

Trifuhalol A Suppresses Allergic Inflammation through Dual Inhibition of TAK1 and MK2 Mediated by IgE and IL-33
 and
and 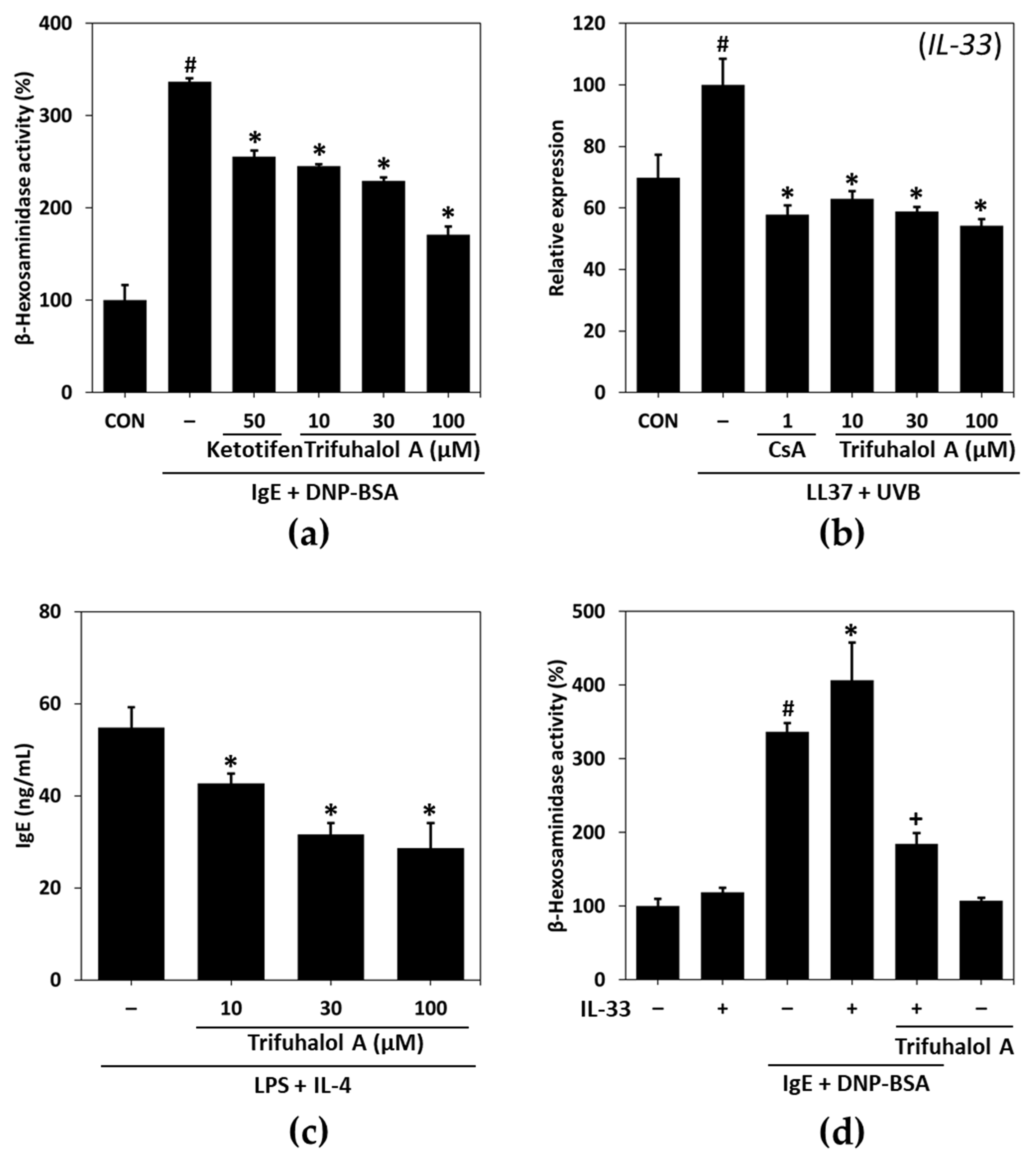{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Trifuhalol A on Allergic Inflammation-Related Mediators
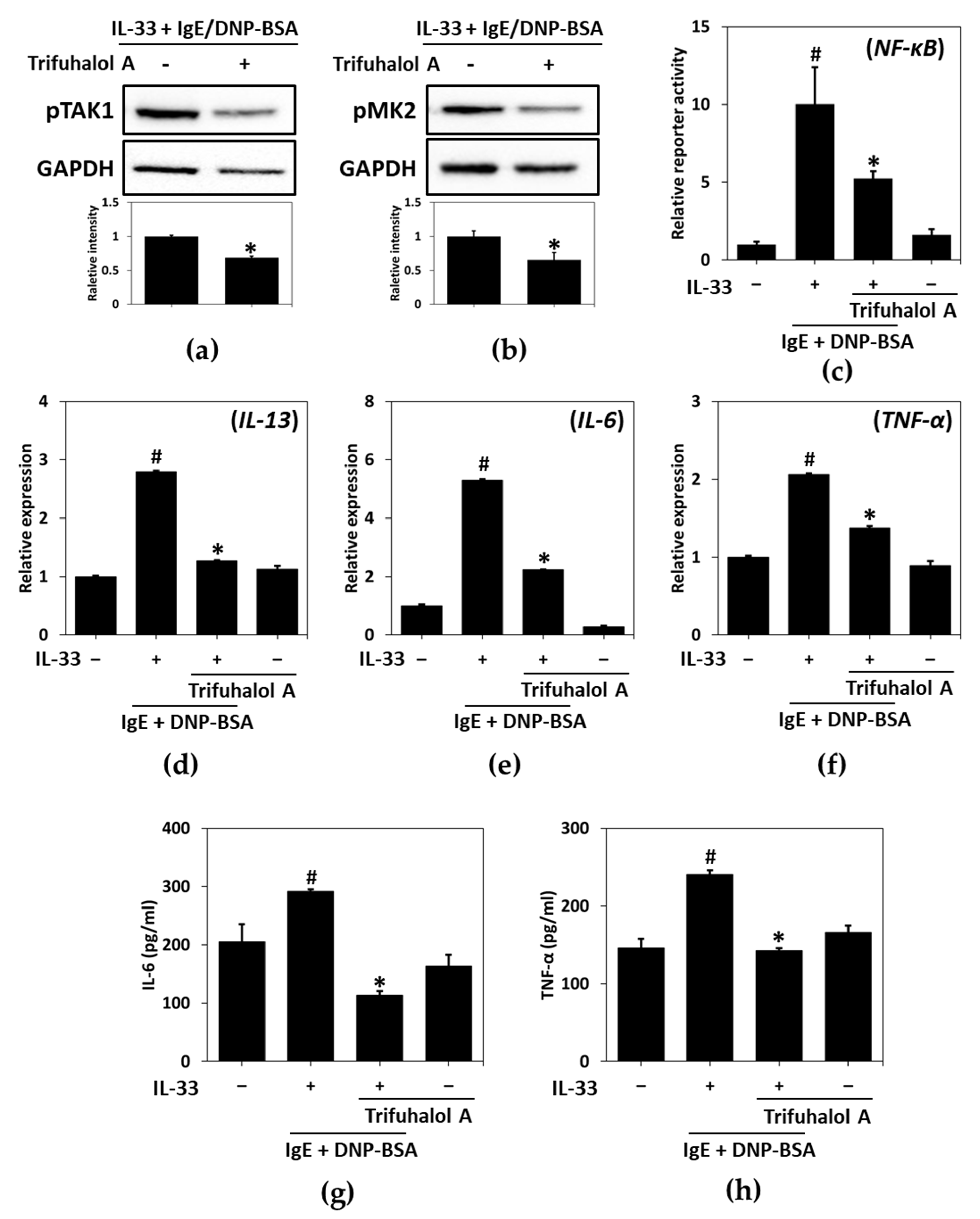
2.2. Effects of Trifuhalol A on IL-33- and IgE-Mediated Immune Cell Activation
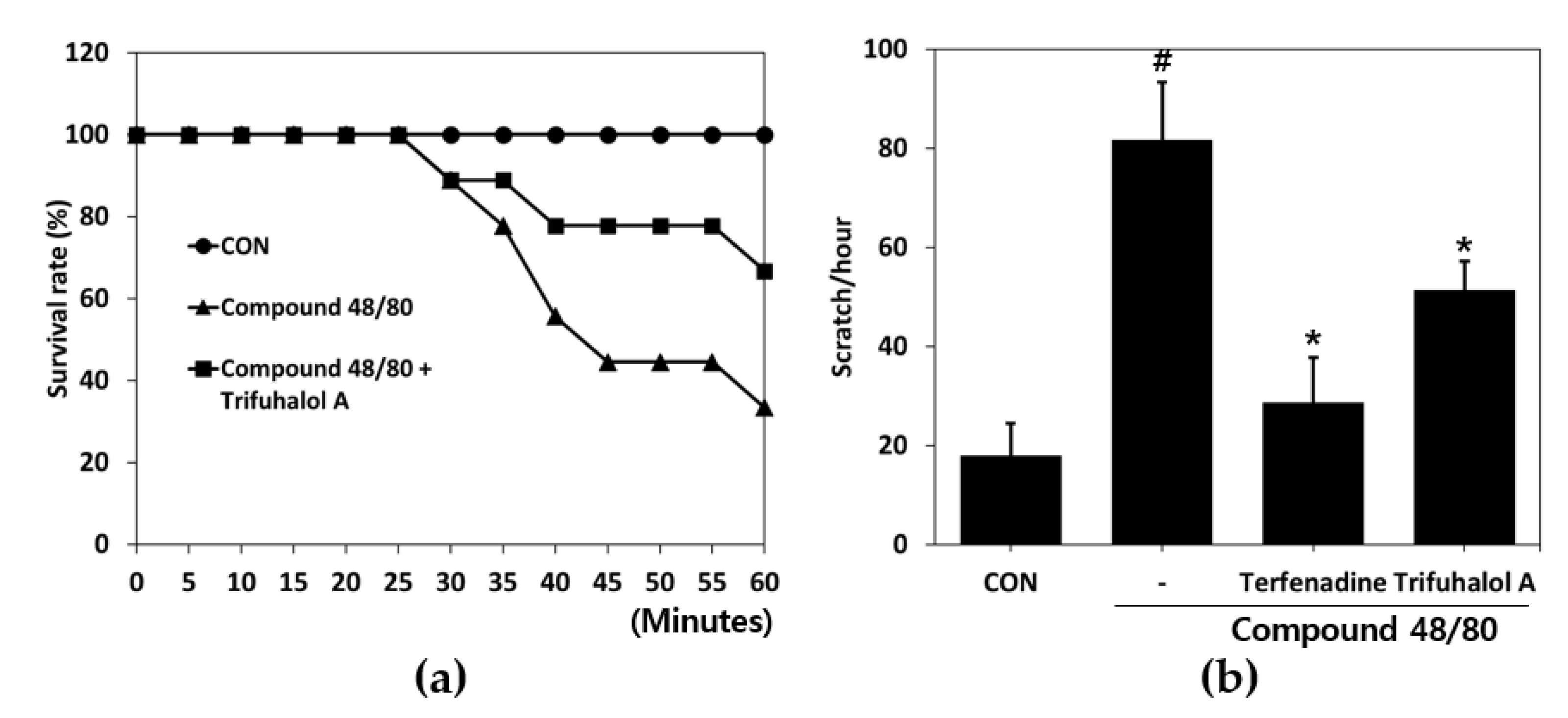
2.3. Effect of Trifuhalol A on Systemic Anaphylaxis and Itching
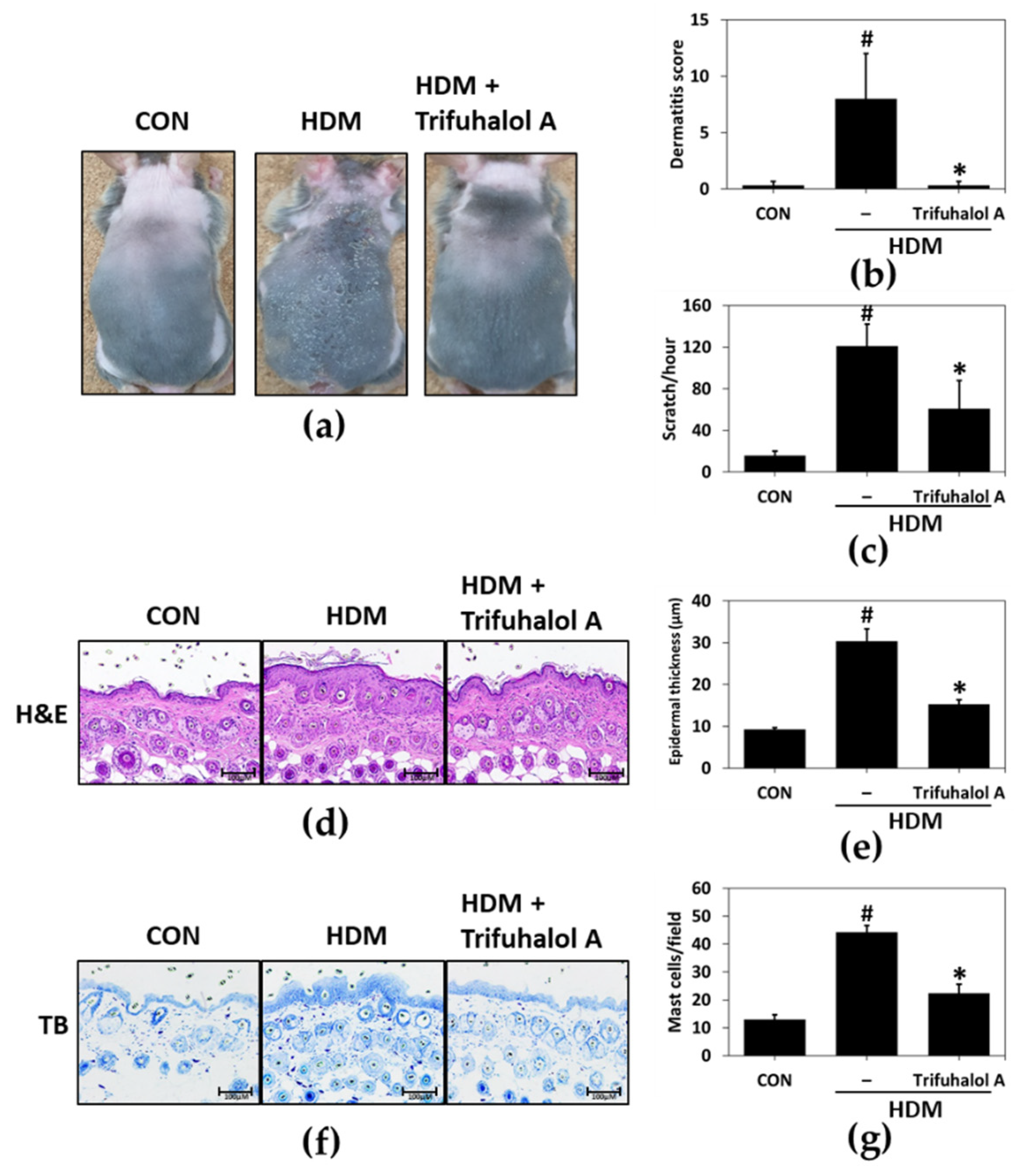
2.4. Effect of Trifuhalol A on HDM-Induced AD-like Lesions
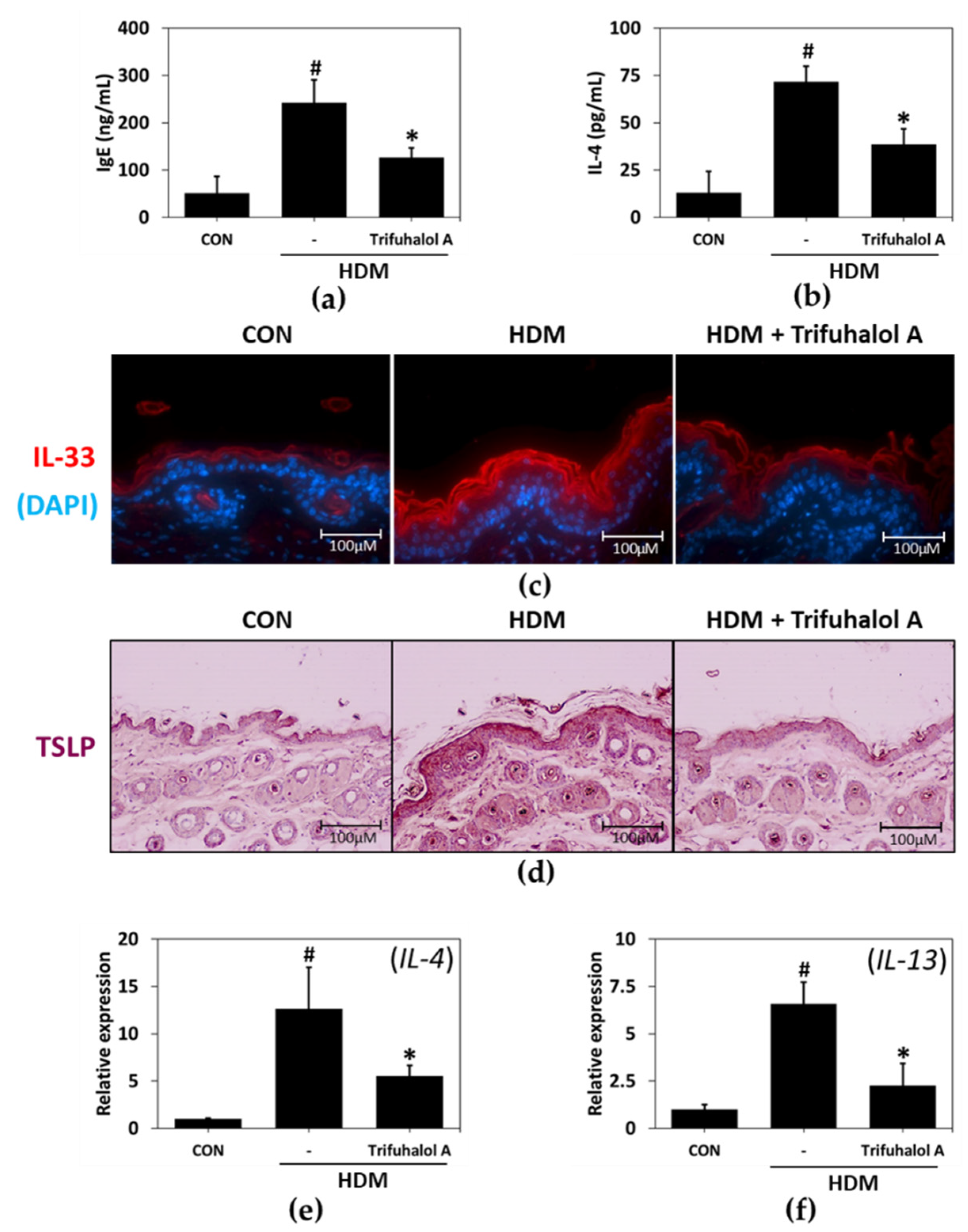
2.5. Effects of Trifuhalol A on Serum and Tissue Factors Associated with Allergic Inflammation in HDM-Induced AD-like Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals and Cells
4.2. Animals
4.3. β-Hexosaminidase Release from RBL-2H3 Cells
4.4. Detection of IL-33 in UVB-Irradiated and LL-37-Treated HaCaT cells
4.5. Assay of IgE from U266B1 Cells
4.6. Luciferase Reporter Gene Assay of NF-κB in RBL-2H3 Cells
4.7. Quantitative Real-Time PCR
4.8. Compound 48/80-Induced Systemic Anaphylactic Reaction
4.9. Compound 48/80-Induced Scratching Behavior Tests
4.10. Induction of AD by HDM in NC/Nga Mice
4.11. Measurement of Scratching Behavior Tests in NC/Nga Mice
4.12. Evaluation of the Skin Lesion (Dermatitis Score)
4.13. Histological Examination
4.14. Measurement of Total Serum IL-4 and IgE Levels
4.15. Measurement of Cytokine Levels
4.16. Immunohistochemistry
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fireman, P. Understanding asthma pathophysiology. Allergy Asthma Proc. 2003, 24, 79. [Google Scholar]
- Hansen, I.; Klimek, L.; Mösges, R.; Hörmann, K. Mediators of inflammation in the early and the late phase of allergic rhinitis. Curr. Opin. Allergy Clin. Immunol. 2004, 4, 159–163. [Google Scholar] [CrossRef]
- Katelaris, C. Ocular allergy: Implications for the clinical immunologist. Ann. Allergy Asthma Immunol. 2003, 90, 23–27. [Google Scholar] [CrossRef]
- Trocme, S.D.; Sra, K.K. Spectrum of ocular allergy. Curr. Opin. Allergy Clin. Immunol. 2002, 2, 423–427. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Adrian, M. Piliponsky AM The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef]
- Wang, Z.; Guhl, S.; Franke, K.; Artuc, M.; Zuberbier, T.; Babina, M. IL-33 and MRGPRX2-triggered activation of human skin mast cells—elimination of receptor expression on chronic exposure, but reinforced degranulation on acute priming. Cells 2019, 8, 341. [Google Scholar] [CrossRef]
- Hakim, M.M.; Patel, I.C. A review on phytoconstituents of marine brown algae. Future J. Pharm. Sci. 2020, 6, 129. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.; Mateus, N.; Cardoso, S.M. Optimization of phlorotannins extraction from Fucus vesiculosus and evaluation of their potential to prevent metabolic disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef]
- Phasanasophon, K.; Kim, S.M. Antioxidant and cosmeceutical activities of Agarum cribrosum phlorotannin extracted by ultrasound treatment. Nat. Prod. Commun. 2018, 13. [Google Scholar] [CrossRef]
- Phasanasophon, K.; Kim, S.M. Anti-inflammatory activity of the phlorotannin trifuhalol A using LPS-stimulated RAW264. 7 cells through NF-κB and MAPK main signaling pathways. Nat. Prod. Commun. 2019, 14, 1934578X19849798. [Google Scholar] [CrossRef]
- Cho, M.; Lee, D.-J.; Kim, J.-K.; You, S. Molecular characterization and immunomodulatory activity of sulfated fucans from Agarum cribrosum. Carbohydr. Polym. 2014, 113, 507–514. [Google Scholar] [CrossRef]
- Liao, B.-C.; Hou, R.C.-W.; Wang, J.-S.; Jeng, K.-C.G. Enhancement of the release of inflammatory mediators by substance P in rat basophilic leukemia RBL-2H3 cells. J. Biomed. Sci. 2006, 13, 613–619. [Google Scholar] [CrossRef]
- Rukwied, R.; Lischetzki, G.; McGlone, F.; Heyer, G.; Schmelz, M. Mast cell mediators other than histamine induce pruritus in atopic dermatitis patients: A dermal microdialysis study. Br. J. Dermatol. 2000, 142, 1114–1120. [Google Scholar] [CrossRef]
- Ohmura, T.; Hayashi, T.; Satoh, Y.; Konomi, A.; Jung, B.; Satoh, H. Involvement of substance P in scratching behaviour in an atopic dermatitis model. Eur. J. Pharmacol. 2004, 491, 191–194. [Google Scholar] [CrossRef]
- Yang, G.; Oh, J.-W.; Lee, H.E.; Lee, B.H.; Lim, K.-M.; Lee, J.Y. Topical Application of Dieckol Ameliorates Atopic Dermatitis in NC/Nga Mice by Suppressing Thymic Stromal Lymphopoietin Production. J. Investig. Dermatol. 2016, 136, 1062–1066. [Google Scholar] [CrossRef]
- Brandt, E.B.; Sivaprasad, U. Th2 cytokines and atopic dermatitis. J. Clin. Cell. Immunol. 2011, 2, 110. [Google Scholar] [CrossRef]
- Modena, B.D.; Dazy, K.; White, A.A. Emerging concepts: Mast cell involvement in allergic diseases. Transl. Res. 2016, 174, 98–121. [Google Scholar] [CrossRef]
- Adamczewski, M.; Kinet, J.-P. The high-affinity receptor for immunoglobulin E. Lymph. Act. 1994, 59, 173–190. [Google Scholar]
- Shin, T.-Y.; Kim, S.-H.; Suk, K.; Ha, J.-H.; Kim, I.; Lee, M.-G.; Jun, C.-D.; Kim, S.-Y.; Lim, J.-P.; Eun, J.-S. Anti-allergic effects of Lycopus lucidus on mast cell-mediated allergy model. Toxicol. Appl. Pharmacol. 2005, 209, 255–262. [Google Scholar] [CrossRef]
- Navinés-Ferrer, A.; Ainsua-Enrich, E.; Serrano-Candelas, E.; Proaño-Pérez, E.; Muñoz-Cano, R.; Gastaminza, G.; Olivera, A.; Martin, M. MYO1F Regulates IgE and MRGPRX2-Dependent Mast Cell Exocytosis. J. Immunol. 2021, 206, 2277–2289. [Google Scholar] [CrossRef]
- Andrade, M.V.; Iwaki, S.; Ropert, C.; Gazzinelli, R.T.; Cunha-Melo, J.R.; Beaven, M.A. Amplification of cytokine production through synergistic activation of NFAT and AP-1 following stimulation of mast cells with antigen and IL-33. Eur. J. Immunol. 2011, 41, 760–772. [Google Scholar] [CrossRef]
- Nakajima, S.; Ishimaru, K.; Kobayashi, A.; Yu, G.; Nakamura, Y.; Oh-Oka, K.; Suzuki-Inoue, K.; Kono, K.; Nakao, A. Resveratrol inhibits IL-33–mediated mast cell activation by targeting the MK2/3–PI3K/Akt axis. Sci. Rep. 2019, 9, 18423. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.-R.; Lee, A.Y.; Song, J.-H.; Yang, S.; Park, I.; Lim, J.-O.; Jung, T.-Y.; Ko, J.-W.; Kim, J.-C.; Lim, K.S. Scrophularia buergeriana attenuates allergic inflammation by reducing NF-κB activation. Phytomedicine 2020, 67, 153159. [Google Scholar] [CrossRef]
- Decker, W.W.; Campbell, R.L.; Manivannan, V.; Luke, A.; Sauver, J.L.S.; Weaver, A.; Bellolio, M.F.; Bergstralh, E.J.; Stead, L.G.; Li, J.T. The etiology and incidence of anaphylaxis in Rochester, Minnesota: A report from the Rochester Epidemiology Project. J. Allergy Clin. Immunol. 2008, 122, 1161–1165. [Google Scholar] [CrossRef]
- Rothschild, A. Mechanisms of histamine release by compound 48/80. Br. J. Pharmacol. 1970, 38, 253. [Google Scholar] [CrossRef]
- Dombrowicz, D.; Flamand, V.; Brigman, K.K.; Koller, B.H.; Kinet, J.-P. Abolition of anaphylaxis by targeted disruption of the high affinity immunoglobulin E receptor α chain gene. Cell 1993, 75, 969–976. [Google Scholar] [CrossRef]
- Takubo, M.; Ueda, Y.; Yatsuzuka, R.; Jiang, S.; Fujii, Y.; Kamei, C. Characteristics of scratching behavior induced by some chemical mediators in hairless mice. J. Pharmacol. Sci. 2006, 100, 285–288. [Google Scholar] [CrossRef]
- Renz, H.; Jujo, K.; Bradley, K.L.; Domenico, J.; Gelfand, E.W.; Leung, D.Y. Enhanced IL-4 production and IL-4 receptor expression in atopic dermatitis and their modulation by interferon-gamma. J. Investig. Dermatol. 1992, 99, 403–408. [Google Scholar] [CrossRef]
- Hvid, M.; Vestergaard, C.; Kemp, K.; Christensen, G.B.; Deleuran, B.; Deleuran, M. IL-25 in atopic dermatitis: A possible link between inflammation and skin barrier dysfunction? J. Investig. Dermatol. 2011, 131, 150–157. [Google Scholar] [CrossRef]
- Jujo, K.; Renz, H.; Abe, J.; Gelfand, E.W.; Leung, D.Y. Decreased interferon gamma and increased interleukin-4 production in atopic dermatitis promotes IgE synthesis. J. Allergy Clin. Immunol. 1992, 90, 323–331. [Google Scholar]
- Traidl, C.; Jugert, F.; Merk, H.; Krieg, T.; Hunzelmann, N. Inhibition of Allergic Contact Dermatitis to DNCB But Not to Oxazolone in Interleukin-4-Deficient Mice. J. Investig. Dermatol. 1999, 112, 476–482. [Google Scholar] [PubMed]
- Leung, D.Y.; Soter, N.A. Cellular and immunologic mechanisms in atopic dermatitis. J. Am. Acad. Dermatol. 2001, 44, S1–S12. [Google Scholar] [PubMed]
- Huang, W.-C.; Huang, C.-H.; Hu, S.; Peng, H.-L.; Wu, S.-J. Topical spilanthol inhibits MAPK signaling and ameliorates allergic inflammation in DNCB-induced atopic dermatitis in mice. Int. J. Mol. Sci. 2019, 20, 2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.-W.; Cheng, C.-C.; Hwang, T.-S.; Huang, W.-C.; Liou, C.-J.; Chen, W.-C.; Wu, S.-J. Danggui buxue tang inhibits 2, 4-dinitrochlorobenzene: Induced atopic dermatitis in mice. Evid. -Based Complementary Altern. Med. 2015, 2015, 672891. [Google Scholar]
- Jegal, J.; Park, N.-J.; Jo, B.-G.; Kim, T.-Y.; Bong, S.-K.; Choi, S.; Paik, J.-H.; Kim, J.-W.; Kim, S.-N.; Yang, M.H. Wikstroemia ganpi Extract Improved Atopic Dermatitis-Like Skin Lesions via Suppression of Interleukin-4 in 2, 4-Dinitrochlorobenzene-Induced SKH-1 Hairless Mice. Molecules 2021, 26, 2016. [Google Scholar]
- Yamamoto, M.; Haruna, T.; Yasui, K.; Takahashi, H.; Iduhara, M.; Takaki, S.; Deguchi, M.; Arimura, A. A novel atopic dermatitis model induced by topical application with dermatophagoides farinae extract in NC/Nga mice. Allergol. Int. 2007, 56, 139–148. [Google Scholar]
- Hashimoto, Y.; Arai, I.; Takano, N.; Tanaka, M.; Nakaike, S. Induction of scratching behaviour and dermatitis in various strains of mice cohabiting with NC/Nga mice with chronic dermatitis. Br. J. Dermatol. 2006, 154, 28–33. [Google Scholar] [CrossRef]
- Soumelis, V.; Liu, Y.-J. Human thymic stromal lymphopoietin: A novel epithelial cell-derived cytokine and a potential key player in the induction of allergic inflammation. Semin. Immunopathol. 2004, 25, 325–333. [Google Scholar] [CrossRef]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A. Human epithelial cells trigger dendritic cell–mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef]
- Lüthi, A.U.; Cullen, S.P.; McNeela, E.A.; Duriez, P.J.; Afonina, I.S.; Sheridan, C.; Brumatti, G.; Taylor, R.C.; Kersse, K.; Vandenabeele, P. Suppression of interleukin-33 bioactivity through proteolysis by apoptotic caspases. Immunity 2009, 31, 84–98. [Google Scholar] [PubMed]
- Chan, B.C.; Lam, C.W.; Tam, L.-S.; Wong, C.K. IL33: Roles in allergic inflammation and therapeutic perspectives. Front. Immunol. 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Paul, W.E.; Zhu, J. How are TH2-type immune responses initiated and amplified? Nat. Rev. Immunol. 2010, 10, 225–235. [Google Scholar]
- Oliphant, C.J.; Barlow, J.L.; McKenzie, A.N. Insights into the initiation of type 2 immune responses. Immunology 2011, 134, 378–385. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bong, S.-K.; Park, N.-J.; Lee, S.H.; Lee, J.W.; Kim, A.T.; Liu, X.; Kim, S.M.; Yang, M.H.; Kim, Y.K.; Kim, S.-N. Trifuhalol A Suppresses Allergic Inflammation through Dual Inhibition of TAK1 and MK2 Mediated by IgE and IL-33. Int. J. Mol. Sci. 2022, 23, 10163. https://doi.org/10.3390/ijms231710163
Bong S-K, Park N-J, Lee SH, Lee JW, Kim AT, Liu X, Kim SM, Yang MH, Kim YK, Kim S-N. Trifuhalol A Suppresses Allergic Inflammation through Dual Inhibition of TAK1 and MK2 Mediated by IgE and IL-33. International Journal of Molecular Sciences. 2022; 23(17):10163. https://doi.org/10.3390/ijms231710163
Chicago/Turabian StyleBong, Sim-Kyu, No-June Park, Sang Heon Lee, Jin Woo Lee, Aaron Taehwan Kim, Xiaoyong Liu, Sang Moo Kim, Min Hye Yang, Yong Kee Kim, and Su-Nam Kim. 2022. "Trifuhalol A Suppresses Allergic Inflammation through Dual Inhibition of TAK1 and MK2 Mediated by IgE and IL-33" International Journal of Molecular Sciences 23, no. 17: 10163. https://doi.org/10.3390/ijms231710163