
The Antibiotic Fosfomycin Mimics the Effects of the Intermediate Metabolites Phosphoenolpyruvate and Glyceraldehyde-3-Phosphate on the Stenotrophomonas maltophilia Transcriptome


Abstract
:1. Introduction
2. Results
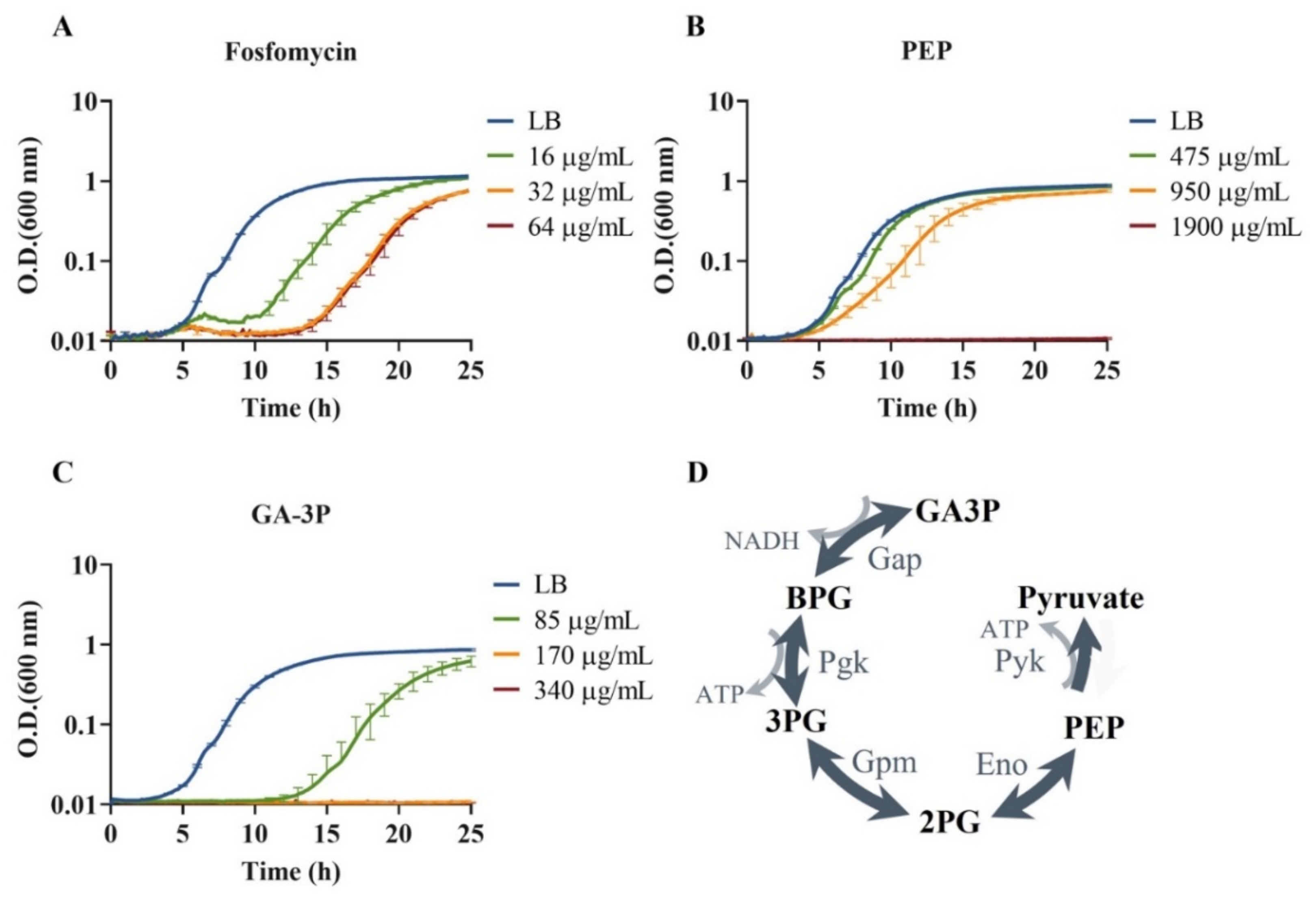
2.1. S. maltophilia’s Susceptibility to Fosfomycin, PEP, and GA-3P
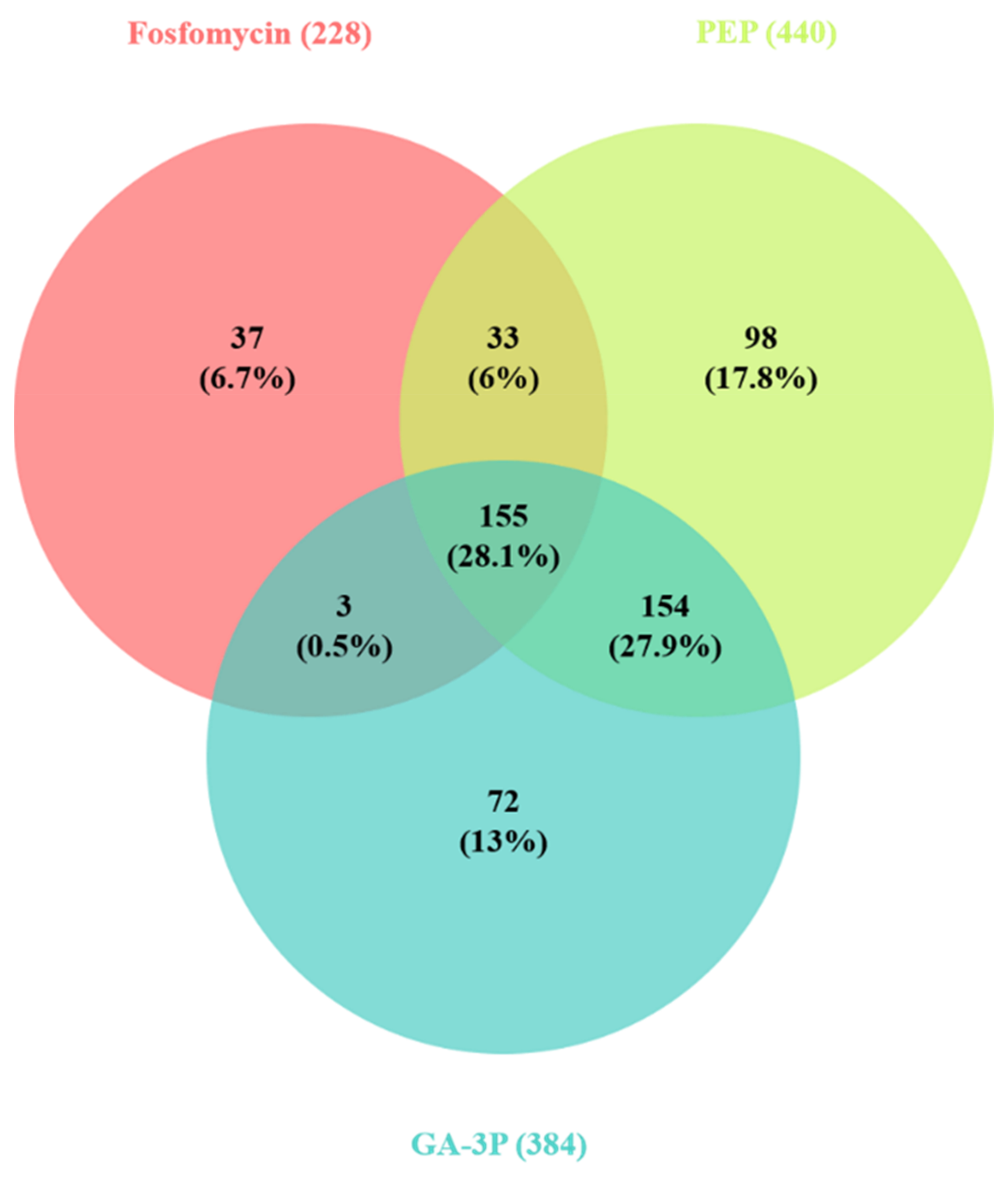
2.2. Effects of Fosfomycin, PEP, and GA-3P on S. maltophilia’s Transcriptome
2.3. Stress Responses Are Strongly Affected by Fosfomycin, PEP, and GA-3P Treatments
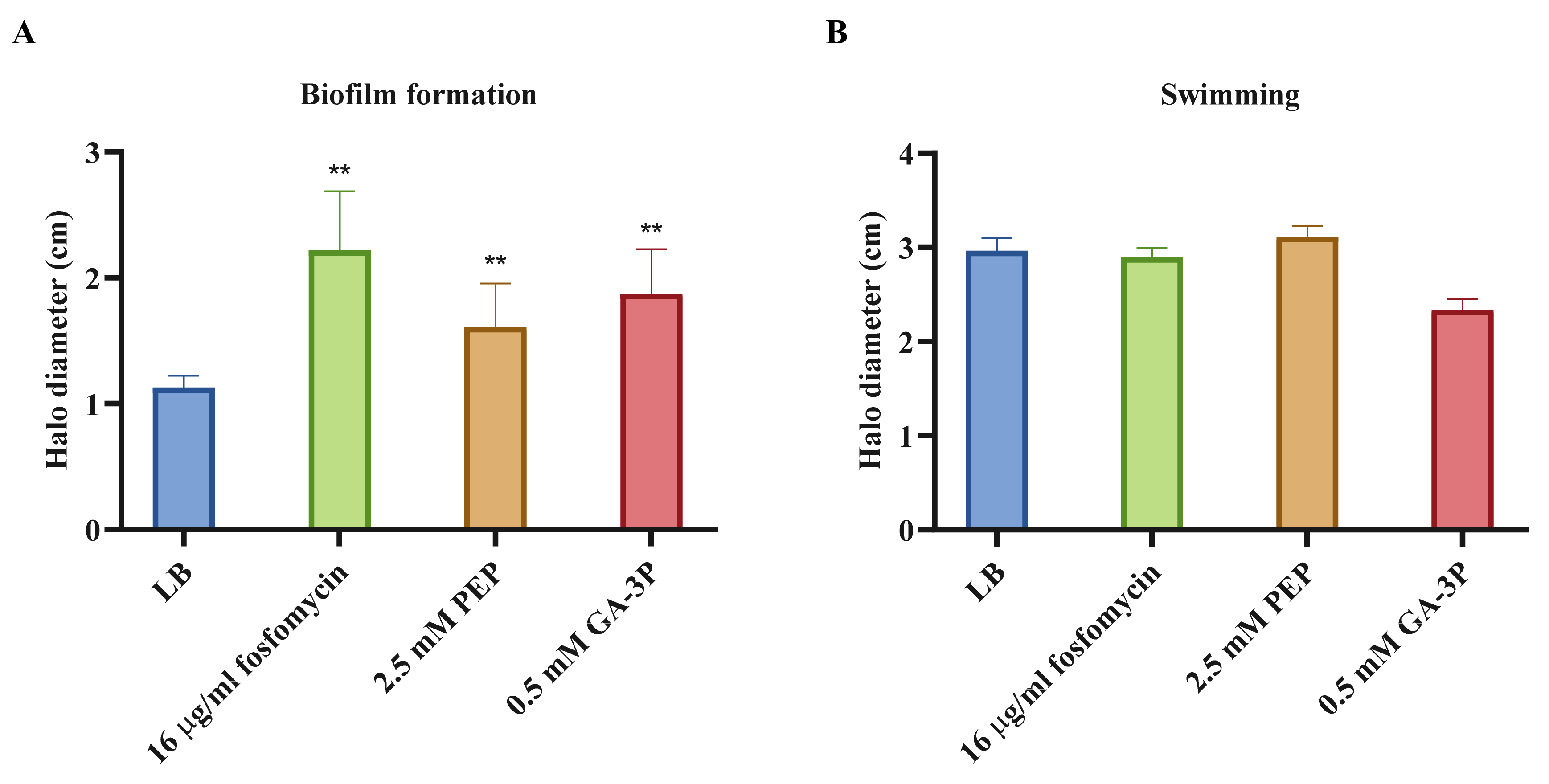
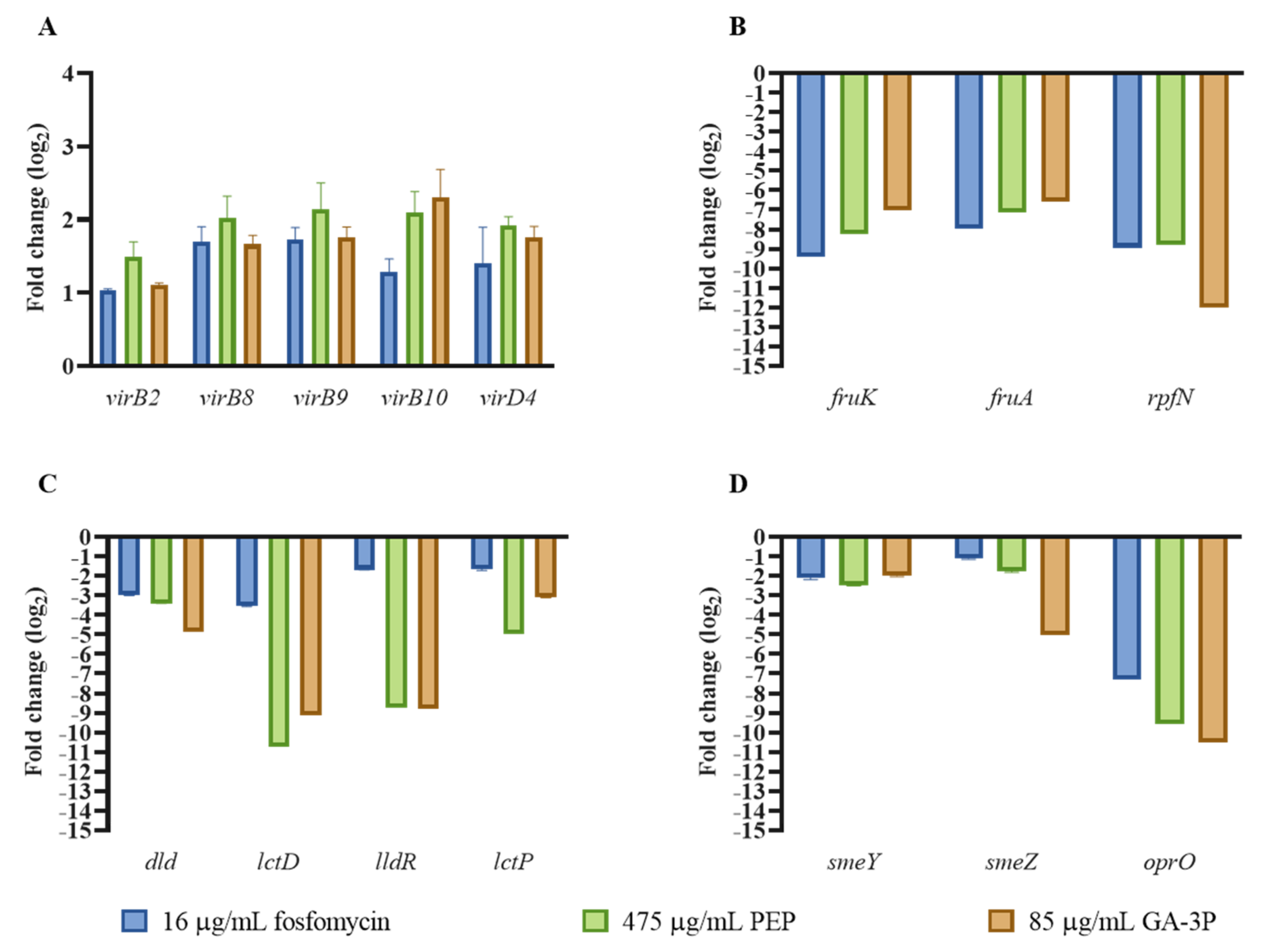
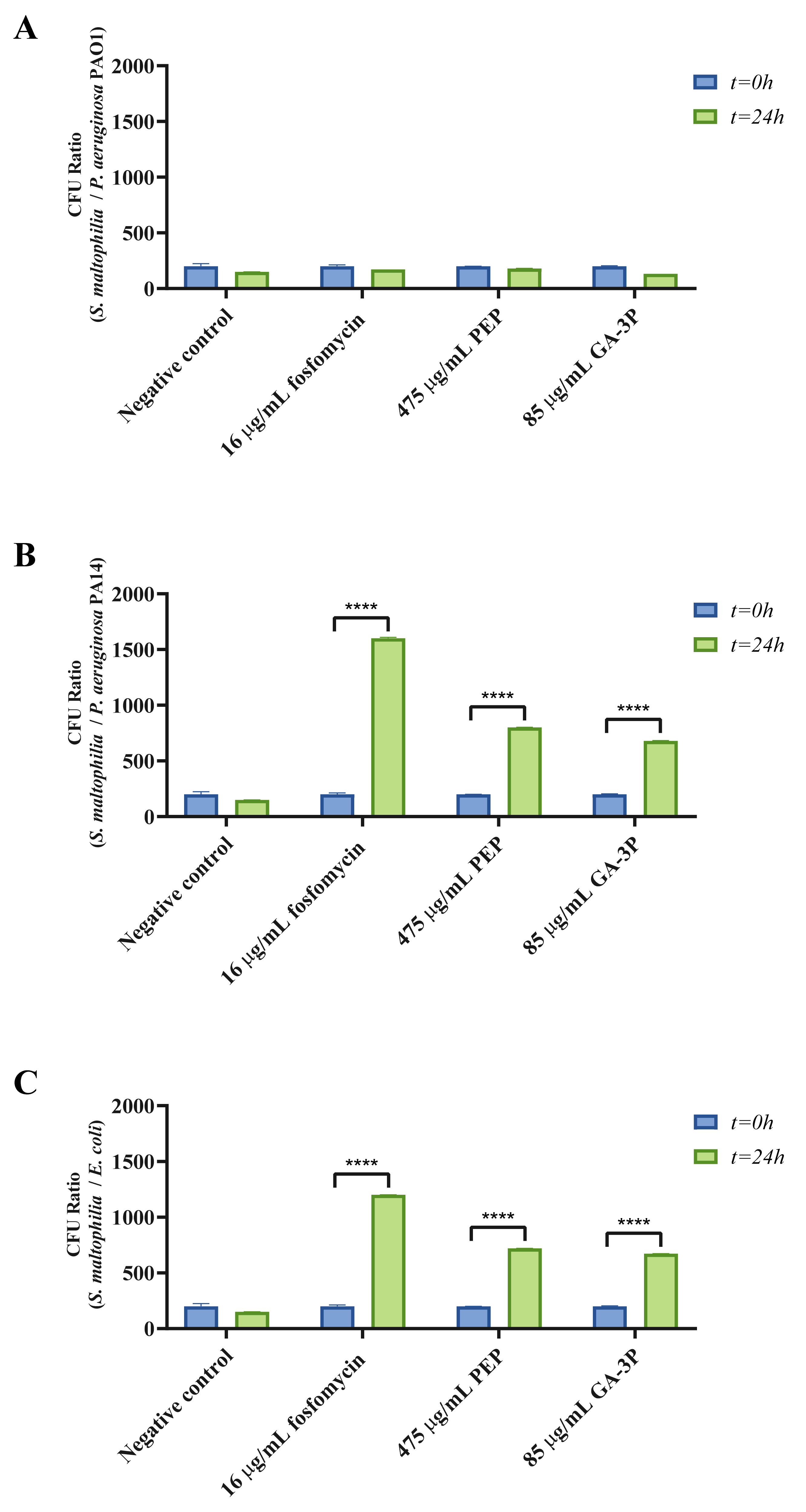
2.4. Motility, Chemotaxis, and Other Virulence-Related Genes Affected by the Three Different Treatments
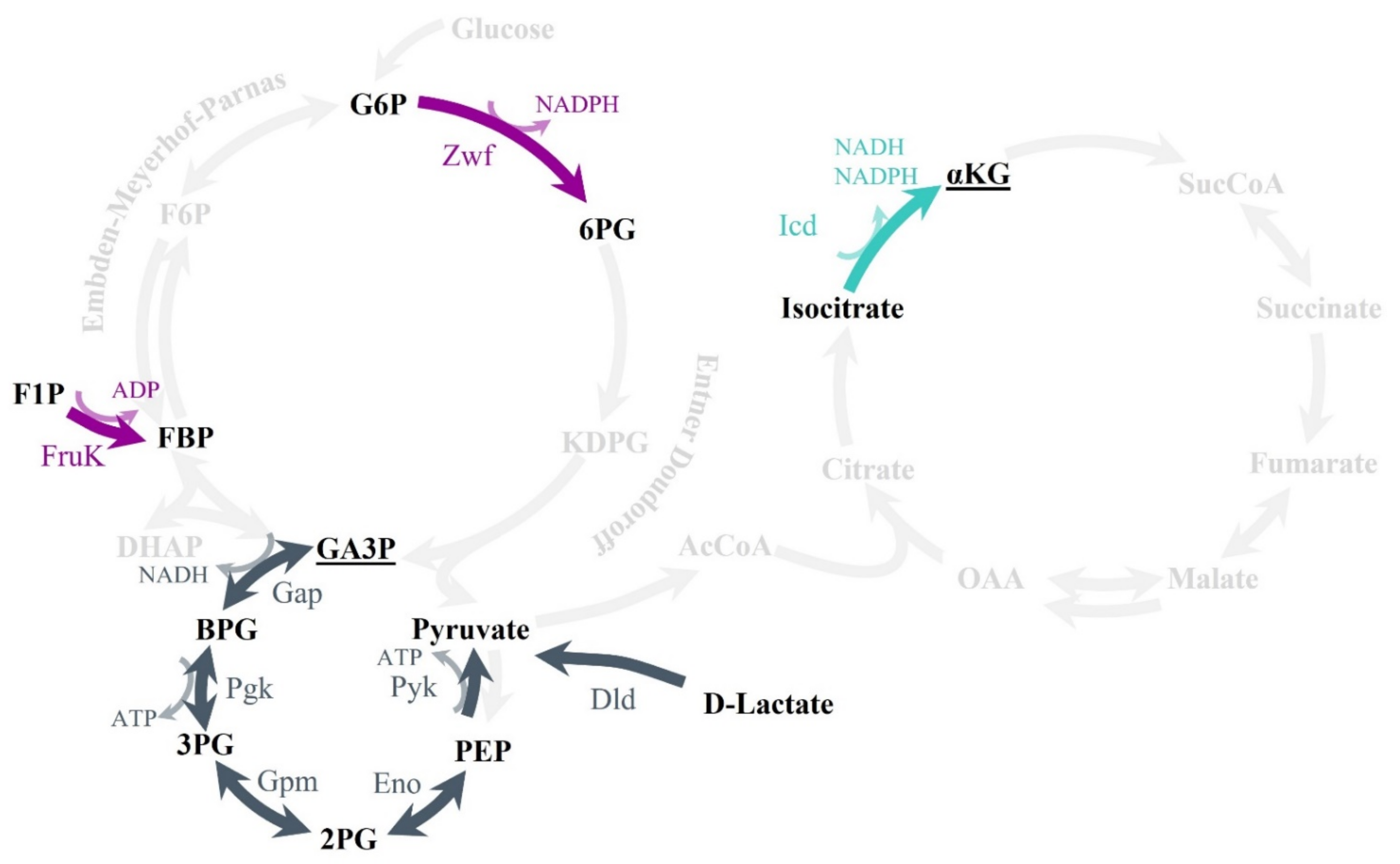
2.5. Metabolic Pathways Affected by Fosfomycin, PEP, and GA-3P Treatments
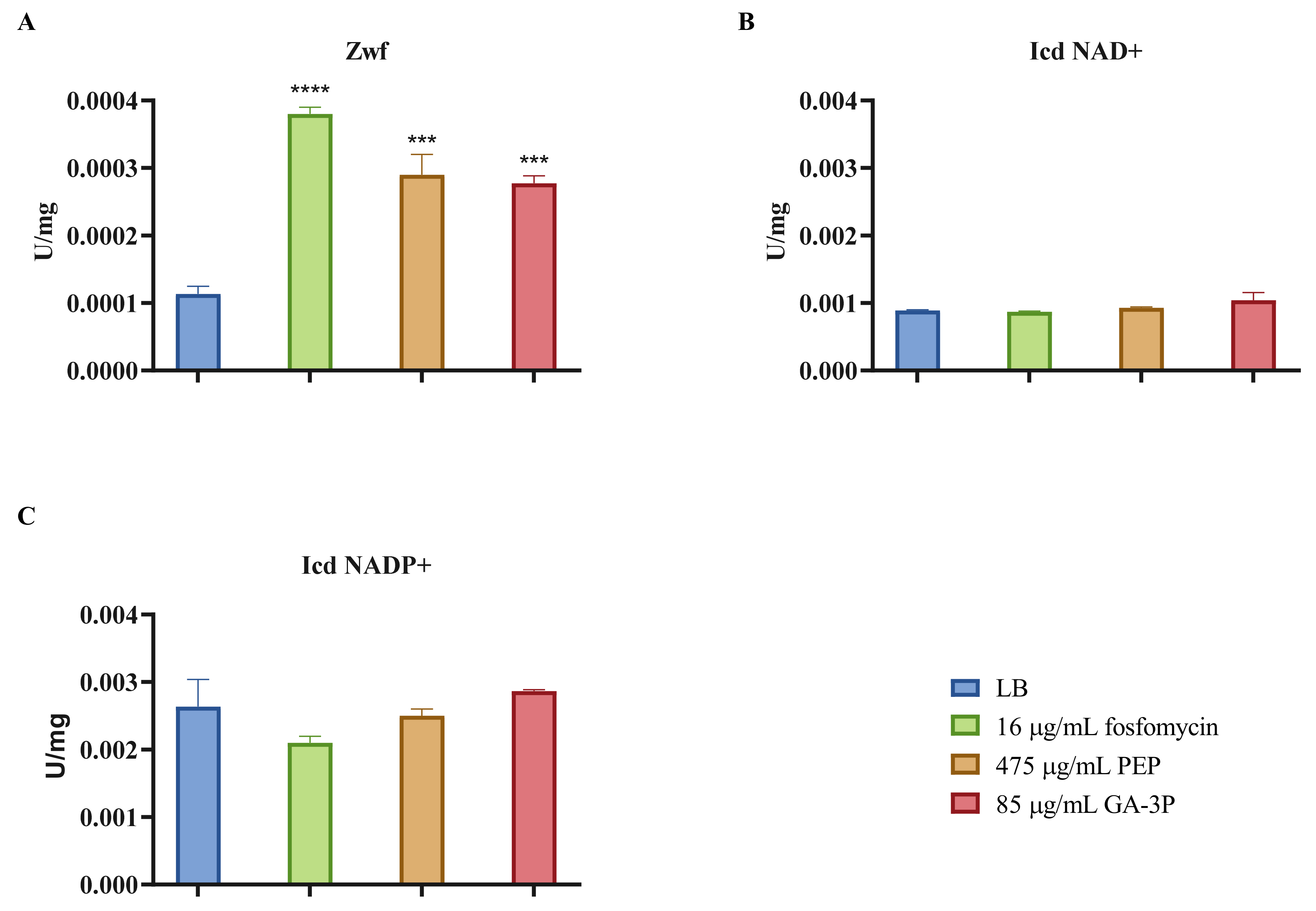
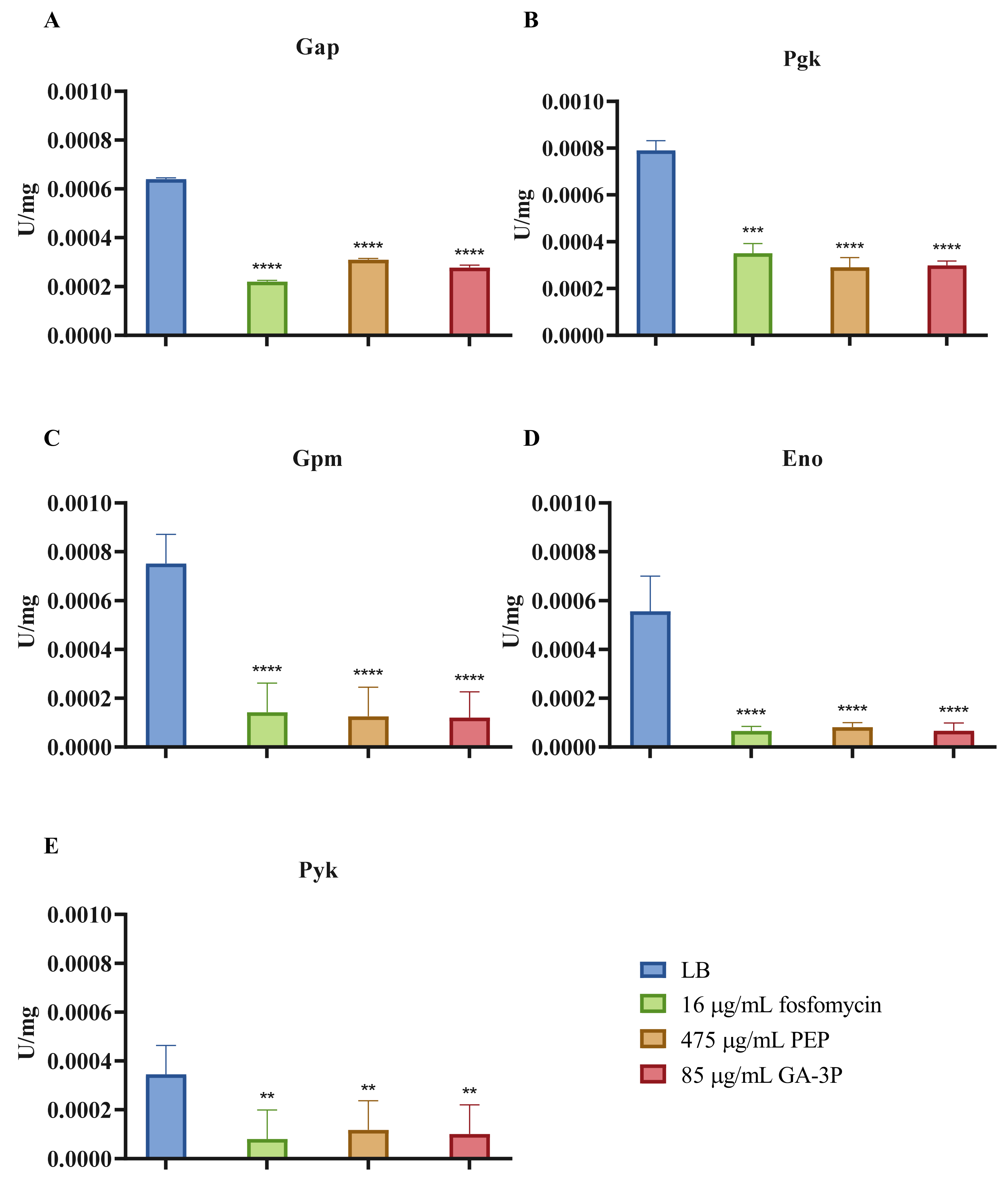
2.6. Effects of Fosfomycin, PEP, and GA-3P on the Central Metabolism: Enzymatic Activity of the Main Dehydrogenases and Lower Glycolysis Enzymes
2.7. General Transport Is Downregulated by Fosfomycin, PEP, and GA-3P
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Conditions
4.2. Bacterial Growth Measurement
4.3. Protein Quantification
4.4. In Vitro Activity Assays of the Enzymes of the Lower Glycolytic Pathway and Dehydrogenases
4.5. RNA Extraction and RNA-Seq
4.6. Quantitative Reverse Transcription PCR (qRT-PCR)
4.7. Biofilm Formation Assay
4.8. Swimming Assay
4.9. Bacterial Competition Assays
4.10. Antimicrobial Susceptibility Assays
4.11. BLASTp Search
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brazas, M.D.; Hancock, R.E. Using microarray gene signatures to elucidate mechanisms of antibiotic action and resistance. Drug Discov. Today 2005, 10, 1245–1252. [Google Scholar] [CrossRef]
- Fajardo, A.; Martinez-Martin, N.; Mercadillo, M.; Galan, J.C.; Ghysels, B.; Matthijs, S.; Cornelis, P.; Wiehlmann, L.; Tummler, B.; Baquero, F.; et al. The neglected intrinsic resistome of bacterial pathogens. PLoS ONE 2008, 3, e1619. [Google Scholar] [CrossRef] [Green Version]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as intermicrobial signaling agents instead of weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 379–411. [Google Scholar] [CrossRef]
- Davies, J.; Spiegelman, G.B.; Yim, G. The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 2006, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Gmuender, H.; Kuratli, K.; Di Padova, K.; Gray, C.P.; Keck, W.; Evers, S. Gene expression changes triggered by exposure of Haemophilus influenzae to novobiocin or ciprofloxacin: Combined transcription and translation analysis. Genome Res. 2001, 11, 28–42. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.L. Antibiotics and antibiotic resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [Green Version]
- Groschel, M.I.; Meehan, C.J.; Barilar, I.; Diricks, M.; Gonzaga, A.; Steglich, M.; Conchillo-Sole, O.; Scherer, I.C.; Mamat, U.; Luz, C.F.; et al. The phylogenetic landscape and nosocomial spread of the multidrug-resistant opportunist Stenotrophomonas maltophilia. Nat. Commun. 2020, 11, 2044. [Google Scholar] [CrossRef]
- Sader, H.S.; Jones, R.N. Antimicrobial susceptibility of uncommonly isolated non-enteric Gram-negative bacilli. Int. J. Antimicrob. Agents 2005, 25, 95–109. [Google Scholar] [CrossRef]
- Sanchez, M.B.; Hernandez, A.; Martinez, J.L. Stenotrophomonas maltophilia drug resistance. Future Microbiol. 2009, 4, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Hendlin, D.; Stapley, E.O.; Jackson, M.; Wallick, H.; Miller, A.K.; Wolf, F.J.; Miller, T.W.; Chaiet, L.; Kahan, F.M.; Foltz, E.L.; et al. Phosphonomycin, a new antibiotic produced by strains of streptomyces. Science 1969, 166, 122–123. [Google Scholar] [CrossRef]
- Kahan, F.M.; Kahan, J.S.; Cassidy, P.J.; Kropp, H. The mechanism of action of fosfomycin (phosphonomycin). Ann. N. Y. Acad. Sci. 1974, 235, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Skarzynski, T.; Mistry, A.; Wonacott, A.; Hutchinson, S.E.; Kelly, V.A.; Duncan, K. Structure of UDP-N-acetylglucosamine enolpyruvyl transferase, an enzyme essential for the synthesis of bacterial peptidoglycan, complexed with substrate UDP-N-acetylglucosamine and the drug fosfomycin. Structure 1996, 4, 1465–1474. [Google Scholar] [CrossRef] [Green Version]
- Falagas, M.E.; Vouloumanou, E.K.; Samonis, G.; Vardakas, K.Z. Fosfomycin. Clin. Microbiol. Rev. 2016, 29, 321–347. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Sandlin, R.C.; Maurelli, A.T. In vitro and in vivo functional activity of Chlamydia MurA, a UDP-N-acetylglucosamine enolpyruvyl transferase involved in peptidoglycan synthesis and fosfomycin resistance. J. Bacteriol. 2003, 185, 1218–1228. [Google Scholar] [CrossRef] [Green Version]
- De Smet, K.A.L.; Kempsell, K.E.; Gallagher, A.; Duncan, K.; Young, D.B. Alteration of a single amino acid residue reverses fosfomycin resistance of recombinant MurA from Mycobacterium tuberculosis. Microbiology 1999, 145 Pt 11, 3177–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Gilpin, M.E.; Attia, M.; Ting, Y.L.; Berti, P.J. Lyme disease enolpyruvyl-UDP-GlcNAc synthase: Fosfomycin-resistant MurA from Borrelia burgdorferi, a fosfomycin-sensitive mutant, and the catalytic role of the active site Asp. Biochemistry 2011, 50, 2205–2212. [Google Scholar] [CrossRef]
- Kim, D.H.; Lees, W.J.; Kempsell, K.E.; Lane, W.S.; Duncan, K.; Walsh, C.T. Characterization of a Cys115 to Asp substitution in the Escherichia coli cell wall biosynthetic enzyme UDP-GlcNAc enolpyruvyl transferase (MurA) that confers resistance to inactivation by the antibiotic fosfomycin. Biochemistry 1996, 35, 4923–4928. [Google Scholar] [CrossRef]
- Venkateswaran, P.S.; Wu, H.C. Isolation and characterization of a phosphonomycin-resistant mutant of Escherichia coli K-12. J. Bacteriol. 1972, 110, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Couce, A.; Briales, A.; Rodriguez-Rojas, A.; Costas, C.; Pascual, A.; Blazquez, J. Genomewide overexpression screen for fosfomycin resistance in Escherichia coli: MurA confers clinical resistance at low fitness cost. Antimicrob. Agents Chemother. 2012, 56, 2767–2769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Parvathi, A.; Hernandez, R.L.; Cadle, K.M.; Varela, M.F. Identification of a novel UDP-N-acetylglucosamine enolpyruvyl transferase (MurA) from Vibrio fischeri that confers high fosfomycin resistance in Escherichia coli. Arch. Microbiol. 2009, 191, 425–429. [Google Scholar] [CrossRef] [Green Version]
- Gisin, J.; Schneider, A.; Nagele, B.; Borisova, M.; Mayer, C. A cell wall recycling shortcut that bypasses peptidoglycan de novo biosynthesis. Nat. Chem. Biol. 2013, 9, 491–493. [Google Scholar] [CrossRef]
- Kadner, R.J.; Winkler, H.H. Isolation and characterization of mutations affecting the transport of hexose phosphates in Escherichia coli. J. Bacteriol. 1973, 113, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Tsuruoka, T.; Yamada, Y. Charactertization of spontaneous fosfomycin (phosphonomycin)-resistant cells of Escherichia coli B in vitro. J. Antibiot. 1975, 28, 906–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sass, P.; Jansen, A.; Szekat, C.; Sass, V.; Sahl, H.G.; Bierbaum, G. The lantibiotic mersacidin is a strong inducer of the cell wall stress response of Staphylococcus aureus. BMC Microbiol. 2008, 8, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigsby, R.E.; Fillgrove, K.L.; Beihoffer, L.A.; Armstrong, R.N. Fosfomycin resistance proteins: A nexus of glutathione transferases and epoxide hydrolases in a metalloenzyme superfamily. Methods Enzymol. 2005, 401, 367–379. [Google Scholar]
- Etienne, J.; Gerbaud, G.; Fleurette, J.; Courvalin, P. Characterization of staphylococcal plasmids hybridizing with the fosfomycin resistance gene fosB. FEMS Microbiol. Lett. 1991, 68, 119–122. [Google Scholar] [CrossRef]
- Bernat, B.A.; Laughlin, L.T.; Armstrong, R.N. Fosfomycin resistance protein (FosA) is a manganese metalloglutathione transferase related to glyoxalase I and the extradiol dioxygenases. Biochemistry 1997, 36, 3050–3055. [Google Scholar] [CrossRef]
- Fillgrove, K.L.; Pakhomova, S.; Schaab, M.R.; Newcomer, M.E.; Armstrong, R.N. Structure and mechanism of the genomically encoded fosfomycin resistance protein, FosX, from Listeria monocytogenes. Biochemistry 2007, 46, 8110–8120. [Google Scholar] [CrossRef]
- Gil-Gil, T.; Corona, F.; Martinez, J.L.; Bernardini, A. The Inactivation of Enzymes Belonging to the Central Carbon Metabolism Is a Novel Mechanism of Developing Antibiotic Resistance. mSystems 2020, 5, e00282-20. [Google Scholar] [CrossRef]
- Romano, A.H.; Conway, T. Evolution of carbohydrate metabolic pathways. Res. Microbiol. 1996, 147, 448–455. [Google Scholar] [CrossRef]
- Tang, C.T.; Engel, R.; Tropp, B.E. L-Glyceraldehude 3-phosphate, a bactericidal agent. Antimicrob. Agents Chemother. 1977, 11, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petek, M.; Baebler, S.; Kuzman, D.; Rotter, A.; Podlesek, Z.; Gruden, K.; Ravnikar, M.; Urleb, U. Revealing fosfomycin primary effect on Staphylococcus aureus transcriptome: Modulation of cell envelope biosynthesis and phosphoenolpyruvate induced starvation. BMC Microbiol. 2010, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baily, C.N.; Cason, R.W.; Vadvalkar, S.S.; Matsuzaki, S.; Humphries, K.M. Inhibition of mitochondrial respiration by phosphoenolpyruvate. Arch. Biochem. Biophys. 2011, 514, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utaida, S.; Dunman, P.M.; Macapagal, D.; Murphy, E.; Projan, S.J.; Singh, V.K.; Jayaswal, R.K.; Wilkinson, B.J. Genome-wide transcriptional profiling of the response of Staphylococcus aureus to cell-wall-active antibiotics reveals a cell-wall-stress stimulon. Microbiology 2003, 149, 2719–2732. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Jayaswal, R.K.; Wilkinson, B.J. Cell wall-active antibiotic induced proteins of Staphylococcus aureus identified using a proteomic approach. FEMS Microbiol. Lett. 2001, 199, 79–84. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Tomoyasu, T.; Takaya, A.; Morioka, M.; Yamamoto, T. Effects of disruption of heat shock genes on susceptibility of Escherichia coli to fluoroquinolones. BMC Microbiol. 2003, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Bayer-Santos, E.; Cenens, W.; Matsuyama, B.Y.; Oka, G.U.; Di Sessa, G.; Mininel, I.D.V.; Alves, T.L.; Farah, C.S. The opportunistic pathogen Stenotrophomonas maltophilia utilizes a type IV secretion system for interbacterial killing. PLoS Pathog. 2019, 15, e1007651. [Google Scholar] [CrossRef] [Green Version]
- Nas, M.Y.; White, R.C.; DuMont, A.L.; Lopez, A.E.; Cianciotto, N.P. Stenotrophomonas maltophilia Encodes a VirB/VirD4 Type IV Secretion System That Modulates Apoptosis in Human Cells and Promotes Competition against Heterologous Bacteria, Including Pseudomonas aeruginosa. Infect. Immun. 2019, 87, e00457-19. [Google Scholar] [CrossRef] [Green Version]
- Alegria, M.C.; Souza, D.P.; Andrade, M.O.; Docena, C.; Khater, L.; Ramos, C.H.; da Silva, A.C.; Farah, C.S. Identification of new protein-protein interactions involving the products of the chromosome- and plasmid-encoded type IV secretion loci of the phytopathogen Xanthomonas axonopodis pv. citri. J. Bacteriol. 2005, 187, 2315–2325. [Google Scholar] [CrossRef] [Green Version]
- Deutscher, J.; Francke, C.; Postma, P.W. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939–1031. [Google Scholar] [CrossRef] [Green Version]
- Cordaro, J.C.; Melton, T.; Stratis, J.P.; Atagun, M.; Gladding, C.; Hartman, P.E.; Roseman, S. Fosfomycin resistance: Selection method for internal and extended deletions of the phosphoenolpyruvate: Sugar phosphotransferase genes of Salmonella typhimurium. J. Bacteriol. 1976, 128, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Rabus, R.; Reizer, J.; Paulsen, I.; Saier, M.H., Jr. Enzyme I(Ntr) from Escherichia coli. A novel enzyme of the phosphoenolpyruvate-dependent phosphotransferase system exhibiting strict specificity for its phosphoryl acceptor, NPr. J. Biol. Chem. 1999, 274, 26185–26191. [Google Scholar] [CrossRef] [Green Version]
- Chavarría, M.; Goñi-Moreno, Á.; de Lorenzo, V.; Nikel, P.I. A Metabolic Widget Adjusts the Phosphoenolpyruvate-Dependent Fructose Influx in Pseudomonas putida. mSystems 2016, 1, e00154-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc collection of microbial genomes and metabolic pathways. Brief. Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Citak, F.; Ghai, I.; Rosenkotter, F.; Benier, L.; Winterhalter, M.; Wagner, R. Probing transport of fosfomycin through substrate specific OprO and OprP from Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2018, 495, 1454–1460. [Google Scholar] [CrossRef]
- Hancock, R.E.; Brinkman, F.S. Function of pseudomonas porins in uptake and efflux. Annu. Rev. Microbiol. 2002, 56, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Pechous, R.; Ledala, N.; Wilkinson, B.J.; Jayaswal, R.K. Regulation of the expression of cell wall stress stimulon member gene msrA1 in methicillin-susceptible or -resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2004, 48, 3057–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, W. NAD+/NADH and NADP+/NADPH in cellular functions and cell death: Regulation and biological consequences. Antioxid Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [Green Version]
- Tomoyasu, T.; Mogk, A.; Langen, H.; Goloubinoff, P.; Bukau, B. Genetic dissection of the roles of chaperones and proteases in protein folding and degradation in the Escherichia coli cytosol. Mol. Microbiol. 2001, 40, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Tomoyasu, T.; Goloubinoff, P.; Rudiger, S.; Roder, D.; Langen, H.; Bukau, B. Identification of thermolabile Escherichia coli proteins: Prevention and reversion of aggregation by DnaK and ClpB. EMBO J. 1999, 18, 6934–6949. [Google Scholar] [CrossRef]
- Thomas, J.G.; Baneyx, F. ClpB and HtpG facilitate de novo protein folding in stressed Escherichia coli cells. Mol. Microbiol. 2000, 36, 1360–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahn, R.; Buckle, A.M.; Perrett, S.; Johnson, C.M.; Corrales, F.J.; Golbik, R.; Fersht, A.R. Chaperone activity and structure of monomeric polypeptide binding domains of GroEL. Proc. Natl. Acad. Sci. USA 1996, 93, 15024–15029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, K.L.; O’Neill, A.J.; Mengin-Lecreulx, D.; Henderson, P.J.; Bostock, J.M.; Dunsmore, C.J.; Simmons, K.J.; Fishwick, C.W.; Leeds, J.A.; Chopra, I. The nature of Staphylococcus aureus MurA and MurZ and approaches for detection of peptidoglycan biosynthesis inhibitors. Mol. Microbiol. 2009, 72, 335–343. [Google Scholar] [CrossRef]
- Prior, T.I.; Kornberg, H.L. Nucleotide sequence of fruA, the gene specifying enzyme IIfru of the phosphoenolpyruvate-dependent sugar phosphotransferase system in Escherichia coli K12. J. Gen. Microbiol. 1988, 134, 2757–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanover, J.A.; Krause, M.W.; Love, D.C. The hexosamine signaling pathway: O-GlcNAc cycling in feast or famine. Biochim. Biophys. Acta 2010, 1800, 80–95. [Google Scholar] [CrossRef] [Green Version]
- Brauer, M.J.; Yuan, J.; Bennett, B.D.; Lu, W.; Kimball, E.; Botstein, D.; Rabinowitz, J.D. Conservation of the metabolomic response to starvation across two divergent microbes. Proc. Natl. Acad. Sci. USA 2006, 103, 19302–19307. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.T.; Huang, Y.W.; Chen, S.J.; Chang, C.W.; Yang, T.C. The SmeYZ Efflux Pump of Stenotrophomonas maltophilia Contributes to Drug Resistance, Virulence-Related Characteristics, and Virulence in Mice. Antimicrob. Agents Chemother. 2015, 59, 4067–4073. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.C.; Tsai, Y.L.; Huang, Y.W.; Chen, H.Y.; Hsueh, P.R.; Lai, S.Y.; Chen, L.C.; Chou, Y.H.; Lin, W.Y.; Liaw, S.J. Stenotrophomonas maltophilia PhoP, a Two-Component Response Regulator, Involved in Antimicrobial Susceptibilities. PLoS ONE 2016, 11, e0153753. [Google Scholar]
- Blanco, P.; Corona, F.; Martinez, J.L. Biolog Phenotype Microarray Is a Tool for the Identification of Multidrug Resistance Efflux Pump Inducers. Antimicrob. Agents Chemother. 2018, 62, e01263-18. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Zhang, L.; McKay, G.A.; Poole, K. Role of the acetyltransferase AAC(6′)-Iz modifying enzyme in aminoglycoside resistance in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2003, 51, 803–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.L.; Baquero, F. Interactions among strategies associated with bacterial infection: Pathogenicity, epidemicity, and antibiotic resistance. Clin. Microbiol. Rev. 2002, 15, 647–679. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Martínez, J.L. Interventions on Metabolism: Making Antibiotic-Susceptible Bacteria. mBio 2017, 8, e01950-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, A.; Martinez, J.L. Multiple antibiotic resistance in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 1997, 41, 1140–1142. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Martinez, J.L. Expression of multidrug efflux pump SmeDEF by clinical isolates of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2001, 45, 1879–1881. [Google Scholar] [CrossRef] [Green Version]
- Pawluk, A.; Scopes, R.K.; Griffiths-Smith, K. Isolation and properties of the glycolytic enzymes from Zymomonas mobilis. The five enzymes from glyceraldehyde-3-phosphate dehydrogenase through to pyruvate kinase. Biochem. J. 1986, 238, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Tag, M.H.; Wahab, S.M. Classification of the bovine farcy organism. Trop. Anim. Health Prod. 1977, 9, 124. [Google Scholar] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: https://bioinfogp.cnb.csic.es/tools/venny/ (accessed on 15 November 2021).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Category | Total Genes | Fosfomycin Transcriptome (Specific) | PEP Transcriptome (Specific) | GA-3P Transcriptome (Specific) |
|---|---|---|---|---|
| Stress response | 55 | 25 * (0 **) | 47 (5) | 49 (8) |
| Chemotaxis and motility | 50 | 28 (9) | 39 (2) | 34 (2) |
| Secretion systems | 6 | 0 (0) | 6 (1) | 5 (0) |
| Resistance | 5 | 1 (1) | 3 (1) | 3 (1) |
| Transport | 68 | 33 (6) | 55 (17) | 41 (7) |
| General metabolism | 51 | 11 (7) | 36 (31) | 40 (30) |
| Amino acid metabolism | 24 | 14 (2) | 19 (1) | 18 (3) |
| Outer membrane | 11 | 2 (1) | 9 (2) | 8 (1) |
| Regulators | 44 | 18 (3) | 31 (7) | 31 (10) |
| Replication and transcription | 21 | 6 (2) | 17 (6) | 13 (2) |
| Translation | 47 | 32 (4) | 35 (3) | 40 (7) |
| Iron-related proteins | 37 | 11 (2) | 32 (17) | 16 (3) |
| Other genes | 138 | 47 (8) | 111 (31) | 86 (19) |
| Fold Change (log2) | ||||
|---|---|---|---|---|
| ID—Gene | Fosfomycin | PEP | GA-3P | |
| SMD_RS00495 | 1.17 | 1.11 | 1.03 | |
| SMD_RS00575 | 0.97 | 1.21 | 1.12 | |
| SMD_RS02500 | 0.86 | 1.39 | 1.18 | |
| SMD_RS02505 | 0.67 | 1.13 | 1.12 | |
| SMD_RS02645 | msrB | 1.94 | 2.48 | 2.45 |
| SMD_RS03830 | msrA | 1.06 | 1.58 | 1.62 |
| SMD_RS04070 | 0.42 | 1.05 | 1.04 | |
| SMD_RS04515 | grxC | 0.83 | 1.05 | 1.06 |
| SMD_RS04605 | lon | 1.16 | 1.41 | 1.4 |
| SMD_RS05310 | soxR | 1.21 | 1.59 | 1.22 |
| SMD_RS06170 | 0.99 | 1.27 | 1.32 | |
| SMD_RS06465 | 1.21 | 1.38 | 1.38 | |
| SMD_RS09215 | htpG | 1.4 | 1.75 | 1.79 |
| SMD_RS09380 | 0.64 | 0.95 | 1.16 | |
| SMD_RS09475 | hrcA | 0.97 | 1.31 | 1.36 |
| SMD_RS09480 | grpE | 1.19 | 1.55 | 1.56 |
| SMD_RS09485 | dnaK | 1.08 | 1.36 | 1.39 |
| SMD_RS09490 | dnaJ | 0.88 | 1.05 | 0.99 |
| SMD_RS11005 | 1.46 | 1.23 | 1.04 | |
| SMD_RS11055 | clpA | 0.95 | 1.35 | 1.42 |
| SMD_RS11800 | 0.97 | 1.06 | 0.89 | |
| SMD_RS13010 | −0.28 | −1.07 | −1.15 | |
| SMD_RS13025 | −0.02 | 0.98 | 1.24 | |
| SMD_RS13030 | −0.18 | 0.87 | 1.12 | |
| SMD_RS13045 | 1.13 | 1.47 | 1.41 | |
| SMD_RS13060 | −1.01 | −1.25 | −0.77 | |
| SMD_RS14070 | 1.35 | 1.59 | 1.52 | |
| SMD_RS14255 | sufA | 0.79 | 1.02 | 1.01 |
| SMD_RS14660 | sodA | 1.2 | 1.59 | 1.64 |
| SMD_RS15050 | htpX | 1.01 | 1.08 | 1.22 |
| SMD_RS15740 | 0.8 | 1.19 | 1.01 | |
| SMD_RS15845 | tldD | 0.8 | 0.99 | 1 |
| SMD_RS15930 | trxA | 1.71 | 1.98 | 1.91 |
| SMD_RS16120 | sphB | 0.87 | 1.36 | 1.12 |
| SMD_RS16195 | 1.23 | 1.52 | 1.48 | |
| SMD_RS16245 | 1.24 | 1.41 | 1.56 | |
| SMD_RS16285 | cstA | 0.61 | 0.25 | 1.12 |
| SMD_RS16290 | 0.37 | 0.18 | 1.03 | |
| SMD_RS16330 | 0.92 | 1.09 | 0.91 | |
| SMD_RS16640 | hslO | 1.35 | 1.52 | 1.6 |
| SMD_RS17335 | clpB | 1.24 | 1.74 | 1.77 |
| SMD_RS17345 | 0.74 | 1.13 | 1.42 | |
| SMD_RS18465 | 1.8 | 2.12 | 1.85 | |
| SMD_RS18955 | hslU | 1.79 | 2.14 | 2.2 |
| SMD_RS18960 | hslV | 1.71 | 2.19 | 2.2 |
| SMD_RS18990 | ptrB | 1.08 | 1.46 | 1.42 |
| SMD_RS19000 | 0.91 | 1.02 | 0.88 | |
| SMD_RS19045 | prlC | 1.39 | 1.78 | 1.77 |
| SMD_RS19280 | 0.67 | 1.12 | 1.16 | |
| SMD_RS19325 | 0.9 | 1.48 | 0.94 | |
| SMD_RS19815 | groL | 0.76 | 0.99 | 1.05 |
| SMD_RS19820 | groES | 1.08 | 1.41 | 1.39 |
| SMD_RS21040 | gst6 | 0.77 | 1.18 | 1.19 |
| SMD_RS21540 | 1.09 | 1.32 | 1.73 | |
| SMD_RS18790 | 0.62 | 0.64 | 1.12 | |
| Fold Change (log2) | ||||
|---|---|---|---|---|
| ID—Gene | Fosfomycin | PEP | GA-3P | |
| SMD_RS13770 | virB2 | 0.83 | 1.17 | 1.06 |
| SMD_RS13785 | virB10 | 0.94 | 1.3 | 1.11 |
| SMD_RS13790 | virB9 | 0.98 | 1.4 | 1.24 |
| SMD_RS13795 | virB8 | 0.92 | 1.23 | 1.09 |
| SMD_RS13805 | virD4 | 0.64 | 1.16 | 1.03 |
| Fold Change (log2) | ||||
|---|---|---|---|---|
| ID—Gene or Product | Fosfomycin | PEP | GA-3P | |
| SMD_RS11695 | ptsP | −4.26 | −2.63 | −1.6 |
| SMD_RS11700 | fruK | −5.08 | −5.11 | −1.76 |
| SMD_RS11705 | fruA | −5.06 | −5.09 | −1.78 |
| SMD_RS11710 | rpfN | −3.53 | −4.93 | −2.05 |
| SMD_RS13320 | dld | −1.53 | −3.25 | −0.74 |
| SMD_RS13325 | lctD | −1.4 | −2.9 | −0.67 |
| SMD_RS13330 | lldR | −1.42 | −2.3 | −0.77 |
| SMD_RS13335 | lctP | −1.34 | −3.77 | −0.56 |
| SMD_RS14045 | hutF | 1.11 | 1.29 | 1 |
| SMD_RS14050 | hutI | 1.08 | 1.59 | 1.33 |
| SMD_RS14055 | hutH | 1.3 | 1.53 | 1.45 |
| SMD_RS14060 | hutG | 1.38 | 1.85 | 1.81 |
| SMD_RS14065 | hutU | 2.2 | 2.51 | 2.52 |
| Fold Change (log2) | ||||
|---|---|---|---|---|
| ID—Gene | Fosfomycin | PEP | GA-3P | |
| SMD_RS09135 | zwf | −0.21 | −0.02 | 0.15 |
| SMD_RS04525 | icd (NAD+) | 0.24 | 0.15 | 0.17 |
| SMD_RS20085 | icd (NADP+) | 0.56 | 0.47 | 0.38 |
| SMD_RS17680 | gap | 0.24 | 0.35 | 0.36 |
| SMD_RS17665 | pgk | 0.48 | 0.4 | 0.31 |
| SMD_RS06650 | gpmA | 0.26 | 0.33 | 0.39 |
| SMD_RS04430 | gpm | −0.53 | −1.11 | −1.21 |
| SMD_RS08765 | eno | 0.64 | 0.69 | 0.6 |
| SMD_RS17655 | pyk | 0.22 | 0.43 | 0.14 |
| Fold Change (log2) | ||||
|---|---|---|---|---|
| ID—Gene | Fosfomycin | PEP | GA-3P | |
| SMD_RS10440 | smeY | −0.99 | −1.77 | −1.76 |
| SMD_RS10445 | smeZ | −1.09 | −1.78 | −1.65 |
| SMD_RS18400 | oprO | −6.52 | −6.55 | −2.8 |
| MIC (μg/mL) | ||||
|---|---|---|---|---|
| LB | Fosfomycin (16 μg/mL) | PEP (475 μg/mL) | GA-3P (85 μg/mL) | |
| Amikacin | 6 | 3 | 3 | 3 |
| Tobramycin | 4 | 2 | 1.5 | 2 |
| Gentamicin | 2 | 1.25 | 1 | 1.25 |
| Tetracycline | 2 | 1.5 | 0.75 | 1 |
| SXT | 0.3 | 0.19 | 0.125 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Gil, T.; Ochoa-Sánchez, L.E.; Martínez, J.L. The Antibiotic Fosfomycin Mimics the Effects of the Intermediate Metabolites Phosphoenolpyruvate and Glyceraldehyde-3-Phosphate on the Stenotrophomonas maltophilia Transcriptome. Int. J. Mol. Sci. 2022, 23, 159. https://doi.org/10.3390/ijms23010159
Gil-Gil T, Ochoa-Sánchez LE, Martínez JL. The Antibiotic Fosfomycin Mimics the Effects of the Intermediate Metabolites Phosphoenolpyruvate and Glyceraldehyde-3-Phosphate on the Stenotrophomonas maltophilia Transcriptome. International Journal of Molecular Sciences. 2022; 23(1):159. https://doi.org/10.3390/ijms23010159
Chicago/Turabian StyleGil-Gil, Teresa, Luz Edith Ochoa-Sánchez, and José Luis Martínez. 2022. "The Antibiotic Fosfomycin Mimics the Effects of the Intermediate Metabolites Phosphoenolpyruvate and Glyceraldehyde-3-Phosphate on the Stenotrophomonas maltophilia Transcriptome" International Journal of Molecular Sciences 23, no. 1: 159. https://doi.org/10.3390/ijms23010159