of a nucleosome compaction region disrupts Polycomb-mediated axial patterning. Science 2017, 355, 1081–1084. [Google Scholar] [CrossRef] [Green Version]
- Frangini, A.; Sjoberg, M.; Roman-Trufero, M.; Dharmalingam, G.; Haberle, V.; Bartke, T.; Lenhard, B.; Malumbres, M.; Vidal, M.; Dillon, N. The aurora B kinase and the polycomb protein ring1B combine to regulate active promoters in quiescent lymphocytes. Mol. Cell 2013, 51, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Huang, Y.; Zhang, J.; Liu, M.; Ji, H.; Wang, C.; Cao, N.; Li, C.; Xia, Y.; Jiang, Q.; et al. Polycomb group RING finger proteins 3/5 activate transcription via an interaction with the pluripotency factor Tex10 in embryonic stem cells. J. Biol. Chem. 2017, 292, 21527–21537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Lee, P.; Stafford, J.M.; von Schimmelmann, M.; Schaefer, A.; Reinberg, D. An AUTS2-Polycomb complex activates gene expression in the CNS. Nature 2014, 516, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Geng, Z.; Gong, Y.; Warren, K.; Zheng, H.; Imamura, Y.; Gao, Z. WDR68 is essential for the transcriptional activation of the PRC1-AUTS2 complex and neuronal differentiation of mouse embryonic stem cells. Stem Cell Res. 2018, 33, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Maezawa, S.; Hasegawa, K.; Yukawa, M.; Sakashita, A.; Alavattam, K.G.; Andreassen, P.R.; Vidal, M.; Koseki, H.; Barski, A.; Namekawa, S.H. Polycomb directs timely activation of germline genes in spermatogenesis. Genes Dev. 2017, 31, 1693–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loubiere, V.; Papadopoulos, G.L.; Szabo, Q.; Martinez, A.M.; Cavalli, G. Widespread activation of developmental gene expression characterized by PRC1-dependent chromatin looping. Sci. Adv. 2020, 6, eaax4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Mazor, T.; Huang, H.; Huang, H.T.; Kathrein, K.L.; Woo, A.J.; Chouinard, C.R.; Labadorf, A.; Akie, T.E.; Moran, T.B.; et al. Direct recruitment of polycomb repressive complex 1 to chromatin by core binding transcription factors. Mol. Cell 2012, 45, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Creppe, C.; Palau, A.; Malinverni, R.; Valero, V.; Buschbeck, M. A Cbx8-containing polycomb complex facilitates the transition to gene activation during ES cell differentiation. PLoS Genet. 2014, 10, e1004851. [Google Scholar] [CrossRef]
- Yan, Y.; Zhao, W.; Huang, Y.; Tong, H.; Xia, Y.; Jiang, Q.; Qin, J. Loss of Polycomb Group Protein Pcgf1 Severely Compromises Proper Differentiation of Embryonic Stem Cells. Sci. Rep. 2017, 7, 46276. [Google Scholar] [CrossRef] [Green Version]
- Cohen, I.; Zhao, D.; Bar, C.; Valdes, V.J.; Dauber-Decker, K.L.; Nguyen, M.B.; Nakayama, M.; Rendl, M.; Bickmore, W.A.; Koseki, H.; et al. PRC1 Fine-tunes Gene Repression and Activation to Safeguard Skin Development and Stem Cell Specification. Cell Stem Cell 2018, 22, 726–739.e7. [Google Scholar] [CrossRef] [Green Version]
- Monderer-Rothkoff, G.; Tal, N.; Risman, M.; Shani, O.; Nissim-Rafinia, M.; Malki-Feldman, L.; Medvedeva, V.; Groszer, M.; Meshorer, E.; Shifman, S. AUTS2 isoforms control neuronal differentiation. Mol. Psychiatry 2021, 26, 666–681. [Google Scholar] [CrossRef]
- Russo, D.; Della Ragione, F.; Rizzo, R.; Sugiyama, E.; Scalabri, F.; Hori, K.; Capasso, S.; Sticco, L.; Fioriniello, S.; De Gregorio, R.; et al. Glycosphingolipid metabolic reprogramming drives neural differentiation. EMBO J. 2018, 37, e97674. [Google Scholar] [CrossRef]
- Hauri, S.; Comoglio, F.; Seimiya, M.; Gerstung, M.; Glatter, T.; Hansen, K.; Aebersold, R.; Paro, R.; Gstaiger, M.; Beisel, C. A High-Density Map for Navigating the Human Polycomb Complexome. Cell Rep. 2016, 17, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Hart, C.M.; Francis, N.J.; Vargas, M.L.; Sengupta, A.; Wild, B.; Miller, E.L.; O’Connor, M.B.; Kingston, R.E.; Simon, J.A. Histone Methyltransferase Activity of a Drosophila Polycomb Group Repressor Complex. Cell 2002, 111, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Czermin, B.; Melfi, R.; McCabe, D.; Seitz, V.; Imhof, A.; Pirrotta, V. Drosophila Enhancer of Zeste/ESC Complexes Have a Histone H3 Methyltransferase Activity that Marks Chromosomal Polycomb Sites. Cell 2002, 111, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Li, G.; Sarma, K.; Blais, A.; Zavadil, J.; Woodcock, C.L.; Dynlacht, B.D.; Reinberg, D. Ezh1 and Ezh2 maintain repressive chromatin through different mechanisms. Mol. Cell 2008, 32, 503–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alekseyenko, A.A.; Gorchakov, A.A.; Kharchenko, P.V.; Kuroda, M.I. Reciprocal interactions of human C10orf12 and C17orf96 with PRC2 revealed by BioTAP-XL cross-linking and affinity purification. Proc. Natl. Acad. Sci. USA 2014, 111, 2488–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liefke, R.; Karwacki-Neisius, V.; Shi, Y. EPOP Interacts with Elongin BC and USP7 to Modulate the Chromatin Landscape. Mol. Cell 2016, 64, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Grijzenhout, A.; Godwin, J.; Koseki, H.; Gdula, M.R.; Szumska, D.; McGouran, J.F.; Bhattacharya, S.; Kessler, B.M.; Brockdorff, N.; Cooper, S. Functional analysis of AEBP2, a PRC2 Polycomb protein, reveals a Trithorax phenotype in embryonic development and in ESCs. Development 2016, 143, 2716–2723. [Google Scholar] [CrossRef] [Green Version]
- Kirmizis, A.; Bartley, S.M.; Kuzmichev, A.; Margueron, R.; Reinberg, D.; Green, R.; Farnham, P.J. Silencing of human polycomb target genes is associated with methylation of histone H3 Lys 27. Genes Dev. 2004, 18, 1592–1605. [Google Scholar] [CrossRef] [Green Version]
- Zee, B.M.; Levin, R.S.; Xu, B.; LeRoy, G.; Wingreen, N.S.; Garcia, B.A. In vivo residue-specific histone methylation dynamics. J. Biol. Chem. 2010, 285, 3341–3350. [Google Scholar] [CrossRef] [Green Version]
- Sarma, K.; Margueron, R.; Ivanov, A.; Pirrotta, V.; Reinberg, D. Ezh2 requires PHF1 to efficiently catalyze H3 lysine 27 trimethylation in vivo. Mol. Cell. Biol. 2008, 28, 2718–2731. [Google Scholar] [CrossRef] [Green Version]
- Tie, F.; Banerjee, R.; Stratton, C.A.; Prasad-Sinha, J.; Stepanik, V.; Zlobin, A.; Diaz, M.O.; Scacheri, P.C.; Harte, P.J. CBP-mediated acetylation of histone H3 lysine 27 antagonizes Drosophila Polycomb silencing. Development 2009, 136, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- Cui, K.; Zang, C.; Roh, T.Y.; Schones, D.E.; Childs, R.W.; Peng, W.; Zhao, K. Chromatin signatures in multipotent human hematopoietic stem cells indicate the fate of bivalent genes during differentiation. Cell Stem Cell 2009, 4, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swigut, T.; Wysocka, J. H3K27 demethylases, at long last. Cell 2007, 131, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.; Zhang, Y. SUZ12 is required for both the histone methyltransferase activity and the silencing function of the EED-EZH2 complex. Mol. Cell 2004, 15, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Pasini, D.; Bracken, A.P.; Jensen, M.R.; Lazzerini Denchi, E.; Helin, K. Suz12 is essential for mouse development and for EZH2 histone methyltransferase activity. EMBO J. 2004, 23, 4061–4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, N.D.; Yee, D.; Chen, A.; Kalantry, S.; Chamberlain, S.J.; Otte, A.P.; Magnuson, T. The murine polycomb group protein Eed is required for global histone H3 lysine-27 methylation. Curr. Biol. 2005, 15, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Liu, X. Structural basis of histone H3K27 trimethylation by an active polycomb repressive complex 2. Science 2015, 350, aac4383. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Justin, N.; Ohno, K.; Sharpe, M.L.; Son, J.; Drury, W.J.; Voigt, P.; Martin, S.R.; Taylor, W.R.; De Marco, V.; et al. Role of the polycomb protein EED in the propagation of repressive histone marks. Nature 2009, 461, 762–767. [Google Scholar] [CrossRef] [Green Version]
- Oksuz, O.; Narendra, V.; Lee, C.H.; Descostes, N.; LeRoy, G.; Raviram, R.; Blumenberg, L.; Karch, K.; Rocha, P.P.; Garcia, B.A.; et al. Capturing the Onset of PRC2-Mediated Repressive Domain Formation. Mol. Cell 2018, 70, 1149–1162.e5. [Google Scholar] [CrossRef]
- Poepsel, S.; Kasinath, V.; Nogales, E. Cryo-EM structures of PRC2 simultaneously engaged with two functionally distinct nucleosomes. Nat. Struct. Mol. Biol. 2018, 25, 154–162. [Google Scholar] [CrossRef]
- Youmans, D.T.; Schmidt, J.C.; Cech, T.R. Live-cell imaging reveals the dynamics of PRC2 and recruitment to chromatin by SUZ12-associated subunits. Genes Dev. 2018, 32, 794–805. [Google Scholar] [CrossRef]
- Chen, S.; Jiao, L.; Liu, X.; Yang, X.; Liu, X. A Dimeric Structural Scaffold for PRC2-PCL Targeting to CpG Island Chromatin. Mol. Cell 2020, 77, 1265–1278.e7. [Google Scholar] [CrossRef] [PubMed]
- Hojfeldt, J.W.; Laugesen, A.; Willumsen, B.M.; Damhofer, H.; Hedehus, L.; Tvardovskiy, A.; Mohammad, F.; Jensen, O.N.; Helin, K. Accurate H3K27 methylation can be established de novo by SUZ12-directed PRC2. Nat. Struct. Mol. Biol. 2018, 25, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, L.; Bouveret, R.; Gruissem, W. MSI1-like proteins: An escort service for chromatin assembly and remodeling complexes. Trends Cell Biol. 2005, 15, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, M.; Wild, B.; Muller, J. Nucleosome binding and histone methyltransferase activity of Drosophila PRC2. EMBO Rep. 2005, 6, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Sanulli, S.; Justin, N.; Teissandier, A.; Ancelin, K.; Portoso, M.; Caron, M.; Michaud, A.; Lombard, B.; da Rocha, S.T.; Offer, J.; et al. Jarid2 Methylation via the PRC2 Complex Regulates H3K27me3 Deposition during Cell Differentiation. Mol. Cell 2015, 57, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Holder, M.; Grau, D.; Saldana-Meyer, R.; Yu, J.R.; Ganai, R.A.; Zhang, J.; Wang, M.; LeRoy, G.; Dobenecker, M.W.; et al. Distinct Stimulatory Mechanisms Regulate the Catalytic Activity of Polycomb Repressive Complex 2. Mol. Cell 2018, 70, 435–448.e435. [Google Scholar] [CrossRef] [Green Version]
- Conway, E.; Jerman, E.; Healy, E.; Ito, S.; Holoch, D.; Oliviero, G.; Deevy, O.; Glancy, E.; Fitzpatrick, D.J.; Mucha, M.; et al. A Family of Vertebrate-Specific Polycombs Encoded by the LCOR/LCORL Genes Balance PRC2 Subtype Activities. Mol. Cell 2018, 70, 408–421.e8. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; Gonzalez-Ramirez, M.; Alcaine-Colet, A.; Aranda, S.; Di Croce, L. The Bivalent Genome: Characterization, Structure, and Regulation. Trends Genet. 2020, 36, 118–131. [Google Scholar] [CrossRef]
- Kuzmichev, A.; Nishioka, K.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Histone methyltransferase activity associated with a human multiprotein complex containing the Enhancer of Zeste protein. Genes Dev. 2002, 16, 2893–2905. [Google Scholar] [CrossRef] [Green Version]
- Piunti, A.; Shilatifard, A. Epigenetic balance of gene expression by Polycomb and COMPASS families. Science 2016, 352, aad9780. [Google Scholar] [CrossRef] [Green Version]
- Beringer, M.; Pisano, P.; Di Carlo, V.; Blanco, E.; Chammas, P.; Vizan, P.; Gutierrez, A.; Aranda, S.; Payer, B.; Wierer, M.; et al. EPOP Functionally Links Elongin and Polycomb in Pluripotent Stem Cells. Mol. Cell 2016, 64, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brien, G.L.; Gambero, G.; O’Connell, D.J.; Jerman, E.; Turner, S.A.; Egan, C.M.; Dunne, E.J.; Jurgens, M.C.; Wynne, K.; Piao, L.; et al. Polycomb PHF19 binds H3K36me3 and recruits PRC2 and demethylase NO66 to embryonic stem cell genes during differentiation. Nat. Struct. Mol. Biol. 2012, 19, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Ballare, C.; Lange, M.; Lapinaite, A.; Martin, G.M.; Morey, L.; Pascual, G.; Liefke, R.; Simon, B.; Shi, Y.; Gozani, O.; et al. Phf19 links methylated Lys36 of histone H3 to regulation of Polycomb activity. Nat. Struct. Mol. Biol. 2012, 19, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Rothbart, S.B.; Lu, R.; Xu, B.; Chen, W.Y.; Tripathy, A.; Rockowitz, S.; Zheng, D.; Patel, D.J.; Allis, C.D.; et al. An H3K36 methylation-engaging Tudor motif of polycomb-like proteins mediates PRC2 complex targeting. Mol. Cell 2013, 49, 571–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatchalian, J.; Kingsley, M.C.; Moslet, S.D.; Rosas Ospina, R.D.; Kutateladze, T.G. An aromatic cage is required but not sufficient for binding of Tudor domains of the Polycomblike protein family to H3K36me3. Epigenetics 2015, 10, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musselman, C.A.; Avvakumov, N.; Watanabe, R.; Abraham, C.G.; Lalonde, M.E.; Hong, Z.; Allen, C.; Roy, S.; Nunez, J.K.; Nickoloff, J.; et al. Molecular basis for H3K36me3 recognition by the Tudor domain of PHF1. Nat. Struct. Mol. Biol. 2012, 19, 1266–1272. [Google Scholar] [CrossRef]
- Casanova, M.; Preissner, T.; Cerase, A.; Poot, R.; Yamada, D.; Li, X.; Appanah, R.; Bezstarosti, K.; Demmers, J.; Koseki, H.; et al. Polycomblike 2 facilitates the recruitment of PRC2 Polycomb group complexes to the inactive X chromosome and to target loci in embryonic stem cells. Development 2011, 138, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Perino, M.; van Mierlo, G.; Karemaker, I.D.; van Genesen, S.; Vermeulen, M.; Marks, H.; van Heeringen, S.J.; Veenstra, G.J.C. MTF2 recruits Polycomb Repressive Complex 2 by helical-shape-selective DNA binding. Nat. Genet. 2018, 50, 1002–1010. [Google Scholar] [CrossRef]
- Li, H.; Liefke, R.; Jiang, J.; Kurland, J.V.; Tian, W.; Deng, P.; Zhang, W.; He, Q.; Patel, D.J.; Bulyk, M.L.; et al. Polycomb-like proteins link the PRC2 complex to CpG islands. Nature 2017, 549, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Hunkapiller, J.; Shen, Y.; Diaz, A.; Cagney, G.; McCleary, D.; Ramalho-Santos, M.; Krogan, N.; Ren, B.; Song, J.S.; Reiter, J.F. Polycomb-like 3 promotes polycomb repressive complex 2 binding to CpG islands and embryonic stem cell self-renewal. PLoS Genet. 2012, 8, e1002576. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Bachmann, A.L.; Tauscher, K.; Benda, C.; Fierz, B.; Muller, J. DNA binding by PHF1 prolongs PRC2 residence time on chromatin and thereby promotes H3K27 methylation. Nat. Struct. Mol. Biol. 2017, 24, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, G.; Brien, G.L.; Waston, A.; Streubel, G.; Jerman, E.; Andrews, D.; Doyle, B.; Munawar, N.; Wynne, K.; Crean, J.; et al. Dynamic Protein Interactions of the Polycomb Repressive Complex 2 during Differentiation of Pluripotent Cells. Mol. Cell Proteom. 2016, 15, 3450–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloet, S.L.; Makowski, M.M.; Baymaz, H.I.; van Voorthuijsen, L.; Karemaker, I.D.; Santanach, A.; Jansen, P.; Di Croce, L.; Vermeulen, M. The dynamic interactome and genomic targets of Polycomb complexes during stem-cell differentiation. Nat. Struct. Mol. Biol. 2016, 23, 682–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.; Shen, S.S.; Margueron, R.; Reinberg, D. Nucleosome-binding activities within JARID2 and EZH1 regulate the function of PRC2 on chromatin. Genes Dev. 2013, 27, 2663–2677. [Google Scholar] [CrossRef] [Green Version]
- Justin, N.; Zhang, Y.; Tarricone, C.; Martin, S.R.; Chen, S.; Underwood, E.; De Marco, V.; Haire, L.F.; Walker, P.A.; Reinberg, D.; et al. Structural basis of oncogenic histone H3K27M inhibition of human polycomb repressive complex 2. Nat. Commun. 2016, 7, 11316. [Google Scholar] [CrossRef] [Green Version]
- Kasinath, V.; Faini, M.; Poepsel, S.; Reif, D.; Feng, X.A.; Stjepanovic, G.; Aebersold, R.; Nogales, E. Structures of human PRC2 with its cofactors AEBP2 and JARID2. Science 2018, 359, 940–944. [Google Scholar] [CrossRef] [Green Version]
- Pajtler, K.W.; Wen, J.; Sill, M.; Lin, T.; Orisme, W.; Tang, B.; Hubner, J.M.; Ramaswamy, V.; Jia, S.; Dalton, J.D.; et al. Molecular heterogeneity and CXorf67 alterations in posterior fossa group A (PFA) ependymomas. Acta Neuropathol. 2018, 136, 211–226. [Google Scholar] [CrossRef]
- Piunti, A.; Smith, E.R.; Morgan, M.A.J.; Ugarenko, M.; Khaltyan, N.; Helmin, K.A.; Ryan, C.A.; Murray, D.C.; Rickels, R.A.; Yilmaz, B.D.; et al. CATACOMB: An endogenous inducible gene that antagonizes H3K27 methylation activity of Polycomb repressive complex 2 via an H3K27M-like mechanism. Sci. Adv. 2019, 5, eaax2887. [Google Scholar] [CrossRef] [Green Version]
- Ragazzini, R.; Perez-Palacios, R.; Baymaz, I.H.; Diop, S.; Ancelin, K.; Zielinski, D.; Michaud, A.; Givelet, M.; Borsos, M.; Aflaki, S.; et al. EZHIP constrains Polycomb Repressive Complex 2 activity in germ cells. Nat. Commun. 2019, 10, 3858. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.U.; Do, T.J.; Lund, P.J.; Rashoff, A.Q.; Diehl, K.L.; Cieslik, M.; Bajic, A.; Juretic, N.; Deshmukh, S.; Venneti, S.; et al. PFA ependymoma-associated protein EZHIP inhibits PRC2 activity through a H3 K27M-like mechanism. Nat. Commun. 2019, 10, 2146. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Beato, M.; Sanchez, E.; Gonzalez-Carrero, J.; Morente, M.; Diez, A.; Sanchez-Verde, L.; Martin, M.C.; Cigudosa, J.C.; Vidal, M.; Piris, M.A. Variability in the expression of polycomb proteins in different normal and tumoral tissues. A pilot study using tissue microarrays. Mod. Pathol. 2006, 19, 684–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sun, Y.; Shang, C.; Chen, L.; Chen, H.; Wang, D.; Zeng, X. Distinct Ring1b complexes defined by DEAD-box helicases and EMT transcription factors synergistically enhance E-cadherin silencing in breast cancer. Cell Death Dis. 2021, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.L.; Beckedorff, F.; Zhang, Y.; Garcia-Huidobro, J.; Jiang, H.; Colaprico, A.; Bilbao, D.; Figueroa, M.E.; LaCava, J.; Shiekhattar, R.; et al. Polycomb complexes associate with enhancers and promote oncogenic transcriptional programs in cancer through multiple mechanisms. Nat. Commun. 2018, 9, 3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chan, H.L.; Garcia-Martinez, L.; Karl, D.L.; Weich, N.; Slingerland, J.M.; Verdun, R.E.; Morey, L. Estrogen induces dynamic ERα and RING1B recruitment to control gene and enhancer activities in luminal breast cancer. Sci. Adv. 2020, 6, eaaz7249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, T.; Yuan, F.; Garcia-Martinez, L.; Lee, K.D.; Stransky, S.; Sidoli, S.; Verdun, R.E.; Zhang, Y.; Wang, Z.; et al. The Polycomb protein RING1B enables estrogen-mediated gene expression by promoting enhancer-promoter interaction and R-loop formation. Nucleic Acids Res. 2021, 49, 9768–9782. [Google Scholar] [CrossRef] [PubMed]
- Su, W.J.; Fang, J.S.; Cheng, F.; Liu, C.; Zhou, F.; Zhang, J. RNF2/Ring1b negatively regulates p53 expression in selective cancer cell types to promote tumor development. Proc. Natl. Acad. Sci. USA 2013, 110, 1720–1725. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Li, P.; Shao, X.; Yang, Y.; Liu, X.; Feng, M.; Yu, Q.; Hu, R.; Wang, Z. The E3 Ligase RING1 Targets p53 for Degradation and Promotes Cancer Cell Proliferation and Survival. Cancer Res. 2018, 78, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Lee, J.Y.; Shin, D.H.; Jang, K.S.; Kim, H.J.; Kong, G. Loss of Mel-18 induces tumor angiogenesis through enhancing the activity and expression of HIF-1alpha mediated by the PTEN/PI3K/Akt pathway. Oncogene 2011, 30, 4578–4589. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Park, M.K.; Park, J.H.; Lee, H.J.; Shin, D.H.; Kang, Y.; Lee, C.H.; Kong, G. Loss of the polycomb protein Mel-18 enhances the epithelial-mesenchymal transition by ZEB1 and ZEB2 expression through the downregulation of miR-205 in breast cancer. Oncogene 2014, 33, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Sheng, Y.P.; Li, Q.; Qin, W.; Lu, Y.W.; Cheng, Y.F.; Liu, B.Y.; Zhang, F.C.; Li, J.; Dimri, G.P.; et al. BMI1 and Mel-18 oppositely regulate carcinogenesis and progression of gastric cancer. Mol. Cancer 2010, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.J.; Kieboom, K.; Marino, S.; DePinho, R.A.; van Lohuizen, M. The oncogene and Polycomb-group gene bmi-1 regulates cell proliferation and senescence through the ink4a locus. Nature 1999, 397, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Althobiti, M.; Muftah, A.A.; Aleskandarany, M.A.; Joseph, C.; Toss, M.S.; Green, A.; Rakha, E. The prognostic significance of BMI1 expression in invasive breast cancer is dependent on its molecular subtypes. Breast Cancer Res. Treat. 2020, 182, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Vonlanthen, S.; Heighway, J.; Altermatt, H.J.; Gugger, M.; Kappeler, A.; Borner, M.M.; Van Lohuizen, M.; Betticher, D.C. The bmi-1 oncoprotein is differentially expressed in non-small cell lung cancer and correlates with INK4A-ARF locus expression. Br. J. Cancer 2001, 84, 1372–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, M.; Bharali, D.J.; Sudha, T.; Khedr, M.; Guest, I.; Sell, S.; Glinsky, G.V.; Mousa, S.A. Downregulation of Bmi1 in breast cancer stem cells suppresses tumor growth and proliferation. Oncotarget 2017, 8, 38731–38742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, T.; Seki, A.; Aoki, R.; Ichikawa, H.; Negishi, M.; Miyagi, S.; Oguro, H.; Saraya, A.; Kamiya, A.; Nakauchi, H.; et al. Bmi1 promotes hepatic stem cell expansion and tumorigenicity in both Ink4a/Arf-dependent and -independent manners in mice. Hepatology 2010, 52, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.H.; Feng, Y.; Zhang, R.; Xu, L.H.; Li, M.Z.; Kung, H.F.; Song, L.B.; Zeng, M.S. Bmi-1 promotes invasion and metastasis, and its elevated expression is correlated with an advanced stage of breast cancer. Mol. Cancer 2011, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Song, L.B.; Li, J.; Liao, W.T.; Feng, Y.; Yu, C.P.; Hu, L.J.; Kong, Q.L.; Xu, L.H.; Zhang, X.; Liu, W.L.; et al. The polycomb group protein Bmi-1 represses the tumor suppressor PTEN and induces epithelial-mesenchymal transition in human nasopharyngeal epithelial cells. J. Clin. Investig. 2009, 119, 3626–3636. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Zhao, D.; Yan, L.; Jiang, W.; Kim, J.S.; Gu, B.; Liu, Q.; Wang, R.; Xia, B.; Zhao, J.C.; et al. BMI1 regulates androgen receptor in prostate cancer independently of the polycomb repressive complex 1. Nat. Commun. 2018, 9, 500. [Google Scholar] [CrossRef]
- Zhu, S.; Zhao, D.; Li, C.; Li, Q.; Jiang, W.; Liu, Q.; Wang, R.; Fazli, L.; Li, Y.; Zhang, L.; et al. BMI1 is directly regulated by androgen receptor to promote castration-resistance in prostate cancer. Oncogene 2020, 39, 17–29. [Google Scholar] [CrossRef]
- Deshpande, A.M.; Akunowicz, J.D.; Reveles, X.T.; Patel, B.B.; Saria, E.A.; Gorlick, R.G.; Naylor, S.L.; Leach, R.J.; Hansen, M.F. PHC3, a component of the hPRC-H complex, associates with E2F6 during G0 and is lost in osteosarcoma tumors. Oncogene 2007, 26, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Iwata, S.; Takenobu, H.; Kageyama, H.; Koseki, H.; Ishii, T.; Nakazawa, A.; Tatezaki, S.; Nakagawara, A.; Kamijo, T. Polycomb group molecule PHC3 regulates polycomb complex composition and prognosis of osteosarcoma. Cancer Sci. 2010, 101, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Li, B.; Yang, L.; Guan, Q. CBX2 depletion inhibits the proliferation, invasion and migration of gastric cancer cells by inactivating the YAP/beta-catenin pathway. Mol. Med. Rep. 2021, 23, 137. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Lv, P.; Su, J.; Miao, K.; Xu, H.; Li, M. Overexpression of CBX2 in breast cancer promotes tumor progression through the PI3K/AKT signaling pathway. Am. J. Transl. Res. 2019, 11, 1668–1682. [Google Scholar] [PubMed]
- Jiao, H.K.; Xu, Y.; Li, J.; Wang, W.; Mei, Z.; Long, X.D.; Chen, G.Q. Prognostic significance of Cbx4 expression and its beneficial effect for transarterial chemoembolization in hepatocellular carcinoma. Cell Death Dis. 2015, 6, e1689. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xu, Y.; Long, X.-D.; Wang, W.; Jiao, H.-K.; Mei, Z.; Yin, Q.-Q.; Ma, L.-N.; Zhou, A.-W.; Wang, L.-S.; et al. Cbx4 governs HIF-1alpha to potentiate angiogenesis of hepatocellular carcinoma by its SUMO E3 ligase activity. Cancer Cell 2014, 25, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Niu, G.; Pan, Y.H.; Pan, W.; Zhang, M.F.; Zhang, C.Z.; Shen, H. CBX4 transcriptionally suppresses KLF6 via interaction with HDAC1 to exert oncogenic activities in clear cell renal cell carcinoma. EBioMedicine 2020, 53, 102692. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Wu, Y.; Zhang, R.; Zhang, M.; Liao, D.; Wang, G.; Qin, G.; Xu, R.H.; Kang, T. CBX4 Suppresses Metastasis via Recruitment of HDAC3 to the Runx2 Promoter in Colorectal Carcinoma. Cancer Res. 2016, 76, 7277–7289. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; He, H.; Jiang, Q.; Wang, Y.; Jia, S. CBX6 Promotes HCC Metastasis Via Transcription Factors Snail/Zeb1-Mediated EMT Mechanism. OncoTargets Ther. 2020, 13, 12489–12500. [Google Scholar] [CrossRef]
- Scott, C.L.; Gil, J.; Hernando, E.; Teruya-Feldstein, J.; Narita, M.; Martínez, D.; Visakorpi, T.; Mu, D.; Cordon-Cardo, C.; Peters, G.; et al. Role of the chromobox protein CBX7 in lymphomagenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 5389–5394. [Google Scholar] [CrossRef] [Green Version]
- Bernard, D.; Martinez-Leal, J.F.; Rizzo, S.; Martinez, D.; Hudson, D.; Visakorpi, T.; Peters, G.; Carnero, A.; Beach, D.; Gil, J. CBX7 controls the growth of normal and tumor-derived prostate cells by repressing the Ink4a/Arf locus. Oncogene 2005, 24, 5543–5551. [Google Scholar] [CrossRef] [Green Version]
- Federico, A.; Pallante, P.; Bianco, M.; Ferraro, A.; Esposito, F.; Monti, M.; Cozzolino, M.; Keller, S.; Fedele, M.; Leone, V.; et al. Chromobox protein homologue 7 protein, with decreased expression in human carcinomas, positively regulates E-cadherin expression by interacting with the histone deacetylase 2 protein. Cancer Res. 2009, 69, 7079–7087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Sepe, R.; Cozzolino, F.; Piccolo, C.; Iannone, C.; Iacobucci, I.; Pucci, P.; Monti, M.; Fusco, A. The complex CBX7-PRMT1 has a critical role in regulating E-cadherin gene expression and cell migration. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Forzati, F.; Federico, A.; Pallante, P.; Fedele, M.; Fusco, A. Tumor suppressor activity of CBX7 in lung carcinogenesis. Cell Cycle 2012, 11, 1888–1891. [Google Scholar] [CrossRef] [PubMed]
- Forzati, F.; Federico, A.; Pallante, P.; Abbate, A.; Esposito, F.; Malapelle, U.; Sepe, R.; Palma, G.; Troncone, G.; Scarfò, M.; et al. CBX7 is a tumor suppressor in mice and humans. J. Clin. Investig. 2012, 122, 612–623. [Google Scholar] [CrossRef]
- Huang, Z.; Yan, Y.; Zhu, Z.; Liu, J.; He, X.; Dalangood, S.; Li, M.; Tan, M.; Cai, J.; Tang, P.; et al. CBX7 suppresses urinary bladder cancer progression via modulating AKR1B10-ERK signaling. Cell Death Dis. 2021, 12, 537. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.; Jian, C.; Li, W.; Ye, K.; Ren, J.; Zhu, L.; Wang, Y.; Jin, X.; Lu, Y. The RNF26/CBX7 axis modulates the TNF pathway to promote cell proliferation and regulate sensitivity to TKIs in ccRCC. Int. J. Biol. Sci. 2022, 18, 2132–2145. [Google Scholar] [CrossRef]
- Lee, S.H.; Um, S.J.; Kim, E.J. CBX8 antagonizes the effect of Sirtinol on premature senescence through the AKT-RB-E2F1 pathway in K562 leukemia cells. Biochem. Biophys. Res. Commun. 2016, 469, 884–890. [Google Scholar] [CrossRef]
- Lee, S.H.; Um, S.J.; Kim, E.J. CBX8 suppresses Sirtinol-induced premature senescence in human breast cancer cells via cooperation with SIRT1. Cancer Lett. 2013, 335, 397–403. [Google Scholar] [CrossRef]
- Tang, B.; Tian, Y.; Liao, Y.; Li, Z.; Yu, S.; Su, H.; Zhong, F.; Yuan, G.; Wang, Y.; Yu, H.; et al. CBX8 exhibits oncogenic properties and serves as a prognostic factor in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Z.; Chen, S.L.; Wang, C.H.; He, Y.F.; Yang, X.; Xie, D.; Yun, J.P. CBX8 Exhibits Oncogenic Activity via AKT/beta-Catenin Activation in Hepatocellular Carcinoma. Cancer Res. 2018, 78, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.Y.; Sun, Z.; Mullokandov, G.; Bosch, A.; Qadeer, Z.A.; Cihan, E.; Rapp, Z.; Parsons, R.; Aguirre-Ghiso, J.A.; Farias, E.F.; et al. Cbx8 Acts Non-canonically with Wdr5 to Promote Mammary Tumorigenesis. Cell Rep. 2016, 16, 472–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kang, M.; Zhang, B.; Meng, F.; Song, J.; Kaneko, H.; Shimamoto, F.; Tang, B. m(6)A modification-mediated CBX8 induction regulates stemness and chemosensitivity of colon cancer via upregulation of LGR5. Mol. Cancer 2019, 18, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Yue, J.; Wu, X.; Wang, X.; Wen, J.; Lu, L.; Peng, X.; Qiang, B.; Yuan, J. NSPc1 is a cell growth regulator that acts as a transcriptional repressor of p21Waf1/Cip1 via the RARE element. Nucleic Acids Res. 2006, 34, 6158–6169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, G.; Zhou, W.; Du, J.; Zhou, J.; Wu, D.; Zhao, M.; Yang, L.; Hao, A. PCGF1 promotes epigenetic activation of stemness markers and colorectal cancer stem cell enrichment. Cell Death Dis. 2021, 12, 633. [Google Scholar] [CrossRef]
- Hu, P.S.; Xia, Q.S.; Wu, F.; Li, D.K.; Qi, Y.J.; Hu, Y.; Wei, Z.Z.; Li, S.S.; Tian, N.Y.; Wei, Q.F.; et al. NSPc1 promotes cancer stem cell self-renewal by repressing the synthesis of all-trans retinoic acid via targeting RDH16 in malignant glioma. Oncogene 2017, 36, 4706–4718. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, Y.; Jiang, X.; Zhang, Y.; Wang, H.; Ren, H.; Xu, Y.; Jiang, J.; Wang, Q.; Su, H.; et al. PCGF3 promotes the proliferation and migration of non-small cell lung cancer cells via the PI3K/AKT signaling pathway. Exp. Cell Res. 2021, 400, 112496. [Google Scholar] [CrossRef]
- Lee, J.H.; Zhao, X.M.; Yoon, I.; Lee, J.Y.; Kwon, N.H.; Wang, Y.Y.; Lee, K.M.; Lee, M.J.; Kim, J.; Moon, H.G.; et al. Integrative analysis of mutational and transcriptional profiles reveals driver mutations of metastatic breast cancers. Cell Discov. 2016, 2, 16025. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Li, Q.; Wang, A.; Jiao, J. YY1 regulates melanoma tumorigenesis through a miR-9 ~ RYBP axis. J. Exp. Clin. Cancer Res. 2015, 34, 66. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.; Zhang, X.; Cong, X.; Huang, B.; Chen, H.; Chen, D. Tumor suppressor RYBP harbors three nuclear localization signals and its cytoplasm-located mutant exerts more potent anti-cancer activities than corresponding wild type. Cell. Signal. 2017, 29, 127–137. [Google Scholar] [CrossRef]
- Zhan, S.; Wang, T.; Ge, W.; Li, J. Multiple roles of Ring 1 and YY1 binding protein in physiology and disease. J. Cell. Mol. Med. 2018, 22, 2046–2054. [Google Scholar] [CrossRef]
- Zhou, H.; Li, J.; Zhang, Z.; Ye, R.; Shao, N.; Cheang, T.; Wang, S. RING1 and YY1 binding protein suppresses breast cancer growth and metastasis. Int. J. Oncol. 2016, 49, 2442–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, A.H.; Tan, J.; Zhang, J.H.; Xu, F.J.; Li, F.Y.; Cao, C.Y. Overexpression of RYBP inhibits proliferation, invasion, and chemoresistance to cisplatin in anaplastic thyroid cancer cells via the EGFR pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22241. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.-F.; Zhao, L.-X.; Guo, S.-P.; Wei, S.; Zhai, J.-F.; Zhou, Q.-H. miR-34b inhibits the migration/invasion and promotes apoptosis of non-small-cell lung cancer cells by YAF2. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2038–2046. [Google Scholar]
- Zhang, S.; Zhang, X.; Guan, X.; Ma, X.; Chen, H.; Huang, B.; Chen, D. YAF2 exerts anti-apoptotic effect in human tumor cells in a FANK1- and phosphorylation-dependent manner. Biochem. Biophys. Res. Commun. 2021, 554, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kottakis, F.; Foltopoulou, P.; Sanidas, I.; Keller, P.; Wronski, A.; Dake, B.T.; Ezell, S.A.; Shen, Z.; Naber, S.P.; Hinds, P.W.; et al. NDY1/KDM2B functions as a master regulator of polycomb complexes and controls self-renewal of breast cancer stem cells. Cancer Res. 2014, 74, 3935–3946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Nguyen, A.T.; Zhang, Y. KDM2b/JHDM1b, an H3K36me2-specific demethylase, is required for initiation and maintenance of acute myeloid leukemia. Blood 2011, 117, 3869–3880. [Google Scholar] [CrossRef] [Green Version]
- Isshiki, Y.; Nakajima-Takagi, Y.; Oshima, M.; Aoyama, K.; Rizk, M.; Kurosawa, S.; Saraya, A.; Kondo, T.; Sakaida, E.; Nakaseko, C.; et al. KDM2B in polycomb repressive complex 1.1 functions as a tumor suppressor in the initiation of T-cell leukemogenesis. Blood Adv. 2019, 3, 2537–2549. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, A.; Fiore, M.; Melchionda, F.; Indio, V.; Bertuccio, S.N.; Pession, A. BCOR involvement in cancer. Epigenomics 2019, 11, 835–855. [Google Scholar] [CrossRef] [Green Version]
- Panagopoulos, I.; Thorsen, J.; Gorunova, L.; Haugom, L.; Bjerkehagen, B.; Davidson, B.; Heim, S.; Micci, F. Fusion of the ZC3H7B and BCOR genes in endometrial stromal sarcomas carrying an X;22-translocation. Genes Chromosomes Cancer 2013, 52, 610–618. [Google Scholar]
- Tanaka, T.; Nakajima-Takagi, Y.; Aoyama, K.; Tara, S.; Oshima, M.; Saraya, A.; Koide, S.; Si, S.; Manabe, I.; Sanada, M.; et al. Internal deletion of BCOR reveals a tumor suppressor function for BCOR in T lymphocyte malignancies. J. Exp. Med. 2017, 214, 2901–2913. [Google Scholar] [CrossRef] [Green Version]
- Denk, D.; Nebral, K.; Bradtke, J.; Pass, G.; Möricke, A.; Attarbaschi, A.; Strehl, S. PAX5-AUTS2: A recurrent fusion gene in childhood B-cell precursor acute lymphoblastic leukemia. Leuk. Res. 2012, 36, e178–e181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.H.; Roberts, C.W. Targeting EZH2 in cancer. Nat. Med. 2016, 22, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Yang, J.Y.; Xia, W.; Chen, C.T.; Xie, X.; Chao, C.H.; Woodward, W.A.; Hsu, J.M.; Hortobagyi, G.N.; Hung, M.C. EZH2 promotes expansion of breast tumor initiating cells through activation of RAF1-beta-catenin signaling. Cancer Cell 2011, 19, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takawa, M.; Masuda, K.; Kunizaki, M.; Daigo, Y.; Takagi, K.; Iwai, Y.; Cho, H.-S.; Toyokawa, G.; Yamane, Y.; Maejima, K.; et al. Validation of the histone methyltransferase EZH2 as a therapeutic target for various types of human cancer and as a prognostic marker. Cancer Sci. 2011, 102, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.D.; Mongan, N.P.; Tickoo, S.K.; Boorjian, S.A.; Scherr, D.S.; Gudas, L.J. Increased expression of the polycomb group gene, EZH2, in transition-al cell carcinoma of the bladder. Clin. Cancer Res. 2005, 11, 8570–8576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Qi, J.; Reyes, J.M.; Li, L.; Rao, P.K.; Li, F.; Lin, C.Y.; Perry, J.A.; Lawlor, M.A.; Federation, A.; et al. Oncogenic Deregulation of EZH2 as an Opportunity for Targeted Therapy in Lung Cancer. Cancer Discov. 2016, 6, 1006–1021. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.M.; Wang, C.G.; Zhu, M.; Xing, R.; Cui, J.T.; Li, W.M.; Yu, D.D.; Wang, S.B.; Zhu, W.; Ye, Y.J.; et al. STAT3 signaling drives EZH2 transcriptional activation and mediates poor prognosis in gastric cancer. Mol. Cancer 2016, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- McCabe, M.T.; Ott, H.M.; Ganji, G.; Korenchuk, S.; Thompson, C.; Van Aller, G.S.; Liu, Y.; Graves, A.P.; Della Pietra, A., 3rd; Diaz, E.; et al. EZH2 inhibition as a therapeutic strategy for lymphoma with EZH2-activating mutations. Nature 2012, 492, 108–112. [Google Scholar] [CrossRef]
- Majer, C.R.; Jin, L.; Scott, M.P.; Knutson, S.K.; Kuntz, K.W.; Keilhack, H.; Smith, J.J.; Moyer, M.P.; Richon, V.M.; Copeland, R.A.; et al. A687V EZH2 is a gain-of-function mutation found in lymphoma patients. FEBS Lett. 2012, 586, 3448–3451. [Google Scholar] [CrossRef] [Green Version]
- Béguelin, W.; Popovic, R.; Teater, M.; Jiang, Y.; Bunting, K.L.; Rosen, M.; Shen, H.; Yang, S.N.; Wang, L.; Ezponda, T.; et al. EZH2 is required for germinal center formation and somatic EZH2 muta-tions promote lymphoid transformation. Cancer Cell 2013, 23, 677–692. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki-Kashio, M.; Aoyama, K.; Sashida, G.; Oshima, M.; Tomioka, T.; Muto, T.; Wang, C.; Iwama, A. Ezh2 loss in hematopoietic stem cells predisposes mice to develop heterogeneous malignancies in an Ezh1-dependent manner. Blood 2015, 126, 1172–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntziachristos, P.; Tsirigos, A.; Van Vlierberghe, P.; Nedjic, J.; Trimarchi, T.; Flaherty, M.S.; Ferres-Marco, D.; da Ros, V.; Tang, Z.; Siegle, J.; et al. Genetic inactivation of the polycomb repressive complex 2 in T cell acute lymphoblastic leukemia. Nat. Med. 2012, 18, 298–301. [Google Scholar] [CrossRef]
- Bardelli, V.; Arniani, S.; Pierini, V.; Di Giacomo, D.; Pierini, T.; Gorello, P.; Mecucci, C.; La Starza, R. T-Cell Acute Lymphoblastic Leukemia: Biomarkers and Their Clinical Use-fulness. Genes 2021, 12, 1118. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Chagraoui, J.; Krosl, J.; Gendron, P.; Wilhelm, B.; Lemieux, S.; Boucher, G.; Chagnon, P.; Drouin, S.; Lambert, R.; et al. A key role for EZH2 and associated genes in mouse and human adult T-cell acute leukemia. Genes Dev. 2012, 26, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassef, M.; Rodilla, V.; Teissandier, A.; Zeitouni, B.; Gruel, N.; Sadacca, B.; Irondelle, M.; Charruel, M.; Ducos, B.; Michaud, A.; et al. Impaired PRC2 activity promotes transcriptional instability and favors breast tumorigenesis. Genes Dev. 2015, 29, 2547–2562. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Wahab, O.; Adli, M.; LaFave, L.M.; Gao, J.; Hricik, T.; Shih, A.H.; Pandey, S.; Patel, J.P.; Chung, Y.R.; Koche, R.; et al. ASXL1 mutations promote myeloid transformation through loss of PRC2-mediated gene repression. Cancer Cell 2012, 22, 180–193. [Google Scholar] [CrossRef] [Green Version]
- Lane, A.A.; Chapuy, B.; Lin, C.Y.; Tivey, T.; Li, H.; Townsend, E.C.; Van Bodegom, D.; Day, T.A.; Wu, S.C.; Liu, H.; et al. Triplication of a 21q22 region contributes to B cell transformation through HMGN1 overexpression and loss of histone H3 Lys27 trimethylation. Nat. Genet. 2014, 46, 618–623. [Google Scholar] [CrossRef] [Green Version]
- Wassef, M.; Luscan, A.; Aflaki, S.; Zielinski, D.; Jansen, P.; Baymaz, H.I.; Battistella, A.; Kersouani, C.; Servant, N.; Wallace, M.R.; et al. EZH1/2 function mostly within canonical PRC2 and exhibit proliferation-dependent redundancy that shapes mutational signatures in cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 6075–6080. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, N.; Tiwari, V.K.; Waldmeier, L.; Balwierz, P.J.; Arnold, P.; Pachkov, M.; Meyer-Schaller, N.; Schübeler, D.; van Nimwegen, E.; Christofori, G. Sox4 is a master regulator of epithelial-mesenchymal transition by con-trolling Ezh2 expression and epigenetic reprogramming. Cancer Cell 2013, 23, 768–783. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Simons, D.L.; Segall, I.; Carcamo-Cavazos, V.; Schwartz, E.J.; Yan, N.; Zuckerman, N.S.; Dirbas, F.M.; Johnson, D.L.; Holmes, S.P.; et al. PRC2/EED-EZH2 complex is up-regulated in breast cancer lymph node metastasis compared to primary tumor and correlates with tumor proliferation in situ. PLoS ONE 2012, 7, e51239. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L.; Gao, X.; Jiang, Y.; Zhang, G.; Sun, Z.C.; Cui, B.B.; Yang, Y.M. Expression and clinicopathological significance of EED, SUZ12 and EZH2 mRNA in colorectal cancer. J. Cancer Res. Clin. Oncol. 2015, 141, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Veneti, Z.; Gkouskou, K.K.; Eliopoulos, A.G. Polycomb Repressor Complex 2 in Genomic Instability and Cancer. Int. J. Mol. Sci. 2017, 18, 1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Selvaraju, S.; Curtin, M.L.; Jakob, C.G.; Zhu, H.; Comess, K.M.; Shaw, B.; The, J.; Lima-Fernandes, E.; Szewczyk, M.M.; et al. The EED protein-protein interaction inhibitor A-395 inactivates the PRC2 complex. Nat. Chem. Biol. 2017, 13, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Nakata, Y.; Nagamachi, A.; Yamasaki, N.; Kanai, A.; Sera, Y.; Sasaki, M.; Matsui, H.; Honda, Z.-I.; Oda, H.; et al. Propagation of trimethylated H3K27 regulated by polycomb protein EED is required for embryogenesis, hematopoietic maintenance, and tumor suppression. Proc. Natl. Acad. Sci. USA 2016, 113, 10370–10375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.L.; Barnash, K.D.; Abramyan, T.M.; Li, F.; The, J.; Engelberg, I.A.; Vedadi, M.; Brown, P.J.; Kireev, D.B.; Arrowsmith, C.H.; et al. Discovery of selective activators of PRC2 mutant EED-I363M. Sci. Rep. 2019, 9, 6524. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Hu, H.; Zhang, W.; Li, Z.; Diao, P.; Wang, D.; Zhang, W.; Wang, Y.; Yang, J.; Cheng, J. SUZ12 is a novel putative oncogene promoting tumorigenesis in head and neck squamous cell carcinoma. J. Cell. Mol. Med. 2018, 22, 3582–3594. [Google Scholar] [CrossRef]
- Li, H.; Cai, Q.; Wu, H.; Vathipadiekal, V.; Dobbin, Z.C.; Li, T.; Hua, X.; Landen, C.N.; Birrer, M.J.; Sánchez-Beato, M.; et al. SUZ12 promotes human epithelial ovarian cancer by suppressing apoptosis via silencing HRK. Mol. Cancer Res. 2012, 10, 1462–1472. [Google Scholar] [CrossRef] [Green Version]
- Iliopoulos, D.; Lindahl-Allen, M.; Polytarchou, C.; Hirsch, H.A.; Tsichlis, P.N.; Struhl, K. Loss of miR-200 inhibition of Suz12 leads to poly-comb-mediated repression required for the formation and maintenance of cancer stem cells. Mol. Cell 2010, 39, 761–772. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Wang, K.; Jiang, X.; Gu, C.; Yu, G.; Zhong, Y.; Liu, S.; Nie, Y.; Zhou, Y.; Yang, H. The Down-Regulation of SUZ12 Accelerates the Migration and Invasion of Liver Cancer Cells via Activating ERK1/2 Pathway. J. Cancer 2019, 10, 1375–1384. [Google Scholar] [CrossRef]
- De Raedt, T.; Beert, E.; Pasmant, E.; Luscan, A.; Brems, H.; Ortonne, N.; Helin, K.; Hornick, J.L.; Mautner, V.; Kehrer-Sawatzki, H.; et al. PRC2 loss amplifies Ras-driven transcription and confers sensitivity to BRD4-based therapies. Nature 2014, 514, 247–251. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Jones, S.; Sausen, M.; McMahon, K.; Sharma, R.; Wang, Q.; Belzberg, A.J.; Chaichana, K.; Gallia, G.L.; et al. Somatic mutations of SUZ12 in malignant peripheral nerve sheath tumors. Nat. Genet. 2014, 46, 1170–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ding, L.; Holmfeldt, L.; Wu, G.; Heatley, S.L.; Payne-Turner, D.; Easton, J.; Chen, X.; Wang, J.; Rusch, M.; et al. The genetic basis of early T-cell precursor acute lymphoblastic leukaemia. Nature 2012, 481, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, H.H.; Tseng, Y.F.; Hsu, Y.C.; Lan, S.H.; Wu, S.Y.; Raghavaraju, G.; Cheng, D.E.; Lee, Y.R.; Chang, T.Y.; Chow, N.H.; et al. Ras induces experimental lung metastasis through up-regulation of RbAp46 to suppress RECK promoter activity. BMC Cancer 2015, 15, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; He, C.; Gao, P.; Wang, S.; Lv, R.; Zhou, H.; Zhou, Q.; Zhang, K.; Sun, J.; Fan, C.; et al. HNF1B-mediated repression of SLUG is suppressed by EZH2 in aggressive prostate cancer. Oncogene 2020, 39, 1335–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebre-Medhin, S.; Nord, K.H.; Moller, E.; Mandahl, N.; Magnusson, L.; Nilsson, J.; Jo, V.Y.; Vult von Steyern, F.; Brosjo, O.; Larsson, O.; et al. Recurrent rearrangement of the PHF1 gene in ossifying fibromyxoid tumors. Am. J. Pathol. 2012, 181, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.; Ali, R.; Melnyk, N.; McAlpine, J.N.; Huntsman, D.G.; Gilks, C.B.; Lee, C.-H.; Oliva, E. Frequency of known gene rearrangements in endometrial stromal tumors. Am. J. Surg. Pathol. 2011, 35, 1364–1372. [Google Scholar] [CrossRef]
- Hofvander, J.; Jo, V.Y.; Fletcher, C.D.; Puls, F.; Flucke, U.; Nilsson, J.; Magnusson, L.; Mertens, F. PHF1 fusions cause distinct gene expression and chromatin accessibil-ity profiles in ossifying fibromyxoid tumors and mesenchymal cells. Mod. Pathol. 2020, 33, 1331–1340. [Google Scholar] [CrossRef]
- Micci, F.; Panagopoulos, I.; Bjerkehagen, B.; Heim, S. Consistent rearrangement of chromosomal band 6p21 with genera-tion of fusion genes JAZF1/PHF1 and EPC1/PHF1 in endometrial stromal sarcoma. Cancer Res. 2006, 66, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Gao, Y.; Lv, Y.; Wu, Y.; Guo, Y.; Du, F.; Wang, S.; Yu, J.; Cao, X.; Li, P.A. Polycomb-like 2 regulates PRC2 components to affect proliferation in glioma cells. J. Neurooncol. 2020, 148, 259–271. [Google Scholar] [CrossRef]
- Brien, G.L.; Healy, E.; Jerman, E.; Conway, E.; Fadda, E.; O’Donovan, D.; Krivtsov, A.V.; Rice, A.M.; Kearney, C.J.; Flaus, A.; et al. A chromatin-independent role of Polycomb-like 1 to stabilize p53 and promote cellular quiescence. Genes Dev. 2015, 29, 2231–2243. [Google Scholar] [CrossRef] [Green Version]
- Maganti, H.B.; Jrade, H.; Cafariello, C.; Manias Rothberg, J.L.; Porter, C.J.; Yockell-Lelièvre, J.; Battaion, H.L.; Khan, S.T.; Howard, J.P.; Li, Y.; et al. Targeting the MTF2-MDM2 Axis Sensitizes Refractory Acute Myeloid Leukemia to Chemotherapy. Cancer Discov. 2018, 8, 1376–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.; Ahn, J.H.; Liu, H.; Tsai, Y.H.; Bhanu, N.V.; Koss, B.; Allison, D.F.; Ma, A.; Storey, A.J.; Wang, P.; et al. PHF19 promotes multiple myeloma tumorigenicity through PRC2 activation and broad H3K27me3 domain formation. Blood 2019, 134, 1176–1189. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Hou, J.; Feng, L.; Lv, A.; Ke, X.; Liang, H.; Wang, F.; Zhang, K.; Chen, K.; Cui, H. PHF19 promotes the proliferation, migration, and chemosensitivity of glioblastoma to doxorubicin through modulation of the SIAH1/beta-catenin axis. Cell Death Dis. 2018, 9, 1049. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Hu, Y.W.; Zhao, J.Y.; Hu, X.M.; Li, S.F.; Wang, Y.C.; Gao, J.J.; Sha, Y.H.; Kang, C.M.; Lin, L.; et al. MicroRNA-195-5p acts as an anti-oncogene by targeting PHF19 in hepatocellular carcinoma. Oncol. Rep. 2015, 34, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Jain, P.; Ballare, C.; Blanco, E.; Vizan, P.; Di Croce, L. PHF19 mediated regulation of proliferation and invasiveness in prostate cancer cells. Elife 2020, 9, e51373. [Google Scholar] [CrossRef] [PubMed]
- Ghislin, S.; Deshayes, F.; Middendorp, S.; Boggetto, N.; Alcaïde-Loridan, C. PHF19 and Akt control the switch between proliferative and invasive states in melanoma. Cell Cycle 2012, 11, 1634–1645. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Li, H.; Liu, G.; Han, S.; Xu, P. Knockdown of JARID2 inhibits the proliferation and invasion of ovarian cancer through the PI3K/Akt signaling pathway. Mol. Med. Rep. 2017, 16, 3600–3605. [Google Scholar] [CrossRef]
- Walters, Z.S.; Villarejo-Balcells, B.; Olmos, D.; Buist, T.W.; Missiaglia, E.; Allen, R.; Al-Lazikani, B.; Garrett, M.D.; Blagg, J.; Shipley, J. JARID2 is a direct target of the PAX3-FOXO1 fusion protein and inhibits myogenic differentiation of rhabdomyosarcoma cells. Oncogene 2014, 33, 1148–1157. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xu, C.; Gao, M.; Ding, B.; Wei, X.; Ji, N. Reduced Expression of Jumonji AT-Rich Interactive Domain 2 (JARID2) in Glioma Inhib-its Tumor Growth In Vitro and In Vivo. Oncol. Res. 2017, 25, 365–372. [Google Scholar] [CrossRef]
- Celik, H.; Koh, W.K.; Kramer, A.C.; Ostrander, E.L.; Mallaney, C.; Fisher, D.A.C.; Xiang, J.; Wilson, W.C.; Martens, A.; Kothari, A.; et al. JARID2 Functions as a Tumor Suppressor in Myeloid Neoplasms by Repressing Self-Renewal in Hematopoietic Progenitor Cells. Cancer Cell 2018, 34, 741–756.e748. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, W.; Gao, Q. β-TRCP-mediated AEBP2 ubiquitination and destruction controls cisplatin resistance in ovarian cancer. Biochem. Biophys. Res. Commun. 2020, 523, 274–279. [Google Scholar] [CrossRef] [PubMed]
- van Lohuizen, M.; Verbeek, S.; Scheijen, B.; Wientjens, E.; van der Gulden, H.; Berns, A. Identification of cooperating oncogenes in E mu-myc transgenic mice by provirus tagging. Cell 1991, 65, 737–752. [Google Scholar] [CrossRef]
- Haupt, Y.; Alexander, W.S.; Barri, G.; Klinken, S.P.; Adams, J.M. Novel zinc finger gene implicated as myc collaborator by retrovirally accelerated lymphomagenesis in E mu-myc transgenic mice. Cell 1991, 65, 753–763. [Google Scholar] [CrossRef]
- Jacobs, J.J.; Scheijen, B.; Voncken, J.W.; Kieboom, K.; Berns, A.; van Lohuizen, M. Bmi-1 collaborates with c-Myc in tumorigenesis by inhibiting c-Myc-induced apoptosis via INK4a/ARF. Genes Dev. 1999, 13, 2678–2690. [Google Scholar] [CrossRef]
- Gunster, M.J.; Satijn, D.P.; Hamer, K.M.; den Blaauwen, J.L.; de Bruijn, D.; Alkema, M.J.; van Lohuizen, M.; van Driel, R.; Otte, A.P. Identification and characterization of interactions between the vertebrate polycomb-group protein BMI1 and human homologs of polyhomeotic. Mol. Cell. Biol. 1997, 17, 2326–2335. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.J.; Datta, S.; Band, V.; Dimri, G.P. Mel-18, a polycomb group protein, regulates cell proliferation and senescence via transcriptional repression of Bmi-1 and c-Myc oncoproteins. Mol. Biol. Cell 2007, 18, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Y.; Tan, Y.; Ma, K.; Cui, J. Bmi-1 is critical for the proliferation and invasiveness of gastric carcinoma cells. J. Gastroenterol. Hepatol. 2010, 25, 568–575. [Google Scholar] [CrossRef]
- Bracken, A.P.; Kleine-Kohlbrecher, D.; Dietrich, N.; Pasini, D.; Gargiulo, G.; Beekman, C.; Theilgaard-Monch, K.; Minucci, S.; Porse, B.T.; Marine, J.C.; et al. The Polycomb group proteins bind throughout the INK4A-ARF locus and are disassociated in senescent cells. Genes Dev. 2007, 21, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Qian, Q.; Cao, D.; Yang, J.; Gui, T.; Shen, K. Role of BMI1 in epithelial ovarian cancer: Investigated via the CRISPR/Cas9 system and RNA sequencing. J. Ovarian Res. 2018, 11, 31. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Liu, J.; Zhang, P.; An, M.; Han, C.; Li, Y.; Guan, X.; Zhang, K. O-GlcNAcylation modulates Bmi-1 protein stability and potential oncogenic function in prostate cancer. Oncogene 2017, 36, 6293–6305. [Google Scholar] [CrossRef]
- Xia, P.; Wang, S.; Huang, G.; Du, Y.; Zhu, P.; Li, M.; Fan, Z. RNF2 is recruited by WASH to ubiquitinate AMBRA1 leading to downregulation of autophagy. Cell Res. 2014, 24, 943–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetsu, O.; Ishihara, H.; Kanno, R.; Kamiyasu, M.; Inoue, H.; Tokuhisa, T.; Taniguchi, M.; Kanno, M. mel-18 negatively regulates cell cycle progression upon B cell antigen receptor stimulation through a cascade leading to c-myc/cdc25. Immunity 1998, 9, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Won, H.Y.; Lee, J.Y.; Shin, D.H.; Park, J.H.; Nam, J.S.; Kim, H.C.; Kong, G. Loss of Mel-18 enhances breast cancer stem cell activity and tumorigenicity through activating Notch signaling mediated by the Wnt/TCF pathway. FASEB J. 2012, 26, 5002–5013. [Google Scholar] [CrossRef]
- Qian, T.; Lee, J.Y.; Park, J.H.; Kim, H.J.; Kong, G. Id1 enhances RING1b E3 ubiquitin ligase activity through the Mel-18/Bmi-1 polycomb group complex. Oncogene 2010, 29, 5818–5827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Won, H.Y.; Park, J.H.; Kim, H.Y.; Choi, H.J.; Shin, D.H.; Kang, J.H.; Woo, J.K.; Oh, S.H.; Son, T.; et al. MEL-18 loss mediates estrogen receptor-alpha downregulation and hormone independence. J. Clin. Investig. 2015, 125, 1801–1814. [Google Scholar] [CrossRef] [Green Version]
- Blackledge, N.P.; Klose, R.J. The molecular principles of gene regulation by Polycomb repressive complexes. Nat. Rev. Mol. Cell Biol. 2021, 22, 815–833. [Google Scholar] [CrossRef]
- Shukla, S.; Ying, W.; Gray, F.; Yao, Y.; Simes, M.L.; Zhao, Q.; Miao, H.; Cho, H.J.; Gonzalez-Alonso, P.; Winkler, A.; et al. Small-molecule inhibitors targeting Polycomb repressive complex 1 RING domain. Nat. Chem. Biol. 2021, 17, 784–793. [Google Scholar] [CrossRef]
- Bolomsky, A.; Schlangen, K.; Schreiner, W.; Zojer, N.; Ludwig, H. Targeting of BMI-1 with PTC-209 shows potent anti-myeloma activity and impairs the tumour microenvironment. J. Hematol. Oncol. 2016, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, Y.; Arai, M.A.; Hara, Y.; Ishibashi, M. Identification of BMI1 promoter inhibitors from Streptomyces sp. IFM-11958. Bioorg. Med. Chem. 2019, 27, 2998–3003. [Google Scholar] [CrossRef]
- Ismail, I.H.; McDonald, D.; Strickfaden, H.; Xu, Z.; Hendzel, M.J. A small molecule inhibitor of polycomb repressive complex 1 inhibits ubiquitin signaling at DNA double-strand breaks. J. Biol. Chem. 2013, 288, 26944–26954. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.; Du, W.; Guo, W. EZH2: A novel target for cancer treatment. J. Hematol. Oncol. 2020, 13, 104. [Google Scholar] [CrossRef]
- Wan, J.; Zhan, J.; Li, S.; Ma, J.; Xu, W.; Liu, C.; Xue, X.; Xie, Y.; Fang, W.; Chin, Y.E.; et al. PCAF-primed EZH2 acetylation regulates its stability and promotes lung adenocarcinoma progression. Nucleic Acids Res. 2015, 43, 3591–3604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, P.W.; Shie, J.J.; Chen, C.H.; Wu, C.Y.; Hsu, T.L.; Wong, C.H. O-GlcNAcylation regulates the stability and enzymatic activity of the histone methyltransferase EZH2. Proc. Natl. Acad. Sci. USA 2018, 115, 7302–7307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Lee, J.S.; Kim, H.; Kim, K.; Park, H.; Kim, J.Y.; Lee, S.H.; Kim, I.S.; Kim, J.; Lee, M.; et al. EZH2 generates a methyl degron that is recognized by the DCAF1/DDB1/CUL4 E3 ubiquitin ligase complex. Mol. Cell 2012, 48, 572–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Kim, M.; Woo, D.H.; Shin, Y.; Shin, J.; Chang, N.; Oh, Y.T.; Kim, H.; Rheey, J.; Nakano, I.; et al. Phosphorylation of EZH2 activates STAT3 signaling via STAT3 methylation and promotes tumorigenicity of glioblastoma stem-like cells. Cancer Cell 2013, 23, 839–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, B.; Liang, J.; Yang, X.; Wang, Y.; Zhao, Y.; Wu, H.; Sun, L.; Zhang, Y.; Chen, Y.; Li, R.; et al. Integration of estrogen and Wnt signaling circuits by the polycomb group protein EZH2 in breast cancer cells. Mol. Cell. Biol. 2007, 27, 5105–5119. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.Y.; Jun, S.; Lee, M.; Kim, H.C.; Wang, X.; Ji, H.; McCrea, P.D.; Park, J.I. PAF and EZH2 induce Wnt/β-catenin signaling hyperactivation. Mol. Cell 2013, 52, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Han, Y.; Wang, X.; Li, N.; Liu, Q.; Yin, Y.; Wang, H.; Pan, L.; Li, L.; Song, K.; et al. SETD2 Restricts Prostate Cancer Metastasis by Integrating EZH2 and AMPK Signaling Pathways. Cancer Cell 2020, 38, 350–365.e7. [Google Scholar] [CrossRef]
- Jin, X.; Yang, C.; Fan, P.; Xiao, J.; Zhang, W.; Zhan, S.; Liu, T.; Wang, D.; Wu, H. CDK5/FBW7-dependent ubiquitination and degradation of EZH2 inhibits pancreatic cancer cell migration and invasion. J. Biol. Chem. 2017, 292, 6269–6280. [Google Scholar] [CrossRef] [Green Version]
- Marchione, D.M.; Lisby, A.; Viaene, A.N.; Santi, M.; Nasrallah, M.; Wang, L.P.; Williams, E.A.; Larque, A.B.; Chebib, I.; Garcia, B.A.; et al. Histone H3K27 dimethyl loss is highly specific for malignant peripheral nerve sheath tumor and distinguishes true PRC2 loss from isolated H3K27 trimethyl loss. Mod. Pathol. 2019, 32, 1434–1446. [Google Scholar] [CrossRef]
- Broux, M.; Prieto, C.; Demeyer, S.; Vanden Bempt, M.; Alberti-Servera, L.; Lodewijckx, I.; Vandepoel, R.; Mentens, N.; Gielen, O.; Jacobs, K.; et al. Suz12 inactivation cooperates with JAK3 mutant signaling in the development of T-cell acute lymphoblastic leukemia. Blood 2019, 134, 1323–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neff, T.; Sinha, A.U.; Kluk, M.J.; Zhu, N.; Khattab, M.H.; Stein, L.; Xie, H.; Orkin, S.H.; Armstrong, S.A. Polycomb repressive complex 2 is required for MLL-AF9 leukemia. Proc. Natl. Acad. Sci. USA 2012, 109, 5028–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessard, J.; Schumacher, A.; Thorsteinsdottir, U.; van Lohuizen, M.; Magnuson, T.; Sauvageau, G. Functional antagonism of the Polycomb-Group genes eed and Bmi1 in hemopoietic cell proliferation. Genes Dev. 1999, 13, 2691–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Phipson, B.; Hyland, C.D.; Leong, H.S.; Allan, R.S.; Lun, A.; Hilton, D.J.; Nutt, S.L.; Blewitt, M.E.; Smyth, G.K.; et al. Polycomb repressive complex 2 (PRC2) suppresses Eμ-myc lymphoma. Blood 2013, 122, 2654–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sashida, G.; Harada, H.; Matsui, H.; Oshima, M.; Yui, M.; Harada, Y.; Tanaka, S.; Mochizuki-Kashio, M.; Wang, C.; Saraya, A.; et al. Ezh2 loss promotes development of myelodysplastic syndrome but attenuates its predisposition to leukaemic transformation. Nat. Commun. 2014, 5, 4177. [Google Scholar] [CrossRef] [Green Version]
- Baude, A.; Lindroth, A.M.; Plass, C. PRC2 loss amplifies Ras signaling in cancer. Nat. Genet. 2014, 46, 1154–1155. [Google Scholar] [CrossRef] [PubMed]
- Comet, I.; Riising, E.M.; Leblanc, B.; Helin, K. Maintaining cell identity: PRC2-mediated regulation of transcription and cancer. Nat. Rev. Cancer 2016, 16, 803–810. [Google Scholar] [CrossRef]
- Fillmore, C.M.; Xu, C.; Desai, P.T.; Berry, J.M.; Rowbotham, S.P.; Lin, Y.J.; Zhang, H.; Marquez, V.E.; Hammerman, P.S.; Wong, K.K.; et al. EZH2 inhibition sensitizes BRG1 and EGFR mutant lung tumours to TopoII inhibitors. Nature 2015, 520, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Serresi, M.; Gargiulo, G.; Proost, N.; Siteur, B.; Cesaroni, M.; Koppens, M.; Xie, H.; Sutherland, K.D.; Hulsman, D.; Citterio, E.; et al. Polycomb Repressive Complex 2 Is a Barrier to KRAS-Driven Inflammation and Epithelial-Mesenchymal Transition in Non-Small-Cell Lung Cancer. Cancer Cell 2016, 29, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Chan, H.L.; Morey, L. Emerging Roles for Polycomb-Group Proteins in Stem Cells and Cancer. Trends Biochem. Sci. 2019, 44, 688–700. [Google Scholar] [CrossRef]
- Martin, M.C.; Zeng, G.; Yu, J.; Schiltz, G.E. Small Molecule Approaches for Targeting the Polycomb Repressive Complex 2 (PRC2) in Cancer. J. Med. Chem. 2020, 63, 15344–15370. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.K.; Marquez, V.E.; Fuller, R.W.; Goldstein, B.M.; Haines, D.R.; McPherson, H.; Parsons, J.L.; Shannon, W.M.; Arnett, G.; Hollingshead, M. Synthesis of 3-deazaneplanocin A, a powerful inhibitor of S-adenosylhomocysteine hydrolase with potent and selective in vitro and in vivo antiviral activities. J. Med. Chem. 1989, 32, 1442–1446. [Google Scholar] [CrossRef]
- Tan, J.; Yang, X.; Zhuang, L.; Jiang, X.; Chen, W.; Lee, P.L.; Karuturi, R.K.; Tan, P.B.; Liu, E.T.; Yu, Q. Pharmacologic disruption of Polycomb-repressive complex 2-mediated gene repression selectively induces apoptosis in cancer cells. Genes Dev. 2007, 21, 1050–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, T.B.; Cortez, C.C.; Yoo, C.B.; Liang, G.; Abe, M.; Kelly, T.K.; Marquez, V.E.; Jones, P.A. DZNep is a global histone methylation inhibitor that reactivates developmental genes not silenced by DNA methylation. Mol. Cancer Ther. 2009, 8, 1579–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, S.K.; Wigle, T.J.; Warholic, N.M.; Sneeringer, C.J.; Allain, C.J.; Klaus, C.R.; Sacks, J.D.; Raimondi, A.; Majer, C.R.; Song, J.; et al. A selective inhibitor of EZH2 blocks H3K27 methylation and kills mutant lymphoma cells. Nat. Chem. Biol. 2012, 8, 890–896. [Google Scholar] [CrossRef]
- Knutson, S.K.; Kawano, S.; Minoshima, Y.; Warholic, N.M.; Huang, K.C.; Xiao, Y.; Kadowaki, T.; Uesugi, M.; Kuznetsov, G.; Kumar, N.; et al. Selective inhibition of EZH2 by EPZ-6438 leads to potent antitumor activity in EZH2-mutant non-Hodgkin lymphoma. Mol. Cancer Ther. 2014, 13, 842–854. [Google Scholar] [CrossRef] [Green Version]
- Hoy, S.M. Tazemetostat: First Approval. Drugs 2020, 80, 513–521. [Google Scholar] [CrossRef]
- Qi, W.; Zhao, K.; Gu, J.; Huang, Y.; Wang, Y.; Zhang, H.; Zhang, M.; Zhang, J.; Yu, Z.; Li, L.; et al. An allosteric PRC2 inhibitor targeting the H3K27me3 binding pocket of EED. Nat. Chem. Biol. 2017, 13, 381–388. [Google Scholar] [CrossRef]
- Rej, R.K.; Wang, C.; Lu, J.; Wang, M.; Petrunak, E.; Zawacki, K.P.; McEachern, D.; Fernandez-Salas, E.; Yang, C.Y.; Wang, L.; et al. EEDi-5285: An Exceptionally Potent, Efficacious, and Orally Active Small-Molecule Inhibitor of Embryonic Ectoderm Development. J. Med. Chem. 2020, 63, 7252–7267. [Google Scholar] [CrossRef]
- Huang, Y.; Sendzik, M.; Zhang, J.; Gao, Z.; Sun, Y.; Wang, L.; Gu, J.; Zhao, K.; Yu, Z.; Zhang, L.; et al. Discovery of the Clinical Candidate MAK683: An EED-Directed, Allosteric, and Selective PRC2 Inhibitor for the Treatment of Advanced Malignancies. J. Med. Chem. 2022, 65, 5317–5333. [Google Scholar] [CrossRef]
- Kim, W.; Bird, G.H.; Neff, T.; Guo, G.; Kerenyi, M.A.; Walensky, L.D.; Orkin, S.H. Targeted disruption of the EZH2-EED complex inhibits EZH2-dependent cancer. Nat. Chem. Biol. 2013, 9, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Chen, L.; Jiao, L.; Jiang, X.; Lian, F.; Lu, J.; Zhu, K.; Du, D.; Liu, J.; Ding, H.; et al. Astemizole arrests the proliferation of cancer cells by disrupting the EZH2-EED interaction of polycomb repressive complex 2. J. Med. Chem. 2014, 57, 9512–9521. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gao, S.; Li, J.; Liu, D.; Sheng, C.; Yao, C.; Jiang, W.; Wu, J.; Chen, S.; Huang, W. Wedelolactone disrupts the interaction of EZH2-EED complex and inhibits PRC2-dependent cancer. Oncotarget 2015, 6, 13049–13059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parreno, V.; Martinez, A.M.; Cavalli, G. Mechanisms of Polycomb group protein function in cancer. Cell Res. 2022, 32, 231–253. [Google Scholar] [CrossRef]
Figure 1.
The classification of mammalian Polycomb repressive complexes. (A) The RING1A/B and Polycomb group RING finger proteins (PCGF1–6) are the catalytic core of PRC1 and they interact with a range of accessory subunits to create different PRC1 variants, mainly divided into canonical PRC1 (cRPC1) and non-canonical PRC1 (ncPRC1). cPRC1 complexes (top) assemble around PCGF2/4 and include Chromobox proteins (CBX2/4/6/7/8), Polyhomeotic protein (PHC1/2/3), and SCM proteins (SCML1/2 or SCMH1). By contrast, ncPRC1 complexes (bottom) can assemble around all six PCGF proteins and contain YY1-associated factor 2 (YAF2) and RING and YY1 binding protein (RYBP). Depending on its PCGF protein, it can be divided into PRC1.1–1.6 variants (also known as ncPRC1.1–1.6). (B) PRC2 is composed of four core subunits, EZH1/2, EED, SUZ12, and RBAP46/48 (also referred to as RBBP4/7), and is divided into PRC2.1 (top) and PRC2.2 (bottom) depending on their auxiliary subunits, of which PRC2.1 can be further subdivided into EPOP-containing and PALI-containing PRC2 variants (top).
Figure 1.
The classification of mammalian Polycomb repressive complexes. (A) The RING1A/B and Polycomb group RING finger proteins (PCGF1–6) are the catalytic core of PRC1 and they interact with a range of accessory subunits to create different PRC1 variants, mainly divided into canonical PRC1 (cRPC1) and non-canonical PRC1 (ncPRC1). cPRC1 complexes (top) assemble around PCGF2/4 and include Chromobox proteins (CBX2/4/6/7/8), Polyhomeotic protein (PHC1/2/3), and SCM proteins (SCML1/2 or SCMH1). By contrast, ncPRC1 complexes (bottom) can assemble around all six PCGF proteins and contain YY1-associated factor 2 (YAF2) and RING and YY1 binding protein (RYBP). Depending on its PCGF protein, it can be divided into PRC1.1–1.6 variants (also known as ncPRC1.1–1.6). (B) PRC2 is composed of four core subunits, EZH1/2, EED, SUZ12, and RBAP46/48 (also referred to as RBBP4/7), and is divided into PRC2.1 (top) and PRC2.2 (bottom) depending on their auxiliary subunits, of which PRC2.1 can be further subdivided into EPOP-containing and PALI-containing PRC2 variants (top).

Figure 2.
PRC1-mediated transcriptional repression and activation. (A) Crosstalk between cPRC1.2/4 and PRC2. PRC2 catalyzes trimethylation of histone H3 at Lys27 (H3K27me3), followed by CBX proteins recognition of H3K27me3, drives cPRC1.2/4 to PRC2 pre-occupied gene loci, and mediates mono-ubiquitination of histone H2A at Lys119 (H2AK119ub1), promoting transcriptional repression. (B) Crosstalk between ncPRC1 and PRC2.2. ncPRC1 first catalyzes H2AK119ub1, then PRC2 recognizes and occupies the same gene locus via JARID2, catalyzes H3K27me3, and promotes transcriptional repression. (C) The cPRC1.2/4 complexes mediate chromatin compaction to repress gene transcription, which is mediated by the interaction of the positively charged region of the CBX proteins with nucleosomes. (D) AUTS2 of ncPRC1.3/5 recruits Tex10 and P300 to acetylate histone H3 at Lys27 (H3K27ac), while promoting transcriptional activation. (E) Subunits or associated proteins of PRC1 directly or indirectly neutralize the E3 enzymatic activity of RING1A/B of PRC1, promoting transcriptional activation. CK2 inhibited its ubiquitination activity directly by phosphorylating RING1A/B, whereas Aurora kinases inhibited H2A ubiquitination indirectly by phosphorylating UBE2D3 and USP16.
Figure 2.
PRC1-mediated transcriptional repression and activation. (A) Crosstalk between cPRC1.2/4 and PRC2. PRC2 catalyzes trimethylation of histone H3 at Lys27 (H3K27me3), followed by CBX proteins recognition of H3K27me3, drives cPRC1.2/4 to PRC2 pre-occupied gene loci, and mediates mono-ubiquitination of histone H2A at Lys119 (H2AK119ub1), promoting transcriptional repression. (B) Crosstalk between ncPRC1 and PRC2.2. ncPRC1 first catalyzes H2AK119ub1, then PRC2 recognizes and occupies the same gene locus via JARID2, catalyzes H3K27me3, and promotes transcriptional repression. (C) The cPRC1.2/4 complexes mediate chromatin compaction to repress gene transcription, which is mediated by the interaction of the positively charged region of the CBX proteins with nucleosomes. (D) AUTS2 of ncPRC1.3/5 recruits Tex10 and P300 to acetylate histone H3 at Lys27 (H3K27ac), while promoting transcriptional activation. (E) Subunits or associated proteins of PRC1 directly or indirectly neutralize the E3 enzymatic activity of RING1A/B of PRC1, promoting transcriptional activation. CK2 inhibited its ubiquitination activity directly by phosphorylating RING1A/B, whereas Aurora kinases inhibited H2A ubiquitination indirectly by phosphorylating UBE2D3 and USP16.

Figure 3.
PRC2-mediated transcriptional regulation. (A) PRC2.1 in transcriptional regulation. All PCL and PALI1/2 proteins stimulate the methyltransferase activity of PRC2.1. PCL1/3 can bind to H3K36me3 and recruit PRC2.1 to gene loci containing high H3K36me3 levels. PCL2 recruits PRC2.1 to unmethylated CpG islands. Moreover, PCL3 can recruit the demethylase NO66 to erase the trimethylation modification of H3K36, allowing the deposition of H3K27me3 and further recruitment PRC2, promoting PcG-mediated transcriptional repression of target genes (top). EPOP associates with Elongin B/C (EloB/C), which binds Elongin A (EloA) and fine-tunes the transcriptional repression function of PRC2 on target genes (bottom). (B) PRC2.2 in transcriptional regulation. Both JARID2 and AEBP2 enhance the methyltransferase activity of EZH2 and can recruit PRC2.2 to PcG-targeted gene loci by binding H2AK119ub1. JARID2 is methylated at Lys116 by PRC2.2 and then recognized by EED, which allosterically activates the catalytic activity of EZH2, whereas AEBP2 stimulates the catalytic activity of EZH2 by increasing the stability of PRC2.2 (top). EED allosterically activates the catalytic activity of EZH2 by recognizing H3K27me3 and enhances PRC2.2-mediated transcriptional repression, whereas EZH inhibitory protein (EZHIP) can bind to allosterically activated PRC2.2 and inhibit its methyltransferase activity (bottom).
Figure 3.
PRC2-mediated transcriptional regulation. (A) PRC2.1 in transcriptional regulation. All PCL and PALI1/2 proteins stimulate the methyltransferase activity of PRC2.1. PCL1/3 can bind to H3K36me3 and recruit PRC2.1 to gene loci containing high H3K36me3 levels. PCL2 recruits PRC2.1 to unmethylated CpG islands. Moreover, PCL3 can recruit the demethylase NO66 to erase the trimethylation modification of H3K36, allowing the deposition of H3K27me3 and further recruitment PRC2, promoting PcG-mediated transcriptional repression of target genes (top). EPOP associates with Elongin B/C (EloB/C), which binds Elongin A (EloA) and fine-tunes the transcriptional repression function of PRC2 on target genes (bottom). (B) PRC2.2 in transcriptional regulation. Both JARID2 and AEBP2 enhance the methyltransferase activity of EZH2 and can recruit PRC2.2 to PcG-targeted gene loci by binding H2AK119ub1. JARID2 is methylated at Lys116 by PRC2.2 and then recognized by EED, which allosterically activates the catalytic activity of EZH2, whereas AEBP2 stimulates the catalytic activity of EZH2 by increasing the stability of PRC2.2 (top). EED allosterically activates the catalytic activity of EZH2 by recognizing H3K27me3 and enhances PRC2.2-mediated transcriptional repression, whereas EZH inhibitory protein (EZHIP) can bind to allosterically activated PRC2.2 and inhibit its methyltransferase activity (bottom).

Figure 4.
Multifaceted roles of PRC1 in cancer. (A) The c-Myc protein leads to increased PCGF4 (BMI-1) expression and promotes the transcription repression of target genes p19ARF and p16 repressed at the Ink4a/ARF locus, promoting the proliferation of cancer cells. In contrast, PCGF2 (MEL-18) plays a tumor suppressor role by inhibiting the transcription of c-Myc. (B) PTMs of PRC1 subunits can promote tumorigenesis. Deposition of O-GlcNAcylation on BMI-1 improves its protein stability and inhibits its degradation. Increased levels of BMI-1 protein enhance transcriptional silencing of downstream target genes such as p53, p16, and PTEN, thereby promoting cancer cell proliferation, metastasis, and angiogenesis. (C) PRC1 may also be involved in tumorigenesis and development in a PRC2-independent manner. Interestingly, PRC1 is also found on specific targets that lack the H3K27me3 mark, and these gene loci deposit active marks such as H3K27ac and H3K4me3. The PRC1 subunits PCGF1, PCGF3, and PCGF6 were reported to activate related mechanisms such as PI3K/Akt pathway, EMT, proliferation, and metastasis, thus promoting tumorigenesis. (D) The catalytic subunit of PRC1, RING1B, promotes tumorigenesis through a dual role of gene repression and activation. It promotes cell proliferation and metastasis by activating the transcription of genes such as CCND2 and ZEB2. Meanwhile, it can also promote cell proliferation, invasion, and metastasis by inhibiting the transcription of genes such as LTBP2 and E-cadherin. (E) Overexpression of MEL-18 has been found to play a tumor suppressor role in a variety of cancers. MEL-18 negatively regulates cancer cell proliferation, angiogenesis, invasion, and metastasis by inhibiting the expression of genes such as cyclin D1, Jagged-1, HIF-1α, and ZEB1/2.
Figure 4.
Multifaceted roles of PRC1 in cancer. (A) The c-Myc protein leads to increased PCGF4 (BMI-1) expression and promotes the transcription repression of target genes p19ARF and p16 repressed at the Ink4a/ARF locus, promoting the proliferation of cancer cells. In contrast, PCGF2 (MEL-18) plays a tumor suppressor role by inhibiting the transcription of c-Myc. (B) PTMs of PRC1 subunits can promote tumorigenesis. Deposition of O-GlcNAcylation on BMI-1 improves its protein stability and inhibits its degradation. Increased levels of BMI-1 protein enhance transcriptional silencing of downstream target genes such as p53, p16, and PTEN, thereby promoting cancer cell proliferation, metastasis, and angiogenesis. (C) PRC1 may also be involved in tumorigenesis and development in a PRC2-independent manner. Interestingly, PRC1 is also found on specific targets that lack the H3K27me3 mark, and these gene loci deposit active marks such as H3K27ac and H3K4me3. The PRC1 subunits PCGF1, PCGF3, and PCGF6 were reported to activate related mechanisms such as PI3K/Akt pathway, EMT, proliferation, and metastasis, thus promoting tumorigenesis. (D) The catalytic subunit of PRC1, RING1B, promotes tumorigenesis through a dual role of gene repression and activation. It promotes cell proliferation and metastasis by activating the transcription of genes such as CCND2 and ZEB2. Meanwhile, it can also promote cell proliferation, invasion, and metastasis by inhibiting the transcription of genes such as LTBP2 and E-cadherin. (E) Overexpression of MEL-18 has been found to play a tumor suppressor role in a variety of cancers. MEL-18 negatively regulates cancer cell proliferation, angiogenesis, invasion, and metastasis by inhibiting the expression of genes such as cyclin D1, Jagged-1, HIF-1α, and ZEB1/2.

Figure 5.
Multifaceted roles of PRC2 in cancer. (A) PRC2-mediated gene silencing was found to have both oncogenic and tumor-suppressive roles. Overexpression or gain-of-function mutations of EZH2 enhance the catalytic activity of PRC2 and its transcriptional inhibition of E-cadherin, p21, p16, cyclinD1, and other genes, thus promoting the growth, invasion, and metastasis of tumor cells, which is also related to drug resistance and poor prognosis. Surprisingly, loss-of-function mutants of EZH2 are also found in certain cancers and promote tumorigenesis, implicating the tumor suppressor role of PRC2 as well. (B) PTMs of EZH2 can promote or inhibit tumorigenesis. Deposition of O-GlcNAcylation and Acetylation on EZH2 inhibits its degradation and enhances the catalytic activity of PRC2. Conversely, deposition of methylation and phosphorylation on EZH2 promotes its degradation and inhibits the catalytic activity of PRC2. (C) Methylation of non-histone proteins mediated by EZH2 promotes tumorigenesis in a PRC2-independent manner. Methylation of RORα mediated by EZH2 promotes its degradation and promotes tumor cell proliferation, metastasis, and invasion. In addition, phosphorylated EZH2-mediated STAT3 methylation promotes the expression of SOCS3, c-Myc, and other genes, thereby promoting the self-renewal of stem cell populations (D) EZH2 binds to other proteins to promote the expression of tumor-related genes. EZH2 interacts with ERα and β-catenin and activates c-Myc and cyclin D1 expression, which promotes cell proliferation. EZH2 interacts with ERα and β-catenin and stimulates the expression of Wnt pathway-related genes, thus promoting tumorigenesis.
Figure 5.
Multifaceted roles of PRC2 in cancer. (A) PRC2-mediated gene silencing was found to have both oncogenic and tumor-suppressive roles. Overexpression or gain-of-function mutations of EZH2 enhance the catalytic activity of PRC2 and its transcriptional inhibition of E-cadherin, p21, p16, cyclinD1, and other genes, thus promoting the growth, invasion, and metastasis of tumor cells, which is also related to drug resistance and poor prognosis. Surprisingly, loss-of-function mutants of EZH2 are also found in certain cancers and promote tumorigenesis, implicating the tumor suppressor role of PRC2 as well. (B) PTMs of EZH2 can promote or inhibit tumorigenesis. Deposition of O-GlcNAcylation and Acetylation on EZH2 inhibits its degradation and enhances the catalytic activity of PRC2. Conversely, deposition of methylation and phosphorylation on EZH2 promotes its degradation and inhibits the catalytic activity of PRC2. (C) Methylation of non-histone proteins mediated by EZH2 promotes tumorigenesis in a PRC2-independent manner. Methylation of RORα mediated by EZH2 promotes its degradation and promotes tumor cell proliferation, metastasis, and invasion. In addition, phosphorylated EZH2-mediated STAT3 methylation promotes the expression of SOCS3, c-Myc, and other genes, thereby promoting the self-renewal of stem cell populations (D) EZH2 binds to other proteins to promote the expression of tumor-related genes. EZH2 interacts with ERα and β-catenin and activates c-Myc and cyclin D1 expression, which promotes cell proliferation. EZH2 interacts with ERα and β-catenin and stimulates the expression of Wnt pathway-related genes, thus promoting tumorigenesis.

Figure 6.
Inhibitor structures targeting PRC1 or PRC2.


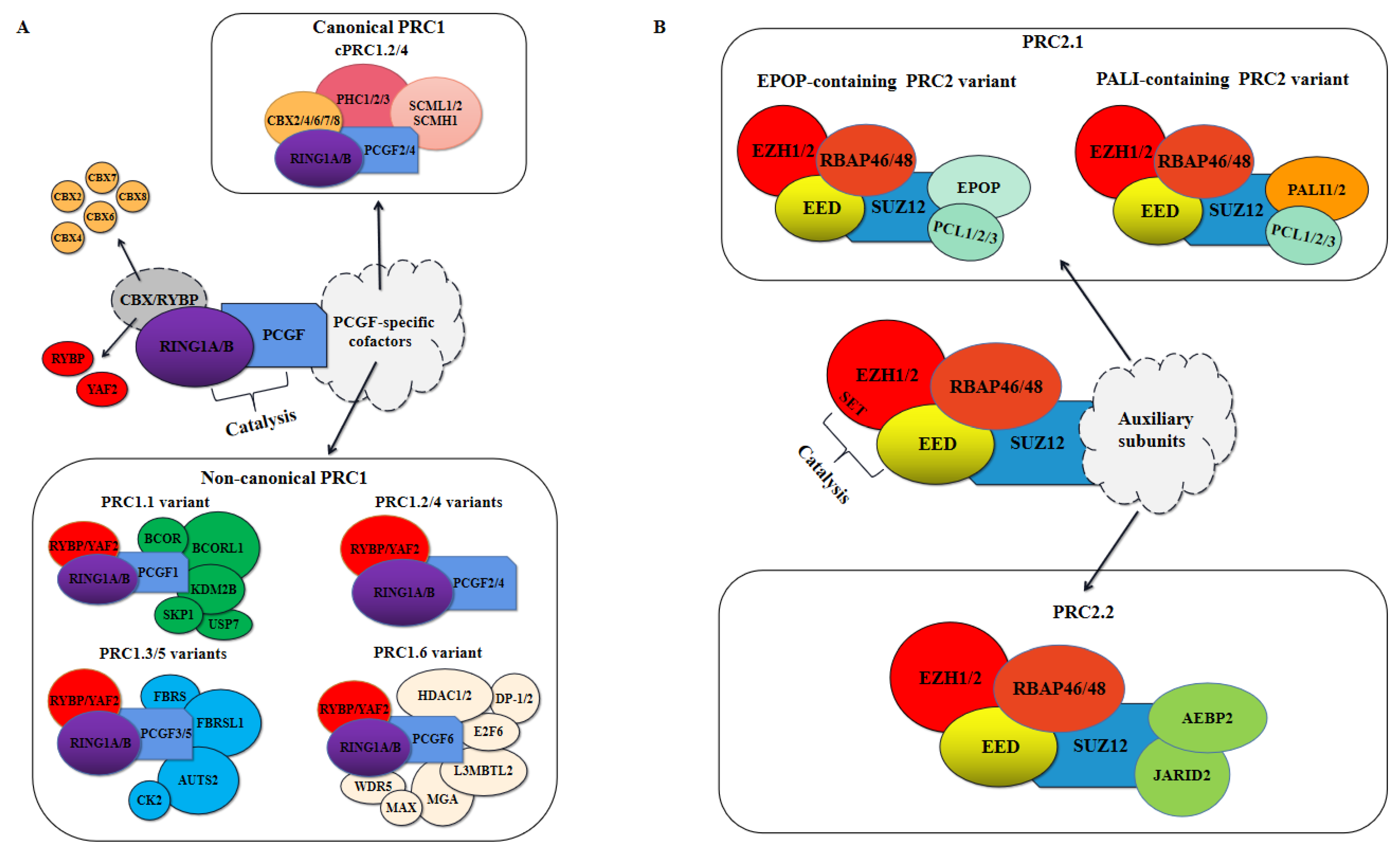{kind=link}
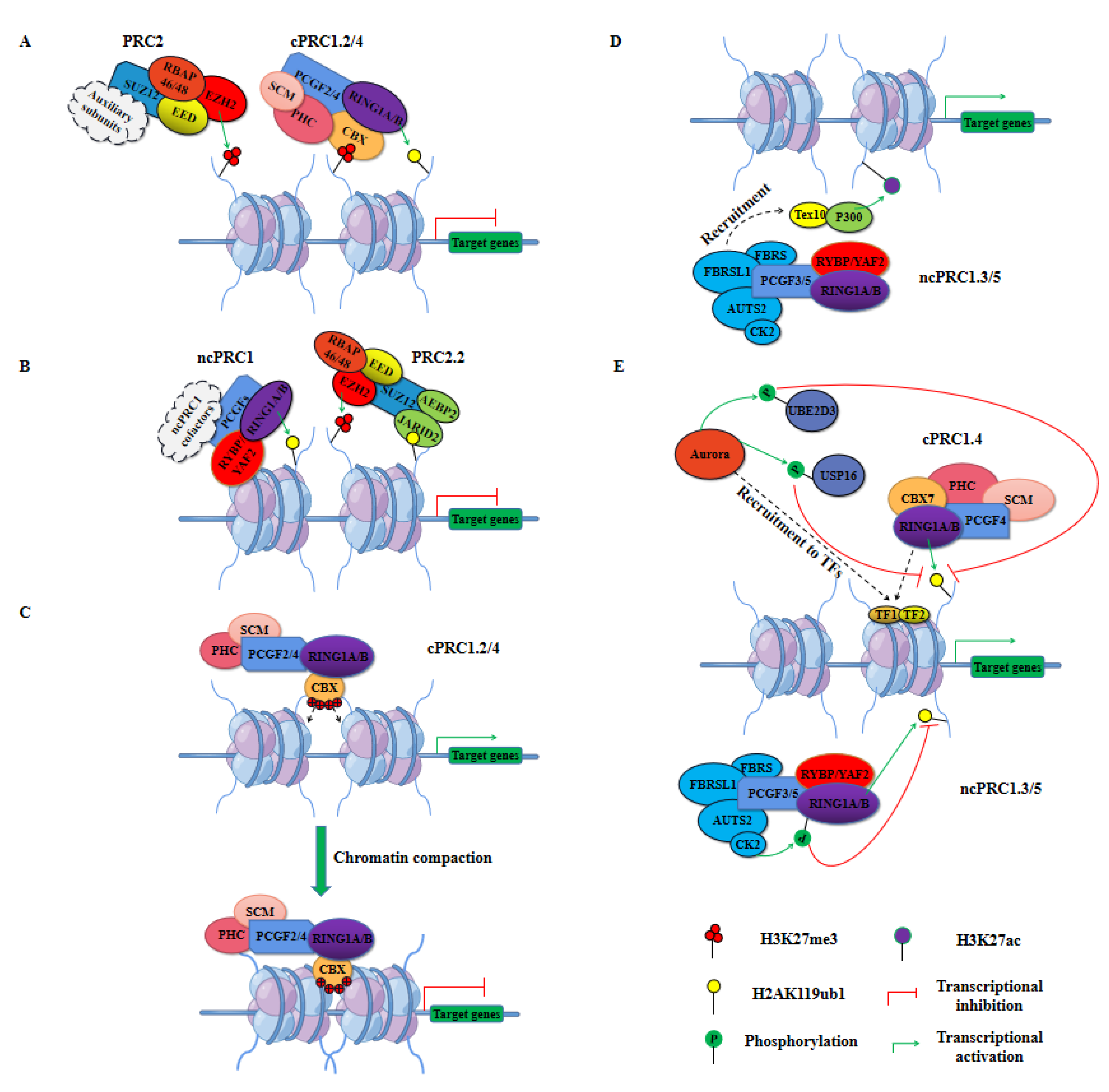{kind=link}
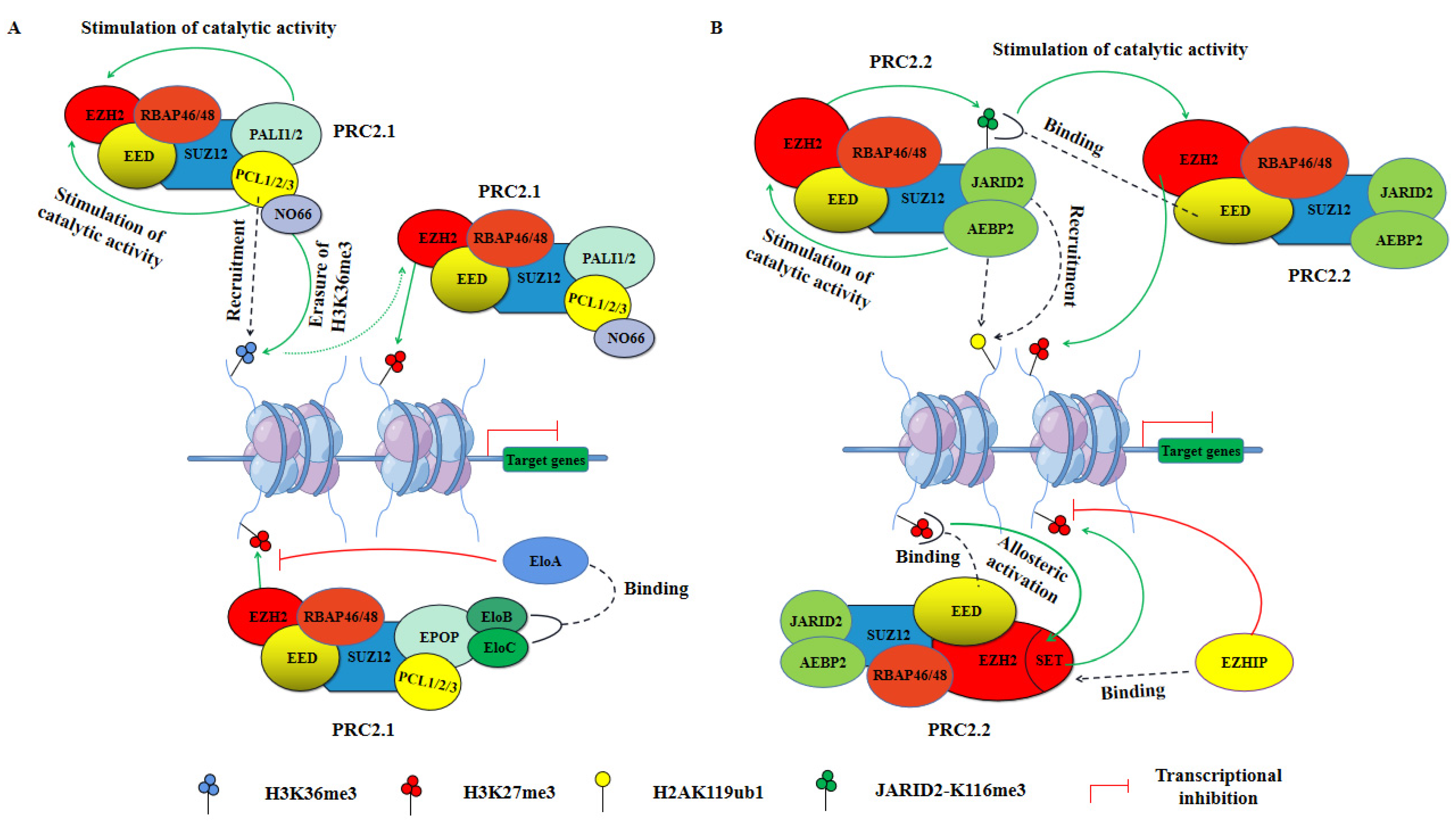{kind=link}
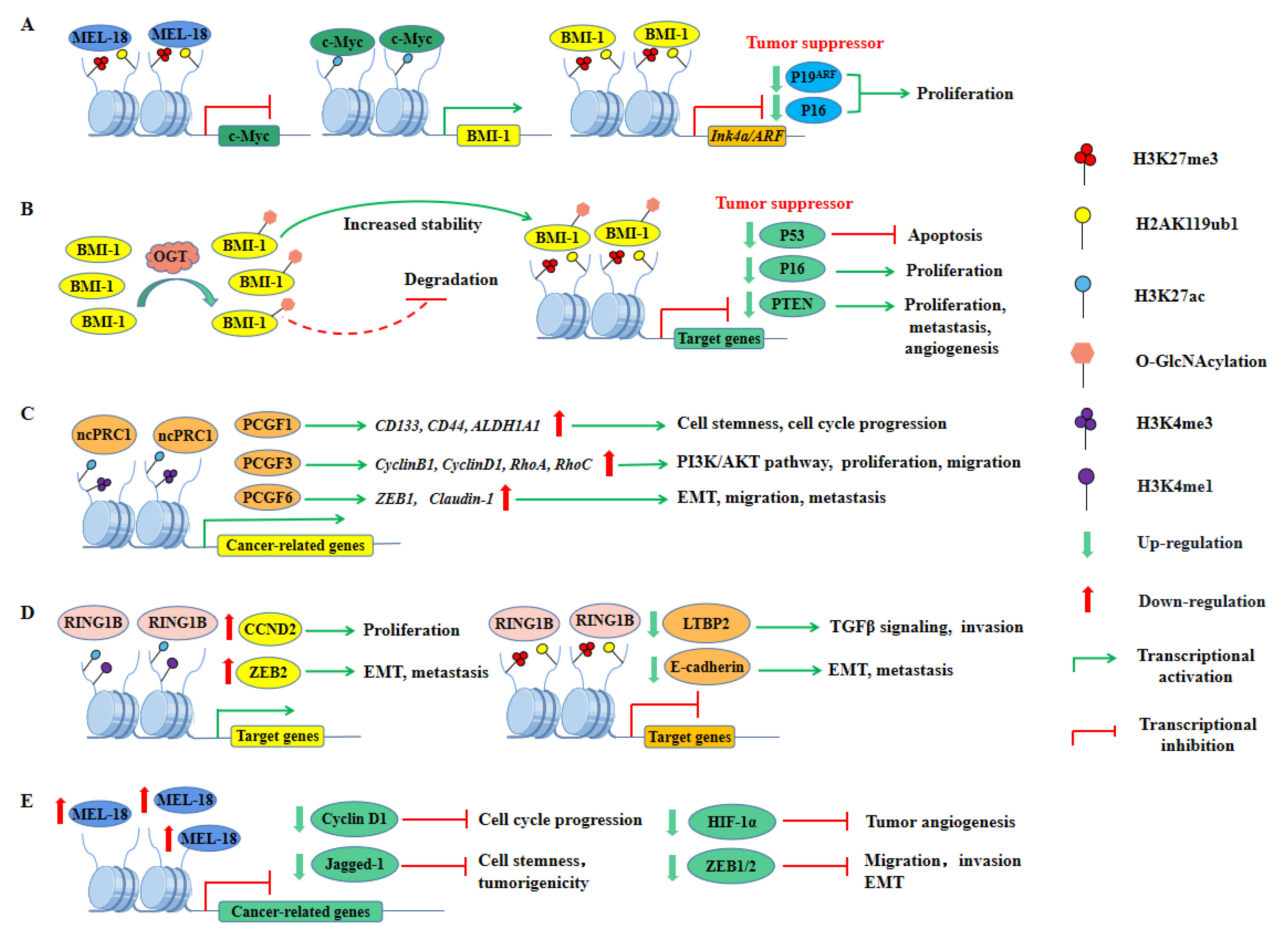{kind=link}
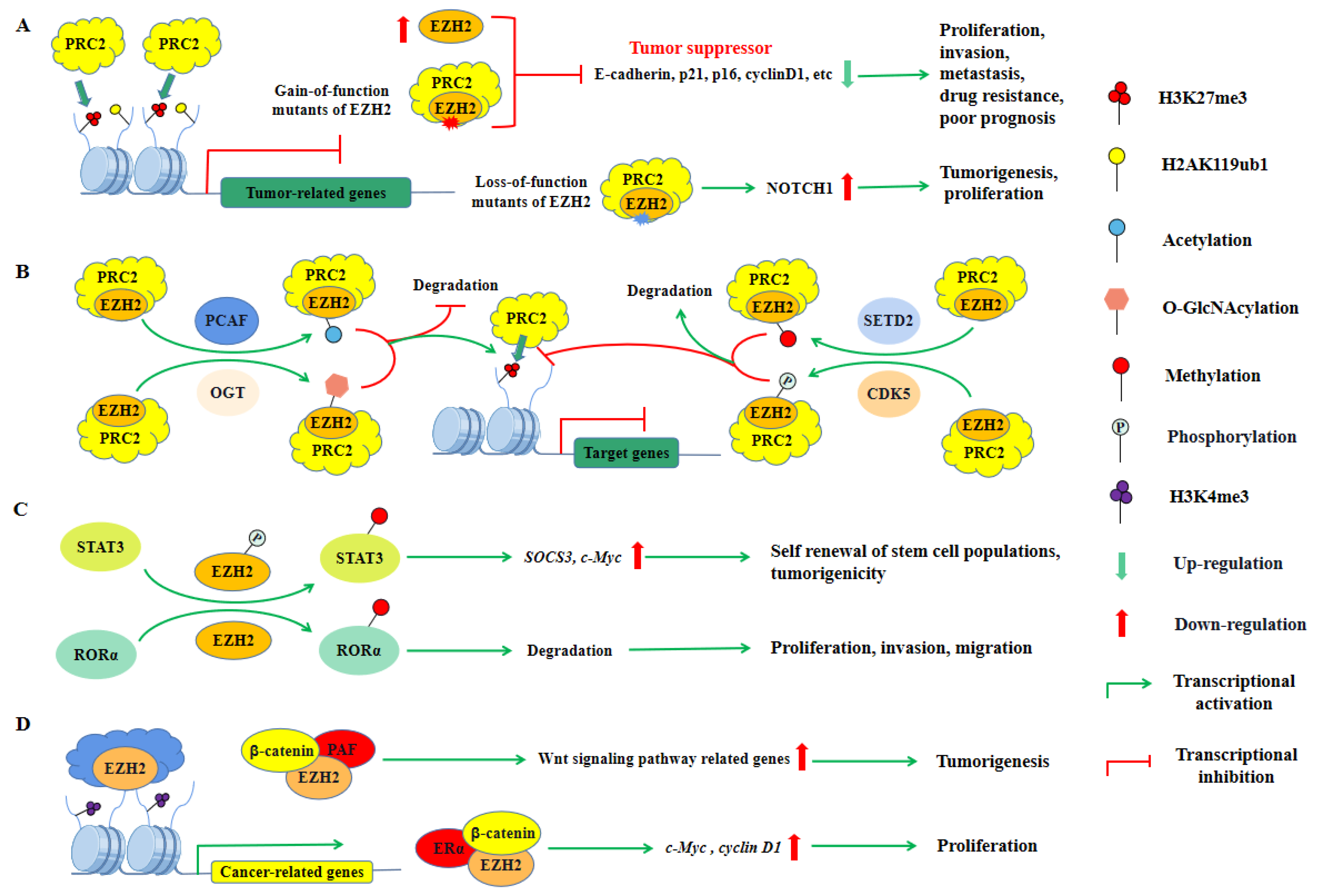{kind=link}
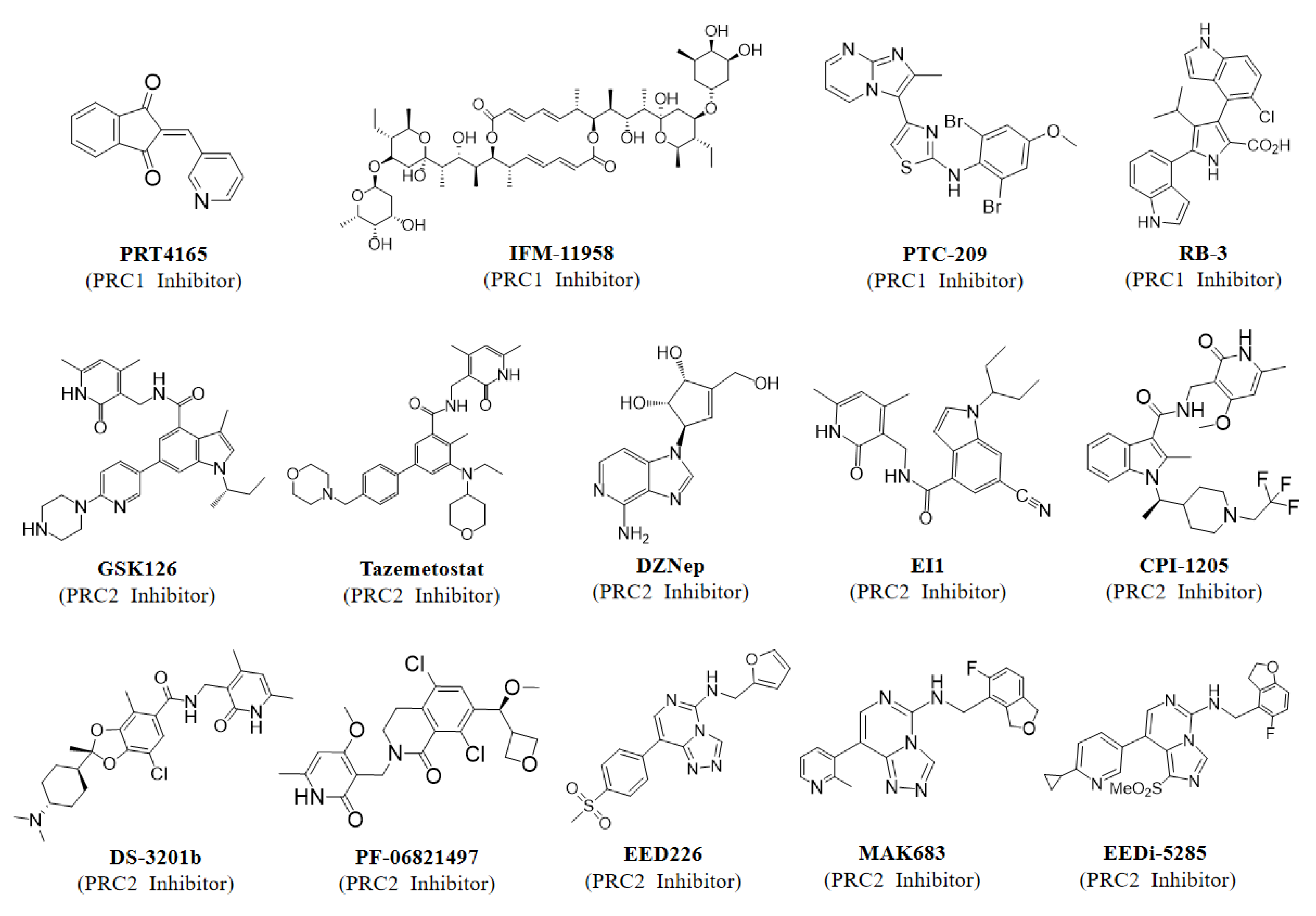{kind=link}
Table 1.
PRC1 components and their roles in cancer.
| Complex a | Subunit b | Function | Descriptions | Cancer type | Refs. |
|---|---|---|---|---|---|
| cPRC1 or ncPRC1 | RING1A | Oncogenic | Overexpression of RING1A promotes oncogene expression | Acute myeloid leukemia | [25] |
| RING1B (RNF2) | Oncogenic | Overexpression of RING1B promotes oncogene expression | Acute myeloid leukemia, | [25,121,122] | |
| RING1B drives proliferation by upregulating transcription of cell cycle regulators | Melanoma | [24] | |||
| RING1B not only promotes the expression of oncogenes but also regulates chromatin accessibility, and improves enhancer activity thus facilitating gene transcription | Breast cancer | [123,124,125] | |||
| RING1B promotes p53 protein degradation | Colorectal cancer, Hepatocellular carcinoma | [126,127] | |||
| PCGF2 (MEL-18) | Tumor suppressor | Overexpression of PCGF2 suppresses oncogene expression | Gastric cancer, Breast cancer | [31,128,129,130] | |
| PCGF4 (BMI-1) | Oncogenic | PCGF4 represses transcription of the Ink4a/ARF locus to promote cellular immortality | Non-small cell lung cancer Lymphoid, Breast cancer, | [131,132,133,134] | |
| PCGF4 promotes stem cell expansion and tumorigenicity in an Ink4a/ARF independent manner | Hepatocellular carcinoma, Glioma | [28,135] | |||
| Ectopic expression of PCGF4 induces EMT and enhances tumor cell invasion and metastasis | Breast cancer, Nasopharyngeal cancer | [132,136,137] | |||
| PCGF4 binds the androgen receptor (AR) and increases its stability, enhancing AR signaling in prostate cancer cells in a PRC1-independent manner | Prostate cancer | [138,139] | |||
| cPRC1 | PHC3 | Tumor suppressor | Missense mutations in PHC3 promote cell proliferation (G201C) | Osteosarcoma | [140,141] |
| cPRC1 | CBX2 | Oncogenic | Loss of CBX2 inhibits cell proliferation, invasion, and migration | Gastric cancer, Breast cancer | [142,143] |
| Knockdown of CBX2 inhibits cell proliferation and promotes apoptosis | Ovarian cancer | [26] | |||
| CBX4 | Oncogenic | Promotes angiogenesis and metastasis of tumors | Hepatocellular carcinoma | [144,145] | |
| CBX4 significantly promotes tumors growth and metastasis | Clear cell renal cell carcinoma | [146] | |||
| Tumor suppressor | Overexpression of CBX4 inhibits cell migration, invasion, and metastasis | Colorectal carcinoma | [147] | ||
| CBX6 | Oncogenic | Overexpression of CBX6 promotes EMT | Hepatocellular carcinoma | [148] | |
| Tumor suppressor | Exogenous overexpression of CBX6 inhibits cell proliferation, migration, and invasion, and induces cell cycle arrest | Breast cancer | [29] | ||
| CBX7 | Oncogenic | CBX7 represses transcription of the Ink4a/ARF locus and promotes tumorigenesis and invasion | Lymphoma, Prostate cell | [149,150] | |
| Overexpression of CBX7 promotes E-cadherin expression and is required for cell migration and invasion | Thyroid neoplasia, Cervical cancer | [151,152] | |||
| Tumor suppressor | Reduced expression of CBX7 in cancer promotes cell progression and proliferation | Lung cancer, Bladder cancer | [153,154,155] | ||
| CBX7 inhibits tumor proliferation by inactivating the tumor necrosis factor (TNF) signaling pathway | Clear cell renal cell carcinoma | [156] | |||
| CBX8 | Oncogenic | Premature senescence and growth arrest of cancer cells are suppressed by CBX8 | Breast cancer, Leukemia | [157,158] | |
| Ectopic expression of CBX8 promoted tumor metastasis and growth, and overexpression of CBX8 in hepatocellular carcinoma cells activated Akt/β-catenin signaling | Hepatocellular carcinoma | [159,160] | |||
| cPRC1 | CBX8 | Oncogenic | CBX8 contributes to tumorigenesis or promotes stemness in specific tumors by acting non canonically | Mammary carcinoma, Colon cancer | [161,162] |
| ncPRC1 | PCGF1 | Oncogenic | Overexpression of PCGF1 promotes tumor cell cycle progression and cell proliferation | Cervical carcinoma | [163] |
| PCGF1 activated stemness markers and promoted stem cell enrichment and self-renewal | Colorectal cancer, Malignant glioma | [164,165] | |||
| PCGF3 | Oncogenic | Overexpression of PCGF3 promoted cancer cell proliferation and migration. | Non-small cell lung cancer | [166] | |
| PCGF6 | Oncogenic | PCGF6 promotes cell migration and metastasis by driving EMT | Breast cancer | [167] | |
| RYBP | Oncogenic | Silencing of RYBP inhibits melanoma cell proliferation, migration, and invasion | Melanoma | [168] | |
| Tumor suppressor | Overexpression of RYBP inhibits the degradation of tumor suppressor proteins and reduces cancer cell proliferation, migration, and metastasis | Breast cancer, Colon cancer, Lung cancer, Thyroid cancer, Hepatocellular carcinoma | [169,170,171,172] | ||
| YAF2 | Oncogenic | YAF2 is overexpressed in a variety of cancers and has been found to inhibit apoptosis | Non-small cell lung cancer, Breast cancer, Colon cancer | [173,174] | |
| KDM2B | Oncogenic | Overexpression of KDM2B Promotes the self-renewal of cancer stem cells | Breast cancer, Acute myeloid leukemia | [175,176] | |
| Tumor suppressor | Represses the expression of Notch pathway-related genes | T-ALL | [177] | ||
| BCOR | Oncogenic | BCOR promotes PRC1-mediated dysregulation of expression through gene fusion | Endometrial stromal sarcoma | [178,179] | |
| ncPRC1 | BCOR | Tumor suppressor | Overexpression of BCOR inhibits cancer stem cell proliferation and self-renewal | T-ALL | [180] |
| AUTS2 | Oncogenic | The fusion PAX5-AUTS2 is a recurrent fusion gene in B-cell acute lymphoblastic leukemia | B-ALL | [181] |
Complex a, different variants of PRC1; Subunit b, defined as a Polycomb group (PcG) protein present in the specific variant; Italicized format, gene name; Superscript numbers, references.
Table 2.
PRC2 components and their roles in cancer.
| Complex a | Subunit b | Function | Descriptions | Cancer type | Refs. |
|---|---|---|---|---|---|
| PRC2.1 or PRC2.2 | EZH2 | Oncogenic | EZH2 is overexpressed in tumors and promotes tumor cell proliferation and invasion | Prostate cancer, Breast cancer, Bladder cancer, Gastric cancer, Lymphoma, etc. | [15,16,182,183,184,185,186,187,188] |
| Gain-of-function mutations in EZH2 increase the methylation levels of PcG-targeted genes and promote cell proliferation | Lymphoma | [18,20,189,190] | |||
| Tumor suppressor | Loss-of-function (LOF) mutations or deletions of EZH2 promote oncogene expression | T-ALL, MPN | [191,192,193,194] | ||
| EZH2 inhibits cell proliferation | Breast cancer, AML, B-ALL | [195,196,197] | |||
| EZH1 | Oncogenic | EZH1 and EZH2 are required for epithelial-mesenchymal transition (EMT) and cell proliferation | Breast cancer, MPNST | [198,199] | |
| EED | Oncogenic | Overexpression of EED in tumors promotes EMT and is also required for cell proliferation | Lymphoma, Breast cancer, Colorectal cancer | [17,23,200,201,202,203] | |
| LOF mutation (I363M) in EED reduces PRC2 catalytic activity and causes increased susceptibility to myeloid cancers | Myeloid cancers | [204,205] | |||
| Tumor suppressor | Recurrent inactivation of EED or SUZ12 is found in malignant peripheral nerve sheath tumors (MPNST) | MPNST | [23] | ||
| SUZ12 | Oncogenic | SUZ12 is overexpressed in a variety of tumors and exerts oncogenic functions by repressing tumor suppressor genes and promoting oncogene expression | Ovarian cancer, Colorectal cancer, HNSCC, Breast cancer | [201,206,207,208] | |
| Tumor suppressor | SUZ12 inhibits tumor migration, invasion, and development | Liver cancer, Gliomas, Melanomas | [209,210] | ||
| Inactivation of SUZ12 is found in MPNST and T-cell acute lymphoblastic leukemia (T-ALL) | MPNST, T-ALL | [23,192,211,212] | |||
| PRC2.1 or PRC2.2 | RBAP46 (RBBP7) | Oncogenic | RBAP46 is required for tumorigenesis | Bladder cancer, Prostate cancer | [213,214] |
| PRC2.1 | EPOP | Oncogenic | The oncogenic role may be mediated through its interaction with EloB/C and USP7 to modulate the chromatin landscape | Colon cancer, Breast cancer | [75] |
| PCL1 (PHF1) | Oncogenic | Might contribute to oncogene expression by fusions in chromosomal translocations that alter chromatin accessibility | Ossifying fibromyxoid tumor, Endometrial stromal tumor | [215,216,217,218] | |
| PCL2 (MTF2) | Oncogenic | Upregulates the expression levels of EED and EZH2 and increases the catalytic activity of PRC2 | Gliomas | [219] | |
| Tumor suppressor | Overexpression of PCL2 stabilizes p53 to promote cellular quiescence | Myeloid leukemia | [220,221] | ||
| PCL3 (PHF19) | Oncogenic | Increases PRC2 activity | Multiple myeloma, Hepatocellular carcinoma, Glioblastoma | [222,223,224] | |
| Tumor suppressor | Inhibits angiogenesis and invasion of tumor cells | Prostate cancer, Melanoma | [225,226] | ||
| PRC2.2 | JARID2 | Oncogenic | Overexpression of JARID2 promotes invasion and metastasis | Ovarian cancer, Glioma Rhabdomyosarcoma | [227,228,229] |
| Tumor suppressor | Inhibition of self-renewal pathways | Myeloid neoplasms | [230] | ||
| AEBP2 | Oncogenic | Inactivation of AEBP2 inhibits proliferation and reduces chemoresistance in ovarian cancer cells | Ovarian cancer | [231] |
Complex a, different variants of PRC2; Subunit b, defined as a Polycomb group (PcG) protein present in the specific variant; Superscript numbers, references.
Table 3.
PcG subunit inhibitors undergoing clinical trials.
| Target | Drug | Cancer | Phase | NCT ID |
|---|---|---|---|---|
| EZH2 | Tazemetostat (EPZ-6438, E7438) CAS: 1403254-99-8 | Follicular lymphoma | 3 | NCT04224493 |
| Advanced epithelioid sarcoma | 3 | NCT04204941 | ||
| Follicular Lymphoma | 2 | NCT05152459 | ||
| Diffuse large B-cell lymphoma, Hematologic neoplasms, Mantle cell lymphoma | 2 | NCT05205252 | ||
| Nasal Cancer | 2 | NCT05151588 | ||
| Synovial sarcoma, Epithelioid sarcoma | 2 | NCT02601950 | ||
| Malignant peripheral nerve sheath tumors | 2 | NCT04917042 | ||
| Metastatic melanoma | 2 | NCT04557956 | ||
| Small cell lung cancer (SCLC) | 1 | NCT05353439 | ||
| Metastatic prostate cancer | 1 | NCT04846478 | ||
| Advanced solid tumors, Hematologic tumors | 1 | NCT04537715 | ||
| B-cell non-Hodgkin lymphoma | 1 | NCT03009344 | ||
| Lirametostat (CPI-1205) CAS: 1621862-70-1 | Metastatic castration-resistant prostate cancer | 2 | NCT03480646 | |
| B-cell lymphoma | 1 | NCT02395601 | ||
| Advanced solid tumors | 1 | NCT03525795 | ||
| Valemetostat (DS-3201b) CAS: 1809336-39-7 | T-cell lymphoma | 2 | NCT04703192 | |
| B-cell lymphoma | 2 | NCT04842877 | ||
| SCLC | 2 | NCT03879798 | ||
| Lymphomas | 1 | NCT02732275 | ||
| Metastatic prostate cancer, Urothelial carcinoma, Renal cell carcinoma | 1 | NCT04388852 | ||
| SHR-2554 | Advanced solid tumors, B-cell lymphomas | 2 | NCT04407741 | |
| Advanced breast cancer | 2 | NCT04355858 | ||
| Lymphomas | 1 | NCT03603951 | ||
| Lymphomas | 1 | NCT05049083 | ||
| CPI-0209 | Solid tumors | 2 | NCT04104776 | |
| PF-06821497 CAS: 1844849-11-1 | Castration-resistant prostate cancer, Follicular lymphoma, SCLC | 1 | NCT03460977 | |
| EED | MAK-683 CAS: 1951408-58-4 | Diffuse large B-cell lymphoma | 2 | NCT02900651 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dong, G.-J.; Xu, J.-L.; Qi, Y.-R.; Yuan, Z.-Q.; Zhao, W. Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer. Int. J. Mol. Sci. 2022, 23, 9574. https://doi.org/10.3390/ijms23179574
AMA Style
Dong G-J, Xu J-L, Qi Y-R, Yuan Z-Q, Zhao W. Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer. International Journal of Molecular Sciences. 2022; 23(17):9574. https://doi.org/10.3390/ijms23179574
Chicago/Turabian StyleDong, Guan-Jun, Jia-Le Xu, Yu-Ruo Qi, Zi-Qiao Yuan, and Wen Zhao. 2022. "Critical Roles of Polycomb Repressive Complexes in Transcription and Cancer" International Journal of Molecular Sciences 23, no. 17: 9574. https://doi.org/10.3390/ijms23179574
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.